
Effects of in ovo feeding of carbohydrates and beta-hydroxy-beta-methylbutyrate on the development...
6
Effects of In Ovo Feeding of Carbohydrates and β-Hydroxy-β-Methylbutyrate on the Development of Chicken Intestine 1 E. Tako,* P. R. Ferket,† and Z. Uni* ,2 *The Faculty of Agricultural, Food and Environmental Quality Sciences, the Department of Animal Sciences, Hebrew University of Jerusalem, PO Box 12, Rehovot, 76100, Isarel; and †North Carolina State University, Department of Poultry Science, College of Agriculture and Life Sciences, Raleigh, North Carolina 27695-7608 ABSTRACT Early development of the digestive tract is crucial for achieving maximal growth and development of chickens. Because the late-term embryo naturally con- sumes the amniotic fluids, insertion of a nutrient solution into the embryonic amniotic fluid [in ovo (IO) feeding] may enhance development. This study examined the ef- fect of IO feeding on d 17.5 of incubation of carbohydrates (CHO) and β-hydroxy-β-methylbutyrate (HMB) on small intestinal development of chickens during the pre and posthatch periods. Results shows that 48 h post-IO feed- ing procedure all IO feeding treatments exhibited in- creased villus width and surface area compared with the control group. At d 3 posthatch the surface area of an average villi was increased by 45% for the HMB IO group and by 33% for the CHO and CHO+HMB IO groups compared with controls (noninjected fertile eggs). The activity of jejunal sucrase-isomaltase (SI) was higher (P (Key words: chicken, in ovo feeding, small intestine, sucrase-isomaltase) 2004 Poultry Science 83:2023–2028 INTRODUCTION The intestinal epithelium is a complex system of multiple cell types undergoing continual renewal and change with well-orchestrated patterns of gene expression during the processes of development and differentiation. This system has a major role in determining the developmental poten- tial of the hatched chick (Bayer et al., 1975; Uni et al., 1995, 1998). Chick growth and development are dependent upon nutrient digestion and absorption, which is a direct result of the functional and morphological development of the small intestine (Baranylova and Holman, 1976; Sell et al., 1991; Akiba and Murakami, 1995; Yamauchi et al., 1996; Uni, 1999). In chickens, the first days after hatch are a critical period for development because a major change 2004 Poultry Science Association, Inc. Received for publication April 14, 2004. Accepted for publication August 31, 2004. 1 This research was supported by Research Grant Award IS-3311-02 from BARD, The United States-Israel Binational Agriculture Research and Development. 2 To whom correspondence should be addressed: [email protected]. 2023 < 0.05) 48 h after IO feeding in all the IO fed embryos, whereas at day of hatch and at d 3 the CHO+HMB IO group had the highest maltase activity (P < 0.05), which was approximately 50% greater than control embryos. These observations indicated that small intestines of IO fed hatchlings were functionally at a similar stage of de- velopment as a conventionally fed 2-d-old chick. Body weight of all IO fed hatchlings was greater than controls, and these differences (P < 0.05) were sustained until the end of the experiment (10 d). At d 10 chicks that were IO fed with CHO had BW that were 2.2% higher, whereas HMB and CHO+HMB IO fed chicks showed 5 to 6.2% BW increase, respectively, compared with controls. The current study shows that the administration of exogenous nutrients into the amnion enhanced intestinal develop- ment by increasing the size of the villi and by increasing the intestinal capacity to digest disaccharides. This advan- tage probably leads to higher BW in IO fed chicks. occurs in the source of nutrients as the yolk is replaced by an exogenous diet (Noble and Ogunyemi, 1989; Noy and Sklan, 1998). Because the intestine is the nutrient supply organ, the sooner it achieves its functional capacity the earlier the young chick can use dietary nutrients and grow according to its genetic potential (Uni et al., 2003b; Uni and Ferket, 2004). Chicken embryos have limited ability to digest and ab- sorb nutrients prior to hatch, as reflected by relatively low mRNA levels of sucrase-isomaltase (SI) and l-aminopepti- dase and the ATPase and sodium glucose transporter (SGLT-1) in the small intestinal mucosa (Uni et al., 2003b). This absorption capability increases close to hatch and con- tinues to increase during the first few days posthatch (Uni et al., 1999; Sklan, 2001; Uni et al., 2003b). Villus height increases by 200 to 300% from d 17 of incubation until hatch. During the posthatch period the small intestine weight Abbreviation Key: CHO = carbohydrates: maltose, sucrose, dextrin, NaCl; CHO+HMB = maltose, sucrose, dextrin, NaCl, β-hydroxy-β-meth- ylbutyrate; HMB = β-hydroxy-β-methylbutyrate, NaCl; HMG-CoA = β- hydroxy-β-methylglutaryl coenzyme A; SI = sucrase-isomaltase.
Transcript of Effects of in ovo feeding of carbohydrates and beta-hydroxy-beta-methylbutyrate on the development...

