
Effects of Chromium Histidinate on Renal Function, Oxidative Stress, and Heat-Shock Proteins in...
9
ORIGINAL RESEARCH Effects of Chromium Histidinate on Renal Function, Oxidative Stress, and Heat-Shock Proteins in Fat-Fed and Streptozotocin-Treated Rats Ayhan Dogukan, MD,* Mehmet Tuzcu, PhD,† Vijaya Juturu, PhD,‡ Gurkan Cikim, MD,§ _ Ibrahim Ozercan, MD,{ James Komorowski, MSc,‡ and Kazim Sahin, DVM, PhD # Objective: Chromium is an essential element for carbohydrate, fat, and protein metabolism. The therapeutic po- tential of chromium histidinate (CrHis) in the treatment of diabetes has been elucidated. The present study investi- gated the effects of CrHis on serum parameters of renal function, on oxidative stress markers (malondialdehyde [MDA] and 8-isoprostane), and on the expression of heat-shock proteins (HSPs) in rats. Methods: Male Wistar rats (n 5 60, 8 weeks old) were divided into four groups. Group 1 received a standard diet (12% of calories as fat). Group 2 received a standard diet, plus CrHis. Group 3 received a high-fat diet (40% of cal- ories as fat) for 2 weeks, and was then injected with streptozotocin (STZ) on day 14 (STZ, 40 mg/kg intraperitoneally). Group 4 was treated in the same way as group 3 (HFD/STZ), but was supplemented with 110 mg CrHis/kg/body weight/day. Oxidative stress in the kidneys of diabetic rats was evidenced by an elevation in levels of MDA and 8- isoprostane. Protein concentrations of HSP60 and HSP70 in renal tissue were determined by Western blot analyses. Results: Chromium histidinate supplementation lowered kidney concentrations of MDA, 8-isoprostane levels, se- rum urea-N, and creatinine, and reduced the severity of renal damage in the STZ-treated group (i.e., the diabetes- induced group). The expression of HSP60 and HSP70 was lower in the STZ group that received CrHis than in the group that did not. No significant effect of CrHis supplementation was detected in regard to the overall measured parameters in the control group. Conclusions: Chromium histidinate significantly decreased lipid peroxidation levels and HSP expression in the kidneys of experimentally induced diabetic rats. This study supported the efficacy of CrHis in reducing renal risk fac- tors and impairment because of diabetes. Ó 2010 by the National Kidney Foundation, Inc. All rights reserved. D IET PLAYS AN IMPORTANT ROLE in the development and prevention of cancer, cardiovascular disease, and diabetes. A high-fat diet can induce obesity and metabolic disorders, insulin resistance, dyslipidemia, and hypertension in rodents and humans. 1,2 High-fat diets might in- duce changes not only in energy metabolism but also in liver, kidney, and brain function. Diabetes is characterized by high levels of blood glucose re- sulting from defects in insulin production or func- tion. The metabolism of carbohydrates, lipids, and proteins is affected under diabetic conditions. 3 This metabolic change can lead to damage of sev- eral different tissues, and particularly those that are insulin-insensitive, such as the kidney. 4 Several *Department of Nephrology, School of Medicine, Firat University, Elazig, Turkey. †Department of Biology, Faculty of Science, Firat University, Elazig, Turkey. ‡Technical Services and Scientific Affairs, Nutrition 21, Inc., Purchase, New York. §Department of Biochemistry, Sarahatun Hospital, Elazig, Turkey. {Department of Pathology, School of Medicine, Firat University, Elazig, Turkey. # Department of Animal Nutrition, Faculty of Veterinary Science, FiratUniversity, Elazig, Turkey. Address reprint requests to Kazim Sahin, DVM, PhD, Depart- ment of Animal Nutrition, Faculty of Veterinary Science, Firat Uni- versity, 23119 Elazig, Turkey. E-mail: [email protected] Ó 2010 by the National Kidney Foundation, Inc. All rights reserved. 1051-2276/10/2002-0008$36.00/0 doi:10.1053/j.jrn.2009.04.009 112 Journal of Renal Nutrition, Vol 20, No 2 (March), 2010: pp 112–120
-
Upload
omniactives -
Category
Documents
-
view
4 -
download
0
Transcript of Effects of Chromium Histidinate on Renal Function, Oxidative Stress, and Heat-Shock Proteins in...

ORIGINAL RESEARCH
Effects of Chromium Histidinateon Renal Function, Oxidative Stress,and Heat-Shock Proteins in Fat-Fedand Streptozotocin-Treated RatsAyhan Dogukan, MD,* Mehmet Tuzcu, PhD,† Vijaya Juturu, PhD,‡
Gurkan Cikim, MD,§ _Ibrahim Ozercan, MD,{ James Komorowski, MSc,‡
and Kazim Sahin, DVM, PhD#
Objective: Chromium is an essential element for carbohydrate, fat, and protein metabolism. The therapeutic po-
tential of chromium histidinate (CrHis) in the treatment of diabetes has been elucidated. The present study investi-
gated the effects of CrHis on serum parameters of renal function, on oxidative stress markers (malondialdehyde
[MDA] and 8-isoprostane), and on the expression of heat-shock proteins (HSPs) in rats.
Methods: Male Wistar rats (n 5 60, 8 weeks old) were divided into four groups. Group 1 received a standard diet
(12% of calories as fat). Group 2 received a standard diet, plus CrHis. Group 3 received a high-fat diet (40% of cal-
ories as fat) for 2 weeks, and was then injected with streptozotocin (STZ) on day 14 (STZ, 40 mg/kg intraperitoneally).
Group 4 was treated in the same way as group 3 (HFD/STZ), but was supplemented with 110 mg CrHis/kg/body
weight/day. Oxidative stress in the kidneys of diabetic rats was evidenced by an elevation in levels of MDA and 8-
isoprostane. Protein concentrations of HSP60 and HSP70 in renal tissue were determined by Western blot analyses.
Results: Chromium histidinate supplementation lowered kidney concentrations of MDA, 8-isoprostane levels, se-
rum urea-N, and creatinine, and reduced the severity of renal damage in the STZ-treated group (i.e., the diabetes-
induced group). The expression of HSP60 and HSP70 was lower in the STZ group that received CrHis than in the
group that did not. No significant effect of CrHis supplementation was detected in regard to the overall measured
parameters in the control group.
Conclusions: Chromium histidinate significantly decreased lipid peroxidation levels and HSP expression in the
kidneys of experimentally induced diabetic rats. This study supported the efficacy of CrHis in reducing renal risk fac-
tors and impairment because of diabetes.
� 2010 by the National Kidney Foundation, Inc. All rights reserved.
*Department of Nephrology, School of Medicine, Firat University,
Elazig, Turkey.
†Department of Biology, Faculty of Science, Firat University,
Elazig, Turkey.
‡Technical Services and Scientific Affairs, Nutrition 21, Inc.,
Purchase, New York.
§Department of Biochemistry, Sarahatun Hospital, Elazig,
Turkey.
