
Spies Wearing Purple Hats: The use of social computing to ...
Upload
independentCategory
view
0download
0
ORIGINAL ARTICLE
Takato Kobayashi Æ Masahiko Kitahara
Eri Tanaka
Effects of habitat fragmentation on the three-way interaction amongants, aphids and larvae of the giant purple emperor,Sasakia charonda (Hewitson), a near-threatened butterfly
Received: 6 June 2006 / Accepted: 11 May 2007 / Published online: 2 August 2007� The Ecological Society of Japan 2007
Abstract Survival rates of both early and middle instarlarvae of the nymphalid butterfly, Sasakia charonda,were estimated to be lowest on test trees planted in ameadow (site A), intermediate in a small, narrow sec-ondary deciduous broadleaf forest (small patch, site B)and highest in a large secondary deciduous broadleafforest (large forest, site C). The larval mortality ratesdue to predation by tree-climbing predators from theground (tree climbing predator) such as ants and thelarvae of carabids were estimated to be greater at sites Aand B than those at site C. The number of predatoryants climbing test trees was significantly greater at sitesA and B than at site C, and the ants harvested honeydewfrom aphids living on tree leaves at those two sites.Aphid densities were significantly higher on trees at sitesA and B than at site C, and aphid densities and numbersof predatory ants were significantly and positively cor-related at sites A and B. In an experiment controllingaphid density per branch on test trees, the numbers ofants and the mortality rates of S. charonda larvae weregreater on branches with high aphid densities than onthose with low aphid densities at both sites A and B.These results suggest that the aphid density per host treewas higher in the meadow and the small patch than in
the large forest; at both sites these higher aphid densitiesattracted higher numbers of predatory ants to test trees,and as a result, mortality rates of S. charonda larvaewere increased.
Keywords Habitat fragmentation Æ Sasakia charonda ÆAnts Æ Aphids Æ Three-way interaction
Introduction
In Japan, secondary broadleaf deciduous forests inagroecosystems used to be managed for production offirewood and charcoal. However management practicesof these forests have all but been abandoned in recentdecades, and the forests have become extremely frag-mented through human land development. In general,habitat fragmentation is thought to be one of themajor causes of species extinction (Wilcox and Mur-phy 1985). Thus, wildlife inhabitation of secondarybroadleaf deciduous forests in agroecosystems nearurban areas is predicted to be declining due to forestfragmentation.
It is also well known that habitat fragmentation altersthe community structure of wildlife (e.g., MacGarvin1982; Robertson et al. 1995). Indeed, several researchersreported that habitat fragmentation affected bioticinteractions among the component species in a com-munity, and as a result, the densities of the specieschanged (Gates and Gysel 1978; Wilcove 1985; Jenner-sten 1988; Rathcke and Jules 1993; Aizen and Feinsinger1994; Washitani 1996).
In Japan, the nymphalid butterfly, Sasakia charonda(Lepidoptera, Nymphalidae) is one of the typical insectsinhabiting secondary broadleaf deciduous forests. Thisspecies was quite common in secondary broadleafdeciduous forests. However, since the 1960s, corre-sponding to a period of rapid economic growth, thepopulation density of the species has decreased as a
T. Kobayashi Æ E. TanakaLaboratory of Applied Entomology, Faculty of Agriculture,Utsunomiya University, Mine350, Utsunomiya,Tochigi 321-8505, Japan
M. KitaharaLaboratory of Animal Ecology,Yamanashi Institute of Environmental Sciences,5597-1 Kenmarubi, Kamiyoshida, Fujiyoshida,Yamanashi 403-0005, Japan
T. Kobayashi (&)K-101, 2-2-18 Umezono, Tsukuba,Ibaragi 305-0045, JapanE-mail: [email protected].: +81-29-8597230Fax: +81-29-8597230
Ecol Res (2008) 23: 409–420DOI 10.1007/s11284-007-0400-1

direct result of deforestation associated with changes inhuman land use (Higuchi and Sato 1987). Consequently,the species is now designated as a near-threatened insectin Japan (Ministry of the Environment 2006).
Our previous studies on the ecology of the speciesrevealed that: (1) there was a significant positive corre-lation between the density of overwintering S. charondalarvae per host tree and the area of the secondarydeciduous broadleaf forests within a radius of 100 m ofthe host tree (Kobayashi et al. 2004); (2) both in ameadow and a small secondary deciduous broadleafforest (small patch), the major mortality factor of earlyinstar larvae was predation by ants, whereas in a largebroadleaf forest (large forest) it was predation by wasps.In addition, the mortality rate of the early instar larvaewas highest in the meadow, intermediate in the smallpatch and lowest in the large forest (Kobayashi andInaizumi 2002); (3) aphid density per host tree wasgreater both in the meadow and the small patch than inthe large forest. The ants responsible for mortality of S.charonda larvae fed on honeydew secreted from theseaphids (Kobayashi and Inaizumi 2002). Thus, it wasspeculated that the increase of aphid density caused byforest fragmentation increases the mortality rate of earlyand middle instar larvae of S. charonda due to the in-creased predation by ants. However, this speculation hasnot yet been tested experimentally.
In this paper, to clarify the mechanisms depressingS. charonda populations experiencing habitat fragmen-tation, we examined whether the three-way interactionamong ants, aphids and this butterfly varies amongsecondary deciduous broadleaf forests of different sizes.
Materials and methods
Life history of S. charonda
In Japan, S. charonda is univoltine and overwinters inthe larval stage. The host tree is Celtis sinensis or C.jezoensis. On the Kanto Plain of Japan, the flight seasonof adults extends from late June to late August. On theKanto Plain, the minimum duration of the egg stage is5 days (Kobayashi and Inaizumi 2002). The mortalityrates of eggs, early (from first to second instar) andmiddle (from third to fourth instar just before over-wintering) instar larvae evaluated in the previous studywere 10–25, >90 and 70–80%, respectively (Kobayashiand Inaizumi 2002). The mortality rate of overwinteringlarvae was about 30% (Kobayashi and Inaizumi 2003),and the mortality rate between the completion of over-wintering and the emergence of adults was greater than80% (Kobayashi and Inaizumi 1999, 2000).
Study sites
We established the following three study sites, A, B andC, on the Utsunomiya University Farm, Mooka City,
Tochigi Prefecture (Fig. 1). We planted ten 3.0-m tall C.sinensis trees, with a diameter of about 30 mm at breastheight in late February 1999, in a row at intervals of2.5 m at each site. Site A was located at the center of ameadow, and a secondary deciduous broadleaf forestcovered 5% of the area within a 100-m radius of theplanted trees. Site B was located at the edge of a smallbroadleaf forest surrounded by an asphalt road and cropfield. The area of the small forest was 0.2 ha, and abroadleaf forest occupied 20% of the area within a 100-m radius of the planted trees. Site C was located along awide path through a large broadleaf forest, and 79 and8% of the area within a 100-m radius of the planted treesconsisted of broadleaf and coniferous forests, respec-tively. The planted trees used in the experiments of thisstudy were pruned every year from 1999 to 2002 so thatthe crowns would not overlap each other. Secondarydeciduous broadleaf forests near the three study sitesconsisted of patches of three types of forests: (1) mixedforest dominated by Castanea crenata, Quercus seratta,Swida controversa and Styrax japonica; (2) plantationsof Q. acutissima or (3) plantations of Q. serrata. Theconiferous forests near the sites consisted of plantationsof Cryptomeria japonica. Near the three sites, the mixedforests were more widely distributed than at the plan-tations of Q. acutissima, Q. serrata or C. japonica (Ko-bayashi et al. 2004). The secondary deciduous broadleaf
Fig. 1 Map of the study area for Sasakia charonda in Mooka City,Tochigi Prefecture, central Japan. Ten experimental Celtis sinensissaplings were planted at each of three sites A–C
410

forests within a 100-m radius of the test trees at the threesites were all plantations of Q. serrata; there was nodifference in the construction of the tree species and sizeamong the sites. Because the longest distance betweentwo sites was approximately only 1,000 m (between Aand B; Fig. 1), it is likely that weather effects on larvaeat the three sites were the same. A factor that differedamong the three sites was the area of secondary decid-uous broadleaf forests within a 100-m radius of theplanted test trees.
Methods
In all experiments described hereafter, we inoculated thelarvae, which were progeny of adult female S. charondacaptured near the study sites, onto the test trees. Weinoculated all larvae on the branches or leaves of testtrees using a calligraphy brush (inoculation).