Effects of In Ovo Feeding of Carbohydrates and β-Hydroxy-β-Methylbutyrateon the Development of Chicken Intestine1
E. Tako,* P. R. Ferket,† and Z. Uni*,2
*The Faculty of Agricultural, Food and Environmental Quality Sciences, the Department of Animal Sciences,Hebrew University of Jerusalem, PO Box 12, Rehovot, 76100, Isarel; and †North Carolina State University,
Department of Poultry Science, College of Agriculture and Life Sciences, Raleigh, North Carolina 27695-7608
ABSTRACT Early development of the digestive tract iscrucial for achieving maximal growth and developmentof chickens. Because the late-term embryo naturally con-sumes the amniotic fluids, insertion of a nutrient solutioninto the embryonic amniotic fluid [in ovo (IO) feeding]may enhance development. This study examined the ef-fect of IO feeding on d 17.5 of incubation of carbohydrates(CHO) and β-hydroxy-β-methylbutyrate (HMB) on smallintestinal development of chickens during the pre andposthatch periods. Results shows that 48 h post-IO feed-ing procedure all IO feeding treatments exhibited in-creased villus width and surface area compared with thecontrol group. At d 3 posthatch the surface area of anaverage villi was increased by 45% for the HMB IO groupand by 33% for the CHO and CHO+HMB IO groupscompared with controls (noninjected fertile eggs). Theactivity of jejunal sucrase-isomaltase (SI) was higher (P
(Key words: chicken, in ovo feeding, small intestine, sucrase-isomaltase)
2004 Poultry Science 83:2023–2028
INTRODUCTION
The intestinal epithelium is a complex system of multiplecell types undergoing continual renewal and change withwell-orchestrated patterns of gene expression during theprocesses of development and differentiation. This systemhas a major role in determining the developmental poten-tial of the hatched chick (Bayer et al., 1975; Uni et al., 1995,1998). Chick growth and development are dependent uponnutrient digestion and absorption, which is a direct resultof the functional and morphological development of thesmall intestine (Baranylova and Holman, 1976; Sell et al.,1991; Akiba and Murakami, 1995; Yamauchi et al., 1996;Uni, 1999). In chickens, the first days after hatch are acritical period for development because a major change
2004 Poultry Science Association, Inc.Received for publication April 14, 2004.Accepted for publication August 31, 2004.1This research was supported by Research Grant Award IS-3311-02
from BARD, The United States-Israel Binational Agriculture Researchand Development.
2To whom correspondence should be addressed: [email protected].
2023
< 0.05) 48 h after IO feeding in all the IO fed embryos,whereas at day of hatch and at d 3 the CHO+HMB IOgroup had the highest maltase activity (P < 0.05), whichwas approximately 50% greater than control embryos.These observations indicated that small intestines of IOfed hatchlings were functionally at a similar stage of de-velopment as a conventionally fed 2-d-old chick. Bodyweight of all IO fed hatchlings was greater than controls,and these differences (P < 0.05) were sustained until theend of the experiment (10 d). At d 10 chicks that wereIO fed with CHO had BW that were 2.2% higher, whereasHMB and CHO+HMB IO fed chicks showed 5 to 6.2%BW increase, respectively, compared with controls. Thecurrent study shows that the administration of exogenousnutrients into the amnion enhanced intestinal develop-ment by increasing the size of the villi and by increasingthe intestinal capacity to digest disaccharides. This advan-tage probably leads to higher BW in IO fed chicks.
occurs in the source of nutrients as the yolk is replaced byan exogenous diet (Noble and Ogunyemi, 1989; Noy andSklan, 1998). Because the intestine is the nutrient supplyorgan, the sooner it achieves its functional capacity theearlier the young chick can use dietary nutrients and growaccording to its genetic potential (Uni et al., 2003b; Uni andFerket, 2004).
Chicken embryos have limited ability to digest and ab-sorb nutrients prior to hatch, as reflected by relatively lowmRNA levels of sucrase-isomaltase (SI) and l-aminopepti-dase and the ATPase and sodium glucose transporter(SGLT-1) in the small intestinal mucosa (Uni et al., 2003b).This absorption capability increases close to hatch and con-tinues to increase during the first few days posthatch (Uniet al., 1999; Sklan, 2001; Uni et al., 2003b). Villus heightincreases by 200 to 300% from d 17 of incubation until hatch.During the posthatch period the small intestine weight
Abbreviation Key: CHO = carbohydrates: maltose, sucrose, dextrin,NaCl; CHO+HMB = maltose, sucrose, dextrin, NaCl, β-hydroxy-β-meth-ylbutyrate; HMB = β-hydroxy-β-methylbutyrate, NaCl; HMG-CoA = β-hydroxy-β-methylglutaryl coenzyme A; SI = sucrase-isomaltase.

TAKO ET AL.2024
increases faster than the body mass (Katanbaf et al., 1988;Sell et al., 1991; Sklan, 2001). This rapid intestinal growthis due to accelerated processes of enterocyte proliferationand differentiation (Geyra et al., 2001a). In addition, theintestinal crypts, which begin to form at hatch, are clearlydefined several days posthatch, increasing in cell numbersand size (Uni et al., 2000; Geyra et al., 2001a).
Previous studies have shown that feeding immediatelyposthatch leads to an acceleration of small intestinal mor-phological development (Noy and Sklan, 1998), whereaslate access to external feed results in delayed developmentof the small intestine mucosal layer (Uni et al., 1998; Geyraet al., 2001a; Uni et al., 2003b). Furthermore, birds deniedaccess to first feed for 24 to 48 h have decreased villi length(Yamauchi et al., 1996), decreased crypt size and crypts pervilli, and decreased enterocyte migration rate (Geyra et al.,2001b). In addition, delayed access to feed for 48 h post-hatch resulted in changes in mucin dynamics, which proba-bly affected the absorptive and protective functions of thesmall intestine (Uni et al., 2003a).
