
Effects of Methylphenidate on Resting-State Functional Connectivity of the Mesocorticolimbic...
12
Effects of Methylphenidate on Resting-State Functional Connectivity of the Mesocorticolimbic Dopamine Pathways in Cocaine Addiction Anna B. Konova, MA; Scott J. Moeller, PhD; Dardo Tomasi, PhD; Nora D. Volkow, MD; Rita Z. Goldstein, PhD IMPORTANCE Cocaine addiction is associated with altered resting-state functional connectivity among regions of the mesocorticolimbic dopamine pathways. Methylphenidate hydrochloride, an indirect dopamine agonist, normalizes task-related regional brain activity and associated behavior in cocaine users; however, the neural systems–level effects of methylphenidate in this population have not yet been described. OBJECTIVE To use resting-state functional magnetic resonance imaging to examine changes in mesocorticolimbic connectivity with methylphenidate and how connectivity of affected pathways relates to severity of cocaine addiction. DESIGN Randomized, placebo-controlled, before-after, crossover study. SETTING Clinical research center. PARTICIPANTS Eighteen nonabstaining individuals with cocaine use disorders. INTERVENTIONS Single doses of oral methylphenidate (20 mg) or placebo were administered at each of 2 study sessions. At each session, resting scans were acquired twice: immediately after drug administration (before the onset of effects [baseline]) and 120 minutes later (within the window of peak effects). MAIN OUTCOMES AND MEASURES Functional connectivity strength was evaluated using a seed voxel correlation approach. Changes in this measure were examined to characterize the neural systems–level effects of methylphenidate; severity of cocaine addiction was assessed by interview and questionnaire. RESULTS Short-term methylphenidate administration reduced an abnormally strong connectivity of the ventral striatum with the dorsal striatum (putamen/globus pallidus), and lower connectivity between these regions during placebo administration uniquely correlated with less severe addiction. In contrast, methylphenidate strengthened several corticolimbic and corticocortical connections. CONCLUSIONS AND RELEVANCE These findings help elucidate the neural systems–level effects of methylphenidate and suggest that short-term methylphenidate can, at least transiently, remodel abnormal circuitry relevant to the pathophysiologic characteristics of cocaine addiction. In particular, the effects of methylphenidate within striatal and cortical pathways constitute a potentially viable mechanism by which methylphenidate could facilitate control of behavior in cocaine addiction. JAMA Psychiatry. 2013;70(8):857-868. doi:10.1001/jamapsychiatry.2013.1129 Published online June 26, 2013. JAMA Psychiatry. 2013;70(8):857-868. doi:10.1001/jamapsychiatry.2013.1129 Published online June 26, 2013. Supplemental content at jamapsychiatry.com Author Affiliations: Departments of Psychiatry and Neuroscience, Icahn School of Medicine at Mount Sinai, New York, New York (Konova, Moeller, Goldstein); Department of Psychology, Stony Brook University, Stony Brook, New York (Konova); National Institute on Alcohol Abuse and Alcoholism, Rockville, Maryland (Tomasi, Volkow); National Institute on Drug Abuse, Bethesda, Maryland (Volkow). Corresponding Author: Rita Z. Goldstein, PhD, Icahn School of Medicine at Mount Sinai, The Leon and Norma Hess Center for Science and Medicine, 1470 Madison Ave., New York, NY 10029 (rita.goldstein @mssm.edu). Research Original Investigation 857 Downloaded From: http://archpsyc.jamanetwork.com/ by a New York University User on 04/16/2015
Transcript of Effects of Methylphenidate on Resting-State Functional Connectivity of the Mesocorticolimbic...

Effects of Methylphenidate on Resting-State FunctionalConnectivity of the Mesocorticolimbic Dopamine Pathwaysin Cocaine AddictionAnna B. Konova, MA; Scott J. Moeller, PhD; Dardo Tomasi, PhD; Nora D. Volkow, MD; Rita Z. Goldstein, PhD
IMPORTANCE Cocaine addiction is associated with altered resting-state functionalconnectivity among regions of the mesocorticolimbic dopamine pathways. Methylphenidatehydrochloride, an indirect dopamine agonist, normalizes task-related regional brain activityand associated behavior in cocaine users; however, the neural systems–level effects ofmethylphenidate in this population have not yet been described.
OBJECTIVE To use resting-state functional magnetic resonance imaging to examine changesin mesocorticolimbic connectivity with methylphenidate and how connectivity of affectedpathways relates to severity of cocaine addiction.
DESIGN Randomized, placebo-controlled, before-after, crossover study.
SETTING Clinical research center.
PARTICIPANTS Eighteen nonabstaining individuals with cocaine use disorders.
INTERVENTIONS Single doses of oral methylphenidate (20 mg) or placebo were administeredat each of 2 study sessions. At each session, resting scans were acquired twice: immediatelyafter drug administration (before the onset of effects [baseline]) and 120 minutes later(within the window of peak effects).
MAIN OUTCOMES AND MEASURES Functional connectivity strength was evaluated using aseed voxel correlation approach. Changes in this measure were examined to characterize theneural systems–level effects of methylphenidate; severity of cocaine addiction was assessedby interview and questionnaire.
RESULTS Short-term methylphenidate administration reduced an abnormally strongconnectivity of the ventral striatum with the dorsal striatum (putamen/globus pallidus), andlower connectivity between these regions during placebo administration uniquely correlatedwith less severe addiction. In contrast, methylphenidate strengthened several corticolimbicand corticocortical connections.
CONCLUSIONS AND RELEVANCE These findings help elucidate the neural systems–level effectsof methylphenidate and suggest that short-term methylphenidate can, at least transiently,remodel abnormal circuitry relevant to the pathophysiologic characteristics of cocaineaddiction. In particular, the effects of methylphenidate within striatal and cortical pathwaysconstitute a potentially viable mechanism by which methylphenidate could facilitate controlof behavior in cocaine addiction.
JAMA Psychiatry. 2013;70(8):857-868. doi:10.1001/jamapsychiatry.2013.1129Published online June 26, 2013.JAMA Psychiatry. 2013;70(8):857-868. doi:10.1001/jamapsychiatry.2013.1129Published online June 26, 2013.
Supplemental content atjamapsychiatry.com
Author Affiliations: Departments ofPsychiatry and Neuroscience, IcahnSchool of Medicine at Mount Sinai,New York, New York (Konova,Moeller, Goldstein); Department ofPsychology, Stony Brook University,Stony Brook, New York (Konova);National Institute on Alcohol Abuseand Alcoholism, Rockville, Maryland(Tomasi, Volkow); National Instituteon Drug Abuse, Bethesda, Maryland(Volkow).
Corresponding Author: Rita Z.Goldstein, PhD, Icahn School ofMedicine at Mount Sinai, The Leonand Norma Hess Center for Scienceand Medicine, 1470 Madison Ave.,New York, NY 10029 ([email protected]).
Research
Original Investigation
857
Downloaded From: http://archpsyc.jamanetwork.com/ by a New York University User on 04/16/2015

R esting-state functional connectivity is a noninvasive andreplicable method for assessing neural circuitry func-tion in neuropsychiatric disorders.1 This method cap-
tures the synchronicity of low-frequency, spontaneous fluc-tuations in blood oxygen level–dependent signals that reflectfluctuations in neuronal activity2 between brain regions in theabsence of external stimulation.3 These synchronous fluctua-tions are confined to gray matter and can be observed for mono-synaptic or polysynaptic anatomic connections.2,4 More im-portant, resting-state connectivity is linked to task-relatedfunctioning of discrete brain regions comprising the samecircuits5 and to corresponding behavior.5,6 Thus, resting-state connectivity can be useful for advancing neural systems–level understanding of the functional and behavioral abnor-malities that characterize neuropsychiatric disorders such asaddiction, with the potential to also serve as a target for thera-peutic interventions.
Perturbations in resting-state connectivity within and be-tween functional brain networks that subserve attentional,emotional, and inhibitory control processes have been ob-served in individuals addicted to nicotine,7,8 opioids,9-11 andcocaine.12,13 Specifically, individuals with cocaine use disor-ders (CUDs) exhibit reduced connectivity of the dorsal fron-toparietal attention network13; in addition, and especially per-tinent to the current study, cocaine users exhibit reducedconnectivity of the mesocorticolimbic dopamine pathways.12
However, thus far it has not been shown whether this abnor-mal connectivity could be modified in individuals with chronic,severe CUD.
Methylphenidate hydrochloride is a psychostimulantwidely used to treat attention-deficit/hyperactivity disorder.Like cocaine, methylphenidate competitively blocks dopa-mine and norepinephrine transporters, thereby increasing ex-tracellular concentration of these neuromodulatoryneurotransmitters.14 However, unlike cocaine, the rate of clear-ance of orally administered methylphenidate from the brainis substantially slower (90-minute half-life compared with 20-minute half-life for cocaine), contributing to its lower abusepotential15 and possible viability as a therapeutic agent in treat-ing cocaine addiction.16 Methylphenidate improves task-related regional brain activation in the dopamine-innervatedventral medial prefrontal and dorsal anterior cingulatecortices17 and corresponding stop signal reaction times inCUD.18 Oral methylphenidate also has been shown19 to attenu-ate changes in glucose metabolism in the lateral orbitofrontaland inferior frontal cortices, hippocampus, and nucleus ac-cumbens when cocaine users are exposed to drug cues.
In the present study, we used a placebo-controlled, before-after, crossover experimental design to examine the effects ofshort-term methylphenidate administration on resting-stateconnectivity in active cocaine users. To our knowledge, theonly study20 that assessed drug-induced changes in resting-state connectivity in individuals with CUD included a smallsample size, used intravenous cocaine, and focused on the sen-sorimotor cortices. We examined mesocorticolimbic connec-tivity using a seed correlation approach based on a more re-cent study12 reporting that cocaine addiction was associatedwith reduced connectivity in several pathways involved in
emotion processing. Given the normalizing effects of meth-ylphenidate on task-related activation and behavior in CUD,17,18
we hypothesized that methylphenidate would strengthen con-nectivity of the medial frontal cortex in pathways underlyingemotion regulation (eg, with amygdala and hippocampus21-23)and inhibitory control (eg, with dorsal and lateral prefrontalcortex18,19). In addition, because methylphenidate attenu-ated brain metabolism following exposure to cocaine cues,19
we expected reduced connectivity between regions underly-ing cue-induced craving (eg, orbitofrontal cortex, ventral anddorsal striatum, and amygdala24-26). Finally, to examinewhether methylphenidate modified connectivity in path-ways directly associated with addictive behavior, we in-spected correlations between connectivity strength and se-verity of cocaine addiction.