Survival rates of early and middle instar larvae
We inoculated 50 freshly hatched larvae of S. charondaonto each test tree at sites A and C on 9 July and at siteB on 14 July 2000. We inoculated all the larvae onto onerandomly selected leaf, because larvae of S. charondahatch from an egg batch simultaneously, and just afterhatching disperse to the neighboring branches (Fukudaet al. 1983). The number of live larvae on each test treewas counted 18 days after inoculation, which corre-sponds to the duration of the first and second instars(16–18 days; Kobayashi and Inaizumi 2002).
To examine the survival rate of middle (third) instarlarvae, we randomly selected eight test trees at each siteand inoculated 30 third instar larvae on each selectedtree just after molting on 9 August 2000. The larvae wereinoculated on 30 randomly selected leaves to distributethem loosely on the test trees, because third instar larvaeof S. charonda typically distribute loosely on the testtrees and never aggregate on a particular branch (Ko-bayashi, unpublished data). The number of live larvaeon each test tree was counted 18 days after inoculation.
Survival rates of early and middle instar larvae in responseto tree climbing predators from the ground
At each site, the ten test trees were randomly dividedinto two groups of five, with each group being assignedto either treatment or control conditions. On 3 July2001, we evaluated the effects of predators that climb thetrees from the ground (tree-climbing predators) on sur-vival of S. charonda larvae by applying a 15-cm-wideband of Tanglefoot (Fuji Tangle�) to the trunks of fivetreatment trees at a height of 1.0 m from the roots. Theremaining five trees were not treated with Tanglefoot.We inoculated 50 newly hatched larvae onto each testtree on 6 July 2001 and 50 reared third instar larvae just
after molting on 26 July. For each stage, the number oflive larvae remaining on each test tree was counted 1, 4,9 and 16 days after inoculation. In this experiment, bothnewly hatched larvae and third instar larvae were inoc-ulated onto one randomly selected leaf. We also re-corded the species of the predators that were observed toeat early and middle instar larvae of S. charonda.
Ant species and the number of each species climbingthe test trees
We identified ant species and counted the number ofindividuals of each species climbing the test trees at eachsite for a 3-min period 12 times (6 times during the earlyday, 2 times in the afternoon/evening and 4 times atnight) during the period from 3 July to 7 August in 2001.The period roughly corresponded to that of the experi-ments described in ‘‘Tanglefoot experiment’’. In thisstudy, early day observations took place between 10:00and 13:00, afternoon/evening observations between15:00 and 18:00, and night observations between 20:00and 23:00. We observed all ten test trees on 3 July 2001,prior to the application of Tanglefoot to half of the tentrees in the study described in ‘‘Tanglefoot experiment’’.After the application of Tanglefoot, we surveyed onlythe five control trees. Tree trunks were marked at aheight of 50 cm from the roots, and the ant species andthe number of individuals of each species that passedthrough this mark from below during a period of 3 minwere recorded. In total, each site was surveyed 65 times(10 test trees surveyed · 1 survey + 5 test trees ·11 surveys).
Density of aphids providing ants with honeydew
On 21 April 2001, we randomly marked ten new shoots(which had sprouted within the previous year) on eachtest tree. On 3 and 23 July and 1 August 2001, we ob-served both leaf surfaces of the first 20 leaves from thetips of each marked shoot, recorded the species of aphidsthat provided the ants with the honeydew and countedindividuals of each species. Thus, we recorded thenumber of aphids per 200 leaves (x) on each test tree (20leaves · 10 shoots). We treated this number as an indi-cator of aphid density per tree.
Relationship between aphid density and the mortalityrates of S. charonda larvae due to predation by ants(net bag experiment)
To determine whether higher aphid density increasepredation of S. charonda larvae by ants, we conductedan experiment at sites A and B. In mid-June 2002, threebranches of each test tree at the two sites were randomlyselected, and the point at which each branch was 1 cm indiameter was marked with vinyl tape. We researched thepart of the branch from the marked point to the tip.
411

Some kinds of animals such as spiders, ants, wasps andfrogs were removed from each branch by hand. Aphidswere killed by spraying a 5% solution of milk dilutedwith water. This solution did not affect larvae of S.charonda (Kobayashi, unpublished).
Next, we covered each branch with a 0.18-mm meshnylon net bag (diameter 60 cm and depth 1.0 m) and tiedits base with a wire. Removal of the aphids and naturalenemies stated above was repeated four times a monthfrom mid-June to mid-July, 2002. Each of the three en-closed branches of each test tree was randomly assignedto high, medium or low aphid density treatment. Weintroduced the aphids, Shivaphis celti, into the enclosuresof branch with varying densities per branch, i.e., 400individuals (high density), 200 individuals (mediumdensity) and four individuals (low density) on 14 July2002 at site A and 15 July at site B. S. celti were sampledfrom leaves on non-experimental branches on each hosttree. To allow ants to enter the net bags, we inserted a 30-cm-long transparent acrylic tube (10 mm in diameter)into each net bag along the surface of the branch.
First, for the experiment with early instar larvae, weinoculated 30 freshly hatched larvae of S. charonda oneach branch on 17 July 2002 at site A and on 18 July2002 at site B. We opened each net bag and counted thenumber of surviving larvae 1, 3, 8 and 16 days afterinoculation. On day 16, we removed all surviving larvae,sprayed each branch with water-diluted milk and re-fastened the net bags for the next experiment.
Second, for the experiment with middle instar larvae,we assigned two aphid density levels to two randomlyselected branches: high (400 S. celti) and low (fourS. celti). On August 13 at site A and August 14 at site B,2002, we inoculated 30 newly molted third instar larvaeonto each branch with a net bag. The number of sur-viving larvae was recorded 1, 3, 8 and 16 days afterinoculation.
To examine the relationship between aphid densityand the number of ants entering the net bag through theacrylic tube, the number of each ant species passingthrough the tube into the bag was recorded for a periodof 3.0 min. For early instar larvae, this observation wasconducted during the early day and at night on the dayof larval inoculation day, and during the early day ondays 3, 8 and 16 after inoculation; for middle instarlarvae, it was conducted during the early day and atnight on day 1, and during the early day on days 4, 9 and16 after inoculation.
Statistical analysis
For each site, the larval survival rate per tree on eachstudy day and the rates obtained were arcsine-trans-formed, while the number of ants (x/3 min/host tree)and aphids (N/200 leaves/host tree) was log(x + 1)- andlog(N + 1)-transformed prior to statistical analysis. Thenumber of ants was the total number of individuals of allant species, detected as a predator of S. charonda.
One-way ANOVA was first used to compare larvalsurvival rates, numbers of ants and aphids among thethree sites or three treatments. When a significant dif-ference was detected in the above analysis, we usedTukey–Kramer multiple comparisons to confirm thetreatments or sites with a significant difference. A t testwas used to compare values between the two sites ortreatments. We used a Pearson correlation (r) betweenlog-transformed data of ants (x + 1) and aphid densityper host tree (N + 1) at each site on 3 July 2001 andthen the significance of the values were tested.
Results
Survival rates of early and middle instar larvae
Average survival rates of both early and middle instarlarvae were lowest at site A (early: 0, middle: 0.03),intermediate at site B (early: 0.07, middle: 0.28) andhighest at site C (early: 0.17, middle: 0.69) (Table 1). Atsite A, all inoculated early instar larvae disappeared. Thedifferences in the survival rates of both early and middleinstar larvae in all pairwise comparisons among thethree sites were significant (Table 1).
Mortality rates of early and middle instar larvaecaused by climbing predators
Early instar larvae
At all sites, we observed rapid disappearance of larvaeon both treatment and control trees by the 1st day afterinoculation, and survival rates on control trees werealways lower than on treatment trees (Fig. 2). Survivalrates on both treatment and control trees at site A de-creased greatly between the day of inoculation and day1, but the decrease was slight between days 1 and 9, andthereafter remained stable until the last day of theexperiment.
Natural enemies including ants and the mortality ratesof S. charonda larvae by them are shown in Table 2; re-sults presented in the table include findings from a previ-ous study (Kobayashi and Inaizumi 2002). The table alsoshows the frequency of attacks and kills on larvae on testtrees by each enemy in 1999, 2000 and 2001.
