
Insecticide susceptibility and resistance development in ...

Eo
HRH
a
ARRAA
KSBERC
1
tiabca2vratt(di
0d
Toxicology 253 (2008) 89–96
Contents lists available at ScienceDirect
Toxicology
journa l homepage: www.e lsev ier .com/ locate / tox ico l
nantioselective cytotoxicity of the insecticide bifenthrinn a human amnion epithelial (FL) cell line
uigang Liu, Meirong Zhao, Cong Zhang, Yun Ma, Weiping Liu ∗
esearch Center of Environmental Science, College of Biological and Environmental Engineering, Zhejiang University of Technology,angzhou 310032, People’s Republic of China
r t i c l e i n f o
rticle history:eceived 11 July 2008eceived in revised form 25 August 2008ccepted 26 August 2008vailable online 7 September 2008
eywords:ynthetic pyrethroidsifenthrinnantioselectivity
a b s t r a c t
Synthetic pyrethroids (SPs) are used in preference to organochlorines and organophosphates due to theirhigh efficiency, low toxicity to mammals, and ready biodegradability. Previous studies reported that enan-tioselective toxicity of SPs occurs in aquatic toxicity. Several studies have indicated that SPs could leadto oxidative damage in humans or animals which was associated with their toxic effects. Little is knownabout the differences in the effects of chronic toxicity induced by individual stereoisomers of chiral SPs.The present study was therefore undertaken to evaluate the enantioselectivity in cytotoxicity, genotoxi-city caused by bifenthrin (BF) on human amnion epithelial (FL) cell lines and pesticidal activity on targetorganism. The cell proliferation and cytoflow analysis indicated that 1S-cis-BF presented more toxic effectsthan 1R-cis-BF above the concentration of 7.5 mg L−1 (p > 0.05). FL cells incubated with 1S-cis-BF exhib-
OSytotoxicity
ited a dose-dependent accumulation of intracellular reactive oxygen species (ROS). In the comet assay,the number of cells with damaged DNA incubated with 1S-cis-BF was more than that with 1R-cis-BF(p < 0.01). While the LC50 values of enantiomer to the target pest on Pieris rapae L. show that 1R-cis-BF was300 times more active than 1S-cis-BF. These results indicate that the enantioselective toxicity and activityof BF between non-target organism and target organism was reversal. These implications together suggestthat assessment of the environmental safety and new pesticides development with chiral centers should
y.Crow
ttairceeicwets
consider enantioselectivit
. Introduction
A chiral compound has two (or more) enantiomers that havehe same physical–chemical properties and identical behaviorsn achiral environments. However, as biological systems gener-lly distinguish chirality, enantiomers of the same pesticide mayehave as two completely different compounds with regard to bio-hemical and physiological activity (Enquist and Arak, 1994; Yoonnd Jacobsen, 2003). Chiral compounds account for approximately5% of all pesticides used commercially and for 26% of the totalalue of the world pesticide market in 1996 (Williams, 1996). Thisatio is increasing as compounds with more complex structuresre introduced into use (Liu et al., 2005c). After a decade, morehan 40% of currently used pesticides in China have chiral struc-
ures because the main classes of them are synthetic pyrethroidsSPs) and organophosphates as insecticides, and metolachlor andiphenyl ethers as herbicides (Liu, 2006). For chiral pesticides, typ-cally only one enantiomer is active against the target pests, while
∗ Corresponding author. Tel: +86 571 8832 0666; fax: +86 571 8832 0884.E-mail address: [email protected] (W. Liu).
fd
lefat
300-483X/$ – see front matter. Crown Copyright © 2008 Published by Elsevier Ireland Ltoi:10.1016/j.tox.2008.08.015
n Copyright © 2008 Published by Elsevier Ireland Ltd. All rights reserved.
he other enantiomer is inactive and serves as an unwanted burdeno the environment. Since most chiral pesticides are still marketeds racemates, it is important to evaluate their enantioselectivitiesn environmental safety, ecotoxicological and health risks. Severalecent studies have focused on the effect of enantioselectivity ofhiral pesticides in environmental fate, acute aquatic toxicity (Liut al., 2005b,c), embryo development (Xu et al., 2008), carcinogen-sis, and endocrine disrupting activities (Wang et al., 2007). Anmportant relationship of some adverse effects of pesticides andytotoxicity has been illustrated in a few previous studies. Mean-hile, the assessment of cytotoxicity is an effective method for
valuating the environmental and ecological toxicities. However,he role of enantioselective toxicity on cell lines is poorly under-tood for pesticides, and the knowledge gap is reflected upon theact that the great majority of chiral pesticides are being used andeveloped as if they were achiral.
SPs are among the most commonly used pesticides for control-
ing agricultural and indoor pests. In particular, SPs are commonlymployed in households to eradicate pests and insects, and there-ore the exposure of humans to SPs may be extensive (Morgan etl., 2007; Ostrea et al., 2006). The liberal use of SPs has increasedhe risk of intoxication in non-target species, such as birds, humans,d. All rights reserved.

90 H. Liu et al. / Toxicology 2
fisooiacttpsfiiIdsBagtpt
cawIit
ietSou
2
2
(c(wpew
dcl
2
(aCdNiJTwtA(c
2
1tE(f
2
(rtzDwogarU
2
p5bsFtaX
2
udi(sflmii2(
2
Fig. 1. Enantiomers of cis-bifenthrin (BF).
shes, and organisms present in soil and water (Hill, 1989). Recenttudies have found that SPs from agricultural and residential run-ffs induced toxicological effects on aquatic organisms. Resultsf some in vitro tests indicated pyrethroids displayed cytotoxic-ty including DNA strand breaks and gene mutation (Surrallés etl., 1995a,b; Majumder et al., 1994). The in vitro tests especially cellulture tests are sensitive tests for the assessment of environmentaloxicity. Bifenthrin (BF) is a new SP with a broad spectrum of insec-icidal and acaricidal activity used to control a wide range of insectests in a variety of applications. BF was detected in 95–100% ofediment samples, with a maximum concentration of 542 �g kg−1
or dry season samples and 122 �g kg−1 for wet season samplesn southern California (Bondarenko et al., 2007). BF is also a chiralnsecticide with one pair of enantiomers: 1S-cis-BF and 1R-cis-BF.n our previous studies, cis-BF showed enantioselective endocrineisruption and uptake, and the estrogenic potency of 1S-cis-BF wastronger than that of 1R-cis-BF (Wang et al., 2007). In recent years,F pesticide toxicity has been extensively investigated in insectsnd animal models, but there are few reports of cytotoxicity andenotoxicity in humans in the in vitro models. To our knowledgehere is no data about the enantioselective cytotoxicity of chiralesticides; so research in this area is very important to evaluatehe health risks of SPs.