{Department of Pathology, School of Medicine, Firat University,
Elazig, Turkey.#Department of Animal Nutrition, Faculty of Veterinary Science,
Firat University, Elazig, Turkey.
Address reprint requests to Kazim Sahin, DVM, PhD, Depart-
ment of Animal Nutrition, Faculty of Veterinary Science, Firat Uni-
versity, 23119 Elazig, Turkey. E-mail: [email protected]� 2010 by the National Kidney Foundation, Inc. All rights
reserved.
1051-2276/10/2002-0008$36.00/0
doi:10.1053/j.jrn.2009.04.009
112 Jour
DIET PLAYS AN IMPORTANT ROLE inthe development and prevention of cancer,
cardiovascular disease, and diabetes. A high-fatdiet can induce obesity and metabolic disorders,insulin resistance, dyslipidemia, and hypertensionin rodents and humans.1,2 High-fat diets might in-duce changes not only in energy metabolism butalso in liver, kidney, and brain function. Diabetesis characterized by high levels of blood glucose re-sulting from defects in insulin production or func-tion. The metabolism of carbohydrates, lipids, andproteins is affected under diabetic conditions.3
This metabolic change can lead to damage of sev-eral different tissues, and particularly those that areinsulin-insensitive, such as the kidney.4 Several
nal of Renal Nutrition, Vol 20, No 2 (March), 2010: pp 112–120

CHROMIUM HISTIDINATE AND RENAL FUNCTION 113
mechanisms were proposed for the development ofrenal injury in diabetes.5–9 High-fat diets can alsocause renal lipid accumulation and renal injury.10
There is an interrelationship between chronic dis-ease and various micronutrients such as chromium(Cr), selenium, and zinc.11–14 A significant alter-ation of these elements in diabetic individualsand animals was attributed to insulin defi-ciency.11–13 Patients with diabetes showed lowerCr, selenium, and zinc levels in their whole bloodor plasma than were found in normal subjects.11 Arelationship between renal Cr deficiency and glu-cose intolerance was reported.15 It was also re-ported that the absorption and excretion of Crare higher in diabetic subjects than in nondiabeticsubjects.16,17
Diabetes may be controlled by the use of severalpharmacological agents. Chromium deficiencyleads to impaired glucose tolerance because of insu-lin resistance, wherein insulin is produced in cor-rect amounts but is not taken up properly.Chromium deficiency can also contribute tohyperglycemia.4,18 Therefore, Cr supplementsmight exert beneficial effects in the managementof type 2 diabetes, as characterized by a decline ininsulin response.4 Further, it was reported thata low plasma concentration of histidine is associatedwith inflammation, oxidative stress, the prevalenceof protein-energy wasting, and mortality inpatients with chronic kidney disease.19 Chromiumis required for the normal metabolism of carbohy-drates and fats. This essential micronutrient ele-mental Cr is also required for optimal insulinactivity. Human studies suggest that Cr supplemen-tation decreases insulin levels and improves glucosedisposal in obese populations.20 Chromium histidi-nate is a stable compound with a better absorptionthan other forms of Cr compounds tested in humansubjects.21 Anderson et al. observed that men andwomen absorbed an average 3.1 mg of Cr fromthe chromium-histidine complex, compared with1.8 mg from chromium picolinate, 0.4 mg fromchromium chloride, and 0.2 mg from chromiumpolynicotinate.22 Inadequate amounts of Cr mayresult in improper functioning of the metabolicprocess, and lead to a number of physiologicaldisorders that increase the risk for diabetes andcardiovascular disease, including elevated circulat-ing insulin, glucose, triglycerides, total cholesterol,reduced high-density lipoprotein cholesterol, andimpaired immune function.23,24 Chromium waspostulated to exert antiobesity effects. These
antiobesity effects include an increase in leanbody mass, a decrease in percent body fat, and anincrease in basal metabolic rate.24 Although nu-merous studies of humans and animals examinedhealth-related aspects of chromium picolinate,few studies investigated the beneficial effects ofchromium histidinate (CrHis) in vivo. This studyinvestigated the effects of a high-fat diet on renaldamage and oxidative stress, to determine ifthis damage could be decreased or repaired byCrHis.
Methods
Male Wistar rats (n 5 60, 8 weeks old), weigh-ing between 200 and 250 g, were purchased fromthe Veterinary Control and Research Institute(Elazig, Turkey). These animals were reared ata temperature of 22 6 2�C, with humidity at55% 6 5%, and a 12/12-hour light/dark cycle.Ourexperimentswere conducted under a protocolapproved by Firat University. All procedures in-volving rats were conducted in strict compliancewith relevant laws, the Animal Welfare Act, PublicHealth Services Policy, and guidelines establishedby the Institutional Animal Care and Use Com-mittee of our university. Rats consumed a standarddiet and tap water ad libitum. A blood sample wascollected from the tail vein of each rat for the mea-surement of biochemical efficacy markers. Ratswere randomly assigned to treatment groups.Rats were fed a standard diet (12% of calories asfat; control diet) or a high-fat diet (HFD; 40% ofcalories as fat) for 12 weeks, and received CrHis(Nutrition 21, Purchase, NY).
Materials
The ingredients and chemical composition ofthe basal diet are listed in Table 1. The dietswere stored at 4�C in a cold chamber. Animalswere fed either a normal diet consisting (as percenttotal kcal) of 12% fat and 26% protein, or theHFD consisting of 40% fat and 18% protein. TheCrHis was supplied by Nutrition 21, Inc. TheCrHis was dissolved in water and administered ata concentration of 110 mg/kg/day ad libitum indrinking water containing 5.53 mg Cr/L for 10weeks, to produce 8 mg Cr/day, which is equiva-lent to a dose of 560 mg Cr for a 70-kg humanadult.
