
Utilization of Agricultural Wastes in the Manufacture of ...
Upload
independentCategory
view
1download
0
MARINE
Marine Environmental Research 60 (2005) 317–337
www.elsevier.com/locate/marenvrev
ENVIRONMENTAL
RESEARCH
Effects of mining wastes on a seagrass ecosystem:metal accumulation and bioavailability,
seagrass dynamics and associatedcommunity structure
Lazaro Marın-Guirao a,*, Arnaldo Marın Atucha a,Javier Lloret Barba a, Emma Martınez Lopez b,
Antonio J. Garcıa Fernandez b
a Departamento de Ecologıa e Hidrologıa, Facultad de Biologıa,
Universidad de Murcia, 30100-Murcia, Spainb Departamento de Toxicologıa, Facultad de Veterinaria,
Universidad de Murcia, 30100-Murcia, Spain
Received 5 March 2004; received in revised form 6 November 2004; accepted 12 November 2004
Abstract
Two different Cymodocea nodosa (Ucria) Ascherson beds growing in mining-contaminated
sediments were compared with two reference beds in the Mar Menor coastal lagoon. The accu-
mulation of Zn, Pb and Cd in different fractions of the plant, the sediment parameters that reg-
ulate the availability of metals, the seabed structure and dynamics of each seagrass bed and its
associated macroinvertebrate community were studied. C. nodosa accumulates metals from the
sediments and reflects their bioavailability for this seagrass. At each station, the metal content
of the rhizomes was lower than that of leaves and roots. The concentration of acid-volatile sul-
fides does not seem to influence the availability of metals to the seagrass, possibly due to oxygen
transport to underground tissues. The highest metal concentration in all the contaminated sta-
tions was found in the leaf-biofilm, due to the formation of complexes between metals and the
extracellular polymeric substances that form the biofilm. All the seagrass beds were seen to be
0141-1136/$ - see front matter � 2004 Elsevier Ltd. All rights reserved.
doi:10.1016/j.marenvres.2004.11.002
* Corresponding author. Tel.: +34 968364977; fax: +34 968363963.
E-mail address: [email protected] (L. Marın-Guirao).

318 L. Marın-Guirao et al. / Marine Environmental Research 60 (2005) 317–337
undergoing expansion, those growing in contaminated sediments accumulating great quantities
of metals and showing highest photosynthetic leaf surface area and highest leaf biomass. How-
ever, these structural parameters were not seen to be responsible for the differences in the faunal
composition observed between contaminated and reference beds. Moreover, the multivariate
analysis identified the metal content of leaves, biofilm and sediments as important variables
that may be responsible for these differences in faunal composition. In this study we have dem-
onstrated that both the seagrass C. nodosa and the biofilm on the plant leaves may be used as
environmental tools in the Mar Menor lagoon. The former is an useful indicator of sediment
contamination, whereas the latter seems to be a good sentinel of water quality.
� 2004 Elsevier Ltd. All rights reserved.
Keywords: Coastal lagoon; Cymodocea nodosa; Heavy metals; Bioconcentration factor; Bioavailability;
Biofilm
1. Introduction
Coastal lagoons are relatively enclosed water bodies under the influence of both
the marine and the terrestrial environment. Environmental conditions in coastal la-
goons are very changeable due to their confined nature and to their shallowness. In
this sense, they might even be considered as stressed environments. Furthermore,
being close to land, they are vulnerable to human disturbance (Hodgkin, 1994; Reiz-opouluo, Thessalou-Legaki, & Nicolaidou, 1996). Sediments not only support a vari-
ety of biota that are important for maintaining ecosystem structures and functions,
but are also a reservoir for insoluble contaminants (Chapman & Wang, 2001). The
bioavailability of sediment-bound contaminants is determined by sediment constitu-
ents, overlying and interstitial water chemistry, and the behaviour of organisms
(Loring, 1991), and so should be considered as a dynamic process. For this reason,
sediment quality assessment in estuarine areas was a priority research area for sev-
eral years. From an ecological point of view, one of the major problems when study-ing marine sediment pollution is the bioavailability of the metals absorbed by marine
sediments. It is therefore important to determine the fraction of the total metal con-
centration that is available for the living organisms and to know the mechanisms
which regulate metal bioavailability (Mountouris, Voutsas, & Tassios, 2002). Metal
bioavailability is controlled by the physico-chemical characteristics of the sediments,
such as the organic matter content (Chapman & Wang, 2001), grain size distribution
(Krumgalz, 1989), organic carbon content (Mahoney et al., 1996) and the acid-vol-
atile sulfide concentration (Chapman, Wang, Janssen, Persoone, & Allen, 1998). Ithas been suggested (Sundelin & Eriksson, 2001) that the most accurate way to study
the bioavailability of non-essential metals is by means of bioaccumulation measure-
ments in field-collected or laboratory-exposed organisms. The process by which a
chemical species is accumulated in the biota from its surrounding phases is known
as bioconcentration. The bioconcentration factor (BCF) used to calculate the distri-
bution of heavy metals between sediment and biota is defined as the ratio between
the concentration of metals in the biota and the concentration of metals in the sed-
iment (Mountouris et al., 2002).

L. Marın-Guirao et al. / Marine Environmental Research 60 (2005) 317–337 319
Marine macrophytes not only play a key role in the cycling of elements in shallow
nearshore ecosystems (often significantly increasing the bioavailability of toxic sub-
stances to higher consumers), but are also, in turn, adversely affected by such pollu-
tants. Several monitoring programmes have successfully used marine macrophytes to
quantify contamination loads with trace metals (e.g. Constantini, Giordano, Ciaralli,& Beccaloni, 1991; Haritonidis & Malea, 1999). Indeed, because plants have a high
capacity to accumulate a range of elements, their widespread use as indicators of
marine pollution appears justified (Lytle & Lytle, 2001). In addition, plants show
a considerable tolerance to fluctuations in environmental conditions and thus exhibit
one of the fundamental properties that are demanded of ‘‘good’’ bioindicator spe-
cies. Seagrasses, being primary producers, may be used as a first level indicators to
monitor trace metal levels in coastal marine environments (Prange & Dennison,
2000). Cymodocea nodosa is a common and highly productive seagrass species inthe Mediterranean that is often dominant in shallow coastal lagoons (Terrados &
Ros, 1992). The strong capacity to accumulate heavy metals by seagrasses may result
in a negative impact on their associated fauna. The structural complexity provided
by a seagrass bed is responsible for the high species diversity compared with local
unvegetated areas. Many studies have addressed the effects of seagrass complexity
on macrofauna community composition (e.g. Edgar, 1990; Edgar & Robertson,
1992; Edgar, Shaw, Watsona, & Hammond, 1994; Frost, Rowden, & Attrill,
1999). Nevertheless, no ecological studies have investigated the macroinvertebratecommunities associated with C. nodosa beds growing in mining-contaminated sedi-
ments, or the plant metal concentrations that may act as a factor to explain the com-
munity structure. Measures of in situ benthic community conditions reflect the
sensitivities of multiple component species to longer-term exposure and potential
interactions with a variety of natural environmental factors. Thus in situ benthic
conditions may serve as a sensitive indicator of the biological significance of ob-
served contaminant levels (Hyland, Van Dolah, & Snoots, 1999).
The purpose of the present study was to determine the potential impact of land-derived heavymetals on a seagrass ecosystem. Themain objectives were: (i) to quantify
the extent of cadmium, lead and zinc accumulation in different plant fractions (roots,
rhizomes and leaves) and leaf-biofilmofC. nodosa growing inmetal-contaminated sed-
iments, (ii) to analyse existing relations between plant metal contents and the physico-
chemical properties of sediments, (iii) to detect differences in the state of health of theC.
nodosameadows through various meadow parameters and by reconstructing seagrass
dynamics, and (iv) to determine the structure of seagrass communities associated with
each seagrass bed and the main environmental factors that affect such structures.
2. Materials and methods
2.1. Study area and sampling dates
The Mar Menor is a hypersaline coastal lagoon of 135 km2, located in SE Spain.
Sediments of the Mar Menor present high concentrations of heavy metals due to
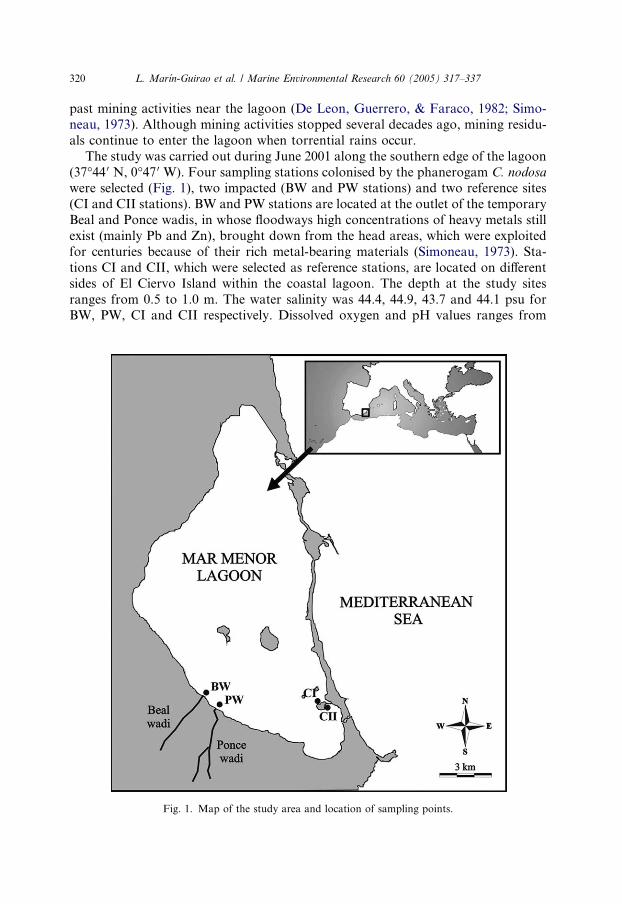
320 L. Marın-Guirao et al. / Marine Environmental Research 60 (2005) 317–337
past mining activities near the lagoon (De Leon, Guerrero, & Faraco, 1982; Simo-
neau, 1973). Although mining activities stopped several decades ago, mining residu-
als continue to enter the lagoon when torrential rains occur.
The study was carried out during June 2001 along the southern edge of the lagoon
(37�44 0 N, 0�47 0 W). Four sampling stations colonised by the phanerogam C. nodosa
were selected (Fig. 1), two impacted (BW and PW stations) and two reference sites
(CI and CII stations). BW and PW stations are located at the outlet of the temporary
Beal and Ponce wadis, in whose floodways high concentrations of heavy metals still
exist (mainly Pb and Zn), brought down from the head areas, which were exploited
for centuries because of their rich metal-bearing materials (Simoneau, 1973). Sta-
tions CI and CII, which were selected as reference stations, are located on different
sides of El Ciervo Island within the coastal lagoon. The depth at the study sites
ranges from 0.5 to 1.0 m. The water salinity was 44.4, 44.9, 43.7 and 44.1 psu forBW, PW, CI and CII respectively. Dissolved oxygen and pH values ranges from
Fig. 1. Map of the study area and location of sampling points.