MethodsParticipantsParticipants were 18 nontreatment-seeking individuals withCUD, recruited from advertisements in local newspapers andby word of mouth. All participants were right-handed and na-tive English speakers and provided their written consent to par-ticipate in the study in accordance with the Stony Brook Uni-versity institutional review board. Participants were otherwisehealthy and not taking any medications, as ascertained dur-ing a full physical and neurologic examination by a neurolo-gist and a diagnostic interview by a clinical psychologist. Thislatter interview included the Structured Clinical Interview forDSM-IV Axis I disorders (research version27,28) and the Addic-tion Severity Index.29 All participants were currently using co-caine and identified cocaine as their primary drug of choice,meeting criteria for current cocaine dependence (n = 17) orabuse (n = 1) (eAppendix in the Supplement). Current comor-bidities included heroin dependence (n = 1), marijuana abuse(n = 1), alcohol abuse (n = 1), and nicotine dependence (n = 14).Exclusion criteria were (1) history of head trauma or loss of con-sciousness (>30 minutes) or other neurologic disease, (2) ab-normal vital signs at the time of screening, (3) history of ma-jor medical conditions, (4) history of major psychiatric disorder(other than substance abuse or dependence), (5) pregnancy asconfirmed with a urine test in all women, (6) contraindica-tions to the magnetic resonance imaging (MRI) environment,(7) history of glaucoma, and (8) except for cocaine, positiveurine screen results for psychoactive drugs or their metabo-lites (amphetamine or methamphetamine, phencyclidine, ben-zodiazepines, cannabis, opiates, barbiturates, and inhal-ants). Nine participants tested positive for cocaine onmethylphenidate day and 8 tested positive for cocaine on pla-cebo day (see the eAppendix in the Supplement for addi-tional information). Demographic information is reported inTable 1.
Cocaine withdrawal symptoms were assessed prior tomedication administration at each study day with the Co-caine Selective Severity Assessment scale32; participants alsocompleted the Cocaine Craving Questionnaire33 and the Se-verity of Dependence Scale,34 which captures perceived con-
Research Original Investigation Effects of Methylphenidate in Cocaine Addiction
858 JAMA Psychiatry August 2013 Volume 70, Number 8 jamapsychiatry.com
Downloaded From: http://archpsyc.jamanetwork.com/ by a New York University User on 04/16/2015

trol over drug-taking and difficulty with stopping drug use dur-ing the past year. For each participant, the severity of addictionwas quantified as a composite score (average Z value) of theseverity of withdrawal, craving, and dependence as assessedon placebo day and the frequency of use of cocaine in the past30 days as assessed during the clinical interview. The sever-ity of withdrawal and dependence did not differ significantlybetween the study days (P > .11); however, participants re-ported more recent use of cocaine and more severe craving onmethylphenidate day than on placebo day (Table 2).
Study SessionsAt each of the 2 study sessions (conducted at mean [SD],8.9 [4.0] days apart), participants were randomized to re-ceive a single oral dose of methylphenidate (20 mg) or pla-cebo (lactose). This methylphenidate dose has been shown toaffect task-related brain activation and behavior in CUD.17,19
Because the effects of methylphenidate in individuals withCUD are not well characterized (particularly vis-à-vis poten-tial cardiovascular complications), the study was initially per-formed such that only participants were blinded to the admin-
istered challenge (n = 13). Once it became clear that risks wereminimal, we transitioned to double-blind administration(n = 5), with study personnel also blinded to the medication.
The methylphenidate and placebo sessions consisted ofidentical study procedures. Resting scans were acquired twice,shortly after medication administration (before the onset ofdrug effects) and approximately 120 minutes later (aftercompletion of 2 functional MRI tasks, a rewarded drug cue sus-tained attention task36 and a classic color word Stroop task37),within the window of peak methylphenidate effects.38 To in-spect whether methylphenidate affected cardiovascular reac-tivity and mood state, we assessed participants’ heart rate,blood pressure, desire for methylphenidate, and feelings ofbeing alert, anxious, high, or restless throughout the study ses-sions (eAppendix in the Supplement).
Image AcquisitionFunctional MRI was performed (4-T Varian/Siemens MRI scan-ner) using a coronal T2*-weighted single-shot gradient-echo echoplanar imaging sequence (echo time/repetition time, 20/1600milliseconds; 3.125 × 3.125 mm2 in-plane resolution; 4-mm slicethickness; 1-mm gap; 33 coronal slices; 20-cm field of view;64 × 64 matrix size; 90°-flip angle; and 200-kHz bandwidth withramp sampling). Padding was used to minimize motion, and ear-plugs and headphones were used to minimize the influence ofscanner noise on brain activation.39,40 Participants were in-structed to keep their eyes open, lie as still as possible, and re-main awake during the resting scans. No video corroborationthat participants adhered to the instructions could be ob-tained. Each resting scan was approximately 8 minutes in du-ration (representing 311 consecutive data points after the first10 data points were removed to account for signal stabiliza-tion). This length is comparable to that in previous studies ofthis type7,8 and within the recommended range for optimal as-sessment of seed-based connectivity.41 Of 21 participants ini-tially scanned, 3 were excluded because of missing data (n = 2)or signal dropout (n = 1) on at least 1 of the 4 scans.
Table 1. Demographic and Clinical Characteristics of 18 Individuals WithCocaine Use Disorders
Characteristic Data
Sex, No.
Male 16
Female 2
Race, No.
African American 15
Othera 3
Age, mean (SD), y 45.6 (7.3)
Educational level, mean (SD), y 12.9 (1.8)
Verbal IQ, mean (SD)b 91.8 (9.2)
Nonverbal IQ, mean (SD)c 9.3 (3.1)
Socioeconomic status: Hollingshead Index,mean (SD)
35.8 (8.4)
Drug use history
Cigarette smokers, No.
Current or past 14
Nonsmokers 4
Cigarettes per day, mean (SD), No. (12 currentsmokers only)
8.0 (4.2)
Age of onset of cocaine use, mean (SD), y 26.9 (6.3)
Duration of cocaine use, mean (SD), y 15.3 (7.5)
Preferred route of cocaine administration,d No.
Smoking 14
Intranasal 1
Intravenous 1
Days of cocaine use per week, mean (SD),No. in past 30 de 2.7 (2.1)
Abbreviation: WASI, Wechsler Abbreviated Scale of Intelligence.a White, Hispanic, or Asian.b Determined with the Wide Range Achievement Test III–Reading Scale.30
c Determined with the WASI–Matrix Reasoning Scale.31
d Missing data for 2 participants.e Variables used to compute the addiction severity composite score.
Table 2. Daily Baseline Assessment
Characteristic Methylphenidate PlaceboCocaine urine status, No.
Positive 9 8
Negative 9 10
Current cocaine abstinence,mean (SD), d since last usea
5.4 (6.5) 7.8 (9.5)
Depression score, mean (SD)b 6.9 (5.5) 6.1 (4.8)
Withdrawal symptomsc,d 15.9 (10.6) 12.2 (7.8)
Severity of Dependence Scaled,e 7.4 (2.5) 7.3 (2.6)
Cocaine cravingd,f,g 22.0 (13.4) 17.4 (12.7)
a P < .05.b Determined with the Beck Depression Inventory II35 (score range, 0-63).c Determined with the 18-item Cocaine Selective Severity Assessment32 (score
range, 0-126).d Variables used to compute the addiction severity composite score.e Determined with the Severity of Dependence Scale34 (score range, 0-15).f Determined with the 5-item Craving Questionnaire33 (score range, 0-45).g P < .01.