Thus, because immediate access to feed after hatch iscritical for the development of the intestine, nutrient supplyduring the prehatch period [at d 17 or 18 of incubation,using in ovo (IO) feeding] (Uni and Ferket, 2004) wouldbe expected to enhance development of the small intestine.Many possible nutrient supplements could be included inthe IO feeding solution. Carbohydrates (CHO) would beused as a source for glucose, which is crucial for hatchlingdevelopment (Moran, 1985). Sodium ions are important forthe brush border transporter action (Gal-Gerber et al., 2000;Currid et al., 2004). In addition other nutrients, such asminerals, vitamins, and enteric modulators are good candi-dates for IO feeding (Uni and Ferket 2004).
From the research reported by Nissen et al., (1994) β-hydroxy-β-methylbutyrate (HMB), a leucine metabolite,decreased chicken mortality and increased carcass yield.HMB is produced through the metabolism of α-ketoisoca-proate, which is oxidized by α-ketoisocaproate dioxygen-ase to produce free HMB within the cell cytosol (Sabourinand Bieber, 1982; Sabourin and Bieber, 1983). Studies haveshown (Van Koevering and Nissen, 1992) that smallamounts of HMB are synthesized endogenously in thebody, as approximately 5% of leucine metabolism normallyproceeds through this pathway. It was proposed by Nissenand Abumarad (1997) that HMB is converted to β-hydroxy-β-methylglutaryl coenzyme A (HMG-CoA) in some tissuesand serves as a key carbon source for de novo cholesterolsynthesis in tissues, which is necessary to maintain maxi-mal cell function. Furthermore, these researchers suggestedthat in situations of stimulated growth or differentiationHMG-CoA may be rate limiting for cholesterol synthesis,which could limit cell growth or function. Therefore, it wasproposed that feeding HMB could provide a saturatingsource of cytosolic HMG-CoA for cholesterol synthesis and,in turn, allow maximal cell growth and function.
3Gold Kist Hatchery, Silver City, NC.
The present study examined the hypothesis that IO feed-ing will accelerate development of the digestive tract dur-ing the pre- and posthatch periods. The effect of providingfeeding solutions, which contained sodium chloride, CHO,and HMB on intestinal development was studied from thelast quarter of embryonic incubation until d 3 posthatchby measuring morphological parameters of the intestinalmucosal layer and by evaluating activity of SI, a majorbrush border digestion enzyme,
MATERIALS AND METHODS
Incubation and IO Feeding
Fertile eggs (Ross × Ross) were obtained from a commer-cial hatchery3 from a maternal flock 35 wk in lay. The eggswere incubated under optimal conditions at the Depart-ment of Poultry Science at North Carolina State University.Ten eggs from each group were sampled at 18, 19, and 20d of incubation, at hatch, and at d 3 posthatch. At 17.5 dof incubation, 400 eggs containing viable embryos wereweighed and divided into 4 groups with equal weightfrequency distribution of 100 eggs each with average eggweight of 58 × 1.1g. Each group was then injected withsuitable IO feed treatment solutions (1 mL/egg) with a21-ga needle inserted into the amniotic fluid, which wasidentified by candling. The 3 treatment groups includedthe following: 1) CHO solution: 25 g of maltose/L, 25 g ofsucrose/L, 200 g of dextrin/L, and 5 g of NaCl/L, 2) HMBsolution: 1 g of HMB/L in 5 g of NaCl/L, 3) CHO+HMBsolution: 25 g of maltose/L, 25 g of sucrose/L, 200 g ofdextrin/L, 1 g of HMB/L in 5 g of NaCl/L. The controlwas a noninjected group that paralleled routine proceduresin commercial hatcheries. In addition, preliminary experi-ments conducted in our laboratory indicated that injectionof 1 mL of 5 g of NaCl/L did not affect (P < 0.05) embryoand chick BW, intestinal development, or brush bordermembrane enzymatic activity. After the eggs were injected,the injection holes were sealed with cellophane tape, andeggs were placed in hatching trays such that each treatmentwas equally represented in each location of the incubator.
Collecting Intestinal Segments
The jejunum was removed as previously described (Uniet al., 2003a) and 1 cm long segments were taken and placedin 2 separate tubes: 1) fixed in 4% neutral-buffered formalinsolution for histology, and 2) stored at −20°C for determina-tion of SI activity.