The cytotoxicity assay is one kind of in vitro assay to test the toxi-ity of environmental chemicals using the effect on cell growth andpoptosis in target cells. The human amnion epithelial (FL) cell lineas originally derived from normal amnion (Fogh and Lund, 1957).
n recent years, the cell line has been used to assay the cytotox-city of some environmental chemicals, such as microcystins andributyltin (Liu and Xu, 2007; Fu et al., 2005).
In this study, the cytotoxicity of enantiomers of BF was assessedn FL cells in vitro using cell growth and apoptosis assays. Enantios-lectivity was further evaluated for BF in genetic damage due tohe induction of oxidative stress by BF. Given that most of the chiralPs are used in their racemic forms, knowledge of the cytotoxicityf individual enantiomers will be of great value for improving ournderstanding of their associated ecotoxicological and health risks.
. Materials and methods
.1. Chemicals
Analytical standard of racemic cis-BF (99.5%, 2-methylbiphenyl-3-ylmethyl-Z)-(1RS)-cis-3-(2-chloro-3,3,3-trifluoroprop-1-enyl)-2,2-dimethylcyclopropane
arboxylate, Fig. 1), 2′ ,7′-dichlorofluorescein diacetate (DCFH-DA), phalloidinfluorescein isothiocyanate labeled), and Annexin-V-FITC Apoptosis Detection Kitas from Sigma (St. Louis, MO, USA); thiazolyl blue (MTT) solution (5 mg mL−1hosphate-buffered saline (PBS)) was from Amresco (Solon, OH, USA). Minimalssential medium (MEM) was from Gibco (Paisley, UK); fetal bovine serum (FBS)as from Shijiqing Reagent Company, Hangzhou, China. All test compounds were
twOm
53 (2008) 89–96
issolved in ethanol, with the solvent content at <0.1% in the final volume. Otherhemicals or solvents used in this study were of cell culture, high-performanceiquid chromatography (HPLC), or analytical grade.
.2. Preparation of BF enantiomers
The enantiomers of cis-BF were resolved on a Jasco LC-2000 series HPLC systemJasco, Tokyo, Japan) equipped with a PU-2089 intelligent quaternary gradient pump,mobile-phase vacuum degasser, an AS-1559 autosampler with a 100 �L loop, a
O-2060 column temperature-control compartment, a UV-2075 ultraviolet–visibleetector, a variable wavelength CD-2095 circular dichroism (CD) detector, and an LC-et II/ADC data collector (Jasco, Tokyo, Japan). The Chiralcel OJ (250 mm × 4.6 mm
.d., cellulose tris(4-methyl benzoate) column (Daicel Chemical Industries, Tokyo,apan)] was used. The mobile phase was made of n-hexane fortified with ethanol.he detection wavelength of the CD detector was 230 nm. The resolved enantiomersere individually collected at the HPLC outlet, evaporated to dryness, and used in
he following bioassays. Concentrations of the enantiomers were determined on angilent 6890N gas chromatograph (GC) equipped with an electron capture detector
ECD), assuming the same response factor for enantiomers as for the racemate ofis-BF.
.3. Cell culture and treatments
FL cells (human amniotic cells, ATCC) were cultured in MEM supplemented with0% FBS (Gibco BRL), bovine serum albumin (0.2%), and PBS (10%, v/v). Cells were cul-ured at 37 ◦C with 5% CO2 in 25 cm2 flasks, trypsinized (0.25% sterile trypsin/0.02%DTA in PBS) and subcultured every 2–5 days. Cells were seeded in 96-well platesCostar, Cambridge, MA, USA) at a density of 2 × 105 cells mL−1 and allowed to adhereor 24 h prior to assaying.
.4. Assessment of cell viability
Cell viability was measured by quantitative colorimetric assay with MTT (3-4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide). The medium waseplaced with the experimental medium along with one of five concentrations ofhe test compound. After 24 h the medium was removed from the wells, and thia-olyl blue (MTT) solution was added. After 4 h, the solution was removed and 150 �LMSO per well was added. After 10 min shaking with a micro-mixer, the absorbanceas measured at 490 nm with a Bio-Rad Model 680 microplate reader (Bio-Rad Lab-ratories, Hercules, CA, USA). Results were calculated as the ratio of each exposureroup to the vehicle control (0.1% ethanol, onefold) and as the mean ± standard devi-tion (S.D.) of four independent measurements performed in three replicates. Theesults were analyzed using the t-test with Origin (OriginLab, Northampton, MA,SA).
.5. FITC-Annexin-V/propidium iodide staining assay
Cell apoptosis was assessed by flow cytometry according to the manufacturer’srotocol (Annexin-V-FITC Apoptosis Detection Kit, Sigma). Briefly, the FL cells at× 106 cells mL−1 were treated with BF or vehicle for 24 h at 37 ◦C in a 5% CO2 incu-ator, and then harvested and washed twice with PBS. Then the treated cells wereuspended in binding buffer at 3 × 106 cells mL−1, and supplemented with 5 �L ofITC-Annexin-V and 10 �L of PI, and incubated for 20 min at room temperature inhe dark. All experiments were repeated at least three times and flow cytometricnalysis was performed with a FAC Scan flow cytometer (Beckman Coulter EPICSL).
.6. Assay of the cellular contents of reactive oxygen species
The intracellular generation of reactive oxygen species (ROS) was measuredsing 2′ ,7′-dichlorofluorescein, which is oxidized to the fluorescent form 2′ ,7′-ichlorofluorescein by H2O2 and other ROS. DCFH-DA is known to enter the cell,
s hydrolyzed enzymatically to a non-fluorescent analog, 2′ ,7′-dichlorofluoresceneDCF-H), and is trapped in the cell. In the presence of certain ROS intermediates,uch as H2O2 or low-molecular-weight peroxyl radicals, DCF-H is oxidized to highlyuorescent 2′ ,7′-dichlorofluorescein (Wang and Joseph, 1999). Briefly, after treat-ent with BF or vehicle for 5 h, the treated cells were washed three times with
ce-cold PBS and then incubated with 10 �M DCFH-DA (100 mM in dimethyl sulfox-de) for 30 min at 37 ◦C. The cellular free radical content was assayed by measuring′ ,7′-dichlorofluorescein fluorescence by using a fluorescent spectrophotometerexcitation at 485 nm/emission at 535 nm, Tecan Infinite M200, Switzerland).