Effects of Methylphenidate in Cocaine Addiction Original Investigation Research
jamapsychiatry.com JAMA Psychiatry August 2013 Volume 70, Number 8 859
Downloaded From: http://archpsyc.jamanetwork.com/ by a New York University User on 04/16/2015
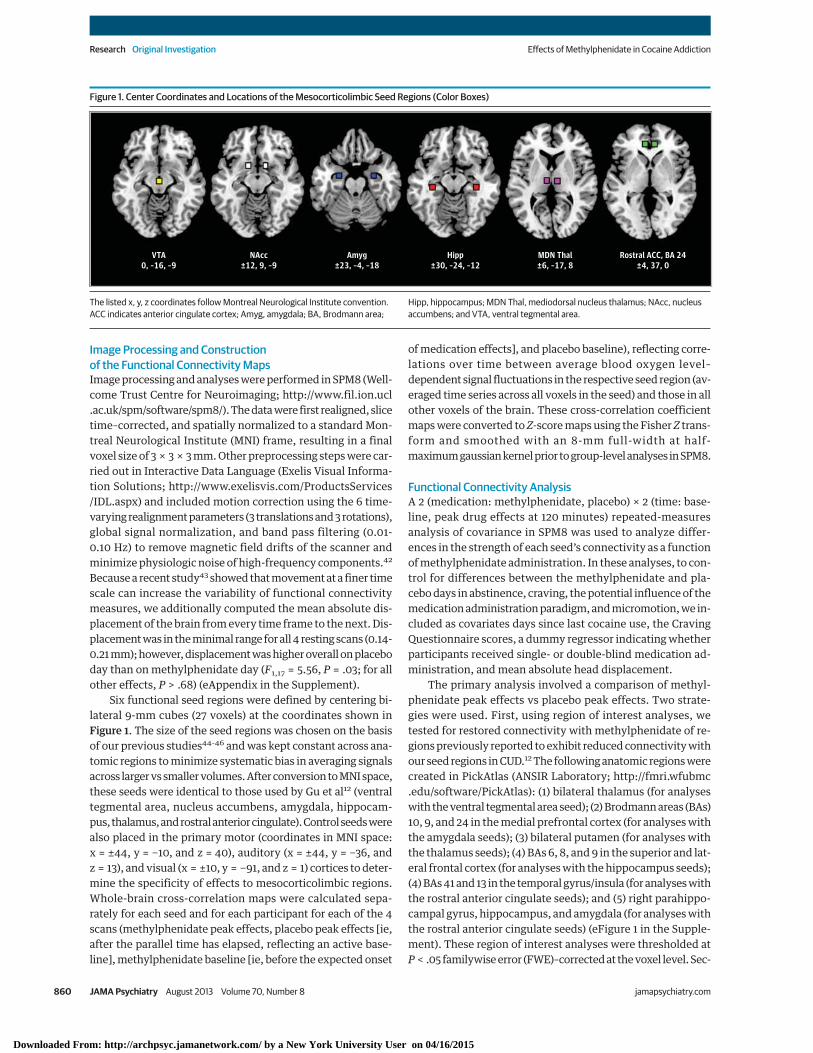
Image Processing and Constructionof the Functional Connectivity MapsImage processing and analyses were performed in SPM8 (Well-come Trust Centre for Neuroimaging; http://www.fil.ion.ucl.ac.uk/spm/software/spm8/). The data were first realigned, slicetime–corrected, and spatially normalized to a standard Mon-treal Neurological Institute (MNI) frame, resulting in a finalvoxel size of 3 × 3 × 3 mm. Other preprocessing steps were car-ried out in Interactive Data Language (Exelis Visual Informa-tion Solutions; http://www.exelisvis.com/ProductsServices/IDL.aspx) and included motion correction using the 6 time-varying realignment parameters (3 translations and 3 rotations),global signal normalization, and band pass filtering (0.01-0.10 Hz) to remove magnetic field drifts of the scanner andminimize physiologic noise of high-frequency components.42
Because a recent study43 showed that movement at a finer timescale can increase the variability of functional connectivitymeasures, we additionally computed the mean absolute dis-placement of the brain from every time frame to the next. Dis-placement was in the minimal range for all 4 resting scans (0.14-0.21 mm); however, displacement was higher overall on placeboday than on methylphenidate day (F1,17 = 5.56, P = .03; for allother effects, P > .68) (eAppendix in the Supplement).
Six functional seed regions were defined by centering bi-lateral 9-mm cubes (27 voxels) at the coordinates shown inFigure 1. The size of the seed regions was chosen on the basisof our previous studies44-46 and was kept constant across ana-tomic regions to minimize systematic bias in averaging signalsacross larger vs smaller volumes. After conversion to MNI space,these seeds were identical to those used by Gu et al12 (ventraltegmental area, nucleus accumbens, amygdala, hippocam-pus, thalamus, and rostral anterior cingulate). Control seeds werealso placed in the primary motor (coordinates in MNI space:x = ±44, y = −10, and z = 40), auditory (x = ±44, y = −36, andz = 13), and visual (x = ±10, y = −91, and z = 1) cortices to deter-mine the specificity of effects to mesocorticolimbic regions.Whole-brain cross-correlation maps were calculated sepa-rately for each seed and for each participant for each of the 4scans (methylphenidate peak effects, placebo peak effects [ie,after the parallel time has elapsed, reflecting an active base-line], methylphenidate baseline [ie, before the expected onset
of medication effects], and placebo baseline), reflecting corre-lations over time between average blood oxygen level–dependent signal fluctuations in the respective seed region (av-eraged time series across all voxels in the seed) and those in allother voxels of the brain. These cross-correlation coefficientmaps were converted to Z-score maps using the Fisher Z trans-form and smoothed with an 8-mm full-width at half-maximum gaussian kernel prior to group-level analyses in SPM8.
Functional Connectivity AnalysisA 2 (medication: methylphenidate, placebo) × 2 (time: base-line, peak drug effects at 120 minutes) repeated-measuresanalysis of covariance in SPM8 was used to analyze differ-ences in the strength of each seed’s connectivity as a functionof methylphenidate administration. In these analyses, to con-trol for differences between the methylphenidate and pla-cebo days in abstinence, craving, the potential influence of themedication administration paradigm, and micromotion, we in-cluded as covariates days since last cocaine use, the CravingQuestionnaire scores, a dummy regressor indicating whetherparticipants received single- or double-blind medication ad-ministration, and mean absolute head displacement.
The primary analysis involved a comparison of methyl-phenidate peak effects vs placebo peak effects. Two strate-gies were used. First, using region of interest analyses, wetested for restored connectivity with methylphenidate of re-gions previously reported to exhibit reduced connectivity withour seed regions in CUD.12 The following anatomic regions werecreated in PickAtlas (ANSIR Laboratory; http://fmri.wfubmc.edu/software/PickAtlas): (1) bilateral thalamus (for analyseswith the ventral tegmental area seed); (2) Brodmann areas (BAs)10, 9, and 24 in the medial prefrontal cortex (for analyses withthe amygdala seeds); (3) bilateral putamen (for analyses withthe thalamus seeds); (4) BAs 6, 8, and 9 in the superior and lat-eral frontal cortex (for analyses with the hippocampus seeds);(4) BAs 41 and 13 in the temporal gyrus/insula (for analyses withthe rostral anterior cingulate seeds); and (5) right parahippo-campal gyrus, hippocampus, and amygdala (for analyses withthe rostral anterior cingulate seeds) (eFigure 1 in the Supple-ment). These region of interest analyses were thresholded atP < .05 familywise error (FWE)–corrected at the voxel level. Sec-
Figure 1. Center Coordinates and Locations of the Mesocorticolimbic Seed Regions (Color Boxes)
VTA0, –16, –9
NAcc±12, 9, –9
Amyg±23, –4, –18
Hipp±30, –24, –12
MDN Thal±6, –17, 8
Rostral ACC, BA 24±4, 37, 0
The listed x, y, z coordinates follow Montreal Neurological Institute convention.ACC indicates anterior cingulate cortex; Amyg, amygdala; BA, Brodmann area;
Hipp, hippocampus; MDN Thal, mediodorsal nucleus thalamus; NAcc, nucleusaccumbens; and VTA, ventral tegmental area.
Research Original Investigation Effects of Methylphenidate in Cocaine Addiction
860 JAMA Psychiatry August 2013 Volume 70, Number 8 jamapsychiatry.com
Downloaded From: http://archpsyc.jamanetwork.com/ by a New York University User on 04/16/2015

ond, we explored whole-brain changes in connectivity usinga cluster-level P < .05 FWE-corrected threshold (see the eAp-pendix in the Supplement). Here, we selected a priori a mini-mum height (P < .005 uncorrected) and cluster extent (20 ad-jacent voxels) threshold. We then applied the FWE correction(at the cluster level) as implemented in SPM8 to determine theprobability of obtaining a given cluster size assuming a ran-dom gaussian field distribution. Significant regions from themethylphenidate vs placebo peak contrast were extracted as3-mm radius spheres, chosen according to the image smooth-ness (ie, the volume of the resolution elements47), using theEasyROI toolbox (http://www.sbirc.ed.ac.uk/LCL/LCL_M1.html). The extracted average signal in these regions was usedfor visual representation of the data, comparison with healthycontrols, and multiple regression analysis with addiction se-verity, described below. Anatomic specificity was deter-mined with the Anatomy toolbox.48
Secondary (control) analyses compared methylphenidatepeak effects vs same-day baseline and methylphenidate same-day baseline vs placebo same-day baseline, both used to ruleout ancillary factors particular to each study session (eg, pre-medication administration cocaine craving and days since lastcocaine use) (Table 2). Results from the control analyses weremasked by the methylphenidate peak effects vs placebo peakeffects contrast (P < .05 uncorrected) and are reported at P < .005uncorrected with a minimum cluster extent of 20 adjacentvoxels.
Finally, we sought to determine whether methylpheni-date modified connectivity in participants with CUD to a levelthat no longer significantly differs from that of healthy indi-viduals. For this purpose, we compared connectivity strengthduring placebo and, separately, during methylphenidate withthat of an independent sample of 16 healthy control partici-pants for whom we acquired resting-state data under placeboconditions (see the eAppendix in the Supplement for addi-tional information).
Association of Connectivity Strength With Severityof Cocaine AddictionTo determine whether methylphenidate modified connectiv-ity between regions that, under active baseline conditions (ie,placebo peak effects), were directly associated with severityof cocaine addiction, we used the composite addiction sever-ity score (Table 2) as the dependent variable in a multiple re-gression analysis in SPSS 18.0 (SPSS, Inc) restricting the pre-dictors to include connectivity measures in regions that bothchanged after methylphenidate and differed significantly fromhealthy controls (either in the present study [eTable 2 in theSupplement] or as reported by Gu et al12) (6 total predictors).
ResultsEffects of Methylphenidate on Cardiovascular Reactivityand Subjective MoodExcept for diastolic blood pressure (which was higher after meth-ylphenidate than after placebo), changes in cardiovascular re-activity and mood did not differ significantly between the meth-
ylphenidate and placebo study sessions (eTable 1 in theSupplement). Consistent with prior studies using 20 mg of oralmethylphenidate,17,19 self-reports of cocaine wanting also didnot differ significantly (eAppendix in the Supplement).