Table 1 Average of survival rates for early (1st and 2nd) andmiddle (3rd and 4th) instar larvae of Sasakia charonda at sites A, Band C
Site Pa
A B C
Early instar (n = 10) 0a 0.07b 0.17c F = 27.1, P < 0.001Middle instar (n = 8) 0.03a 0.28b 0.69c F = 52.4, P < 0.001
Within each instar, values followed by different letters are signifi-cantly different. (Tukey–Kramer multiple comparisons, P < 0.05;survival rates were arcsine-transformed)a One-way ANOVA
412
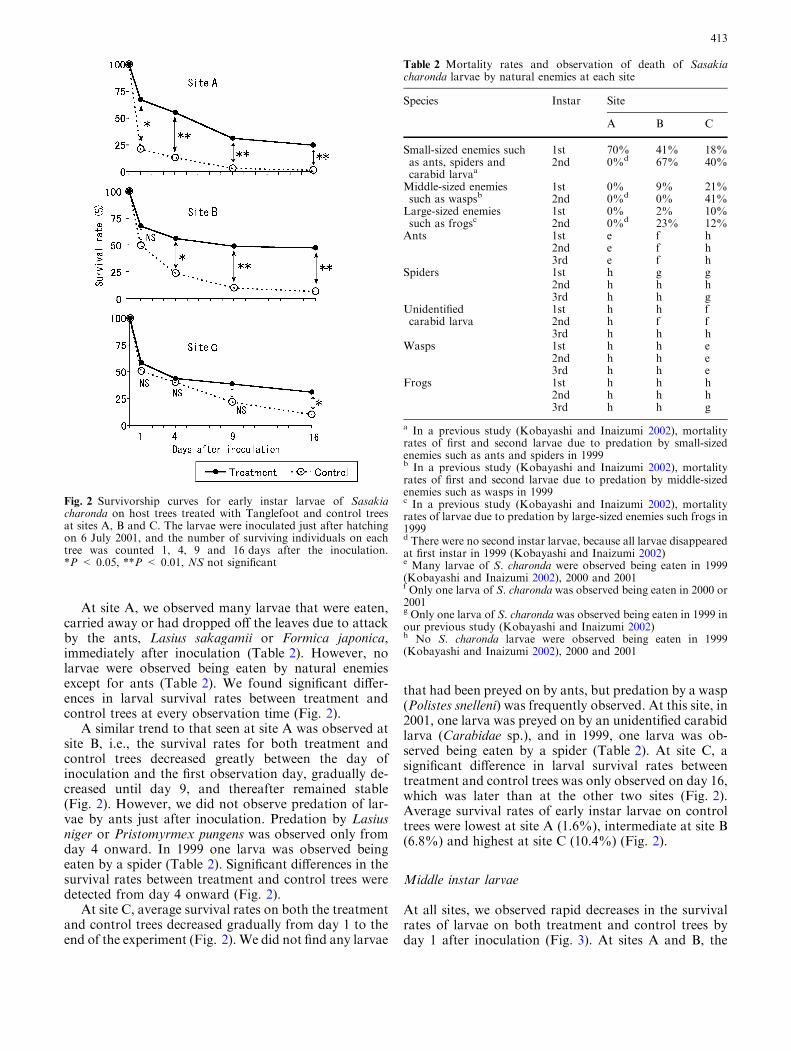
At site A, we observed many larvae that were eaten,carried away or had dropped off the leaves due to attackby the ants, Lasius sakagamii or Formica japonica,immediately after inoculation (Table 2). However, nolarvae were observed being eaten by natural enemiesexcept for ants (Table 2). We found significant differ-ences in larval survival rates between treatment andcontrol trees at every observation time (Fig. 2).
A similar trend to that seen at site A was observed atsite B, i.e., the survival rates for both treatment andcontrol trees decreased greatly between the day ofinoculation and the first observation day, gradually de-creased until day 9, and thereafter remained stable(Fig. 2). However, we did not observe predation of lar-vae by ants just after inoculation. Predation by Lasiusniger or Pristomyrmex pungens was observed only fromday 4 onward. In 1999 one larva was observed beingeaten by a spider (Table 2). Significant differences in thesurvival rates between treatment and control trees weredetected from day 4 onward (Fig. 2).
At site C, average survival rates on both the treatmentand control trees decreased gradually from day 1 to theend of the experiment (Fig. 2). We did not find any larvae
that had been preyed on by ants, but predation by a wasp(Polistes snelleni) was frequently observed. At this site, in2001, one larva was preyed on by an unidentified carabidlarva (Carabidae sp.), and in 1999, one larva was ob-served being eaten by a spider (Table 2). At site C, asignificant difference in larval survival rates betweentreatment and control trees was only observed on day 16,which was later than at the other two sites (Fig. 2).Average survival rates of early instar larvae on controltrees were lowest at site A (1.6%), intermediate at site B(6.8%) and highest at site C (10.4%) (Fig. 2).
Middle instar larvae
At all sites, we observed rapid decreases in the survivalrates of larvae on both treatment and control trees byday 1 after inoculation (Fig. 3). At sites A and B, the
Fig. 2 Survivorship curves for early instar larvae of Sasakiacharonda on host trees treated with Tanglefoot and control treesat sites A, B and C. The larvae were inoculated just after hatchingon 6 July 2001, and the number of surviving individuals on eachtree was counted 1, 4, 9 and 16 days after the inoculation.*P < 0.05, **P < 0.01, NS not significant
Table 2 Mortality rates and observation of death of Sasakiacharonda larvae by natural enemies at each site
Species Instar Site
A B C
Small-sized enemies suchas ants, spiders andcarabid larvaa
1st 70% 41% 18%2nd 0%d 67% 40%
Middle-sized enemiessuch as waspsb
1st 0% 9% 21%2nd 0%d 0% 41%
Large-sized enemiessuch as frogsc
1st 0% 2% 10%2nd 0%d 23% 12%
Ants 1st e f h2nd e f h3rd e f h
Spiders 1st h g g2nd h h h3rd h h g
Unidentifiedcarabid larva
1st h h f2nd h f f3rd h h h
Wasps 1st h h e2nd h h e3rd h h e
Frogs 1st h h h2nd h h h3rd h h g
a In a previous study (Kobayashi and Inaizumi 2002), mortalityrates of first and second larvae due to predation by small-sizedenemies such as ants and spiders in 1999b In a previous study (Kobayashi and Inaizumi 2002), mortalityrates of first and second larvae due to predation by middle-sizedenemies such as wasps in 1999c In a previous study (Kobayashi and Inaizumi 2002), mortalityrates of larvae due to predation by large-sized enemies such frogs in1999d There were no second instar larvae, because all larvae disappearedat first instar in 1999 (Kobayashi and Inaizumi 2002)e Many larvae of S. charonda were observed being eaten in 1999(Kobayashi and Inaizumi 2002), 2000 and 2001f Only one larva of S. charonda was observed being eaten in 2000 or2001g Only one larva of S. charonda was observed being eaten in 1999 inour previous study (Kobayashi and Inaizumi 2002)h No S. charonda larvae were observed being eaten in 1999(Kobayashi and Inaizumi 2002), 2000 and 2001
413
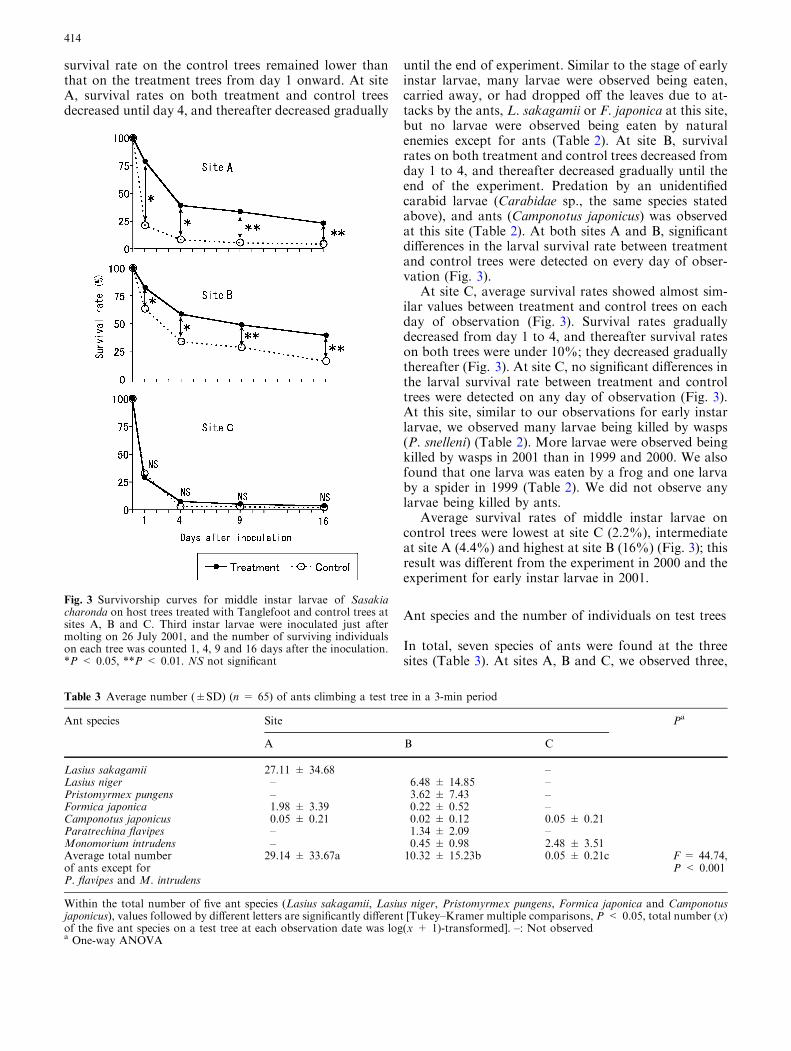
survival rate on the control trees remained lower thanthat on the treatment trees from day 1 onward. At siteA, survival rates on both treatment and control treesdecreased until day 4, and thereafter decreased gradually
until the end of experiment. Similar to the stage of earlyinstar larvae, many larvae were observed being eaten,carried away, or had dropped off the leaves due to at-tacks by the ants, L. sakagamii or F. japonica at this site,but no larvae were observed being eaten by naturalenemies except for ants (Table 2). At site B, survivalrates on both treatment and control trees decreased fromday 1 to 4, and thereafter decreased gradually until theend of the experiment. Predation by an unidentifiedcarabid larvae (Carabidae sp., the same species statedabove), and ants (Camponotus japonicus) was observedat this site (Table 2). At both sites A and B, significantdifferences in the larval survival rate between treatmentand control trees were detected on every day of obser-vation (Fig. 3).
