
Effects of geothermal effluents on macrobenthic communities in a pristine sub-arctic lake.
12
146 Inland Waters (2011) 1, pp. 146-157 © International Society of Limnology 2011 DOI: 10.5268/IW-1.3.363 Article Effects of geothermal effluents on macrobenthic communities in a pristine sub-arctic lake Sigurður S. Snorrason 1 , Hilmar J. Malmquist 2 *, Hrefna B. Ingólfsdóttir 1 , Þórey Ingimundardóttir 2 , and Jón S. Ólafsson 1,3 1 University of Iceland, Askja, Sturlugata 7, IS–101 Reykjavík, Iceland 2 Natural History Museum of Kópavogur, Hamraborg 6a, IS–200 Kópavogur, Iceland 3 Insititute of Freshwater Fisheries, Keldnaholt, IS–112 Reykjavík, Iceland * Corresponding author email: [email protected] Received 11 October 2010; accepted 22 July 2011; published 31 October 2011 Abstract We studied the effects of warm effluents from the Nesjavellir geothermal power plant on benthic invertebrate communities along a depth gradient of 4 m in lake Þingvallavatn, Iceland. Four study sites were selected: 2 heat- affected sites and 2 cold reference sites. Thermal pollution was detected down to 0.4 m at both heat-affected sites, which were 7–12 °C higher than at the same depth at the reference sites. Sustained temperatures ≥27 °C, which is 10–17 °C above ambient water temperatures, seems to be detrimental to the gastropod Radix peregra and several of the littoral chironomid species. Such effects were, however, confined to a small area inside a sheltered rift pool at one of the heat-affected sites. Moderate warming up to ~10 °C above ambient lake temperature in summer, seems to benefit R. peregra, potentially through both a higher survival rate and a shorter life cycle. At the heat-affected sites, sustained high temperatures transformed a relatively diverse chironomid community to a species-poor community, dominated by Paratanytarsus sp. and Cricotopus sylvestris. In contrast, the chironomid Eukiefferiella minor was much more prominent at one of the reference sites. To separate thermal from biological effects, more studies are needed on species- specific attributes, including time-series data on growth, survival, and reproduction of the main littoral zoobenthic species. Although our research examined short-term responses of aquatic invertebrates to local thermal stress, our results may also be relevant to long-term responses of freshwater ecosystems at high latitudes to climate change and warming. Key words: Chironomidae, geothermal effects, Iceland, lake zoobenthos, Radix peregra, thermal stress Introduction With increasing awareness of the negative effects of CO 2 emissions from burning fossil fuels, the pressure to use clean and renewable energy sources low in CO 2 such as hydropower and geothermal energy has increased worldwide (IEA 2003, 2009). Utilization of hydropower and particularly geothermal energy has been praised as environmentally friendly, low impact industry (e.g., Rybach 2005, Valfells et al. 2005). While this may be true to some extent, various environmental impacts from geothermal resources are of concern (Arnórsson 2004), such as airborne pollution, mainly sulphur gases H 2 S and SO 2 (e.g., Kristmannsdóttir and Ármannsson 2003); thermal pollution (e.g., Langford 1990); contamination by trace elements, some with ecotoxicological potentials such as arsenic (As), boron (B), and mercury (Hg; e.g., Aksoy al. 2009); and unsustainable, excessive energy production with reduced thermal capacity of the field under question (Axelsson et al. 2005). Due to the geology of Iceland, situated on the Mid- Atlantic Ocean ridge characterized by high volcanic activity and extensive high-energy geothermal systems, around 62% of the island’s primary energy needs in 2008
Transcript of Effects of geothermal effluents on macrobenthic communities in a pristine sub-arctic lake.

146
Inland Waters (2011) 1, pp. 146-157
© International Society of Limnology 2011
DOI: 10.5268/IW-1.3.363
Article
Effects of geothermal effluents on macrobenthic communities in a pristine sub-arctic lakeSigurður S. Snorrason1, Hilmar J. Malmquist2*, Hrefna B. Ingólfsdóttir1, Þórey Ingimundardóttir2, and Jón S. Ólafsson1,3
1 University of Iceland, Askja, Sturlugata 7, IS–101 Reykjavík, Iceland2 Natural History Museum of Kópavogur, Hamraborg 6a, IS–200 Kópavogur, Iceland3 Insititute of Freshwater Fisheries, Keldnaholt, IS–112 Reykjavík, Iceland* Corresponding author email: [email protected]
Received 11 October 2010; accepted 22 July 2011; published 31 October 2011
Abstract
We studied the effects of warm effluents from the Nesjavellir geothermal power plant on benthic invertebrate communities along a depth gradient of 4 m in lake Þingvallavatn, Iceland. Four study sites were selected: 2 heat- affected sites and 2 cold reference sites. Thermal pollution was detected down to 0.4 m at both heat-affected sites, which were 7–12 °C higher than at the same depth at the reference sites. Sustained temperatures ≥27 °C, which is 10–17 °C above ambient water temperatures, seems to be detrimental to the gastropod Radix peregra and several of the littoral chironomid species. Such effects were, however, confined to a small area inside a sheltered rift pool at one of the heat-affected sites. Moderate warming up to ~10 °C above ambient lake temperature in summer, seems to benefit R. peregra, potentially through both a higher survival rate and a shorter life cycle. At the heat-affected sites, sustained high temperatures transformed a relatively diverse chironomid community to a species-poor community, dominated by Paratanytarsus sp. and Cricotopus sylvestris. In contrast, the chironomid Eukiefferiella minor was much more prominent at one of the reference sites. To separate thermal from biological effects, more studies are needed on species-specific attributes, including time-series data on growth, survival, and reproduction of the main littoral zoobenthic species. Although our research examined short-term responses of aquatic invertebrates to local thermal stress, our results may also be relevant to long-term responses of freshwater ecosystems at high latitudes to climate change and warming.
Key words: Chironomidae, geothermal effects, Iceland, lake zoobenthos, Radix peregra, thermal stress
Introduction
With increasing awareness of the negative effects of CO2 emissions from burning fossil fuels, the pressure to use clean and renewable energy sources low in CO2 such as hydropower and geothermal energy has increased worldwide (IEA 2003, 2009). Utilization of hydropower and particularly geothermal energy has been praised as environmentally friendly, low impact industry (e.g., Rybach 2005, Valfells et al. 2005). While this may be true to some extent, various environmental impacts from geothermal resources are of concern (Arnórsson 2004),
such as airborne pollution, mainly sulphur gases H2S and SO2 (e.g., Kristmannsdóttir and Ármannsson 2003); thermal pollution (e.g., Langford 1990); contamination by trace elements, some with ecotoxicological potentials such as arsenic (As), boron (B), and mercury (Hg; e.g., Aksoy al. 2009); and unsustainable, excessive energy production with reduced thermal capacity of the field under question (Axelsson et al. 2005).