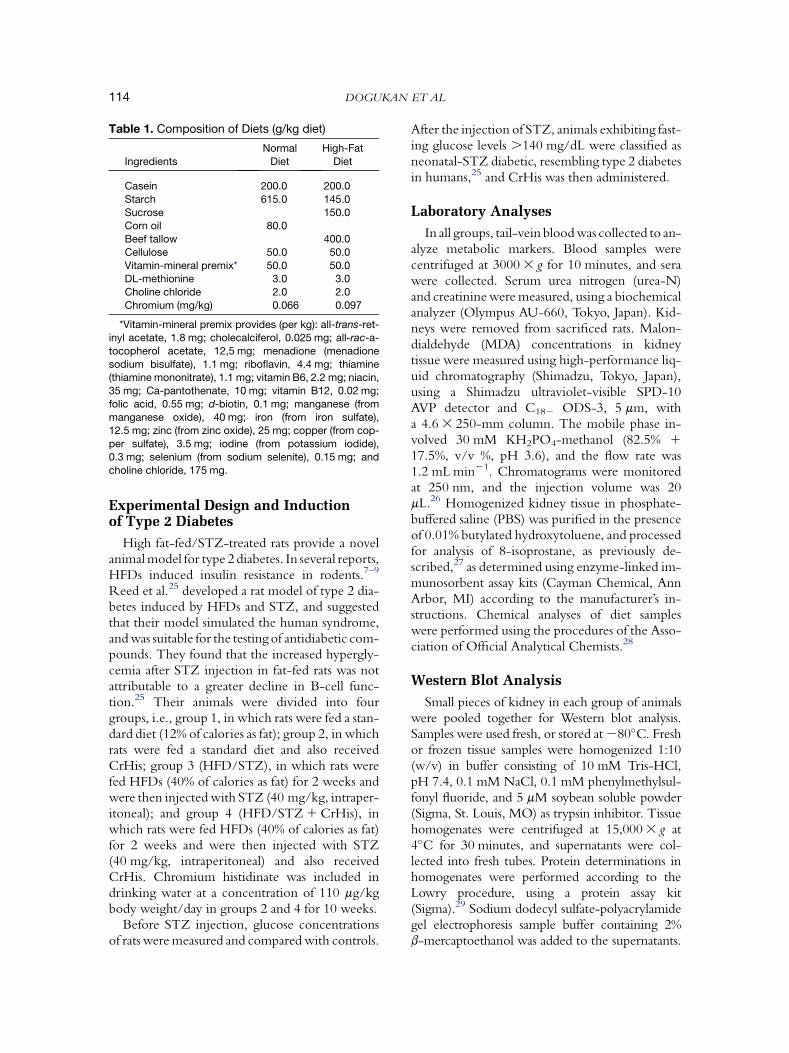
Table 1. Composition of Diets (g/kg diet)
Ingredients
Normal
Diet
High-Fat
Diet
Casein 200.0 200.0
Starch 615.0 145.0
Sucrose 150.0Corn oil 80.0
Beef tallow 400.0
Cellulose 50.0 50.0
Vitamin-mineral premix* 50.0 50.0DL-methionine 3.0 3.0
Choline chloride 2.0 2.0
Chromium (mg/kg) 0.066 0.097
*Vitamin-mineral premix provides (per kg): all-trans-ret-inyl acetate, 1.8 mg; cholecalciferol, 0.025 mg; all-rac-a-
tocopherol acetate, 12,5 mg; menadione (menadione
sodium bisulfate), 1.1 mg; riboflavin, 4.4 mg; thiamine(thiamine mononitrate), 1.1 mg; vitamin B6, 2.2 mg; niacin,
35 mg; Ca-pantothenate, 10 mg; vitamin B12, 0.02 mg;
folic acid, 0.55 mg; d-biotin, 0.1 mg; manganese (from
manganese oxide), 40 mg; iron (from iron sulfate),12.5 mg; zinc (from zinc oxide), 25 mg; copper (from cop-
per sulfate), 3.5 mg; iodine (from potassium iodide),
0.3 mg; selenium (from sodium selenite), 0.15 mg; and
choline chloride, 175 mg.
DOGUKAN ET AL114
Experimental Design and Inductionof Type 2 Diabetes
High fat-fed/STZ-treated rats provide a novelanimal model for type 2 diabetes. In several reports,HFDs induced insulin resistance in rodents.7–9
Reed et al.25 developed a rat model of type 2 dia-betes induced by HFDs and STZ, and suggestedthat their model simulated the human syndrome,and was suitable for the testing of antidiabetic com-pounds. They found that the increased hypergly-cemia after STZ injection in fat-fed rats was notattributable to a greater decline in B-cell func-tion.25 Their animals were divided into fourgroups, i.e., group 1, in which rats were fed a stan-dard diet (12% of calories as fat); group 2, in whichrats were fed a standard diet and also receivedCrHis; group 3 (HFD/STZ), in which rats werefed HFDs (40% of calories as fat) for 2 weeks andwere then injected with STZ (40 mg/kg, intraper-itoneal); and group 4 (HFD/STZ 1 CrHis), inwhich rats were fed HFDs (40% of calories as fat)for 2 weeks and were then injected with STZ(40 mg/kg, intraperitoneal) and also receivedCrHis. Chromium histidinate was included indrinking water at a concentration of 110 mg/kgbody weight/day in groups 2 and 4 for 10 weeks.
Before STZ injection, glucose concentrationsof rats were measured and compared with controls.
After the injection of STZ, animals exhibiting fast-ing glucose levels .140 mg/dL were classified asneonatal-STZ diabetic, resembling type 2 diabetesin humans,25 and CrHis was then administered.
Laboratory Analyses
In all groups, tail-vein blood was collected to an-alyze metabolic markers. Blood samples werecentrifuged at 3000 3 g for 10 minutes, and serawere collected. Serum urea nitrogen (urea-N)and creatinine were measured, using a biochemicalanalyzer (Olympus AU-660, Tokyo, Japan). Kid-neys were removed from sacrificed rats. Malon-dialdehyde (MDA) concentrations in kidneytissue were measured using high-performance liq-uid chromatography (Shimadzu, Tokyo, Japan),using a Shimadzu ultraviolet-visible SPD-10AVP detector and C182 ODS-3, 5 mm, witha 4.6 3 250-mm column. The mobile phase in-volved 30 mM KH2PO4-methanol (82.5% 1
17.5%, v/v %, pH 3.6), and the flow rate was1.2 mL min21. Chromatograms were monitoredat 250 nm, and the injection volume was 20mL.26 Homogenized kidney tissue in phosphate-buffered saline (PBS) was purified in the presenceof 0.01% butylated hydroxytoluene, and processedfor analysis of 8-isoprostane, as previously de-scribed,27 as determined using enzyme-linked im-munosorbent assay kits (Cayman Chemical, AnnArbor, MI) according to the manufacturer’s in-structions. Chemical analyses of diet sampleswere performed using the procedures of the Asso-ciation of Official Analytical Chemists.28
Western Blot Analysis
Small pieces of kidney in each group of animalswere pooled together for Western blot analysis.Samples were used fresh, or stored at 280�C. Freshor frozen tissue samples were homogenized 1:10(w/v) in buffer consisting of 10 mM Tris-HCl,pH 7.4, 0.1 mM NaCl, 0.1 mM phenylmethylsul-fonyl fluoride, and 5 mM soybean soluble powder(Sigma, St. Louis, MO) as trypsin inhibitor. Tissuehomogenates were centrifuged at 15,000 3 g at4�C for 30 minutes, and supernatants were col-lected into fresh tubes. Protein determinations inhomogenates were performed according to theLowry procedure, using a protein assay kit(Sigma).29 Sodium dodecyl sulfate-polyacrylamidegel electrophoresis sample buffer containing 2%b-mercaptoethanol was added to the supernatants.