At site C, average survival rates showed almost sim-ilar values between treatment and control trees on eachday of observation (Fig. 3). Survival rates graduallydecreased from day 1 to 4, and thereafter survival rateson both trees were under 10%; they decreased graduallythereafter (Fig. 3). At site C, no significant differences inthe larval survival rate between treatment and controltrees were detected on any day of observation (Fig. 3).At this site, similar to our observations for early instarlarvae, we observed many larvae being killed by wasps(P. snelleni) (Table 2). More larvae were observed beingkilled by wasps in 2001 than in 1999 and 2000. We alsofound that one larva was eaten by a frog and one larvaby a spider in 1999 (Table 2). We did not observe anylarvae being killed by ants.
Average survival rates of middle instar larvae oncontrol trees were lowest at site C (2.2%), intermediateat site A (4.4%) and highest at site B (16%) (Fig. 3); thisresult was different from the experiment in 2000 and theexperiment for early instar larvae in 2001.
Ant species and the number of individuals on test trees
In total, seven species of ants were found at the threesites (Table 3). At sites A, B and C, we observed three,
Table 3 Average number (±SD) (n = 65) of ants climbing a test tree in a 3-min period
Ant species Site Pa
A B C
Lasius sakagamii 27.11 ± 34.68 –Lasius niger – 6.48 ± 14.85 –Pristomyrmex pungens – 3.62 ± 7.43 –Formica japonica 1.98 ± 3.39 0.22 ± 0.52 –Camponotus japonicus 0.05 ± 0.21 0.02 ± 0.12 0.05 ± 0.21Paratrechina flavipes – 1.34 ± 2.09 –Monomorium intrudens – 0.45 ± 0.98 2.48 ± 3.51Average total numberof ants except forP. flavipes and M. intrudens
29.14 ± 33.67a 10.32 ± 15.23b 0.05 ± 0.21c F = 44.74,P < 0.001
Within the total number of five ant species (Lasius sakagamii, Lasius niger, Pristomyrmex pungens, Formica japonica and Camponotusjaponicus), values followed by different letters are significantly different [Tukey–Kramer multiple comparisons, P < 0.05, total number (x)of the five ant species on a test tree at each observation date was log(x + 1)-transformed]. –: Not observeda One-way ANOVA
Fig. 3 Survivorship curves for middle instar larvae of Sasakiacharonda on host trees treated with Tanglefoot and control trees atsites A, B and C. Third instar larvae were inoculated just aftermolting on 26 July 2001, and the number of surviving individualson each tree was counted 1, 4, 9 and 16 days after the inoculation.*P < 0.05, **P < 0.01. NS not significant
414
six and two species of ants, and L. sakagamii, L. nigerand Monomorium intrudens were the dominant species,respectively. Among the seven species, five ant species(L. sakagamii, L. niger, P. pungens, F. japonica and C.japonicus) were seen to prey on larvae of S. charonda,and the remaining two species (Paratrechina flavipes andM. intrudens) were not seen. The average for the totalnumber of the five ant species per tree observed over a 3-min period is shown at the bottom of Table 3. Theaverage for the total number of ants was highest at siteA (29.14), intermediate at site B (10.32) and lowest atsite C (0.05); the differences in the average of the totalnumber in all pairwise comparisons among the threesites were significant (Table 3).
Aphid species that provided ants with honeydewand the density of each species
All aphid species found in this study were S. celti, Aphisfabae and Aphis spiraecola. S. celti and A. spiraecolawere found at site A, S. celti and A. fabae at site B andonly S. celti was found at site C (Table 4). Among thesespecies, S. celti was most abundant at each site. Weobserved that each aphid species provided honeydew to
ants at sites A and B, but we could not confirm this atsite C. The mean total number of aphid species per 200leaves was greatest at site A (310.4), intermediate at siteB (62.7) and lowest at site C (15.4); the differences in thevalue for all pairwise comparisons among the three siteswere significant (Table 4).
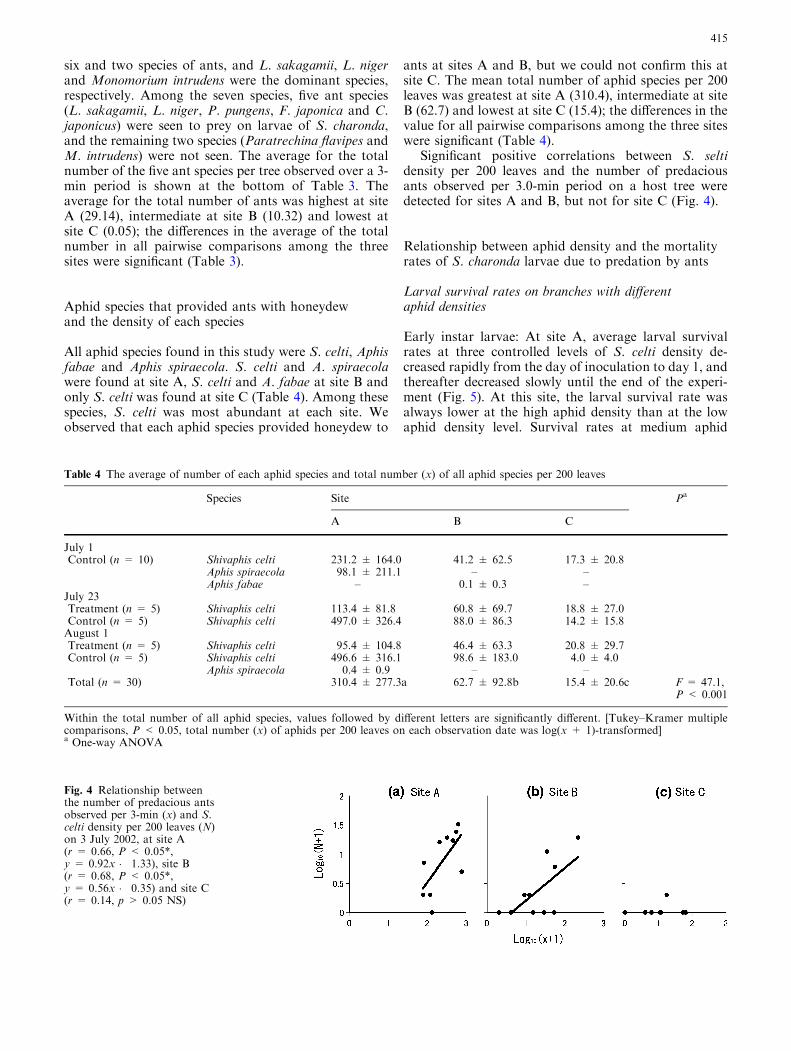
Significant positive correlations between S. seltidensity per 200 leaves and the number of predaciousants observed per 3.0-min period on a host tree weredetected for sites A and B, but not for site C (Fig. 4).