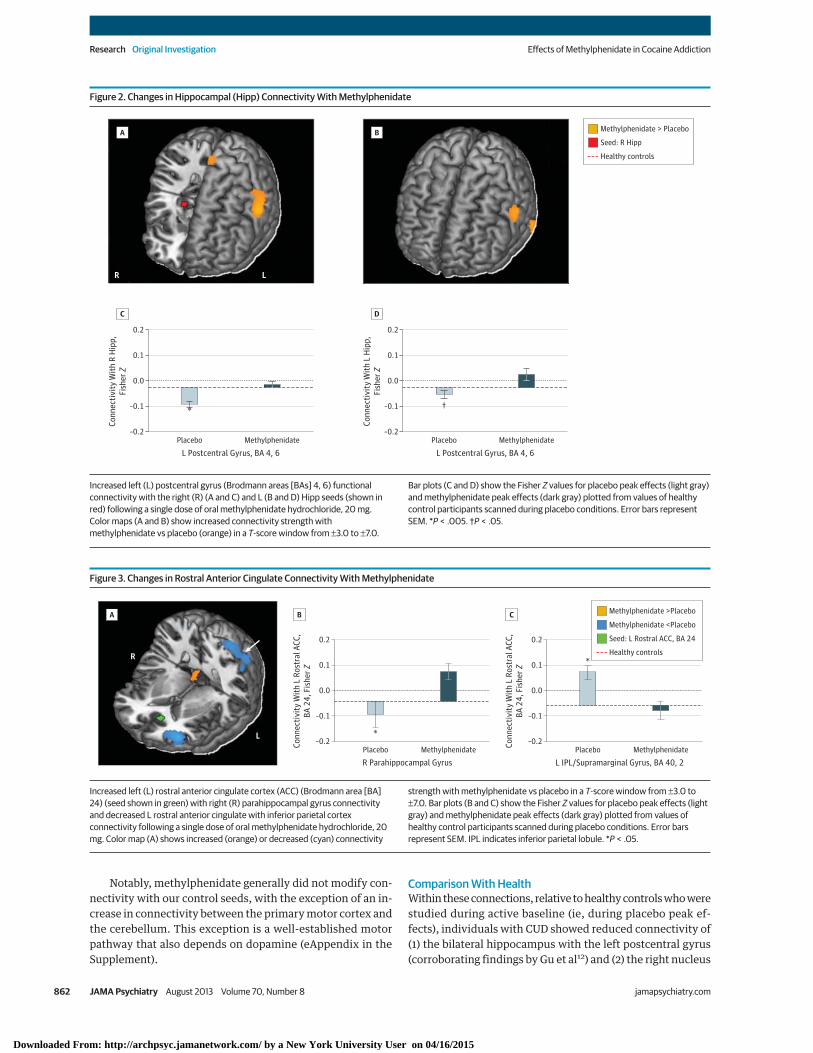
Effects of Methylphenidate on Connectivity StrengthPrimary Analyses: Methylphenidate vs Placebo Peak EffectsThe region of interest analyses indicated that methylpheni-date modified 2 corticolimbic connections that were previ-ously reported to be disrupted in CUD12: compared with pla-cebo, methylphenidate increased the connectivity of (1) theright and left hippocampus with the left postcentral gyrus (BAs4, 6) (peak coordinate in MNI space: x = −60, y = −3, and z = 33,Z = 4.39, FWE-corrected P = .008; and x = −63, y = 3, and z = 30,Z = 4.05, FWE-corrected P = .03, respectively) (Figure 2) and(2) the left rostral anterior cingulate with the right parahippo-campal gyrus (x = 15, y = −36, and z = −9, Z = 3.57, FWE-corrected P = .02) (Figure 3A and B).
Whole-brain analyses revealed additional effects of meth-ylphenidate in connections relevant to self-control and crav-ing (Table 3 and eFigure 2 in the Supplement). Specifically, com-pared with placebo, methylphenidate increased theconnectivity of (1) the right rostral anterior cingulate with theleft dorsal cingulate (eFigure 3 in the Supplement), (2) the rightthalamus with the medial orbitofrontal cortex, and (3) the rightnucleus accumbens with the medial orbitofrontal cortex andright superior temporal gyrus extending to the postcentral gy-rus and rolandic operculum (Figure 4A and C). Methylpheni-date also reduced the connectivity of (1) the ventral tegmen-tal area with the right caudate and putamen; (2) the lefthippocampus with the left insula, thalamus, and putamen; (3)the right thalamus with the bilateral putamen; and (4) the rightnucleus accumbens with the left putamen/globus pallidus(Figure 4A and D). Other methylphenidate-induced changesincluded reduced connectivity of the ventral tegmental area,right hippocampus, and left rostral anterior cingulate with thecerebellum and reduced connectivity of the left rostral ante-rior cingulate (Figure 3C), right hippocampus, and right nucleusaccumbens with the inferior parietal cortex extending to theangular gyrus and precuneus. No significant changes in con-nectivity strength were observed for the bilateral amygdala,left thalamus, or left nucleus accumbens seeds in either theregion of interest or the whole-brain analyses.
Control AnalysesSupporting the idea that these changes in connectivity were dueto the pharmacologic effects of methylphenidate (and not dif-ferences between the study days or the participants’ expecta-tion to receive methylphenidate), baseline connectivity did notdiffer significantly between the 2 study days for any of the seeds(as inspected with the methylphenidate baseline vs placebobaseline contrast, masked by the methylphenidate peak > pla-cebo peak or methylphenidate peak < placebo peak contrastsreported above). The methylphenidate peak effect vs same-day baseline contrast was similarly examined and, in contrast,revealed significant results in most brain regions identified bythe methylphenidate peak vs placebo peak contrast (regionsidentified in both analyses are indicated in Table 3).
Effects of Methylphenidate in Cocaine Addiction Original Investigation Research
jamapsychiatry.com JAMA Psychiatry August 2013 Volume 70, Number 8 861
Downloaded From: http://archpsyc.jamanetwork.com/ by a New York University User on 04/16/2015
Notably, methylphenidate generally did not modify con-nectivity with our control seeds, with the exception of an in-crease in connectivity between the primary motor cortex andthe cerebellum. This exception is a well-established motorpathway that also depends on dopamine (eAppendix in theSupplement).
Comparison With HealthWithin these connections, relative to healthy controls who werestudied during active baseline (ie, during placebo peak ef-fects), individuals with CUD showed reduced connectivity of(1) the bilateral hippocampus with the left postcentral gyrus(corroborating findings by Gu et al12) and (2) the right nucleus
Figure 2. Changes in Hippocampal (Hipp) Connectivity With Methylphenidate
LR
Methylphenidate > Placebo
Seed: R Hipp
Healthy controls
Conn
ectiv
ity W
ith R
Hip
p,Fi
sher
Z
–0.2
–0.1
0.1
0.0
0.2
–0.2
–0.1
0.1
0.0
0.2
L Postcentral Gyrus, BA 4, 6Placebo
*
Methylphenidate
Conn
ectiv
ity W
ith L
Hip
p,Fi
sher
Z
L Postcentral Gyrus, BA 4, 6Placebo
†
Methylphenidate
A
C
B
D
Increased left (L) postcentral gyrus (Brodmann areas [BAs] 4, 6) functionalconnectivity with the right (R) (A and C) and L (B and D) Hipp seeds (shown inred) following a single dose of oral methylphenidate hydrochloride, 20 mg.Color maps (A and B) show increased connectivity strength withmethylphenidate vs placebo (orange) in a T-score window from ±3.0 to ±7.0.
Bar plots (C and D) show the Fisher Z values for placebo peak effects (light gray)and methylphenidate peak effects (dark gray) plotted from values of healthycontrol participants scanned during placebo conditions. Error bars representSEM. *P < .005. †P < .05.
Figure 3. Changes in Rostral Anterior Cingulate Connectivity With Methylphenidate
L
R
–0.2
0.2
Conn
ectiv
ity W
ith L
Ros
tral
ACC
,BA
24,
Fis
her Z
0.0
–0.1
0.1
R Parahippocampal GyrusPlacebo
*
Methylphenidate–0.2
0.1
0.2
Conn
ectiv
ity W
ith L
Ros
tral
ACC
,BA
24,
Fis
her Z
0.0
–0.1
L IPL/Supramarginal Gyrus, BA 40, 2Placebo
*
Methylphenidate
A B C Methylphenidate >Placebo
Seed: L Rostral ACC, BA 24
Methylphenidate <Placebo
Healthy controls
Increased left (L) rostral anterior cingulate cortex (ACC) (Brodmann area [BA]24) (seed shown in green) with right (R) parahippocampal gyrus connectivityand decreased L rostral anterior cingulate with inferior parietal cortexconnectivity following a single dose of oral methylphenidate hydrochloride, 20mg. Color map (A) shows increased (orange) or decreased (cyan) connectivity
strength with methylphenidate vs placebo in a T-score window from ±3.0 to±7.0. Bar plots (B and C) show the Fisher Z values for placebo peak effects (lightgray) and methylphenidate peak effects (dark gray) plotted from values ofhealthy control participants scanned during placebo conditions. Error barsrepresent SEM. IPL indicates inferior parietal lobule. *P < .05.