CHROMIUM HISTIDINATE AND RENAL FUNCTION 115
Equal amounts of total protein (10 mg/sample) wereapplied in each lane, as described previously.30 Sep-arated proteins were transferred to nitrocellulosemembranes (Schleicher and Schuell, Inc., Keene,NH). Nitrocellulose blots were washed twice for5 minutes each in PBS, and blocked with 1% bovineserum albumin in PBS for 1 hour before applicationof the primary antibody. Primary antibody (anti-heat-shock protein [HSP]60 and anti-HSP70monoclonal antibody, Santa Cruz Biotechnology,Inc., Santa Cruz, CA) was diluted (1:1000) in thesame buffer containing 0.05% Tween-20. Thenitrocellulose membrane was incubated overnightat 4�C with protein antibody. The blots werewashed and incubated for 1 hour with a secondaryantibody, either an anti-mouse or anti-rat immuno-globulin G peroxidase conjugated (Sigma, Dorset,United Kingdom). Specific binding was detectedusing diaminobenzidine and H2O2 as substrates.31
Protein loading was controlled using a monoclonalmouse antibody against b-actin antibody (A5316;Sigma). Blots were performed at least three timesto confirm data reproducibility.32,33 Protein levelswere analyzed densitometrically, using an image-analysis system (Image J, National Institutes ofHealth, Bethesda, MD).
Histopathology
The kidney of each rat was examined grossly.Tissues were removed for histological study,washed with normal saline, and immersion-fixedin 10% buffered formalin immediately. Theywere gradually dehydrated, embedded in paraffin,cut into 5-mM sections, and stained with hema-toxylin and eosin for histological examination,according to standard procedures.34 Vacuolardegeneration, tubular atrophy and dilatation, in-terstitial inflammation, interstitial fibrosis, and glo-merular hypercellularity were semiquantitativelyevaluated for histopathological changes by a pa-thologist unaware of the type of treatment. A min-imum of 10 fields for each kidney slide wasexamined and scored for severity of changes, usingthe following scale: 2, none; 1, mild damage;11, moderate damage; and 111, severedamage.
Statistical Analysis
Data are presented as means (6 SEM). Samplesize was calculated based on a power of 85% anda P value of .05. A sample size of 10 per treatment
was calculated. Data were analyzed using the gen-eral linear model (GLM) procedure of SAS software(SAS, Inc., Cary, NC).35 Least-squares treatmentswere compared if a significant F statistic (5% levelof P) was detected by analysis of variance.Treatments were also compared using Student’sunpaired t-test for comparisons of individual treat-ments. P , .05 was considered statistically signifi-cant. Between-group differences in latencies wereanalyzed by analysis of variance for repeated mea-surements, followed by Fisher’s post hoc test forall groups.
Results
Table 2 shows the effects of CrHis on serum pa-rameters of renal function. The serum urea-N andcreatinine levels increased significantly in the STZand HFD groups. After oral administration ofCrHis, serum urea-N and creatinine levels werenot altered in rats fed a regular diet, but decreasedsignificantly in diabetic rats. In STZ-treated rats,a significant increase in levels of MDA and 8-iso-prostane was evident in kidney tissue when com-pared with control rats, as indicated in Table 3.A significant reduction in the level of MDA wasevident in CrHis-treated rats when comparedwith STZ-treated rats. Protein concentrations ofHSP60 and HSP70 in renal tissue were deter-mined by Western blot analyses. The results arepresented in Figures 1 and 2. Based on band den-sities, renal HSP60 and HSP70 were significantlyelevated in diabetic rats compared with controlrats. However, the extent of protein expression de-clined in rats treated with CrHis. No significantdifference in level of HSP was found in group 1versus group 2.
In groups 1 and 2, normal renal tubular archi-tecture and normal glomeruli were observed(Fig. 3A,B). In HFD/STZ rats, there was a signif-icant vacuolar degeneration of renal tubular cells,with displaced pyknotic nuclei. Renal glomeruliwith mesangial hyperplasia were evident. A diffuseinfiltration of monomorphonuclear cells, tubu-lointerstitial fibrosis, and tubular atrophy was ob-served (Fig. 3C). In HFD/STZ 1 CrHis-treatedrats, mild tubular atrophy with no major glomeru-lar pathology was observed (Fig. 3D and Table 4).
Discussion
Diabetes is a metabolic disorder associated withthe impairment of glucose utilization, caused by
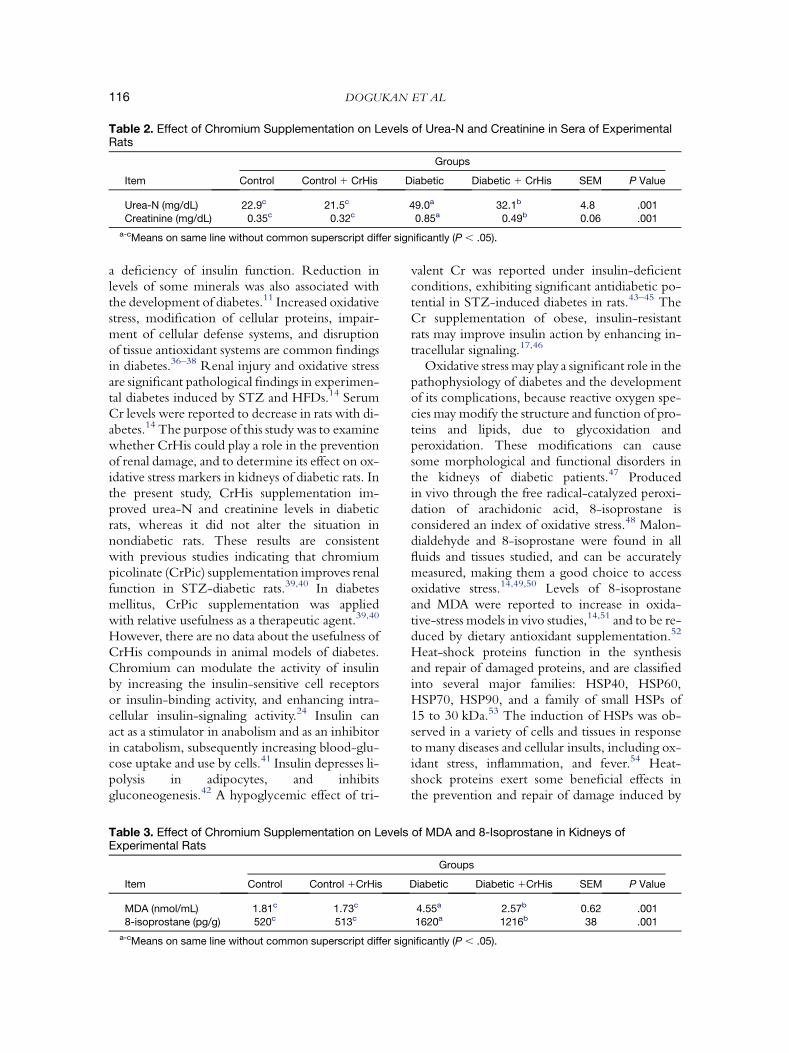
Table 2. Effect of Chromium Supplementation on Levels of Urea-N and Creatinine in Sera of ExperimentalRats
Groups
Item Control Control 1 CrHis Diabetic Diabetic 1 CrHis SEM P Value
Urea-N (mg/dL) 22.9c 21.5c 49.0a 32.1b 4.8 .001
Creatinine (mg/dL) 0.35c 0.32c 0.85a 0.49b 0.06 .001
a-cMeans on same line without common superscript differ significantly (P , .05).