L. Marın-Guirao et al. / Marine Environmental Research 60 (2005) 317–337 321
8.11 to 8.8 mg/l and from 8.4 to 8.55 respectively. Suspended solids and chlorophyll a
concentrations presented values of 22.13 mg/l and 3.18 mg/m3 respectively in BW; 13
mg/l and 5.17 mg/m3 in PW; 10.53 mg/l and 5.15 mg/m3 in CI and 12 mg/l and 5.47
mg/m3 in CII station.
2.2. C. nodosa (Ucria) Ascherson
To estimate metal accumulation in different C. nodosa tissues, sediment and mac-
rophyte samples were collected by snorkel diving. At each sampling station entire
plants and the top 5 cm of sediment were collected. The plant material was placed
in re-sealable clean polythene bags and transported to the laboratory on ice. In
the laboratory, phanerogam leaves were scraped with a glass slide to separate the
particulate material, extracellular polymeric substances and the epibionts that formthe biofilm present on the leaf surfaces. No calcareous Rhodophyceae were observed
on the plant leaves probably due to the high salinity of the lagoon waters. To remove
the adhering particulate material and sediments from the rest of the plant tissues,
roots and rhizomes were rubbed off with fingers and with a soft tooth-brush, washed
with ultrapure nitric acid (2%) and then rinsed with distilled water. The effectiveness
of the procedure was checked observing the plant tissues with a scanning electron
microscope (Jed JSM T-300). Plants were divided into the following fractions: roots,
rhizomes and leaves. When bacteria and microalgae are associated with surfaces (e.g.sediment particles or plant surfaces) they secrete a matrix of mucilaginous extracel-
lular polymeric substances (EPS) to form a ‘‘microbial biofilm’’ (Decho, 2000). Bio-
film consists of attached algae, bacteria and associated fine detrital material that
adheres to substrates in water bodies and is a food source for invertebrates that
scrape mineral and organic surfaces (Farag, Woodward, Goldstein, Brumbaugh,
& Meyer, 1998). The metal content in the leaf-biofilm was also determined. Each
sample was dried at 60 �C until constant weight. Prior to digestion samples were
powdered in a porcelain mortar and homogenized. Digestions were carried out intriplicate on the homogenized total-sediment, plant fraction and leaf-biofilm. About
0.2 g of sample was digested, adding 1 ml of a nitric, perchloric and sulphuric acid
mixture (8:8:1). Samples were heat-digested by a step-wise increase of temperature to
380 �C until total evaporation. Samples were rediluted with de-ionized water in an
ultrasonic bath, acidified with suprapure chloridric acid. Zn, Cd, and Pb concentra-
tions were measured polarographically by anodic stripping voltammetry (Metrohm
646 VA Processor) with hanging mercury drop.
To determine the structure of each C. nodosa bed, the entire plant content cover-ing an area of 0.09 m2 was sampled (n = 5). In each sample, the width and length of
all the leaves on the shoots collected were measured and the following parameters
were determined: number of leaves per shoot, specific leaf area (cm2), density of
shoots (shoots/m2), leaf biomass (g d.w./m2), leaf index or leaf surface area (m2
leaves/m2 sediment) (Terrados, 1991).
The state of health of each seagrass bed was determined reconstructing seagrass
dynamics from one single sampling. Twelve rhizomes were randomly extracted with
care from the sediments in each station. The reconstruction techniques employed are

322 L. Marın-Guirao et al. / Marine Environmental Research 60 (2005) 317–337
described by Duarte et al. (1994). Briefly, living and dead shoots were counted and
fitted to the model: Nt = N0 e�Mt, where Nt is the number of shoots older than modal
shoots by a time t; N0 is the number of shoots with an age equal to the mode. The
exponential decrease of shoot abundance with increasing age results from shoot
mortality and allows an estimation of the exponential shoot mortality rates (M). An-nual shoot recruitment rates can also be calculated from the total number of shoots
and the number of shoots older than a year in the shoot population. Mortality and
recruitment estimates are useful for evaluating the status of seagrass meadows and as
a forecast of their development (Duarte & Sand-Jensen, 1990).
2.3. Sediments
The sediment parameters, organic matter content, percentage of silt-clay, total or-ganic carbon and total volatile sulfides, were analyzed, in order to run a series of cor-
relation and multiple regression analyses between the plant and sediment variables.
Sediment samples for determining the acid volatile sulfides (AVS) and simulta-
neously extracted metals (SEM) were frozen without airhead space to minimise sul-
fide oxidation. Sediment samples (n = 4) were analyzed for AVS by a cold-acid
purge-and-trap technique described in detail by Allen, Gongmin, and Deng
(1993). The sediment solid-phase sulfide was converted to hydrogen sulfide by adding
6 M hydrochloric acid to the reaction flask. The hydrogen sulfide was purged withnitrogen and trapped in a 2 M NaOH solution. The amount of sulfide was deter-
mined with the help of an ion selective silver/sulfide electrode (ThermoOrion, model
9616). The sediment–water–hydrochloric acid slurry remaining in the reaction flask
was immediately filtered, and Zn, Pb and Cd concentrations were polarographically
measured by anodic stripping voltammetry (Metrohm 646 VA Processor) with hang-
ing mercury drop.
Sediment particle size distribution was determined by mechanical dry sieving (Bu-
chanan, 1984). Samples of oven-dried sediments were sieved through a stacked set ofgraded sieves within the range 2000–62 lm. The sediment organic matter content
was determined as the percentage of weight lost upon ignition of dry sediment in
a muffle furnace at 450 �C for 6 h. The dried sediments were finely ground and care-
fully sieved through a 63 lm stainless steel mesh; the total organic carbon (TOC)
content was then determined with a Carlo Erba Instruments (EA1108) elemental
analyser following sample preparation with 1 N HCl to decompose carbonate (Ver-
dardo, Forelich, & McIntyre, 1990).
2.4. Faunal community
To detect differences between the invertebrate communities of each seagrass bed,
four replicates were taken in each station. Invertebrates associated with the macro-
phytes were directly collected by the blade-clipping method (Edgar, 1990), sampling
an area of 0.09 m2, cutting the macrophytes into mesh-bag of 0.5 mm and taking the
top 3 cm of sediments. The replicate samples from each station were processed to
characterize the infaunal assemblages. Animals were sorted from sample debris