Relationship between aphid density and the mortalityrates of S. charonda larvae due to predation by ants
Larval survival rates on branches with differentaphid densities
Early instar larvae: At site A, average larval survivalrates at three controlled levels of S. celti density de-creased rapidly from the day of inoculation to day 1, andthereafter decreased slowly until the end of the experi-ment (Fig. 5). At this site, the larval survival rate wasalways lower at the high aphid density than at the lowaphid density level. Survival rates at medium aphid
Fig. 4 Relationship betweenthe number of predacious antsobserved per 3-min (x) and S.celti density per 200 leaves (N)on 3 July 2002, at site A(r = 0.66, P < 0.05*,y = 0.92x � 1.33), site B(r = 0.68, P < 0.05*,y = 0.56x � 0.35) and site C(r = 0.14, p > 0.05 NS)
Table 4 The average of number of each aphid species and total number (x) of all aphid species per 200 leaves
Species Site Pa
A B C
July 1Control (n = 10) Shivaphis celti 231.2 ± 164.0 41.2 ± 62.5 17.3 ± 20.8
Aphis spiraecola 98.1 ± 211.1 – –Aphis fabae – 0.1 ± 0.3 –
July 23Treatment (n = 5) Shivaphis celti 113.4 ± 81.8 60.8 ± 69.7 18.8 ± 27.0Control (n = 5) Shivaphis celti 497.0 ± 326.4 88.0 ± 86.3 14.2 ± 15.8August 1Treatment (n = 5) Shivaphis celti 95.4 ± 104.8 46.4 ± 63.3 20.8 ± 29.7Control (n = 5) Shivaphis celti 496.6 ± 316.1 98.6 ± 183.0 4.0 ± 4.0
Aphis spiraecola 0.4 ± 0.9 – –Total (n = 30) 310.4 ± 277.3a 62.7 ± 92.8b 15.4 ± 20.6c F = 47.1,
P < 0.001
Within the total number of all aphid species, values followed by different letters are significantly different. [Tukey–Kramer multiplecomparisons, P < 0.05, total number (x) of aphids per 200 leaves on each observation date was log(x + 1)-transformed]a One-way ANOVA
415
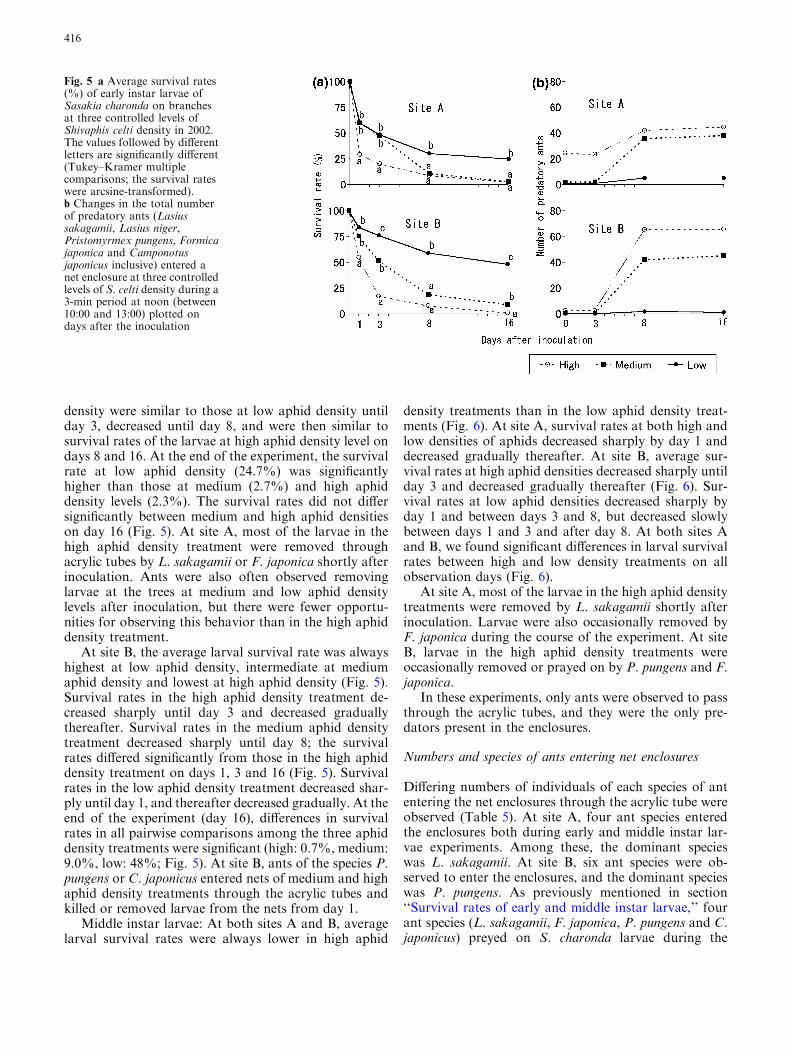
density were similar to those at low aphid density untilday 3, decreased until day 8, and were then similar tosurvival rates of the larvae at high aphid density level ondays 8 and 16. At the end of the experiment, the survivalrate at low aphid density (24.7%) was significantlyhigher than those at medium (2.7%) and high aphiddensity levels (2.3%). The survival rates did not differsignificantly between medium and high aphid densitieson day 16 (Fig. 5). At site A, most of the larvae in thehigh aphid density treatment were removed throughacrylic tubes by L. sakagamii or F. japonica shortly afterinoculation. Ants were also often observed removinglarvae at the trees at medium and low aphid densitylevels after inoculation, but there were fewer opportu-nities for observing this behavior than in the high aphiddensity treatment.
At site B, the average larval survival rate was alwayshighest at low aphid density, intermediate at mediumaphid density and lowest at high aphid density (Fig. 5).Survival rates in the high aphid density treatment de-creased sharply until day 3 and decreased graduallythereafter. Survival rates in the medium aphid densitytreatment decreased sharply until day 8; the survivalrates differed significantly from those in the high aphiddensity treatment on days 1, 3 and 16 (Fig. 5). Survivalrates in the low aphid density treatment decreased shar-ply until day 1, and thereafter decreased gradually. At theend of the experiment (day 16), differences in survivalrates in all pairwise comparisons among the three aphiddensity treatments were significant (high: 0.7%, medium:9.0%, low: 48%; Fig. 5). At site B, ants of the species P.pungens or C. japonicus entered nets of medium and highaphid density treatments through the acrylic tubes andkilled or removed larvae from the nets from day 1.
Middle instar larvae: At both sites A and B, averagelarval survival rates were always lower in high aphid
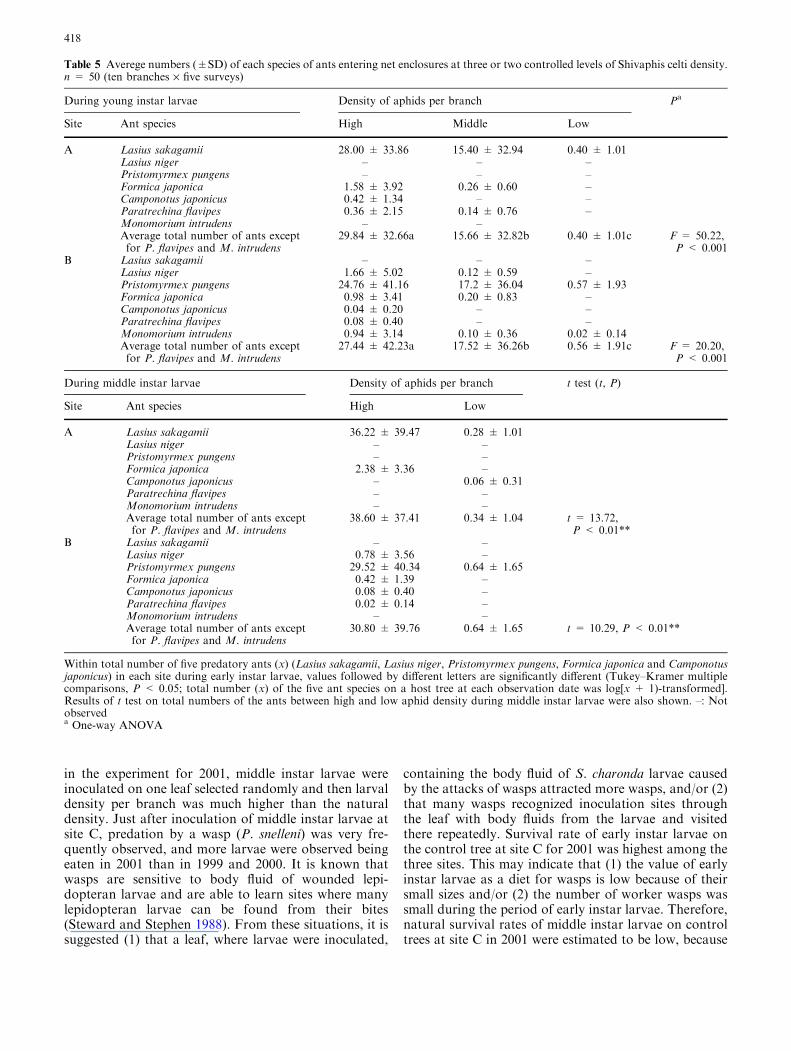
density treatments than in the low aphid density treat-ments (Fig. 6). At site A, survival rates at both high andlow densities of aphids decreased sharply by day 1 anddecreased gradually thereafter. At site B, average sur-vival rates at high aphid densities decreased sharply untilday 3 and decreased gradually thereafter (Fig. 6). Sur-vival rates at low aphid densities decreased sharply byday 1 and between days 3 and 8, but decreased slowlybetween days 1 and 3 and after day 8. At both sites Aand B, we found significant differences in larval survivalrates between high and low density treatments on allobservation days (Fig. 6).
