
Effects of day-length and temperature on floral structure
10
Effects of day-length and temperature on floral structure and fertility restoration in a season-dependent male-sterile Solanum villosum Mill. mutant Christopher O. Ojiewo Kenji Murakami Peter W. Masinde Stephen G. Agong Masaharu Masuda Received: 6 November 2006 / Accepted: 17 April 2007 / Published online: 22 May 2007 Ó Springer Science+Business Media B.V. 2007 Abstract Solanum villosum is an important African leafy vegetable whose yield is limited mainly by competition from early and excess fruit-set. Induced male-sterility is a potential tool to reduce this competition and enhance yields. This study was conducted to investigate the influence of photope- riod and temperature on the floral dynamics of a season-dependent male-sterile mutant. The mutant, named T-5, has flowers which are sepaloid, mostly stamenless, indeterminate and partially restored in winter, late-spring, summer and autumn, respec- tively. Floral organ restoration was found to be largely independent of photoperiod conditions. Day/ night temperatures of 25/25 and 30/208C were found to favour restoration of the floral organ but most flowers were stamenless and infertile. High night temperature favoured the formation of indeterminate flowers both in the growth chamber (308C) and in the greenhouse (>258C). On the other hand, low growth chamber (108C) and greenhouse (<158C) night temperature favoured the formation of sepal- oid flowers. The optimum temperatures for floral structure and fertility restoration were between 208C and 258C (day) and 15–208C (night). Propagation of T-5 mutant can thus be achieved by growing in regions or seasons with such temperature ranges. Under temperatures unfavourable for fruit-set, leaf productivity is expected to be high. Keywords Abnormal floral organ Fertility Restoration Indeterminate flower Introduction Solanum nigrum complex is a conglomeration of several Solanaceous taxa widely consumed as leafy vegetables in most parts of Africa and South–East Asia. The taxa in this section of African origin are commonly referred to as African nightshade of which S. villosum shows the widest distribution in the tropics (Edmonds and Chweya 1997). The leaves contain high levels of vitamins (especially A, B and C), mineral fibres (such as iron, calcium and phos- phorus), carbohydrates and proteins. They also con- tain phenolics and alkaloids, such as nicotine, quinine, cocaine, and morphine, which are known for their medicinal attributes (Kokwaro 1993). The demand for foods containing substances with phyto- therapeutic (herbal medicine) activity is on the rise C. O. Ojiewo K. Murakami P. W. Masinde S. G. Agong M. Masuda (&) Graduate School of Natural Science and Technology, Okayama University, 1-1-1 Tsushima Naka, Okayama, Okayama 700-0080, Japan e-mail: [email protected] P. W. Masinde S. G. Agong Department of Horticulture, Jomo Kenyatta University of Agriculture and Technology, P. O. Box 62000, Nairobi 00200, Kenya 123 Euphytica (2007) 158:231–240 DOI 10.1007/s10681-007-9445-z
Transcript of Effects of day-length and temperature on floral structure

Effects of day-length and temperature on floral structureand fertility restoration in a season-dependent male-sterileSolanum villosum Mill. mutant
Christopher O. Ojiewo Æ Kenji Murakami ÆPeter W. Masinde Æ Stephen G. Agong ÆMasaharu Masuda
Received: 6 November 2006 / Accepted: 17 April 2007 / Published online: 22 May 2007
� Springer Science+Business Media B.V. 2007
Abstract Solanum villosum is an important African
leafy vegetable whose yield is limited mainly by
competition from early and excess fruit-set. Induced
male-sterility is a potential tool to reduce this
competition and enhance yields. This study was
conducted to investigate the influence of photope-
riod and temperature on the floral dynamics of a
season-dependent male-sterile mutant. The mutant,
named T-5, has flowers which are sepaloid, mostly
stamenless, indeterminate and partially restored in
winter, late-spring, summer and autumn, respec-
tively. Floral organ restoration was found to be
largely independent of photoperiod conditions. Day/
night temperatures of 25/25 and 30/208C were found
to favour restoration of the floral organ but most
flowers were stamenless and infertile. High night
temperature favoured the formation of indeterminate
flowers both in the growth chamber (308C) and in
the greenhouse (>258C). On the other hand, low
growth chamber (108C) and greenhouse (<158C)
night temperature favoured the formation of sepal-
oid flowers. The optimum temperatures for floral
structure and fertility restoration were between 208Cand 258C (day) and 15–208C (night). Propagation of
T-5 mutant can thus be achieved by growing in
regions or seasons with such temperature ranges.
Under temperatures unfavourable for fruit-set, leaf
productivity is expected to be high.