L. Marın-Guirao et al. / Marine Environmental Research 60 (2005) 317–337 323
and plants under a binocular magnifying glass and classified into major constituent
taxa and preserved in 70% alcohol. Taxonomic identification of Crustacea, Mollusca
and Polychaeta was then performed at species level. The following community
descriptive parameters were calculated for each sample and then summarized for
each station: total abundance, species richness (Margelef�s d), Shannon–Wienerdiversity (natural log, H 0), evenness (Pielou�s J), Simpson�s dominance index (D).
The numerical contribution of major taxonomic groups (Polychaeta, Mollusca,
Crustacea) was calculated as the proportion of taxa abundance to total abundance
for each sample and for each station; mean proportions were also determined and
expressed as percentages.
2.5. Data analysis
Differences in the variables measured, including C. nodosa meadow parameters,
sediment physico-chemical variables and metal concentration in different plant frac-
tions and leaf-biofilm, were compared between sites using one-way ANOVA (Dun-
net�s test). Statistical analyses (one-way analysis of variance, correlation coefficients)
were performed on log-transformed data in order to eliminate the dependence of var-
iance on the means (computer application Statistica v4.5). BCFs were calculated by
dividing the metal concentration in each of the plant fractions and in the leaf-biofilm
by the metal concentration in the sediment (total and simultaneously extracted tosulfides). A series of correlation analyses between the BFCs and sediment variables
was run in order to detect the importance of these sediment parameters and the
availability of metals to C. nodosa.
Benthic community analysis was performed using the various routines available
within the PRIMER (Plymouth Routines In Marine Ecological Research) software
package (Clarke & Gorley, 2001). The significance of differences between points
was tested using one-way ANOVA. A ranked triangular similarity matrix was con-
structed using the Bray–Curtis similarity measure on transformed (double root)invertebrate abundance data. Subsequent ordination of the stations was made from
the similarity matrix by non-metric multidimensional scaling (MDS). Formal sig-
nificance tests for differences between samples were preformed using the ANOSIM
permutation test (Clarke & Green, 1988). The taxa contributing to any dissimilar-
ity between samples were investigated by using the similarity percentages procedure
SIMPER (Clarke, 1993). The relationship between community structure and envi-
ronmental variables was explored applying a canonical correspondence analysis
(CCA, unimodal model) (ter Braak, 1986). The ordination technique was per-formed using the program CANOCO version 4.0 (ter Braak & Smilauer, 1998).
Species abundance data, the percentages of organic matter content, total organic
carbon and silt–clay fraction were square root-transformed. Variables related with
the meadow structure and with the metal content in sediments and in the different
plan fractions were log-transformed. To avoid colinearity between variables, the
original data set of environmental variables (n = 28) were reduced to 14. The leaf
surface, leaves biomass and the percentage of organic matter were eliminated from
the analysis since we found highly significant correlation (r > 0.75) with other

324 L. Marın-Guirao et al. / Marine Environmental Research 60 (2005) 317–337
environmental variables. Two groups of highly inter-correlated environmental vari-
ables, one related with sediment metal concentrations and the other with the plant
metal content, were summarized, applying PCA to each group (ter Braak, 1995).
This technique identifies the orthogonal vectors describing a significant portion
of the variance within the original group of variables. The resulting componentscan be treated as traditional variables for further statistical analysis. The benefits
of PCA include the reduction of many variables to a few components, the inclusion
of all original variables in each component so that further analysis is not based on
only a single variable but all variables of the data group, and generation of orthog-
onal, non-correlated, components (Jenerette, Lee, Waller, & Carlson, 2002). Two
new variables labelled SMC (sediment metal content) and PMC (plant metal con-
tent) were obtained from these analyses, selecting the first axis of both PCAs
(explaining 97% and 89% of the variability of these groups of variables, respec-tively) and were included as independent variables in the CCA. The Monte Carlo
permutation test (999 permutations) served us to evaluate the significance of each
environmental variable in the faunal assemblage.
3. Results
3.1. Seagrass biology
Although the number of leaves per shoot was similar in the four sites studied, the
leaf-specific area for the reference siteswas smaller than for the impactedones (Table 1).
This implies that the photosynthetic leaf-surface area per shoot was higher in both
wadi sites. Shoot density showed a different pattern, the highest density correspond-
ing to PW (1538 ± 326 shoots/m2) and the lowest to BW (802 ± 41 shoots/m2), while
the reference sites presented intermediate values. The highest leaf biomass corre-
sponded to the wadi stations (BW and PW).
Table 1
Summary of Cymodocea nodosameadow descriptive parameters (mean ± SD) and parameters employed to
reconstruct seagrass dynamics for each of the meadows sampled
Parameters Impacted stations Non-impacted stations
BW PW CI CII
Leaves per shoot 3.5 ± 0.3 4.1 ± 0.3 4.2 ± 0.2 4.2 ± 0.4
Leaf-specific area (cm2) 14.5 ± 1.3 12.3 ± 0.7 7.7 ± 0.8 5.4 ± 1.4
Shoot density (no. shoots/m2) 802 ± 41 1538 ± 326 1344 ± 58 1029 ± 430
Leaf biomass (g d.w./m2) 96.7 ± 33.6 89.6 ± 13.9 58.9 ± 9.5 30.1 ± 4.5
Leaf index (m2/m2) 1.37 ± 0.3 2.03 ± 0.33 1.05 ± 0.11 0.5 ± 0.06
Gross recruitment (R) 0.62 0.7 0.44 0.82
Mortality (ln units/year) (M) 0.14 0.02 0.04 0.04
Net recruitment (R�M) +0.48* +0.68* +0.4* +0.51*
Biofilm biomass (g d.w./m2 leaf) 0.40 ± 0.20 0.17 ± 0.14 0.20 ± 0.08 0.07 ± 0.01
* Net recruitment (R�M) positive values indicate expanding seagrass beds, negative values indicate the
regression of beds.

L. Marın-Guirao et al. / Marine Environmental Research 60 (2005) 317–337 325
A comparison of recruitment and mortality rates for the individual populations
showed that recruitment exceeded mortality in every meadow (Table 1). Shoot mor-
tality rates differed greatly between sites, BW presenting the highest mortality rates
(0.14 ln units/year) and Ponce wadi the lowest (0.02 ln units/year). The net rate of
population change calculated from the gross recruitment and mortality rates indi-cates that all seagrass beds are expanding, the highest net recruitment values being
shown by the CII bed. The descriptive parameters of each seagrass bed (leaf surface,
number of leaves per shoot, shoot density, leaf biomass and leaf index) showed sig-
nificant differences (p < 0.05) between sampling stations.
3.2. Sediment analysis
As regards general sediment characteristics, stations BW and CII had a largersilt–clay fraction (10.5% and 8.4%, respectively) than station PW and CI (0.6%
and 0.2%, respectively). The percentage of organic matter was highest in BW
(8.2%), as was the percentage of total organic carbon (1.62%). As regards the
other stations, PW, CI and CII, the percentages were 2.4, 3.2 and 3 for organic
matter, and 1.4, 1.2 and 1.3 for total organic carbon, respectively. ANOVA tests
confirmed that there were significant differences (p < 0.05) between stations for
these sediment physico-chemical variables, including the concentration of metals
(total concentration and simultaneously extracted to AVS) and the concentrationof acid volatile sulfide (370.4 lg S/g sed dw for BW, 175.2 for PW, 47 for CI and
90.8 for CII).
3.3. C. nodosa metal accumulation
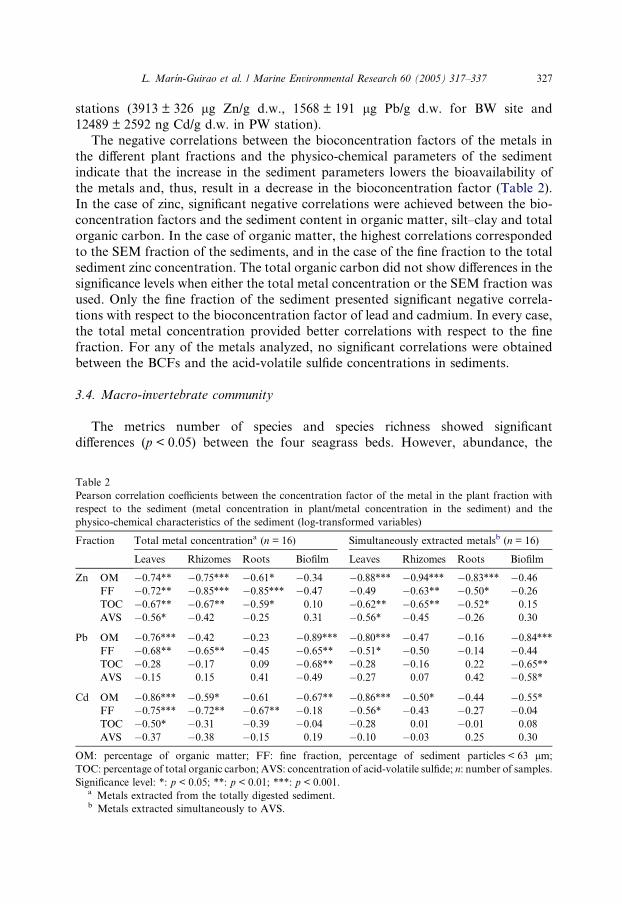
Both the sediments and C. nodosa plants collected from the Mar Menor contained
zinc, lead and cadmium (Fig. 2). There were significant differences (p < 0.05) in each
plant fraction metal concentration between sampling stations. Stations BW and PW,which are located at the outlets of the desert-streams, presented the highest metal
contents in both sediments and in plants. Sediments, biofilm and the plant fractions
of PW station presented higher zinc concentrations than lead and cadmium,
whereas, in BW, the highest concentration in sediments and biofilm corresponded
to zinc, while the lead concentration was slightly higher in roots, rhizomes and
leaves. The metal concentrations determined in the sediments of the selected stations
were higher than in leaves, rhizomes and roots for the three metals determined. The
reference stations (CI and CII) showed similar metal concentrations for the threemetals in the plant fractions. In the case of cadmium, the sediments showed the high-
est values, except in station PW, where the leaf-biofilm showed the greatest concen-
tration. Different patterns were observed between both wadi stations as regards the
plant metal concentrations of all three metals studied, the highest plant metal con-
tent being found in the roots in BW station, but in the leaves of PW plants. The me-
tal content of the rhizomes was lower than in leaves and roots for every station and
metal investigated. The assemblage of mucous substances, algae and microorganisms
that form the leaf-biofilm showed the highest metal concentrations in the polluted