.7. Comet assay
The protocol used for the comet assay with cells from peripheral blood followedhe guidelines of Singh et al. (1988). To improve the repeatability of the test, slidesere washed three times (5 min each) with distilled water prior to DNA unwinding.bservations were made at a final magnification of 400× using an epifluorescenceicroscope (Zeiss) equipped with filters suitable for ethidium bromide. A number
H. Liu et al. / Toxicology 2
Fm
oea
2
pl3tPfm
2
2nCi11f
2
pvfa
3
3
etrcfbbtK
tp1noG
3
tuvdrJt(
wtp(ctuptwcs(
3
icals play an important role in cytotoxicity. FL cells were incubatedwith individual stereoisomers or racemate of BF for 4 h at con-centrations of 0, 25, 5, 10, and 20 mg L−1, followed by ROS leveldetermination. ROS levels were expressed by relative fluorescenceintensity. Fig. 5 shows that FL cells incubated with 1S-cis-BF or
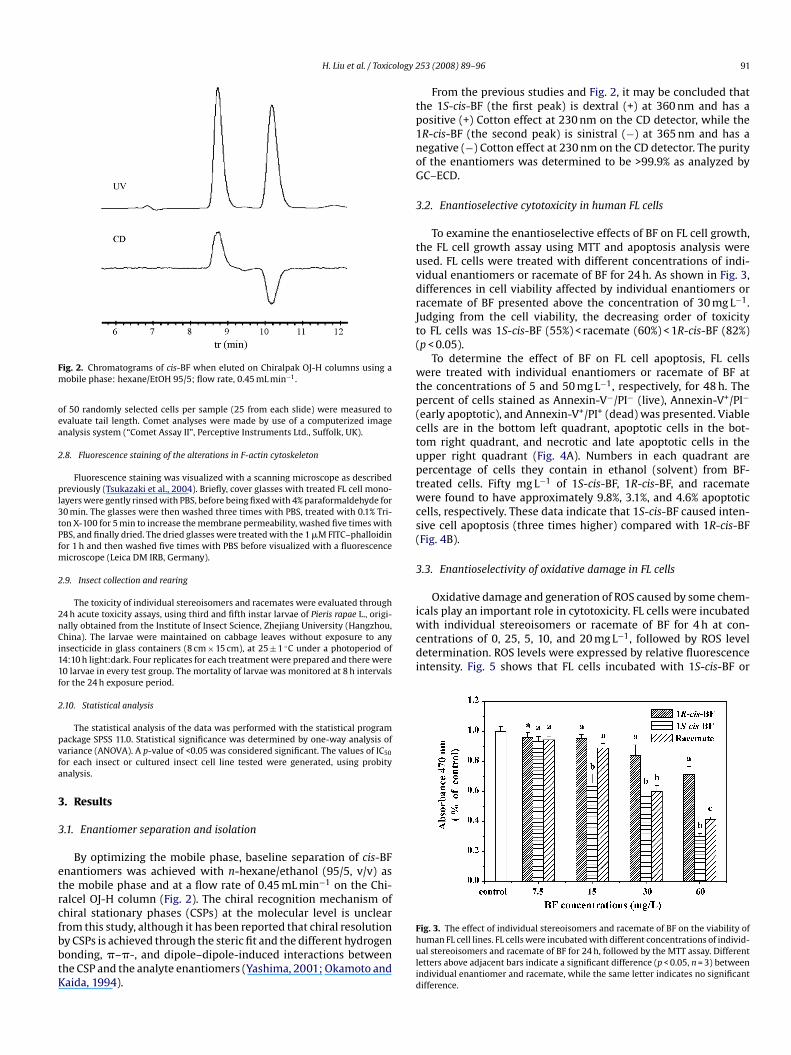
ig. 2. Chromatograms of cis-BF when eluted on Chiralpak OJ-H columns using aobile phase: hexane/EtOH 95/5; flow rate, 0.45 mL min−1.
f 50 randomly selected cells per sample (25 from each slide) were measured tovaluate tail length. Comet analyses were made by use of a computerized imagenalysis system (“Comet Assay II”, Perceptive Instruments Ltd., Suffolk, UK).
.8. Fluorescence staining of the alterations in F-actin cytoskeleton
Fluorescence staining was visualized with a scanning microscope as describedreviously (Tsukazaki et al., 2004). Briefly, cover glasses with treated FL cell mono-
ayers were gently rinsed with PBS, before being fixed with 4% paraformaldehyde for0 min. The glasses were then washed three times with PBS, treated with 0.1% Tri-on X-100 for 5 min to increase the membrane permeability, washed five times withBS, and finally dried. The dried glasses were treated with the 1 �M FITC–phalloidinor 1 h and then washed five times with PBS before visualized with a fluorescence
icroscope (Leica DM IRB, Germany).
.9. Insect collection and rearing
The toxicity of individual stereoisomers and racemates were evaluated through4 h acute toxicity assays, using third and fifth instar larvae of Pieris rapae L., origi-ally obtained from the Institute of Insect Science, Zhejiang University (Hangzhou,hina). The larvae were maintained on cabbage leaves without exposure to any
nsecticide in glass containers (8 cm × 15 cm), at 25 ± 1 ◦C under a photoperiod of4:10 h light:dark. Four replicates for each treatment were prepared and there were0 larvae in every test group. The mortality of larvae was monitored at 8 h intervalsor the 24 h exposure period.
.10. Statistical analysis
The statistical analysis of the data was performed with the statistical programackage SPSS 11.0. Statistical significance was determined by one-way analysis ofariance (ANOVA). A p-value of <0.05 was considered significant. The values of IC50
or each insect or cultured insect cell line tested were generated, using probitynalysis.
. Results
.1. Enantiomer separation and isolation
By optimizing the mobile phase, baseline separation of cis-BFnantiomers was achieved with n-hexane/ethanol (95/5, v/v) ashe mobile phase and at a flow rate of 0.45 mL min−1 on the Chi-alcel OJ-H column (Fig. 2). The chiral recognition mechanism ofhiral stationary phases (CSPs) at the molecular level is unclear
rom this study, although it has been reported that chiral resolutiony CSPs is achieved through the steric fit and the different hydrogenonding, �–�-, and dipole–dipole-induced interactions betweenhe CSP and the analyte enantiomers (Yashima, 2001; Okamoto andaida, 1994).Fhulid
53 (2008) 89–96 91
From the previous studies and Fig. 2, it may be concluded thathe 1S-cis-BF (the first peak) is dextral (+) at 360 nm and has aositive (+) Cotton effect at 230 nm on the CD detector, while theR-cis-BF (the second peak) is sinistral (−) at 365 nm and has aegative (−) Cotton effect at 230 nm on the CD detector. The purityf the enantiomers was determined to be >99.9% as analyzed byC–ECD.