Bird Housing
Upon hatch (hatchability was 91% in all groups), chicksof each treatment were randomly assigned to 7 pens pertreatment (10 hatchlings/m2 pen) and housed in a totalconfinement building at North Carolina State University.Each pen was equipped with an automatic nipple drinkerand manual self-feeder. The concrete floor was beddedwith wood shavings, and supplemental incandescent heat
EFFECTS OF IN OVO FEEDING ON INTESTINE DEVELOPMENT 2025
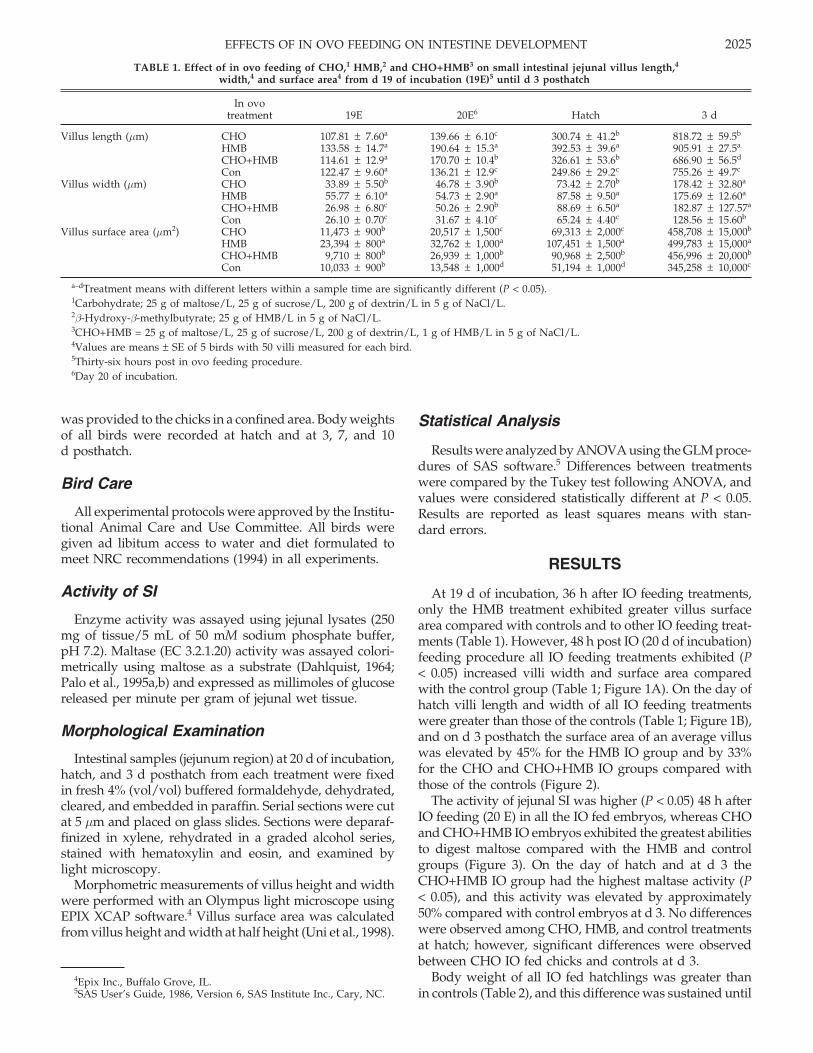
TABLE 1. Effect of in ovo feeding of CHO,1 HMB,2 and CHO+HMB3 on small intestinal jejunal villus length,4
width,4 and surface area4 from d 19 of incubation (19E)5 until d 3 posthatch
In ovotreatment 19E 20E6 Hatch 3 d
Villus length (µm) CHO 107.81 ± 7.60a 139.66 ± 6.10c 300.74 ± 41.2b 818.72 ± 59.5b
HMB 133.58 ± 14.7a 190.64 ± 15.3a 392.53 ± 39.6a 905.91 ± 27.5a
CHO+HMB 114.61 ± 12.9a 170.70 ± 10.4b 326.61 ± 53.6b 686.90 ± 56.5d
Con 122.47 ± 9.60a 136.21 ± 12.9c 249.86 ± 29.2c 755.26 ± 49.7c
Villus width (µm) CHO 33.89 ± 5.50b 46.78 ± 3.90b 73.42 ± 2.70b 178.42 ± 32.80a
HMB 55.77 ± 6.10a 54.73 ± 2.90a 87.58 ± 9.50a 175.69 ± 12.60a
CHO+HMB 26.98 ± 6.80c 50.26 ± 2.90b 88.69 ± 6.50a 182.87 ± 127.57a
Con 26.10 ± 0.70c 31.67 ± 4.10c 65.24 ± 4.40c 128.56 ± 15.60b
Villus surface area (µm2) CHO 11,473 ± 900b 20,517 ± 1,500c 69,313 ± 2,000c 458,708 ± 15,000b
HMB 23,394 ± 800a 32,762 ± 1,000a 107,451 ± 1,500a 499,783 ± 15,000a
CHO+HMB 9,710 ± 800b 26,939 ± 1,000b 90,968 ± 2,500b 456,996 ± 20,000b
Con 10,033 ± 900b 13,548 ± 1,000d 51,194 ± 1,000d 345,258 ± 10,000c
a–dTreatment means with different letters within a sample time are significantly different (P < 0.05).1Carbohydrate; 25 g of maltose/L, 25 g of sucrose/L, 200 g of dextrin/L in 5 g of NaCl/L.2β-Hydroxy-β-methylbutyrate; 25 g of HMB/L in 5 g of NaCl/L.3CHO+HMB = 25 g of maltose/L, 25 g of sucrose/L, 200 g of dextrin/L, 1 g of HMB/L in 5 g of NaCl/L.4Values are means ± SE of 5 birds with 50 villi measured for each bird.5Thirty-six hours post in ovo feeding procedure.6Day 20 of incubation.
was provided to the chicks in a confined area. Body weightsof all birds were recorded at hatch and at 3, 7, and 10d posthatch.
Bird Care
All experimental protocols were approved by the Institu-tional Animal Care and Use Committee. All birds weregiven ad libitum access to water and diet formulated tomeet NRC recommendations (1994) in all experiments.
Activity of SI
Enzyme activity was assayed using jejunal lysates (250mg of tissue/5 mL of 50 mM sodium phosphate buffer,pH 7.2). Maltase (EC 3.2.1.20) activity was assayed colori-metrically using maltose as a substrate (Dahlquist, 1964;Palo et al., 1995a,b) and expressed as millimoles of glucosereleased per minute per gram of jejunal wet tissue.
Morphological Examination
Intestinal samples (jejunum region) at 20 d of incubation,hatch, and 3 d posthatch from each treatment were fixedin fresh 4% (vol/vol) buffered formaldehyde, dehydrated,cleared, and embedded in paraffin. Serial sections were cutat 5 µm and placed on glass slides. Sections were deparaf-finized in xylene, rehydrated in a graded alcohol series,stained with hematoxylin and eosin, and examined bylight microscopy.