DOGUKAN ET AL116
a deficiency of insulin function. Reduction inlevels of some minerals was also associated withthe development of diabetes.11 Increased oxidativestress, modification of cellular proteins, impair-ment of cellular defense systems, and disruptionof tissue antioxidant systems are common findingsin diabetes.36–38 Renal injury and oxidative stressare significant pathological findings in experimen-tal diabetes induced by STZ and HFDs.14 SerumCr levels were reported to decrease in rats with di-abetes.14 The purpose of this study was to examinewhether CrHis could play a role in the preventionof renal damage, and to determine its effect on ox-idative stress markers in kidneys of diabetic rats. Inthe present study, CrHis supplementation im-proved urea-N and creatinine levels in diabeticrats, whereas it did not alter the situation innondiabetic rats. These results are consistentwith previous studies indicating that chromiumpicolinate (CrPic) supplementation improves renalfunction in STZ-diabetic rats.39,40 In diabetesmellitus, CrPic supplementation was appliedwith relative usefulness as a therapeutic agent.39,40
However, there are no data about the usefulness ofCrHis compounds in animal models of diabetes.Chromium can modulate the activity of insulinby increasing the insulin-sensitive cell receptorsor insulin-binding activity, and enhancing intra-cellular insulin-signaling activity.24 Insulin canact as a stimulator in anabolism and as an inhibitorin catabolism, subsequently increasing blood-glu-cose uptake and use by cells.41 Insulin depresses li-polysis in adipocytes, and inhibitsgluconeogenesis.42 A hypoglycemic effect of tri-
Table 3. Effect of Chromium Supplementation on LevelsExperimental Rats
Item Control Control 1CrHis
MDA (nmol/mL) 1.81c 1.73c
8-isoprostane (pg/g) 520c 513c
a-cMeans on same line without common superscript differ sig
valent Cr was reported under insulin-deficientconditions, exhibiting significant antidiabetic po-tential in STZ-induced diabetes in rats.43–45 TheCr supplementation of obese, insulin-resistantrats may improve insulin action by enhancing in-tracellular signaling.17,46
Oxidative stress may play a significant role in thepathophysiology of diabetes and the developmentof its complications, because reactive oxygen spe-cies may modify the structure and function of pro-teins and lipids, due to glycoxidation andperoxidation. These modifications can causesome morphological and functional disorders inthe kidneys of diabetic patients.47 Producedin vivo through the free radical-catalyzed peroxi-dation of arachidonic acid, 8-isoprostane isconsidered an index of oxidative stress.48 Malon-dialdehyde and 8-isoprostane were found in allfluids and tissues studied, and can be accuratelymeasured, making them a good choice to accessoxidative stress.14,49,50 Levels of 8-isoprostaneand MDA were reported to increase in oxida-tive-stress models in vivo studies,14,51 and to be re-duced by dietary antioxidant supplementation.52
Heat-shock proteins function in the synthesisand repair of damaged proteins, and are classifiedinto several major families: HSP40, HSP60,HSP70, HSP90, and a family of small HSPs of15 to 30 kDa.53 The induction of HSPs was ob-served in a variety of cells and tissues in responseto many diseases and cellular insults, including ox-idant stress, inflammation, and fever.54 Heat-shock proteins exert some beneficial effects inthe prevention and repair of damage induced by
of MDA and 8-Isoprostane in Kidneys of
Groups
Diabetic Diabetic 1CrHis SEM P Value
4.55a 2.57b 0.62 .001
1620a 1216b 38 .001
nificantly (P , .05).
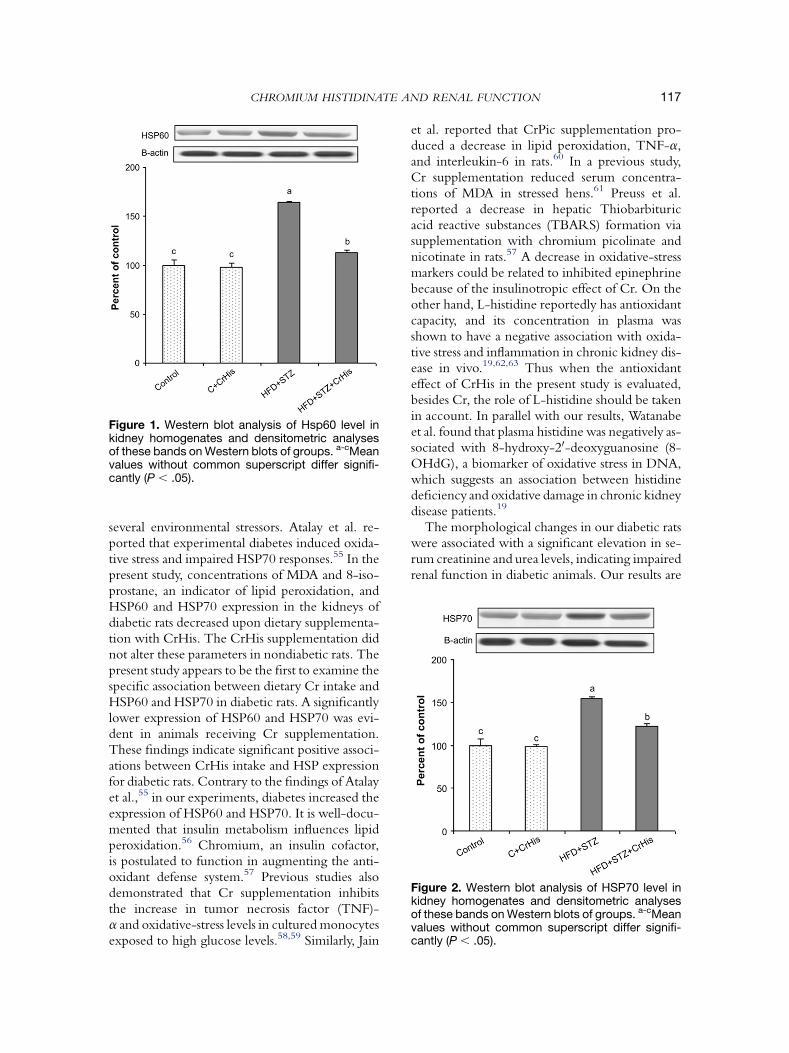
Figure 1. Western blot analysis of Hsp60 level inkidney homogenates and densitometric analysesof these bands on Western blots of groups. a-cMeanvalues without common superscript differ signifi-cantly (P , .05).
Figure 2. Western blot analysis of HSP70 level inkidney homogenates and densitometric analysesof these bands on Western blots of groups. a-cMeanvalues without common superscript differ signifi-cantly (P , .05).