.2. Enantioselective cytotoxicity in human FL cells
To examine the enantioselective effects of BF on FL cell growth,he FL cell growth assay using MTT and apoptosis analysis weresed. FL cells were treated with different concentrations of indi-idual enantiomers or racemate of BF for 24 h. As shown in Fig. 3,ifferences in cell viability affected by individual enantiomers oracemate of BF presented above the concentration of 30 mg L−1.udging from the cell viability, the decreasing order of toxicityo FL cells was 1S-cis-BF (55%) < racemate (60%) < 1R-cis-BF (82%)p < 0.05).
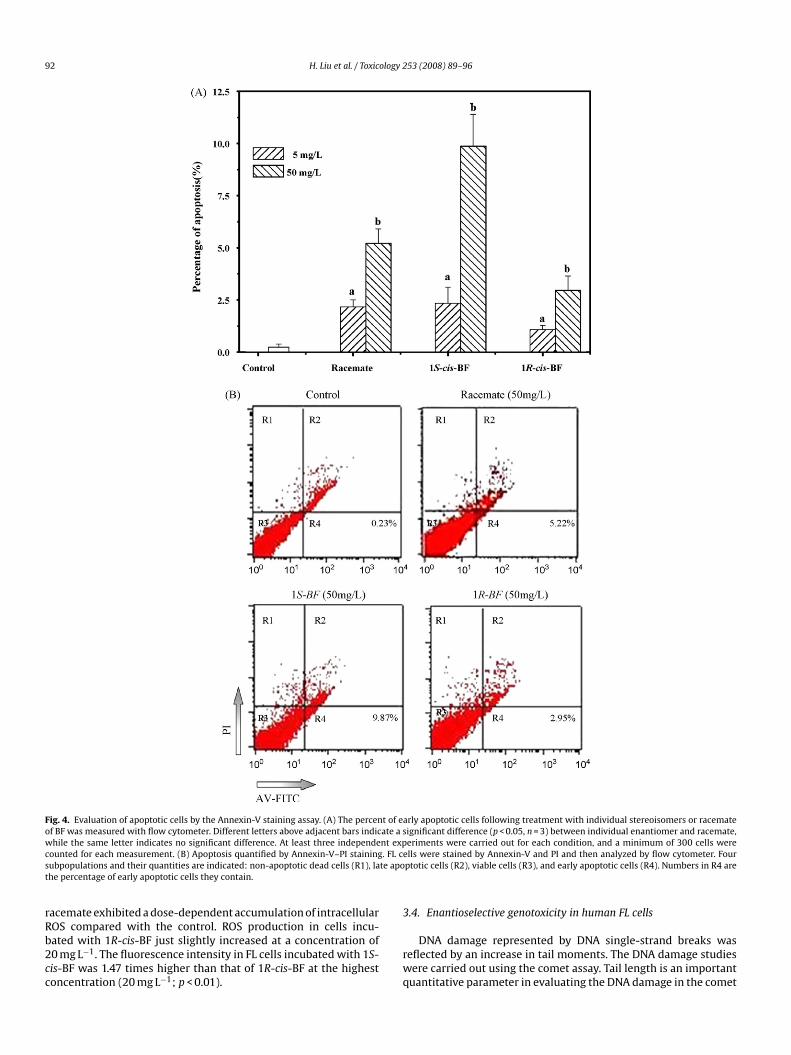
To determine the effect of BF on FL cell apoptosis, FL cellsere treated with individual enantiomers or racemate of BF at
he concentrations of 5 and 50 mg L−1, respectively, for 48 h. Theercent of cells stained as Annexin-V−/PI− (live), Annexin-V+/PI−
early apoptotic), and Annexin-V+/PI+ (dead) was presented. Viableells are in the bottom left quadrant, apoptotic cells in the bot-om right quadrant, and necrotic and late apoptotic cells in thepper right quadrant (Fig. 4A). Numbers in each quadrant areercentage of cells they contain in ethanol (solvent) from BF-reated cells. Fifty mg L−1 of 1S-cis-BF, 1R-cis-BF, and racemateere found to have approximately 9.8%, 3.1%, and 4.6% apoptotic
ells, respectively. These data indicate that 1S-cis-BF caused inten-ive cell apoptosis (three times higher) compared with 1R-cis-BFFig. 4B).
.3. Enantioselectivity of oxidative damage in FL cells
Oxidative damage and generation of ROS caused by some chem-
ig. 3. The effect of individual stereoisomers and racemate of BF on the viability ofuman FL cell lines. FL cells were incubated with different concentrations of individ-al stereoisomers and racemate of BF for 24 h, followed by the MTT assay. Different
etters above adjacent bars indicate a significant difference (p < 0.05, n = 3) betweenndividual enantiomer and racemate, while the same letter indicates no significantifference.
92 H. Liu et al. / Toxicology 253 (2008) 89–96
Fig. 4. Evaluation of apoptotic cells by the Annexin-V staining assay. (A) The percent of early apoptotic cells following treatment with individual stereoisomers or racemateof BF was measured with flow cytometer. Different letters above adjacent bars indicate a significant difference (p < 0.05, n = 3) between individual enantiomer and racemate,while the same letter indicates no significant difference. At least three independent experiments were carried out for each condition, and a minimum of 300 cells werec . FL ces te apot
rRb2cc
3
ounted for each measurement. (B) Apoptosis quantified by Annexin-V–PI stainingubpopulations and their quantities are indicated: non-apoptotic dead cells (R1), lahe percentage of early apoptotic cells they contain.