Keywords Abnormal floral organ � Fertility
Restoration � Indeterminate flower
Introduction
Solanum nigrum complex is a conglomeration of
several Solanaceous taxa widely consumed as leafy
vegetables in most parts of Africa and South–East
Asia. The taxa in this section of African origin are
commonly referred to as African nightshade of which
S. villosum shows the widest distribution in the
tropics (Edmonds and Chweya 1997). The leaves
contain high levels of vitamins (especially A, B and
C), mineral fibres (such as iron, calcium and phos-
phorus), carbohydrates and proteins. They also con-
tain phenolics and alkaloids, such as nicotine,
quinine, cocaine, and morphine, which are known
for their medicinal attributes (Kokwaro 1993). The
demand for foods containing substances with phyto-
therapeutic (herbal medicine) activity is on the rise
C. O. Ojiewo � K. Murakami � P. W. Masinde �S. G. Agong � M. Masuda (&)
Graduate School of Natural Science and Technology,
Okayama University, 1-1-1 Tsushima Naka, Okayama,
Okayama 700-0080, Japan
e-mail: [email protected]
P. W. Masinde � S. G. Agong
Department of Horticulture,
Jomo Kenyatta University of Agriculture and Technology,
P. O. Box 62000, Nairobi 00200, Kenya
123
Euphytica (2007) 158:231–240
DOI 10.1007/s10681-007-9445-z

worldwide (Duke 1990; Prohens et al. 2003). In
Kenya, as well as most parts of Africa and South–
East Asia, the perception of traditional leafy vegeta-
bles as ‘‘healthy’’ foods has accounted for a sudden
upsurge in their consumption, making them promis-
ing alternative cash crops locally and for export
(Schippers 2000).
Limited by a prolific early flowering and exces-
sive fruit-set which compete with leaf productivity,
the leaf yields of these important vegetables are,
however, relatively low. Induction and introduction
of male-sterile varieties has been proposed as an
economically viable and technically feasible strat-
egy to circumvent the post-anthesis sink-source
imbalance, improved and stabilize leaf yields
Ojiewo et al. (2005). We have induced four genic
male-sterile mutant types through seed irradiation
with c-ray namely: acetocarmine-stained unviable
pollen (T-1), defective pollen not stainable with
acetocarmine (T-2), pollenless (T-3), and low pollen
producing (T-4) types (Ojiewo et al. 2005). In
addition a novel male-sterile mutant with abnormal
floral organs (T-5) was isolated from one line
irradiated with 20 Gy 12C5+ (Ojiewo et al. 2006a).
Genic male-sterile lines pose a challenge in their
maintenance and propagation. Their maintenance
requires backcrossing with a heterozygote line, but
the progeny produced is 50% fertile and 50% male-
sterile. Partial fertility restoration of the M2 prog-
eny, desirable for maintenance and propagation,
was observed in some of the T-1*T-4 lines in
autumn, but the restoration was enhanced in the
subsequent M3 progeny, posing a threat in the use
of these lines as new varieties with high leaf yields
(Ojiewo et al. 2006b).
The abnormal floral organ mutant (T-5) has
exhibited a stable season-dependent structural
dynamics and fertility restoration. The M2, M3 and
M4 flowers were sepaloid from late-winter to mid-
spring, mostly stamenless in late-spring, indetermi-
nate in summer and partially restored with fruit-set
and seed-set in autumn. The objectives of the current
study were to (1) examine the possible influence of
day-length and/or temperature on this structural
dynamism and the eventual fertility restoration; (2)
determine the critical daylength and /or temperature
for fertility restoration to enable synchronizing of
seed (for propagation) and leaf production with
environmental conditions.
Materials and methods
Effects of daylength on floral organ structure and
fertility restoration
Seed from M3 generation of T-5 plants were sown in
cell trays containing vermiculite on 1 March 2005
and germinated seedlings raised in pots containing
commercial forest soil in a greenhouse. At floral bud
initiation, 3 plants each were transferred to 3 growth
chambers set at a constant day/night temperature of
25/258C and daylengths of 8, 12 and 16 h. All initial
floral buds were removed to give a uniform start. On
each of the plants, 15 floral trusses were sampled
from newly differentiating inflorescences and scored
for floral organ abnormalities, fruit-set and number of
seeds per fruit between 1 May and 15 June, 2005.
Although most flowers opened, fruit-set was negligi-
ble in all treatments. Many flowers at anthesis were
either stamenless or had distorted stamens, which
aborted a few days after anthesis.
In vitro pollen germination capacity, pollen
fertilization capacity on-stigma and stigma
viability
Pollen was collected at anthesis from normal looking
stamens and from greenhouse grown wild-type plants
for comparison. For in-vitro pollen germination, the
pollen grains were cultured in drops of a modified BK
medium (Brewbaker and Kwack 1963) consisting of
10% sucrose and 50 ppm H3BO3 in deionized
distilled H2O supplemented with 100 ppm CaCl2.
The pH of the medium was adjusted to 6.0. Hanging
drop cultures were prepared by placing three 30 ll
drops of the growth medium on 30 mm Petri-dishes.
The anthers were held with forceps and gently tapped
onto each drop to release small amounts of pollen.
The Petri dishes were then gently overturned onto
Petri dish covers with moist filter papers. Pollen was
left to germinate in an incubator set at 258C and
observed under a light microscope after 1.5*2 h.
Pollen was considered to have germinated when the
pollen tube was equal to or more than the pollen grain
diameter (Heslop-Harrison et al. 1984). For each
drop, about 100 pollen grains were scored and for
each daylength treatment three replicates were used.
Flowers from 5 trusses each of 3 wild-type plants
raised in the greenhouse were emasculated 2 days
232 Euphytica (2007) 158:231–240
123

before anthesis and pollinated at anthesis with pollen
from each of the daylength treatments. At the same
time, 5 floral trusses were selected from each
daylength treatment and stamenless flowers polli-
nated with pollen from the wild-type plants. The rate
of fruit-set, number of seeds per fruit and the
germination capacity of F1 seed resulting from the
each pollination were examined.
