
Efficacy of inactivated vaccines against H5N1 avian influenza infection in ducks
6
Efficacy of inactivated vaccines against H5N1 avian influenza infection in ducks Deborah Middleton, John Bingham, Paul Selleck, Sue Lowther, Laurie Gleeson 1 , Philip Lehrbach 2 , Simon Robinson 2 , Jeff Rodenberg 3 , Mahesh Kumar 3 , Marion Andrew ⁎ Australian Animal Health Laboratory, CSIRO Livestock Industries, PO Bag 24, Geelong, Victoria, 3220 Australia Received 26 May 2006; returned to author for revision 6 July 2006; accepted 29 August 2006 Available online 9 October 2006 Abstract The current Asian H5N1 highly pathogenic avian influenza virus has spread over much of Asia and into Europe and Africa. As well as affecting village and commercial chicken operations in many South East Asian countries, it differs from past H5 avian influenza viruses in that it causes morbidity and mortalities in other domesticated birds, such as ducks and turkeys and in wild waterbirds. Effective vaccines that can prevent infection, as well as disease, and be used in a variety of avian species are needed for field use. In this report, a bivalent H5N9 + H7N1 oil emulsion vaccine is compared, in ducks, to a monovalent H5N3 oil emulsion vaccine that has been derived by reverse genetics with an H5 from A/chicken/ Vietnam/C58/04. While both vaccines protected against morbidity, the monovalent vaccine provided effective protection, with no evidence of shedding of the challenge virus and no serological response to the H5N1 challenge virus. © 2006 Elsevier Inc. All rights reserved. Keywords: Avian influenza; H5N1; Vaccine; Ducks Introduction The major control strategy for highly pathogenic avian influenza (HPAI) outbreaks in poultry has traditionally been one of eradication via movement restrictions and slaughter of affected and at-risk birds and improved biosecurity at poultry facilities to prevent initial infection. However, with the widespread presence of the current Asian H5N1 virus in village poultry, including ducks and turkeys, and in wildlife species, particularly migrating birds, alternate control strategies must be considered, with vaccines likely to be a key component. Currently available commercial vaccines for avian influenza are oil emulsion, killed virus vaccines. They have mostly been used to control endemic low pathogenic avian influenza (LPAI) in chickens and turkeys, or HPAI outbreaks in Pakistan and Mexico (Halvorson, 2002; Naeem, 1998; Swayne and Suarez, 2000). The European Union has approved use of inactivated oil emulsion vaccines for use in Italy, provided they allow for differentiation of infected versus vaccinated birds (Capua et al., 2002). Control of mildly pathogenic avian influenza in Mexico has been undertaken with both killed vaccines and recombinant fowlpox vaccines (Swayne and Suarez, 2000). While it has been demonstrated that an inactivated oil emulsion vaccine could interrupt transmission of H7N7 (Van der Goot et al., 2005) or the current H5N1 HPAI (Ellis et al., 2004), concerns still exist that these vaccines may not have 100% efficacy in the field and will not totally prevent shedding of virus. Additionally, their use does not allow the differentiation between infected and vaccinated birds, which interferes with Virology 359 (2007) 66 – 71 www.elsevier.com/locate/yviro ⁎ Corresponding author. Fax: +61 3 5227 5555. E-mail addresses: [email protected] (D. Middleton), [email protected] (J. Bingham), [email protected] (P. Selleck), [email protected] (S. Lowther), [email protected] (L. Gleeson), [email protected] (P. Lehrbach), [email protected] (S. Robinson), [email protected] (J. Rodenberg), [email protected] (M. Kumar), [email protected] (M. Andrew). 1 Current address: FAO Regional Office, Manivanh Mansion, 39 Phra Atit Rd, Banaglampu, Bangkok 10200, Thailand. 2 Fort Dodge Australia Pty. Limited PO Box 6024 Baulkham Hills BC NSW 2153, Australia. 3 Fort Dodge Animal Health, Division of Wyeth 800 Fifth Street NW Fort Dodge, Iowa 50501, USA. 0042-6822/$ - see front matter © 2006 Elsevier Inc. All rights reserved. doi:10.1016/j.virol.2006.08.046
Transcript of Efficacy of inactivated vaccines against H5N1 avian influenza infection in ducks

07) 66–71www.elsevier.com/locate/yviro
Virology 359 (20
Efficacy of inactivated vaccines against H5N1 avian influenzainfection in ducks
Deborah Middleton, John Bingham, Paul Selleck, Sue Lowther, Laurie Gleeson 1, Philip Lehrbach 2,Simon Robinson2, Jeff Rodenberg 3, Mahesh Kumar 3, Marion Andrew⁎
Australian Animal Health Laboratory, CSIRO Livestock Industries, PO Bag 24, Geelong, Victoria, 3220 Australia
Received 26 May 2006; returned to author for revision 6 July 2006; accepted 29 August 2006Available online 9 October 2006
Abstract
The current Asian H5N1 highly pathogenic avian influenza virus has spread over much of Asia and into Europe and Africa. As well asaffecting village and commercial chicken operations in many South East Asian countries, it differs from past H5 avian influenza viruses in that itcauses morbidity and mortalities in other domesticated birds, such as ducks and turkeys and in wild waterbirds. Effective vaccines that can preventinfection, as well as disease, and be used in a variety of avian species are needed for field use. In this report, a bivalent H5N9+H7N1 oil emulsionvaccine is compared, in ducks, to a monovalent H5N3 oil emulsion vaccine that has been derived by reverse genetics with an H5 from A/chicken/Vietnam/C58/04. While both vaccines protected against morbidity, the monovalent vaccine provided effective protection, with no evidence ofshedding of the challenge virus and no serological response to the H5N1 challenge virus.© 2006 Elsevier Inc. All rights reserved.