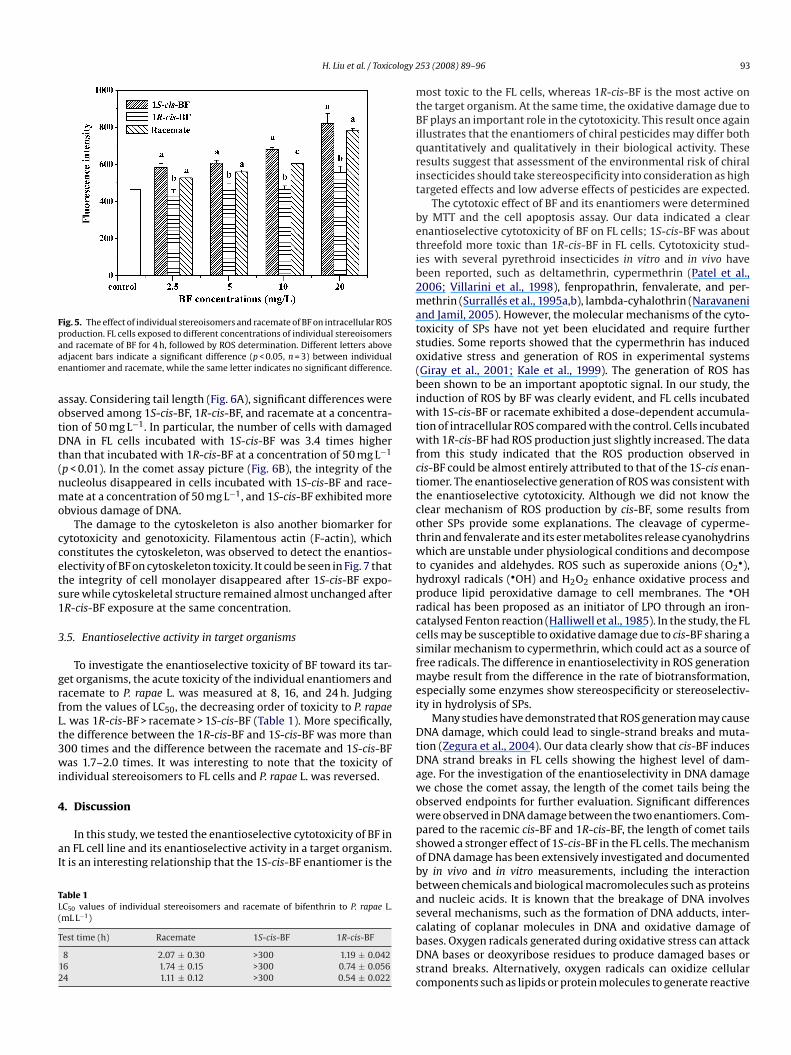
acemate exhibited a dose-dependent accumulation of intracellular
OS compared with the control. ROS production in cells incu-ated with 1R-cis-BF just slightly increased at a concentration of0 mg L−1. The fluorescence intensity in FL cells incubated with 1S-is-BF was 1.47 times higher than that of 1R-cis-BF at the highestoncentration (20 mg L−1; p < 0.01).rwq
lls were stained by Annexin-V and PI and then analyzed by flow cytometer. Fourptotic cells (R2), viable cells (R3), and early apoptotic cells (R4). Numbers in R4 are
.4. Enantioselective genotoxicity in human FL cells
DNA damage represented by DNA single-strand breaks waseflected by an increase in tail moments. The DNA damage studiesere carried out using the comet assay. Tail length is an important
uantitative parameter in evaluating the DNA damage in the comet
H. Liu et al. / Toxicology 2
Fig. 5. The effect of individual stereoisomers and racemate of BF on intracellular ROSpaae
aotDt(nmo
ccets1
3
grfLt3wi
4
aI
TL(
T
2
mtBiqrit
betib2matso(biwtwfcttcotwthprccsfmei
DtDawo
roduction. FL cells exposed to different concentrations of individual stereoisomersnd racemate of BF for 4 h, followed by ROS determination. Different letters abovedjacent bars indicate a significant difference (p < 0.05, n = 3) between individualnantiomer and racemate, while the same letter indicates no significant difference.
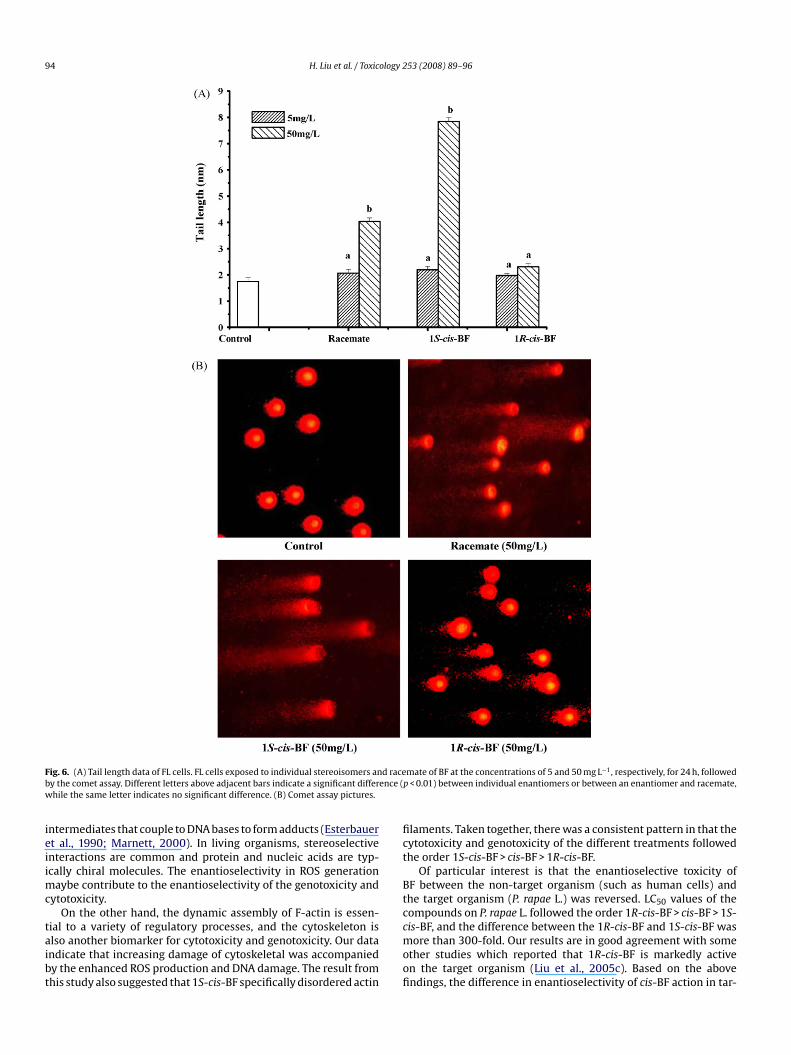
ssay. Considering tail length (Fig. 6A), significant differences werebserved among 1S-cis-BF, 1R-cis-BF, and racemate at a concentra-ion of 50 mg L−1. In particular, the number of cells with damagedNA in FL cells incubated with 1S-cis-BF was 3.4 times higher
han that incubated with 1R-cis-BF at a concentration of 50 mg L−1
p < 0.01). In the comet assay picture (Fig. 6B), the integrity of theucleolus disappeared in cells incubated with 1S-cis-BF and race-ate at a concentration of 50 mg L−1, and 1S-cis-BF exhibited more
bvious damage of DNA.The damage to the cytoskeleton is also another biomarker for
ytotoxicity and genotoxicity. Filamentous actin (F-actin), whichonstitutes the cytoskeleton, was observed to detect the enantios-lectivity of BF on cytoskeleton toxicity. It could be seen in Fig. 7 thathe integrity of cell monolayer disappeared after 1S-cis-BF expo-ure while cytoskeletal structure remained almost unchanged afterR-cis-BF exposure at the same concentration.