Effect of temperature on floral organ structure and
fertility restoration
To examine the effect of night temperature, plants
maintained by cutback were transferred to growth
chambers where night temperatures were set at 30, 20
and 108C. A neutral daylength (12/12 h) was
maintained and day temperature was set at 308C in
all the 3 chambers. There was no fruit-set in any of
the set-ups. To determine the optimum day/night
temperature integrals for fruit-set, new seedlings
were raised as above in a greenhouse where mini-
mum temperature was maintained at 128C between
15 Jan and 28 Feb 2006. Just before flower bud
initiation, three plants each were transferred to 3
separate growth chambers where a neutral daylength
(12/12 h) was maintained and day/night temperatures
were set at 25/20, 25/15 and 20/158C, respectively.
The rate of occurrence of floral organ abnormalities
and fruit- and seed-set in restored flowers were scored
from inflorescences sampled as above. The remaining
plants were raised in the greenhouse where similar
data was collected from 3 plants at the beginning,
middle and end of March, April and May. In addition,
in vitro pollen germination capacity of greenhouse
raised T-5 plants was investigated and compared with
wild-type normal pollen.
Statistical analysis
With only 3 phytochambers each with a different
lighting and temperature regime there was no true
replication in this experiment, so variability between
plants within compartments was used as a proxy
error. The experiment assumes a completely random-
ized design (CRD) with 3 replicate plants per
treatment. For ease and consistency in data analysis,
a similar set up was repeated in the greenhouse
experiments. All statistical analyses were performed
using SPSS. Analysis of variance (ANOVA) was
used to determine if the means of the treatments were
significantly different. Discrete variables, such as the
number of seeds per fruit, were analyzed after square
root transformation and proportions were analyzed
after logarithmic transformation to stabilize the
variance among groups. Differences in the parame-
ters measured between the different lighting and
temperature regimes were evaluated based on LSD
(P � 0.05). Results are presented as the mean ± SE
for n = 3. Means within the same column and
experiment followed with different letters were
significantly different.
Results
Effects of daylength on floral organ structure and
fertility restoration
All floral buds in the growth chamber as well as the
greenhouse were sepaloid at initiation regardless of
daylength or temperature treatment. While the wild-
type flowers opened 2–3 days after flower bud
initiation, T-5 floral organ differentiation occurred
2 weeks after initiation of the sepaloid buds. Some
flower buds aborted before any differentiation was
observed, while others aborted at ‘anthesis’ (Fig. 1).
However, all computations were based on the orig-
inally initiated flower buds (5–8 flowers per inflores-
cence) and the differences discussed on the basis of
their significance in agricultural application.
While maintaining the day and night temperature
constant at 258C and only varying the photoperiod,
Fig. 1 Wild-type floral
buds at anthesis (a), T-5
flowers initiated at the same
time as the wild-type still in
sepaloid state (b) and
differentiating T-5 flowers
after 2 weeks (c)
Euphytica (2007) 158:231–240 233
123

there were negligible incidences of sepaloid or
indeterminate flowers (Table 1). Apart from a few
flowers that aborted, more than 90% of the remaining
flowers opened, regardless of the photoperiod. How-
ever, considering that fruit-set of wild-type plants in
the greenhouse was perfect (100%), the fruit-set in
the open T-5 flowers was considered negligible
(0*4%) in all the 3 set-ups. Most of the fruits were
malformed with no seeds even at maturity (Fig. 2a, b)
while the seeded fruits (Fig. 2c, d) had very few seeds
per fruit (9*13) compared to >60 seeds per fruit in
wild-type plants.
A closer observation of the floral structure
revealed that 30–40% of the flowers had malformed
stamens (Fig. 3a) that dropped off within 3 days after
anthesis. A significant proportion of the opening
flowers were, thus, left stamenless. In addition, the
unaborted stamens exhibited other abnormalities such
as fusion to petals (Fig. 3b) or fusion to carpels
(Fig. 3c). Besides, there were also malformations
such as splitting of the styles and stigmas (Fig. 3d),
unfused multiple styles with no distinct stigmas
(Fig. 3e) and unfused multiple styles and stigmas
(Fig. 3f). To this end it was hypothesized that failure
of fruit-set was due to these stamen and carpel
abnormalities.
