
EFFECTS OF CANAL PLUGGING ON THE VESTIBULO-OCULAR REFLEX AND VESTIBULAR NERVE DISCHARGE DURING...
24
80:3077-3099, 1998. ; J Neurophysiol Sergei B. Yakushin, Theodore Raphan, Jun-Ichi Suzuki, Yasuko Arai and Bernard Cohen Reflex in Monkey: Effects of Canal Plugging Dynamics and Kinematics of the Angular Vestibulo-Ocular You might find this additional info useful... 60 articles, 15 of which you can access for free at: This article cites http://jn.physiology.org/content/80/6/3077.full#ref-list-1 13 other HighWire-hosted articles: This article has been cited by http://jn.physiology.org/content/80/6/3077#cited-by including high resolution figures, can be found at: Updated information and services http://jn.physiology.org/content/80/6/3077.full can be found at: Journal of Neurophysiology about Additional material and information http://www.the-aps.org/publications/jn This information is current as of June 11, 2013. http://www.the-aps.org/. 1998 the American Physiological Society. ISSN: 0022-3077, ESSN: 1522-1598. Visit our website at © a year (monthly) by the American Physiological Society, 9650 Rockville Pike, Bethesda MD 20814-3991. Copyright publishes original articles on the function of the nervous system. It is published 12 times Journal of Neurophysiology by guest on June 11, 2013 http://jn.physiology.org/ Downloaded from
Transcript of EFFECTS OF CANAL PLUGGING ON THE VESTIBULO-OCULAR REFLEX AND VESTIBULAR NERVE DISCHARGE DURING...
80:3077-3099, 1998. ;J Neurophysiol Sergei B. Yakushin, Theodore Raphan, Jun-Ichi Suzuki, Yasuko Arai and Bernard CohenReflex in Monkey: Effects of Canal PluggingDynamics and Kinematics of the Angular Vestibulo-Ocular
You might find this additional info useful...
60 articles, 15 of which you can access for free at: This article citeshttp://jn.physiology.org/content/80/6/3077.full#ref-list-1
13 other HighWire-hosted articles: This article has been cited by http://jn.physiology.org/content/80/6/3077#cited-by
including high resolution figures, can be found at: Updated information and serviceshttp://jn.physiology.org/content/80/6/3077.full
can be found at: Journal of Neurophysiology about Additional material and informationhttp://www.the-aps.org/publications/jn
This information is current as of June 11, 2013.
http://www.the-aps.org/. 1998 the American Physiological Society. ISSN: 0022-3077, ESSN: 1522-1598. Visit our website at
©a year (monthly) by the American Physiological Society, 9650 Rockville Pike, Bethesda MD 20814-3991. Copyright publishes original articles on the function of the nervous system. It is published 12 timesJournal of Neurophysiology
by guest on June 11, 2013http://jn.physiology.org/
Dow
nloaded from

Dynamics and Kinematics of the Angular Vestibulo-Ocular Reflex inMonkey: Effects of Canal Plugging
SERGEI B. YAKUSHIN,1 THEODORE RAPHAN,2 JUN-ICHI SUZUKI,3 YASUKO ARAI,4
AND BERNARD COHEN1
1Departments of Neurology and Physiology and Biophysics, Mount Sinai School of Medicine, New York, New York10029; 2Department of Computer and Information Science, Brooklyn College of the City University of New York,Brooklyn, New York 11210; 3Department of Otolaryngology, Teikyo University and 4Tokyo Women’s Medical College,Tokyo 117-0003, Japan
Yakushin, Sergei B., Theodore Raphan, Jun-Ichi Suzuki, Ya- results in three push-pull pairs, right and left lateral (RLLL),suko Arai, and Bernard Cohen. Dynamics and kinematics of the right anterior and left posterior (RALP), and left anteriorangular vestibulo-ocular reflex in monkey: effects of canal plug- and right posterior (LARP), that code all angular headging. J. Neurophysiol. 80: 3077–3099, 1998. Horizontal and roll movements. The precise angles of the individual semicircu-components of the angular vestibulo-ocular reflex (aVOR) were lar canals have been estimated in several studies, whichelicited by sinusoidal rotation at frequencies from 0.2 Hz (607 /s)
indicate that the canal planes form a nonorthogonal basisto 4.0 Hz (É67 /s) in cynomolgus monkeys. Animals had bothfor sensing head acceleration (Blanks et al. 1985; Curthoyslateral canals plugged (VC, vertical canals intact) , both lateralet al. 1977; Dickman 1996; Reisine et al. 1985, 1988). Thecanals and one pair of the vertical canals plugged (RALP, rightprecise contribution of the canal pairs has been evaluatedanterior and left posterior canals intact; LARP, left anterior and
right posterior canal intact) , or all six semicircular canal plugged using the technique of canal plugging (Ewald 1892; Money(NC, no canals) . In normal animals, horizontal and roll eye veloc- and Scott 1962). After plugging, bone completely obliteratesity was in phase with head velocity and peak horizontal and roll the canal space for several millimeters. It generally is as-gains were É0.8 and 0.6 in upright and 907 pitch, respectively. sumed that there is no flow of endolymph when the canalsNC animals had small aVOR gains at 0.2 Hz, and the temporal are plugged adequately. The implication is that a pluggedphases were shifted É907 toward acceleration. As the frequency
canal can be modeled as having no cupula deflection and noincreased to 4 Hz, aVOR temporal gains and phases tended toafferent modulation when the head is rotated. Using canalnormalize. Findings were similar for the LARP, RALP, and VCplugging, it was shown that the semicircular canal pairsanimals when they were rotated in the planes of the plugged canals.contribute to the aVOR gain according to the geometricThat is, they tended to normalize at higher frequencies. A model
was developed incorporating the geometric organization of the relationship between the canals and stereotaxic head coordi-canals and first order canal-endolymph dynamics. Canal plugging nate frame in which eye movements are measured (Angelakiwas modeled as an alteration in the low frequency 3-db roll-off and Hess 1996; Angelaki et al. 1996; Baker et al. 1987;and corresponding dominant time constant. The shift in the low- Bohmer et al. 1985; Yakushin et al. 1995). From the nonor-frequency 3-dB roll-off was seen in the temporal responses as a thogonal geometry of the canals and from parameters identi-phase lead of the aVOR toward acceleration at higher frequencies.
fied with data from normal monkeys, a model of the aVORThe phase shifted toward stimulus velocity as the frequency in-was constructed that predicted the responses to 0.2-Hz sinu-creased toward 4.0 Hz. By incorporating a dynamic model of thesoidal rotation after canal plugging simply by setting thecanals into the three-dimensional canal system, the spatial re-response from plugged canals to zero (Yakushin et al. 1995).sponses were predicted at all frequencies. Animals were also stimu-
lated with steps of velocity in planes parallel to the plugged lateral This demonstrated the contribution of the individual recipro-canals. This induced a response with a short time constant and low cal canal pairs quantitatively. It also indicated that when apeak velocity in each monkey. Gains were normalized for step specific canal pair is plugged, no adaptation takes place inrotation with respect to time constant as (steady state eye velocity)/ the spatial eye velocity response generated by the remaining(stimulus acceleration 1 time constant) . Using this procedure, the semicircular canals at a midband frequency of 0.2 Hz (Ya-gains were the same in canal plugged as in normal animals and
kushin et al. 1995).corresponded to gains obtained in the frequency analysis. The studyThe conclusion that there was no adaptation after canalsuggests that canal plugging does not block the afferent response
plugging has been challenged. The gains and phases of theto rotation, it merely shifts the dynamic response to higher frequen-cies. aVOR after lateral canal plugging were closer to those of
the normal animal when tested with a higher frequency (1.1Hz) (Angelaki and Hess 1996; Angelaki et al. 1996). This
I N T R O D U C T I O N was explained as an adaptive response or spatial ‘‘recalibra-tion’’ of the central vestibular system. That is, by producingThe semicircular canal system, which comprises threea stronger horizontal response from the vertical canals, thepairs of complementary canals, forms the sensory basis forresponse plane of the vertical canals effectively would bethe angular vestibulo-ocular reflex (aVOR). The two lateralrotated. Such recalibrations in response plane of the intactsemicircular canals can be approximated by a single plane,
as can one anterior and the contralateral posterior canal. This canal system have been observed after ‘‘cross-axis adapta-
30770022-3077/98 $5.00 Copyright q 1998 The American Physiological Society
J1032-7/ 9k2e$$no07 12-09-98 07:00:21 neupa LP-Neurophys
by guest on June 11, 2013http://jn.physiology.org/
Dow
nloaded from

YAKUSHIN, RAPHAN, SUZUKI, ARAI, AND COHEN3078
the animals were upright during testing, the normal to the hori-tion.’’ Schultheis and Robinson (1981) demonstrated thatzontal stereotaxic plane was along the spatial vertical. Eye move-cats could be adapted with continued sinusoidal pitchingments were recorded with scleral search coils. Two coils werewhile an optokinetic surround sinusoidally oscillated in aimplanted on the left eye. One coil was used to measure horizontalhorizontal plane. When animals were later tested in darknessand vertical eye position (Judge et al. 1980). A second coil waswith a pitching stimulus, eye velocity was oblique with a placed approximately orthogonal to the frontal coil to measure roll
horizontal component. The magnitude of the eye velocity eye position (Cohen et al. 1992a; Dai et al. 1994; Yakushin et al.also could be adaptively reduced or increased, depending on 1995). Both coils were sutured to the sclera at the time of surgery.the phase relationship between the adapting vestibular and Postmortem, the coils were embedded firmly in connective tissueoptokinetic stimuli, and cross-axis adaptation was greater that was attached to the sclera. From this, we assume that there
was no movement of the coils relative to the globe during eyewhen the otolith organs were reoriented relative to a spatialmovement.vertical during head rotation. Other types of cross-axis adap-
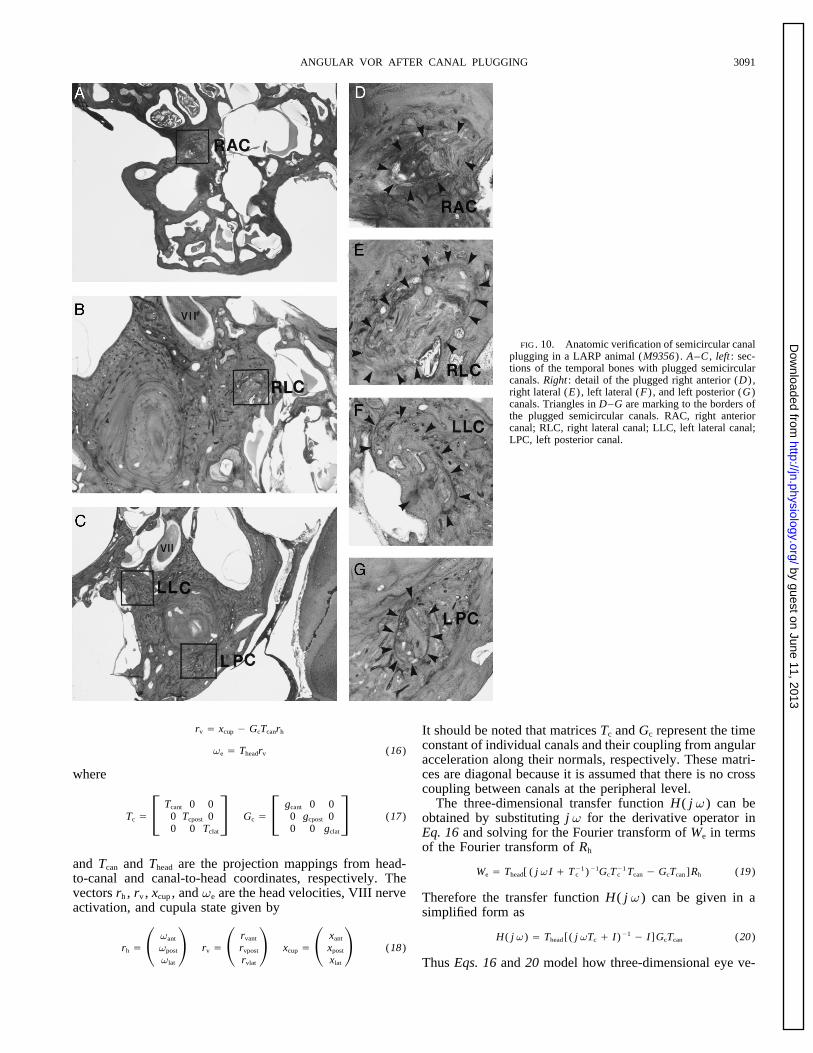
About 1 mo after coil implantation, the semicircular canals weretation also have been described (Baker et al. 1986, 1987;plugged by grinding across the bony and membranous canals andHarrison et al. 1986; Peng et al. 1994; Peterson et al. 1991).packing the orifices with bone dust (Cohen et al. 1964, 1965;That there might be adaptation after canal plugging alsoMoney and Scott 1962; Suzuki and Cohen 1966; Suzuki et al.could be inferred from the classic studies of Ewald (1892). 1964; Yakushin et al. 1995). The plugging was performed on the
After single canals were plugged in pigeons, the birds still side opposite to the ampulla. This left the hair cells of the canalscould fly to the ceiling of their cage on recovery from anes- and otoliths intact. After recovery, the bone fused to provide anthesia. These results were different from the behavior of impenetrable block to the flow of endolymph (Fig. 10).pigeons after unilateral labyrinthectomy. These animals were Two animals had all six semicircular canal plugged (no canal
animals, NC: M9308 and M9357) . One animal had both lateralunable to fly. In addition, they had sustained torsion of thecanal plugged (vertical canals intact, VC: M9354) and two animalshead and held abnormal postures for prolonged periodshad only one reciprocal vertical canal pair intact: right anterior and(Ewald 1892). It is striking, that even after extensive canalleft posterior canals intact (RALP: M9355) or left anterior andplugging in the monkey, animals regain relative posturalright posterior canals intact (LARP: M9306) . Partial data on stepstability and the ability to move rapidly in space after aresponses and time constants also were obtained from five otherperiod of several weeks (Yakushin et al. 1995). The basis animals with canal plugging (LC, M9008; VC, M9003; LARP,
for this adaptation is unknown. One obvious difference be- M9006 and M9356; RALP, M9223) . Data presented in this studytween the canal-plugged and labyrinthectomized animals is were obtained from 2 wk to 2 yr after canal plugging. Deficits inthat the spontaneous discharge of the primary afferents is postural control and uncoordinated head movements in the planemaintained after canal plugging (Goldberg and Fernandez of the plugged canals that were observed in the first weeks after
surgery had largely disappeared by the time of testing. Thus the1975), whereas the spontaneous input is lost after labyrin-results that will be reported were from animals that had recoveredthectomy.from the acute effects of operation.In preliminary studies, we have found that there was a
Canal plugging was verified anatomically in several animalsmeasurable aVOR response when monkeys were rotated inand physiologically in others by the characteristic alteration in thethe plane of plugged canals. This also was present in anresponse to sinusoidal rotation at 0.2-Hz, 607 /s peak velocityanimal with all semicircular canals plugged (Yakushin et al. around an axis perpendicular to the average plane of the plugged
1997), ruling out the possibility that there was reorganiza- canals (Yakushin et al. 1995).tion of vertical canal input to support yaw eye movements.The purpose of this study was to clarify the etiology of the
Data collection and processingresponses to high-frequency sinusoids and steps of velocityin animals with plugged canals. Our aim was to develop During testing, the monkey’s head was fixed in a rigid 15 cm
frame made of 12.2-mm Plexiglas. The frame held two sets of 13-a three-dimensional dynamic and kinematic model of thecm field coils that generated orthogonal oscillating magnetic fieldssemicircular canals that predicted the quantitative responsesat a frequency of 24 kHz. The axes of the field coils were alongbefore and after plugging to low- and high-frequency rota-the interaural and dorsoventral axes of the head, establishing ations and to steps of constant velocity.head fixed reference frame for measuring the orientation of thesearch coils in front and on top of the left eye. The head was fixed
M E T H O D S relative to the field coils with the left eye centered in the magneticfields. Eye velocities were calibrated by rotating the animals in
Experiments were performed on six cynomolgus monkeys. In light at 307 /s about the pitch, roll, and yaw axis. It was assumedfive animals, one or more reciprocal semicircular canal pairs were that horizontal and vertical gains were unity in this conditionplugged. One other animal was used to obtain control data. Partial (Crawford and Vilis 1991; Dai et al. 1991; Raphan et al. 1979;data also were obtained from five other canal-plugged animals. Robinson 1963). Roll gains were assumed to be 0.6 when rotationThe experiments conformed to the Guide for the Care and Use of was around a naso-occipital axis aligned with the spatial verticalLaboratory Animals (National Research Council 1996) and were (Crawford and Vilis 1991; Henn et al. 1992; Yakushin et al. 1995).approved by the Institutional Animal Care and Use Committee. This agrees with roll gains determined for monkeys using other
techniques (Dai et al. 1994; Telford et al. 1996; Yue et al. 1994).In this paper, we use the terms ‘‘horizontal’’ and ‘‘yaw’’ inter-Surgical procedureschangeably. Eye velocities to the left, down, and counterclockwise(from the animal’s point of view) are represented by downwardThe surgical procedures used in these experiments have been
described in detail (Yakushin et al. 1995). Briefly, head bolts deflections in the velocity traces in the figures. Data were recordedwith amplifiers having a band-pass of DC to 40 Hz. The equipmentwere implanted on the skull in dental acrylic cement under general
anesthesia in sterile surgical conditions. This provided painless was controlled and data were acquired with a computer. Voltageswere digitized at 600 Hz/channel with 12 bit resolution and storedfixation of the head in stereotaxic coordinates during testing. When
J1032-7/ 9k2e$$no07 12-09-98 07:00:21 neupa LP-Neurophys
by guest on June 11, 2013http://jn.physiology.org/
Dow
nloaded from