.5. Enantioselective activity in target organisms
To investigate the enantioselective toxicity of BF toward its tar-et organisms, the acute toxicity of the individual enantiomers andacemate to P. rapae L. was measured at 8, 16, and 24 h. Judgingrom the values of LC50, the decreasing order of toxicity to P. rapae. was 1R-cis-BF > racemate > 1S-cis-BF (Table 1). More specifically,he difference between the 1R-cis-BF and 1S-cis-BF was more than00 times and the difference between the racemate and 1S-cis-BFas 1.7–2.0 times. It was interesting to note that the toxicity of
ndividual stereoisomers to FL cells and P. rapae L. was reversed.
. Discussion
In this study, we tested the enantioselective cytotoxicity of BF inn FL cell line and its enantioselective activity in a target organism.t is an interesting relationship that the 1S-cis-BF enantiomer is the
able 1C50 values of individual stereoisomers and racemate of bifenthrin to P. rapae L.mL L−1)
est time (h) Racemate 1S-cis-BF 1R-cis-BF
8 2.07 ± 0.30 >300 1.19 ± 0.04216 1.74 ± 0.15 >300 0.74 ± 0.0564 1.11 ± 0.12 >300 0.54 ± 0.022
wpsobbascbDsc
53 (2008) 89–96 93
ost toxic to the FL cells, whereas 1R-cis-BF is the most active onhe target organism. At the same time, the oxidative damage due toF plays an important role in the cytotoxicity. This result once again
llustrates that the enantiomers of chiral pesticides may differ bothuantitatively and qualitatively in their biological activity. Theseesults suggest that assessment of the environmental risk of chiralnsecticides should take stereospecificity into consideration as highargeted effects and low adverse effects of pesticides are expected.
The cytotoxic effect of BF and its enantiomers were determinedy MTT and the cell apoptosis assay. Our data indicated a clearnantioselective cytotoxicity of BF on FL cells; 1S-cis-BF was abouthreefold more toxic than 1R-cis-BF in FL cells. Cytotoxicity stud-es with several pyrethroid insecticides in vitro and in vivo haveeen reported, such as deltamethrin, cypermethrin (Patel et al.,006; Villarini et al., 1998), fenpropathrin, fenvalerate, and per-ethrin (Surrallés et al., 1995a,b), lambda-cyhalothrin (Naravaneni
nd Jamil, 2005). However, the molecular mechanisms of the cyto-oxicity of SPs have not yet been elucidated and require furthertudies. Some reports showed that the cypermethrin has inducedxidative stress and generation of ROS in experimental systemsGiray et al., 2001; Kale et al., 1999). The generation of ROS haseen shown to be an important apoptotic signal. In our study, the
nduction of ROS by BF was clearly evident, and FL cells incubatedith 1S-cis-BF or racemate exhibited a dose-dependent accumula-
ion of intracellular ROS compared with the control. Cells incubatedith 1R-cis-BF had ROS production just slightly increased. The data
rom this study indicated that the ROS production observed inis-BF could be almost entirely attributed to that of the 1S-cis enan-iomer. The enantioselective generation of ROS was consistent withhe enantioselective cytotoxicity. Although we did not know thelear mechanism of ROS production by cis-BF, some results fromther SPs provide some explanations. The cleavage of cyperme-hrin and fenvalerate and its ester metabolites release cyanohydrinshich are unstable under physiological conditions and decompose
o cyanides and aldehydes. ROS such as superoxide anions (O2•),
ydroxyl radicals (•OH) and H2O2 enhance oxidative process androduce lipid peroxidative damage to cell membranes. The •OHadical has been proposed as an initiator of LPO through an iron-atalysed Fenton reaction (Halliwell et al., 1985). In the study, the FLells may be susceptible to oxidative damage due to cis-BF sharing aimilar mechanism to cypermethrin, which could act as a source ofree radicals. The difference in enantioselectivity in ROS generation
aybe result from the difference in the rate of biotransformation,specially some enzymes show stereospecificity or stereoselectiv-ty in hydrolysis of SPs.
Many studies have demonstrated that ROS generation may causeNA damage, which could lead to single-strand breaks and muta-
ion (Zegura et al., 2004). Our data clearly show that cis-BF inducesNA strand breaks in FL cells showing the highest level of dam-ge. For the investigation of the enantioselectivity in DNA damagee chose the comet assay, the length of the comet tails being the
bserved endpoints for further evaluation. Significant differencesere observed in DNA damage between the two enantiomers. Com-ared to the racemic cis-BF and 1R-cis-BF, the length of comet tailshowed a stronger effect of 1S-cis-BF in the FL cells. The mechanismf DNA damage has been extensively investigated and documentedy in vivo and in vitro measurements, including the interactionetween chemicals and biological macromolecules such as proteinsnd nucleic acids. It is known that the breakage of DNA involveseveral mechanisms, such as the formation of DNA adducts, inter-
alating of coplanar molecules in DNA and oxidative damage ofases. Oxygen radicals generated during oxidative stress can attackNA bases or deoxyribose residues to produce damaged bases ortrand breaks. Alternatively, oxygen radicals can oxidize cellularomponents such as lipids or protein molecules to generate reactive
94 H. Liu et al. / Toxicology 253 (2008) 89–96
F d raceb ence (w
ieiimc
taibt
fict
Btc
ig. 6. (A) Tail length data of FL cells. FL cells exposed to individual stereoisomers any the comet assay. Different letters above adjacent bars indicate a significant differhile the same letter indicates no significant difference. (B) Comet assay pictures.
ntermediates that couple to DNA bases to form adducts (Esterbauert al., 1990; Marnett, 2000). In living organisms, stereoselectiventeractions are common and protein and nucleic acids are typ-cally chiral molecules. The enantioselectivity in ROS generation
aybe contribute to the enantioselectivity of the genotoxicity andytotoxicity.
On the other hand, the dynamic assembly of F-actin is essen-
ial to a variety of regulatory processes, and the cytoskeleton islso another biomarker for cytotoxicity and genotoxicity. Our datandicate that increasing damage of cytoskeletal was accompaniedy the enhanced ROS production and DNA damage. The result fromhis study also suggested that 1S-cis-BF specifically disordered actincmoofi
mate of BF at the concentrations of 5 and 50 mg L−1, respectively, for 24 h, followedp < 0.01) between individual enantiomers or between an enantiomer and racemate,
laments. Taken together, there was a consistent pattern in that theytotoxicity and genotoxicity of the different treatments followedhe order 1S-cis-BF > cis-BF > 1R-cis-BF.