In vitro pollen germination capacity, pollen
fertilization capacity on-stigma and stigma
viability
To test the viability of the T-5 pollen, in-vitro
germination tests were carried out and compared
with wild-type pollen. At the same time the pollen
was used to pollinate emasculated flowers of wild-
type plants in the greenhouse. To test the viability
of the T-5 carpels, stamenless flowers were polli-
nated with wild-type pollen. T-5 pollen germination
Table 1 Effect of daylength on the occurrence of floral organ abnormalities, fruit- and seed-set
Daylength (hr)z Frequency of abnormalities (%) Fruit-set (%) No. seeds per fruit
Sepaloid Stamenless Indeterminate
8 1.3 ± 0.0a 40.2 ± 2.3a 7.0 ± 0.2a 0.0 –
12 3.1 ± 0.1a 30.5 ± 1.8b 4.2 ± 0.1a 1.4 ± 1.0a 13.2 ± 0.6a
16 3.5 ± 0.1a 40.8 ± 2.1a 0.0 3.9 ± 1.2a 9.3 ± 1.1a
z Growth chamber day and night temperature was maintained constant at 258C throughout the experiment. Data are means ± SE;
n = 3. Means within the same column sharing the same letters are not significantly different based on LSD0.05
Fig. 2 Fruit abnormalities
observed under constant
day/night temperature of
25/258C with varying
photoperiods. Some
malformed fruits with
vegetative parts attached (a)
remained small and seedless
even at maturity (b), while
others had very few seeds
(c and d)
234 Euphytica (2007) 158:231–240
123

in-vitro was significantly lower (11*22%) than that
of wild-type pollen (55*61%). In addition, fruit-set
with T-5 pollen on W-T flowers was considerably
low (16*27%), with only a few seeds per fruit
(12*19). On the other hand, there was a high rate
of fruit-set (85*94%) when wild-type pollen was
used to pollinate the open T-5 flowers, with the
mean number of seeds per fruit ranging from 48 to
63. There were minimal fruit abnormalities (Fig. 4b,
c), though not ultimately absent. Save for many
large sepals still attached to the fruit-stalk, most of
the fruits were properly formed (Fig. 4d) with a
near wild-type fruit shape (Fig. 4a). The germina-
tion capacity of both seed lots was high (84*91%)
with no differences of agricultural significance
(Table 2).
Effect of temperature on floral organ structure and
fertility restoration
There were significant differences (P < 0.05) between
the three night temperature regimes with regard to
floral organ structure abnormalities and its restora-
tion. Maintaining the daylength at 12 h and the day
temperature at 308C, high night temperature (308C)
resulted mostly in indeterminate flowers, low night
temperature (108C) in sepaloid flowers, while mod-
erate night temperature (208C) resulted in some
anthesis (Table 3). Open flowers under 30/208C had
similar abnormalities observed during the photope-
riod experiment under 25/258C, with 13.3% of the
flowers being stamenless. However, there was no
fruit-set in any of the 3 night temperature set-ups.
With day and night temperature integrals of 25/158Cand 20/158C and 25/208C, there was minimal flower
abortion before anthesis. Although all initial flowers
were sepaloid, floral organ differentiation occurred
within 2 weeks as expected with no flowers retaining
the sepaloid form, or becoming indeterminate and the
frequency of stamenless open flowers was negligible
(1–2%). Although not comparable to the wild-type
plants, fruit-set in all the three set-ups was signifi-
cantly high (40*50%), with 25/208C being the
highest. At the same time, cases of fruit malformation
were minimal. The mean number of seeds per fruit
was also significantly higher (&40) than the earlier
set-ups (Table 4). There were no significant differ-
ences in the floral organ restoration and the rate of
fruit- and seed-set among the 3 day and night
temperature integrals.
Fig. 3 Types of stamen and
carpel abnormalities
observed under day/night
temperature of 30/208C and
25/258C with various
photoperiods. Top row, left
to right: stamen abortion
(a), fusion to petals (b) and
fusion to carpels (c). Lower
row, left to right: carpel
splitting (d), multiple
carpels with no distinct
stigmas (e) and unfused
carpels and stigmas (f)
Fig. 4 Varying degrees of fruit abnormalities (b and c) observed when normal pollen was used to pollinate T-5 plants raised under
25/258C with various photoperiods. Most fruits were normal (d) and comparable to wild type fruits (a)
Euphytica (2007) 158:231–240 235
123

In the greenhouse, the frequency of sepaloid
flowers was high at the beginning of March and
continued to reduce up to late April when the night
temperature was still <158C (Table 5). However,
from early May when the night temperature had risen
beyond 158C, no flower retained the sepaloid form
beyond 2 weeks after initiation of floral buds.
Stamenless flowers were observed from mid-April
and the frequency continued to increase up to early-
May, and then reduced thereafter. Indeterminate
flowers started to appear in early-May when the day
temperatures had risen above 258C and continued to
increase thereafter as summer approached. In-vitro
pollen germination capacity and the corresponding
fruit-set increased from early-March to late-April,
and then decreased thereafter. The number of seeds
per fruit followed the same trend but started declining
between early and mid April and continued in the
same trend till the end of the experiment in late May.