At site A, most of the larvae in the high aphid densitytreatments were removed by L. sakagamii shortly afterinoculation. Larvae were also occasionally removed byF. japonica during the course of the experiment. At siteB, larvae in the high aphid density treatments wereoccasionally removed or prayed on by P. pungens and F.japonica.
In these experiments, only ants were observed to passthrough the acrylic tubes, and they were the only pre-dators present in the enclosures.
Numbers and species of ants entering net enclosures
Differing numbers of individuals of each species of antentering the net enclosures through the acrylic tube wereobserved (Table 5). At site A, four ant species enteredthe enclosures both during early and middle instar lar-vae experiments. Among these, the dominant specieswas L. sakagamii. At site B, six ant species were ob-served to enter the enclosures, and the dominant specieswas P. pungens. As previously mentioned in section‘‘Survival rates of early and middle instar larvae,’’ fourant species (L. sakagamii, F. japonica, P. pungens and C.japonicus) preyed on S. charonda larvae during the
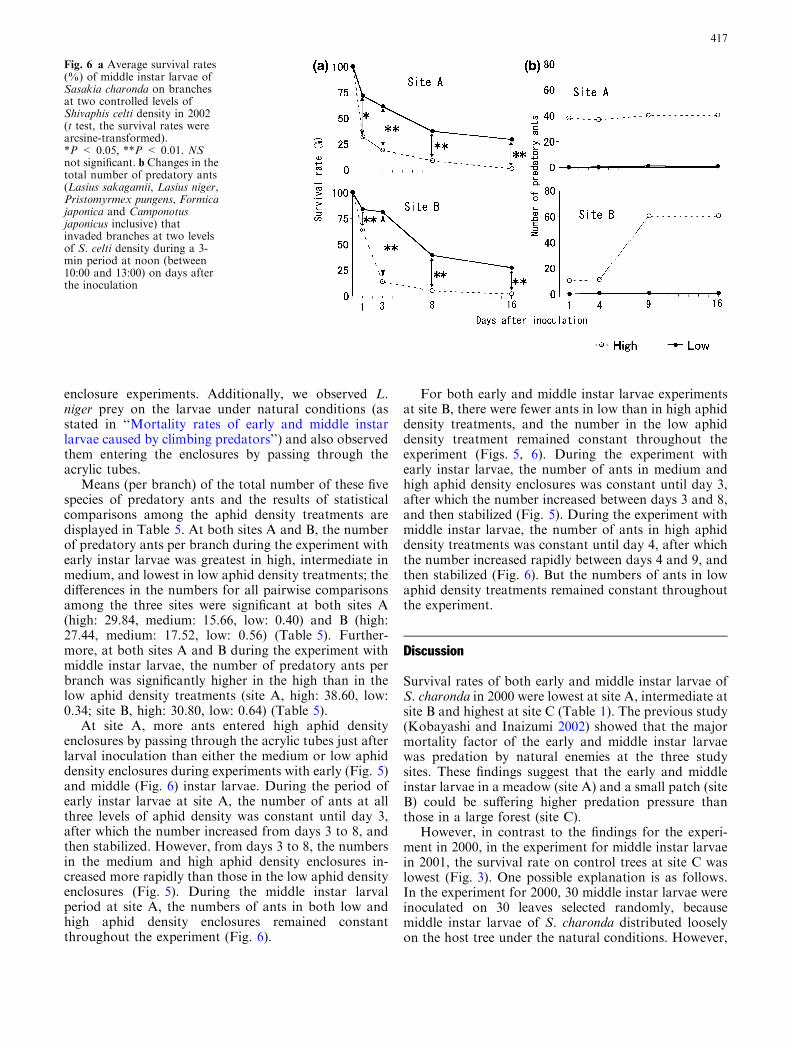
Fig. 5 a Average survival rates(%) of early instar larvae ofSasakia charonda on branchesat three controlled levels ofShivaphis celti density in 2002.The values followed by differentletters are significantly different(Tukey–Kramer multiplecomparisons; the survival rateswere arcsine-transformed).b Changes in the total numberof predatory ants (Lasiussakagamii, Lasius niger,Pristomyrmex pungens, Formicajaponica and Camponotusjaponicus inclusive) entered anet enclosure at three controlledlevels of S. celti density during a3-min period at noon (between10:00 and 13:00) plotted ondays after the inoculation
416
enclosure experiments. Additionally, we observed L.niger prey on the larvae under natural conditions (asstated in ‘‘Mortality rates of early and middle instarlarvae caused by climbing predators’’) and also observedthem entering the enclosures by passing through theacrylic tubes.
Means (per branch) of the total number of these fivespecies of predatory ants and the results of statisticalcomparisons among the aphid density treatments aredisplayed in Table 5. At both sites A and B, the numberof predatory ants per branch during the experiment withearly instar larvae was greatest in high, intermediate inmedium, and lowest in low aphid density treatments; thedifferences in the numbers for all pairwise comparisonsamong the three sites were significant at both sites A(high: 29.84, medium: 15.66, low: 0.40) and B (high:27.44, medium: 17.52, low: 0.56) (Table 5). Further-more, at both sites A and B during the experiment withmiddle instar larvae, the number of predatory ants perbranch was significantly higher in the high than in thelow aphid density treatments (site A, high: 38.60, low:0.34; site B, high: 30.80, low: 0.64) (Table 5).
At site A, more ants entered high aphid densityenclosures by passing through the acrylic tubes just afterlarval inoculation than either the medium or low aphiddensity enclosures during experiments with early (Fig. 5)and middle (Fig. 6) instar larvae. During the period ofearly instar larvae at site A, the number of ants at allthree levels of aphid density was constant until day 3,after which the number increased from days 3 to 8, andthen stabilized. However, from days 3 to 8, the numbersin the medium and high aphid density enclosures in-creased more rapidly than those in the low aphid densityenclosures (Fig. 5). During the middle instar larvalperiod at site A, the numbers of ants in both low andhigh aphid density enclosures remained constantthroughout the experiment (Fig. 6).
For both early and middle instar larvae experimentsat site B, there were fewer ants in low than in high aphiddensity treatments, and the number in the low aphiddensity treatment remained constant throughout theexperiment (Figs. 5, 6). During the experiment withearly instar larvae, the number of ants in medium andhigh aphid density enclosures was constant until day 3,after which the number increased between days 3 and 8,and then stabilized (Fig. 5). During the experiment withmiddle instar larvae, the number of ants in high aphiddensity treatments was constant until day 4, after whichthe number increased rapidly between days 4 and 9, andthen stabilized (Fig. 6). But the numbers of ants in lowaphid density treatments remained constant throughoutthe experiment.
Discussion
Survival rates of both early and middle instar larvae ofS. charonda in 2000 were lowest at site A, intermediate atsite B and highest at site C (Table 1). The previous study(Kobayashi and Inaizumi 2002) showed that the majormortality factor of the early and middle instar larvaewas predation by natural enemies at the three studysites. These findings suggest that the early and middleinstar larvae in a meadow (site A) and a small patch (siteB) could be suffering higher predation pressure thanthose in a large forest (site C).