Keywords: Avian influenza; H5N1; Vaccine; Ducks
Introduction
The major control strategy for highly pathogenic avianinfluenza (HPAI) outbreaks in poultry has traditionally been oneof eradication via movement restrictions and slaughter ofaffected and at-risk birds and improved biosecurity at poultryfacilities to prevent initial infection. However, with thewidespread presence of the current Asian H5N1 virus in village
⁎ Corresponding author. Fax: +61 3 5227 5555.E-mail addresses: [email protected] (D. Middleton),
[email protected] (J. Bingham), [email protected] (P. Selleck),[email protected] (S. Lowther), [email protected] (L. Gleeson),[email protected] (P. Lehrbach), [email protected](S. Robinson), [email protected] (J. Rodenberg), [email protected](M. Kumar), [email protected] (M. Andrew).1 Current address: FAO Regional Office, Manivanh Mansion, 39 Phra Atit Rd,
Banaglampu, Bangkok 10200, Thailand.2 Fort Dodge Australia Pty. Limited PO Box 6024 Baulkham Hills BC NSW
2153, Australia.3 Fort Dodge Animal Health, Division of Wyeth 800 Fifth Street NW Fort
Dodge, Iowa 50501, USA.
0042-6822/$ - see front matter © 2006 Elsevier Inc. All rights reserved.doi:10.1016/j.virol.2006.08.046
poultry, including ducks and turkeys, and in wildlife species,particularly migrating birds, alternate control strategies must beconsidered, with vaccines likely to be a key component.
Currently available commercial vaccines for avian influenzaare oil emulsion, killed virus vaccines. They have mostly beenused to control endemic low pathogenic avian influenza (LPAI)in chickens and turkeys, or HPAI outbreaks in Pakistan andMexico (Halvorson, 2002; Naeem, 1998; Swayne and Suarez,2000). The European Union has approved use of inactivated oilemulsion vaccines for use in Italy, provided they allow fordifferentiation of infected versus vaccinated birds (Capua et al.,2002). Control of mildly pathogenic avian influenza in Mexicohas been undertaken with both killed vaccines and recombinantfowlpox vaccines (Swayne and Suarez, 2000).
While it has been demonstrated that an inactivated oilemulsion vaccine could interrupt transmission of H7N7 (Van derGoot et al., 2005) or the current H5N1 HPAI (Ellis et al., 2004),concerns still exist that these vaccines may not have 100%efficacy in the field and will not totally prevent shedding ofvirus. Additionally, their use does not allow the differentiationbetween infected and vaccinated birds, which interferes with

Table 1Titration of A/Muscovy duck/Vietnam/453/2004 in Pekin ducks
Dose(EID50)
First signof morbidityon day:
Time ofdeath oreuthanasia (day)
Virus isolation Antibodytiter a
Day 4b Day 7b
106.7 3 5 +/− nd/ndc nd3 – +/− −/− 1283 4 +/+ nd/nd nd3d 4 +/+ nd/nd nd3d 5 −/− nd/nd nd
105.7 3d 4 +/− nd/nd nd3 – −/− +/+ 1283d 4 +/− nd/nd nd3d 4 +/− nd/nd nd3 8 +/− −/− nd
104.7 9 9 −/+ +/+ nd9 9 −/− −/− nd8 9 −/− +/+ nd8 8 −/− +/− nd7 8 −/− +/+ nd
103.7 6 6 −/− nd/nd nd– – −/− +/− 810 10 −/− +/− nd– – −/+ −/+ 6410 10 −/− +/+ nd
102.7 – – −/− −/− <2– – −/− −/− <2– – −/− −/− <2– – −/− −/− <2– – −/− −/− <2
a Antibody titer on day 10 post challenge.b Positive or negative for virus in the pharynx/cloaca.c Not done.d Nervous clinical signs observed in addition to depression.
67D. Middleton et al. / Virology 359 (2007) 66–71
monitoring the disease status within flocks and regions; nor havethey been formulated or tested for vaccination efficacy in ducks.Vaccines based on the current Asian H5N1 strain and whichovercome these problems and for use in a variety of poultryspecies are needed urgently.