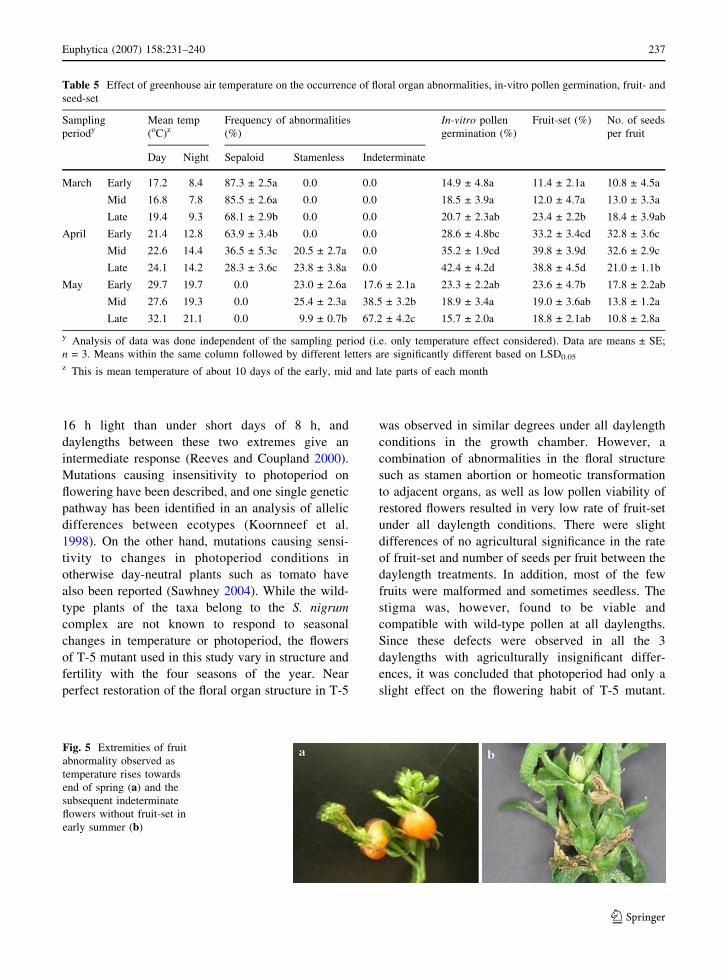
Incidentally, the proportion of malformed fruits
increased concomitantly with increase in the green-
house temperature from early March to late May,
with the latest fruits being seedless (Fig. 5a), fol-
lowed by no fruit-set (Fig. 5b).
Discussion
Many species have an obligatory requirement for the
correct photoperiod for transition from vegetative to
reproductive phase. Flowering in the plant model
Arabidopsis occurs much earlier under long days of
Table 2 Effect of daylength on in-vitro pollen germination, fertilization and seed germination capacity
Source of T-5 Cross In vitro pollen germination (%)z Fruit-set (%) No. of seeds per fruit Seed germination (%)
8h T-5 x WT 61.2 ± 2.6a 85.4 ± 4.2a 56.3 ± 3.3ab 84.3 ± 2.9a
WT x T-5 11.3 ± 1.1c 16.8 ± 1.2c 13.2 ± 1.4c 90.7 ± 2.4a
12h T-5 x WT 58.7 ± 1.6a 94.1 ± 2.6a 48.9 ± 4.1b 89.3 ± 5.6a
WT x T-5 13.1 ± .08c 27.5 ± 3.4b 19.4 ± 2.4c 89.7 ± 2.3a
16h T-5 x WT 55.5 ± 2.2a 89.5 ± 6.7a 62.8 ± 5.6a 89.3 ± 2.8a
WT x T-5 22.4 ± 2.3b 24.3 ± 1.7bc 12.6 ± 2.3c 87.0 ± 6.4a
z This is the pollen germination of the pollen parent (not the seed parent). Data are means ± SE; n = 3. Means within the same
column sharing the same letters are not significantly different based on LSD0.05
Table 3 Effect of night temperature on the occurrence of floral organ abnormalities, fruit- and seed-set
Night temp. (8C)z Frequency of abnormalities (%) Fruit-set No. seeds per fruit
Sepaloid Stamenless Indeterminate
10 75.8 ± 9.7a 0.0 14.2 ± 1.9a 0.0 –
20 34.6 ± 3.6b 13.3 ± 3.6 28.4 ± 2.1b 0.0 –
30 24.3 ± 5.6c 0.0 65.7 ± 3.4c 0.0 –
z Growth chamber day temperature was maintained constant at 308C while the daylength was neutral (12 h) throughout the
experiment. Data are means ± SE; n = 3. Means within the same column followed by different letters are significantly different based
on LSD0.05
Table 4 Combined effect of day and night temperature on the occurrence of floral organ abnormalities, fruit- and seed-set
Temperature (oC) Frequency of abnormalities (%) Fruit-set (%) No. seeds per fruit
Day (12h) Night (12h) Sepaloid Stamenless Indeterminate
20 15 0.0 2.1 ± 0.3a 0.0 38.6 ± 4.9a 37.0 ± 2.5a
25 15 0.0 1.4 ± 0.1a 0.0 41.7 ± 5.6a 38.7 ± 3.6a
25 20 0.0 0.0 0.0 54.5 ± 3.3b 37.5 ± 4.0a
236 Euphytica (2007) 158:231–240
123
16 h light than under short days of 8 h, and
daylengths between these two extremes give an
intermediate response (Reeves and Coupland 2000).
Mutations causing insensitivity to photoperiod on
flowering have been described, and one single genetic
pathway has been identified in an analysis of allelic
differences between ecotypes (Koornneef et al.