Due to the geology of Iceland, situated on the Mid- Atlantic Ocean ridge characterized by high volcanic activity and extensive high-energy geothermal systems, around 62% of the island’s primary energy needs in 2008

DOI: 10.5268/IW-1.3.363
147Effects of geothermal effluents on macrobenthic communities
Inland Waters (2011) 1, pp. 146-157
were from indigenous geothermal sources and 20% from hydropower (Björnsson 2010). The Nesjavellir geothermal field in southwest Iceland is a high enthalpy geothermal system within the Hengill central volcano, one of the largest high enthalpy geothermal complexes in Iceland (Gunnlaugsson et al. 2000). The area is situated within the catchment of Lake Þingvallavatn, the largest lake in Iceland, a cold, spring-fed, oligotrophic rift lake of high conservation value (Jónasson 1992, 1993, Jónasson and Hersteinsson 2011). For the past 30 years, warm wastewater from a geothermal power plant at Nesjavellir has been discharged into a nearby brook and then later into shallow sink wells and deep drill holes close by the power station (Gunnlaugsson et al. 2000, Zarandi and Ivarsson 2010). The warm effluents mix with groundwater, which flows through the porous lava bedrock before entering the southwest shore of Þingvallavatn (Ólafsson 1992, Wetang’ula and Snorrason 2005, Zarandi and Ivarsson 2010). The chemical constituents in both geothermal wastewater and biological samples at outlet and reference sites in Þingvallavatn have been analysed on several occasions since 1984 (Ólafsson 1992, Wetang’ula and Snorrason 2005, Zarandi and Ivarsson 2010). Both chemical and, in particular, thermal pollution have been observed in effluents at some of the outlet sites, especially after the start of electrical production in 1998. Chemical components that have increased include As, aluminium (Al), B, silicon dioxide (SiO2), sulphate (SO4), sodium (Na), potassium (K), and calcium (Ca; Wetang’ula and Snorrason 2005, Zarandi and Ivarsson 2010). Despite this pollution, with up to a 7-fold increase in concentration for As (0.7–4.97 μg L−1) at one heat-affected site (Zarandi and Ivarsson 2010), the concentrations of trace elements have in most cases been low and within acceptable inter-national environmental quality guidelines for the protection of aquatic life (Wetang’ula and Snorrason 2005, Zarandi and Ivarsson 2010). Nevertheless, from an ecotoxicological perspective, As, Al, and B have been considered to be potential threats to the biological communities at the outflow sites in the lake (Wetang’ula and Snorrason 2005). However, chemical analysis of biological samples, including sediment, plants, inverte-brates, and fish, collected at outflow and reference sites in the lake have revealed no effects that can be related to the wastewater effluents from the Nesjavellir cogeneration power plant (Wetang’ula and Snorrason 2005, Sturlaugs-son et al. 2009, Auðunsson et al. 2010).
Regardless of chemical pollution, extensive changes have occurred in thermal regimes at outflow sites in the lake, with a rise in temperature up to 30 °C above the ambient lake temperature at one heat-affected site (Wetang’ula and Snorrason 2005, Zarandi and Ivarsson 2010). This anthropogenic thermal impact of the
Nesjavellir cogeneration power plant presents an opportunity to study responses of aquatic organisms to the effects of thermal stress and changes in the environment. The study may also have a bearing on freshwater ecosystem responses at high latitudes in the context of climate change and warming of freshwater systems (Hughes 2000, Walther et al. 2002, Malmquist et al. 2009, Jeppesen et al. 2010, Woodward et al. 2010).
In this study we assess the short-term effects of elevated water temperature on benthic invertebrate communities in the littoral zone from heat-affected and unaffected reference sites in Þingvallavatn. Based on general physiological considerations, we assumed that elevated temperature would have local effects on the zoobenthos in at least 2 ways. First, at the species level we expected that elevated temperature would affect life history traits of eurythermic species (e.g., by accelerating growth rates and shortening generation times; Nebeker 1971, Lamberti and Resch 1983, Thomas and McClintock 1996). Second, at the community level we expected that elevated water temperature would increase the number and abundance of eurythermic species at the expense of stenothermic species (Walther et al. 2002, Daufresne et al. 2003).
Study area
The lake
Lake Þingvallavatn (Fig. 1.) is situated in southwest Iceland at 100.5 m a.s.l., with surface area 83 km2, lake volume 2.855 Gl, maximum depth 114 m, and mean depth 34 m (Aðalsteinsson et al. 1992). The catchment area is 1300 km2, 50% of which is characterized by permeable, postglacial basaltic lava bedrock. Around 90% of the lake inflow is cold (2.8–3.5 °C) subsurface spring water entering the lake via small inlets and large rifts, primarily at the northern shore (Aðalsteinsson et al. 1992, Ólafsson 1992). The mean discharge at the outlet river Efra-Sog in the southeast corner of the lake is 100 m3 s−1, and the calculated retention time is about 330 d with the water running from north to south toward the outlet (Aðalsteins-son et al. 1992).
Due to the cold spring-fed nature of the lake and its large volume, maximum water temperatures in late July–August usually only vary between 10 and 12 °C and rarely exceed 13 °C (Aðalsteinsson et al. 1992, Malmquist et al. 2010a, 2010b). Temperatures during November–May usually range between 0.1 and 5.0 °C throughout the whole lake. For the period 1951–2008 the lake was ice-covered an average of 73 d, with ice-on in the second week of January and ice-off usually in middle to late April (Aðalsteinsson et al. 1992, Sveinbjörnsson 2009). For the

148
DOI: 10.5268/IW-1.3.363
Sigurður Snorrason et al.
© International Society of Limnology 2011
past 10–15 years, levels of ice cover in Þingvallavatn have been low due to climate warming (Sveinbjörnsson 2009).
Before exploitation of the Nesjavellir geothermal field started in the late 1980s (Gunnlaugsson et al. 2000, Zarandi and Ivarsson 2010), warm ground water (7.5–10.7 °C) affected by natural geothermal run-off entered the lake as a warm spring outlet within the Varmagjá rift on the southwest lake shore (Ólafsson 1992, Aðalsteinsson et al. 1992). The natural groundwater flow through the Nesjahraun lava field was estimated to be ~1.6 m3 s−1 (Ólafsson 1992), but due to exploitation the outflow from the Nesjavellir geothermal field to Þingval-lavatn tripled between 1990 and 2000 (Gunnlaugsson et al. 2000).
Geothermal exploitation
Operation of the Nesjavellir geothermal power plant, 5–6 km south of Þingvallavatn (Fig 1), started in 1990, but test drilling and local use of hot water on the site began in 1946 (Zarandi and Ivarsson 2010). Three types of wastewaters are produced at the Nesjavellir power plant: condensate water and separator water derived from high enthalpy wells, and heated fresh groundwater derived from shallow cold water wells. The Nesjavellir power plant produces electricity in steam turbines and hot water for space heating by heating cold groundwater in heat exchangers (Zarandi and Ivarsson 2010). The cold water (5–7 °C, ~2000 L s−1) to be heated is pumped from a shallow freshwater aquifer in the lava field close to the southwest shore of Þingvallavatn (Fig. 1).
In 2003 when the present study was undertaken, the discharged effluents from the Nesjavellir power plant consisted of used and unused brine (46–100 °C, 115–143 kg s−1), cooling water (49–69 °C, 343–1776 L s−1), and condensate (48–68 °C, 126–140 kg s−1). In 1998 the operation of the power plant, which had produced only warm water (100 MWth) since 1990, was transformed to a cogeneration facility producing both warm water and electricity (60 MWe). In 2001, electrical production was increased to 90 MWe, and in 2005 the Nesjavellir power plant produced 120 MWe and around 280 MWth (Zarandi and Ivarsson 2010). As a consequence of increased discharge of warm effluents from the Nesjavellir power plant, water temperatures at several groundwater outlet sites along a 2 km stretch of the southwest shore of Þingvallavatn have increased significantly above ambient water temperature. In 2003, the water temperature at 3 outlet sites in the lake had been elevated on average by 10–18 °C above ambient water temperature (Wetang’ula 2004, Wetang’ula and Snorrason 2005, Zarandi and Ivarsson 2010).
Methods
Sampling sites
Benthic invertebrates were sampled in Þingvallavatn in late August 2003 at 0.2, 0.4, 1.0, 2.0, and 4.0 m along depth transects at 2 heat-affected sites, Varmagjá and Eldvík, and 2 unaffected reference sites, Miðfell and Vatnskot (Fig. 1). Varmagjá and Eldvík were selected as representatives for heat-affected sites because of good physico-chemical and biological background data compared to other heat-affected sites. In addition, inverte-brate community information was available from Varmagjá before and after exploitation of the Nesjavellir geothermal field (Snorrason 1982, Lindegaard 1992, Ólafsson 1992, Snorrason and Jónsson 2000, Wetang’ula
Fig 1. Lake Þingvallavatn (64°10´N, 21°10´W) and the 2 heat- affected sites, Varmagjá and Eldvík, and the 2 reference sites, Miðfell and Vatnskot, marked in black dots. The thick arrow indicates direction of flow of warm effluents from the Nesjavellir geothermal power plant.