Of particular interest is that the enantioselective toxicity ofF between the non-target organism (such as human cells) andhe target organism (P. rapae L.) was reversed. LC50 values of theompounds on P. rapae L. followed the order 1R-cis-BF > cis-BF > 1S-
is-BF, and the difference between the 1R-cis-BF and 1S-cis-BF wasore than 300-fold. Our results are in good agreement with somether studies which reported that 1R-cis-BF is markedly activen the target organism (Liu et al., 2005c). Based on the abovendings, the difference in enantioselectivity of cis-BF action in tar-
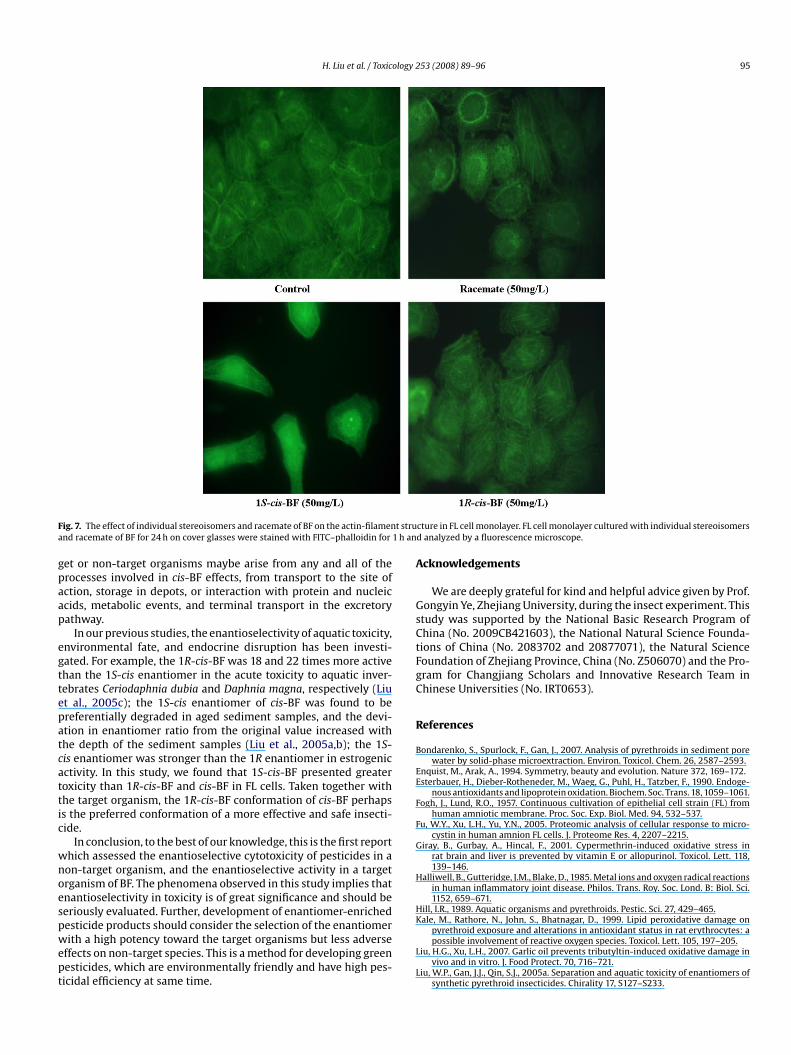
H. Liu et al. / Toxicology 253 (2008) 89–96 95
F t strua 1 h an
gpaap
egttepatcattic
wnoespwept
A
GsCtFgC
R
B
EE
F
F
G
H
HK
ig. 7. The effect of individual stereoisomers and racemate of BF on the actin-filamennd racemate of BF for 24 h on cover glasses were stained with FITC–phalloidin for
et or non-target organisms maybe arise from any and all of therocesses involved in cis-BF effects, from transport to the site ofction, storage in depots, or interaction with protein and nucleiccids, metabolic events, and terminal transport in the excretoryathway.
In our previous studies, the enantioselectivity of aquatic toxicity,nvironmental fate, and endocrine disruption has been investi-ated. For example, the 1R-cis-BF was 18 and 22 times more activehan the 1S-cis enantiomer in the acute toxicity to aquatic inver-ebrates Ceriodaphnia dubia and Daphnia magna, respectively (Liut al., 2005c); the 1S-cis enantiomer of cis-BF was found to bereferentially degraded in aged sediment samples, and the devi-tion in enantiomer ratio from the original value increased withhe depth of the sediment samples (Liu et al., 2005a,b); the 1S-is enantiomer was stronger than the 1R enantiomer in estrogenicctivity. In this study, we found that 1S-cis-BF presented greateroxicity than 1R-cis-BF and cis-BF in FL cells. Taken together withhe target organism, the 1R-cis-BF conformation of cis-BF perhapss the preferred conformation of a more effective and safe insecti-ide.
In conclusion, to the best of our knowledge, this is the first reporthich assessed the enantioselective cytotoxicity of pesticides in aon-target organism, and the enantioselective activity in a targetrganism of BF. The phenomena observed in this study implies thatnantioselectivity in toxicity is of great significance and should beeriously evaluated. Further, development of enantiomer-enriched
esticide products should consider the selection of the enantiomerith a high potency toward the target organisms but less adverseffects on non-target species. This is a method for developing greenesticides, which are environmentally friendly and have high pes-icidal efficiency at same time.
L
L
cture in FL cell monolayer. FL cell monolayer cultured with individual stereoisomersd analyzed by a fluorescence microscope.
cknowledgements
We are deeply grateful for kind and helpful advice given by Prof.ongyin Ye, Zhejiang University, during the insect experiment. Thistudy was supported by the National Basic Research Program ofhina (No. 2009CB421603), the National Natural Science Founda-ions of China (No. 2083702 and 20877071), the Natural Scienceoundation of Zhejiang Province, China (No. Z506070) and the Pro-ram for Changjiang Scholars and Innovative Research Team inhinese Universities (No. IRT0653).
eferences
ondarenko, S., Spurlock, F., Gan, J., 2007. Analysis of pyrethroids in sediment porewater by solid-phase microextraction. Environ. Toxicol. Chem. 26, 2587–2593.
nquist, M., Arak, A., 1994. Symmetry, beauty and evolution. Nature 372, 169–172.sterbauer, H., Dieber-Rotheneder, M., Waeg, G., Puhl, H., Tatzber, F., 1990. Endoge-
nous antioxidants and lipoprotein oxidation. Biochem. Soc. Trans. 18, 1059–1061.ogh, J., Lund, R.O., 1957. Continuous cultivation of epithelial cell strain (FL) from
human amniotic membrane. Proc. Soc. Exp. Biol. Med. 94, 532–537.u, W.Y., Xu, L.H., Yu, Y.N., 2005. Proteomic analysis of cellular response to micro-
cystin in human amnion FL cells. J. Proteome Res. 4, 2207–2215.iray, B., Gurbay, A., Hincal, F., 2001. Cypermethrin-induced oxidative stress in
rat brain and liver is prevented by vitamin E or allopurinol. Toxicol. Lett. 118,139–146.