The Asian H5N1 virus cannot be grown to high titer in eggs,which is the traditional method of virus production for humanand avian influenza vaccines. Thus, alternatives to homologousvirus vaccines are being developed. Live vectored vaccines canprovide additional safety and the ability to differentiate betweeninfected and vaccinated birds. Fowlpox (Qiao et al., 2003),infectious laryngotracheitis virus (Luschow et al., 2001),Newcastle disease virus (Park et al., 2006) and adenovirus(Gao et al., 2006) vectors expressing the hemagglutinin, H5,have all been assessed and shown to have protective efficacy,with reduction, but not complete elimination, of virus shedding.Other influenza strains, particularly those that share the H5hemagglutinin type, but with a different neuraminidase, havebeen shown to have some efficacy in the field (Ellis et al., 2004),but the most promising approach is the use of reverse genetics tocreate an influenza virus that has the current H5 in a geneticbackground that allows growth to high titer in eggs, but has lowpathogenicity in either avian or mammalian species.
Reverse genetics has been used to create influenza virusreassortants with the hemagglutinin and neuraminidase genesfrom either the human H5N1 isolate, A/HK/491/97 (Subbarao etal., 2003), or the avian H5N1 isolate, A/Goose/Guangdong/96(Tian et al., 2005). Both these reassortants are apathogenic inchickens and the reassortant virus with H5 and N1 from A/Goose/Guangdong/96 has been tested as formalin inactivatedpreparation for protective efficacy against the parent HPAIH5N1 in specific pathogen-free (SPF) chickens, and in non-SPFgeese and ducks. Those studies demonstrated that the reassortantvaccine could prevent mortality and reduce shedding of thechallenge virus.
Inclusion of a different neuraminidase subtype in thereassortant vaccine allows differentiation of infected versusvaccinated birds. This principle has been demonstrated withreassortants using hemagglutinin genes from H5 and H7 LPAIviruses and the remaining genes, including the N1 fromA/WSN/33 (Lee et al., 2004). Oil emulsion vaccines with thesereassortants reduced replication of the parental H5 and H7LPAI strains in SPF chickens. A reassortant virus with H5 fromA/Goose/Hong Kong/437-4/99 and N3 from A/Duck/Germany/1215/73 has been constructed (Liu et al., 2003). Whenformulated as an oil emulsion, the vaccine was able to protectSPF chickens against mortality and markedly reduce virusshedding following challenge with HPAI H5N1 virus but did notprovide sterilizing immunity. In this study a similar reassortmentvirus where the hemagglutinin gene was derived from A/chicken/Vietnam/C58/04 (H5N1) virus was used in an inacti-vated oil emulsion formulation proprietary to Fort DodgeAnimal Health.
As ducks appear to be playing a critical role in theamplification and spread of H5N1 influenza virus in Asia,vaccines that are effective in ducks will provide an importanttool for control of the disease. The current study, reported here,
compares, in ducks, the efficacy of the H5N3 reassortant virusvaccine to that of a bivalent H5N9+H7N1 vaccine anddemonstrates that the reassortant virus provides superiorprotection, with no evidence of shedding of the challengevirus on days 4 and 7 post challenge and no antibody response tothe challenge virus, indicating little, if any, viral replication.
Results
Pathogenicity of A/Muscovy/Vietnam/453/2004 in ducks