Fig. 2. Metal concentrations in sediments (total and SEM), in the different plant fractions employed in the
study (leaves, rhizomes and roots) and in the leaf-biofilm.
326 L. Marın-Guirao et al. / Marine Environmental Research 60 (2005) 317–337
L. Marın-Guirao et al. / Marine Environmental Research 60 (2005) 317–337 327
stations (3913 ± 326 lg Zn/g d.w., 1568 ± 191 lg Pb/g d.w. for BW site and
12489 ± 2592 ng Cd/g d.w. in PW station).
The negative correlations between the bioconcentration factors of the metals in
the different plant fractions and the physico-chemical parameters of the sediment
indicate that the increase in the sediment parameters lowers the bioavailability ofthe metals and, thus, result in a decrease in the bioconcentration factor (Table 2).
In the case of zinc, significant negative correlations were achieved between the bio-
concentration factors and the sediment content in organic matter, silt–clay and total
organic carbon. In the case of organic matter, the highest correlations corresponded
to the SEM fraction of the sediments, and in the case of the fine fraction to the total
sediment zinc concentration. The total organic carbon did not show differences in the
significance levels when either the total metal concentration or the SEM fraction was
used. Only the fine fraction of the sediment presented significant negative correla-tions with respect to the bioconcentration factor of lead and cadmium. In every case,
the total metal concentration provided better correlations with respect to the fine
fraction. For any of the metals analyzed, no significant correlations were obtained
between the BCFs and the acid-volatile sulfide concentrations in sediments.
3.4. Macro-invertebrate community
The metrics number of species and species richness showed significantdifferences (p < 0.05) between the four seagrass beds. However, abundance, the
Table 2
Pearson correlation coefficients between the concentration factor of the metal in the plant fraction with
respect to the sediment (metal concentration in plant/metal concentration in the sediment) and the
physico-chemical characteristics of the sediment (log-transformed variables)
Fraction Total metal concentrationa (n = 16) Simultaneously extracted metalsb (n = 16)
Leaves Rhizomes Roots Biofilm Leaves Rhizomes Roots Biofilm
Zn OM �0.74** �0.75*** �0.61* �0.34 �0.88*** �0.94*** �0.83*** �0.46
FF �0.72** �0.85*** �0.85*** �0.47 �0.49 �0.63** �0.50* �0.26
TOC �0.67** �0.67** �0.59* 0.10 �0.62** �0.65** �0.52* 0.15
AVS �0.56* �0.42 �0.25 0.31 �0.56* �0.45 �0.26 0.30
Pb OM �0.76*** �0.42 �0.23 �0.89*** �0.80*** �0.47 �0.16 �0.84***
FF �0.68** �0.65** �0.45 �0.65** �0.51* �0.50 �0.14 �0.44
TOC �0.28 �0.17 0.09 �0.68** �0.28 �0.16 0.22 �0.65**
AVS �0.15 0.15 0.41 �0.49 �0.27 0.07 0.42 �0.58*
Cd OM �0.86*** �0.59* �0.61 �0.67** �0.86*** �0.50* �0.44 �0.55*
FF �0.75*** �0.72** �0.67** �0.18 �0.56* �0.43 �0.27 �0.04
TOC �0.50* �0.31 �0.39 �0.04 �0.28 0.01 �0.01 0.08
AVS �0.37 �0.38 �0.15 0.19 �0.10 �0.03 0.25 0.30
OM: percentage of organic matter; FF: fine fraction, percentage of sediment particles < 63 lm;
TOC: percentage of total organic carbon; AVS: concentration of acid-volatile sulfide; n: number of samples.
Significance level: *: p < 0.05; **: p < 0.01; ***: p < 0.001.a Metals extracted from the totally digested sediment.b Metals extracted simultaneously to AVS.
328 L. Marın-Guirao et al. / Marine Environmental Research 60 (2005) 317–337
Shannon–Wiener diversity index, the Pielou evenness and the Simpson dominance
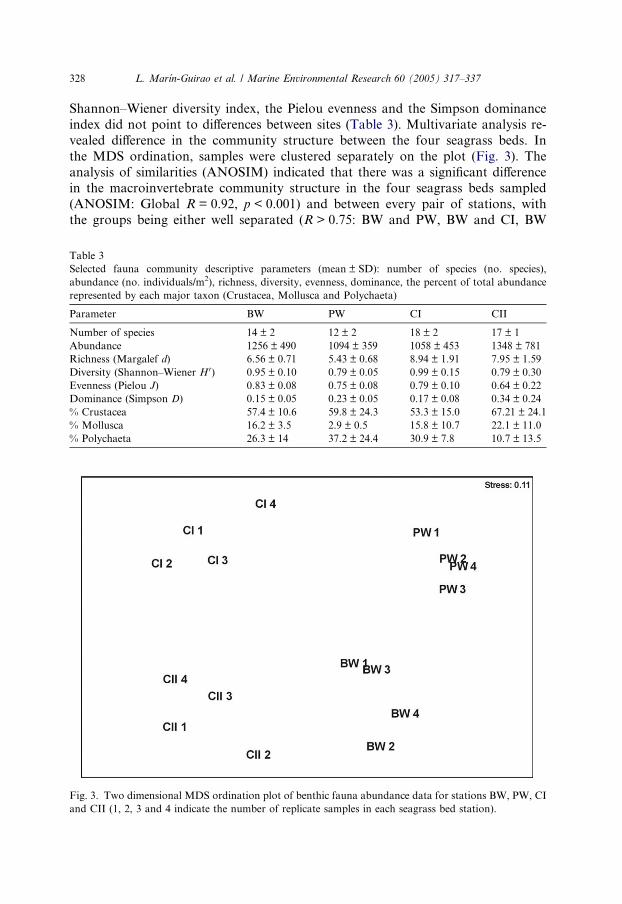
index did not point to differences between sites (Table 3). Multivariate analysis re-
vealed difference in the community structure between the four seagrass beds. In
the MDS ordination, samples were clustered separately on the plot (Fig. 3). The
analysis of similarities (ANOSIM) indicated that there was a significant differencein the macroinvertebrate community structure in the four seagrass beds sampled
(ANOSIM: Global R = 0.92, p < 0.001) and between every pair of stations, with
the groups being either well separated (R > 0.75: BW and PW, BW and CI, BW
Table 3
Selected fauna community descriptive parameters (mean ± SD): number of species (no. species),
abundance (no. individuals/m2), richness, diversity, evenness, dominance, the percent of total abundance
represented by each major taxon (Crustacea, Mollusca and Polychaeta)
Parameter BW PW CI CII
Number of species 14 ± 2 12 ± 2 18 ± 2 17 ± 1
Abundance 1256 ± 490 1094 ± 359 1058 ± 453 1348 ± 781
Richness (Margalef d) 6.56 ± 0.71 5.43 ± 0.68 8.94 ± 1.91 7.95 ± 1.59
Diversity (Shannon–Wiener H0) 0.95 ± 0.10 0.79 ± 0.05 0.99 ± 0.15 0.79 ± 0.30
Evenness (Pielou J) 0.83 ± 0.08 0.75 ± 0.08 0.79 ± 0.10 0.64 ± 0.22
Dominance (Simpson D) 0.15 ± 0.05 0.23 ± 0.05 0.17 ± 0.08 0.34 ± 0.24
% Crustacea 57.4 ± 10.6 59.8 ± 24.3 53.3 ± 15.0 67.21 ± 24.1
% Mollusca 16.2 ± 3.5 2.9 ± 0.5 15.8 ± 10.7 22.1 ± 11.0
% Polychaeta 26.3 ± 14 37.2 ± 24.4 30.9 ± 7.8 10.7 ± 13.5
Fig. 3. Two dimensional MDS ordination plot of benthic fauna abundance data for stations BW, PW, CI
and CII (1, 2, 3 and 4 indicate the number of replicate samples in each seagrass bed station).