DOI: 10.5268/IW-1.3.363
149Effects of geothermal effluents on macrobenthic communities
Inland Waters (2011) 1, pp. 146-157
2004). The 2 cold reference sites were selected because they were quite distant from the geothermaly affected outlets and because ecological data were available over the same time period as for Varmagjá (e.g., Snorrason 1982, Lindegaard 1992, Ólafsson 1992, Snorrason and Jónsson 2000).
VarmagjáThe site is characterized by a small (10 × 35 m), shallow (≤1 m) rift pool connected to Þingvallavatn via a 2 m wide and 0.1–0.6 m deep opening, making it somewhat isolated from the lake and shielded from wave exposure. Thus, the temperature effects are sustained inside the pool irrespec-tive of wave action that quickly mixes warm and cold water at exposed shores. At Varmagjá, samples were collected at 0.1 m depth within the rift pool at the main outflow point of geothermal effluents and at 5 depths along a transect outside the rift pool. Prior to exploitation of the Nesavellir field, a natural geothermal signature with temperatures of 7.5–10.7 °C had been observed for some time within the rift pool (Ólafsson 1992, Aðalsteinsson et al. 1992, Snorrason and Jónsson 2000). As the power plant was transformed into a cogeneration facility, the discharge of excess cooling water led to rapid increase in water temperature at Varmagjá. For the period 1999–2002, the average temperature within the rift pool had risen to 19.2 °C, and a further rise to an average of 28.6 °C was seen for the period 2003–2006 (Zarandi and Ivarsson 2010). Temperatures outside the rift pool followed a different and more dynamic pattern. Measurements in calm weather along vertical and horizontal transects in the summer 2003 and winter 2004 showed that the summer temperature may reach ~26 °C close to the surface just outside the mouth of the rift pool but cools down rapidly outside the mouth. At 10 m distance from the mouth, tem-peratures within the topmost 0.3 m surface water had decreased to 14–25 °C, at 25 m to 14–16.5 °C, and at 50 m to 13–14 °C (Wetang’ula 2004). At 75 m distance from the mouth of the rift pool the temperature above 1.5 m was close to 13 °C (Wetang’ula 2004), similar to the natural maximum summer temperature in the lake (Aðal-steinsson et al. 1992, Malmquist et al. 2010a, 2010b). Thus, thermal pollution during summer outside the rift pool at Varmagjá seems to be confined more or less to a 30–50 m long, ~30 m wide, and 20–30 cm thin layer of warm water floating on top of the cooler, denser lake water. Water within the rift pool never freezes.
EldvíkThe site is 0.3 km east of Varmagjá. In terms of discharge, this site resembles Varmagjá, but they differ in topography. There is no rift pool at Eldvík, so the oulets are exposed to wave actions. Before operation of the Nesjavellir power
plant there were no records of geothermal influences at the Eldvík site. As the operation of the power plant has been extended, water temperatures have risen markedly at 2–3 distinct outflow springs on the lake shoreline (Wetang’ula 2004, Zarandi and Ivarsson 2010). During 1999–2002 average temperatures at the outflow sites almost doubled as compared to the period before 1990 (17.2 vs. 9.8 °C). For the period 2003–2006, temperatures rose to 19.7 °C on average (Zarandi and Ivarsson 2010). Temperature profiles in summer 2003 at Eldvík along vertical and horizontal transects resemble those at Varmagjá, although the size of the area affected by thermal pollution at Eldvík appears somewhat smaller (Wetang’ula 2004). Only sporadic temperature measurements are available in winter at Eldvík. In winter 2004, temperature at one outflow site at the lake shoreline measured 27.3 °C (Wetang’ula 2004), and in February 2009 temperature at another outflow site measured 15.8 °C (Zarandi and Ivarsson 2010). As at Varmagjá, an ice-free area of 500–1000 m2 extending from the outflow sites has been present at Eldvík for the past 10–15 years.
MiðfellThis cold reference site is located ~6.5 km east of Varmagjá and well outside the impact area of the Nesjavellir power plant. Maximum summer temperatures rarely exceed 13 °C, and for most of the year temperatures range between 1 and 6 °C. There are no known inlets of spring-fed water at the site, and the site is usually frozen between January and March.
Vatnskot This cold reference site is located on the north shore of the lake within the Þingvellir National Park and the buffer zone of UNESCO World Cultural and Natural Heritage List, some 12 km northeast and upstream from the impact area of the Nesjavellir power plant. The site is close to the main inlets of spring-fed water into the lake, where cold groundwater (2.8–3.5 °C) seeps from the north through porous lava fields and enters in the littoral zone as small inlets or through rifts. Maximum summer temperatures at the site are usually close to 12–13 °C in July–August, but for most of the year temperatures are within the range of 1–6 °C. The site is usually ice-free in winter because of the stable inflow of spring-fed water of constant temperature.
Samples and data collection
At all 4 sampling sites, 6 lava stones 4–20 cm in diameter and 5–12 cm in height were collected randomly at each sampling depth by a scuba diver. The stones were carefully removed from the bottom and put in a bucket of

150
DOI: 10.5268/IW-1.3.363
Sigurður Snorrason et al.
© International Society of Limnology 2011
water, closed with a lid, and lifted to the surface. Each stone was thoroughly brushed clean in prefiltered water, the content sieved through a 250 μm sieve, and the retained invertebrates preserved in 3–4% formalin. Overhead projections of the stones were drawn on paper and the area was measured for subsequent calculations of invertebrate density per unit area. A single measurement of water temperature was performed at each sampling depth with a mercury thermometer (0.1 °C accuracy). The stony lava substrate on the bottom at all sampling sites was similar in size, shape, and surface texture, as was the bottom slope, ranging between 5–15°. All sampling sites except the rift pool at Varmagjá were similarly exposed to the open lake and were assumed to be affected to a similar degree by wave exposure, albeit from different directions. Due to small lava reefs, the shallowest sampling depths at Eldvík and Vatnskot study sites may be slightly less exposed than at Miðfell.
To explore thermal effects on life history traits at the species level we used the pulmonate gastropod Radix peregra, which is known to be adapted to a variety of thermal regimes in Icelandic freshwaters, including warm ponds (Tuxen 1944, Þórðarson 1983), indicating that it is an eurythermic species. Radix peregra is among the most abundant macroinvertebrates in the lake (Snorrason 1982, 2000, Lindegaard 1992) and an important food for 2 of the 4 morphs of Arctic charr (Salvelinus alpinus) occupying the lake (Malmquist et al. 1992). In Þingvallavatn, the snails reach sexual maturity in 2 years. They mate and reproduce in late May–early June, after which the adults die; the eggs hatch in late June–early August (Snorrason 1982, 2000, Lindegaard 1992). This means that at all times of the year the size distribution of the snails exhibits 2 peaks representing the juvenile and subadult cohorts. To test for changes in the cohort size structure, the length of R. peregra shells from the apex to the distal rim of the shell aperture was measured under a stereo microscope to the nearest 0.5 mm. For comparisons of size distributions of R. peregra before and after wastewater discharge from the Nesjavellir power plant, we used data from the 0.4 m and 4.0 m depths at Varmagjá, Miðfell, and Vatnskot obtained in August–September 1977 (Snorrason 1982).
To examine effects of elevated water temperature at the community level, we assessed the community structure of chironomid larvae, the most abundant group of littoral macroinvertebrates in Þingvallavatn (Lindegaard 1992) as well as generally in Icelandic lakes (Malmquist et al. 2000). Chironomid larvae are also an imporant food for Arctic charr and threespine stickleback (Gasterosteus aquleatus) in the lake (Malmquist et al. 1992, Sandlund et al. 1992).
Chironomidae larvae were identified to species, if possible. Abundance (density) of chironomid larvae and
the gastropod R. peregra is presented as average number of individuals per square meter. The averages are based on random selection of 6 of 10 stones from each depth.