Morphometric measurements of villus height and widthwere performed with an Olympus light microscope usingEPIX XCAP software.4 Villus surface area was calculatedfrom villus height and width at half height (Uni et al., 1998).
4Epix Inc., Buffalo Grove, IL.5SAS User’s Guide, 1986, Version 6, SAS Institute Inc., Cary, NC.
Statistical Analysis
Results were analyzed by ANOVA using the GLM proce-dures of SAS software.5 Differences between treatmentswere compared by the Tukey test following ANOVA, andvalues were considered statistically different at P < 0.05.Results are reported as least squares means with stan-dard errors.
RESULTS
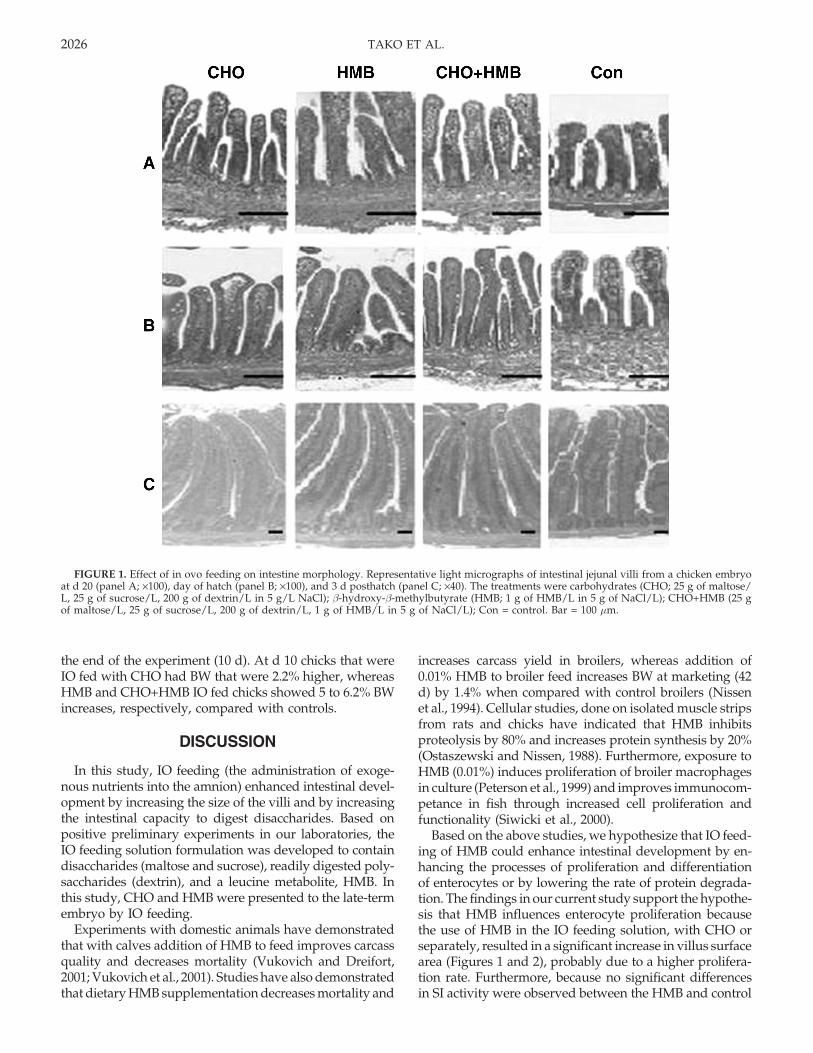
At 19 d of incubation, 36 h after IO feeding treatments,only the HMB treatment exhibited greater villus surfacearea compared with controls and to other IO feeding treat-ments (Table 1). However, 48 h post IO (20 d of incubation)feeding procedure all IO feeding treatments exhibited (P< 0.05) increased villi width and surface area comparedwith the control group (Table 1; Figure 1A). On the day ofhatch villi length and width of all IO feeding treatmentswere greater than those of the controls (Table 1; Figure 1B),and on d 3 posthatch the surface area of an average villuswas elevated by 45% for the HMB IO group and by 33%for the CHO and CHO+HMB IO groups compared withthose of the controls (Figure 2).
The activity of jejunal SI was higher (P < 0.05) 48 h afterIO feeding (20 E) in all the IO fed embryos, whereas CHOand CHO+HMB IO embryos exhibited the greatest abilitiesto digest maltose compared with the HMB and controlgroups (Figure 3). On the day of hatch and at d 3 theCHO+HMB IO group had the highest maltase activity (P< 0.05), and this activity was elevated by approximately50% compared with control embryos at d 3. No differenceswere observed among CHO, HMB, and control treatmentsat hatch; however, significant differences were observedbetween CHO IO fed chicks and controls at d 3.
Body weight of all IO fed hatchlings was greater thanin controls (Table 2), and this difference was sustained until
TAKO ET AL.2026
FIGURE 1. Effect of in ovo feeding on intestine morphology. Representative light micrographs of intestinal jejunal villi from a chicken embryoat d 20 (panel A; ×100), day of hatch (panel B; ×100), and 3 d posthatch (panel C; ×40). The treatments were carbohydrates (CHO; 25 g of maltose/L, 25 g of sucrose/L, 200 g of dextrin/L in 5 g/L NaCl); β-hydroxy-β-methylbutyrate (HMB; 1 g of HMB/L in 5 g of NaCl/L); CHO+HMB (25 gof maltose/L, 25 g of sucrose/L, 200 g of dextrin/L, 1 g of HMB/L in 5 g of NaCl/L); Con = control. Bar = 100 µm.
the end of the experiment (10 d). At d 10 chicks that wereIO fed with CHO had BW that were 2.2% higher, whereasHMB and CHO+HMB IO fed chicks showed 5 to 6.2% BWincreases, respectively, compared with controls.