ANGULAR VOR AFTER CANAL PLUGGING 3079
on optical disk. Eye position voltages were smoothed and digitally stability of the head was not different from that used in the othermonkeys in this series. Tape was placed around the head over thedifferentiated by finding the slope of the least squares linear fit,
corresponding to a filter with a 3-dB cutoff ú40 Hz, the cutoff eyes at the level of the forehead to reduce possible skin movement,and a coil was attached to the tape. No modulation in voltage wasfrequency of the filters used for data acquisition. Saccades were
eliminated using an order statistic filter (Engelken and Stevens induced by applied frequencies of oscillation °4 Hz or by stepsof velocity. We conclude that there was neither deformation of the1990; Engelken et al. 1996).
During testing animals sat in a primate chair in a multiaxis coil box nor slip of the monkey’s head relative to the coil fieldduring rotation at high frequency or during steps.vestibular stimulator (Neurokinetics) that has been described pre-
viously (Dai et al. 1991; Reisine and Raphan 1992). In brief,the stimulator is composed of three gimbaled axes for rotation, a Step response analysishorizontal axis parallel to the earth horizontal, a nested yaw axis,
Animals were tested with approximate ramps of velocity. Rota-and a doubly nested inner pitch/roll axis. The yaw and pitch/rolltion began with an approximate step of angular acceleration fromaxes are enclosed in a light-tight optokinetic cylinder, 91 cm in0 to 2707 /s2 (Fig. 9B) . Peak acceleration was reached after 33 msdiameter with 107 black and white stripes. The axis of the OKNand was maintained for 195 ms, after which the animals werecylinder also is controlled independently and is collinear with thedecelerated with the same slope to 07 /s2 . This generated a rampyaw axis. Each axis went through the center of rotation of the headof velocity over 260 ms up to a peak of 607 /s (Fig. 9A) . Toand was computer controlled. The pitch/roll and horizontal axesmeasure the horizontal aVOR gain, rotation was held for 5 s inwere controlled by position servos and the yaw and OKN axes bydarkness. Rotation was stopped with the animal in light for ¢5 svelocity servos. The peak acceleration of the primate axis wasto damp any postrotatory response (Raphan et al. 1979). Alternate2707 /s2 . Monkeys sat in the primate chair with their heads fixedrotations to the left and right were repeated 10 times.to a box that held the field coils. When animals were rotated in
The interest in this paper was in the canal dynamics. This in-light, they had full field optokinetic stimulation. When the monkeyscludes the contributions of the endolymph flow, the elastic andwere upright, the lateral semicircular plane formed an angle of 15–viscous properties of the canal structure, and the spike encoding227 with earth horizontal (Blanks et al. 1985; Reisine et al. 1988).process that transduces the mechanical motion to nerve impulsesIn these experiments, the animals sat so that the interaural axisalong the eighth nerve (Fernandez and Goldberg 1971; Highsteinwas aligned with the pitch/roll axis. They were upright or wereet al. 1996; Landolt and Correia 1980). The canal dynamics cantilted to a fixed pitch position and rotated about a spatial verticalbe described as a first-order model given byaxis. This paradigm is similar to that used in previous studies of
horizontal eye movements induced after semicircular canal plug- ddt
rv /1Tc
rv Å 0gcddtvc (1)ging (Angelaki and Hess 1996; Angelaki et al. 1996; Baker and
Peterson 1991; Baker et al. 1982, 1986; Bohmer et al. 1985; Minorwhere gc is the coupling relating canal output to angular accelera-and Goldberg 1990; Yakushin et al. 1995).tion input d/dt(vc ) (Raphan and Cohen 1981). The parameter,Tc , is the dominant time constant of the canal dynamics, and rv is
Sinusoidal analysis the canal afferent output. We will refer to Tc as the canal timeconstant. Because the acceleration lasted for only 260 ms, only theTesting was performed with animals upright (07) and staticallycanal dynamics and direct pathway would be activated during thetilted forward (nose down, /) or backward (nose up, 0) in 107period of acceleration. Eye velocity was assumed to be proportionalincrements up to {907. Animals were rotated sinusoidally about ato the afferent output of the canals about the axis of rotation. Itspatial vertical axis in darkness. This stimulus induced yaw eyewas assumed further that there was no contribution of velocitymovements when animals were upright and both yaw and roll eyestorage. This can be approximated by an equivalent direct pathwaymovements when the animals were pitched forward or back (Yaku-gain, gd , referenced to an equivalent composite canal activation,shin et al. 1995). Animals with normal semicircular canals wererveq . Therefore eye velocity about a particular axis, ve , can beused as controls, and their data were compared with data obtainedgiven byfrom canal-plugged animals. At least 10 cycles were collected for
each of the 19 test positions. Two normal and the five canal-plugged ddt
rveq /1Tc
rveq Å 0gceqddtvhanimals were tested during sinusoidal rotation at a variety of frequen-
cies (0.2 and 0.5 Hz at peak velocity 607 /s, 1.0 Hz at É337 /s, 2.0ve Å gdrveq (2)Hz atÉ167 /s, and 4.0 Hz atÉ67/s). Desaccaded eye velocities were
fit with a sinusoid at the frequency of oscillation using a least mean where gceq is the coupling from head acceleration to rreq . The gainsquare algorithm. From this, the average value of peak eye velocity of the reflex, gVOR, then can be given by the equationand the phase relative to the stimulus (temporal phase) were deter-
gVOR Å gceqgd (3)mined. The maximum and minimum values of the data for eachindividual cycle of eye velocity were obtained at the times of the Using Eqs. 2 and 3, the gain, gVOR, can be measured in a number
of ways dependent on the magnitude of the dominant time constant.peaks of the fitted curves. Temporal gains of the aVOR were deter-mined for each cycle as (peak-to-peak eye velocity)/(peak-to-peak In the normal monkey, Tc was assumed equal to 4 s (3–6 s)
(Buttner and Waespe 1981; Correia et al. 1992; Goldberg andstimulus velocity). Mean gain and standard deviations were obtainedover all peak values. Fernandez 1971; Reisine and Henn 1984). During the 260 ms of
acceleration, using Eqs. 1–3, eye velocity about a given axis canWe evaluated whether there was an artifactual component in theeye velocity as a result of deformation of the Plexiglas box in be related to head velocity and will be approximately linear withwhich the field coils were embedded. An eye coil of the same d
dtve Å 0gVOR
ddt
vh (4)diameter and number of turns as that placed on the eye of themonkey was mounted on a Plexiglas plate and fixed to the coilbox in the center of the field. No modulation in voltage was induced Therefore, using Eq. 4, the gain, gVOR, could be computed as a
ratio of eye acceleration to head acceleration or as a ratio of theby applied frequencies of oscillation°4 Hz or by steps of velocity.We also tested whether there was movement of the animal’s head peak eye velocity to stimulus velocity (vh) .
For canal time constants Tc , which were much less than 4 s, andrelative to the coil box, using animals with the Sirota head implan-tation technique (Sirota et al. 1988). In this technique, the lateral for which eye velocity responses reached a steady-state value
J1032-7/ 9k2e$$no07 12-09-98 07:00:21 neupa LP-Neurophys
by guest on June 11, 2013http://jn.physiology.org/
Dow
nloaded from

YAKUSHIN, RAPHAN, SUZUKI, ARAI, AND COHEN3080
within the 200 ms from onset of acceleration (Fig. 9D) , d/dt(rveq )(Eq. 2) is close to zero. In these instances, the gain, gVOR, couldbe computed from
1Tc
ve Å 0gVORddt
vh (5)
as the ratio of the steady-state eye velocity to (time constantmultiplied by stimulus acceleration, d/dt(vh)) . For normal ani-mals, gains computed from the steady-state and acceleration peri-ods are equivalent; measurements were made from slow phasevelocity during the steady state. In the canal-plugged animals, thegains were determined from the calculation shown in Eq. 5. (Fora more complete description of the computation, see section onModeling the three-dimensional kinematics and dynamics of thesemicircular canals: effects of plugging below).
The gain and canal time constant (Tc ) for plugged lateral canalwas obtained from responses to rotation when the head was tilted30–407 forward, which approximately eliminated the contributionof the vertical canals. Desaccaded eye velocities in response tosteps of rotation at 607 /s were synchronized to the beginning ofrotation. The average value of eye velocity divided by averagestimulus velocity over the first 20 ms that contained no saccadesafter the ramp of velocity had stabilized was taken as the gain ofthe individual response. Individual gains were averaged over 10responses to obtain the average gain and standard deviation of thestep response. Time constants were computed by fitting individualresponse curves with a single exponential rising to the ‘‘steady-state’’ value and finding their average value and standard deviation.
Both per- and postrotatory responses produced by constant ve-locity rotation in darkness were used to measure the central orvelocity storage time constant of the horizontal aVOR. After theinitial 260-ms period of acceleration, the velocity of rotation washeld constant for a prolonged period with the animal in darknessuntil the slow phase velocity had decayed to zero. A similar acceler-ation profile was used to stop rotation, generating a postrotatoryresponse. The velocity storage time constant for the normal mon-keys was calculated from a double exponential technique pre-viously described (Raphan et al. 1979), assuming a 4-s canal timeconstant. The time constant of velocity storage also was obtainedfrom optokinetic after-nystagmus (OKAN) by fitting the decliningvelocity with a single exponential (Cohen et al. 1977).
FIG. 1. A : position of the left labyrinth of a monkey. Coordinate frameCoordinate notationof the head, taken as the stereotaxic frame, is the coordinate frame in which
In previous studies from our laboratory, we used the coordinate roll ( torsion), pitch (vertical) , and yaw (horizontal) eye movements werenotation originally used by Fernandez and Goldberg (1976). This measured. X represents the roll axis, Y the pitch axis, and Z the yaw axis.reference frame has the pitch axis (eX ) along an interaural axis B : representation of the relative orientation of the coordinate axes of the
stereotaxic frame (X–Z ) and the coordinate axes determined by the normalsfrom the left ear. The roll axis (eY ) lies along the naso-occipitalto the semicircular canal planes. Positive directions of these normals wereaxis and points out the back of the head, and the yaw axis (eZ ) isdetermined by using a right hand rule for the rotation direction, whichout the top of the head. This frame was utilized in Yakushin etexcited an individual canal. Xc represents the positive direction for theal. (1995) in which three-dimensional eye responses from canal-anterior canal, Yc the posterior canal, and Zc the lateral canal. Ul is theplugged monkeys were studied at 0.2 Hz (Yakushin et al. 1995), angle between stereotaxic vertical axis (Z ) and the average direction of the
and in Wearne et al. (1996–1998), which referenced much of the lateral canal axis (Zc ) . The Xc axis is obtained by rotating the X axis aboutprevious work. In the current study, we have used a different the Y axis through an angle 0Ua and then about the intermediate Z axisreference frame, one that is rotated 907 relative to that used pre- through an angle Ca . The Yc axis is obtained by rotating the Y axis aboutviously (Fig. 1) . This frame has been used in psychophysical the Y axis through an angle 0Up ( this corresponds to no rotation of the
vector but only orients the intermediate Z axis) and then about the intermedi-studies of the vestibular system (Guedry 1974), and recently hasate Z axis through an angle Cp . Each canal axis is therefore characterizedbecome a general standard in vestibular studies. It is also com-by 2 generalized coordinates that determine its orientation.monly used in aerospace engineering. The major differences are
that the basis vectors for the head coordinate frame are eX (roll) ,way, the most general transformation between the two basis setseY (pitch) , and eZ (yaw), corresponding to the X , Y, and Z axes,can be given by six angles that define how the canal afferents coderespectively. The canal basis unit vectors are defined as beforethe head acceleration or velocity signals (see modeling section for(Yakushin et al. 1995). These are the normals to the anterior canala complete derivation).(ea ) , posterior canal (ep) , and lateral canal (el ) , corresponding to
the Xc , Yc , and Zc axes. These basis vectors do not form an orthonor-Convention of temporal gain and phase representationmal set.
Each canal basis vector can be given as a rotation of one of the Because of the vector nature of the head and eye velocities, theprojections of the stimulus velocity, which is along the spatialhead basis vectors and can be defined by two Euler angles. In this
J1032-7/ 9k2e$$no07 12-09-98 07:00:21 neupa LP-Neurophys
by guest on June 11, 2013http://jn.physiology.org/
Dow
nloaded from