Research Original Investigation Effects of Methylphenidate in Cocaine Addiction
862 JAMA Psychiatry August 2013 Volume 70, Number 8 jamapsychiatry.com
Downloaded From: http://archpsyc.jamanetwork.com/ by a New York University User on 04/16/2015
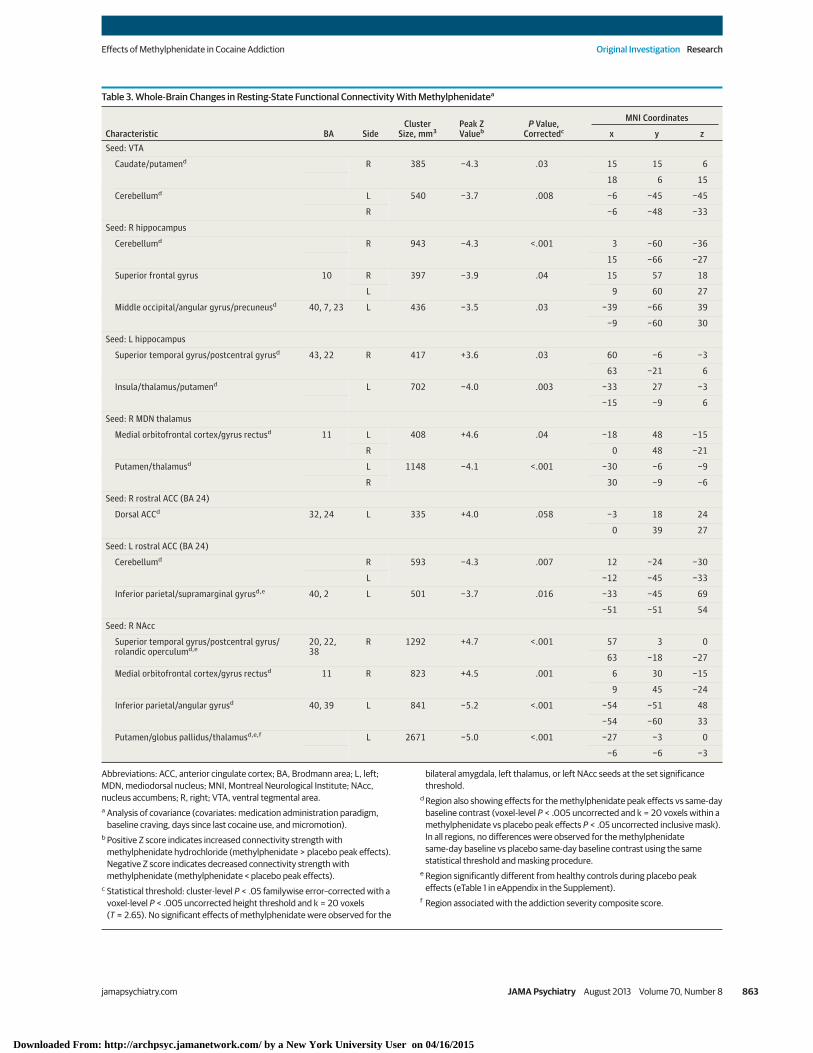
Table 3. Whole-Brain Changes in Resting-State Functional Connectivity With Methylphenidatea
Characteristic BA SideCluster
Size, mm3Peak ZValueb
P Value,Correctedc
MNI Coordinates
x y zSeed: VTA
Caudate/putamend R 385 −4.3 .03 15 15 6
18 6 15
Cerebellumd L 540 −3.7 .008 −6 −45 −45
R −6 −48 −33
Seed: R hippocampus
Cerebellumd R 943 −4.3 <.001 3 −60 −36
15 −66 −27
Superior frontal gyrus 10 R 397 −3.9 .04 15 57 18
L 9 60 27
Middle occipital/angular gyrus/precuneusd 40, 7, 23 L 436 −3.5 .03 −39 −66 39
−9 −60 30
Seed: L hippocampus
Superior temporal gyrus/postcentral gyrusd 43, 22 R 417 +3.6 .03 60 −6 −3
63 −21 6
Insula/thalamus/putamend L 702 −4.0 .003 −33 27 −3
−15 −9 6
Seed: R MDN thalamus
Medial orbitofrontal cortex/gyrus rectusd 11 L 408 +4.6 .04 −18 48 −15
R 0 48 −21
Putamen/thalamusd L 1148 −4.1 <.001 −30 −6 −9
R 30 −9 −6
Seed: R rostral ACC (BA 24)
Dorsal ACCd 32, 24 L 335 +4.0 .058 −3 18 24
0 39 27
Seed: L rostral ACC (BA 24)
Cerebellumd R 593 −4.3 .007 12 −24 −30
L −12 −45 −33
Inferior parietal/supramarginal gyrusd,e 40, 2 L 501 −3.7 .016 −33 −45 69
−51 −51 54
Seed: R NAcc
Superior temporal gyrus/postcentral gyrus/rolandic operculumd,e
20, 22,38
R 1292 +4.7 <.001 57 3 0
63 −18 −27
Medial orbitofrontal cortex/gyrus rectusd 11 R 823 +4.5 .001 6 30 −15
9 45 −24
Inferior parietal/angular gyrusd 40, 39 L 841 −5.2 <.001 −54 −51 48
−54 −60 33
Putamen/globus pallidus/thalamusd,e,f L 2671 −5.0 <.001 −27 −3 0
−6 −6 −3
Abbreviations: ACC, anterior cingulate cortex; BA, Brodmann area; L, left;MDN, mediodorsal nucleus; MNI, Montreal Neurological Institute; NAcc,nucleus accumbens; R, right; VTA, ventral tegmental area.a Analysis of covariance (covariates: medication administration paradigm,
baseline craving, days since last cocaine use, and micromotion).b Positive Z score indicates increased connectivity strength with
methylphenidate hydrochloride (methylphenidate > placebo peak effects).Negative Z score indicates decreased connectivity strength withmethylphenidate (methylphenidate < placebo peak effects).
c Statistical threshold: cluster-level P < .05 familywise error–corrected with avoxel-level P < .005 uncorrected height threshold and k = 20 voxels(T = 2.65). No significant effects of methylphenidate were observed for the
bilateral amygdala, left thalamus, or left NAcc seeds at the set significancethreshold.
d Region also showing effects for the methylphenidate peak effects vs same-daybaseline contrast (voxel-level P < .005 uncorrected and k = 20 voxels within amethylphenidate vs placebo peak effects P < .05 uncorrected inclusive mask).In all regions, no differences were observed for the methylphenidatesame-day baseline vs placebo same-day baseline contrast using the samestatistical threshold and masking procedure.
e Region significantly different from healthy controls during placebo peakeffects (eTable 1 in eAppendix in the Supplement).
f Region associated with the addiction severity composite score.
Effects of Methylphenidate in Cocaine Addiction Original Investigation Research
jamapsychiatry.com JAMA Psychiatry August 2013 Volume 70, Number 8 863
Downloaded From: http://archpsyc.jamanetwork.com/ by a New York University User on 04/16/2015
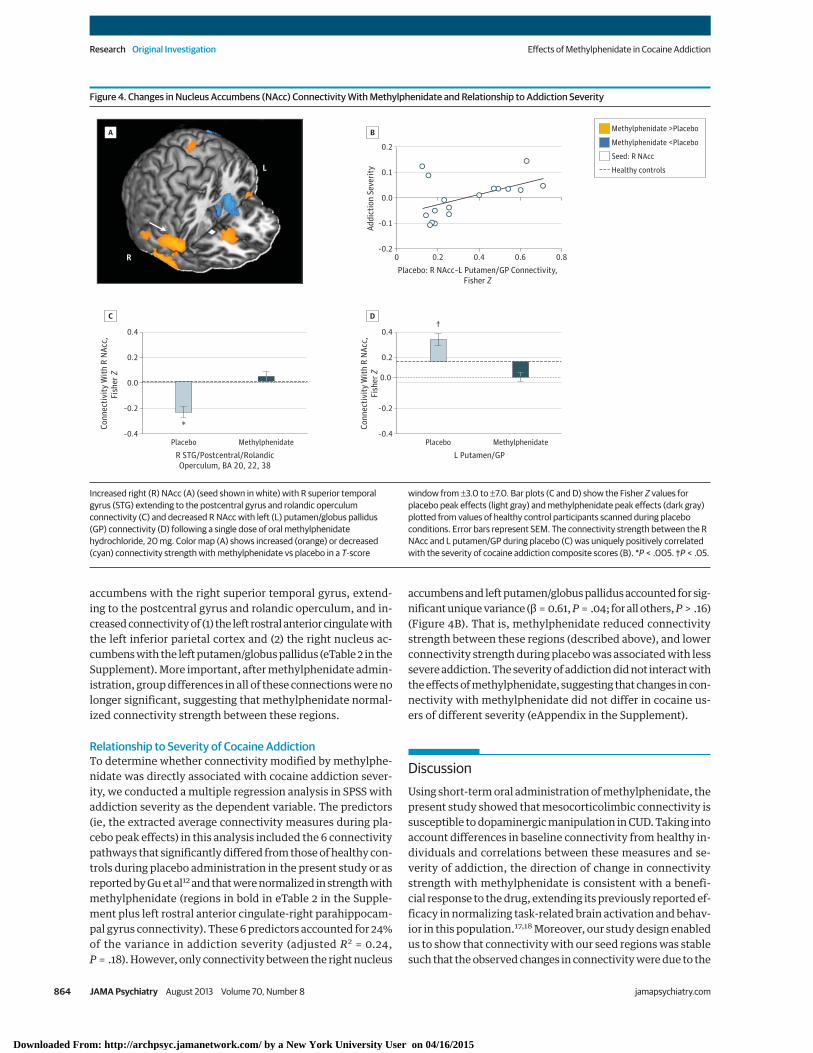
accumbens with the right superior temporal gyrus, extend-ing to the postcentral gyrus and rolandic operculum, and in-creased connectivity of (1) the left rostral anterior cingulate withthe left inferior parietal cortex and (2) the right nucleus ac-cumbens with the left putamen/globus pallidus (eTable 2 in theSupplement). More important, after methylphenidate admin-istration, group differences in all of these connections were nolonger significant, suggesting that methylphenidate normal-ized connectivity strength between these regions.
Relationship to Severity of Cocaine AddictionTo determine whether connectivity modified by methylphe-nidate was directly associated with cocaine addiction sever-ity, we conducted a multiple regression analysis in SPSS withaddiction severity as the dependent variable. The predictors(ie, the extracted average connectivity measures during pla-cebo peak effects) in this analysis included the 6 connectivitypathways that significantly differed from those of healthy con-trols during placebo administration in the present study or asreported by Gu et al12 and that were normalized in strength withmethylphenidate (regions in bold in eTable 2 in the Supple-ment plus left rostral anterior cingulate-right parahippocam-pal gyrus connectivity). These 6 predictors accounted for 24%of the variance in addiction severity (adjusted R2 = 0.24,P = .18). However, only connectivity between the right nucleus
accumbens and left putamen/globus pallidus accounted for sig-nificant unique variance (β = 0.61, P = .04; for all others, P > .16)(Figure 4B). That is, methylphenidate reduced connectivitystrength between these regions (described above), and lowerconnectivity strength during placebo was associated with lesssevere addiction. The severity of addiction did not interact withthe effects of methylphenidate, suggesting that changes in con-nectivity with methylphenidate did not differ in cocaine us-ers of different severity (eAppendix in the Supplement).