DISCUSSION
In this study, IO feeding (the administration of exoge-nous nutrients into the amnion) enhanced intestinal devel-opment by increasing the size of the villi and by increasingthe intestinal capacity to digest disaccharides. Based onpositive preliminary experiments in our laboratories, theIO feeding solution formulation was developed to containdisaccharides (maltose and sucrose), readily digested poly-saccharides (dextrin), and a leucine metabolite, HMB. Inthis study, CHO and HMB were presented to the late-termembryo by IO feeding.
Experiments with domestic animals have demonstratedthat with calves addition of HMB to feed improves carcassquality and decreases mortality (Vukovich and Dreifort,2001; Vukovich et al., 2001). Studies have also demonstratedthat dietary HMB supplementation decreases mortality and
increases carcass yield in broilers, whereas addition of0.01% HMB to broiler feed increases BW at marketing (42d) by 1.4% when compared with control broilers (Nissenet al., 1994). Cellular studies, done on isolated muscle stripsfrom rats and chicks have indicated that HMB inhibitsproteolysis by 80% and increases protein synthesis by 20%(Ostaszewski and Nissen, 1988). Furthermore, exposure toHMB (0.01%) induces proliferation of broiler macrophagesin culture (Peterson et al., 1999) and improves immunocom-petance in fish through increased cell proliferation andfunctionality (Siwicki et al., 2000).
Based on the above studies, we hypothesize that IO feed-ing of HMB could enhance intestinal development by en-hancing the processes of proliferation and differentiationof enterocytes or by lowering the rate of protein degrada-tion. The findings in our current study support the hypothe-sis that HMB influences enterocyte proliferation becausethe use of HMB in the IO feeding solution, with CHO orseparately, resulted in a significant increase in villus surfacearea (Figures 1 and 2), probably due to a higher prolifera-tion rate. Furthermore, because no significant differencesin SI activity were observed between the HMB and control

EFFECTS OF IN OVO FEEDING ON INTESTINE DEVELOPMENT 2027
TABLE 2. Body weight1 (g) of embryos at d 19 of incubation (19E) and hatchlings (hatch and d 3, 7,and 10) from 3 in ovo feeding treatments: CHO,2 HMB,3 CHO+HMB,4 and control
Body weight (g)
In ovo treatment 19E Hatch Day 3 Day 7 Day 10
CHO 46.34 ± 0.44a 46.03 ± 0.76a 88.53 ± 3.74a 234.30 ± 0.91a 245.10 ± 2.27b
HMB 46.74 ± 0.90a 46.07 ± 0.47a 84.66 ± 1.87b 233.90 ± 1.15a 251.01 ± 2.4a
CHO+HMB 46.88 ± 0.54a 46.68 ± 0.52a 86.24 ± 2.18a 234.20 ± 2.35a 254.06 ± 4.04a
Con 45.92 ± 0.88a 44.59 ± 0.48b 84.01 ± 0.83b 231.10 ± 1.49b 239.70 ± 4.11c
a–dTreatment means with different letters within a sample time are significantly different (P < 0.05).1Weight values are means ± SE of 10 embryos at 19E, 70 birds at hatch, and 50 birds at d 3, 7, and 10.2Carbohydrate; 25 g of maltose/L, 25 g of sucrose/L, 200 g of dextrin/L in 5 g of NaCl/L.3β-Hydroxy-β-methylbutyrate; 25 g of HMB/L in 5 g of NaCl/L.4CHO+HMB = 25 g of maltose/L, 25 g of sucrose/L, 200 g of dextrin/L, 1 g of HMB/L in 5 g of NaCl/L.
chicks at day of hatch and at d 3, it is suggested that HMBdoes not have a direct critical effect on intestinal func-tionality.
Carbohydrates were chosen as an important componentof the IO feed solution because they are critical during thefinal stage of chick embryo development, prior to emer-gence from the shell, and very little CHO remains in theegg before hatch (Christensen et al., 1993). In addition,several studies have demonstrated that higher levels of thesubstrate in the intestine elevate activity of disaccharidases.In rats, after 48 h of fasting the sucrase activity decreasedand increased during refeeding (Holt and Yeh, 1992). Asolution containing free amino acids and glucose that wasgiven to premature neonatal piglets has been reported toenhance maltase and sucrase activities (Petersen et al., 2002)and the feeding of maternal feeding solutions containingvarious levels of sucrose, glucose, and galactose increasedvilli length and crypt formation in piglets (Pluske et al.,1996).