Allantoic fluid from SPF eggs infected with the H5N1 strain,A/Muscovy duck/Vietnam/453/2004, was diluted in ten-foldsteps in PBS and used to infect groups of five ducks. The viruswas administered intraocularly, intranasally and orally, eachbird receiving from 102.7 to 106.7 EID50 of virus. Mortality,morbidity, virus re-isolation and antibody levels were measuredover the dose range (Table 1). Four out of five birds died in eachof the two highest dose groups (106.7 and105.7 EID50); all birdsdied at the 104.7 EID50 dose; three of five at the next lowest doseand all birds survived the lowest dose of 102.7 EID50. Thedevelopment of morbidity was delayed in the lower doses andthe time to death was also longer in the lower doses. Theobserved clinical signs of morbidity were reduced food intake,reduced water and grooming activity and nervous signs. Thenervous signs (head waving, high-stepping gait, falling and
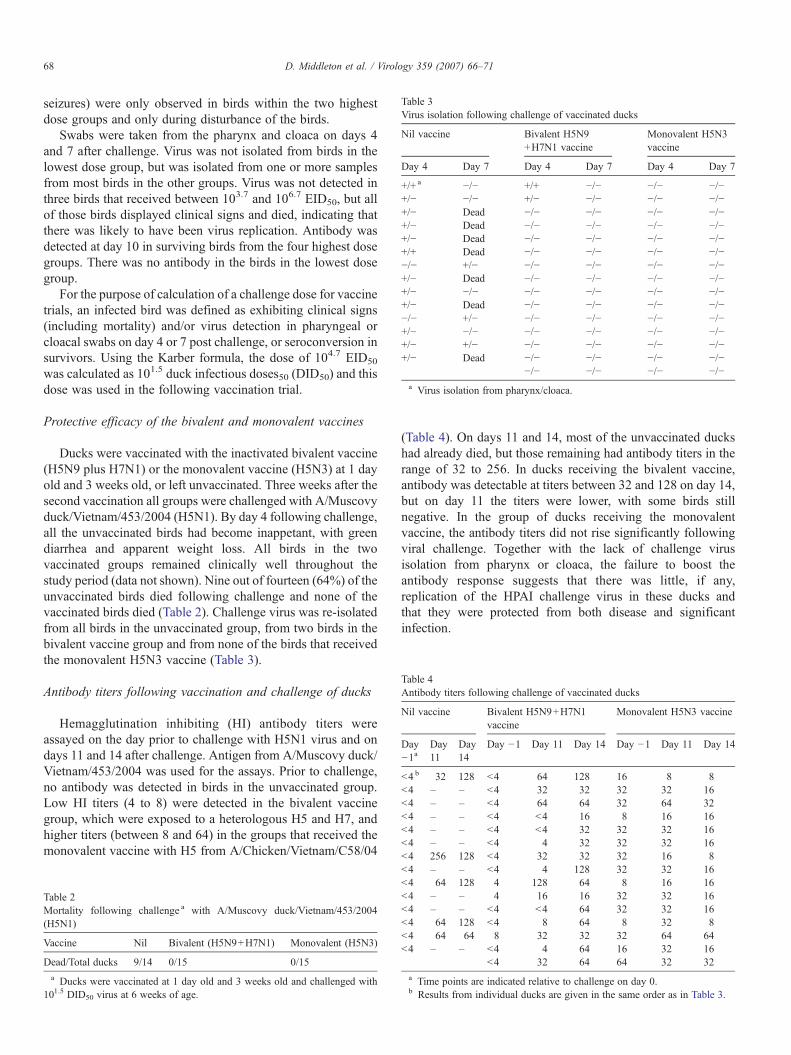
Table 3Virus isolation following challenge of vaccinated ducks
Nil vaccine Bivalent H5N9+H7N1 vaccine
Monovalent H5N3vaccine
Day 4 Day 7 Day 4 Day 7 Day 4 Day 7
+/+ a −/− +/+ −/− −/− −/−+/− −/− +/− −/− −/− −/−+/− Dead −/− −/− −/− −/−+/− Dead −/− −/− −/− −/−+/− Dead −/− −/− −/− −/−+/+ Dead −/− −/− −/− −/−−/− +/− −/− −/− −/− −/−+/− Dead −/− −/− −/− −/−+/− −/− −/− −/− −/− −/−+/− Dead −/− −/− −/− −/−−/− +/− −/− −/− −/− −/−+/− −/− −/− −/− −/− −/−+/− +/− −/− −/− −/− −/−+/− Dead −/− −/− −/− −/−
−/− −/− −/− −/−a Virus isolation from pharynx/cloaca.
Table 4Antibody titers following challenge of vaccinated ducks
Nil vaccine Bivalent H5N9+H7N1vaccine
Monovalent H5N3 vaccine
Day−1a
Day11
Day14
Day −1 Day 11 Day 14 Day −1 Day 11 Day 14
<4 b 32 128 <4 64 128 16 8 8<4 – – <4 32 32 32 32 16<4 – – <4 64 64 32 64 32<4 – – <4 <4 16 8 16 16<4 – – <4 <4 32 32 32 16<4 – – <4 4 32 32 32 16<4 256 128 <4 32 32 32 16 8
68 D. Middleton et al. / Virology 359 (2007) 66–71
seizures) were only observed in birds within the two highestdose groups and only during disturbance of the birds.
Swabs were taken from the pharynx and cloaca on days 4and 7 after challenge. Virus was not isolated from birds in thelowest dose group, but was isolated from one or more samplesfrom most birds in the other groups. Virus was not detected inthree birds that received between 103.7 and 106.7 EID50, but allof those birds displayed clinical signs and died, indicating thatthere was likely to have been virus replication. Antibody wasdetected at day 10 in surviving birds from the four highest dosegroups. There was no antibody in the birds in the lowest dosegroup.
For the purpose of calculation of a challenge dose for vaccinetrials, an infected bird was defined as exhibiting clinical signs(including mortality) and/or virus detection in pharyngeal orcloacal swabs on day 4 or 7 post challenge, or seroconversion insurvivors. Using the Karber formula, the dose of 104.7 EID50
was calculated as 101.5 duck infectious doses50 (DID50) and thisdose was used in the following vaccination trial.
Protective efficacy of the bivalent and monovalent vaccines
Ducks were vaccinated with the inactivated bivalent vaccine(H5N9 plus H7N1) or the monovalent vaccine (H5N3) at 1 dayold and 3 weeks old, or left unvaccinated. Three weeks after thesecond vaccination all groups were challenged with A/Muscovyduck/Vietnam/453/2004 (H5N1). By day 4 following challenge,all the unvaccinated birds had become inappetant, with greendiarrhea and apparent weight loss. All birds in the twovaccinated groups remained clinically well throughout thestudy period (data not shown). Nine out of fourteen (64%) of theunvaccinated birds died following challenge and none of thevaccinated birds died (Table 2). Challenge virus was re-isolatedfrom all birds in the unvaccinated group, from two birds in thebivalent vaccine group and from none of the birds that receivedthe monovalent H5N3 vaccine (Table 3).