CHROMIUM HISTIDINATE AND RENAL FUNCTION 117
several environmental stressors. Atalay et al. re-ported that experimental diabetes induced oxida-tive stress and impaired HSP70 responses.55 In thepresent study, concentrations of MDA and 8-iso-prostane, an indicator of lipid peroxidation, andHSP60 and HSP70 expression in the kidneys ofdiabetic rats decreased upon dietary supplementa-tion with CrHis. The CrHis supplementation didnot alter these parameters in nondiabetic rats. Thepresent study appears to be the first to examine thespecific association between dietary Cr intake andHSP60 and HSP70 in diabetic rats. A significantlylower expression of HSP60 and HSP70 was evi-dent in animals receiving Cr supplementation.These findings indicate significant positive associ-ations between CrHis intake and HSP expressionfor diabetic rats. Contrary to the findings of Atalayet al.,55 in our experiments, diabetes increased theexpression of HSP60 and HSP70. It is well-docu-mented that insulin metabolism influences lipidperoxidation.56 Chromium, an insulin cofactor,is postulated to function in augmenting the anti-oxidant defense system.57 Previous studies alsodemonstrated that Cr supplementation inhibitsthe increase in tumor necrosis factor (TNF)-a and oxidative-stress levels in cultured monocytesexposed to high glucose levels.58,59 Similarly, Jain
et al. reported that CrPic supplementation pro-duced a decrease in lipid peroxidation, TNF-a,and interleukin-6 in rats.60 In a previous study,Cr supplementation reduced serum concentra-tions of MDA in stressed hens.61 Preuss et al.reported a decrease in hepatic Thiobarbituricacid reactive substances (TBARS) formation viasupplementation with chromium picolinate andnicotinate in rats.57 A decrease in oxidative-stressmarkers could be related to inhibited epinephrinebecause of the insulinotropic effect of Cr. On theother hand, L-histidine reportedly has antioxidantcapacity, and its concentration in plasma wasshown to have a negative association with oxida-tive stress and inflammation in chronic kidney dis-ease in vivo.19,62,63 Thus when the antioxidanteffect of CrHis in the present study is evaluated,besides Cr, the role of L-histidine should be takenin account. In parallel with our results, Watanabeet al. found that plasma histidine was negatively as-sociated with 8-hydroxy-20-deoxyguanosine (8-OHdG), a biomarker of oxidative stress in DNA,which suggests an association between histidinedeficiency and oxidative damage in chronic kidneydisease patients.19
The morphological changes in our diabetic ratswere associated with a significant elevation in se-rum creatinine and urea levels, indicating impairedrenal function in diabetic animals. Our results are
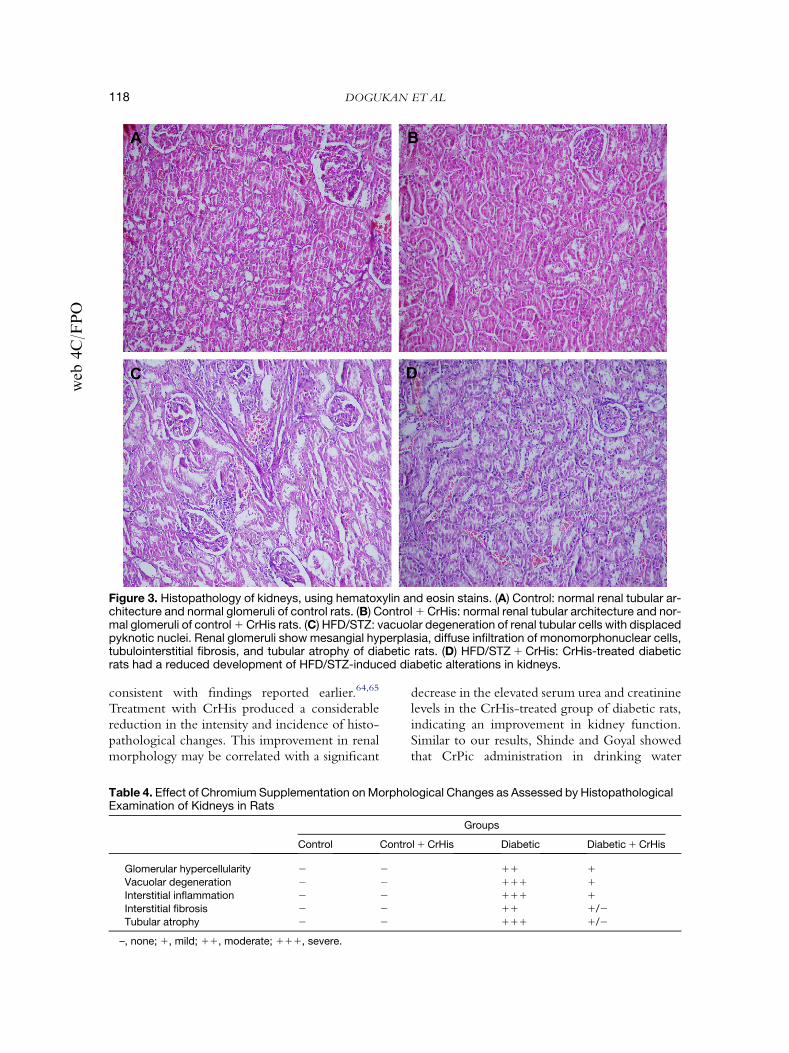
Figure 3. Histopathology of kidneys, using hematoxylin and eosin stains. (A) Control: normal renal tubular ar-chitecture and normal glomeruli of control rats. (B) Control 1 CrHis: normal renal tubular architecture and nor-mal glomeruli of control 1 CrHis rats. (C) HFD/STZ: vacuolar degeneration of renal tubular cells with displacedpyknotic nuclei. Renal glomeruli show mesangial hyperplasia, diffuse infiltration of monomorphonuclear cells,tubulointerstitial fibrosis, and tubular atrophy of diabetic rats. (D) HFD/STZ 1 CrHis: CrHis-treated diabeticrats had a reduced development of HFD/STZ-induced diabetic alterations in kidneys.
web
4C/FPO
DOGUKAN ET AL118
consistent with findings reported earlier.64,65
Treatment with CrHis produced a considerablereduction in the intensity and incidence of histo-pathological changes. This improvement in renalmorphology may be correlated with a significant
Table 4. Effect of Chromium Supplementation on MorphoExamination of Kidneys in Rats
Control Contro
Glomerular hypercellularity 2 2
Vacuolar degeneration 2 2
Interstitial inflammation 2 2
Interstitial fibrosis 2 2
Tubular atrophy 2 2
–, none; 1, mild; 11, moderate; 111, severe.
decrease in the elevated serum urea and creatininelevels in the CrHis-treated group of diabetic rats,indicating an improvement in kidney function.Similar to our results, Shinde and Goyal showedthat CrPic administration in drinking water
logical Changes as Assessed by Histopathological
Groups
l 1 CrHis Diabetic Diabetic 1 CrHis
11 1
111 1
111 1
11 1/2
111 1/2

CHROMIUM HISTIDINATE AND RENAL FUNCTION 119
(8 mg/mL) for 6 weeks decreased vacuolar degen-eration, cellular infiltration, and hypertrophy ofkidneys in STZ-diabetic rats, whereas CrPic didnot alter normal function or morphology incontrol rats.39 Mita et al. reported that supplemen-tation with CrPic recovered renal Cr concentra-tions and improved renal function in type 2diabetic mice.40 No toxic effects were evidentfor in vivo models at different doses,23,66 or in di-abetic models in vivo.39,67
In conclusion, CrHis-treated rats (our group 4)had decreased lipid-peroxidation levels and HSPexpression. These results suggest that CrHis mayprevent the renal impairment associated with dia-betes. Further studies are required to explore theefficacy of CrHis in preventing or reverting thekidney disorders associated with diabetes.