L. Marın-Guirao et al. / Marine Environmental Research 60 (2005) 317–337 329
and CII, PW and CI, PW and CII) or overlapping but clearly different (R > 0.5: CI
and CII). The SIMPER analysis was used to determine which organisms contributed
to the similarity/dissimilarity observed. Within the four seagrass beds studied, sam-
ples from CII site exhibited the lowest similarity (BW = 67.76; PW = 71.12;
CI = 65.61; CII = 56.58). No single species had a large contributory influence tothe dissimilarities between the seagrass beds. The maximum contributions came from
three species of amphipod, Microdeutopus sp. (10.98%), Siphonoecetes sabatieri
(8.57%) and Gammarus sp. (7.70%). The largest dissimilarity between each pair of
stations was found between PW and CII (65.58%), which was mainly due to amphi-
pod species. Samples from BW and CII showed the lowest percentage of dissimilarity
(49.54%), the main organisms responsible for the dissimilarity being the amphipod
Siphonoecetes sabatieri, some members of the Spionidae Polychaeta family and the
gastropod Bittium paludosum. For the rest of the pairs of stations, the percentageof dissimilarity were 50.53 for BW and PW, 61.50 for BW and CI, 62.35 for PW
and CI and 51.64 for CI and CII.
The CCA graph (Fig. 4) relating environmental variables to the faunal assem-
blages showed each of the four stations located in each of the four quadrants of
the plot. The vectors depicted in the plot are scaled to the coordinates of the Pearson
product moment correlations of the various environmental variables calculated indi-
vidually with the two axes developed by the correspondence analysis. Each vector
represents a gradient through the biological data, with the arrow pointing to the areaof higher value for the variable, except the two new variables SMC and PMC ob-
tained with the PCAs, whose arrows points to the areas of lower values. Vertical axis
II clearly separates two groups of stations, each composed of one polluted and one
reference site. This axis was positively correlated with the epiphyte biomass and with
the silt–clay fraction of the sediments and negatively correlated with the shoots den-
sity, the leaf index and with the lower sediment metal content. Horizontal axis I,
which clearly separated the reference sites (CI and CII) from the polluted ones
(BW and PW), was positively correlated with the AVS concentration and the concen-tration of Cd in epiphytes and negatively correlated with the lower plant and sedi-
ment metal content. A clearly marked gradient was identified since the first
eigenvalue was 0.324 (Table 4). For this analysis 60% of the macroinvertebrate com-
munity variability was explained by the environmental variables as represented by
the first two axes depicted in Fig. 4. The test of significance for the first axis and over-
all CCA was highly significant (P < 0.005; Monte Carlo permutation test).
4. Discussion
Contaminants in sediments are distributed between the solid particles and the
water filling the interstices between the particles (pore water). This partitioning of
the contaminants is determined by specific combinations of chemical, biological
and environmental parameters, which make it a dynamic process. C. nodosa, as other
plants, accumulates the contaminants of the sediments from the dissolved fraction
(pore water) but can also induce changes in the chemistry of the sediment and
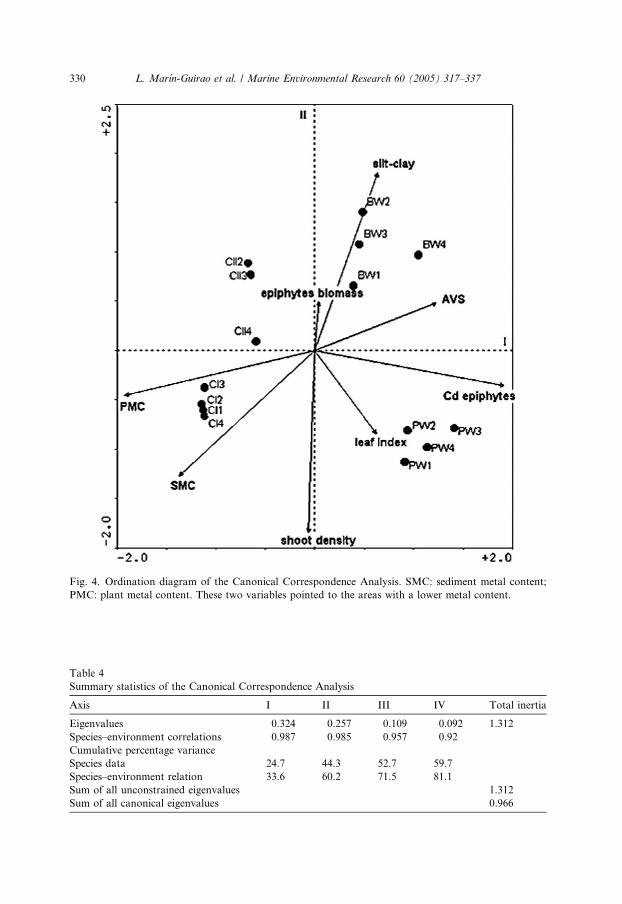
Fig. 4. Ordination diagram of the Canonical Correspondence Analysis. SMC: sediment metal content;
PMC: plant metal content. These two variables pointed to the areas with a lower metal content.
Table 4
Summary statistics of the Canonical Correspondence Analysis
Axis I II III IV Total inertia
Eigenvalues 0.324 0.257 0.109 0.092 1.312
Species–environment correlations 0.987 0.985 0.957 0.92
Cumulative percentage variance
Species data 24.7 44.3 52.7 59.7
Species–environment relation 33.6 60.2 71.5 81.1
Sum of all unconstrained eigenvalues 1.312
Sum of all canonical eigenvalues 0.966
330 L. Marın-Guirao et al. / Marine Environmental Research 60 (2005) 317–337

L. Marın-Guirao et al. / Marine Environmental Research 60 (2005) 317–337 331
sediment pore water, affecting the sorption of toxicants and their equilibrium parti-
tioning and thus their bioavailability. As is reflected in this study, C. nodosa accumu-
lates metals from the sediments, reflecting their bioavailability for plants growing in
the contaminated sediments of the south basin of the Mar Menor coastal lagoon.
Cadmium, lead and zinc were selected in this study because they are the main metalsproduced in the mining activities of the surrounding area, and are the metals present
in highest concentrations in the sediments (De Leon et al., 1982). Zinc is a trace me-
tal required by plants for survival, whereas cadmium and lead are both toxic metals
that are not required by plants. The accumulation patterns of trace metals differ be-
tween parts of C. nodosa, as was seen in the seagrass Posidonia oceanica (Schlacher-
Hoenlinger & Schlacher, 1998) and in the eelgrass Zostera marina (Lyngby & Brix,
1989). The zinc concentrations recorded in our study for C. nodosa exceeded the
background level for non-contaminated areas in stations BW and PW (Moore &Ramamoorthy, 1984). In these stations zinc and lead levels in different parts of
the plant were higher than in other studies previously published (Capone, Mascia,
Porcu, & Tagliasacchi-Masala, 1983; Malea & Haritonidis, 1994; Prange & Denni-
son, 2000; Sanchiz, Garcıa-Carrascosa, & Pastor, 2000). The highest Zn concentra-
tion in the three plant fractions studied at all the study sites, except BW station, was
located in the leaf material. Nicolaidou and Nott (1998) obtained similar patterns in
a nickel smelter in Greece. Root uptake with subsequent translocation to above-
ground tissues has been demonstrated for a variety of elements, including metals(Jackson, Rowan, Cornett, & Kalff, 1994) and may be an important vector out of
the sediments for elements that would otherwise remain buried.
Although there is some solid evidence that much of the Cd and Pb uptake from
waters into seagrass leaves and epiphytes follows a passive, surface area-dependent
process (Ward, 1989), differences in the absolute uptake kinetics between plant or-
gans may be appreciable. In our case, the highest Pb and Cd levels were found in
the leaves of C. nodosa, except in the case BW station, where roots presented higher
values than leaves. The differences observed between metal concentrations in leavesand roots from both contaminated stations might be due to the great SEM-AVS mo-
lar difference. In BW station the SEM-AVS molar difference (106.18 ± 5.18 lmol/g
dry sed) indicates that great quantities of divalent metals are not bound to sulfides
and thus remain potentially bioavailable, whereas in PW the difference were close
to 0 (0.10 ± 0.08 lmol/g dry sed), indicating that most of the metals form insoluble
metal–sulfide complexes, thus reducing their bioavailability. In this sense, something
similar to that of PW station might occur in the reference stations, where the SEM-
AVS molar difference was negative, indicating that metals are bound to sulfides(�0.005 and �1.57 lmol/g dry sed for CI and CII, respectively).
In the wadi stations (BW and PW) the biofilm showed lead and zinc concentrations
that were about one order of magnitude higher than in the plant. These high metal
concentrations found in the leaf-biofilm in BW and PW stations are probably the re-
sult of the high metal absorption capacity of the EPS. In a laboratory experiment Suh,
Yun, and Kim (1999) found that the excreted EPS was responsible for more than 90%
of Pb2+ accumulation of the total lead dissolved, while Schlekat, Decho, and Chan-
dler (1998) found a very strong cadmium chelation capacity across a wide range of