Antibody titers following vaccination and challenge of ducks
Hemagglutination inhibiting (HI) antibody titers wereassayed on the day prior to challenge with H5N1 virus and ondays 11 and 14 after challenge. Antigen from A/Muscovy duck/Vietnam/453/2004 was used for the assays. Prior to challenge,no antibody was detected in birds in the unvaccinated group.Low HI titers (4 to 8) were detected in the bivalent vaccinegroup, which were exposed to a heterologous H5 and H7, andhigher titers (between 8 and 64) in the groups that received themonovalent vaccine with H5 from A/Chicken/Vietnam/C58/04
Table 2Mortality following challenge a with A/Muscovy duck/Vietnam/453/2004(H5N1)
Vaccine Nil Bivalent (H5N9+H7N1) Monovalent (H5N3)
Dead/Total ducks 9/14 0/15 0/15
a Ducks were vaccinated at 1 day old and 3 weeks old and challenged with101.5 DID50 virus at 6 weeks of age.
(Table 4). On days 11 and 14, most of the unvaccinated duckshad already died, but those remaining had antibody titers in therange of 32 to 256. In ducks receiving the bivalent vaccine,antibody was detectable at titers between 32 and 128 on day 14,but on day 11 the titers were lower, with some birds stillnegative. In the group of ducks receiving the monovalentvaccine, the antibody titers did not rise significantly followingviral challenge. Together with the lack of challenge virusisolation from pharynx or cloaca, the failure to boost theantibody response suggests that there was little, if any,replication of the HPAI challenge virus in these ducks andthat they were protected from both disease and significantinfection.
<4 – – <4 4 128 32 32 16<4 64 128 4 128 64 8 16 16<4 – – 4 16 16 32 32 16<4 – – <4 <4 64 32 32 16<4 64 128 <4 8 64 8 32 8<4 64 64 8 32 32 32 64 64<4 – – <4 4 64 16 32 16
<4 32 64 64 32 32
a Time points are indicated relative to challenge on day 0.b Results from individual ducks are given in the same order as in Table 3.

69D. Middleton et al. / Virology 359 (2007) 66–71
Discussion
As different isolates of the Asian H5N1 HPAI viruses havebeen shown to vary in their virulence for different avian andmammalian species (Govorkova et al., 2005; Nguyen et al.,2005), it was important to titrate A/Muscovy duck/Vietnam/453/2004 in the population of ducks that were to be used forthe vaccination trial. Compared to the higher dose groups, thegroups that received the intermediate doses of virus (104.7 and103.7 EID50) display a delayed onset of morbidity andmortality, but, interestingly, there were no observations ofnervous signs in these two intermediate dose groups. Thissuggests that the spread of virus within the duck and thepathogenic process is qualitatively different depending on thesize of the infecting dose. However, with the limited numberof birds and of samples taken for virus re-isolation in thisexperiment, correlations between the sites or timing of virusre-isolation and birds with or without nervous signs were notobserved.
The low or undetectable antibody titers in ducks vaccinatedwith the bivalent vaccine, which had H5 from strain A/chicken/Italy/22A/98 demonstrates the importance of matching thevaccine hemagglutinin antigen to the influenza strain currentlycirculating and is the reason why human influenza vaccines arereformulated each year. Despite the failure to stimulatesignificant HI titers, the bivalent vaccine did offer protectionas no ducks showed morbidity or mortality. However, the rise inantibody titers following challenge is indicative of some virusreplication within these birds.
The hemagglutinin used in the monovalent vaccine wasderived by reverse genetics from A/chicken/Vietnam/C58/04, avirus closely related to the challenge strain. During construc-tion, the hemagglutinin was modified by deletion of thenucleotides encoding four amino acids at the HA1–HA2cleavage site to create a cleavage site that is similar to that ofnon pathogenic viruses (Hatta et al., 2001). This modificationappears not to have affected the immunogenicity of the H5.
Clinical signs and virus isolation occurred earlier in thevaccination trial than they had in the corresponding group in thetitration experiment. As the ducks were sourced from the samesupplier and were challenged at the same age, there is no obviousreason for this variation in the timecourse of infection. However,given the earlier disease progression in the unvaccinated ducks,it may have been possible that virus could have been detectedbefore day 4 in the vaccinated birds. However, the failure of thechallenge H5N1 virus to stimulate a secondary antibodyresponse in the ducks vaccinated with the closely related H5 inthe monovalent vaccine is very strong evidence for minimal, tono, replication of the challenge virus in those ducks.
In both the titration study and vaccine trial, virus re-isolationpost challenge occurred more frequently from the pharynx ofbirds than from the cloaca. However, in some birds virus wasisolated from the cloaca and not the pharynx on particular days.The optimum time of virus re-isolation was largely, but notcompletely, dependent on the challenge dose, and at all doses,sampling at a single time point would have given an incompletepicture, demonstrating the danger of assuming that there is no
viral shedding after obtaining a negative result at a single timepoint.