ANGULAR VOR AFTER CANAL PLUGGING 3081
curves represented in our previous work on canal plugging at 0.2Hz (Yakushin et al. 1995). For perfect compensation, phases weredefined as 01807 for yaw under all conditions and for roll whentilted back (Fig. 2, E, F, and H) . For roll in the tilted forwardcondition, the phase for perfect compensation was defined as 07(Fig. 2G) . After canal plugging, there were increases in phaseshifts at higher frequencies.
Gain (the ratio of eye velocity in head to stimulus velocity inspace) was defined as positive or negative depending on the posi-tive direction for the unit vector of the head coordinate relative tostimulus velocity (X , Z, Fig. 2, A and B) (Yakushin et al. 1995).When the animal was tilted õ907 forward or backward, yaw eyevelocity was considered to have a positive gain when it was in theshaded region around the phase of01807 relative to stimulus veloc-ity (compare Fig. 2, C and D, and E and F) . For roll, gain wasconsidered positive when tilted forward (Fig. 2G) and negativewhen tilted back (Fig. 2H) .
Data analysis
The spatial gains and phases of the horizontal and roll aVOR werethe variables of interest of this study. An equal number of individualgain values was obtained for each tilt position. The gain values as afunction of tilt angle were fit, using a minimum mean square errorcriteria, with a sinusoid y Å A∗ cos(x / B), where x is the tilt angle.The peak value (A, spatial gain) and its phase relative to the upright(B, spatial phase) were obtained from the fit.
Changes in eye position as a function of tilt angle can introduceerrors in the measurement of eye velocity (Yakushin et al. 1995).To determine the approximate size of the errors, we evaluated theeffect of static head pitch on horizontal, vertical, and roll compo-nents of eye position in the two normal and four canal-pluggedanimals used in this study. Head tilt had no effect on horizontaleye position in the normal animals (M9357 and M9358) when theywere tested at different frequencies from 0.2 to 4.0 Hz (0 { 27) .The vertical component of eye position varied as a function of tiltangle. The eyes were minimally deviated in the upright and {907tilt positions and maximally deviated for {457 tilts (M9358, 77;
FIG. 2. Conventions for describing eye velocity in the head. A and B :M9357, 47) . The torsional component of eye position was notpositive directions associated with the vector components of head and eyeaffected by forward tilts in either normal animal. It increased as amovements referenced to a head-based coordinate frame. A positive valuefunction of backward tilt in M9358, being maximal (up to 0207)along the reference directions corresponds to a rotation according to a right
hand rule. Thus a positive value for Z corresponds to a leftward movement when the animal was tilted ¢507. In the canal-plugged animals,and a positive value for X corresponds to a clockwise rotation from the horizontal and roll eye position did not vary as a function of headanimal’s viewpoint. Reference sinusoid for computing gains was along the tilt. The vertical component deviated up to {107 for the RALPspatial vertical (C and D) . E and G : for the tilted-forward condition, the and LARP animals. The eye deviation was up with forward tiltscomponents of head velocity along the yaw (Z ) and roll (X ) axes are and down with backward tilts. Thus in agreement with our previouspositive, and the corresponding compensatory eye velocity components findings (Yakushin et al. 1995), eye deviations within {157 didwere taken as 01807 out of phase (E) or in phase (07) (G) with reference
not introduce significant errors between eye velocities and thosesinusoid. This has been taken as a positive gain. F and H : with the headcomputed as a derivative of coil voltages.tilted backward, the component of head velocity along the yaw axis is
positive and the component along the roll axis is negative. This correspondsto yaw (F) and roll (H) eye velocities, which have phases of 01807 with Statistical analysis of datathe reference sinusoid. Gain of the yaw component therefore was taken aspositive, whereas the gain of the roll component was taken as negative. A standard unpaired t-test was used to compare two groups ofPeak temporal phases were varied from optimal in or out of phase position data. For more than two groups of data, an analysis of variance(arrow, E–H) after canal plugging when tested at different frequencies. (ANOVA) was used. If the general ANOVA showed significantPhase was considered to be positive or negative if it did not deviate more
differences between data sets, then each between-group degree ofthan {907 from corresponding optimal phase (shadow area, E–H) .freedom was analyzed separately by developing orthogonal con-trasts. In this case, results of the test were adjusted with a Scheffeapproach (Keppel 1991). In the statistical analyses for goodnessvertical, is related by a cosine of the angle between the stimulus
axis and the axis of measurement. The positive Z axis always of fit, the null hypothesis was that the mean gain of the data foreach tilt angle is equal to the value obtained from the optimal fitmaintains an angle °907 relative to the positive stimulus axis for
head tilts between {907 (Fig. 2, A and B) . Therefore the projection to the data over all angles of tilt. Hypotheses were tested by exam-ining the ratio of the variance of the data relative to the mean andof the stimulus onto the yaw axis of the head is always positive.
The positive X axis has an angle ú907 when tilted forward and the variance relative to the fitted value. Because each measurementof gain at each tilt angle was done independently, the ratio followsõ907 when tilted back (Fig. 2, A and B) and the projection changes
sign. Therefore references for gains and phases of eye velocity an F distribution (Keppel 1991; Yakushin et al. 1995). Data inthis paper are described by means { SD.were defined independently to be consistent with the spatial gain
J1032-7/ 9k2e$$no07 12-09-98 07:00:21 neupa LP-Neurophys
by guest on June 11, 2013http://jn.physiology.org/
Dow
nloaded from

YAKUSHIN, RAPHAN, SUZUKI, ARAI, AND COHEN3082
J1032-7/ 9k2e$$no07 12-09-98 07:00:21 neupa LP-Neurophys
by guest on June 11, 2013http://jn.physiology.org/
Dow
nloaded from

ANGULAR VOR AFTER CANAL PLUGGING 3083
R E S U L T S Therefore contributions of frequencies other than 4 Hz tothe stimulus and response were not significant. Because the
Sinusoidal analyses amplitude of the responses and the signal to noise ratio waslowest at 4 Hz, this conclusion can be extended to the lower
TEMPORAL CHARACTERISTICS OF THE aVOR BEFORE AND frequencies, as well.AFTER CANAL PLUGGING. In response to sinusoidal oscilla- The responses of the NC animal were compared with thetion at 0.2 Hz, the gain of the aVOR is negligible for rotation
horizontal and roll temporal phases of two normal animalsin a plane orthogonal to the average plane of the remainingas a function of head tilt about the interaural (pitch) axis atcanals (Yakushin et al. 1995). The effects of frequency onvarious frequencies. For the normal animals, the horizontalthe gain of the responses were striking. In two animals withcomponent of the aVOR was compensatory, being 01807all six semicircular canals plugged (NC animals; M9308 andout of phase with stimulus velocity at each frequency fromM9357) , aVOR gains during rotation at 0.2 Hz about any0.2 to 4.0 Hz for tilts in the range of {907 (Fig. 4A) . Theaxis was close to zero (Fig. 3A) . As stimulus frequencytemporal phases of the roll components were also compensa-increased, the gain of both the horizontal and roll compo-tory, being about 01807 relative to stimulus velocity whennents increased concurrently. In M9308, the peak gains ofthe normal animals were tilted backward and 07 when tiltedthe horizontal component, tested in the upright position, roseforward (Fig. 4D ; also see phase convention, Fig. 2) . Therefrom 0.05 { 0.04 at 0.2 Hz to 0.52 { 0.10 at 4.0 Hz (Fig.was a range of uncertainty approximately equal to {1073, A–D, left) . In M9357, the peak horizontal gain at a fre-around the upright position for the roll temporal phases. Thisquency of 0.2 Hz was significantly different from the gainis a region of head tilt where the gain is close to zero andat a frequency of 4.0 Hz (0.02 { 0.03 and 0.37 { 0.17,the phase is uncertain. This was largely due to the smallrespectively, P õ 0.6∗1008 using 2-tailed t-test) . The rollgains that were elicited in these head orientations. At eachresponses rose from 0.03 { 0.03 at 0.2 Hz to 0.47 { 0.14frequency, the temporal phases of the horizontal and roll com-at 4.0 Hz in M9308 when tested in the 907 tilt forwardponents were not significantly different from being compensa-condition (Fig. 3, A–D, right) and from 0.02 { 0.04 totory at any tilt angle (P ú 0.20, ANOVA). Therefore, data0.28 { 0.16 when M9357 was tilted 907 forward (P õat a given frequency were pooled for all tilt angles, and an0.0008). It should be noted that the peak velocity of theANOVA was performed to determine whether there was vari-stimulus was different for sinusoids at different frequencies.ation of temporal phase as a function of stimulus frequency.Peak stimulus acceleration was maintained at 2007 /s2 acrossVariations in temporal phase of the horizontal and roll compo-the frequency spectrum, which kept the system in the linearnents were small and insignificant for both animals (ANOVA,range for the normal animal. We assumed that linearity alsoScheffe post hoc adjustment; P ú 0.05). Thus temporalwas maintained for the canal-plugged animals.phases were not significantly different from being compensa-As the gains rose, the temporal phases of the horizontaltory at any tilt angle or frequency in the normal animals.sinusoids and roll sinusoids shifted from being in phase with
When the NC animals were tested in different head orien-head acceleration õ1.0 Hz (Fig. 3, B and E) toward beingtations, the temporal phases tended to be relatively invariantin phase with head velocity at 4 Hz (Fig. 3, C–E) . Thusover the range of tilt angles for all frequencies, with thethe gains and phases of the responses tended to normalizeexception of the uncertainty region (Fig. 4, B and E) . Thewhen tested at higher frequencies. Baker et al. (1982) alsotemporal responses of the horizontal and roll eye velocityreported canal responses in two cats with all six semicircularled the normal response by 907 when tested at 0.2 Hz. Thiscanal plugged. Their animals had small, but consistent aVORis demonstrated in Fig. 4 for M9308 (compare thin lines,gains (0.07 and 0.08) when they were tested at 2.5 Hz. TheFig. 4, A and B, 092 { 217 for horizontal and Fig. 4, D andtemporal responses had a phase lead ofÉ907 relative to headE, 072 { 217 for roll phases) . As the frequency increased,acceleration (Baker et al. 1982), consistent with findings inthe average temporal phase shifted toward 01807, whichthis study.was closer to being compensatory for head velocity, as inBecause the peak stimulus velocity at 4.0 Hz was lowthe normal animal (Figs. 3E and 4, 1 Hz: 0119 { 267 for(É67 /s) , other frequency components of the stimulus orhorizontal and 0107 { 97 for roll; 4 Hz: 0148 { 347 fornoise could have distorted the results. We evaluated thehorizontal and 0162 { 77 for roll) . The change in temporalspectral composition of the stimulus and the response bothphase as a function of frequency was similar for M9357before and after canal plugging. The stimulus had an approx-(Fig. 3E) .imate Gaussian spectral density distribution with a mean at
In the VC animal, the intact vertical canals would domi-4{ 0.2 Hz. Eye velocity responses before and after pluggingnate the response when the animal was tilted back and rotatedhad the same spectral distribution. Both had large peaks in(Yakushin et al. 1995). In this position, the temporal phasestheir spectra at 4 Hz with a standard deviation approximatelyof the horizontal component of the aVOR were similar toequal to that of the stimulus. The spectral distribution, boththose of the normal animals (negative values on abscissa,before and after plugging was close to zero, 4 SD from
where the spectrum of the stimulus was at a peak at 4 Hz. Fig. 4, C and F) . When the VC animal was tilted forward
FIG. 3. Horizontal ( left column) and roll (right column) eye velocity of one of the animals in which all 6 canals wereplugged (NC, M9308) tested at 0.2 (A) , 1.0 (B) , 2.0 (C) , and 4.0 Hz (D) . Monkey was upright when the horizontal eyevelocities were recorded ( left) and tilted forward 907 when the roll eye velocities were recorded (right) . rrr, sinusoidalline on each graph represents head velocity with reversed polarity to facilitate comparison. E : when stimulus frequencyincreased, the temporal phases of yaw and roll shifted from being approximately in phase with stimulus acceleration to beingin phase with stimulus velocity (from 090 to 01807) .
J1032-7/ 9k2e$$no07 12-09-98 07:00:21 neupa LP-Neurophys
by guest on June 11, 2013http://jn.physiology.org/
Dow
nloaded from
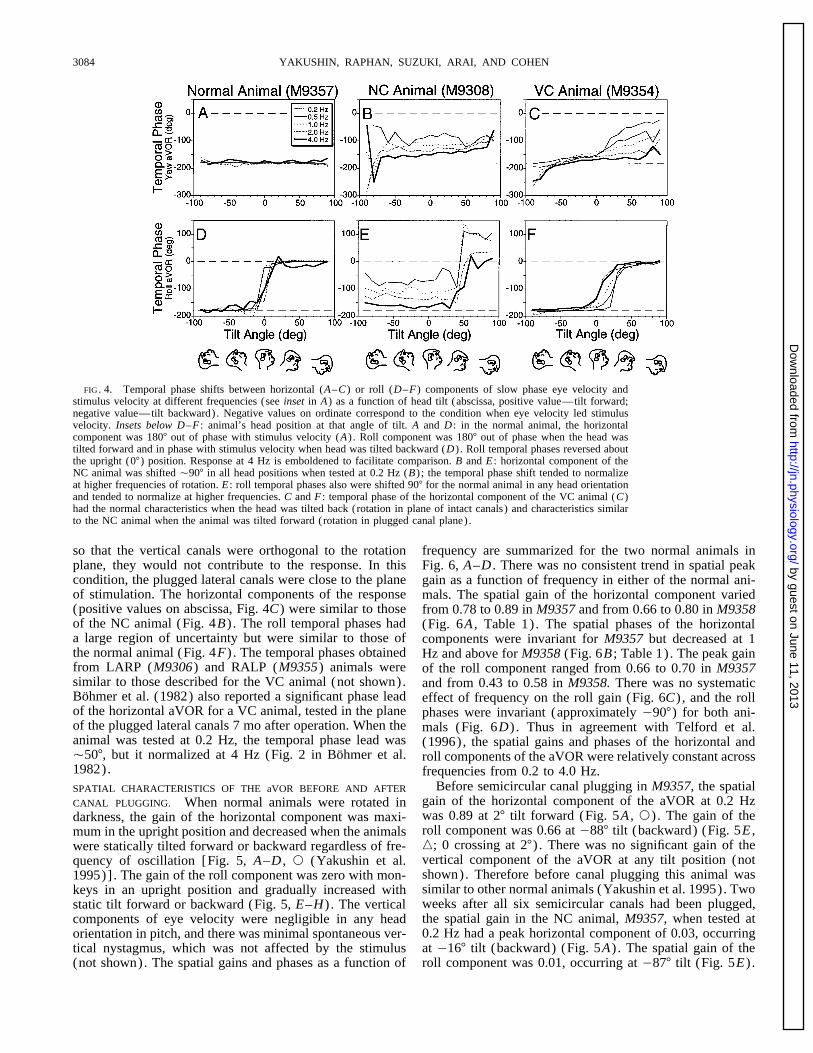
YAKUSHIN, RAPHAN, SUZUKI, ARAI, AND COHEN3084
FIG. 4. Temporal phase shifts between horizontal (A–C) or roll (D–F) components of slow phase eye velocity andstimulus velocity at different frequencies (see inset in A) as a function of head tilt (abscissa, positive value—tilt forward;negative value—tilt backward). Negative values on ordinate correspond to the condition when eye velocity led stimulusvelocity. Insets below D–F : animal’s head position at that angle of tilt. A and D : in the normal animal, the horizontalcomponent was 1807 out of phase with stimulus velocity (A) . Roll component was 1807 out of phase when the head wastilted forward and in phase with stimulus velocity when head was tilted backward (D) . Roll temporal phases reversed aboutthe upright (07) position. Response at 4 Hz is emboldened to facilitate comparison. B and E : horizontal component of theNC animal was shifted Ç907 in all head positions when tested at 0.2 Hz (B) ; the temporal phase shift tended to normalizeat higher frequencies of rotation. E : roll temporal phases also were shifted 907 for the normal animal in any head orientationand tended to normalize at higher frequencies. C and F : temporal phase of the horizontal component of the VC animal (C)had the normal characteristics when the head was tilted back (rotation in plane of intact canals) and characteristics similarto the NC animal when the animal was tilted forward (rotation in plugged canal plane).
so that the vertical canals were orthogonal to the rotation frequency are summarized for the two normal animals inplane, they would not contribute to the response. In this Fig. 6, A–D . There was no consistent trend in spatial peakcondition, the plugged lateral canals were close to the plane gain as a function of frequency in either of the normal ani-of stimulation. The horizontal components of the response mals. The spatial gain of the horizontal component varied(positive values on abscissa, Fig. 4C) were similar to those from 0.78 to 0.89 in M9357 and from 0.66 to 0.80 in M9358of the NC animal (Fig. 4B) . The roll temporal phases had (Fig. 6A , Table 1). The spatial phases of the horizontala large region of uncertainty but were similar to those of components were invariant for M9357 but decreased at 1the normal animal (Fig. 4F) . The temporal phases obtained Hz and above for M9358 (Fig. 6B ; Table 1). The peak gainfrom LARP (M9306) and RALP (M9355) animals were of the roll component ranged from 0.66 to 0.70 in M9357similar to those described for the VC animal (not shown). and from 0.43 to 0.58 in M9358. There was no systematicBohmer et al. (1982) also reported a significant phase lead effect of frequency on the roll gain (Fig. 6C) , and the rollof the horizontal aVOR for a VC animal, tested in the plane phases were invariant (approximately 0907) for both ani-of the plugged lateral canals 7 mo after operation. When the mals (Fig. 6D) . Thus in agreement with Telford et al.animal was tested at 0.2 Hz, the temporal phase lead was (1996), the spatial gains and phases of the horizontal andÇ507, but it normalized at 4 Hz (Fig. 2 in Bohmer et al. roll components of the aVOR were relatively constant across1982). frequencies from 0.2 to 4.0 Hz.
Before semicircular canal plugging in M9357, the spatialSPATIAL CHARACTERISTICS OF THE aVOR BEFORE AND AFTERgain of the horizontal component of the aVOR at 0.2 HzCANAL PLUGGING. When normal animals were rotated inwas 0.89 at 27 tilt forward (Fig. 5A , s) . The gain of thedarkness, the gain of the horizontal component was maxi-roll component was 0.66 at 0887 tilt (backward) (Fig. 5E ,mum in the upright position and decreased when the animalsn; 0 crossing at 27) . There was no significant gain of thewere statically tilted forward or backward regardless of fre-vertical component of the aVOR at any tilt position (notquency of oscillation [Fig. 5, A–D , s (Yakushin et al.shown). Therefore before canal plugging this animal was1995)] . The gain of the roll component was zero with mon-similar to other normal animals (Yakushin et al. 1995). Twokeys in an upright position and gradually increased withweeks after all six semicircular canals had been plugged,static tilt forward or backward (Fig. 5, E–H) . The verticalthe spatial gain in the NC animal, M9357, when tested atcomponents of eye velocity were negligible in any head0.2 Hz had a peak horizontal component of 0.03, occurringorientation in pitch, and there was minimal spontaneous ver-at 0167 tilt (backward) (Fig. 5A) . The spatial gain of thetical nystagmus, which was not affected by the stimulus
(not shown). The spatial gains and phases as a function of roll component was 0.01, occurring at 0877 tilt (Fig. 5E) .
J1032-7/ 9k2e$$no07 12-09-98 07:00:21 neupa LP-Neurophys
by guest on June 11, 2013http://jn.physiology.org/
Dow
nloaded from
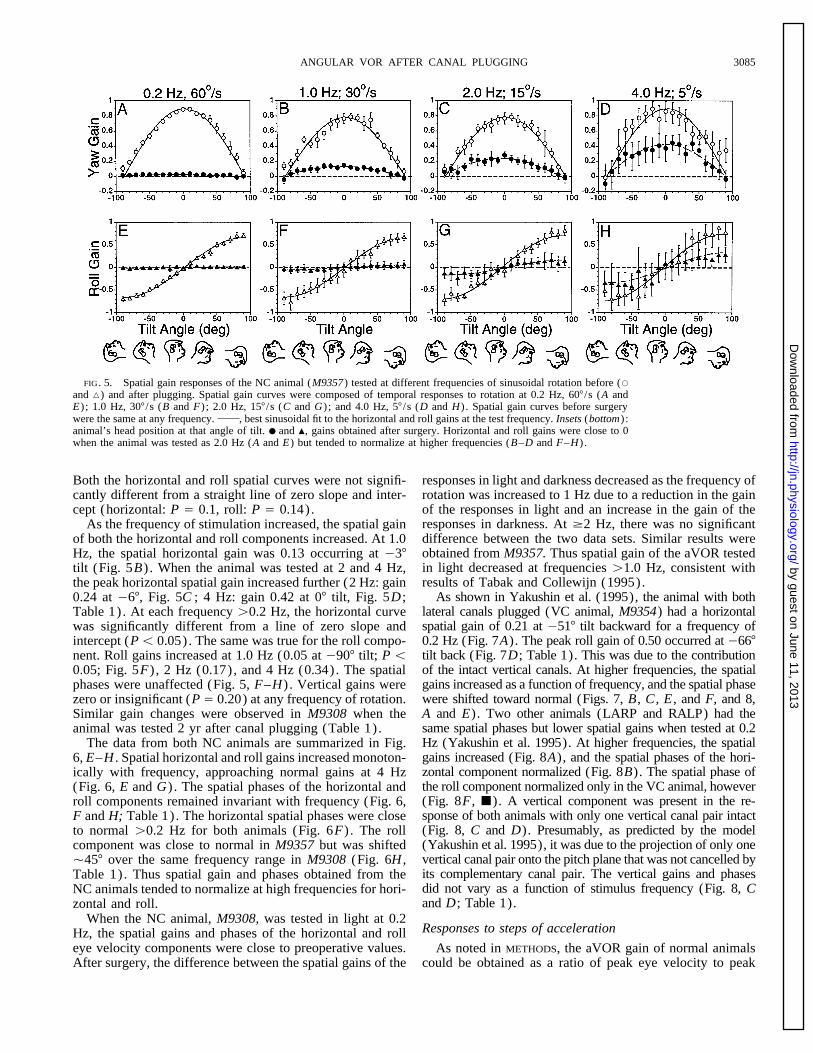
ANGULAR VOR AFTER CANAL PLUGGING 3085
FIG. 5. Spatial gain responses of the NC animal (M9357) tested at different frequencies of sinusoidal rotation before (s
and n) and after plugging. Spatial gain curves were composed of temporal responses to rotation at 0.2 Hz, 607 /s (A andE) ; 1.0 Hz, 307 /s (B and F) ; 2.0 Hz, 157 /s (C and G) ; and 4.0 Hz, 57 /s (D and H) . Spatial gain curves before surgerywere the same at any frequency. , best sinusoidal fit to the horizontal and roll gains at the test frequency. Insets (bottom) :animal’s head position at that angle of tilt. ● and m, gains obtained after surgery. Horizontal and roll gains were close to 0when the animal was tested as 2.0 Hz (A and E) but tended to normalize at higher frequencies (B–D and F–H) .
Both the horizontal and roll spatial curves were not signifi- responses in light and darkness decreased as the frequency ofrotation was increased to 1 Hz due to a reduction in the gaincantly different from a straight line of zero slope and inter-
cept (horizontal: P Å 0.1, roll: P Å 0.14). of the responses in light and an increase in the gain of theresponses in darkness. At ¢2 Hz, there was no significantAs the frequency of stimulation increased, the spatial gaindifference between the two data sets. Similar results wereof both the horizontal and roll components increased. At 1.0obtained from M9357. Thus spatial gain of the aVOR testedHz, the spatial horizontal gain was 0.13 occurring at 037in light decreased at frequencies ú1.0 Hz, consistent withtilt (Fig. 5B) . When the animal was tested at 2 and 4 Hz,results of Tabak and Collewijn (1995).the peak horizontal spatial gain increased further (2 Hz: gain
As shown in Yakushin et al. (1995), the animal with both0.24 at 067, Fig. 5C ; 4 Hz: gain 0.42 at 07 tilt, Fig. 5D ;lateral canals plugged (VC animal, M9354) had a horizontalTable 1). At each frequency ú0.2 Hz, the horizontal curvespatial gain of 0.21 at 0517 tilt backward for a frequency ofwas significantly different from a line of zero slope and0.2 Hz (Fig. 7A). The peak roll gain of 0.50 occurred at 0667intercept (Põ 0.05). The same was true for the roll compo-tilt back (Fig. 7D; Table 1). This was due to the contributionnent. Roll gains increased at 1.0 Hz (0.05 at 0907 tilt; P õof the intact vertical canals. At higher frequencies, the spatial0.05; Fig. 5F) , 2 Hz (0.17), and 4 Hz (0.34). The spatialgains increased as a function of frequency, and the spatial phasephases were unaffected (Fig. 5, F–H) . Vertical gains werewere shifted toward normal (Figs. 7, B, C , E , and F, and 8,zero or insignificant (P Å 0.20) at any frequency of rotation.A and E). Two other animals (LARP and RALP) had theSimilar gain changes were observed in M9308 when thesame spatial phases but lower spatial gains when tested at 0.2animal was tested 2 yr after canal plugging (Table 1).Hz (Yakushin et al. 1995). At higher frequencies, the spatialThe data from both NC animals are summarized in Fig.gains increased (Fig. 8A), and the spatial phases of the hori-6, E–H . Spatial horizontal and roll gains increased monoton-zontal component normalized (Fig. 8B). The spatial phase ofically with frequency, approaching normal gains at 4 Hzthe roll component normalized only in the VC animal, however(Fig. 6, E and G) . The spatial phases of the horizontal and(Fig. 8F, j). A vertical component was present in the re-roll components remained invariant with frequency (Fig. 6,sponse of both animals with only one vertical canal pair intactF and H; Table 1). The horizontal spatial phases were close(Fig. 8, C and D). Presumably, as predicted by the modelto normal ú0.2 Hz for both animals (Fig. 6F) . The roll(Yakushin et al. 1995), it was due to the projection of only onecomponent was close to normal in M9357 but was shiftedvertical canal pair onto the pitch plane that was not cancelled byÇ457 over the same frequency range in M9308 (Fig. 6H ,its complementary canal pair. The vertical gains and phasesTable 1). Thus spatial gain and phases obtained from thedid not vary as a function of stimulus frequency (Fig. 8, CNC animals tended to normalize at high frequencies for hori-and D; Table 1).zontal and roll.
When the NC animal, M9308, was tested in light at 0.2Responses to steps of accelerationHz, the spatial gains and phases of the horizontal and roll
eye velocity components were close to preoperative values. As noted in METHODS, the aVOR gain of normal animalscould be obtained as a ratio of peak eye velocity to peakAfter surgery, the difference between the spatial gains of the
J1032-7/ 9k2e$$no07 12-09-98 07:00:21 neupa LP-Neurophys
by guest on June 11, 2013http://jn.physiology.org/
Dow
nloaded from