1998). On the other hand, mutations causing sensi-
tivity to changes in photoperiod conditions in
otherwise day-neutral plants such as tomato have
also been reported (Sawhney 2004). While the wild-
type plants of the taxa belong to the S. nigrum
complex are not known to respond to seasonal
changes in temperature or photoperiod, the flowers
of T-5 mutant used in this study vary in structure and
fertility with the four seasons of the year. Near
perfect restoration of the floral organ structure in T-5
was observed in similar degrees under all daylength
conditions in the growth chamber. However, a
combination of abnormalities in the floral structure
such as stamen abortion or homeotic transformation
to adjacent organs, as well as low pollen viability of
restored flowers resulted in very low rate of fruit-set
under all daylength conditions. There were slight
differences of no agricultural significance in the rate
of fruit-set and number of seeds per fruit between the
daylength treatments. In addition, most of the few
fruits were malformed and sometimes seedless. The
stigma was, however, found to be viable and
compatible with wild-type pollen at all daylengths.
Since these defects were observed in all the 3
daylengths with agriculturally insignificant differ-
ences, it was concluded that photoperiod had only a
slight effect on the flowering habit of T-5 mutant.
Table 5 Effect of greenhouse air temperature on the occurrence of floral organ abnormalities, in-vitro pollen germination, fruit- and
seed-set
Sampling
periodyMean temp
(oC)zFrequency of abnormalities
(%)
In-vitro pollen
germination (%)
Fruit-set (%) No. of seeds
per fruit
Day Night Sepaloid Stamenless Indeterminate
March Early 17.2 8.4 87.3 ± 2.5a 0.0 0.0 14.9 ± 4.8a 11.4 ± 2.1a 10.8 ± 4.5a
Mid 16.8 7.8 85.5 ± 2.6a 0.0 0.0 18.5 ± 3.9a 12.0 ± 4.7a 13.0 ± 3.3a
Late 19.4 9.3 68.1 ± 2.9b 0.0 0.0 20.7 ± 2.3ab 23.4 ± 2.2b 18.4 ± 3.9ab
April Early 21.4 12.8 63.9 ± 3.4b 0.0 0.0 28.6 ± 4.8bc 33.2 ± 3.4cd 32.8 ± 3.6c
Mid 22.6 14.4 36.5 ± 5.3c 20.5 ± 2.7a 0.0 35.2 ± 1.9cd 39.8 ± 3.9d 32.6 ± 2.9c
Late 24.1 14.2 28.3 ± 3.6c 23.8 ± 3.8a 0.0 42.4 ± 4.2d 38.8 ± 4.5d 21.0 ± 1.1b
May Early 29.7 19.7 0.0 23.0 ± 2.6a 17.6 ± 2.1a 23.3 ± 2.2ab 23.6 ± 4.7b 17.8 ± 2.2ab
Mid 27.6 19.3 0.0 25.4 ± 2.3a 38.5 ± 3.2b 18.9 ± 3.4a 19.0 ± 3.6ab 13.8 ± 1.2a
Late 32.1 21.1 0.0 9.9 ± 0.7b 67.2 ± 4.2c 15.7 ± 2.0a 18.8 ± 2.1ab 10.8 ± 2.8a
y Analysis of data was done independent of the sampling period (i.e. only temperature effect considered). Data are means ± SE;
n = 3. Means within the same column followed by different letters are significantly different based on LSD0.05
z This is mean temperature of about 10 days of the early, mid and late parts of each month
Fig. 5 Extremities of fruit
abnormality observed as
temperature rises towards
end of spring (a) and the
subsequent indeterminate
flowers without fruit-set in
early summer (b)
Euphytica (2007) 158:231–240 237
123

Further, it was postulated that the seasonal changes in
T-5 flower were dependent on temperature, and that a
constant day/night temperature of 25/258C could
favour partial floral organ structure but not fertility
restoration.
Temperature conditions are known to influence
the development of floral organs and sex expression
in many species of flowering plants. When tomato
and pepper plants (two Solanaceous vegetables)
were exposed to low temperatures, they produced
flowers showing alterations in the number, morphol-
ogy, and pattern of fusion of floral organs. Conse-
quently, malformed fruits were produced from these
flowers (Sawhney 1983; Polowick and Sawhney
1985). However, there were striking differences in
the response of tomato and pepper to low temper-
ature. For example, in pepper flowers, low temper-
ature inhibited the enlargement of petals and
stamens but enhanced the same in tomato. In
addition, low temperature induced male-sterility in
pepper but not in tomato. On the other hand, low
temperature resulted in increased number of petals,
stamens and locules in tomato but not in pepper.
These results suggest that temperature conditions
may induce different responses in different species
within the same family. Our results under day/night
temperatures of 258C and 30/208C have some
similarities to these observations, though the tem-
perature was not low. However, at low temperatures
our results are completely different from these
reports.