References
1. Barnard RJ, Roberts CK, Varon SM, Berger JJ: Diet-in-
duced insulin resistance precedes other aspects of the metabolic
syndrome. J Appl Physiol 84:1311-1315, 1998
2. Buettner R, Scholmerich J, Bollheimer LC: High-fat diet:
modeling the metabolic disorders of human obesity in rodents.
Obesity 15:798-808, 2007
3. Broadhurst CL, Domenico P: Clinical studies on chromium
picolinate supplementation in diabetes mellitus—a review. Diabe-
tes Technol Ther 8:677-687, 2006
4. Bartlett HE, Eperjesi F: Nutritional supplementation for
type 2 diabetes: a systematic review. Ophthalmic Physiol Opt
28:503-523, 2008
5. Praga M: Obesity: a neglected culprit in renal disease. Neph-
rol Dial Transplant 17:1157-1159, 2002
6. Abrass CK: Overview: obesity—what does it have to do
with kidney disease? J Am Soc Nephrol 15:2768-2772, 2004
7. Bagby SP: Obesity-initiated metabolic syndrome and the
kidney: a recipe for chronic kidney disease? J Am Soc Nephrol
15:2775-2791, 2004
8. El-Atat FA, Stas SN, McFarlane SI, et al: The relationship
between hyperinsulinemia, hypertension and progressive renal
disease. J Am Soc Nephrol 15:2816-2827, 2004
9. Kume S, Uzu T, Araki S, Sugimoto T, et al: Role of altered
renal lipid metabolism in the development of renal injury induced
by a high-fat diet. J Am Soc Nephrol 18:2715-2723, 2007
10. Jiang T, Wang Z, Proctor G, et al: Diet-induced obesity in
C57BL/6J mice causes increased renal lipid accumulation and glo-
merulosclerosis via a sterol regulatory element-binding protein-
1c-dependent pathway. J Biol Chem 280:32317-32325, 2005
11. Wallach S: Clinical and biochemical aspects of chromium
deficiency. J Am Coll Nutr 4:107-120, 1985
12. Becker DJ, Reul B, Ozeelikay A, et al: Oral selenate im-
proves glucose homeostasis and partly reverses abnormal expres-
sion of liver glycolytic and gluconeagenic enzymes in diabetic
rats. Diabetologia 39:3-11, 1996
13. Faure P, Roussel A, Coudray C, Richard MJ, Halimi S,
Favier A: Zinc and insulin sensitivity. Biol Trace Elem Res 32:
305-310, 1992
14. Sahin K, Onderci M, Tuzcu M, et al: Effect of chromium
on carbohydrate and lipid metabolism in a rat model of type 2 di-
abetes mellitus: the fat-fed, streptozotocin-treated rat. Metabolism
56:1233-1240, 2007
15. Feng W, Ding W, Qian Q, et al: Comparison of the chro-
mium distribution in organs and subcellular fractions of normal
and diabetic rats by using enriched stable isotope Cr-50 tracer
technique. Biol Trace Elem Res 71:121-129, 1999
16. Ding W, Chai Z, Duan P, et al: Serum and urine chromium
concentrations in elderly diabetics. Biol Trace Elem Res 63:
231-237, 1998
17. Cefalu WT, Wang ZQ, Zhang XH, et al: Oral chromium
picolinate improves carbohydrate and lipid metabolism and en-
hances skeletal muscle Glut-4 translocation in obese, hyperinsuli-
nemic (JCR-LA corpulent) rats. J Nutr 132:1107-1114, 2002
18. Cefalu WT, Hu FB: Role of chromium in human health
and in diabetes. Diabetes Care 27:2741-2751, 2004
19. Watanabe M, Suliman ME, Qureshi AR, et al: Conse-
quences of low plasma histidine in chronic kidney disease patients:
associations with inflammation, oxidative stress, and mortality. Am
J Clin Nutr 87:1860-1866, 2008
20. Cefalu WT, Bell-Farrow AD, Stegner J, et al: Effect of
chromium picolinate on insulin sensitivity in vivo. J Trace Elem
Exp Med 12:71-83, 1999
21. Anderson RA, Polansky MM, Bryden NA: Chromium-
histidine complexes as nutrient supplements. US patent issued,
February 10, 2004
22. Anderson RA, Polansky MM, Bryden NA: Stability and
absorption of chromium and absorption of chromium histidinate
complexes by humans. Biol Trace Elem Res 101:211-218, 2004
23. Anderson RA: Nutritional factors influencing the glucose/
insulin system: chromium. J Am Coll Nutr 16:404-410, 1997
24. Anderson RA: Chromium, glucose intolerance and diabe-
tes. J Am Coll Nutr 17:548-555, 1998
25. Reed MJ, Meszaros K, Entes LJ, et al: A new rat model of
type 2 diabetes: the fat-fed, streptozotocin-treated rat. Metabolism
49:1390-1394, 2000
26. Karatepe M: Simultaneous determination of ascorbic acid
and free malondialdehyde in human serum by HPLC/UV. LC-
GC North Am 22:362-365, 2004
27. Wong YT, Ruan R, Tay FEH: Relationship between levels
of oxidative DNA damage, lipid peroxidation and mitochondrial
membrane potential in young and old F344 rats. Free Radic Res
40:393-402, 2006
28. Association of Official Analytical Chemists: Official
Methods of Analysis, 15th Ed. Arglinton, VA: Association of
Official Analytical Chemists, 1990.
29. Lowry OH, Rosebrough NJ, Farr AL, et al: Protein mea-
surement with the Folin phenol reagent. J Biol Chem 193:
265-275, 1951
30. Laemmli UK: Cleavage of structural proteins during the
assembly of the head of bacteriophage T4. Nature 227:680-685,
1970
31. Baydas G, Tuzcu M: Protective effects of melatonin against
ethanol-induced reactive gliosis in hippocampus and cortex of
young and aged rats. Exp Neurol 194:175-181, 2005
32. Kilic E, Kilic U, Soliz J, Bassetti CL, Gassmann M,
Hermann DM: Brain-derived erythropoietin protects from focal
cerebral ischemia by dual activation of ERK-1/-2 and Akt path-
ways. FASEB J 19:2026-2028, 2005
33. Kilic E, Kilic U, Wang Y, Bassetti CL, Marti HH,
Hermann DM: The phosphatidylinositol-3 kinase/Akt pathway

DOGUKAN ET AL120
mediates VEGF’s neuroprotective activity and induces blood brain
barrier permeability after focal cerebral ischemia. FASEB J 20:
1185-1187, 2006
34. Ross MH, Reith EJ, Romrell LJ: Histology—A Text and
Atlas. Baltimore: Williams and Wilkins, 1989
35. SAS Institute: Inc.: SAS User’s Guide: Statistics. Cary, NC:
SAS Institute, Inc., 1999
36. Atalay M, Laaksonen DE, Niskanen L, et al: Altered anti-
oxidant enzyme defenses in insulin-dependent diabetic men
with increased resting and exercise-induced oxidative stress.