YAKUSHIN, RAPHAN, SUZUKI, ARAI, AND COHEN3086
FIG. 6. Spatial gains and phases of the normal (A–D) and NC animals (E–H) tested at different frequencies.A–D : spatial gains (A and C) and phases (B and D) ofthe horizontal (A and B) and roll (C and D) componentsof the angular vestibulo-ocular reflex (aVOR) of 2 normalmonkey (s, M9357; n, M9358) tested in darkness atdifferent frequencies. E–H : spatial gains (E and G) andphases (F and H) of the horizontal (E and F) and roll(G and H) components of the aVOR of the NC animals(M9308 and M9357) tested in darkness at frequencies of0.2–4.0 Hz. – – – , correspond to 07 in B and F and0907 in D and H, the normal values for the horizontaland roll components. In general, gains and phases werestable across the different stimulus frequencies. Spatialphases at 0.2 Hz (F and H) were omitted because gainswere close to 0 and, therefore, phases were not meaningful(see Table 1).
stimulus velocity or as a ratio of eye acceleration to stimulus velocity of this exponential increase was symmetrical, andthere was no significant difference between the response toacceleration using a velocity ramp stimulus with a high ac-
celeration (Fig. 9, A and C ) . This is because eye velocity either side (left: 14.7 { 3.37 / s, right 14.6 { 3.37 / s; P õ0.05). For one NC animal, M9308, aVOR gain, gVOR, [eyeessentially follows stimulus velocity during the ramp. After
the semicircular canals were plugged, the eye velocity re- velocity/ (stimulus acceleration * time constant)] was0.81 { 0.15 to the left and 0.72 { 0.15 to the right (P Åsponse did not follow stimulus velocity during acceleration.
Stimulus velocity rose linearly, but eye velocity had an expo- 0.33). Both were not significantly different from normal(P Å 0.480 and P Å 0.479 for rotation to the left and to thenential increase which reached a steady-state value when
stimulus velocity was still increasing (Fig. 9, A and D) . The right, respectively; Table 2). In the other NC animal, M9357,the values of Tc and steady-state eye velocities were similartime constant (Tc ) of the exponential increase in animal
M9308 was 69 { 13 ms for slow phase eye velocity to the (Table 2), and computed gains were 0.76{ 0.21 for rotationto the left and 0.81 { 0.30 for rotation to the right. Similarleft and 78 { 15 ms for slow phase eye velocity to the right
(P Å 0.37) when the animal was rotated in the plane of the results were obtained for one VC, two LARP, and two RALPanimals (Table 2). That is, the canal-plugged gains wereplugged lateral canal (307 tilt forward). Steady-state eye
J1032-7/ 9k2e$$no07 12-09-98 07:00:21 neupa LP-Neurophys
by guest on June 11, 2013http://jn.physiology.org/
Dow
nloaded from