The data presented here are total densities of macroin-vertebrates, chironomids, and R. peregra and total number of taxa. For statistical analyses, student t-test or Mann-Whitney rank test were used for 2 sample comparisons and one-way ANOVA or Kruskal-Wallis for multiple comparisons. Nonparametric tests were applied when data did not meet the assumptions of normality. Student-New-man-Keuls (SNK) and Dunn’s post hoc test were used to isolate differences.
Results
Temperature
Water temperature at the heat-affected and reference sites differed down to ~1.0 m, with the most profound differences at 0.2 m and 0.4 m, respectively (Fig. 2). At the Varmagjá heat-affected site, the water temperature was 27.3 °C within the rift pool and 23.2–12.0 °C outside the rift pool, cooling rapidly below 0.4 m. At the Eldvík heat-affected site, the water temperature was 19.3–12.2 °C along the transect, with a sharp decrease below 0.4 m. The temperature ranged between 13.5–8.5 °C at the Miðfell reference site and between 12.7–12.1 °C at the Vatnskot reference site. For all depths pooled, excluding the site within the rift pool at Varmagjá, the water temperature was neither significantly different between the 2 heat-affected sites (Mann-Whitney: T = 0.255, df = 8, p = 0.805) nor between the 2 reference sites (T = 0.544, df = 8, p = 0.601), but significantly higher at the heat-affected sites than the reference sites (T = 2.724, df = 18, p = 0.014).
Invertebrates
Within each of the 4 study sites, the total densities of mac-roinvertebrates differed significantly among sampling depths (Kruskal-Wallis results for Varmagjá (0.1 m excluded): H = 16.92, df = 4, p = 0.002; Eldvík: H = 23.12, df = 4, p < 0.001; Miðfell: H = 23.38, df = 4, p < 0.001; and Vatnskot: H = 16.03, df = 4, p = 0.003). In general, densities of invertebrates were higher at deeper parts at each study site (Fig. 3). At each sampling depth, total densities of macroinvertebrates differed significantly among the 4 study sites (Kruskal Wallis results at 0.2 m: H = 17.59, p < 0.001; 0.4 m: H = 15.93, p = 0.001; 1.0 m: H = 12.64, p = 0.005; 2.0 m: H = 28.38, p < 0.001; and 4.0 m: H = 13.07, p = 0.004). The total densities of mac-roinvertebrates were in all cases lowest at the Miðfell reference site. For the 0.2 m sampling depth, total mac-

DOI: 10.5268/IW-1.3.363
151Effects of geothermal effluents on macrobenthic communities
Inland Waters (2011) 1, pp. 146-157
roinvertebrate densities were significantly higher at both the heat-affected sites than the 2 reference sites (SNK post hoc test; p < 0.05). The difference was not significant between the 2 heat-affected sites, however (p > 0.05). At a depth of 0.4 m the total macroinvertebrate densities were still highest at the 2 heat-affected sites, with significantly higher density in Varmagjá than Eldvík (SNK post hoc test: p < 0.05). At depths between 1.0 and 4.0 m there was no evident pattern in macroinvertebrate densities in comparisons of heat-affected and reference sites (Fig. 3). For all 4 study sites, except the Vatnskot reference site, the total macroinvertebrate taxa richness was highest at the greatest sampling depth and tended to increase with increasing depth (Fig. 4), but the association was only significant at the Varmagjá heat-affected site (Kruskal-Wallis: H = 21.08, df = 5, p < 0.001) and the Miðfell reference site (Kruskal-Wallis: H = 14.35, df = 4, p = 0.006). Within each sampling depth, taxa richness only differed significantly among the 4 study sites at the 2.0 m depth (ANOVA; F = 3.73, df = 3, p = 0.028). There, the highest richness was at the Vatnskot reference site and lowest in the Eldvík heat-affected site (Fig. 4).
Densities of R. peregra at the 2 heat-affected sites in Varmagjá and Eldvík were significantly higher at 0.4 (Mann-Whitney; p < 0.001) and 1.0 m sampling depths (Mann-Whitney; p < 0.001) as compared to the 2 reference sites at Miðfell and Vatnskot. At Varmagjá, the density of R. peregra was significantly different among sampling depths (Kruskal-Wallis: H = 29.65, df = 5, p < 0.001), and in all pair-wise comparisons the density was significantly lower inside the rift pool than outside for all sampling depths (SNK post hoc test; p < 0.05; Fig. 5).
Comparisons of size distributions of R. peregra in 1977 (before development of geothermal utilization) from the 2 reference sites and from the transect outside the Varmagjá heat-affected site clearly show the presence of 2 cohorts expected at this time of the year (i.e., the newly hatched snails and 1-year-old subadults; Snorrason 1982, Lindegaard 1992). This pattern is clear both in the surf zone (0.4 m) and deeper along the sampling transect (4.0 m; Fig. 6), the exception being the shallower sampling depths (represented by 0.4 m in Fig. 6) closest to the outflow of the rift pool. Here the bimodality is not clear. A natural geothermal influence in Varmagjá, with water of 7.5–10.7 °C, might influence growth and maturity age in snails near the mouth of the Varmagjá rift pool and cause local irregularities in size distributions (i.e., making cohort size distributions more variable and distinctions between cohorts less clear; Snorrason 1982).
The data from 2003 show a similar pattern in terms of cohort structure (Fig. 6). At the Varmagjá heat-affected site the size distribution is unimodal and spread at the near-shore depths (represented by 0.4 m), but at 4.0 m the
data show presence of larger snails, presumably 1-year-old subadults. The size distributions at the shallow sampling depths at the reference site Vatnskot are considerably displaced compared to 1977, which was unexpected considering that the 1977 sample was taken 4 weeks later in the year than the 2003 samples.
Fig. 2. Water temperature at the 4 study sites in calm weather 22–29 Aug 2003.
Fig. 3. Densities (mean no. ind./m2, ± s.e.) of macroinvertebrates at the 2 heat-affected sites, Varmagjá and Eldvík, and the 2 reference sites, Miðfell and Vatnskot. The rift pool at Varmagjá is labelled at 0.1 m.
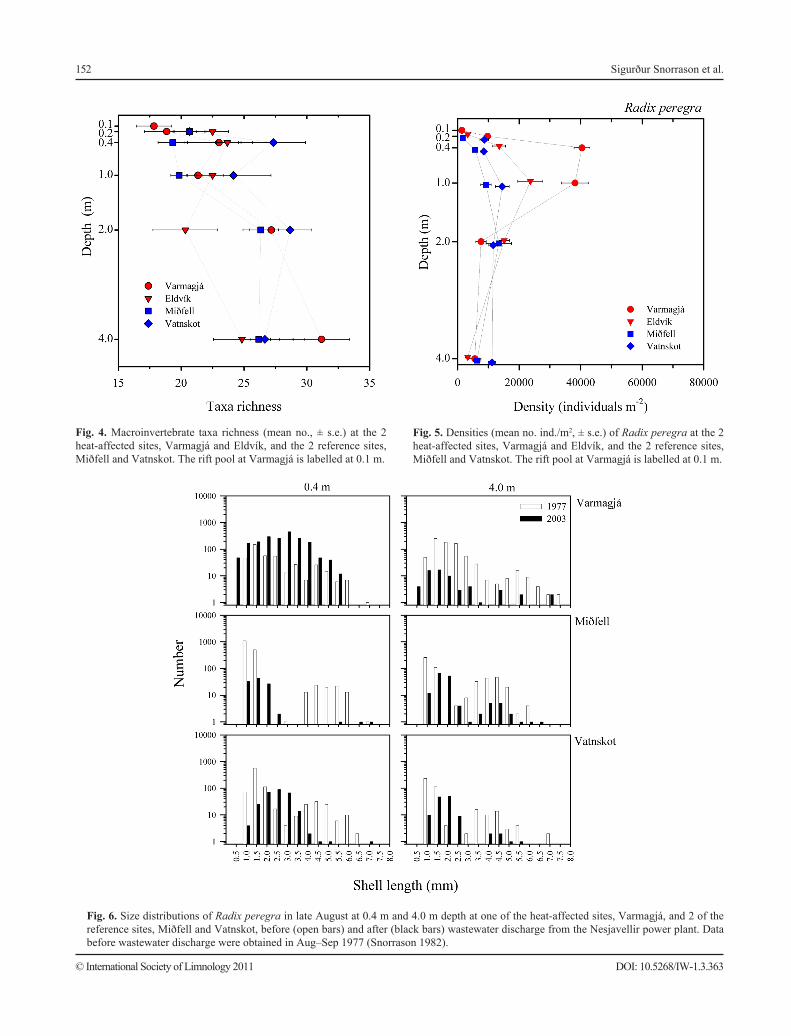
152
DOI: 10.5268/IW-1.3.363
Sigurður Snorrason et al.