332 L. Marın-Guirao et al. / Marine Environmental Research 60 (2005) 317–337
salinity and pH conditions in an isolated estuarine bacterium which produced EPS. In
addition, since biofilm, by definition, is a complex of functional consortia made up of
living microorganisms and of inorganic and organic solids, the high metal concentra-
tion measured in the biofilm might also be due to the deposition of metal-absorbed
particulate material introduced through both desert-streams. When torrential rainsintroduced heavy metals into the sea, the greatest part of these metals was associated
with suspended particulate material and resulted in the deposition of metals (Bradley
& Lewin, 1982; Polh, Hennings, Siegel, & Bachor, 2002). In this sense, the swell might
cause the resuspension of the sediments, increasing the amount of dissolved and par-
ticulate metals that would be absorbed over the biofilm. The high leaf-biofilm lead
concentration found in the reference stations may reflect the effect of other contam-
ination sources in the Mar Menor lagoon (maritime traffic, the proximity of a road or
the atmospheric deposition from military flights in the area). In the light of the above,the biofilm could reflect pulse metal inputs into the aquatic systems, especially in
hypersaline coastal lagoons, where metals remain dissolved for a short time.
The differences in metal concentrations found between plants and leaf-biofilm
suggest that leaves and their associated biofilm-epibiota cannot be considered as a
homogeneous compartment (Schlacher-Hoenlinger & Schlacher, 1998) and may be
considered a convenient tool for environmental monitoring and employed as a sen-
tinel accumulator.
The substantial Zn, Pb and Cd concentrations in the plant and in sediments sug-gest that tissue concentrations reflect the availability of metals in the environment. A
positive relationship between metal contents in the sediments and in C. nodosa and
other seagrasses was also mentioned by other authors (Malea & Haritonidis, 1994;
Malea, Haritonidis, & Kevrekidis, 1994).
The negative correlations between the BCFs of Zn, Pb and Cd in the different
plant fractions in relation to the total sediment concentrations and the fine fraction
of the sediments indicate that an increased fine fraction lowers the bioavailability of
metals and, thus, decreases the BCF. Dense canopies function as effective traps offine-grained particles and contribute to maintaining sediment redox potentials at lev-
els that promote phase partitioning to the ionic, and thus bioavailable form of the
metals (Jackson, 1998). This fine trapped sediment fraction will affect the absorption
of metals and thus their bioavailability. In the case of zinc, significant negative cor-
relations were also found with the organic matter and the total organic carbon con-
tent. As indicated by Campbell et al. (1988), the metals that preferentially bind to
organic ligands, like Zn, are probably less available to rooted macrophytes when
such ligands are available for binding. Other sediment components, such as the vol-atile sulfides, do not seem to influence the availability of metals to the seagrass, pos-
sibly due to oxygen transport to underground tissues, which would change the
physico-chemical conditions of the sediments and oxidize the sulfides. Marba and
Duarte (2001) suggested that the release of oxygen into the sediments by C. nodosa,
may increase the oxidation of reduced compounds (e.g. sulfide) by creating a suboxic
zone, where redox potentials would be higher than those in the reduced zone. This
increase in redox conditions within the seagrass rhizosphere may help to avoid the
accumulation of phytotoxins such as sulfide in the sediment but may increase the

L. Marın-Guirao et al. / Marine Environmental Research 60 (2005) 317–337 333
availability of metals to the plant. Recently Jacob and Otte (2004) indicated that liv-
ing plants in permanently flooded areas significantly decrease concentrations of acid-
volatile sulfides, indicating root-induced sediment oxidation.
Although both seagrass beds growing in the metal-enriched sediments influenced
by desert-streams (BW and PW) accumulated great quantities of metals, no detri-mental effects were observed on the plants growing. Furthermore, these two seagrass
beds presented the highest photosynthetic leaf surface and the highest leaf biomass.
It has previously been seen that mining tailings with a high heavy metal content
mixed with stream sediments, enriched in organic matter, result in floodplain soils
that may even be attractive as amendments to increase the productivity of home gar-
dens (Cobb, Sands, Waters, Wixson, & Dorward-King, 2000; Samantaray, Rout, &
Das, 2001). Something similar may be occurring in both seagrass beds, where an
increment in the supply of trace elements and nutrients introduced through desertstreams will increase plant productivity (Perez, Duarte, Romero, Sand-Jensen, &
Alcoverro, 1994). The shoot density was higher in PW and lower in BW, which
was closely related with shoot mortality. The higher mortality rate found in BW than
in the PW bed may be due to the greater quantity of leaf-biofilm in BW plants, which
would diminish the light intensity available for plant survival (Ruiz, Marın, Calvo, &
Ramirez, 1993; Ruiz & Romero, 2003).
The univariate metrics (except species richness) applied in the present study to the
seagrass-associated macroinvertebrate communities were not significantly differentbetween the four seagrass beds studied. However, multivariate analysis, which is of-
ten recognized as being more sensitive for this sort of data (Clarke & Warwick,
2001), revealed a significant difference in community composition. Something similar
was observed by Frost et al. (1999) when studying the macroinvertebrate communi-
ties associated with the seagrass Z. marina.
As reflected in the CCA, those variables related with the seagrass habitat structure
(shoot density, silt–clay fraction, leaf index and epiphyte biomass) separates two
groups of stations, each composed of one contaminated and one reference site. Manystudies have demonstrated that faunal community structures associated with sea-
grasses are closely correlated with seagrass physical parameters such as shoot density,
leaf surface and leaf biomass since they provide the physical structure usable as living
space and protection from predators (Edgar & Barret, 2002; Edgar & Robertson,
1992; and cites therein). Furthermore, they increase the food resources such as dia-
tom/bacterial mats (biofilm) associated with plant surfaces (Edgar, 1990; Orth &
Van Montfrans, 1984; Parker, Duffy, & Orth, 2001) and the sediment and seagrass
detritus retained as a result of the reduction in hydrodynamic effects (De Troch, Fiers,& Vincx, 2003; Edgar et al., 1994; Jenkins, Walker-Smith, & Hamer, 2002). These
important parameters, which are closely related with the faunal structures associated
with seagrasses, do not seem tomark the differences between the contaminated and the
reference seagrass beds. However, those variables related with the metal content in the
different plant fractions (including the biofilm) and in sediments, appeared to best ex-
plain the separation of the contaminated stations from the reference ones. Most of
macroinvertebrate species associated with seagrass beds do not directly feed on the
plant, but on the biofilm-epiphyte growing on the leaf surface, which is more palatable