DiscussionUsing short-term oral administration of methylphenidate, thepresent study showed that mesocorticolimbic connectivity issusceptible to dopaminergic manipulation in CUD. Taking intoaccount differences in baseline connectivity from healthy in-dividuals and correlations between these measures and se-verity of addiction, the direction of change in connectivitystrength with methylphenidate is consistent with a benefi-cial response to the drug, extending its previously reported ef-ficacy in normalizing task-related brain activation and behav-ior in this population.17,18 Moreover, our study design enabledus to show that connectivity with our seed regions was stablesuch that the observed changes in connectivity were due to the
Figure 4. Changes in Nucleus Accumbens (NAcc) Connectivity With Methylphenidate and Relationship to Addiction Severity
0.0
–0.4
0.4
Conn
ectiv
ity W
ith R
NAc
c,Fi
sher
Z
0.0
–0.2
0.2
R STG/Postcentral/RolandicOperculum, BA 20, 22, 38
Placebo
*
Methylphenidate
L
R
–0.4
0.2
0.4
Conn
ectiv
ity W
ith R
NAc
c,Fi
sher
Z
–0.2
L Putamen/GPPlacebo
†
Methylphenidate
–0.20 0.2 0.4 0.6 0.8
0.1
0.2
Addi
ctio
n Se
verit
y
0.0
–0.1
Placebo: R NAcc–L Putamen/GP Connectivity,Fisher Z
Methylphenidate >Placebo
Seed: R NAcc
Methylphenidate <Placebo
Healthy controls
A
C
B
D
Increased right (R) NAcc (A) (seed shown in white) with R superior temporalgyrus (STG) extending to the postcentral gyrus and rolandic operculumconnectivity (C) and decreased R NAcc with left (L) putamen/globus pallidus(GP) connectivity (D) following a single dose of oral methylphenidatehydrochloride, 20 mg. Color map (A) shows increased (orange) or decreased(cyan) connectivity strength with methylphenidate vs placebo in a T-score
window from ±3.0 to ±7.0. Bar plots (C and D) show the Fisher Z values forplacebo peak effects (light gray) and methylphenidate peak effects (dark gray)plotted from values of healthy control participants scanned during placeboconditions. Error bars represent SEM. The connectivity strength between the RNAcc and L putamen/GP during placebo (C) was uniquely positively correlatedwith the severity of cocaine addiction composite scores (B). *P < .005. †P < .05.
Research Original Investigation Effects of Methylphenidate in Cocaine Addiction
864 JAMA Psychiatry August 2013 Volume 70, Number 8 jamapsychiatry.com
Downloaded From: http://archpsyc.jamanetwork.com/ by a New York University User on 04/16/2015

effects of methylphenidate and not ancillary factors particu-lar to each study session.
Using predefined regions of interest, we found that meth-ylphenidate strengthened connectivity of the bilateral hippo-campus with the postcentral gyrus and of the rostral anteriorcingulate with the parahippocampal gyrus–corticolimbic con-nections suggested to underlie successful emotionregulation21-23 that were reported to be disrupted in cocaineaddiction.12 In particular, the postcentral gyrus is involved incraving suppression,49 and both the postcentral gyrus and thehippocampus fail to normally activate when cocaine users areexposed to stress, abnormalities that could underlie stress-related vulnerability to cocaine relapse.50 Strengthened con-nectivity in emotion processing and memory formation path-ways with methylphenidate may contribute to enhancedretention of emotional associative learning (as has been shownin rodents51,52) and better control over emotional distur-bances and emotional memories (eg, when combined with ex-posure therapy), particularly those associated with with-drawal symptoms and conditioned responses that frequentlylead to relapse in addiction.53
In addition to strengthened corticolimbic connectivity,methylphenidate strengthened connectivity of the rostralanterior cingulate with the dorsal cingulate. These cingulateregions have differential anatomic connections with emo-tion and cognitive control networks,54 and reduced correla-tions have been observed between these regions during pro-cessing of salient cues in cocaine users.55 Therefore,strengthened frontal cortical connectivity with methylphe-nidate may point to the mechanism that contributes to themethylphenidate-induced improvements in behavioral andneural measures of self-control on both neutral18 andsalient17 tasks of executive function in CUD. This finding isalso important in view of tractography studies in CUD, inwhich higher fractional anisotropy in regions of the frontalcortex and the rostral corpus callosum linking these regionspredicts longer abstinence.56 Although methylphenidateincreased connectivity of the rostral with dorsal cingulate,an executive control and attention network region, methyl-phenidate reduced connectivity of the rostral cingulate withthe inferior parietal cortex/supramarginal gyrus, a “defaultmode” region, which may play a role in methylphenidate’sattention-enhancing properties.57
Most notably, methylphenidate reduced the connectiv-ity of several subcortical regions, including the ventral teg-mental area, hippocampus, thalamus, and nucleus accum-bens, with the dorsal striatum. In particular, connectivitywithin striatal circuits, possibly instantiated via the spiral-ing dopamine connections through the midbrain that linkthe nucleus accumbens with the dorsolateral striatum or viaother nodes of the cortico-thalamic-striatal loops,58 isstrongly implicated in drug-seeking.59 Because the progres-sion of cocaine addiction involves a shift in striatal circuitsfrom ventral to dorsal,26,60-62 the strength of connectivitybetween these regions may be marking individual differ-ences in disease severity. Indeed, higher baseline connectiv-ity of the nucleus accumbens with the putamen/globus pal-lidus uniquely correlated with more severe addiction. The
finding that short-term methylphenidate can modify thisconnection may be clinically relevant given that blockingstriato-midbrain-striatal serial connectivity selectivelydecreased drug-seeking in rats trained to habitually self-administer cocaine.59 Future longer-term intervention stud-ies should test whether systematic, prolonged weakening ofthis connection helps restore control over drug-seekingbehavior in humans. Other effects of methylphenidate mayalso contribute to increased control over craving and drug-seeking, including methylphenidate-strengthened connec-tivity of the nucleus accumbens with a region encompass-ing the superior temporal and postcentral gyri and therolandic operculum. Indeed, cognitive control of craving isassociated with inverse coupling between these corticalregions and the nucleus accumbens in addicted individualsusing different strategies to resist craving.63,64
Although not hypothesized a priori or identified as abnor-mal relative to healthy controls, connectivity in other path-ways modified by methylphenidate also may be relevant to ad-diction; further research is needed to clarify the precisemechanisms and consequences of these changes. For ex-ample, because the locus ceruleus, which is in close proximityto our ventral tegmental area seed, is the main source of nor-adrenergic innervation to the cerebellum,65 connectivity re-ductions in this pathway could be the result of differential ef-fects of methylphenidate on norepinephrine. Other effects mayalso have noradrenergic underpinnings, including methylphe-nidate-strengthened connectivity of the thalamus with the me-dial orbitofrontal cortex, potentially signaling normalization ofnoradrenergic deficits in this region in cocaine abusers.66
Several caveats should be considered when interpretingthe current results. First, like most stimulants, methylpheni-date produced cardiovascular changes that differed from thosewith placebo. However, because blood oxygen level–dependent fluctuations correlating with heart rate and respi-ration are global,67,68 these changes are not likely to signifi-cantly influence our results. At the neurochemical level, asecond concern is that the effects of methylphenidate are notspecific to dopamine because methylphenidate also blocks thenorepinephrine transporter69 (and may account for some of itseffects, as discussed above). Nevertheless, even if effects arethe result of norepinephrine transporter blockade, the under-lying mechanism could still be dopaminergic, since dopa-mine also has high affinity for norepinephrine transporters—particularly in regions where norepinephrine transporters aremore abundant than dopamine transporters, such as the fron-tal cortex.70 Third, similar to prior studies, we detected a num-ber of negative relationships with our seed regions. One con-cern that arises is the possible contribution of analyticalprocedures (eg, global signal normalization).41,71,72 However,prior work suggests that negative relationships cannot sim-ply be attributed to correction for the global signal73 and thatremoving global signal from the data, which is primarily lo-calized to gray matter, can actually improve neuroanatomicspecificity of positive relationships.74 Nevertheless, becauseour contrast of interest was a within-subject change in con-nectivity, this issue is unlikely to modify our conclusions.Fourth, although seed-based methods for connectivity are one
Effects of Methylphenidate in Cocaine Addiction Original Investigation Research
jamapsychiatry.com JAMA Psychiatry August 2013 Volume 70, Number 8 865
Downloaded From: http://archpsyc.jamanetwork.com/ by a New York University User on 04/16/2015

of the most commonly used approaches, in part because thesemethods are reliable75 and especially appropriate when re-searchers have specific a priori hypotheses,76 they are lim-ited by (1) the univariate nature of the analysis, which limitsthe scope of network-level conclusions that can be drawn,76
and (2) susceptibility to biases related to variations in seedpositioning.76,77 We addressed this latter limitation by usingseeds identical to those used in previous research.12 Futurestudies could also apply complementary methods (eg, thosebased on graph theory42 or independent component analy-sis) to capture the effects of methylphenidate on brain net-works. Last, we cannot at present speak to whether methyl-phenidate-induced modulation of connectivity is a viable targetfor treatment approaches in CUD. Future studies would needto examine the effects of methylphenidate using dose-dependent and/or prolonged administration (ie, occurring overdays or weeks). In addition, because methylphenidate as astand-alone treatment may be insufficient to achieve a posi-tive clinical outcome in cocaine addiction,78,79 except for in in-stances of comorbidity with attention-deficit/hyperactivitydisorder,80 the therapeutic effects of methylphenidate shouldbe explored in conjunction with behavioral interventions tar-geted toward increasing emotion regulation and self-controland decreasing conditioned associations.
In summary, our findings provide novel evidence that me-socorticolimbic connectivity is susceptible to modification bypharmacologic agents targeting dopamine in individuals withchronic, severe CUD. Methylphenidate primarily strength-ened connections between regions underlying emotionregulation and cognitive control and reduced connections be-tween regions underlying habits, including compulsive drug-seeking and craving. Although the precise mechanism of theseeffects remains to be determined, our data suggest that meth-ylphenidate may transiently, and independently of task de-mands, modify striatal and cortical synchronous activity withconnected brain regions. These changes could serve to facili-tate behavior or make cortical processing underlying behav-ior more efficient57,81-83 or less difficult to override.84 By high-lighting effects in mesocorticolimbic pathways in CUD, ourresults extend prior work that has shown pharmacologic modu-lation of default mode and executive control network cir-cuitry in nicotine dependence (via nicotine patch7,8) and lim-bic circuitry in depression (via the antidepressant sertraline).A better understanding of the potentially therapeutic effectsof methylphenidate on neural circuitry function in CUD (eg,with future studies evaluating the clinical efficacy of methyl-phenidate) may promote the development of improved treat-ment options for stimulant addictions.