There have been a number of published trials reporting onvaccines that provide protective immunity against HPAI.However, many of those trials have been conducted in SPFpoultry and past experience has shown that vaccine immuno-genicity, and therefore efficacy, is often lower in conventionalanimals than in SPF animals (Heine et al., 1997). While SPFanimals are valuable in eliminating variables during earlystudies, it is important to test vaccines intended for eventualfield use, in animals with similar genetics and rearingconditions. Tian et al. (2005) have also demonstrated that avaccine, produced by reverse genetics to express an H5 antigenfrom an Asian H5N1 virus, can protect field ducks and geesefrom mortality and morbidity, with very reduced shedding of thechallenge virus. However, they did not test whether thechallenge virus boosted the antibody response. When aNewcastle disease virus expressing H7 was used to vaccinatenon-SPF chickens, 90% of the birds survived a lethal challengewith HPAI H7N7 virus, but the challenge virus stimulated hightiters of antibody in the surviving birds, suggesting that therehad been significant viral replication, if not shedding (Park etal., 2006). The apparently less solid protection with theNewcastle disease vectored vaccine raises the possibility thatother influenza virus gene products have contributed toprotection in the work described here, but differences in thechallenge virus may also have contributed to the differentoutcome.
The results presented here demonstrate that a killedmonovalent H5N3 vaccine is able to provide solid protectionin ducks against a challenge with an HPAI virus with a highlyrelated H5. The absence of any virus re-isolation, tested overseveral days, plus the failure to boost the antibody response isstrong evidence that the birds were protected against bothinfection and disease. This reassortant vaccine virus can providea safe and effective tool for the control of the Asian H5N1influenza virus in village and commercial ducks, as it willprevent virus shedding and also will permit the differentiationbetween vaccinated and infected birds.
Materials and methods
Viruses and vaccines
A/Muscovy duck/Vietnam/453/2004 H5N1 was obtainedfrom the Regional Animal Health Centre, Ho Chi Minh City,Vietnam. This isolate was obtained from diseased ducks andwas passaged twice in SPF chicken eggs (Charles RiverLaboratories, Australia). Prior to use, the virus was titrated inSPF chicken eggs which were incubated at 37 °C for up to 5days before assessment of allantoic fluid for hemagglutinationactivity. The titer was calculated as 108 EID50/ml.
The bivalent vaccine, Poulvac i-AI H5N9, H7N1 wassupplied by Fort dodge Australia Pty. Limited 1 MaitlandPlace, Baulkham Hills NSW 2153 Australia, as an inactivatedoil emulsion vaccine formulated with inactivated allantoicfluids infected with avian influenza virus type A subtypes H5N9

70 D. Middleton et al. / Virology 359 (2007) 66–71
(strain A/chicken/italy/22A/98) and H7N1 (strain A/chicken/Italy/1067/H7N1/1999). The monovalent vaccine, Poulvac i-AIH5N3 RG was derived by reverse genetics by a method similarto that described by Liu et al. (2003). Briefly, the H5hemagglutinin from A/chicken/Vietnam/C58/04 was modifiedby deletion of twelve nucleotides at the cleavage site betweenHA1 and HA2. This gene was combined with N3 from A/Duck/Germany/1215/73 and the internal genes from A/PR/8/34. Bothvaccines were formulated as water in oil emulsions.
Animals and accommodation
One-day- or six-week-old Pekin ducks of mixed sex werepurchased from a commercial supplier and housed on tiled floorswith access towood shavings andwater for bathing. Theywere fedcommercial rations and also had access to drinking water. Anumbered leg band identified individual ducks. All animalexperimentation was performed with the approval of aninstitutional Animal Ethics Committee. Prior to inoculation ofvirus, a blood samplewas taken from the jugular vein of each duck.
Titration of virus in ducks
Infectious allantoic fluid from eggs inoculated with A/Muscovy duck/Vietnam/453/2004 was diluted in ten-fold stepsin sterile phosphate-buffered saline (PBS). Dilutions from 10−1
to 10− 5 were inoculated intraocularly (50 μl per eye),intranasally (200 μl) and orally (200 μl) into each of fiveducks per dilution, giving a total dose of 106.7 EID50 per duck inthe highest dose group. Following inoculation of virus dilutionsinto ducks, samples were back titrated into embryonated eggs toconfirm their titer.
Ducks were monitored twice daily for clinical signs,particularly depression, inappetence and nervous signs. Duckswere killed with intravenous barbiturate on welfare grounds ifthey demonstrated moderate clinical signs that confirmedinfection (reduced interaction with pen mates, slight head tiltor twitching). Pharyngeal and cloacal swabs (EUROTUBOcotton swabs) were taken from all surviving ducks on days 4and stored at −80 °C in 2 ml PBS+ antibiotics. Blood sampleswere taken from the wing vein of each surviving duck at day 10after challenge. Serum was stored at −20 °C.
All median duck infectious doses (DID50) were calculatedusing the Karber formula (Lennette, 1995).