ANGULAR VOR AFTER CANAL PLUGGING 3087
TABLE 1. Spatial gain and phases of the horizontal and roll aVOR components
Normal Animals NC Animals VC LARP RALPFrequency, Animal Animal Animal
Hz M9357 M9358 M9308 M9357 M9354 M9306 M9355
HorizontalSpatial gain 0.2 0.89 0.80 0.05 0.03 0.21 0.06 0.15
0.5 0.88 0.79 — 0.06 0.30 0.17 0.311.0 0.78 0.66 0.22 0.13 0.39 0.26 0.372.0 0.78 0.73 0.38 0.24 0.53 0.39 0.564.0 0.89 0.71 0.55 0.42 0.53 0.55 0.62
Spatial phase 0.2 2 10 22 016 051 029 0210.5 0 11 — 08 015 09 091.0 2 09 3 03 03 07 52.0 1 09 10 06 03 5 014.0 2 05 10 0 3 6 01
RollSpatial gain 0.2 0.66 0.43 0.04 0.01 0.50 0.21 0.42
0.5 0.70 0.49 — 0.03 0.51 0.34 0.491.0 0.66 0.45 0.22 0.05 0.49 0.31 0.512.0 0.70 0.45 0.38 0.17 0.58 0.36 0.524.0 0.74 0.58 0.58 0.34 0.65 0.42 0.64
Spatial phase 0.2 088 089 067 087 066 078 0600.5 090 089 — 084 067 067 0551.0 089 090 043 090 070 071 0712.0 094 093 047 086 096 068 0534.0 092 088 045 084 099 074 042
aVOR, angular vestibulo-ocular reflex; NC, no canals; VC, vertical canals intact; LARP, left anterior and right posterior canals intact; RALP, rightanterior and left posterior canals intact.
close to preoperative values. The results indicate that canal time constants before operation varied between 40 and 50s. After plugging, OKAN time constants on average wereplugging shortens the canal time constant by almost two
orders of magnitude but does not affect its gain. Thus the Ç10 s (Table 3). The reduction of the integrator time con-stant would raise the low-frequency 3-dB cutoff to Ç0.02response of the unplugged canals to a ramp of velocity over
260 ms was close to the velocity of the stimulus (Fig. 9, A Hz, which is still well below the frequency range consideredin this study. This justified neglecting the contribution ofand C ) . For the plugged canals, the time constant was much
shorter, and the response to the same ramp of velocity was the velocity storage integrator.exponential and was closer to the acceleration of the stimulus
Morphological and physiological verification of canal(Fig. 9, B and D) .pluggingCanal plugging also was associated with a reduction in
the dominant time constant of the aVOR and of OKAN. Morphology was available for several of the animals thedata of which are presented in this study. An example fromThe average central aVOR and OKAN (velocity storage)
FIG. 7. Spatial responses of the mon-key with both lateral canals plugged (VCanimal) tested at different frequency of si-nusoidal rotation. Average gains of the yaw(A–C) and roll (D–F) components, testedat 0.2 Hz and 607 /s (A and D) , at 1.0 Hzand 337 /s (B and E) , and at 4.0 Hz, 77 /s(C and F) . , minimum mean squareerror fit to the horizontal and torsional gainsat the tested frequencies. Insets (bottom) :animal’s head position at that angle of tilt.When this animal was tested after surgery,the responses (gains and spatial phases)tended to normalize as the testing fre-quency increased.
J1032-7/ 9k2e$$no07 12-09-98 07:00:21 neupa LP-Neurophys
by guest on June 11, 2013http://jn.physiology.org/
Dow
nloaded from
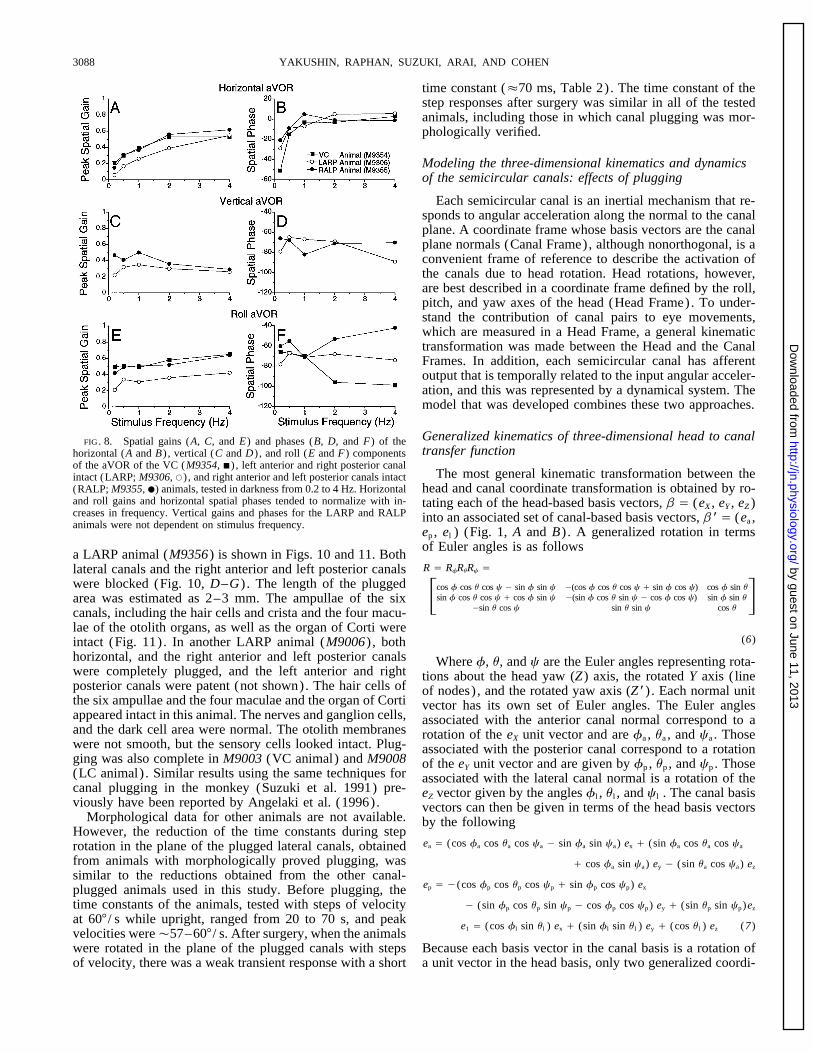
YAKUSHIN, RAPHAN, SUZUKI, ARAI, AND COHEN3088
time constant (É70 ms, Table 2). The time constant of thestep responses after surgery was similar in all of the testedanimals, including those in which canal plugging was mor-phologically verified.
Modeling the three-dimensional kinematics and dynamicsof the semicircular canals: effects of plugging
Each semicircular canal is an inertial mechanism that re-sponds to angular acceleration along the normal to the canalplane. A coordinate frame whose basis vectors are the canalplane normals (Canal Frame), although nonorthogonal, is aconvenient frame of reference to describe the activation ofthe canals due to head rotation. Head rotations, however,are best described in a coordinate frame defined by the roll,pitch, and yaw axes of the head (Head Frame). To under-stand the contribution of canal pairs to eye movements,which are measured in a Head Frame, a general kinematictransformation was made between the Head and the CanalFrames. In addition, each semicircular canal has afferentoutput that is temporally related to the input angular acceler-ation, and this was represented by a dynamical system. Themodel that was developed combines these two approaches.
Generalized kinematics of three-dimensional head to canalFIG. 8. Spatial gains (A, C, and E) and phases (B, D, and F) of thetransfer functionhorizontal (A and B) , vertical (C and D) , and roll (E and F) components
of the aVOR of the VC (M9354, j) , left anterior and right posterior canalThe most general kinematic transformation between theintact (LARP; M9306, s) , and right anterior and left posterior canals intact
(RALP; M9355, ●) animals, tested in darkness from 0.2 to 4 Hz. Horizontal head and canal coordinate transformation is obtained by ro-and roll gains and horizontal spatial phases tended to normalize with in- tating each of the head-based basis vectors, b Å (eX , eY , eZ )creases in frequency. Vertical gains and phases for the LARP and RALP into an associated set of canal-based basis vectors, b * Å (ea ,animals were not dependent on stimulus frequency.
ep , el ) (Fig. 1, A and B) . A generalized rotation in termsof Euler angles is as follows
a LARP animal (M9356) is shown in Figs. 10 and 11. BothR Å RfRuRc Ålateral canals and the right anterior and left posterior canals
were blocked (Fig. 10, D–G) . The length of the plugged Fcos f cos u cos c 0 sin f sin c 0(cos f cos u cos c / sin f cos c) cos f sin usin f cos u cos c / cos f sin c 0(sin f cos u sin c 0 cos f cos c) sin f sin u
0sin u cos c sin u sin c cos uGarea was estimated as 2–3 mm. The ampullae of the six
canals, including the hair cells and crista and the four macu-lae of the otolith organs, as well as the organ of Corti were
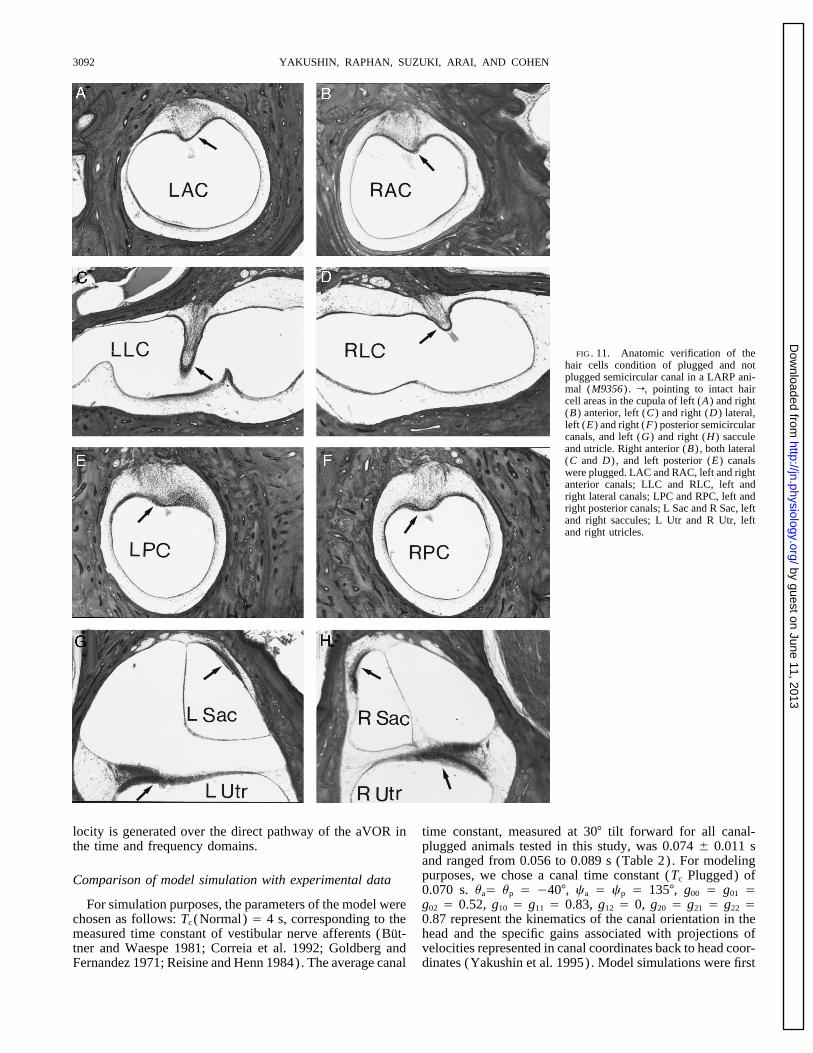
(6)intact (Fig. 11). In another LARP animal (M9006) , bothhorizontal, and the right anterior and left posterior canals Where f, u, and c are the Euler angles representing rota-were completely plugged, and the left anterior and right tions about the head yaw (Z) axis, the rotated Y axis ( lineposterior canals were patent (not shown). The hair cells of of nodes) , and the rotated yaw axis (Z*) . Each normal unitthe six ampullae and the four maculae and the organ of Corti vector has its own set of Euler angles. The Euler anglesappeared intact in this animal. The nerves and ganglion cells, associated with the anterior canal normal correspond to aand the dark cell area were normal. The otolith membranes rotation of the eX unit vector and are fa , ua , and ca . Thosewere not smooth, but the sensory cells looked intact. Plug- associated with the posterior canal correspond to a rotationging was also complete in M9003 (VC animal) and M9008 of the eY unit vector and are given by fp , up , and cp . Those(LC animal) . Similar results using the same techniques for associated with the lateral canal normal is a rotation of thecanal plugging in the monkey (Suzuki et al. 1991) pre- eZ vector given by the angles fl , ul , and cl . The canal basisviously have been reported by Angelaki et al. (1996). vectors can then be given in terms of the head basis vectors
Morphological data for other animals are not available. by the followingHowever, the reduction of the time constants during step
ea Å (cos fa cos ua cos ca 0 sin fa sin ca ) ex / (sin fa cos ua cos carotation in the plane of the plugged lateral canals, obtainedfrom animals with morphologically proved plugging, was / cos fa sin ca ) ey 0 (sin ua cos ca ) ez
similar to the reductions obtained from the other canal-ep Å 0(cos fp cos up cos cp / sin fp cos cp) explugged animals used in this study. Before plugging, the
time constants of the animals, tested with steps of velocity 0 (sin fp cos up sin cp 0 cos fp cos cp) ey / (sin up sin cp)ez
at 607 / s while upright, ranged from 20 to 70 s, and peake1 Å (cos fl sin ul ) ex / (sin fl sin ul ) ey / (cos ul ) ez (7)
velocities wereÇ57–607 / s. After surgery, when the animalswere rotated in the plane of the plugged canals with steps Because each basis vector in the canal basis is a rotation of
a unit vector in the head basis, only two generalized coordi-of velocity, there was a weak transient response with a short
J1032-7/ 9k2e$$no07 12-09-98 07:00:21 neupa LP-Neurophys
by guest on June 11, 2013http://jn.physiology.org/
Dow
nloaded from