© International Society of Limnology 2011
Fig. 4. Macroinvertebrate taxa richness (mean no., ± s.e.) at the 2 heat-affected sites, Varmagjá and Eldvík, and the 2 reference sites, Miðfell and Vatnskot. The rift pool at Varmagjá is labelled at 0.1 m.
Fig. 5. Densities (mean no. ind./m2, ± s.e.) of Radix peregra at the 2 heat-affected sites, Varmagjá and Eldvík, and the 2 reference sites, Miðfell and Vatnskot. The rift pool at Varmagjá is labelled at 0.1 m.
Fig. 6. Size distributions of Radix peregra in late August at 0.4 m and 4.0 m depth at one of the heat-affected sites, Varmagjá, and 2 of the reference sites, Miðfell and Vatnskot, before (open bars) and after (black bars) wastewater discharge from the Nesjavellir power plant. Data before wastewater discharge were obtained in Aug–Sep 1977 (Snorrason 1982).
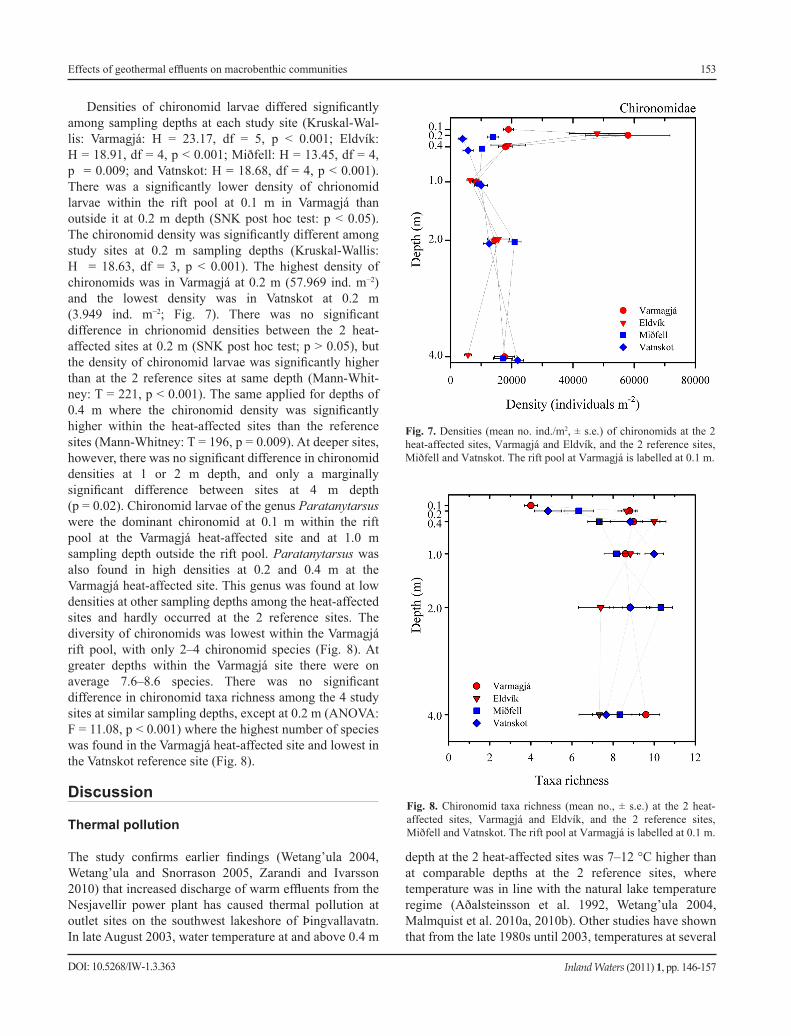
DOI: 10.5268/IW-1.3.363
153Effects of geothermal effluents on macrobenthic communities
Inland Waters (2011) 1, pp. 146-157
depth at the 2 heat-affected sites was 7–12 °C higher than at comparable depths at the 2 reference sites, where temperature was in line with the natural lake temperature regime (Aðalsteinsson et al. 1992, Wetang’ula 2004, Malmquist et al. 2010a, 2010b). Other studies have shown that from the late 1980s until 2003, temperatures at several
Densities of chironomid larvae differed significantly among sampling depths at each study site (Kruskal-Wal-lis: Varmagjá: H = 23.17, df = 5, p < 0.001; Eldvík: H = 18.91, df = 4, p < 0.001; Miðfell: H = 13.45, df = 4, p = 0.009; and Vatnskot: H = 18.68, df = 4, p < 0.001). There was a significantly lower density of chrionomid larvae within the rift pool at 0.1 m in Varmagjá than outside it at 0.2 m depth (SNK post hoc test: p < 0.05). The chironomid density was significantly different among study sites at 0.2 m sampling depths (Kruskal-Wallis: H = 18.63, df = 3, p < 0.001). The highest density of chironomids was in Varmagjá at 0.2 m (57.969 ind. m−2) and the lowest density was in Vatnskot at 0.2 m (3.949 ind. m−2; Fig. 7). There was no significant difference in chrionomid densities between the 2 heat- affected sites at 0.2 m (SNK post hoc test; p > 0.05), but the density of chironomid larvae was significantly higher than at the 2 reference sites at same depth (Mann-Whit-ney: T = 221, p < 0.001). The same applied for depths of 0.4 m where the chironomid density was significantly higher within the heat-affected sites than the reference sites (Mann-Whitney: T = 196, p = 0.009). At deeper sites, however, there was no significant difference in chironomid densities at 1 or 2 m depth, and only a marginally significant difference between sites at 4 m depth (p = 0.02). Chironomid larvae of the genus Paratanytarsus were the dominant chironomid at 0.1 m within the rift pool at the Varmagjá heat-affected site and at 1.0 m sampling depth outside the rift pool. Paratanytarsus was also found in high densities at 0.2 and 0.4 m at the Varmagjá heat-affected site. This genus was found at low densities at other sampling depths among the heat-affected sites and hardly occurred at the 2 reference sites. The diversity of chironomids was lowest within the Varmagjá rift pool, with only 2–4 chironomid species (Fig. 8). At greater depths within the Varmagjá site there were on average 7.6–8.6 species. There was no significant difference in chironomid taxa richness among the 4 study sites at similar sampling depths, except at 0.2 m (ANOVA: F = 11.08, p < 0.001) where the highest number of species was found in the Varmagjá heat-affected site and lowest in the Vatnskot reference site (Fig. 8).
Discussion
Thermal pollution
The study confirms earlier findings (Wetang’ula 2004, Wetang’ula and Snorrason 2005, Zarandi and Ivarsson 2010) that increased discharge of warm effluents from the Nesjavellir power plant has caused thermal pollution at outlet sites on the southwest lakeshore of Þingvallavatn. In late August 2003, water temperature at and above 0.4 m
Fig. 7. Densities (mean no. ind./m2, ± s.e.) of chironomids at the 2 heat-affected sites, Varmagjá and Eldvík, and the 2 reference sites, Miðfell and Vatnskot. The rift pool at Varmagjá is labelled at 0.1 m.