ARTICLE INFORMATION
Submitted for Publication: June 14, 2012; finalrevision received September 26, 2012; acceptedNovember 9, 2012.
Published Online: June 26, 2013.doi:10.1001/jamapsychiatry.2013.1129.
Author Contributions: Ms Konova and Drs Moeller,Tomasi, and Goldstein had full access to all the datain the study and take responsibility for the integrityof the data and the accuracy of the data analysis.Study concept and design: Tomasi, Volkow,Goldstein.Acquisition of data: Tomasi.Analysis and interpretation of data: Konova,Moeller, Tomasi, Goldstein.Drafting of the manuscript: Konova, Moeller.Critical revision of the manuscript for importantintellectual content: All authors.Statistical analysis: Konova, Moeller, Tomasi.Obtained funding: Goldstein.Administrative, technical, and material support:Tomasi, Volkow, Goldstein.Study supervision: Tomasi, Volkow, Goldstein.
Conflict of Interest Disclosures: None reported.
Funding/Support: This research was conductedwith grant support from the National Institute onDrug Abuse (1R01DA023579 to Dr Goldstein and1F32DA030017-01 to Dr Moeller).
Additional Contributions: Nelly Alia-Klein, PhD;Daniel Carrero, MS; Tom Maloney, PhD; MuhammadA. Parvaz, PhD; Patricia A. Woicik, PhD; Gene-JackWang, MD; Frank Telang, MD; Barbara Hubbard,RN; and the entire Neuropsychoimaging and PETgroup provided assistance with participantrecruitment and study procedures.
REFERENCES
1. Rosazza C, Minati L. Resting-state brainnetworks: literature review and clinical applications.Neurol Sci. 2011;32(5):773-785.
2. Shmuel A, Leopold DA. Neuronal correlates ofspontaneous fluctuations in fMRI signals in monkeyvisual cortex: implications for functionalconnectivity at rest. Hum Brain Mapp. 2008;29(7):751-761.
3. Fox MD, Raichle ME. Spontaneous fluctuations inbrain activity observed with functional magneticresonance imaging. Nat Rev Neurosci. 2007;8(9):700-711.
4. Damoiseaux JS, Greicius MD. Greater than thesum of its parts: a review of studies combiningstructural connectivity and resting-state functionalconnectivity. Brain Struct Funct. 2009;213(6):525-533.
5. Hampson M, Driesen NR, Skudlarski P, Gore JC,Constable RT. Brain connectivity related to workingmemory performance. J Neurosci. 2006;26(51):13338-13343.
6. Kelly AM, Uddin LQ, Biswal BB, Castellanos FX,Milham MP. Competition between functional brainnetworks mediates behavioral variability.Neuroimage. 2008;39(1):527-537.
7. Cole DM, Beckmann CF, Long CJ, Matthews PM,Durcan MJ, Beaver JD. Nicotine replacement inabstinent smokers improves cognitive withdrawalsymptoms with modulation of resting brainnetwork dynamics. Neuroimage. 2010;52(2):590-599.
8. Hong LE, Gu H, Yang Y, et al. Association ofnicotine addiction and nicotine’s actions withseparate cingulate cortex functional circuits. ArchGen Psychiatry. 2009;66(4):431-441.
9. Ma N, Liu Y, Li N, et al. Addiction relatedalteration in resting-state brain connectivity.Neuroimage. 2010;49(1):738-744.
10. Upadhyay J, Maleki N, Potter J, et al. Alterationsin brain structure and functional connectivity inprescription opioid-dependent patients. Brain.2010;133(pt 7):2098-2114.
11. Ma N, Liu Y, Fu XM, et al. Abnormal braindefault-mode network functional connectivity indrug addicts. PLoS One. 2011;6(1):e16560.doi:10.1371/journal.pone.0016560.
12. Gu H, Salmeron BJ, Ross TJ, et al.Mesocorticolimbic circuits are impaired in chroniccocaine users as demonstrated by resting-statefunctional connectivity. Neuroimage. 2010;53(2):593-601.
13. Kelly C, Zuo XN, Gotimer K, et al. Reducedinterhemispheric resting state functionalconnectivity in cocaine addiction. Biol Psychiatry.2011;69(7):684-692.
14. Kuczenski R, Segal DS. Effects ofmethylphenidate on extracellular dopamine,serotonin, and norepinephrine: comparison withamphetamine. J Neurochem. 1997;68(5):2032-2037.
15. Volkow ND, Fowler JS, Wang GJ, Swanson JM.Dopamine in drug abuse and addiction: results fromimaging studies and treatment implications. MolPsychiatry. 2004;9(6):557-569.
16. Levin FR, Evans SM, Brooks DJ, Garawi F.Treatment of cocaine dependent treatment seekerswith adult ADHD: double-blind comparison ofmethylphenidate and placebo. Drug AlcoholDepend. 2007;87(1):20-29.
17. Goldstein RZ, Woicik PA, Maloney T, et al. Oralmethylphenidate normalizes cingulate activity incocaine addiction during a salient cognitive task.
Research Original Investigation Effects of Methylphenidate in Cocaine Addiction
866 JAMA Psychiatry August 2013 Volume 70, Number 8 jamapsychiatry.com
Downloaded From: http://archpsyc.jamanetwork.com/ by a New York University User on 04/16/2015

Proc Natl Acad Sci U S A. 2010;107(38):16667-16672.
18. Li CS, Morgan PT, Matuskey D, et al. Biologicalmarkers of the effects of intravenousmethylphenidate on improving inhibitory control incocaine-dependent patients. Proc Natl Acad Sci U SA. 2010;107(32):14455-14459.
19. Volkow ND, Wang GJ, Tomasi D, et al.Methylphenidate attenuates limbic brain inhibitionafter cocaine-cues exposure in cocaine abusers.PLoS One. 2010;5(7):e11509. doi:10.1371/journal.pone.0011509.
20. Li SJ, Biswal B, Li Z, et al. Cocaineadministration decreases functional connectivity inhuman primary visual and motor cortex as detectedby functional MRI. Magn Reson Med. 2000;43(1):45-51.
21. Diekhof EK, Geier K, Falkai P, Gruber O. Fear isonly as deep as the mind allows: a coordinate-basedmeta-analysis of neuroimaging studies on theregulation of negative affect. Neuroimage.2011;58(1):275-285.
22. Mak AK, Hu ZG, Zhang JX, Xiao ZW, Lee TM.Neural correlates of regulation of positive andnegative emotions: an fMRI study. Neurosci Lett.2009;457(2):101-106.
23. Kanske P, Heissler J, Schönfelder S, Bongers A,Wessa M. How to regulate emotion? neuralnetworks for reappraisal and distraction. CerebCortex. 2011;21(6):1379-1388.
24. Chase HW, Eickhoff SB, Laird AR, Hogarth L.The neural basis of drug stimulus processing andcraving: an activation likelihood estimationmeta-analysis. Biol Psychiatry. 2011;70(8):785-793.
25. Kühn S, Gallinat J. Common biology of cravingacross legal and illegal drugs—a quantitativemeta-analysis of cue-reactivity brain response. Eur JNeurosci. 2011;33(7):1318-1326.
26. Volkow ND, Wang GJ, Telang F, et al. Cocainecues and dopamine in dorsal striatum: mechanismof craving in cocaine addiction. J Neurosci.2006;26(24):6583-6588.
27. First MB, Spitzer RL, Gibbon M, Williams J.Structured Clinical Interview for DSM-IV Axis IDisorders—Patient Edition (SCID-I/P, Version 2.0).New York: Biometrics Research Dept, New YorkState Psychiatric Institute; 1996.
28. Ventura J, Liberman RP, Green MF, Shaner A,Mintz J. Training and quality assurance with theStructured Clinical Interview for DSM-IV (SCID-I/P).Psychiatry Res. 1998;79(2):163-173.
29. McLellan AT, Kushner H, Metzger D, et al. Thefifth edition of the Addiction Severity Index. J SubstAbuse Treat. 1992;9(3):199-213.
30. Wilkinson G. The Wide-Range Achievement Test3—Administration Manual. Wilmington, DE: WideRange Inc; 1993.
31. Wechsler D. Wechsler Abbreviated Scale ofIntelligence. San Antonio, TX: Psychological Corp;1999.
32. Kampman KM, Volpicelli JR, McGinnis DE, et al.Reliability and validity of the Cocaine SelectiveSeverity Assessment. Addict Behav. 1998;23(4):449-461.
33. Tiffany ST, Singleton E, Haertzen CA,Henningfield JE. The development of a cocainecraving questionnaire. Drug Alcohol Depend.1993;34(1):19-28.
34. Gossop M, Griffiths P, Powis B, Strang J.Severity of dependence and route of administrationof heroin, cocaine and amphetamines. Br JAddiction. 1992;87(11):1527-1536.
35. Beck AT, Steer RA, Brown GK. Beck DepressionInventory Manual. 2nd ed. San Antonio, TX:Psychological Corp; 1996.
36. Goldstein RZ, Tomasi D, Rajaram S, et al. Role ofthe anterior cingulate and medial orbitofrontalcortex in processing drug cues in cocaine addiction.Neuroscience. 2007;144(4):1153-1159.
37. Moeller S, Tomasi D, Honorio J, Volkow ND,Goldstein RZ. Dopaminergic involvement duringmental fatigue in health and cocaine addiction.Transl Psychiatr. 2012;2:e176. doi:10.1038/tp.2012.110.
38. Volkow ND, Wang GJ, Fowler JS, et al.Dopamine transporter occupancies in the humanbrain induced by therapeutic doses of oralmethylphenidate. Am J Psychiatry. 1998;155(10):1325-1331.
39. Tomasi D, Caparelli EC, Chang L, Ernst T.fMRI-acoustic noise alters brain activation duringworking memory tasks. Neuroimage. 2005;27(2):377-386.