ANGULAR VOR AFTER CANAL PLUGGING 3089
FIG. 9. Horizontal (C and D) eye velocity during constant velocity rotation to the right ( left) and to the left (right) whenanimal (M9308) was tested in 307 tilt forward position before (C) and after plugging all 6 canal (D) . Each graph representsthe superposition of several eye velocity responses. A represent velocities and B represent acceleration of the head rotation.
nates are necessary to define the rotation of one unit vector The inverse of the above matrix, describes how the basisvectors eX , eY , and eZ can be given in terms of ea , ep , andinto another. Therefore without loss of generality, the trans-
formation between basis vector can be reduced to the follow- el , and is given bying by setting fa Å fp Å cl Å 0 eX Å Aea / Bep / Cel
ea Å cos ua cos caex / sin caey 0 sin ua cos caezeY Å Dea / Eep / Fel
ep Å 0cos up cos cp / cos cpey / sin up sin cpez
eZ Å Gea / Hep / Iel (10)el Å cos fl sin ulex / sin fl sin ul / cos ulez (8)
The relationship described by Eq. 8 defines the matrix of where T01can is given by
the transformation, Tcan , that project vectors given in headcoordinates along the normals of the canals, which represent T01
can Å S A B CD E FG H I
D (11)the canal coordinate frame. The matrix Tcan is given as fol-lows
Using the inverse matrix, a projection mapping from canalTcan Å S cos ua cos ca sin ca 0sin ua cos ca
0cos up cos cp cos cp sin up sin cp
cos fl sin ul sin fl sin ul cos ul
D (9) coordinates back to head coordinates, Thead can be obtainedas
J1032-7/ 9k2e$$no07 12-09-98 07:00:21 neupa LP-Neurophys
by guest on June 11, 2013http://jn.physiology.org/
Dow
nloaded from
YAKUSHIN, RAPHAN, SUZUKI, ARAI, AND COHEN3090
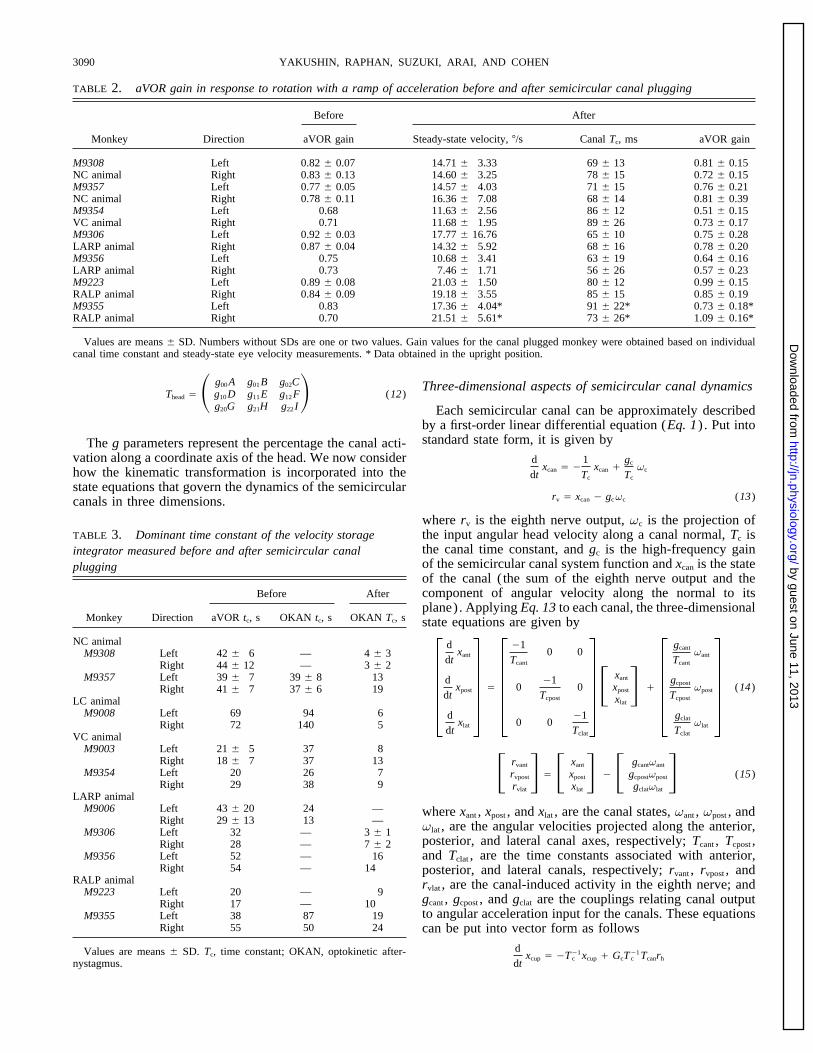
TABLE 2. aVOR gain in response to rotation with a ramp of acceleration before and after semicircular canal plugging
Before After
Monkey Direction aVOR gain Steady-state velocity, 7/s Canal Tc, ms aVOR gain
M9308 Left 0.82 { 0.07 14.71 { 3.33 69 { 13 0.81 { 0.15NC animal Right 0.83 { 0.13 14.60 { 3.25 78 { 15 0.72 { 0.15M9357 Left 0.77 { 0.05 14.57 { 4.03 71 { 15 0.76 { 0.21NC animal Right 0.78 { 0.11 16.36 { 7.08 68 { 14 0.81 { 0.39M9354 Left 0.68 11.63 { 2.56 86 { 12 0.51 { 0.15VC animal Right 0.71 11.68 { 1.95 89 { 26 0.73 { 0.17M9306 Left 0.92 { 0.03 17.77 { 16.76 65 { 10 0.75 { 0.28LARP animal Right 0.87 { 0.04 14.32 { 5.92 68 { 16 0.78 { 0.20M9356 Left 0.75 10.68 { 3.41 63 { 19 0.64 { 0.16LARP animal Right 0.73 7.46 { 1.71 56 { 26 0.57 { 0.23M9223 Left 0.89 { 0.08 21.03 { 1.50 80 { 12 0.99 { 0.15RALP animal Right 0.84 { 0.09 19.18 { 3.55 85 { 15 0.85 { 0.19M9355 Left 0.83 17.36 { 4.04* 91 { 22* 0.73 { 0.18*RALP animal Right 0.70 21.51 { 5.61* 73 { 26* 1.09 { 0.16*
Values are means { SD. Numbers without SDs are one or two values. Gain values for the canal plugged monkey were obtained based on individualcanal time constant and steady-state eye velocity measurements. * Data obtained in the upright position.
Three-dimensional aspects of semicircular canal dynamicsThead Å S g00A g01B g02C
g10D g11E g12Fg20G g21H g22I
D (12)
Each semicircular canal can be approximately describedby a first-order linear differential equation (Eq. 1) . Put intostandard state form, it is given byThe g parameters represent the percentage the canal acti-
vation along a coordinate axis of the head. We now consider ddt
xcan Å 01Tc
xcan /gc
Tc
vchow the kinematic transformation is incorporated into thestate equations that govern the dynamics of the semicircular
rv Å xcan 0 gcvc (13)canals in three dimensions.where rv is the eighth nerve output, vc is the projection ofthe input angular head velocity along a canal normal, Tc isTABLE 3. Dominant time constant of the velocity storagethe canal time constant, and gc is the high-frequency gainintegrator measured before and after semicircular canalof the semicircular canal system function and xcan is the statepluggingof the canal ( the sum of the eighth nerve output and the
Before After component of angular velocity along the normal to itsplane) . Applying Eq. 13 to each canal, the three-dimensional
Monkey Direction aVOR tc, s OKAN tc, s OKAN Tc, s state equations are given byNC animal
M9308 Left 42 { 6 — 4 { 3Right 44 { 12 — 3 { 2
M9357 Left 39 { 7 39 { 8 13
ddt
xant
ddt
xpost
ddt
xlat
Å
01Tcant
0 0
001
Tcpost
0
0 001Tclat
F xant
xpost
xlat
G /
gcant
Tcant
vant
gcpost
Tcpost
vpost
gclat
Tclat
vlat
(14)Right 41 { 7 37 { 6 19LC animal
M9008 Left 69 94 6Right 72 140 5
VC animalM9003 Left 21 { 5 37 8
Right 18 { 7 37 13M9354 Left 20 26 7 F rvant
rvpost
rvlat
G Å F xant
xpost
xlat
G 0 F gcantvant
gcpostvpost
gclatvlat
G (15)Right 29 38 9
LARP animalM9006 Left 43 { 20 24 — where xant , xpost , and xlat , are the canal states, vant , vpost , and
Right 29 { 13 13 — vlat , are the angular velocities projected along the anterior,M9306 Left 32 — 3 { 1posterior, and lateral canal axes, respectively; Tcant , Tcpost ,Right 28 — 7 { 2
M9356 Left 52 — 16 and Tclat , are the time constants associated with anterior,Right 54 — 14 posterior, and lateral canals, respectively; rvant , rvpost , and
RALP animal rvlat , are the canal-induced activity in the eighth nerve; andM9223 Left 20 — 9
gcant , gcpost , and gclat are the couplings relating canal outputRight 17 — 10to angular acceleration input for the canals. These equationsM9355 Left 38 87 19
Right 55 50 24 can be put into vector form as follows
Values are means { SD. Tc, time constant; OKAN, optokinetic after- ddt
xcup Å 0T01c xcup / GcT01
c Tcanrhnystagmus.
J1032-7/ 9k2e$$no07 12-09-98 07:00:21 neupa LP-Neurophys
by guest on June 11, 2013http://jn.physiology.org/
Dow
nloaded from
ANGULAR VOR AFTER CANAL PLUGGING 3091
FIG. 10. Anatomic verification of semicircular canalplugging in a LARP animal (M9356) . A–C , left : sec-tions of the temporal bones with plugged semicircularcanals. Right : detail of the plugged right anterior (D) ,right lateral (E) , left lateral (F) , and left posterior (G)canals. Triangles in D–G are marking to the borders ofthe plugged semicircular canals. RAC, right anteriorcanal; RLC, right lateral canal; LLC, left lateral canal;LPC, left posterior canal.
rv Å xcup 0 GcTcanrh It should be noted that matrices Tc and Gc represent the timeconstant of individual canals and their coupling from angularve Å Theadrv (16)acceleration along their normals, respectively. These matri-ces are diagonal because it is assumed that there is no crosswherecoupling between canals at the peripheral level.
The three-dimensional transfer function H( j v) can beTc Å F Tcant 0 0
0 Tcpost 00 0 Tclat
G Gc Å F gcant 0 00 gcpost 00 0 gclat
G (17) obtained by substituting j v for the derivative operator inEq. 16 and solving for the Fourier transform of We in termsof the Fourier transform of Rh
and Tcan and Thead are the projection mappings from head-We Å Thead[ ( j vI / T01
c )01GcT01c Tcan 0 GcTcan ]Rh (19)to-canal and canal-to-head coordinates, respectively. The
vectors rh , rv , xcup , and ve are the head velocities, VIII nerve Therefore the transfer function H( j v) can be given in aactivation, and cupula state given by simplified form as
H( j v) Å Thead [( j vTc / I)01 0 I]GcTcan (20)rh Å S vant
vpost
vlat
D rv Å S rvant
rvpost
rvlat
D xcup Å S xant
xpost
xlat
D (18)
Thus Eqs. 16 and 20 model how three-dimensional eye ve-
J1032-7/ 9k2e$$no07 12-09-98 07:00:21 neupa LP-Neurophys
by guest on June 11, 2013http://jn.physiology.org/
Dow
nloaded from
YAKUSHIN, RAPHAN, SUZUKI, ARAI, AND COHEN3092
FIG. 11. Anatomic verification of thehair cells condition of plugged and notplugged semicircular canal in a LARP ani-mal (M9356) . r, pointing to intact haircell areas in the cupula of left (A) and right(B) anterior, left (C) and right (D) lateral,left (E) and right (F) posterior semicircularcanals, and left (G) and right (H) sacculeand utricle. Right anterior (B) , both lateral(C and D) , and left posterior (E) canalswere plugged. LAC and RAC, left and rightanterior canals; LLC and RLC, left andright lateral canals; LPC and RPC, left andright posterior canals; L Sac and R Sac, leftand right saccules; L Utr and R Utr, leftand right utricles.
locity is generated over the direct pathway of the aVOR in time constant, measured at 307 tilt forward for all canal-plugged animals tested in this study, was 0.074 { 0.011 sthe time and frequency domains.and ranged from 0.056 to 0.089 s (Table 2). For modelingpurposes, we chose a canal time constant (Tc Plugged) ofComparison of model simulation with experimental data0.070 s. uaÅ up Å 0407, ca Å cp Å 1357, g00 Å g01 Åg02 Å 0.52, g10 Å g11 Å 0.83, g12 Å 0, g20 Å g21 Å g22 ÅFor simulation purposes, the parameters of the model were
chosen as follows: Tc (Normal) Å 4 s, corresponding to the 0.87 represent the kinematics of the canal orientation in thehead and the specific gains associated with projections ofmeasured time constant of vestibular nerve afferents (But-
tner and Waespe 1981; Correia et al. 1992; Goldberg and velocities represented in canal coordinates back to head coor-dinates (Yakushin et al. 1995). Model simulations were firstFernandez 1971; Reisine and Henn 1984). The average canal
J1032-7/ 9k2e$$no07 12-09-98 07:00:21 neupa LP-Neurophys
by guest on June 11, 2013http://jn.physiology.org/
Dow
nloaded from
ANGULAR VOR AFTER CANAL PLUGGING 3093
13, C, D, G, and H) and fit the data from the VC animal(Fig. 4, C and F) . For both the curves could be considereda combination of the responses of the normal and the NCanimal. At 407 tilt forward, the intact vertical canals arenot activated during rotation for the VC, LARP, and RALPanimals. Therefore their responses should have the samevariation of temporal phases as a function of frequency asthe NC animal. This can be seen by examining the temporalphases for the different frequencies along the dotted lines inFig. 13, B–D and F–H) . When the animals were tiltedbackward so that one or both pairs of reciprocal intact verti-cal canals were close to the plane of rotation, the temporalphases of the VC and LARP animals converged to be approx-imately equal to01807, close to values predicted for animalswith normal labyrinths. Thus changing the dominant timeconstant of the canal from 4.0 s (normal) to 0.070 s (canal-plugged condition) could explain the observed phase varia-tions for the canal-plugged animals used in this study at allapplied frequencies.
The model predicted that the temporal gains for the yawand roll components would vary sinusoidally as a function
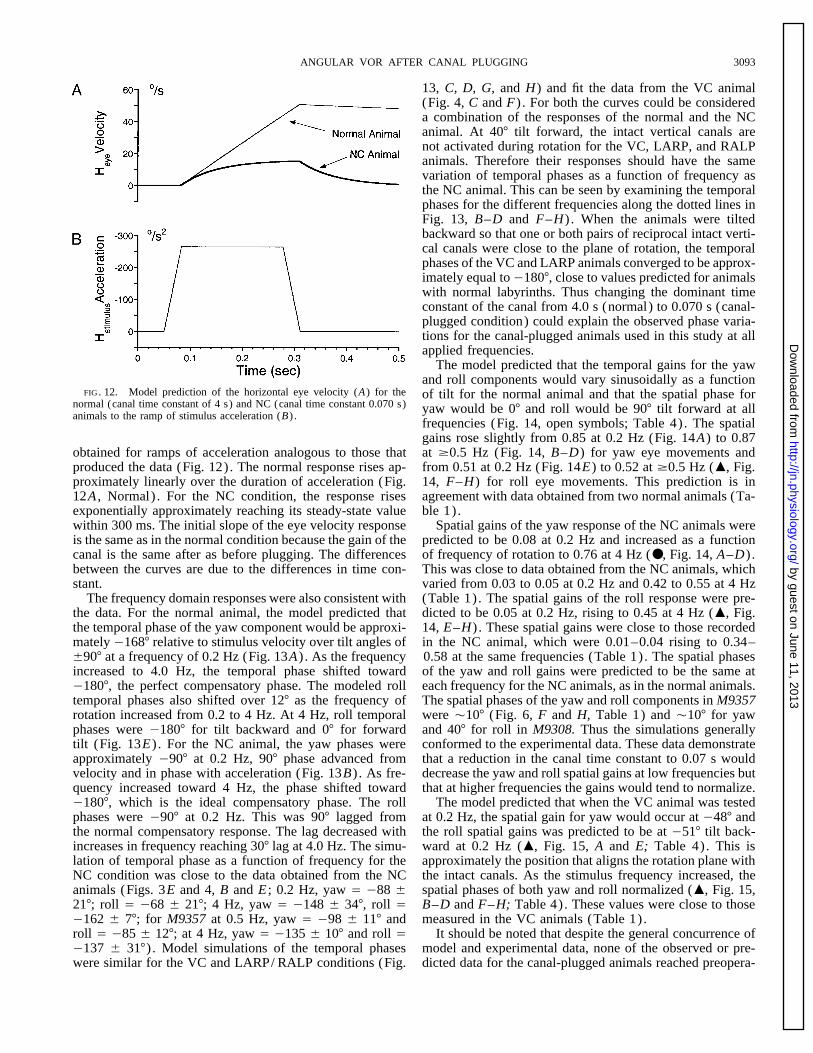
FIG. 12. Model prediction of the horizontal eye velocity (A) for the of tilt for the normal animal and that the spatial phase fornormal (canal time constant of 4 s) and NC (canal time constant 0.070 s) yaw would be 07 and roll would be 907 tilt forward at allanimals to the ramp of stimulus acceleration (B) .
frequencies (Fig. 14, open symbols; Table 4). The spatialgains rose slightly from 0.85 at 0.2 Hz (Fig. 14A) to 0.87at ¢0.5 Hz (Fig. 14, B–D) for yaw eye movements andobtained for ramps of acceleration analogous to those that
produced the data (Fig. 12). The normal response rises ap- from 0.51 at 0.2 Hz (Fig. 14E) to 0.52 at ¢0.5 Hz (m, Fig.14, F–H) for roll eye movements. This prediction is inproximately linearly over the duration of acceleration (Fig.
12A , Normal) . For the NC condition, the response rises agreement with data obtained from two normal animals (Ta-ble 1).exponentially approximately reaching its steady-state value
within 300 ms. The initial slope of the eye velocity response Spatial gains of the yaw response of the NC animals werepredicted to be 0.08 at 0.2 Hz and increased as a functionis the same as in the normal condition because the gain of the
canal is the same after as before plugging. The differences of frequency of rotation to 0.76 at 4 Hz (●, Fig. 14, A–D) .This was close to data obtained from the NC animals, whichbetween the curves are due to the differences in time con-
stant. varied from 0.03 to 0.05 at 0.2 Hz and 0.42 to 0.55 at 4 Hz(Table 1). The spatial gains of the roll response were pre-The frequency domain responses were also consistent with
the data. For the normal animal, the model predicted that dicted to be 0.05 at 0.2 Hz, rising to 0.45 at 4 Hz (m, Fig.14, E–H) . These spatial gains were close to those recordedthe temporal phase of the yaw component would be approxi-
mately 01687 relative to stimulus velocity over tilt angles of in the NC animal, which were 0.01–0.04 rising to 0.34–0.58 at the same frequencies (Table 1). The spatial phases{907 at a frequency of 0.2 Hz (Fig. 13A) . As the frequency
increased to 4.0 Hz, the temporal phase shifted toward of the yaw and roll gains were predicted to be the same ateach frequency for the NC animals, as in the normal animals.01807, the perfect compensatory phase. The modeled roll
temporal phases also shifted over 127 as the frequency of The spatial phases of the yaw and roll components in M9357were Ç107 (Fig. 6, F and H, Table 1) and Ç107 for yawrotation increased from 0.2 to 4 Hz. At 4 Hz, roll temporal
phases were 01807 for tilt backward and 07 for forward and 407 for roll in M9308. Thus the simulations generallyconformed to the experimental data. These data demonstratetilt (Fig. 13E) . For the NC animal, the yaw phases were
approximately 0907 at 0.2 Hz, 907 phase advanced from that a reduction in the canal time constant to 0.07 s woulddecrease the yaw and roll spatial gains at low frequencies butvelocity and in phase with acceleration (Fig. 13B) . As fre-
quency increased toward 4 Hz, the phase shifted toward that at higher frequencies the gains would tend to normalize.The model predicted that when the VC animal was tested01807, which is the ideal compensatory phase. The roll
phases were 0907 at 0.2 Hz. This was 907 lagged from at 0.2 Hz, the spatial gain for yaw would occur at 0487 andthe roll spatial gains was predicted to be at 0517 tilt back-the normal compensatory response. The lag decreased with
increases in frequency reaching 307 lag at 4.0 Hz. The simu- ward at 0.2 Hz (m, Fig. 15, A and E; Table 4). This isapproximately the position that aligns the rotation plane withlation of temporal phase as a function of frequency for the
NC condition was close to the data obtained from the NC the intact canals. As the stimulus frequency increased, thespatial phases of both yaw and roll normalized (m, Fig. 15,animals (Figs. 3E and 4, B and E ; 0.2 Hz, yaw Å 088 {
217; roll Å 068 { 217; 4 Hz, yaw Å 0148 { 347, roll Å B–D and F–H; Table 4). These values were close to thosemeasured in the VC animals (Table 1).0162 { 77; for M9357 at 0.5 Hz, yaw Å 098 { 117 and
roll Å 085 { 127; at 4 Hz, yaw Å 0135 { 107 and roll Å It should be noted that despite the general concurrence ofmodel and experimental data, none of the observed or pre-0137 { 317) . Model simulations of the temporal phases
were similar for the VC and LARP/ RALP conditions (Fig. dicted data for the canal-plugged animals reached preopera-
J1032-7/ 9k2e$$no07 12-09-98 07:00:21 neupa LP-Neurophys
by guest on June 11, 2013http://jn.physiology.org/
Dow
nloaded from
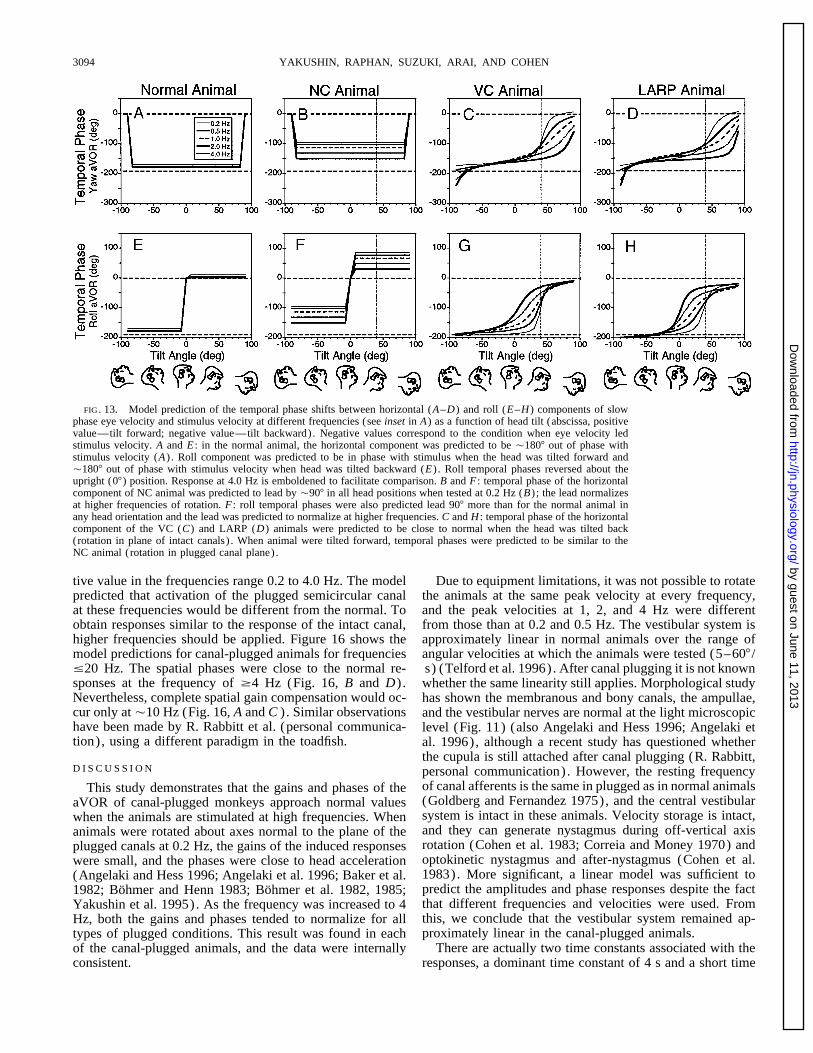
YAKUSHIN, RAPHAN, SUZUKI, ARAI, AND COHEN3094
FIG. 13. Model prediction of the temporal phase shifts between horizontal (A–D) and roll (E–H) components of slowphase eye velocity and stimulus velocity at different frequencies (see inset in A) as a function of head tilt (abscissa, positivevalue—tilt forward; negative value—tilt backward). Negative values correspond to the condition when eye velocity ledstimulus velocity. A and E : in the normal animal, the horizontal component was predicted to be Ç1807 out of phase withstimulus velocity (A) . Roll component was predicted to be in phase with stimulus when the head was tilted forward andÇ1807 out of phase with stimulus velocity when head was tilted backward (E) . Roll temporal phases reversed about theupright (07) position. Response at 4.0 Hz is emboldened to facilitate comparison. B and F : temporal phase of the horizontalcomponent of NC animal was predicted to lead by Ç907 in all head positions when tested at 0.2 Hz (B) ; the lead normalizesat higher frequencies of rotation. F : roll temporal phases were also predicted lead 907 more than for the normal animal inany head orientation and the lead was predicted to normalize at higher frequencies. C and H : temporal phase of the horizontalcomponent of the VC (C) and LARP (D) animals were predicted to be close to normal when the head was tilted back(rotation in plane of intact canals) . When animal were tilted forward, temporal phases were predicted to be similar to theNC animal (rotation in plugged canal plane).
tive value in the frequencies range 0.2 to 4.0 Hz. The model Due to equipment limitations, it was not possible to rotatethe animals at the same peak velocity at every frequency,predicted that activation of the plugged semicircular canal
at these frequencies would be different from the normal. To and the peak velocities at 1, 2, and 4 Hz were differentfrom those than at 0.2 and 0.5 Hz. The vestibular system isobtain responses similar to the response of the intact canal,
higher frequencies should be applied. Figure 16 shows the approximately linear in normal animals over the range ofangular velocities at which the animals were tested (5–607 /model predictions for canal-plugged animals for frequencies
°20 Hz. The spatial phases were close to the normal re- s) (Telford et al. 1996). After canal plugging it is not knownsponses at the frequency of ¢4 Hz (Fig. 16, B and D) . whether the same linearity still applies. Morphological studyNevertheless, complete spatial gain compensation would oc- has shown the membranous and bony canals, the ampullae,cur only atÇ10 Hz (Fig. 16, A and C ) . Similar observations and the vestibular nerves are normal at the light microscopichave been made by R. Rabbitt et al. (personal communica- level (Fig. 11) (also Angelaki and Hess 1996; Angelaki ettion), using a different paradigm in the toadfish. al. 1996), although a recent study has questioned whether
the cupula is still attached after canal plugging (R. Rabbitt,D I S C U S S I O N personal communication). However, the resting frequency
of canal afferents is the same in plugged as in normal animalsThis study demonstrates that the gains and phases of the(Goldberg and Fernandez 1975), and the central vestibularaVOR of canal-plugged monkeys approach normal valuessystem is intact in these animals. Velocity storage is intact,when the animals are stimulated at high frequencies. Whenand they can generate nystagmus during off-vertical axisanimals were rotated about axes normal to the plane of therotation (Cohen et al. 1983; Correia and Money 1970) andplugged canals at 0.2 Hz, the gains of the induced responsesoptokinetic nystagmus and after-nystagmus (Cohen et al.were small, and the phases were close to head acceleration1983). More significant, a linear model was sufficient to(Angelaki and Hess 1996; Angelaki et al. 1996; Baker et al.predict the amplitudes and phase responses despite the fact1982; Bohmer and Henn 1983; Bohmer et al. 1982, 1985;that different frequencies and velocities were used. FromYakushin et al. 1995). As the frequency was increased to 4this, we conclude that the vestibular system remained ap-Hz, both the gains and phases tended to normalize for allproximately linear in the canal-plugged animals.types of plugged conditions. This result was found in each
There are actually two time constants associated with theof the canal-plugged animals, and the data were internallyconsistent. responses, a dominant time constant of 4 s and a short time
J1032-7/ 9k2e$$no07 12-09-98 07:00:21 neupa LP-Neurophys
by guest on June 11, 2013http://jn.physiology.org/
Dow
nloaded from