Fig. 8. Chironomid taxa richness (mean no., ± s.e.) at the 2 heat- affected sites, Varmagjá and Eldvík, and the 2 reference sites, Miðfell and Vatnskot. The rift pool at Varmagjá is labelled at 0.1 m.

154
DOI: 10.5268/IW-1.3.363
Sigurður Snorrason et al.
© International Society of Limnology 2011
outlet sites on the southwest lakeshore of Þingvallavatn increased on average by 10–18 °C above ambient water temperature (Wetang’ula 2004, Wetang’ula and Snorrason 2005, Zarandi and Ivarsson 2010). Before exploitation of the Nesjavellir geothermal field in the late 1980s, a relatively weak, natural geothermal influence was known in Þingvallavatn at only one site, the rift pool at Varmagjá, with warm groundwater (7.5–10.7 °C) entering the rift pool at one distinct outlet (Aðalsteinsson et al. 1992, Ólafsson 1992).
Despite the considerable increase in temperature at lakeshore outlet sites, the thermal pollution was confined spatially on a vertical scale. At both heat-affected sites, thermal effects were more or less restricted to the littoral surf zone at and above 0.4 m depth, with no detectable differences in temperature between the heat-affected and the reference sites at and below 2.0 m. Furthermore, as observed on a calm day in summer 2003 and winter 2004 (Wetang’ula 2004), the thermal pollution at Eldvík and outside the rift pool at Varmagjá was confined to a thin layer (20–30 cm) of warm water, at most 30 m wide and extending 30–50 m out from the outlet sites, floating on top of the cooler, denser lake water. A likely significant factor accounting for the vertical confinement of thermal pollution in Þingvallavatn is a combination of the voluminous, cold nature of the lake (Aðalsteinsson et al. 1992, Ólafsson 1992, Malmquist et al. 2010a, 2010b) along with wind-induced wave actions and thorough mixing of the water as a result of frequent lows and windy climate in the area (Einarsson 1992).
In contrast to limited vertical thermal pollution, thermal effects on a horizontal scale were prominently present in 2003 at several outlet sites along a 2 km stretch on the southwest lakeshore of Þingvallavatn (Wetang’ula 2004, Wetang’ula and Snorrason 2005, Zarandi and Ivarsson 2010). From 2003, temperatures at outlet sites along the southwest lakeshore continued to increase and seemed to peak in 2006, as observed in Varmagjá with a maximum temperature of 31.3 °C (average temperature of 28.6 °C for the period 2003–2006; Zarandi and Ivarsson 2010). Since 2009, indications are that temperatures at outlet sites in Eldvík and Varmagjá have decreased by 2–4 °C (Zarandi and Ivarsson 2010), possibly a result of measurements taken in 2004 at the Nesjavellir power plant, when a cooling tower was built and reinjection of warm wastewater into deep wells reaching below groundwater level began. In 2009–2010, reinjection wells accommodated most, some 130 l s−1, of the separator water and 30 l s−1 of condensate water; however, excess separator water still flows into Nesjavallalækur brook, and about 85% of the condensate is disposed of into shallow wells just north of the power station (Zarandi and Ivarsson 2010).
Chemical pollution at the heat-affected sites does not seem to have affected the biota to any measureable degree. Some increase in concentrations of several chemical components of known ecotoxicological potential, especially in As, Al, B, and SO4, has been recorded, but in most cases the concentrations have been within international, environmental quality guidelines for the protection of aquatic life (Wetang‘ula and Snorrason 2005, Zarandi and Ivarsson 2010). Concordant with this, studies on trace elements in biological samples, including analyses of Al, As, and Hg in sediment, plants, inverte-brates, and fish, show no evidence of effects that can be related to the geothermal wastewater (Snorrason and Jónsson 2000, Sturlaugsson et al. 2009, Auðunsson et al. 2010, Zarandi and Ivarsson 2010).
Thermal effects on macroinvertebrates
The low density of R. peregra inside the rift pool at Varmagjá, where water temperature was high, 27–28 °C, indicates that temperatures ≥27 °C may be detrimental for R. peregra. This fits well with a number of studies on R. peregra showing that it’s upper limit of optimum temperature is around 30 °C (Islam et al. 2001, Morley et al. 2007, Ólafsson et al. 2010, Woodward et al. 2010). Also, our findings that densities of R. peregra outside the rift pool at Varmagjá and at Eldvík, where temperatures were elevated down to 0.4–1 m were higher than at the 2 cold control sites at comparable depths, indicate that R. peregra may benefit from moderate warming effects (e.g., through higher fecundity and or survival rate of immature snails).
The bimodal size distributions of R. peregra from the 2 control sites with data from 1977 at the same sites are typical of a 2 year, semelparous life cycle of the snail in the lake (Snorrason 1982, 2000, Lindegaard 1992). In contrast, the apparent unimodal size distribution at Varmagjá could illustrate a plastic life history response due to heat effect, whereby the snails manage to reproduce after 1 year instead of 2. The higher variation in snail size outside the rift pool at Varmagjá presumably reflects the high degree of temperature fluctuations generated by wind mixing effects. Note that these effects are also highly variable on a small scale, depending on frequency and direction of winds and distance from effluent outlets.
Our data on the chironomids clearly indicate that sustained, high temperatures have transformed a relatively diverse chironomid community to a primarily 2-species community dominated by Paratanytarsus sp. and Cricotopus sylvestris. The occurrence of C. sylvestris at the heat-affected sites is not unexpected. The presence of C. sylvestris in thermal springs in Iceland with tempera-tures around 40 °C has long been known (Tuxen 1944,

DOI: 10.5268/IW-1.3.363
155Effects of geothermal effluents on macrobenthic communities
Inland Waters (2011) 1, pp. 146-157
Pinder 1997), and recent studies in Iceland, including streams in the area of the Hengill central volcano, demonstrate that C. sylvestris thrives well under elvated temperature regimes (Ólafsson et al. 2010, Woodward et al. 2010). Paratanytarsus sp. has not been previously recorded from Þingvallavatn, and only twice before in Iceland where it was found in warm effluents from the Krafla geothermal power plant and in the nearby Víti crater lake (Kristjánsdóttir 2004, Ólafsson et al. 2010). Its presence in a habitat of elevated temperature is, however, congruent with habitat preferences characteristic for many species belonging to the Chironominae subfamily, considered in general to be warm-adapted (Oliver 1971). In effluents from geothermal areas in New Zealand, the chironomid species Cladopelma curtivalva (misidentified as Chironomus cylindricus, ser: Cranston and Martin 2007) was found in 39-41 °C water where the larvae were associated in a dense algae mat on the stream bottom (Winterbourn 1968, 1969). Other species of Chironomus and Tanytarsus have been demonstrated to be very heat tolerant, including survival in thermal water with tempera-tures of 40–48 °C (Lamberti and Resh 1983, Pinder 1997).
During on-land winds, heat-affected water mixes with cooler lake water. This causes temperature fluctuations, especially in the littoral zone (<2 m) ranging from 23 °C in calm weather but dropping to ambient lake temperature, 10–11 °C in August, during on-land winds (Aðalsteinsson et al. 1992, Wetang’ula 2004). The chironomid community of the littoral zone does not seem to be much affected by fluctuating temperatures. Paratanytarsus sp. was almost exclusively restricted to the 2 heat-affected sites, particu-larly Varmagjá where it was the dominant chironomid species. In contrast, E. minor, a wave-tolerant chironomid species otherwise quite abundant in the <1.0 m littoral zone of the lake (Lindegaard 1992), was less prominent at the heat-affected sites than at the Miðfell reference site with similar wind exposure to the 2 heat-affected sites. The low represenation of E. minor and other Orthocladii-nae species, including Corynoneura cf. scutella, at the heat-affected sites but otherwise abundance at other sites supports the notion that these species are thermal genaralists and or cool-water affiliates (Oliver 1971, Olsen et al. 2003, Woodward et al. 2010).