Work with Arabidopsis thaliana and Antirrhinum
majus has allowed the genetic and molecular char-
acterization of several families of regulatory genes
that control flower initiation and development. In
these species and many flowering plants, floral organ
identity is regulated by homeotic functions of genes
mostly of the MADS-box class. A gene-based
‘‘ABCDE’’ model outlining the overlapping activi-
ties of five classes of regulatory genes, which are
responsible for the identity of the five floral organs
has been developed: A and E for the sepals; A, B and
E for the petals; B, C and E for the stamens; C and E
for the carpels; and C, D and E for the ovules
(Theissen 2001). Mutations affecting the proper
function of any of these genes promote homeotic
transformations and, consequently, abnormal floral
organogenesis. Temperature has been reported to
affect the function of floral structure regulatory genes
in Antirrhinum (Zachgo et al. 1995) and Arabidopsis
(Sablowski and Meyerowitz 1998). In both cases,
mutant plants grown at low temperature (15–168C)
produce nearly wild-type flowers, but growth at the
nonpermissive temperature (26*288C) causes the
second-whorl organs to develop as sepals (only A
function present) and stamens to be replaced by
carpelloid organs (only C function left in the third
whorl). In this study, both 30/208C and 25/258C were
found to favour anthesis in T-5 but not fruit-set. High
(308C) and low (108C) night temperature extremes
hampered flower restoration, with the former favour-
ing indeterminate flower development while the latter
favoured sepaloid flowers. An interplay between
night and day temperature was necessary to achieve
both floral organ and fertility restoration, the opti-
mum temperatures being between 208C–258C (day)
and 15–208C (night). The response of the floral organ
development and fertility restoration was similar in
the greenhouse as in the growth chamber.
We have previously described the homeotic
changes in T-5 mutant flower in comparison to the
wild-type flowers (Ojiewo et al. 2006a). As expected
for homeotic mutants, these changes reproduced the
organ identity of adjacent whorls and the intermedi-
ate organs formed kept the growth pattern character-
istic of the whorl they occupy. The observed
homeotic transformations followed normal patterns
in their time of appearance, the kind of transforma-
tion, and the whorls affected. Regarding the time of
appearance, the homeotic transformations took place
late in T-5 floral organ development, giving rise to
chimeric organs such as petaloid sepals and capelloid
stamens. As a result there were more carpels than the
standard single carpel in wild-type flower. However,
observations from this study suggest that splitting
events taking place in the carpel could also contribute
to this phenomenon. Meristematic mutations altering
the number of floral organs have been reported in
Arabidopsis. Genes such as CLAVATA1 (CLV1) and
CLAVATA3 (CLV3) seem to be required to regulate
the size of the shoot and floral meristems, which in
turn affects the number of floral organs that develop
in each whorl. Weak alleles at loci clv1 and clv3
plants develop enlarged shoot and floral meristems,
which give rise to flowers with additional organs in
each whorl (Clark et al. 1993, 1995, 1997). These
phenotypic similarities could suggest an influence of
temperature on the expression or activity of the
238 Euphytica (2007) 158:231–240
123

corresponding T-5 genes in flowers restored under
30/25 or 25/258C. Unfortunately, T-5 orthologs of
any floral meristem or floral organ identity genes
have not been isolated, which precludes any analysis
of the effects of temperature on their expression at the
moment.
Zachgo et al. (1995) reported that sensitivity to
temperature in Antirrhinum def-101 was a property of
a mutant protein which loses its active conformation
or the ability to interact functionally with other
proteins under non-permissive temperatures. On the
other hand, Sablowski and Meyerowitz (1998) dem-
onstrated that temperature sensitivity in Arabidopsis
ap3–1 mutant was caused by a temperature-sensitive
splicing defect resulting in unstable RNA–RNA
interactions. Since there are no reports (to the best
of our knowledge ) of any mutant plant species whose
floral organ structure varies from season to season
with accurate repetition of the pattern every year, the
dynamic nature of the T-5 flower structure is of
potential biochemical and physiological significance
in floral genetics. On the other hand, these charac-
teristics are of various potential applications from a
breeding and agronomic point of view. Seed produc-
tion for maintenance and propagation purposes can be
done by temporal or spatial manipulation to synchro-
nize flowering with temperatures of 20–258C (day)
and 15–208C (night). Taking the case of Kenya (and
most parts of Africa) where the wild-type S. villosum
was obtained, this can be achieved by producing T-5
plants in cooler months of the year such as June–
August or in highland regions which remain cool
throughout the year. When and where fruit-set is
hampered (sepaloid, stamenless, and indeterminate),
the competition between the vegetative and repro-
ductive function is eliminated hence leaf yield is
expected to be high. Regardless of photoperiod
conditions, hybrid seed production using T-5 as the
female parent could potentially be done under
temperatures that favour stamenless flower formation
with minimum contamination risk. Environmental
modulation of male-sterility has been useful in plant
breeding (reviewed by Perez-Prat and Campagne
2002). For example, the use of thermo-photosensitive
genic male-sterile mutants provides the opportunity
to produce rice hybrid seed by a two-line system. For
maintenance, the male-sterile female line can be
propagated by growing it under environmental con-
ditions that restore fertility (Wu et al. 2003).
Conclusions and recommendations
As there were no significant differences between 8,
12 and 16 h daylength with regard to floral organ
structure restoration, fruit- and seed-set, it is plausible
to conclude that the African nightshade T-5 mutant is
photoperiod-insensitive. The results suggest rather
that it is temperature-sensitive, the critical minimum
and maximum temperatures for floral organ and
fertility restoration being 158C and 258C, respec-
tively. The male function of the T-5 mutant flower
can be eliminated when not needed by growing in
restrictive temperatures and restored when needed by
growing in permissive temperatures. Thus, produc-
tion of fruit and seed (which are the major sink of the
plant) can be eliminated to improve leaf productivity
and restored for purposes of propagation. Further
analysis of wild-type and T-5 mutant through gene
cloning and sequencing, gene expression studies and
phylogenetic constructions is necessary to identify
the mutant lesions resulting in the T-5 phenotype. In
addition, the causes of temperature sensitivity in the
affected gene and its interaction with other genes
controlling floral organ development and final iden-
tity should be investigated further.