However, in contrast to the findings for the experi-ment in 2000, in the experiment for middle instar larvaein 2001, the survival rate on control trees at site C waslowest (Fig. 3). One possible explanation is as follows.In the experiment for 2000, 30 middle instar larvae wereinoculated on 30 leaves selected randomly, becausemiddle instar larvae of S. charonda distributed looselyon the host tree under the natural conditions. However,
Fig. 6 a Average survival rates(%) of middle instar larvae ofSasakia charonda on branchesat two controlled levels ofShivaphis celti density in 2002(t test, the survival rates werearcsine-transformed).*P < 0.05, **P < 0.01. NSnot significant. b Changes in thetotal number of predatory ants(Lasius sakagamii, Lasius niger,Pristomyrmex pungens, Formicajaponica and Camponotusjaponicus inclusive) thatinvaded branches at two levelsof S. celti density during a 3-min period at noon (between10:00 and 13:00) on days afterthe inoculation
417
in the experiment for 2001, middle instar larvae wereinoculated on one leaf selected randomly and then larvaldensity per branch was much higher than the naturaldensity. Just after inoculation of middle instar larvae atsite C, predation by a wasp (P. snelleni) was very fre-quently observed, and more larvae were observed beingeaten in 2001 than in 1999 and 2000. It is known thatwasps are sensitive to body fluid of wounded lepi-dopteran larvae and are able to learn sites where manylepidopteran larvae can be found from their bites(Steward and Stephen 1988). From these situations, it issuggested (1) that a leaf, where larvae were inoculated,
containing the body fluid of S. charonda larvae causedby the attacks of wasps attracted more wasps, and/or (2)that many wasps recognized inoculation sites throughthe leaf with body fluids from the larvae and visitedthere repeatedly. Survival rate of early instar larvae onthe control tree at site C for 2001 was highest among thethree sites. This may indicate that (1) the value of earlyinstar larvae as a diet for wasps is low because of theirsmall sizes and/or (2) the number of worker wasps wassmall during the period of early instar larvae. Therefore,natural survival rates of middle instar larvae on controltrees at site C in 2001 were estimated to be low, because
Table 5 Averege numbers (±SD) of each species of ants entering net enclosures at three or two controlled levels of Shivaphis celti density.n = 50 (ten branches · five surveys)
During young instar larvae Density of aphids per branch Pa
Site Ant species High Middle Low
A Lasius sakagamii 28.00 ± 33.86 15.40 ± 32.94 0.40 ± 1.01Lasius niger – – –Pristomyrmex pungens – – –Formica japonica 1.58 ± 3.92 0.26 ± 0.60 –Camponotus japonicus 0.42 ± 1.34 – –Paratrechina flavipes 0.36 ± 2.15 0.14 ± 0.76 –Monomorium intrudens – –Average total number of ants exceptfor P. flavipes and M. intrudens
29.84 ± 32.66a 15.66 ± 32.82b 0.40 ± 1.01c F = 50.22,P < 0.001
B Lasius sakagamii – – –Lasius niger 1.66 ± 5.02 0.12 ± 0.59 –Pristomyrmex pungens 24.76 ± 41.16 17.2 ± 36.04 0.57 ± 1.93Formica japonica 0.98 ± 3.41 0.20 ± 0.83 –Camponotus japonicus 0.04 ± 0.20 – –Paratrechina flavipes 0.08 ± 0.40 – –Monomorium intrudens 0.94 ± 3.14 0.10 ± 0.36 0.02 ± 0.14Average total number of ants exceptfor P. flavipes and M. intrudens
27.44 ± 42.23a 17.52 ± 36.26b 0.56 ± 1.91c F = 20.20,P < 0.001
During middle instar larvae Density of aphids per branch t test (t, P)
Site Ant species High Low
A Lasius sakagamii 36.22 ± 39.47 0.28 ± 1.01Lasius niger – –Pristomyrmex pungens – –Formica japonica 2.38 ± 3.36 –Camponotus japonicus – 0.06 ± 0.31Paratrechina flavipes – –Monomorium intrudens – –Average total number of ants exceptfor P. flavipes and M. intrudens
38.60 ± 37.41 0.34 ± 1.04 t = 13.72,P < 0.01**
B Lasius sakagamii – –Lasius niger 0.78 ± 3.56 –Pristomyrmex pungens 29.52 ± 40.34 0.64 ± 1.65Formica japonica 0.42 ± 1.39 –Camponotus japonicus 0.08 ± 0.40 –Paratrechina flavipes 0.02 ± 0.14 –Monomorium intrudens – –Average total number of ants exceptfor P. flavipes and M. intrudens
30.80 ± 39.76 0.64 ± 1.65 t = 10.29, P < 0.01**
Within total number of five predatory ants (x) (Lasius sakagamii, Lasius niger, Pristomyrmex pungens, Formica japonica and Camponotusjaponicus) in each site during early instar larvae, values followed by different letters are significantly different (Tukey–Kramer multiplecomparisons, P < 0.05; total number (x) of the five ant species on a host tree at each observation date was log[x + 1)-transformed].Results of t test on total numbers of the ants between high and low aphid density during middle instar larvae were also shown. –: Notobserveda One-way ANOVA
418

the inoculation method was controversial and thus thenatural mortality of middle instar larvae was overesti-mated due to predation by wasps. Although there is apossibility of underestimation of survival rates of middleinstar larvae at site C, it is suggested that natural sur-vival rates on early and middle instar larvae of S.charonda at site C might be higher than at sites A and B.
It is known that ants are a major cause of mortality oflepidopteran larvae at early stages of their development(Furuta 1968; Matsui and Kokubo 1974; Hondo 1981;Watanabe 1976, 1981), and wasps are also major mor-tality factors of lepidopteran larvae (Steward and Ste-phen 1988). The previous study (Kobayashi andInaizumi 2002) (Table 2) reported that, at site C, themortality rates of early instar larvae of S. charondapreyed on by medium-sized natural enemies such aswasps were relatively higher than at sites A and B,whereas those preyed on by small-sized natural enemiessuch as ants at sites A and B were relatively higher thansite C. Our results (Figs. 2, 3; Table 2) suggest thatmortalities of early and middle instar larvae of S. char-onda due to predation by tree climbing predators such asants and carabid larvae were higher at sites A and Bthan at site C, whereas mortalities of the larvae due topredation by wasps were higher at site C than at site Aand B.
We observed that all aphid species provided ants withthe honeydew at both sites A and B. It is known thathoneydew produced by homopteran insects such asaphids is one of the major food sources for ants (Room1972; Buckley 1987; Ito and Higashi 1991; Sakata 2000).Fritz (1982) reported similar results, indicating thatthere was a significant positive correlation between thedensity of membracid bugs, which provide ants withtheir honeydew, and the number of ants per host tree.Results of both aphid density and the number of antsclimbing a test tree (Tables 3, 4) and significant positivecorrelation between aphid density and the number ofants at sites A and B (Fig. 4) suggest that the aphiddensity on C. sinensis trees may directly affect the densityof predatory ants climbing on the trees in meadows orsmall patches.
It is reported that, when ants attend homopteraninsects, they eat (Nickerson et al. 1977; Laine and Nie-mela 1980; Buckley 1987) or exclude (Messina 1981; Itoand Higashi 1991) other phytophagous insects such aslepidopteran larva that are feeding on the same hostplants. Results of the net bag experiments (Table 5;Figs. 5, 6) indicated that, in the meadows and smallpatches in the study area, aphid densities on C. sinensistrees were relatively high, promoting an increase in thenumber of ants climbing on test trees, and as a result,leading to an increase in the mortality rates of early andmiddle instar S. charonda larvae.
Our findings can also explain the previous finding ofa significant positive correlation between the density ofoverwintering larvae per host tree and the area of sec-ondary deciduous broadleaf forests within a radius of100 m of the host tree (Kobayashi et al. 2004), sug-
gesting that larval density of S. charonda was negativelyaffected by fragmentation of secondary deciduousbroadleaf forests. That is, it is considered that thefragmentation of secondary forests enhanced the num-ber of aphids on test trees, and thus led to an increase inthe number of predatory ants climbing the trees, andfinally resulted in the lower larval densities due to theirincreased mortality rates by the ants. In fact, it is wellknown that habitats of L. sakagamii, F. japonica and C.japonicus are mainly open lands or fragmented forests(Hayahi et al. 1994; Touyama and Nakagoshi 1994:Terayama 1997), supporting the above results andspeculation.
Until now, the decline and extinction of organisms inresponse to fragmentation of habitat within a specieshave been mainly explained by metapopulationdynamics. In particular, many studies have clarified themetapopulation dynamics of some endangered grass-land butterfly species (e.g., Thomas and Harrison 1992;Thomas et al. 1992; Thomas and Jones 1993; Hill et al.1996). However, recently, changes in the biotic inter-actions between a threatened species and other organ-isms have been pointed out as another major factor inthe decline of the threatened species in response tohabitat fragmentation. For example, it has been re-ported that extinction of some seed plants in fragmentedhabitats resulted from the disappearance of pollinatinginsects due to the fragmented habitats, which have amutually beneficial relationship with the seed plants(Jennersten 1988; Rathcke and Jules 1993; Aizen andFeinsinger 1994; Washitani 1996). On the other hand,the cause of decline of small birds, whose habitats wereforest interiors, was reported to be an increase in pre-dation by natural enemies made possible by forestfragmentation (Gates and Gysel 1978; Wilcove 1985). Inthis case, these enemies had previously not posed aserious threat to the small birds, because they usuallysought their prey at the forest edges rather than insidethe forest. In the present study, we also showed that thedecline of a butterfly species in response to fragmenta-tion of habitat can be explained through the alteredbiotic interactions between the butterfly species andother organisms. Therefore, to predict the long-termeffects of fragmentation of secondary deciduousbroadleaf forests on S. charonda, it is very important toconsider not only the well-known factors affecting theextinction of small local populations such as demo-graphic stochasticity, Allee effect, environmental sto-chasticity, catastrophes, genetic stochasticity andinbreeding depression (e.g., Hunter 1996), but also theeffect of the three-way biotic interactions as shown inthe present study. Consequently, our study could clarifyone of the mechanisms underlying decreases in S.charonda population densities due to fragmentation ofsecondary deciduous broadleaf forests. Perhaps in thefuture, the analytical approaches and results of thisstudy will be very helpful to our understanding of theeffects of fragmentation of secondary forests on avariety of organisms inhabiting them.