Acta Physiol Scand 161:195-201, 1997
37. Ceriello A, Bortolotti N, Falleti E, et al: Total radical trap-
ping antioxidant parameter in NIDDM patients. Diabetes Care
20:194-197, 1997
38. Atalay M, Laaksonen DE: Diabetes, oxidative stress and
physical exercise. J Sports Sci Med 1:1-14, 2002
39. Shinde UA, Goyal RK: Effect of chromium picolinate on
histopathological alterations in STZ and neonatal STZ diabetic
rats. J Cell Mol Med 7:322-329, 2003
40. Mita Y, Ishihara K, Fukuchi Y, et al: Supplementation with
chromium picolinate recovers renal Cr concentration and
improves carbohydrate metabolism and renal function in type 2
diabetic mice. Biol Trace Elem Res 105:229-248, 2005
41. Cupo MA, Donaldson WE: Chromium and vanadium
effects on glucose metabolism and lipid synthesis in the chick.
Poult Sci 66:120-126, 1987
42. Lambert B, Jacquemin C: Inhibition by insulin of the
adrenaline-stimulated adenylate cyclase in rat adipose tissue.
FEBS Lett 105:19-22, 1979
43. Shinde UA, Sharma G, Xu YJ, et al: Anti-diabetic activity
and mechanism of action of chromium chloride. Exp Clin Endo-
crinol Diabetes 112:248-252, 2004
44. Sun Y, Clodfelder BJ, Shute AA, et al: The biomimetic
[Cr(3)O(O(2)CCH(2)CH(3)) (6)(H(2)O)(3)](1) decreases plasma
insulin, cholesterol, and triglycerides in healthy and type II diabetic
rats but not type I diabetic rats. J Biol Inorg Chem 7:852-862, 2002
45. Machalinski B, Walczak M, Syrenicz A, et al: Hypoglyce-
mic potency of novel trivalent chromium in hyperglycemic insu-
lin-deficient rats. J Trace Elem Med Biol 20:33-39, 2006
46. Wang ZQ, Zhang XH, Russell JC, et al: Chromium pico-
linate enhances skeletal muscle cellular insulin signaling in vivo in
obese, insulin-resistant JCR: LA-cp rats. J Nutr 136:415-420,
2006
47. Sundaram RK, Bhaskar A, Vijayalingam S, et al: Antioxi-
dant status and lipid peroxidation in type II diabetes mellitus
with and without complications. Clin Sci 90:255-260, 1996
48. Morrow JD, Minton TA, Badr KF, et al: Evidence that the
F2-isoprostane, 8-epi-prostaglandin F2a, is formed in vivo. Bio-
chim Biophys Acta 1210:244-248, 1994
49. Roberts LJ, Morrow JD: Measurement of F2-isoprostanes
as an index of oxidative stress in vivo. Free Radic Biol Med 28:
505-513, 2000
50. Moore K: Isoprostane and the liver. Chem Phys Lipids 128:
125-133, 2004
51. Basu S: Oxidative injury induced cyclooxygenase activa-
tion in experimental hepatotoxicity. Biochem Biophys Res
Commun 254:764-767, 1999
52. Pratico D, Tangirala RK, Rader DJ, et al: Vitamin E sup-
presses isoprostane generation in vivo and reduces atherosclerosis
in Apo E-deficient mice. Nat Med 4:1189-1192, 1998
53. Powers SK, Locke M, Demirel HA: Exercise, heat shock
proteins, and myocardial protection from I-R injury. Med Sci
Sports Exerc 33:386-392, 2001
54. Burdon RH: Heat shock and the heat shock proteins. Bio-
chem J 240:313-324, 1986
55. Atalay M, Oksala NK, Laaksonen DE, et al: Exercise train-
ing modulates heat shock protein response in diabetic rats. J Appl
Physiol 97:605-611, 2004
56. Gallaher DD, Csallany AS, Shoeman DW, et al: Diabetes
increases excretion of urinary malondehyde conjugates in rats.
Lipids 28:663-666, 1993
57. Preuss HG, Grojec PL, Lieberman S, et al: Effects of differ-
ent chromium compounds on blood pressure and lipid peroxida-
tion in spontaneously hypertensive rats. Clin Nephrol 47:
325-330, 1997
58. Jain SK, Lim G: Chromium chloride inhibits TNF-a and
IL-6 secretion in isolated human blood mononuclear cells exposed
to high glucose. Horm Metab Res 38:60-62, 2006
59. Jain SK, Kannan K: Chromium chloride inhibits oxidative
stress and TNF-a secretion caused by exposure to high glucose in
cultured monocytes. Biochem Biophys Res Commun 289:
687-691, 2001
60. Jain SK, Rains JL, Croad JL: Effect of chromium niacinate
and chromium picolinate supplementation on lipid peroxidation,
TNF-a, IL-6, CRP, glycated hemoglobin, triglycerides and cho-
lesterol levels in blood of streptozotocin-treated diabetic rats. Free
Radic Biol Med 43:1124-1131, 2007
61. Onderci M, Sahin N, Sahin K, et al: The antioxidant prop-
erties of chromium and zinc: in vivo effects on digestibility, lipid
peroxidation, antioxidant vitamins and some minerals under
a low ambient temperature. Biol Trace Elem Res 92:139-150,
2003
62. Kalant N, McCormick S: Inhibition by serum components
of oxidation and collagen-binding of low-density lipoprotein. Bi-
ochim Biophys Acta 1128:211-219, 1992
63. Pisarenko OI: Mechanisms of myocardial protection by
amino acids: facts and hypotheses. Clin Exp Pharmacol Physiol
23:627-633, 1996
64. Jensen PK, Christiansen JS, Steven K, et al: Renal function
in streptozotocin-diabetic rats. Diabetologia 21:409-414, 1981
65. Dai S, Yuen VG, Orvig C, et al: Prevention of diabetes-in-
duced pathology in STZ-diabetic rats by bis(maltolato)oxovana-
dium (IV). Pharmacol Commun 3:311-321, 1993
66. Rhodes MC, Hebert CD, Herbert RA, et al: Absence of
toxic effects in F344/N rats and B6C3F1 mice following sub-
chronic administration of chromium picolinate monohydrate.
Food Chem Toxicol 43:21-29, 2005
67. Mozaffari MS, Patel C, Ballas C, et al: Effects of chronic
chromium picolinate treatment in uninephrectomized rat. Metab-
olism 54:1243-1249, 2005




