40. Tomasi DG, Ernst T. Echo planar imaging at 4Tesla with minimum acoustic noise. J Magn ResonImaging. 2003;18(1):128-130.
41. Van Dijk KR, Hedden T, Venkataraman A, EvansKC, Lazar SW, Buckner RL. Intrinsic functionalconnectivity as a tool for human connectomics:theory, properties, and optimization.J Neurophysiol. 2010;103(1):297-321.
42. Tomasi D, Volkow ND. Functional connectivitydensity mapping. Proc Natl Acad Sci U S A.2010;107(21):9885-9890.
43. Van Dijk KR, Sabuncu MR, Buckner RL. Theinfluence of head motion on intrinsic functionalconnectivity MRI. Neuroimage. 2012;59(1):431-438.
44. Tomasi D, Volkow ND. Language network:segregation, laterality and connectivity. MolPsychiatry. 2012;17(8):759. doi:10.1038/mp.2012.99.
45. Tomasi D, Volkow ND. Abnormal functionalconnectivity in children with attention-deficit/hyperactivity disorder. Biol Psychiatry. 2012;71(5):443-450.
46. Tomasi D, Volkow ND, Wang R, et al. Disruptedfunctional connectivity with dopaminergic midbrainin cocaine abusers. PLoS One. 2010;5(5):e10815.doi:10.1371/journal.pone.0010815.
47. Worsley KJ, Evans AC, Marrett S, Neelin P. Athree-dimensional statistical analysis for CBFactivation studies in human brain. J Cereb BloodFlow Metab. 1992;12(6):900-918.
48. Eickhoff SB, Stephan KE, Mohlberg H, et al. Anew SPM toolbox for combining probabilisticcytoarchitectonic maps and functional imagingdata. Neuroimage. 2005;25(4):1325-1335.
49. Brody AL, Mandelkern MA, Olmstead RE, et al.Neural substrates of resisting craving duringcigarette cue exposure. Biol Psychiatry.2007;62(6):642-651.
50. Sinha R, Lacadie C, Skudlarski P, et al. Neuralactivity associated with stress-induced cocainecraving: a functional magnetic resonance imagingstudy. Psychopharmacology (Berl). 2005;183(2):171-180.
51. Bethancourt JA, Camarena ZZ, Britton GB.Exposure to oral methylphenidate fromadolescence through young adulthood producestransient effects on hippocampal-sensitive memoryin rats. Behav Brain Res. 2009;202(1):50-57.
52. Abraham AD, Cunningham CL, Lattal KM.Methylphenidate enhances extinction of contextualfear. Learn Mem. 2012;19(2):67-72.
53. Koob GF, Le Moal M. Addiction and the brainantireward system. Annu Rev Psychol. 2008;59:29-53.
54. Margulies DS, Kelly AM, Uddin LQ, Biswal BB,Castellanos FX, Milham MP. Mapping the functionalconnectivity of anterior cingulate cortex.Neuroimage. 2007;37(2):579-588.
55. Goldstein RZ, Alia-Klein N, Tomasi D, et al.Anterior cingulate cortex hypoactivations to anemotionally salient task in cocaine addiction. ProcNatl Acad Sci U S A. 2009;106(23):9453-9458.
56. Xu J, DeVito EE, Worhunsky PD, Carroll KM,Rounsaville BJ, Potenza MN. White matter integrityis associated with treatment outcome measures incocaine dependence. Neuropsychopharmacology.2010;35(7):1541-1549.
57. Tomasi D, Volkow ND, Wang GJ, et al.Methylphenidate enhances brain activation anddeactivation responses to visual attention andworking memory tasks in healthy controls.Neuroimage. 2011;54(4):3101-3110.
58. Haber SN, Fudge JL, McFarland NR.Striatonigrostriatal pathways in primates form anascending spiral from the shell to the dorsolateralstriatum. J Neurosci. 2000;20(6):2369-2382.
59. Belin D, Everitt BJ. Cocaine seeking habitsdepend upon dopamine-dependent serialconnectivity linking the ventral with the dorsalstriatum. Neuron. 2008;57(3):432-441.
60. Porrino LJ, Smith HR, Nader MA, Beveridge TJ.The effects of cocaine: a shifting target over thecourse of addiction. Prog NeuropsychopharmacolBiol Psychiatry. 2007;31(8):1593-1600.
61. Everitt BJ, Belin D, Economidou D, Pelloux Y,Dalley JW, Robbins TW. Neural mechanismsunderlying the vulnerability to develop compulsivedrug-seeking habits and addiction. Philos Trans RSoc Lond B Biol Sci. 2008;363(1507):3125-3135.
62. Everitt BJ, Robbins TW. Neural systems ofreinforcement for drug addiction: from actions tohabits to compulsion. Nat Neurosci. 2005;8(11):1481-1489.
63. Volkow ND, Fowler JS, Wang GJ, et al.Cognitive control of drug craving inhibits brainreward regions in cocaine abusers. Neuroimage.2010;49(3):2536-2543.
64. Kober H, Mende-Siedlecki P, Kross EF, et al.Prefrontal-striatal pathway underlies cognitiveregulation of craving. Proc Natl Acad Sci U S A.2010;107(33):14811-14816.
65. Rogawski MA, Aghajanian GK. Modulation oflateral geniculate neurone excitability bynoradrenaline microiontophoresis or locuscoeruleus stimulation. Nature. 1980;287(5784):731-734.
66. Ding YS, Singhal T, Planeta-Wilson B, et al. PETimaging of the effects of age and cocaine on thenorepinephrine transporter in the human brainusing (S,S)-[11C]O-methylreboxetine and HRRT.Synapse. 2010;64(1):30-38.
Effects of Methylphenidate in Cocaine Addiction Original Investigation Research
jamapsychiatry.com JAMA Psychiatry August 2013 Volume 70, Number 8 867
Downloaded From: http://archpsyc.jamanetwork.com/ by a New York University User on 04/16/2015

67. Birn RM, Diamond JB, Smith MA, BandettiniPA. Separating respiratory-variation-relatedfluctuations from neuronal-activity-relatedfluctuations in fMRI. Neuroimage. 2006;31(4):1536-1548.
68. Chang C, Glover GH. Effects of model-basedphysiological noise correction on default modenetwork anti-correlations and correlations.Neuroimage. 2009;47(4):1448-1459.
69. Hannestad J, Gallezot JD, Planeta-Wilson B,et al. Clinically relevant doses of methylphenidatesignificantly occupy norepinephrine transporters inhumans in vivo. Biol Psychiatry. 2010;68(9):854-860.
70. Morón JA, Brockington A, Wise RA, Rocha BA,Hope BT. Dopamine uptake through thenorepinephrine transporter in brain regions withlow levels of the dopamine transporter: evidencefrom knock-out mouse lines. J Neurosci.2002;22(2):389-395.
71. Murphy K, Birn RM, Handwerker DA, Jones TB,Bandettini PA. The impact of global signalregression on resting state correlations: areanti-correlated networks introduced? Neuroimage.2009;44(3):893-905.
72. Saad ZS, Gotts SJ, Murphy K, et al. Trouble atrest: how correlation patterns and groupdifferences become distorted after global signalregression. Brain Connect. 2012;2(1):25-32.
73. Fransson P. Spontaneous low-frequency BOLDsignal fluctuations: an fMRI investigation of theresting-state default mode of brain functionhypothesis. Hum Brain Mapp. 2005;26(1):15-29.
74. Fox MD, Zhang D, Snyder AZ, Raichle ME. Theglobal signal and observed anticorrelated restingstate brain networks. J Neurophysiol. 2009;101(6):3270-3283.
75. Shehzad Z, Kelly AM, Reiss PT, et al. The restingbrain: unconstrained yet reliable. Cereb Cortex.2009;19(10):2209-2229.
76. Cole DM, Smith SM, Beckmann CF. Advancesand pitfalls in the analysis and interpretation ofresting-state FMRI data. Front Syst Neurosci.2010;4:8. doi:10.3389/fnsys.2010.00008.
77. Buckner RL, Andrews-Hanna JR, Schacter DL.The brain’s default network: anatomy, function, andrelevance to disease. Ann N Y Acad Sci. 2008;1124:1-38.
78. Gawin F, Riordan C, Kleber H. Methylphenidatetreatment of cocaine abusers without attentiondeficit disorder: a negative report. Am J DrugAlcohol Abuse. 1985;11(3-4):193-197.
79. Grabowski J, Roache JD, Schmitz JM, RhoadesH, Creson D, Korszun A. Replacement medicationfor cocaine dependence: methylphenidate. J ClinPsychopharmacol. 1997;17(6):485-488.
80. Collins SL, Levin FR, Foltin RW, Kleber HD,Evans SM. Response to cocaine, alone and in
combination with methylphenidate, in cocaineabusers with ADHD. Drug Alcohol Depend.2006;82(2):158-167.
81. Volkow ND, Fowler JS, Wang GJ, et al.Methylphenidate decreased the amount of glucoseneeded by the brain to perform a cognitive task.PLoS One. 2008;3(4):e2017. doi:10.1371/journal.pone.0002017.
82. Marquand AF, De Simoni S, O’Daly OG, WilliamsSC, Mourão-Miranda J, Mehta MA. Patternclassification of working memory networks revealsdifferential effects of methylphenidate,atomoxetine, and placebo in healthy volunteers.Neuropsychopharmacology. 2011;36(6):1237-1247.
83. Mehta MA, Owen AM, Sahakian BJ, MavaddatN, Pickard JD, Robbins TW. Methylphenidateenhances working memory by modulating discretefrontal and parietal lobe regions in the human brain.J Neurosci. 2000;20(6):RC65.
84. Epstein JN, Casey BJ, Tonev ST, et al. ADHD-and medication-related brain activation effects inconcordantly affected parent-child dyads withADHD. J Child Psychol Psychiatry. 2007;48(9):899-913.
Research Original Investigation Effects of Methylphenidate in Cocaine Addiction
868 JAMA Psychiatry August 2013 Volume 70, Number 8 jamapsychiatry.com
Downloaded From: http://archpsyc.jamanetwork.com/ by a New York University User on 04/16/2015




