334 L. Marın-Guirao et al. / Marine Environmental Research 60 (2005) 317–337
to them and which is an important food source for heterotrophs in tropical seagrass
beds (Yamamuro, 1999). The metal content of this food source may be important
in determining the structure of the communities associated with seagrasses and may
transfer metals through dietary ingestion (Boult, Johnson, & Curtis, 1997; Schlekat,
Decho, & Chandler, 2000). Furthermore, during leaf senescence, the seagrass detritusgenerated may transfer metals to the detritivores by detritus ingestion or by liberating
metal to the water column when decomposing. In a study of metal biotransference in a
seagrass ecosystem, Barwick and Maher (2003) showed that metals, such as Zn, Pb,
Cu and Cd, were bioaccumulated and biotransferred through the food chain, princi-
pally in detritivores and herbivores that feed directly the epiphytes, plants or detritus.
Finally, although the metal concentrations in C. nodosa are higher than those
found in the bibliography, the distribution of the seagrass in the coastal lagoon, does
not seem to be influenced by sediment metal contamination but by other environ-mental factors, such as the influence of the desert-streams (Beal and Ponce wadis).
Although seagrass is probably not affected by metal accumulation, we found differ-
ences in the invertebrate communities associated with the seagrass beds, which may
be due to the toxic effects of the metal content of the invertebrates� food sources (sed-
iments, leaves and biofilm), since other environmental parameters, including as salin-
ity, pH, dissolved oxygen concentration, suspended solids and chlorophyll a, did not
show conspicuous differences in the bodywater of the lagoon.
We have evidenced that both the seagrass C. nodosa and the biofilm on the plantleaves may be used as environmental tools in the Mar Menor lagoon. We consider
this marine phanerogam to be a useful indicator of sediment contamination, which
may serve to study the long-term evolution of the contaminant load in sediments,
reflecting the potential metal bioavailability for the biota. On the other hand, in
the case of water contamination, the leaf-biofilm seems to be good sentinel of the
water quality as it rapidly reflects the entrance of metals in the aquatic environment
and can be used as a sentinel accumulator.
Acknowledgements
Special thanks to Dr. Andres Mellado for his helpful guidance in the Canoco
analysis. We are grateful to two anonymous referees for their useful comments
and constructive suggestions. We wish to thank to Philip Thomas for the English
revision of the manuscript. This work was funded by the European Regional Devel-
opment Fund (ERDF), Consejerıa de Agricultura, Agua y Medio Ambiente, Regionde Murcia, Programa Seneca, 2001 (AGR/24/FS/02).
References
Allen, H. E., Gongmin, F., & Deng, B. (1993). Analysis of acid volatile sulfide (AVS) and simultaneously
extracted metals (SEM) for estimation of potential toxicity in aquatic sediments. Environmental
Toxicology and Chemistry, 12, 1441–1453.

L. Marın-Guirao et al. / Marine Environmental Research 60 (2005) 317–337 335
Barwick, M., & Maher, W. (2003). Biotransference and biomagnification of selenium, cooper, cadmium,
zinc, arsenic and lead in a temperate seagrass ecosystem from Lake Macquarie Estuary, NSW,
Australia. Marine Environmental Research, 56, 471–502.
Boult, S., Johnson, N., & Curtis, C. (1997). Recognition of a biofilm at the sediment–water interface of an
acid mine drainage-contaminated stream, and its role in controlling iron flux. Hydrological Processes,
11, 391–399.
Bradley, S. B., & Lewin, J. (1982). Transport of heavy metals on suspended sediments under high flow
conditions in a mineralised region of Wales. Environmental Pollution Series B, 4, 257–267.
Buchanan, J. B. (1984). Sediment analysis. In N. A. Holme & A. D. Mcintyre (Eds.),Methods for the study
of marine benthos (pp. 41–65). Blackwell Scientific Publications.
Campbell, P. G. C., Lewis, A. G., Chapman, P. M., Crowder, A. A., Fletcher, W. K., Imber, B., et al.
(1988). Biologically available metals in sediments. Publ. No. 27694, National Research Council of
Canada, Ottawa, Ont..
Capone, W., Mascia, C., Porcu, M., & Tagliasacchi-Masala, M. L. (1983). Uptake of lead and
chromium by primary producers and consumers in a polluted lagoon. Marine Pollution Bulletin, 3,
97–102.
Chapman, P. M., & Wang, F. (2001). Assessing sediment contamination in estuaries. Environmental
Toxicology and Chemistry, 20, 3–22.
Chapman, P. M., Wang, F., Janssen, C., Persoone, G., & Allen, H. E. (1998). Ecotoxicology of metals in
aquatic sediments: Binding and release, bioavailability, risk assessment, and remediation. Canadian
Journal of Fisheries and Aquatic Sciences, 55, 2221–2243.
Clarke, K. R. (1993). Non-parametric multivariate analyses of changes in community structure. Australian
Journal of Ecology, 18, 117–143.
Clarke, K. R. & Gorley, R. N. (2001). PRIMER v5: User Manual/Tutorial. Plymouth, UK.
Clarke, K. R., & Green, R. H. (1988). Statistical design and analysis for a biological effects study. Marine
Ecology Progress Series, 46, 213–226.
Clarke, K. R., & Warwick, R. M. (2001). Change in marine communities: an approach to statistical analysis
and interpretation (2nd ed.). UK: Plymouth Marine Laboratory.
Cobb, G. P., Sands, K., Waters, M., Wixson, B. G., & Dorward-King, E. (2000). Accumulation of
heavy metals by vegetables grown in mine wastes. Environmental Toxicology and Chemistry, 19,
600–607.
Constantini, S., Giordano, R., Ciaralli, L., & Beccaloni, E. (1991). Mercury, cadmium and lead evaluation
in Posidonia oceanica and Codium tormentosum. Marine Pollution Bulletin, 22, 362–363.
Decho, A. W. (2000). Microbial biofilms in intertidal systems: an overview. Continental Shelf Research, 20,
1257–1273.
De Leon, A. R., Guerrero, J. & Faraco, F. (1982). Evolution of the pollution of the coastal lagoon of Mar
Menor. VI Journees Etud. Pollutions, Cannes, CIESM.
De Troch, M., Fiers, F., & Vincx, M. (2003). Niche segregation and habitat specialisation of harpacticoids
copepods in a tropical seagrass bed. Marine Biology, 142, 345–355.
Duarte, C. M., Marba, N., Agawin, N., Cebrian, J., Enriquez, S., Fortes, M. D., et al. (1994).
Reconstruction of seagrass dynamics: age determinations and associated tools for the seagrass
ecologist. Marine Ecology Progress Series, 107, 195–209.
Duarte, C. M., & Sand-Jensen, K. (1990). Seagrass colonization: Biomass development and shoot
demography in Cymodocea nodosa patches. Marine Ecology Progress Series, 67, 97–103.
Edgar, G. J. (1990). The influence of plant structure on the species richness, biomass and secondary
production of macrofaunal assemblages associated with Western Australian seagrass beds. Journal
of Experimental Marine Biology and Ecology, 137, 215–240.
Edgar, G. J., & Barret, N. S. (2002). Benthic macrofauna in Tasmanian estuaries: scales of distribution and
relationships with environmental variables. Journal of Experimental Marine Biology and Ecology, 270,
1–24.
Edgar, G. J., & Robertson, A. I. (1992). The influence of seagrass structure on the distribution and
abundance of mobile epifauna: pattern amd process in a Western Australian Amphibolis bed. Journal of
Experimental Marine Biology and Ecology, 160, 13–31.