ANGULAR VOR AFTER CANAL PLUGGING 3095
FIG. 14. Model prediction of the horizontal (A–D) and roll (E–H) spatial gains for the normal (s and n) and NC (●
and m) animals tested at 0.2 Hz (A and E) , 1.0 Hz (B and F) , 2.0 Hz (C and G) , and 4.0 Hz (D and H) . Compare withFigs. 6 and 8.
constant estimated to be Ç3 ms (Steinhausen 1933). The dominant time constant to decrease but would cause thesmall time constant to increase. This would produce a lowerdominant time constant is proportional to the viscosity of
the endolymph and inversely proportional to the moment of gain at higher frequencies. However, our results show amonotonic increase in gain with frequency corresponding toinertia. The short time constant is proportional to the viscos-
ity and inversely proportional to the elasticity of the cupula a 50-fold reduction in dominant time constant. An equivalent50-fold increase in the small time constant would cause amembrane. Either an increase in viscosity or a decrease in
the moment of inertia could decrease the canal time constant gain reduction at 1 Hz, which was not observed. Therefore,it is unlikely that the short time constant of the canals wasafter plugging. An increase in the viscosity would cause thechanged. This suggests that the viscosity of the endolymphwas unaltered before and after plugging.TABLE 4. Model prediction of the spatial gain and phases of
There are two techniques that have been described forthe horizontal and roll aVOR componentsplugging the canals. Ewald’s original technique was to openthe perilymphatic space and compress the membranous canalLARP
Stimulus and with gutta percha, a clay-like material. The Money/ Scott /Frequency, Normal NC VC RALP Suzuki technique, which was used in these experiments
Hz Animal Animal Animal Animal (Money and Scott 1962; Suzuki et al. 1991) entails drillingacross the bony and membranous canals and filling the open-Horizontaling with bone chips and soft tissue. Initially after surgerySpatial gain 0.2 0.85 0.08 0.44 0.23
0.5 0.87 0.19 0.47 0.28 there probably is mixing of endolymph and perilymph with1.0 0.87 0.35 0.53 0.40 this plugging technique. The cupula also may be discon-2.0 0.87 0.58 0.66 0.60 nected from the roof of the ampulla (R. Rabbitt, personal4.0 0.87 0.76 0.79 0.76
communication), which may account for the decreased re-Spatial phase 0.2 0 0 048 0450.5 0 0 045 035 sponses seen immediately after surgery (Angelaki and Hess1.0 0 0 032 018 1996; Angelaki et al. 1996). However, in the long term, it2.0 0 0 014 05 is likely that the endolymph and perilymph compartments4.0 0 0 03 01
are separated and that the cupula reattaches. Moreover, aRollmixture of endolymph and perilymph would have a de-Spatial gain 0.2 0.51 0.05 0.45 0.23
0.5 0.52 0.11 0.46 0.25 creased not an increased viscosity, which would produce a1.0 0.52 0.21 0.46 0.29 longer not a shorter time constant. It is more likely that the2.0 0.52 0.34 0.48 0.38 canal plug had reduced the effective moment of inertia in4.0 0.52 0.45 0.50 0.47
response to angular acceleration of the head, although theSpatial phase 0.2 090 090 051 052exact nature of the mechanism is not clear.0.5 090 090 051 057
1.0 090 090 057 068 There has been controversy as to whether there is spatial2.0 090 090 071 082 adaptation of the response of the remaining canals after canal4.0 090 090 082 088 plugging. Yakushin et al. (1995) tested animals at 0.2 Hz
J1032-7/ 9k2e$$no07 12-09-98 07:00:21 neupa LP-Neurophys
by guest on June 11, 2013http://jn.physiology.org/
Dow
nloaded from

YAKUSHIN, RAPHAN, SUZUKI, ARAI, AND COHEN3096
FIG. 15. Model prediction of the horizontal (A–D) and roll (E–H) spatial gains for the normal (s) , VC (m) , and RALP(●) animals tested at 0.2 Hz (A and E) , 1.0 Hz (B and F) , 2.0 Hz (C and G) , and 4.0 Hz (D and H) . Compare with Fig.10.
and concluded that such adaptation did not occur. Angelaki NC animals. At 4 Hz, the responses were close to being1807 out of phase or in phase with head velocity (Figs. 3,and Hess (1996a) reported similar changes in phase when
animals were tested between 0.05 and 1.1 Hz but concluded 4, and 6). The induced eye movements could not have arisenfrom spatial adaptation in the NC animals because therethat spatial adaptation had occurred in the planes of the intact
canals to account for the frequency-dependent responses in were no intact canals left to adapt. The responses in the NCanimals were similar to those of the other animals whenthe planes of the plugged canals (Angelaki and Hess 1996;
Angelaki et al. 1996). The responses of the NC animals rotated around axes orthogonal to the plugged canals. Be-cause the frequency response of the eye movements inducedwere critical in resolving this question. Independent of head
orientation to the axis of rotation, the temporal and spatial at 2–4 Hz is above the peak frequency of eye movementsproduced by activation of neck muscles in plugged canalgains and phases of the horizontal and roll eye velocity
components tended to normalize at higher frequencies in the animals (0.5 Hz) (Bohmer and Henn 1983), the most likelyalternate hypothesis is that the responses arose from theplugged canals.
In support of this alternative hypothesis, the responsesof each of the animals with various combinations of canalplugging formed a consistent set of data over a range offrequencies from 0.2 to 4.0 Hz. In every case, when theanimals were rotated sinusoidally in the plane of the pluggedcanals at lower frequencies, the response was in phase withhead acceleration. As the frequency of rotation increased, itmoved toward being in phase with head velocity. Conse-quently, when the animals were rotated sinusoidally in theplane of the plugged canals at lower frequencies, the gainwas close to zero, but it approached the normal responsegain and phase as the frequency of rotation increased. Gen-eral support for this alternative hypothesis came from thefinding that the spatial gains and phases for the horizontalcomponent could be predicted by combining responses ofthe plugged and the normal canals. This was simply doneusing a model, in which the gain of the aVOR was heldconstant, but the time constant of the canal was reduceddue to changes in the canal dynamics after plugging. Oneproblem, which arose in one of the NC animals (M9308) ,
FIG. 16. Model prediction for the spatial gains (A and C) and phaseswas that the spatial phase of the roll component remained(B and D) as a function of stimulus frequency for the normal animals and
after different canal plugging (see inset in A) . at 0457 (Fig. 6H) and did not approach the 0907 spatial
J1032-7/ 9k2e$$no07 12-09-98 07:00:21 neupa LP-Neurophys
by guest on June 11, 2013http://jn.physiology.org/
Dow
nloaded from