Therefore, we hypothesized that providing disaccharidesand easily digested CHO to the embryonic intestinal tissue,which at d 17.5 E has a low capacity to digest and absorb(Uni et al., 2003b), will elevate the activity of the relevant
FIGURE 2. Differences (%) in average villi surface area betweencontrol and 3 in ovo (IO) treatments. Carbohydrates (CHO; 25 g ofmaltose/L, 25 g of sucrose/L, 200 g of dextrin/L in 5 g of NaCl/L); β-hydroxy-β-methylbutyrate (HMB; 1 g of HMB/L in 5 g of NaCl/L);CHO+HMB (25 g of maltose/L, 25 g of sucrose/L, 200 g of dextrin/L,1 g of HMB/L in 5 g of NaCl/L). 20E = d 20 of incubation.
brush border enzymes. The current results support thishypothesis and show that CHO IO fed embryos had highermaltase activity before and after hatch. Furthermore, addi-tion of HMB to CHO led to the highest levels of sucraseactivity at hatch and at d 3 compared with other IO treat-ments and, therefore, may suggest an enhancing effect ofthis combination. This finding can be explained by theeffect of HMB on enterocyte proliferation and the knowneffect of CHO substrate on brush border enzyme activity.
The morphological measurements of the jejunal villi re-vealed that CHO IO fed embryos had 50% elevation invillus surface area prior to hatch, whereas HMB IO fedembryos exhibited 140% elevation, and the CHO+HMB IOfed embryos were intermediate (Figure 3; Table 1). Thedifferences remained through hatch and d 3 (Table 1),whereas the gradual reduction in the difference betweenIO and control treatments suggested that the effect of IOfeeding on intestine morphological development is maxi-mal at 48 h after the IO procedure.
Because the villus surface area has been shown to becorrelated with growth in the chicken (Sklan, 2001) the
FIGURE 3. Activity of sucrase-isomaltase at 18, 19, and 20 d ofincubation (18E, 19E, and 20E, respectively); hatch; and 3 d posthatch.a–cColumns with different letters differ significantly (P < 0.05). Valuesare means × SE, n = 10. The 3 in ovo (IO) feeding treatments werecarbohydrates (CHO; 25 g of maltose/L, 25 g of sucrose/L, 200 g ofdextrin/L in 5 g of NaCl/L); β-hydroxy-β-methylbutyrate (HMB; 1 gof HMB/L in 5 g of NaCl/L); CHO+HMB (25 g of maltose/L, 25 g ofsucrose/L, 200 g of dextrin/L, 1 g of HMB/L in 5 g of NaCl/L).

TAKO ET AL.2028
larger surface area in the IO treatments probably contrib-uted to increased nutrient digestion and assimilation andresulted in BW elevation (Table 2). The current studyshowed that IO fed chicks had enhanced enteric develop-ment, which probably led to higher BW in IO fed chicks.
REFERENCES
Akiba, Y., and H. Murakami. 1995. Partitioning of energy andprotein during early growth of broiler chicks and contributionof vitelline residue. 10th European Symposium on PoultryNutrition, Antalia, Turkey.
Baranylova, E., and J. Holman. 1976. Morphological changes inthe intestinal wall in fed and fasted chickens in the first weekafter hatching. Acta Vet. (Brno) 45:151–158.
Bayer, R. C., C. B. Chawan, F. H. Bird, and S. D. Musgrave. 1975.Characteristics of the absorptive surface of the small intestineof the chicken from 1 day to 14 weeks of age. Poult. Sci.54:155–169.
Christensen, V. L., W. E. Donaldson, and K. E. Nestor. 1993. Effectof maternal dietary triiodothyronine on embryonic physiologyof turkeys. Poult. Sci. 72:2316–2327.
Currid, A., B. Ortega, and M. A. Valverde. 2004. Chloride secretionin a morphologically differentiated human colonic cell line thatexpresses the epithelial Na+ channel. J. Physiol. 555:241–250.
Dahlquist, A. 1964. Method of assay of intestinal disaccaridases.Anal. Biochem. 7:18–25.
Gal-Gerber, O., S. J. Mabjeesh, D. Sklan, and Z. Uni. 2000. Partialsequence and expression of the gene for and activity of thesodium-glucose transporter in the small intestine of fed,starved and re-fed chickens. J. Nutr. 130:2174–2179.
Geyra, A., Z. Uni, and D. Sklan. 2001a. Enterocyte dynamicsand mucosal development in the posthatch chick. Poult. Sci.80:776–782.
Geyra, A., Z. Uni, and D. Sklan. 2001b. the effect of fasting atdifferent ages on growth and tissue dynamics in the smallintestine of the young chick. Br. J. Nutr. 86:53–61.
Holt, P. R., and K. Y. Yeh. 1992. Effects of starvation and refeedingon jejunal disaccharidase activity. Dig. Dis. Sci. 37:827–832.
Katanbaf, M. N., E. A. Dunnington, and P. B. Siegel. 1988. Allo-morphic relationships from hatching to 56 days in parentallines and F1 crosses of chickens selected 27 generations forhigh and low body weight. Growth Dev. Aging 51:11–21.
Moran, E. T. 1985. Digestion and absorption in fowl and eventsthrough prenatal development. J. Nutr. 115:665–674.
National Research Council. 1994. Nutrient Requirements of Poul-try. 9th ed. National Academy of Science, Washington, DC.
Nissen, S. L., and N. N. Abumrad 1997. Nutritional role of theleucine metabolite β-hydroxy-β-methylbutyrate (HMB). J.Nutr. Biochem. 8:300–311.
Nissen, S., J. C. Fuller, Jr., J. Sell, P. R. Ferket, and D. V. Rives.1994. The effect of beta-hydroxy-beta-methylbutyrate ongrowth, mortality, and carcass qualities of broiler chickens.Poult. Sci. 73:137–155.
Noble, R. C., and D. Ogunyemi. 1989. Lipid changes in the residualyolk and liver of the chick immediately after hatching. Biol.Neonate 56:228–236.