Vaccination trial
The optimal number of ducks per group for the challengewas assessed to be fifteen, using data from previous work withthe challenge virus, but groups of twenty birds were vaccinatedto account for the normal mortality rate observed in youngbirds. The three randomly assigned groups of day-old Pekinducks of mixed sexes were each housed in a microbiologicallycontained and separate room, under the same conditions usedfor the virus titration. Prior to vaccination, ten randomlyselected ducks were screened by c-ELISA which detectsantibodies to all avian influenza A viruses and all ducks were
found to be negative. One group of ducks was inoculated withPBS; one group was vaccinated with the Poulvac i-AI H5N9,H7N1 vaccine and the third group with the Poulvac i-AI H5N3vaccine. As recommended by the manufacturer, the vaccine wasadministered subcutaneously in 200 μl high on the neck. Thevaccinated groups were revaccinated at 3 weeks of age, using500 μl of vaccine delivered by the same route.
Three weeks later, at the time of challenge, only 14unvaccinated birds remained alive, and the other two groupswere reduced to fifteen, in accordance with welfare require-ments. Leg bands applied to each bird. Using a 1 ml graduatedsyringe, all ducks were inoculated with the challenge virus, A/Muscovy duck/Vietnam/453/2004. A dose of 101.5 DID50 wasgiven to each duck, in a total volume of 500 μl, with 200 μlbeing delivered to both nares, 200 μl given orally and 100 μlbeing delivered to the open eyes.
Ducks were observed twice daily for signs of morbidity orfor mortality. Each duck was bled from the wing veinimmediately prior to challenge and on days 11 and 14 afterchallenge to obtain serum for antibody assays. Pharyngeal andcloacal swabs were taken on days 4 and 7 for virus re-isolation.
Virus re-isolation
Virus isolations were performed according to the OfficeInternational des Epizooties (OIE) Manual (Anon., 2004).Briefly, swabs were placed in PBS and an aliquot titrated intoembryonated eggs, which were incubated for 5 days at 37 °Cbefore assay of allantoic fluid for hemagglutinating activity.
Antibody assays
Hemagglutination inhibition antibody titers in serum wereassayed according to the OIE Manual, using A/Muscovy duck/Vietnam/453/2004 as antigen (Alexander, 2004). This assaywas used to detect antibody specific for the H5 hemagglutinin,before and after challenge.
For the c-ELISA, A/PR/8/34 H1N1 influenza virus grown ineggs and diluted in a carbonate buffer was used as antigen. Thetest sera were diluted ten-fold in PBS with 0.05% Tween 20 and1% skim milk powder and added to the antigen-coated platestogether with a monoclonal antibody specific for influenzanucleoprotein. After incubation and then washing, the boundmonoclonal antibody was detected with sheep anti-murineimmunoglobulin (Silenus Cat. DAH) and TMB substrate(Sigma Cat. T-2885). The reaction was stopped with sulphuricacid and the color read at 450 nm. Percent inhibition of bindingof the monoclonal antibody was calculated and less than 40%inhibition was interpreted as negative.
Acknowledgments
Funding for this study was provided by Fort Dodge AnimalHealth. The work was conducted in the laboratories of CSIROLivestock Industries at the Australian Animal Health Labora-tory, Geelong and there are no conflicts of interest issues relatedto this project.

71D. Middleton et al. / Virology 359 (2007) 66–71
References
Alexander, D.J., 2004. Highly pathogenic avian influenza, Manual of DiagnosticTests and Vaccines for Terrestrial Animals, 5th ed. Office Internationale desEpizooties, Paris, pp. 258–269.
Anon., 2004. Highly pathogenic avian influenza, In: Vallat, B., Edwards, S.(Eds.), OIE Manual of Diagnostic Tests and Vaccines for TerrestrialAnimals, 5th ed. World Organisation for Animal Health, Paris, pp. 258–269.
Capua, I., Terregino, C., Cattoli, G., Mutinelli, F., Rodriguez, J.F., 2002.Development of a DIVA (differentiating infected from vaccinated animals)strategy using a vaccine containing heterologous neuraminidase for thecontrol of avian influenza. Avian Pathol. 32, 47–55, doi:10.1080/0307945021000070714.
Ellis, T.M., Leung, C.Y.H.C., Chow, M.K.W., Bissett, L.A., Wong, W., Guan,Y., Peiris, J.S.M., 2004. Vaccination of chickens against H5N1 avianinfluenza in the face of an outbreak interrupts virus transmission. AvianPathol. 33, 405–412, doi:10.1080/03079450410001724012.
Gao, G., Soloff, A.C., Lu, X., Montecalvo, A., Nguyen, D.C., Matsuoka, Y.,Robbins, P.D., Swayne, D.E., Donis, R.O., Katz, J.M., Barratt-Boyes, S.M.,Gambotto, A., 2006. Protection of mice and poultry from lethal H5N1 avianinfluenza virus through adenovirus-based immunization. J. Virol. 80,1959–1964.
Govorkova, E.A., Rehg, J.E., Krauss, S., Yen, H.-L., Guan, Y., Peiris, M.,Nguyen, T.D., Hanh, T.H., Puthavathana, P., Long, H.T., Buranathai, C., Lim,W., Webster, R.G., Hoffmann, E., 2005. Lethality to ferrets of H5N1influenza viruses isolated from humans and poultry in 2004. J. Virol. 79,2191–2198.