336 L. Marın-Guirao et al. / Marine Environmental Research 60 (2005) 317–337
Edgar, G. J., Shaw, C., Watsona, G. F., & Hammond, L. S. (1994). Comparisons of species richness, size-
structure and production of benthos in vegetated and unvegetated habitats in Western Port, Victoria.
Journal of Experimental Marine Biology and Ecology, 176, 201–226.
Farag, A. M., Woodward, D. F., Goldstein, J. N., Brumbaugh, W., & Meyer, J. S. (1998). Concentrations
of metals associated with mining waste in sediments, biofilm, benthic macroinvertebrates, and fish from
the Coeur d�Alene River Basin, Idaho. Archives of Environmental Contamination and Toxicology, 34,
119–127.
Frost, M. T., Rowden, A. A., & Attrill, M. J. (1999). Effect of habitat fragmentation on the
macroinvertebrate infaunal communities associated with the seagrass Zostera marina L. Aquatic
Conservation: Marine and Freshwater Ecosystems, 9, 255–263.
Haritonidis, S., & Malea, P. (1999). Bioaccumulation of metals by the green alga Ulva rigida from
Thermaikos Gulf, Greece. Environmental Pollution, 104, 365–372.
Hodgkin, E. P. (1994). Estuaries and coastal lagoons. In L. S. Hammond & R. N. Synnot (Eds.), Marine
biology (pp. 315–322). Melbourne, Australia: Longman Cheshire.
Hyland, J. L., Van Dolah, R. F., & Snoots, T. R. (1999). Predicting stress in benthic communities of
southeastern US estuaries in relation to chemical contamination of sediments. Environmental
Toxicology and Chemistry, 18, 2557–2564.
Jackson, L. J. (1998). Paradigms of metal accumulation in rooted aquatic vascular plants. The Science of
Total Environment, 219, 223–231.
Jackson, L. J., Rowan, D. J., Cornett, R. J., & Kalff, J. (1994). Myriophyllum spicatum pumps essential
and nonessential trace elements from sediments to epiphytes. Canadian Journal Fish and Aquatic
Science, 51, 1769–1773.
Jacob, D. L., & Otte, M. L. (2004). Long-term effects of submergence and wetland vegetation on metals in
a 90-year old abandoned Pb–Zn mine tailings pond. Environmental Pollution, 130, 337–345.
Jenerette, G. D., Lee, J., Waller, D. W., & Carlson, R. E. (2002). Multivariate analisis for ecoregion
delineation for aquatic systems. Environmental Management, 29, 67–75.
Jenkins, G. P., Walker-Smith, G. K., & Hamer, P. A. (2002). Elements of habitat complexity that influence
harpacticoid copepods associated with seagrass beds in a temperate bay. Oecologia, 131, 598–605.
Krumgalz, B. C. (1989). Unusual grain size effect on trace metals and organic matter in contaminated
sediments. Marine Pollution Bulletin, 20, 608–611.
Loring, D. H. (1991). Normalization of heavy-metal data from estuarine and coastal sediments. ICES
Journal of Marine Science, 48, 101–115.
Lyngby, J. E., & Brix, H. (1989). Heavy metals in eelgrass (Zostera marina L.) during growth and
decomposition. Hydrobiologia, 176/177, 189–196.
Lytle, J. S., & Lytle, T. F. (2001). Use of plants for toxicity assessment of estuarine ecosystems.
Environmental Toxicology and Chemistry, 20, 68–83.
Mahoney, D. J., Di Toro, M. D., Gonzalez, M. A., Curto, M., Dilg, M., De Rosa, D. L., et al. (1996).
Partitioning of metals to sediment organic carbon. Environmental Toxicology and Chemistry, 15,
2187–2197.
Malea, P., & Haritonidis, S. (1994). Local distribution and seasonal variation of Fe, Pb, Zn, Cu, Cd, Na,
K, Ca and Mg concentrations in the seagrass Cymodocea nodosa (Ucria) Aschers. In the Antikyra Gulf,
Greece. Marine Ecology, 16, 41–56.
Malea, P., Haritonidis, S., & Kevrekidis, T. (1994). Seasonal and local variations of metal concentrations
in the seagrass Posidonia oceanica (L.) Delile in the Antikyra Gulf, Greece. The Science of Total
Environment, 153, 225–235.
Marba, N., & Duarte, C. M. (2001). Growth and sediment space occupation by seagrass Cymodocea
nodosa roots. Marine Ecology Progress Series, 224, 291–298.
Moore, J. W., & Ramamoorthy, S. (1984). Heavy metals in natural waters. New York, Berlin, Heidelberg,
Tokyo: Springer.
Mountouris, A., Voutsas, E., & Tassios, D. (2002). Bioconcentration of heavy metals in aquatic
environments: the importance of bioavailability. Marine Pollution Bulletin, 44, 1136–1141.
Nicolaidou, A., & Nott, J. A. (1998). Metals in sediment, seagrass and gastropods near a nickel smelter in
Greece: Possible interactions. Marine Pollution Bulletin, 36, 360–365.

L. Marın-Guirao et al. / Marine Environmental Research 60 (2005) 317–337 337
Orth, R. J., & Van Montfrans, J. (1984). Epiphyte–seagrass relationships with an emphasis on the role of
micrograzing: a review. Aquatic Botany, 18, 43–69.
Parker, J. D., Duffy, J. E., & Orth, R. J. (2001). Plant species diversity and composition: experimental
effects on marine epifaunal assemblages. Marine Ecology Progress Series, 224, 55–67.
Perez, M., Duarte, M. C., Romero, J., Sand-Jensen, K., & Alcoverro, T. (1994). Grouth plasticity in
Cymodocea nodosa stands: the importance of nutrient supply. Aquatic Botany, 47, 249–264.
Polh, C., Hennings, U., Siegel, H., & Bachor, A. (2002). Trace metal impact into the Balthic Sea during the
excepcional Oder flood in summer 1997. Marine Chemistry, 79, 101–111.
Prange, J. A., & Dennison, W. C. (2000). Physiological responses of five seagrass species to trace metals.
Marine Pollution Bulletin, 41, 327–336.
Reizopouluo, S., Thessalou-Legaki, M., & Nicolaidou, A. (1996). Assessment of disturbance in
Mediterranean lagoons: an evaluation of methods. Marine Biology, 125, 189–197.
Ruiz, J. M., Marın, A., Calvo, J. F., & Ramirez, L. (1993). Interactions between a floodway and coastal
constructions in Aguilas Bay (Southeastern Spain). Ocean and Coastal Management, 19, 241–262.
Ruiz, J. M., & Romero, J. (2003). Effects of disturbances caused by coastal constructions on spatial
structure, growth dynamics and photosynthesis of the seagrass Posidonia oceanica. Marine Pollution
Bulletin, 46, 1523–1533.
Samantaray, S., Rout, G. R., & Das, P. (2001). Heavy metal and nutrient concentration in soil and plants
growing on a metalliferous chromite minespoil. Environmental Technology, 22, 1147–1154.
Sanchiz, C., Garcıa-Carrascosa, A. M., & Pastor, A. (2000). Heavy metal contents in soft-bottom marine
macrophytes and sediments along the Mediterranean coast of Spain. Marine Ecology, 21, 1–16.
Schlacher-Hoenlinger, M. A., & Schlacher, T. A. (1998). Accumulation, contamination and seasonal
variability of trace metals in the coastal zone – patterns in a seagrass meadow from the Mediterranean.
Marine Biology, 131, 401–410.
Schlekat, C. E., Decho, A. W., & Chandler, G. T. (1998). Sorption of cadmium to bacterial extracelular
polymeric sediment coatings under estuarine conditions. Environmental Toxicology and Chemistry, 17,
1867–1874.
Schlekat, C. E., Decho, A. W., & Chandler, G. T. (2000). Bioavailability of particle-associated silver,
cadmium and zinc to the estuarine amphipod Leptocheirus plumulosus through dietary ingestion.
Limnology and Oceanography, 45, 11–21.
Simoneau, J. (1973). Mar Menor; Evolution Sedimentologique et Geochimique recente du remplissage.
These presente a l�Universite Paul Sebatier de Tolouse (Sciences). France.
Suh, J. H., Yun, J. W., & Kim, D. S. (1999). Effect of extracelular polymeric substances (EPS) on Pb2+
accumulation by Aureobasidium pullulans. Bioprocess Engineering, 21, 1–4.
Sundelin, B., & Eriksson, A. (2001). Mobility and bioavailability of trace metals in sulfidic coastal
sediments. Environmental Toxicology and Chemistry, 20, 748–756.
ter Braak, C. J. F. (1986). Cannonical correspondence analysis: a new eigenvector technique for
multivariate direct gradient analysis. Ecology, 67, 1167–1179.
ter Braak, C. J. F. (1995). Ordination. In R. H. G. Jongman, C. J. F. ter Braak, & O. F. R. van Tongeren
(Eds.), Data analysis in community and landscape ecology (pp. 91–173). Cambridge, UK: Cambridge
University Press.
terBraak,C. J.F.&Smilauer,P. (1998).CANOCOreferencemanualanduser�s guide toCanoco forWindows
Software for Canonical Community Ordination (version 4). Microcomputer Power, Ithaca, NY.
Terrados, J. (1991). Crecimiento y produccion de las praderas de macrofitos del Mar Menor, Murcia.
Ph.D. Thesis. University of Murcia.
Terrados, J., & Ros, J. D. (1992). Growth and primary production of Cymodocea nodosa (Ucria)
Ascherson in a Mediterranean coastal lagoon. Aquatic Botany, 43, 63–74.
Verdardo, D. J., Forelich, P. N., & McIntyre, A. (1990). Determination of organic carbon and nitrogen in
marine sediments using the Carlo Erba NA-1500 analyzer. Deep Sea Research, 37, 157–165.
Ward, T. J. (1989). The accumulation and effects of metals in seagrass habitats. In A. W. D. Larkum, A. J.
U. McComb, & S. A. Shepherd (Eds.), Biology of seagrasses (pp. 797–820). Amsterdam: Elsevier.
Yamamuro, M. (1999). Importance of epiphytic cyanobacteria as food sources for heterotrophs in a
tropical seagrass bed. Coral Reefs, 18, 263–271.
Copyright © 2022 FDOKUMEN




