Noy, Y., and D. Sklan. 1998. Yolk utilisation in the newly hatchedpoult. Br. Poult. Sci. 39:446–451.
Ostaszewski, P., and S. Nissen. 1988. Effect of hyperglucagonemiaon whole-body leucine metabolism in immature pigs beforeand during a meal. Am. J. Physiol. 254:E372–E377.
Palo, P. E., J. L. Sell, F. J. Piquer, M. F. Soto-Salanova, and L.Vilaseca. 1995a. Effect of early nutrition restriction on broilerchicken: 1. Performance and development of the gastrointesti-nal tract. Poult. Sci. 74:88–101.
Palo, P. E., J. L. Sell, F. J. Piquer, L. Vilaseca, and M. F. Soto-Salanova. 1995b. Effect of early nutrient restriction on broiler
chickens. 2. Performance and digestive enzyme activities.Poult. Sci. 74:1470–1483.
Petersen, Y. M., J. Elnif, M. Schmidt, and P. T. Sangild. 2002.Glucagon-like peptide 2 enhances maltase-glucoamylase andsucrase-isomaltase gene expression and activity in parenterallyfed premature neonatal piglets. Pediatr. Res. 52:498–503.
Peterson, A. L., M. A. Qureshi, P. R. Ferket, and J. C. Fuller, Jr.1999. In vitro exposure with beta-hydroxy-beta-methylbuty-rate enhances chicken macrophage growth and function. Vet.Immunol. Immunopathol. 67:67–78.
Pluske, J. R., M. J. Thompson, C. S. Atwood, P. H. Bird, I. H.Williams, and P. E. Hartmann. 1996. Maintenance of villusheight and crypt depth, and enhancement of disaccharide di-gestion and monosaccharide absorption, in piglets fed on cows’whole milk after weaning. Br. J. Nutr. 76:409–422.
Sabourin, P. J., and L. L. Bieber. 1982. The mechanism of alpha-ketoisocaproate oxygenase. Formation of beta-hydroxyisoval-erate from alpha-ketoisocaproate. J. Biol. Chem. 257:7468–7471.
Sabourin, P. J., and L. L. Bieber. 1983. Formation of beta-hydroxyi-sovalerate by an alpha-ketoisocaproate oxygenase in humanliver. Metabolism 32:160–164.
Sell, J. L., C. R. Angel, F. J. Piquer, E. G. Mallarino, and H. A. al-Batshan. 1991. Developmental patterns of selected characteris-tics of the gastrointestinal tract of young turkeys. Poult. Sci.70:1200–1205.
Sklan, D. 2001. Development of the digestive tract of poultry.Worlds Poult. Sci. J. 57:415–427.
Siwicki, A. K., J. C. Fuller, Jr., S. L. Nissen, P. Ostaszewski, and M.Studnicka. 2000. In vitro effects of β-hydroxy-β-methylbutyrate(HMB) on cell-medidated immunity in fish. Vet. Immunol.Immunopathol. 76(3–4):191–197.
Uni, Z. 1999. Functional development of the small intestine indomestic birds: Cellular and molecular aspects. Poult. AvianBiol. Rev. 10:167–179.
Uni, Z., and P. R. Ferket. 2004. Methods for early nutrition andtheir potential. World’s Poult. Sci. J. 60:101–111.
Uni, Z., S. Ganot, and D. Sklan. 1998. Posthatch developmentof mucosal function in the broiler small intestine. Poult. Sci.77:75–82.
Uni, Z., A. Geyra, H. Ben-Hur, and D. Sklan. 2000. Small intestinaldevelopment in the young chick: Crypt formation and entero-cyte proliferation and migration. Br. Poult. Sci. 41:544–551.
Uni, Z., Y. Noy, and D. Sklan. 1995. Posthatch changes in morphol-ogy and function of the small intestines in heavy- and light-strain chicks. Poult. Sci. 74:1622–1629.
Uni, Z., Y. Noy, and D. Sklan. 1999. Posthatch development ofsmall intestinal function in the poult. Poult. Sci. 78:215–222.
Uni, Z., A. Smirnov, and D. Sklan. 2003a. Pre- and posthatchdevelopment of goblet cells in the broiler small intestine: Effectof delayed access to feed. Poult. Sci. 82:320–327.
Uni, Z., E. Tako, O. Gal-Garber, and D. Sklan. 2003b. Morphologi-cal, molecular, and functional changes in the chicken smallintestine of the late-term embryo. Poult. Sci. 82:1747–1754.
Van Koevering, M., and S. Nissen. 1992. Oxidation of leucine andalpha-ketoisocaproate to beta-hydroxy-beta-methylbutyrate invivo. Am. J. Physiol. 262:E27–E31.
Vukovich, M. D., and G. D. Dreifort. 2001. Effect of beta-hydroxy-beta-methylbutyrate on the onset of blood lactate accumulationand V(O)(2) peak in endurance-trained cyclists. J. StrengthCond. Res. 15:491–497.
Vukovich, M. D., G. Slater, M. B. Macchi, M. J. Turner, K. Fallon,T. Boston, and J. Rathmacher. 2001. Beta-hydroxy-beta-methyl-butyrate (HMB) kinetics and the influence of glucose ingestionin humans. J. Nutr. Biochem. 12:631–639.
Yamauchi, K., H. Kamisoyama, and Y. Isshiki. 1996. Effects offasting and refeeding on structures of the intestinal villi andepithelial cells in White Leghorn hens. Br. Poult. Sci. 37:909–921.




