The thermal effects observed in Þingvallavatn are in line with those observed in some geothermally affected streams in Iceland, where temperature was the dominating factor explaining community structure of the stream benthos (Friberg et al. 2009, Ólafsson et al. 2010, Woodward et al. 2010). Our results are also in agreement with a study by Lamberti and Resh (1983) on geothermal effects on stream benthos, where thermal and chemical effects were experimentally separated. As in the present study, Lamberti and Resh (1983) found that the thermal
component had greater influence than the chemical component in determining community features of the zoobenthos. The study was carried out in a third-order stream at the Geysers, California, USA, where temperature was naturally elevated by ~8 °C from the average midday water temperature of 19 °C to 27 °C, and chemicals naturally elevated by as much as 10 times for B (Lamberti and Resh 1983). Under these circumstances, densities of macroinvertebrates doubled; however, by heating of non-geothermal water by 8 °C (19 °C to 27 °C), macroinverte-brate densities decreased by ~25%, whereas cooling of geothermal water by 14.5 °C (36.5 °C to 22 °C) increased macroinvertebrate densities by 70 times. The study by Lamberti and Resh (1983) and the present one, along with information on chemicals (Snorrason and Jónsson 2000, Wetang’ula 2004, Zarandi and Ivarsson 2010), indicate that a number of macroinvertebrate species are unaffected by moderate elevation of potentially ecotoxicological elements but benefit from moderate elevation of temperature (i.e., an elevation of 8–12 °C above ambient temperature).
In addition, many macroinvertebrate species in both studies (all 44 species indentified by Lamberti and Resh [1983] and 48 species and other taxonomical identies in the present study) seem to have a growth optimum somewhere below 25–30 °C, indicating an absolute upper level of temperature (i.e., 25–30 °C) for growth optimum by many of the macroinvertebrates (see also previous discussion on R. peregra). Such an absolute temperature level for growth optimum common to many organisms may be expected, assuming that the temperature is affecting a vital physiological function—an evolutionary, inert trait basic for life. In support of this, only 3 species of chironomids have been recorded world-wide from thermal springs with temperature ≥40 °C, and apparently no insect in an active state tolerates temperatures >50 °C (Tuxen 1944, Pinder 1997). According to Lamb (1992), the upper limit (lethal temperature) for most insects falls between 25 and 40 °C, and the developmental rate will decrease as this upper limit is approached (Olsen et al. 2003).
Our results generally agree with predictions and obser-vations of the influences of warming on invertebrates, cli-matically induced or otherwise (Lamberti and Resh 1983, Hughes 2000, Walther et al. 2002, Woodward et al. 2010). Moderate warming resulted in increased density of mac-roinvertebrates for the gastropod snail R. peregra and some of the chironomid species, particularly C. sylvestris and Orthocladius oblidens, but species richness remained little or unaffected. At extreme temperatures (>25–30 °C), both density and species richness declined; however, to further separate thermal effects from biological, species-specific attributes, studies are needed that include

156
DOI: 10.5268/IW-1.3.363
Sigurður Snorrason et al.
© International Society of Limnology 2011
time-series data on growth, survival, and reproduction of R. peregra and life history studies on the littoral chironomid assemblages. Moreover, because exploitation of the Nesjavellir gothermal field has increased since the present study was carried out, with accompanying increase in warm effluents along the lakeshore of Þingvallavatn (Zarandi and Ivarsson 2010), monitoring studies on the biota are also urgently needed on a yearly basis. Such monitoring studies are urgent in a broader sense in Iceland considering the increased demand for electricty from the energy-intensive industry, especially the expansion of the country’s aluminum industry, over the past 5 years (Björnsson 2010).
Acknowledgements
We thank Anna K. Gunnarsdóttir, Erlín E. Jóhannsdóttir, Finnur Ingimarsson, and Haraldur R. Ingvason for assistance in the field. The study was carried out with a grant from Reykjavik Energy (Orkuveita Reykjavíkur).
References
Aðalsteinsson H, Jónasson PM, Rist S. 1992. Physical characteristics of Thingvallavatn, Iceland. Oikos. 64:121-135.
Aksoy N, Simsek C, Gunduz O. 2009. Groundwater contamination mechanisms in a geothermal field: A case study of Balcova, Turkey. J Contam Hydrol. 103:13-28.
Arnórsson S. 2004. Envirnonmental impact of geothermal energy utilization. In: Giere R, Stille P, editors. Energy, waste and the environment: a geochemical perspective. London (UK): Geological Society. Special Publications. p. 297-336.
Auðunsson GA, Ólafsson J, Guðbergsson G, Malmquist HJ. 2010. Mercury in brown trout (Salmno trutta) in Icelandic lakes. Poster presented at Reykjavík Energy conference. 14 May 2010. Icelandic.
Axelsson G, Stefánsson V, Björnsson G, Liu J. 2005. Sustainable utilization of geothermal resources for 100–300 years. Proceedings of the World Geothermal Congress 2005. April 2005. Antalya, Turkey. 8 p.
Björnsson S, editor. 2010. Geothermal development and research in Iceland. National Energy Authority (Orkustofnun). 39 p.
Cranston PS, Martin J. 2007 Family Chironomidae. In: Evenhuis NL, editor. Catalog of the Diptera of the Australasian and Oceanian Regions. [cited 3 Jun 2011]. Available from http://hbs.bishopmu-seum.org/aocat/hybotidae.html.
Daufresne M, Roger MC, Capra H, Lamouroux N. 2003. Long-term changes within the invertebrate and fish communities of the Upper Rhône River: effects of climatic factors. Global Change Biol. 10:124-140.
Einarsson MÁ. 1992. Climatic conditions of the Thingvallavatn area. Oikos. 64:96-104.
Friberg N, Dybkjær JB, Olafsson JS, Gislason GM, Larsen SE, Lauridsen TL. 2009. Relationships between structure and function in
streams contrasting in temperature. Freshwater Biol. 4:2051-2068. Gunnlaugsson E, Frimannson H, Sverrisson GA. 2000. District heating
in Reykjavik – 70 years experience. Proceedings World Geothermal Congress 2000, 28 May–10 Jun 2000. Kyushu-Tohoku, Japan. p. 2087-2092.
Hughes L. 2000. Biological consequences of global warming. Is the signal already apparent? Trends Ecol. Evol. 15:56-61.
[IEA] . International Energy Agency. 2003. Energy to 2050: Scenarios for a Sustainable Future. Paris (France).
[IEA] International Energy Agency. 2009. World Energy Outlook, 2009. Paris (France).
Islam, MN, Gordon, RP, McLachlan, AJ. 2001. The biology of Lymnaea peraegra (Muller) (Gastropoda: Pulmonata: Basom-matophora) with special reference to the effects of herbicides on its reproduction. OnLine J Biol Sci. 1:532-540.
Jeppesen E, Meerhoff M, Holmgren K, González-Bergonzoni I, Teixeira-de Mello F, Declerck SAJ, Meester LD, Søndergaard M, Lauridsen TL, Bjerring R, et al. 2010. Impacts of climate warming on lake fish community structure and potential effects on ecosystem function. Hydrobiologia 646:73-90.
Jónasson PM. 1992. Iceland – an island astride the Mid-Atlantic Ridge. Oikos. 64:9-13.
Jónasson PM. 1993. Presidential Address. Continental rifting – energy pathways of two contrasting rift lakes. Verh Internat Verein Limnol. 25:1-14.
Jónasson PM, Hersteinsson P, editors. 2011. Thingvallavatn – A unique world evolving. Opna, Reykjavik (Iceland). 326 p.
Kristjánsdóttir B. 2004. Lífið í Víti; botnlífverur. [Benthic invertebrates of Víti crater lake]. [BSc dissertation]. [city of university (Iceland)]: University of Iceland. Icelandic.
Kristmannsdóttir H, Ármannsson H 2003. Environmental aspects of geothermal energy utilization. Geothermics 32:451–461.
Lamb RJ. 1992. Developmental rate of Acythosiphon pisum (Homoptera: Ephididae) at low temperatures: implications for estimating rate parameters for insects. Environ Entomol. 21:10-19.