Halvorson, D.A., 2002. The control of H5 or H7 mildly pathogenic avianinfluenza: a role for inactivated vaccine. Avian Pathol. 31, 5–12,doi:10.1080/03079450120106570.
Hatta, M., Gao, P., Halfmann, P., Kawaoka, Y., 2001. Molecular basis for highvirulence of Hong Kong H5N1 influenza A viruses. Science 293,1840–1842.
Heine, H.G., Foord, A.J., Young, P.L., Hooper, P.T., Lehrbach, P.R., Boyle, D.B.,1997. Recombinant fowlpox virus vaccines against Australian virulentMarek's disease virus: gene sequence analysis and comparison of vaccineefficacy in specific pathogen free and production chickens. Virus Res. 50,23–33.
Lee, C.-W., Senne, D.A., Suarez, D.L., 2004. Generation of reassortant influenzavaccines by reverse genetics that allows utilization of a DIVA (differentiatinginfected from vaccinated animals) strategy for the control of avian influenza.Vaccine 22, 3175–3181, doi:10.1016/j.vaccine.2004.01.055.
Lennette, D.A., 1995. General principles for laboratory diagnosis of viral,rickettsial, and chlamydial infections, In: Lennette, E.H., Lennette, D.A.,
Lennette, E.T. (Eds.), Diagnostic Procedures for Viral, Rickettsial, andChlamydial Infections, 7th ed. American Public Health Assoc., Washington,pp. 3–25.
Liu, M., Wood, J.M., Ellis, T., Krauss, S., Seiler, P., Johnson, C., Hoffmann, E.,Humberd, J., Hulse, D., Zhang, Y., Webster, R.G., Perez, D.R., 2003.Preparation of a standardized, efficacious agricultural H5N3 vaccine byreverse genetics. Virology 314, 580–590, doi:10.1016/S0042-6822(03)00458-6.
Luschow, D., Werner, O., Mettenleiter, T.C., Fuchs, W., 2001. Protection ofchickens from lethal avian influenza A virus infection by live-virusvaccination with infectious laryngotracheitis virus recombinants expressingthe hemagglutinin (h5) gene. Vaccine 19, 4249–4259.
Naeem, K., 1998. The avian influenza H7N3 outbreak in south central Asia.Proceedings of the 4th International Symposium on Avian Influenza,Athens, Georgia, USA, pp. 31–35.
Nguyen, D.C., Uyeki, T.M., Jadhao, S., Maines, T., Shaw, M., Matsuoka, Y.,Smith, C., Rowe, T., Lu, X., Hall, H., Xu, X., Balish, A., Klimov, A.,Tumpey, T.M., Swayne, D.E., Huynh, L.P.T., Hghiem, H.K., Nguyen,H.H.T., Hoang, L.T., Cox, N.J., Katz, J.M., 2005. Isolation and charac-terisation of avian influenza viruses, including highly pathogenic H5N1,from poultry in live bird markets in Hanoi, Vietnam, in 2001. J. Virol. 79,4201–4212.
Park, M.-S., Steel, J., Garcia-Sastre, A., Swayne, D., Palese, P., 2006.Engineered viral vaccine constructs with dual specificity: avian influenzaand Newcastle disease. Proc. Natl. Acad. Sci. 103, 8203–8208, doi:10.1073/pnas.0602566103.
Qiao, C.-L., Yu, K.-Z., Jiang, Y.-P., Jia, Y.-Q., Tian, G.-B., Liu, M., Deng,G.-H., Wang, X.-R., Meng, Q.-W., Tang, X.-Y., 2003. Protection ofchickens against highly lethal H5N1 and H7H1 avian influenza viruseswith a recombinant fowlpox virus co-expressing H5 haemagglutininand N1 neuraminidase genes. Avian Pathol. 32, 25–31, doi:10.1080/0307945021000070688.
Subbarao, K., Chen, H., Swayne, D., Mingay, L., Fodor, E., Brownlee, G., Xu,X., Lu, X., Katz, J., Cox, N., Matsuoka, Y., 2003. Evaluation of a geneticallymodified reassortant H5N1 influenza Avirus vaccine candidate generated byplasmid-based reverse genetics. Virology 305, 192–200.
Swayne, D.E., Suarez, D.L., 2000. Highly pathogenic avian influenza. Rev. Sci.Tech.-Off. Int. Epizoot. 19, 463–482.
Tian, G., Zhang, S., Li, Y., Bu, Z., Liu, P., Zhou, J., Li, C., Shi, J., Yu, K., Chen,H., 2005. Virology 341, 153–162, doi:10.1016/j.virol.2005.07.011.
Van der Goot, J.A., Koch, G., de Jong, M.C.M., van Boven, M., 2005.Quantification of the effect of vaccination on transmission of avian influenza(H7N7) in chickens. Proc. Natl. Acad. Sci. 102, 18141–18146, doi:10.1073/pnas.0505098102.




