419

Acknowledgments We express sincere thanks to Drs. TakenariInoue and Ken-ichi Ozaki of Forestry and Forest ProductsResearch Institute for their useful comments on this study.
References
Aizen AM, Feinsinger P (1994) Forest fragmentation, pollination,and plant reproduction in a chaco dry forest. Argentina Ecol-ogy 75:330–351
Buckley RC (1987) Ant–plant–homopteran interactions. Adv EcolRes 16:53–85
Fritz RS (1982) An ant–tree hopper mutualism: effects of Formicasubsericea on the survival of Vanduzea arquata. Ecol Entomol7:267–276
Fukuda H, Hama E, Kuzuya K, Takahashi A, Takahashi M,Tanaka B, Tanaka H, Wakabayashi M, Watanabe Y (1983)The butterflies of Japan (II) (in Japanese). Hoikusha, Osaka
Furuta K (1968) The relationship between population density andmortality in the range of latency of Dendrolimus spectabilisBUTLER (in Japanese with English summary). Jpn J ApplEntomol Zool 12:129–136
Gates JE, Gysel L (1978) Avian nest dispersion and fledging successin field-forest ecotones. Ecology 59:871–883
Hayahi K, Hagimoto M, Wada K (1994) Temporal change inextranidal activity pattern of formicid ant, Formica japonica(Formicidae: Hymenoptera) (in Japanese with English sum-mary). Jpn J Ecol 44:171–179
Higuchi H, Sato K (1987) Distribution of Sasakia charonda He-witson in Tochigi Prefecture (in Japanese with English sum-mary). Bull Tochigi Prefect Mus 4:7–14
Hill JK, Thomas CD, Lewis OT (1996) Effects of habitat patch sizeand isolation on dispersal by Hesperia comma butterflies:implications for metapopulation structure. J Anim Ecol 65:725–735
Hondo M (1981) Mortality factors and mortality process of colo-nial stages of the mulberry tiger moth, Spilarctia imparilisBUTLER (Lepidptera: Arctiidae) (in Japanese with Englishsummary). Jpn J Appl Entomol Zool 25:219–228
Hunter Jr ML (1996) Fundamentals of conservation biology.Blackwell, Massachusetts
Ito F, Higashi S (1991) An indirect mutualism between oaks andwood ants via aphids. J Anim Ecol 60: 463–470
Jennersten O (1988) Pollination in Dianthus deltoids (Caryophyll-aceae): effects of habitat fragmentation on visitation and seedset. Conserv Biol 2:359–366
Kobayashi T, Inaizumi M (1999) The rate and factors of mortalityfrom the completion of overwintering in larvae to emergence ofadults in a Nymphalid, Sasakia charonda (Hewitson) (in Japa-nese with English summary). Jpn J Entomol (NS) 2:57–68
Kobayashi T, Inaizumi M (2000) Change of the mortality rates andfactors of larvae after hibernation and pupae of a Nymphalidbutterfly, Sasakia charonda (Hewitson), in relation to size andcommunity structure of the forest surrounding the host plants(in Japanese with English summary). Jpn J Entomol (NS)3:125–138
Kobayashi T, Inaizumi M (2002) Difference in mortality rates andfactors of immatures before overwintering of the NymphalidButterfly, Sasakia charonda (Hewitson), in relation to vegeta-tion shift surrounding the host plants (in Japanese with Englishsummary). Jpn J Entomol 5:35–49
Kobayashi T, Inaizumi M (2003) Mortality factors of overwinter-ing larvae of the Nymphalid butterfly, Sasakia charonda He-witson in Mooka city in Tochigi prefecture. Trans LepidopterolSoc Jpn 54:20–30
Kobayashi T, Tanimoto T, Kitahara M (2004) Relationship be-tween rate of forest area and the densities of the nymphalidbutterfly, Sasakia charonda Hewitson, and the host trees, Celtissinensis Persoon (in Japanese with English summary). Jpn JConserv Ecol 9:1–12
Laine KJ, Niemela P (1980) The influence of ants on the survival ofmountain birches during an Oporinia autumnata (Lep., Geo-metridae) outbreak. Oecologia 47:39–42
MacGarvin M (1982) Species–area relationships of insects on hostplants: herbivores on rosebay willowherb. J Anim Ecol 51:207–223
Matsui H, Kokubo A (1974) Observations on egg masses of thepine-moth, Dendrolimus spectabilis BUTLER attacked by ants(In Japanese). J Jpn Forest Soc 56:182–184
Messina FJ (1981) Plant protection as a consequence of an ant–membracid mutualism: interactions on goldenrod, Salidago sp.Ecology 62:1433–1440
Ministry of the Environment (2006) Threatened wildlife ofJapan—Red Data Book, 2nd edn, vol 5, Insecta. Japan WildlifeReseach Center, Taito-ku, Japan, 246 pp
Nickerson JC, Rolph Kay CA, Buschman LL, Whitcomb WH(1977) The presence of Spissistilus festinus as a factor affectingegg predation by ants in soybeans. Fla Entomol 60:193–199
Rathcke BJ, Jules ES (1993) Habitat fragmentation and plant–pollinator interactions. Curr Sci 65:273–277
Robertson PA, Clarke SA, Warren MS (1995) Woodland man-agement and butterfly diversity. In: Pullin AS (eds) Ecology andconservation of butterflies. Chapman & Hall, London, pp 113–122
Room PM (1972) The fauna of the mistletoe Tapinanthus bangw-ensis (Engl. & K. Krause) growing on cocoa in Ghana: rela-tionship between fauna and mistletoe. J Anim Ecol 41:611–621
Sakata H (2000) Mechanisms restricting ant–aphid mutualism: antforaging strategy and interference among sugar source (inJapanese with English summary). Jpn J Ecol 50:13–22
Steward VB, Stephen FM (1988) Predation by wasps on lepi-dopteran larvae in an Ozark forest canopy Ecol Entomol 13:81–86
Terayama M (1997) The environmental protection from the view-point of biodiversity conservation—focus on the study of antcommunity (in Japanese). Seibutsu-Kagaku 49: 75–83
Thomas CD, Harrison S (1992) Spatial dynamics of a patchilydistributed butterfly species. J Anim Ecol 61:437–446
Thomas CD, Jones TM (1993) Partial recovery of a skipper but-terfly (Hesperia comma) from population refuges: lessons forconservation in a fragmented landscape. J Anim Ecol 62:472–481
Thomas CD, Thomas JA, Warren MS (1992) Distributions ofoccupied and vacant butterfly habitats in fragmented land-scapes. Oecologia 92:563–567
Touyama Y, Nakagoshi N (1994) A comparison of soil arthropodfauna in coniferous plantations and secondary forests (in Jap-anese with English summary). Jpn J Ecol 50:13–22
Washitani I (1996) Predicted genetic consequences of strong fer-tility selection due to pollinator loss in an island population ofPrimula sieboldii. Conserv Biol 10:59–64
Watanabe M (1976) A preliminary study on population dynamicsof the swallowtail butterfly, Papilio xuthus L. in a deforestedarea. Res Popul Ecol 17:200–210
Watanabe M (1981) Population dynamics of the swallowtail but-terfly, Papilio xuthus L., in a deforested area. Res Popul Ecol23:74–93
Wilcove DS (1985) Nest predation in forest tracts and the decline ofmigratory songbirds. Ecology 66:1211–1214
Wilcox B, Murphy D (1985) Conservation strategy: the effects offragmentation on extinction. Am Nat 125: 879–887
420
Copyright © 2022 FDOKUMEN


![Der Herrscher in der Stadt (The Emperor/King in the Medieval [Italian] Town)](https://static.fdokumen.com/doc/165x107/631c02b8665120b3330ba551/der-herrscher-in-der-stadt-the-emperorking-in-the-medieval-italian-town.jpg)

