Lamberti GA, Resch VH. 1983. Geothermal effects on stream benthos: Seperate influences of thermal and chemical components on periphyton and macroinvertebrates. Can J Fish Aquat Sci. 40:1995-2009.
Langford TE. 1990. Ecological effects of thermal discharges. London (UK): Elsevier. 468 p.
Lindegaard C. 1992. Zoobenthos ecology of Thingvallatn: vertical dis-tribution, abundance, population dynamics and production. Oikos. 64:257-304.
Malmquist HJ, Antonsson Þ, Guðbergsson G, Skúlason S, Snorrason SS. 2000. Biodiversity of macroinvertebrates on rocky substrate in the surf zone of Icelandic lakes. Verh Internat Verein Limnol. 27:121-127.
Malmquist HJ, Antonsson Þ, Ingvason HR, Ingimarsson F, Árnason F. 2009. Salmonid fish and warming of shallow Lake Elliðavatn in Southwest Iceland. Verh Internat Verein Limnol. 30:1127-1132.
Malmquist HJ, Ingimarsson F, Ingvason HR, Stefánsson SM. 2010a. Climate change and its effects on lakes in SW-Iceland. In: Andradottir

DOI: 10.5268/IW-1.3.363
157Effects of geothermal effluents on macrobenthic communities
Inland Waters (2011) 1, pp. 146-157
HO, editor. Proceedings of the 14th international workshop on physical processes in natural waters. 28 Jun–1 Jul 2010. Reykjavík (Iceland). p. 34-35.
Malmquist HJ, Ingimarsson F, Ingvason HR, Stefánsson SM. 2010b. Lake Þingvallavatn monitoring project. Report for 2009. Natural History Museum of Kópavogur. Report No. 2-09. 35 p. Icelandic with English summary.
Malmquist HJ, Snorrason SS, Skúlason S, Sandlund OT, Jonsson B, Jónasson PM. 1992. Diet differentiation in polymorphic Arctic charr in Thingvallavatn, Iceland. J Anim Ecol. 61:21-35.
Morley NJ, Adam ME, Lewis JW. 2007. The effects of temperature on the distribution and establishment of Echinoparyphium recuvratum metacercariae in Lymnaea peregra. J Helminthol. 82:311-315.
Nebeker AV. 1971. Effect of high winter water temperature on adult emergence of aquatic insects. Water Res. 5:777-783.
Oliver DR. 1971. Life history of the Chironomidae. Annual review of e-tomology 16:211–230.
Olsen A, Bale JS, Leadbeater BSC, Callow ME, Holden JB. 2003. De-velopmental thresholds and day-degree requirements of Paratanytar-sus grimmii and Corynoneura scutellata (Diptera: Chironomidae): two midges associated with potable water treatment. Physiol Entomol. 28:315-322.
Ólafsson J. 1992. Chemical characteristics and trace elements of Thingvallavatn. Oikos. 64:151-161.
Ólafsson JS, Ingimundardóttir GV, Hansen I, Sigurðardóttir SG. 2010. Macroinvertebrate assemblages in effluent water from the high temperature geothermal areas of Krafla, Ölkelduháls and Miðdalur in Hengill, Iceland. Institute of Freshwater Fisheries, VMST/10019, 64 p. Icelandic with English summary.
Parmesan C. 2006. Ecological and evolutionary responses to recent climate change. Ann Rev Ecol Evol Syst. 37:637-669.
Pinder LCV. 1997. The habitats of chironomid larvae. Chapter 6. In: Armiatage P, Cranston PS, Pinder LCV, editors. The Chironomidae. The biology and ecology of non-biting midges. London (UK): Chapman & Hall. p. 107-135.
Rybach L. 2005. Environmental aspects of geothermal development and utilization, and related legal, institutional and social implications. In: Popovski K, editor. Proceedings of the World Geothermal Congress. Short Pre- and Post-Congress Courses., 24–29 Apr 2005., Antalya (Turkey). p. 297-324.
Sandlund OT, Jónasson PM, Jonsson B, Lindem T, Malmquist HJ, Skúlason S, Snorrason SS. 1992. Threespine stickleback Gasteros-teus aculeatus in Thingvallavatn: habitat and food in a lake dominated by arctic charr Salvelinus alpinus. Oikos. 64:365-370.
Snorrason SS. 1982. The littoral ecosystem and the ecology of Lymnaea peregra in Lake Thingvallavatn, Iceland. [dissertation]. [Liverpool (UK)]:University of Liverpool. 429 p.
Snorrason SS. 2000. Life cycle of the mollusc Lymnaea peregra and its influence on morph formation in Artcic charr, Salvelinus alpinus. Verh Internat Verein Limnol. 27:3214-3217.
Snorrason SS, Jónsson GS. 2000. Assessment of trace elements in biological samples from Varmagjá and Vatnskot. University of Iceland, Institute of Biology. Report no. III. 19 p. Icelandic.
Sturlaugsson J, Jörundsdóttir HÓ, Georgsson F, Gunnlaugsdóttir H. 2009. Mercury and other ecotoxicological trace elements in Lake Þingvallavatn brown trout (Salmo trutta). Matis - Food Research, Innovation & Safety. Report No. 48-09. 28 p. Icelandic wth English summary.
Sveinbjörnsson E. 2009. Winter ice on lake Þingvallavatn – favorable climate indicator. Náttúrufræðingurinn. 78:66-76. Icelandic with English summary.
Thomas DL, McClintock JB. 1996. Aspects of the population dynamics and physiological ecology of the gastropod Physella cubensis (Pulmonata: Physidae) living in a warm-temperate stream and ephemeral pond habitat. Malacologia. 37:333-348.
Þórðarson T. 1983. Økologien til ferskvannssneglene Lymnaea peregra (Müller) og Gyraulus laevis (Alder) i to eutrofe varme kilder på Island; [master‘s thesis]. [Oslo (Norway)]: University of Oslo. Norwegian.
Tuxen SL. 1944. The hot springs, their animal communities and their zoogeographical significance. The Zoology of Iceland. I (II). Copenhagen (Denmark): Einar Munksgaard. 206 p.
Valfells A, Friðleifsson I, Helgason T, Ingimarsson J, Thoroddsson G, Sophusson F. 2005. Sustainable generation and utilization of energy. The case of Iceland. World Energy Council, Congress, 19 Sep 2004. Sydney (Australia). 14 p.
Walther GR, Post E, Convey P, Menzel A, Parmesan C, Beebee TJC, Fromentin J-M, Hoegh-Guldberg O, Bairlein F. 2002. Ecological responses to recent climate change. Nature. 416:389-395.
Wetang’ula G. 2004. Assessment of geothermal wastewater disposal effects: Case studies: Nesjavellir (Iceland) and Olkaria (Kenya). [master’s thesis]. [Reykjavík (Iceland)]: University of Iceland,
Wetang’ula G, Snorrason SS. 2005. Geothermal wastewater disposal: chemical stress assessment – Lake Thingvallavatn, Iceland. Proceedings World Geothermal Congress 2005. 249 April 2005. Antalya (Turkey). p 1-15.
Winterbourn MJ. 1968. The faunas of thermal waters in New Zealand. Tuatara. 16:111-122.
Winterbourn MJ. 1969. The distribution of algae and insects in hot spring thermal gradients at Waimangu, New Zealand. N Z J Mar Freshw Res. 3:459-465.
Woodward G, Dypkær JB, Ólafsson JS, Gíslason GM, Hannesdóttir ER, Friberg N. 2010. Sentinel systems on the razor‘s edge: effects of warming on Arctic geothermal stream ecosystems. Global Change Biol. 16:1979-1991.
Zarandi, SSMM, Ivarsson G. 2010. A review on waste water disposal at the Nesjavellir geothermal power plant. Proceedings World Geothermal Congress 2010. 25–29 Apr 2010. Bali (Indonesia). p. 1-11.




