alliwell, B., Gutteridge, J.M., Blake, D., 1985. Metal ions and oxygen radical reactionsin human inflammatory joint disease. Philos. Trans. Roy. Soc. Lond. B: Biol. Sci.1152, 659–671.
ill, I.R., 1989. Aquatic organisms and pyrethroids. Pestic. Sci. 27, 429–465.ale, M., Rathore, N., John, S., Bhatnagar, D., 1999. Lipid peroxidative damage on
pyrethroid exposure and alterations in antioxidant status in rat erythrocytes: apossible involvement of reactive oxygen species. Toxicol. Lett. 105, 197–205.
iu, H.G., Xu, L.H., 2007. Garlic oil prevents tributyltin-induced oxidative damage invivo and in vitro. J. Food Protect. 70, 716–721.
iu, W.P., Gan, J.J., Qin, S.J., 2005a. Separation and aquatic toxicity of enantiomers ofsynthetic pyrethroid insecticides. Chirality 17, S127–S233.

9 ology 2
L
L
L
M
M
M
N
O
O
P
S
S
S
T
V
W
W
WX
Yashima, E., 2001. Polysaccharide-based chiral stationary phases for high-performance liquid chromatographic enantioseparation. J. Chromatogr. 906,
6 H. Liu et al. / Toxic
iu, W.P., Gan, J.Y., Lee, S.J., Werner, I., 2005b. Isomer selectivity in aquatic toxicityand biodegradation of bifenthrin and permethrin. Environ. Toxicol. Chem. 24,1861–1866.
iu, W.P., Gan, J.Y., Schlenk, D., Jury, W.A., 2005c. Enantioselectivity in environmentalsafety of current chiral insecticides. Proc. Natl. Acad. Sci. U.S.A. 102, 701–706.
iu, W.P., 2006. Pesticide Environmental Chemistry. Chemical Industry Press, Beijing,China, pp. 341–343.
ajumder, S., Chakraborty, A.K., Mandal, T.K., Bhattacharya, A., Basak, D.K., 1994.Subacute toxicity of fenvalerate in broiler chicks: concentration, cytotoxicityand biochemical profiles. Indian J. Exp. Biol. 32, 752–756.
arnett, L.J., 2000. Structure, function and inhibition of cyclo-oxygenases. Ernst.Schering Res. Found Workshop 31, 65–83.
organ, M.K., Sheldon, L.S., Croghan, C.W., Jones, P.A., Chuang, J.C., Wilson, N.K., 2007.An observational study of 127 preschool children at their homes and daycarecenters in Ohio: environmental pathways to cis- and trans-permethrin exposure.Environ. Res. 104, 266–274.
aravaneni, R., Jamil, K., 2005. Evaluation of cytogenetic effects of lambda-cyhalothrin on human lymphocytes. J. Biochem. Mol. Toxicol. 19, 304–310.
kamoto, Y., Kaida, Y., 1994. Resolution by high-performance liquid chromatographyusing polysaccharide carbamates and benzoates as chiral stationary phases. J.Chromatogr. A 666, 403–419.
strea Jr., E.M., Villanueva-Uy, E., Bielawski, D.M., Posecion Jr., N.C., Corrion, M.L.,Jin, Y., Janisse, J.J., Ager, J.W., 2006. Maternal hair—an appropriate matrix fordetecting maternal exposure to pesticides during pregnancy. Environ. Res. 101,311–312.
atel, S., Pandey, A.K., Bajpayee, M., Parmar, D., Dhawan, A., 2006. Cypermethrin-induced DNA damage in organs and tissues of the mouse: evidence from thecomet assay. Mutat. Res. 607, 176–183.
ingh, N.P., McCoy, M.T., Tice, R.R., Schneider, E.L., 1988. A simple technique forquantitation of low levels of DNA damage in individual cells. Exp. Cell Res. 175,184–191.
YZ
53 (2008) 89–96
urrallés, J., Catalán, J., Creus, A., Norppa, H., Xamena, N., Marcos, R., 1995a. Micronu-clei induced by alachlor, mitomycin-C and vinblastine in human lymphocytes:presence of centromeres and kinetochores and influence of staining technique.Mutagenesis 10, 417–423.
urrallés, J., Xamena, N., Creus, A., Catalán, J., Norppa, H., Marcos, R., 1995b. Induc-tion of micronuclei by five pyrethroid insecticides in whole-blood and isolatedhuman lymphocyte cultures. Mutat. Res. 341, 169–184.
sukazaki, M., Satsu, H., Mori, A., Sugita-Konishi, Y., Shimizu, M., 2004. Effects of trib-utyltin on barrier functions in human intestinal Caco-2 cells. Biochem. Biophys.Res. Commun. 315, 991–997.
illarini, M., Moretti, M., Damiani, E., Greci, L., Santroni, A.M., Fedeli, D., Falcioni,G., 1998. Detection of DNA damage in stressed trout nucleated erythrocytesusing the comet assay: protection by nitroxide radicals. Free Radic. Biol. Med.24, 1310–1315.
ang, H., Joseph, J.A., 1999. Quantifying cellular oxidative stress by dichlorofluores-cein assay using microplate reader. Free Radic. Biol. Med. 27, 612–616.
ang, L.M., Liu, W.P., Yang, C.X., Pan, Z.Y., Gan, J.Y., Xu, C., Zhao, M.R., Schlenk, D., 2007.Enantioselectivity in estrogenic potential and uptake of bifenthrin. Environ. Sci.Technol. 41, 6124–6128.
illiams, A., 1996. Opportunities for chiral agrochemicals. Pestic. Sci. 46, 3–9.u, C., Wang, J.J., Liu, W.P., Sheng, D.Y., Tu, Y., Ma, Y., 2008. Separation and aquatic tox-
icity of enantiomers of the pyrethroid insecticide lambda-cyhalothrin. Environ.Toxicol. Chem. 27, 174–181.
105–125.oon, T.P., Jacobsen, E.N., 2003. Privileged chiral catalysts. Science 299, 1691–1693.egura, B., Lah, T.T., Filipic, M., 2004. The role of reactive oxygen species in
microcystin-LR-induced DNA damage. Toxicology 200, 59–68.
Copyright © 2022 FDOKUMEN




