References
Brewbaker JL, Kwack BH (1963) The essential role of calcium
ion in pollen germination and pollen tube growth. Am J
Bot 50:859–886
Clark SE, Running MP, Meyerowitz EM (1993) CLAVATA1, a
regulator of meristem and flower development in Ara-bidopsis. Development 119:397–418
Clark SE, Running MP, Meyerowitz EM (1995) CLAVATA3 is
a specific regulator of shoot and floral meristem devel-
opment affecting the same processes as CLAVATA1.
Development 121:2057–2067
Clark SE, Williams RW, Meyerowitz EM (1997) The CLAV-ATA1 gene encodes a putative receptor kinase that con-
trols shoot and floral meristem size in Arabidopsis. Cell
89:575–585
Duke JA (1990) Promising phytomedicinals. In: Janick J, Si-
mon JE (eds) Advances in new crops. Portland, Timber
Press, OR, USA, pp. 491–498
Edmonds JM , Chewya JA (1997) Black night shades Solanumnigrum L. and related species promoting the conservation
and the use of underutilized and neglected crops 15.
Institute of Plant Genetics and Crop Plant Research,
Gatersleben/international Plant Genetic Resources Insti-
tute, Rome, Italy
Heslop-Harrison J, Heslop-Harrison Y, Shivanna KR (1984)
The evaluation of pollen quality and a further appraisal of
Euphytica (2007) 158:231–240 239
123

the fluorochromatic (FCR) test procedure. Theor Appl
Genet 67:367–375
Kokwaro JO (1993) Medicinal plants of East Africa, 2nd edn.
Kenya Literature Bureau, Nairobi, pp. 58, 157, 224
Koornneef M, Alonso-Blanco C, Blankestijn-de Vries H,
Hanhart CJ, Peeters AJM (1998) Genetic interactions
among late flowering mutants of Arabidopsis. Genetics
148:885–892
Ojiewo CO, Agong SG, Murakami K, Tanaka A, Hase Y,
Masuda M (2005) Male-sterility induced by c-ray irradi-
ation of African nightshade (Solanum nigrum L. ssp.
villosum) seed. J Horticult Sci Biotechnol 80:699–704
Ojiewo CO, Agong SG, Murakami K, Tanaka A, Hase Y
Masuda M (2006a) Biological effect of carbon-ion beam
on mutagenesis and a novel season-dependent floral
homeotic mutant in Solanum villosum Miller. J Horticult
Sci Biotechnol 81:559–564
Ojiewo CO, Murakami K, Masuda M (2006b) A novel male-
sterile mutant with season-dependent floral homeotic trait
induced by 12C5+ ion beam irradiation of African night-
shade (Solanum villosum Mill) seed. Presented at the 27th
International Horticultural Congress. COEX Convention
Centre, Seoul, Korea, 13th–19th August, 2006. IHC 2006
Abstracts. S01-P-139 p 43
Perez-Prat E, Campagne MML (2002) Hybrid seed production
and the challenge of propagating male-sterile plants.
Trends Plant Sci 7:199–203
Polowick PL, Sawhney VK (1985) Temperature effects on
male-fertility and flower and fruit development in Cap-sicum annuum L. Scientia Horticulturae 25:117–127
Prohens J, Rodrı́guez-Burruezo A, Nuez F (2003) New crops:
an alternative for the development of horticulture. Food
Agric Environ 1:75–79
Reeves PH, Coupland G (2000) Response of plant development
to environment: control of flowering by daylength and
temperature. Current Opin Plant Biol 3:37–42
Sawhney VK (1983) Temperature control of male sterility in a
tomato mutant. J Heredity 74:51–54
Sablowski RWM, Meyerowitz EM (1998) Temperature-sensi-
tive splicing in the floral homeotic mutant apetala3–1.
The Plant Cell 10:1453–1463
Sawhney VK (2004) Photoperiod-sensitive male-sterile mutant
in tomato and its potential use in hybrid seed production. J
Horticult Sci Biotechnol 79:138–141
Schippers RR (2000) African indigenous vegetables. An
overview of the cultivated species. Revised CD ROM
Edition. Horticultural Development Services, Chatham,
UK, pp. 25–31, 147–192
Theissen G (2001) Development of floral organ identity: stories
from the MADS house. Current Opin Plant Biol 4:75–85
Wu D, Shen S, Cui H, Xia Y, Shu Q (2003) A novel thermo/
photoperiod-sensitive genic male-sterile (T/PGMS) rice
mutant with green-revertible albino leaf colour marker
induced by gamma irradiation. Field Crops Res 81:141–
147
Zachgo S, Silva EA, Motte P, Trobner W, Saedler H, Schwarz-
Sommer Z (1995) Functional analysis of the Antirrhinumfloral homeotic DEFICIENS gene in vivo and in-vitro
using a temperature sensitive mutant. Development
121:2861–2875
240 Euphytica (2007) 158:231–240
123




