ANGULAR VOR AFTER CANAL PLUGGING 3097
phase of the normal for the frequency range °4 Hz (Fig. dance with Ewald’s Second Law (1892). A similar resultwas obtained from plugged canals, although there was a6D) . The reason for this is not apparent. The roll component
was consistent with model predictions in the other NC ani- significant difference between the canal-plugged and nerve-cut subjects (Aw et al. 1996b). Aw et al. used accelerationsmal (M9357) .
We considered the possibility that activity responsible for of É4,0007 / s2 , which last for É50 ms and then reverse forÉ100 ms, with decelerations being smaller than accelera-eye movements in the planes of the plugged canals may
have arisen in the otolith organs. Goldberg and Fernandez tions. This gives peak head velocities during the accelerationphase of 200–2507 / s. If the time constant of the plugged(1975) demonstrated during centered or on-axis rotation that
‘‘otolith neurons do not respond to even intense angular canal was the same as that we found in the monkey aftercanal plugging (0.07 s) , the model would give a gain closeaccelerations.’’ From this, it is unlikely that the activity
responsible for the eye velocities recorded in the canal to 1. This is because the time constant is larger than the timeof stimulation. However, if the time constant were only 30plugged animals came from the otolith organs. Recently it
has been suggested that movement of the endolymph on the ms in the canal-plugged condition, which is smaller than theperiod of stimulation, the model would give a peak responseutricular side of the cupula could provide a source for fluid
movement even in the presence of plugging (R. Rabbitt, ofõ1207 / s. This would be consistent with theÉ1207 / s thatAw et al. found in the plugged individuals. Correspondingly,personal communication). Our study, which suggests that
plugging does not inactivate the response of a canal but the frequency characteristics would be different. The cut-infrequency would be É6 Hz, as opposed to the 2- to 3-Hzmerely changes its dynamics so that the lower end of the
pass-band is moved to a higher frequency, are in agreement cut-in frequency we found in the monkey (100/ 0.03 ∗ 2p) .Thus the data of Aw et al. (1996a,b) may imply that inwith this suggestion.
It was especially striking that both the sinusoidal and step humans, the canal time constant after posterior canal plug-ging is shorter and the cut-in frequency is higher than in theresponse to short ramps of angular acceleration could be
predicted for canal-plugged and normal animals by modi- monkey. The practical implication is that the type of canalplugging used by Aw et al. (1996a,b) on humans renderedfying a single parameter, the time constant of the afferent
output of the semicircular canals. Normal canals were associ- the canal ineffective even at frequencies close to 20–30 Hz.The model predicted that there would be no changes inated with an afferent time constant of É4 s, while plugged
canals were associated with a time constant of 70 ms. The spatial gain and phase of the yaw and roll components as afunction of frequency in the normal animal. It also predictedreduced time constant would lead to a shift of the low-
frequency cutoff, i.e., the frequency at which the gain is that there would be an 117 temporal phase shift from perfectcompensation. These predictions were based on the assump-down to 0.707 (the 3-dB cutoff) , from 0.039 to 2.270 Hz.
As a result, canal plugging essentially has the effect of tion that there was a 4-s time constant of the canals andthat the contribution of velocity storage was negligible. Theblocking the output of the canals at lower frequencies (°0.2
Hz) while sustaining a normal response at higher frequencies response was limited solely to the direct vestibular pathway,which is predominantly active during frequencies of sinusoi-(¢4 Hz). The shortened time constant also would lead to
the prediction of a reduced steady-state gain for steps of dal rotation between 0.1 and 8 Hz. The gain predictions wererealized in the two normal monkeys used in this study butrotation lasting longer than three time constants (É200 ms).
These conclusions are consistent with studies that demon- none of the normal responses had the 117 phase shift at thelower frequencies. If the contribution of velocity storagestrate reduced eye velocities in response to step rotation in
canal plugged animals (Backous et al. 1996; Bohmer et al. was included, the difference between model predictions andexperimental data would have been reduced significantly.1982). Our interpretation of these results is that it was not
the gain that was reduced, but rather the time constant that This suggests that velocity storage not only contributes tothe aVOR in the low-frequency range (between 0.001 andhad been modified.
For normal animals, the semicircular canal afferents have 0.1) , but it also makes an important contribution to theresponse at midband frequencies (0.2–0.5 Hz) by producinga linear rise over the whole period of constant acceleration.
Thus the time constant can be neglected in calculations of phase lags that reduce the phase leads coming from thecanals. At low frequencies, there would be cross-couplingthe gain (Raphan and Cohen 1981, 1986). In contrast, as a
result of the short time constant in the canal-plugged animal, of velocity storage in tilted positions (Dai et al. 1991; Ra-phan and Sturm 1991; Raphan et al. 1992). This wouldthe increase in eye velocity during constant angular accelera-
tion has an exponential rise, which reaches a close-to-steady- superpose the spatial organization of velocity storage withthe spatial organization of the semicircular canals, therebystate level within the first 200–300 ms. Thus the canal time
constant is a critical factor in aVOR gain calculations (Ra- altering the overall spatial response of the system.It is of interest to speculate why the general motor behav-phan and Cohen 1986). As shown in this paper, if the time
constant is considered, the gain of the aVOR response to a ior of the animals with plugged canals recovered, even inthe NC animals. These monkeys could jump in their cagesstep of velocity is not different in the canal-plugged animals
from those of normal monkeys or from those of sinusoidal and maintain balance. Their only obvious behavioral abnor-mality was a tendency for the head to overshoot brieflyrotation at high frequencies.
High-frequency head rotation is of considerable clinical during gaze shifts in the planes of the plugged canals. Similarfindings were reported by Ewald (1892). He noted that pi-value in detecting abnormalities due to vestibular denerva-
tion (Aw et al. 1996a,b; Halmagyi and Curthoys 1988). geons with plugged canals could fly to the roof of the cageon the same day after plugging as soon as they had recoveredPeak eye velocities are lower and compensation is less when
the head is rotated toward a denervated labyrinth in accor- from anesthesia. Normally, there is dual compensation for
J1032-7/ 9k2e$$no07 12-09-98 07:00:21 neupa LP-Neurophys
by guest on June 11, 2013http://jn.physiology.org/
Dow
nloaded from

YAKUSHIN, RAPHAN, SUZUKI, ARAI, AND COHEN3098
vestibuloocular reflex in response to high-acceleration head rotations. I.head movement by both vision and vestibular input in theResponses in normal subjects. J. Neurophysiol. 76: 4009–4020, 1996b.range of 0.1–1 Hz. This redundancy provides a robustness
BACKOUS, D. D., LASKER, D., AND MINOR, L. B. Asymmetries in the hori-of compensation, which protects against individual variabil- zontal VOR after unilateral plugging of the three semicircular canals inity and effects of lesions. Plugging narrows this range of the squirrel monkey. Soc. Neurosci. Abstr. 22: 1094, 1996.
BAKER, J., GOLDBERG, J., PETERSON, B., AND SCHOR, R. Oculomotor re-overlap, and the canals are essentially ineffective in the rangeflexes after semicircular canal plugging in cats. Brain Res. 252: 151–of 0.1–1 Hz. As we have previously shown, vision substi-155, 1982.tutes for the aVOR during head movements at °0.2 Hz in BAKER, J. F., ISU, N., WICKLAND, C. R., AND PETERSON, B. W. Dynamics
canal-plugged animals (Yakushin et al. 1995). This study of adaptive change in vestibulo-ocular reflex directions. II. Sagittal planedemonstrates that vision also can contribute significantly to rotation. Brain Res. 371: 166–170, 1986.
BAKER, J. F. AND PETERSON, B. W. Excitation of the extraocular musclesnormalization of the aVOR in animals with plugged canalsin decerebrate cats during the vestibulo-ocular reflex in three-dimensionalup to Ç1 Hz. It is at this critical frequency and above that,space. Exp. Brain Res. 84: 266–278, 1991.the aVOR becomes significantly active. Therefore with a BAKER, J., WICKLAND, C., AND PETERSON, B. Dependence of cat vestibulo-
mixture of vision and input from the plugged canals, the ocular reflex direction adaptation on animal orientation during adaptationand rotation in darkness. Brain Res. 408: 339–343, 1987.animals could generate compensatory eye velocity against
BALOH, R. W., HESS, K., HONRUBIA, V., AND YEE, R. D. Low and highhead velocity over a full range of head movements despitefrequency sinusoidal rotational testing in patients with peripheral vestibu-the canal plugging.lar lesions. Acta Otolaryngol. Suppl. (Stockh.) 406: 189–193, 1984.
The reduction in the dominant time constant of the aVOR BLANKS, R.H.I., CURTHOYS, I. S., BENNET, M., AND MARKHAM, C. H. Planarand OKAN after canal plugging has not been noted before. relationships of the semicircular canals in rhesus and squirrel monkeys.
Brain Res. 340: 315–324, 1985.Presumably, it represents habituation of velocity storageBLANKS, R.H.I., CURTHOYS, I. S., AND MARKHAM, C. H. Planar relationshipsafter canal plugging (Cohen et al. 1992b). This habituation
of the semicircular canals in man. Acta Otolaryngol. 80: 185–196, 1975.could account for theÉ50% drop in steady state eye velocity BOHMER, A. AND HENN, V. Horizontal and vertical vestibulo-ocular andnoted during off-vertical axis rotation (OVAR) after canal cervico-ocular reflexes in the monkey during high frequency rotation.
Brain Res. 277: 241–248, 1983.plugging (Cohen et al. 1983; Correia and Money 1968).BOHMER, A., HENN, V., AND SUZUKI, J.-I. Compensatory eye movementsThe reason for this habituation is not known.
in the monkey during high frequency sinusoidal rotations. In: Physiologi-In summary, the data support the hypothesis that pluggingcal and Pathological Aspects of Eye Movements , edited by A. Roucoux.
at a point distant from the cupula may alter but it does not London: Junk, 1982, p. 127–130.block the response of the semicircular canals. Rather it alters BOHMER, A., HENN, V., AND SUZUKI, J.-I. Vestibulo-ocular reflex after
selective plugging of the semicircular canals in the monkey—responsethe dynamic response so that the canals contribute differentlyplane determinations. Brain Res. 326: 291–298, 1985.at different frequencies. The data support the hypothesis that
BUTTNER, U. AND WAESPE, W. Vestibular nerve activity in the alert monkeythe individual canal pairs contribute eye velocity compo- during vestibular and optokinetic nystagmus. Exp. Brain Res. 41: 310–nents according to their vector projection onto the individual 315, 1981.canals, even after the canals are plugged. They suggest that COHEN, B., KOZLOVSKAYA, I. B., RAPHAN, T., SOLOMON, D., HELWIG, D.,
COHEN, N., SIROTA, M. G., AND YAKUSHIN, S. B. Vestibuloocular reflexdespite considerable plasticity in the aVOR, the innate coor-of rhesus monkeys after spaceflight. J. Appl. Physiol. 73: 121S–131S,dinate frame with regard to the head is not altered in adult1992a.
animals by partial inactivation of the individual canals over COHEN, B., SUZUKI, J.-I., AND BENDER, M. B. Eye movements from semicir-a wide range of frequencies. cular canal nerve stimulation in the cat. Ann. Otol. Rhinol. Laryngol. 73:
153–169, 1964.COHEN, B., SUZUKI, J., AND BENDER, M. B. Nystagmus induced by electricWe thank Dr. Minjia Dai for help in data collection and for the use of
stimulation of ampullary nerves. Acta Otolaryngol. 60: 422–436, 1965.data acquisition programs, Dr. Evgeny Buharin for writing the programsCOHEN, B., SUZUKI, J.-I., AND RAPHAN, T. Role of the otolith organs infor data analysis, M. Kunin for implementing the equations for the model
generation of horizontal nystagmus: effects of selective labyrinthine le-dynamics, and V. Rodriguez for technical assistance.sions. Brain Res. 276: 159–164, 1983.This work was supported by National Institutes of Health Grants NS-
COHEN, H., COHEN, B., RAPHAN, T., AND WAESPE, W. Habituation and00294, EY-11812, EY-04148, EY-01867, and DC-03787.adaptation of the vestibulo-ocular reflex: a model of differential controlAddress for reprint requests: S. B. Yakushin, Dept. of Neurology, Boxby the vestibulo-cerebellum. Exp. Brain Res. 90: 526–538, 1992b.1135, Mount Sinai School of Medicine, 1 E. 100th St., New York, NY
CORREIA, M. J. AND MONEY, K. E. The effect of blockage of all six semi-10029.circular canal ducts on nystagmus produced by dynamic linear accelera-
Received 17 December 1997; accepted in final form 13 July 1998. tion in the cat. Acta Otolaryngol. 69: 7–16, 1970.CORREIA, M. J., PERACHIO, A. A., DICKMAN, J. D., KOZLOVSKAYA, I. B.,
SIROTA, M. G., YAKUSHIN, S. B., AND BELOOZEROVA, I. N. Changes inREFERENCESmonkey horizontal semicircular canal afferent responses after spaceflight.J. Appl. Physiol. 73: 112S–120S, 1992.ANGELAKI, D. E. AND HESS, B.J.M. Adaptation of primate vestibular reflex
CRAWFORD, J. D. AND VILIS, T. Axes of eye rotation and Listing’s lawto altered peripheral vestibular inputs. II. Spatiotemporal properties ofduring rotations of the head. J. Neurophysiol. 65: 407–423, 1991.the adapted slow-phase eye velocity. J. Neurophysiol. 76: 2954–2971,
CURTHOYS, I. S., BLANKS, R.H.I., AND MARKHAM, C. H. Semicircular canal1996.functional anatomy in cat, guinea pig and man. Acta Otolaryngol. 83:ANGELAKI, D. E., HESS, B.J.M., ARAI, Y., AND SUZUKI, J.-I. Adaptation258–265, 1977.of primate vestibular reflex to altered peripheral vestibular inputs. I.
DAI, M., MCGARVIE, L., KOZLOVSKAYA, I., RAPHAN, T., AND COHEN, B.Frequency-specific recovery of horizontal VOR after inactivation of theEffects of spaceflight on ocular counterrolling and spatial orientation oflateral semicircular canals. J. Neurophysiol. 76: 2941–2953, 1996.the vestibular system. Exp. Brain Res. 102: 45–56, 1994.AW, S. T., HALMAGYI, G. M., HASLWANTER, T., CURTHOYS, I. S., YAVOR,
DAI, M. J., RAPHAN, T., COHEN, B., AND SCHNABOLK, C. Spatial orientationR. A., AND TODD, M. J. Three-dimensional vector analysis of the humanof velocity storage during post-rotatory nystagmus. Soc. Neurosci. Abstr.vestibuloocular reflex in response to high-acceleration head rotations.17: 314, 1991.II. Responses in subjects with unilateral vestibular loss and selective
DICKMAN, J. D. Spatial orientation of semicircular canals and afferent sensi-semicircular canal occlusion. J. Neurophysiol. 76: 4021–4030, 1996a.tivity vectors in pigeons. Exp. Brain Res. 111: 8–20, 1996.AW, S. T., HASLWANTER, T., HALMAGYI, G. M., CURTHOYS, I. S., YAVOR,
R. A., AND TODD, M. J. Three-dimensional vector analysis of the human ENGELKEN, E. J. AND STEVENS, K. W. A new approach to the analysis of
J1032-7/ 9k2e$$no07 12-09-98 07:00:21 neupa LP-Neurophys
by guest on June 11, 2013http://jn.physiology.org/
Dow
nloaded from

ANGULAR VOR AFTER CANAL PLUGGING 3099
nystagmus: an application for order-statistic filters. Aviat. Space Environ. System , edited by E. L. Keller and D. S. Zee. New York: Pergamon, vol.57, 1986, p. 285–292.Med. 61: 859–864, 1990.
RAPHAN, T., DAI, M., AND COHEN, B. Spatial orientation of the vestibularENGELKEN, E. J., STEVENS, K. W., MCQUEEN, W. J., AND ENDERLE, J. D.system. Ann. NY Acad. Sci. 656: 140–157, 1992.Application of robust data processing methods to he analysis of eye
RAPHAN, T., MATSUO, V., AND COHEN, B. Velocity storage in the vestibulo-movements. Biomed. Sci. Instrum. 32: 7–11, 1996.ocular reflex (VOR). Exp. Brain Res. 35: 229–248, 1979.EWALD, J. R. Physiologische Untersuchungen uber das Endorgan des Ner-
RAPHAN, T. AND STURM, D. Modelling the spatio-temporal organization ofvus Octavus. Weisbaden: 1892.velocity storage in the vestibulo-ocular reflex (VOR) by optokineticFERNANDEZ, C. AND GOLDBERG, J. M. Physiology of peripheral neuronsstudies. J. Neurophysiol. 66: 1410–1421, 1991.innervating semicircular canals of the squirrel monkey. II. Response to
REISINE, H. AND HENN, V. Transformation in three dimensions betweensinusoidal stimulation and dynamics of peripheral vestibular system. J.primary and secondary vestibular neuronal signals in the rhesus monkey.Neurophysiol. 34: 661–675, 1971.Soc. Neurosci. Abstr. 10: 49.7, 1984.FERNANDEZ, C. AND GOLDBERG, J. M. Physiology of peripheral neurons
REISINE, H. AND RAPHAN, T. Neural basis for eye velocity generation ininnervating otolith organs of the squirrel monkey. I. Response to staticthe vestibular nuclei during off-vertical axis rotation (OVAR). Exp.tilts and to long-duration centrifugal force. J. Neurophysiol. 39: 970–Brain Res. 92: 209–226, 1992.995, 1976.
REISINE, H., SIMPSON, J. I., AND HENN, V. A geometric analysis of semicir-GOLDBERG, J. M. AND FERNANDEZ, C. Physiology of peripheral neuronscular canals and induced activity in their peripheral afferents in the rhesusinnervating semicircular canals of the squirrel monkey. I. Resting dis-monkey. Ann. NY Acad. Sci. 545: 10–20, 1988.charge and response to angular accelerations. J. Neurophysiol. 34: 676–
REISINE, H., SIMPSON, J. I., RUDINGER, D., AND HENN, V. Combined anatom-684, 1971.ical and physiological study of semicircular canal orientation in the rhesusGOLDBERG, J. M. AND FERNANDEZ, C. Responses of peripheral vestibular monkey. Soc. Neurosci. Abstr. 11: 97.2, 1985.
neurons to angular and linear accelerations in the squirrel monkey. Acta ROBINSON, D. A. A method of measuring eye movement using a scleralOtolaryngol. (Stockh.) 80: 101–110, 1975. search coil in a magnetic field. IEEE Trans. Bio-Med. Electron. 10: 137–
GUEDRY F. E. Psychophysics of vestibular sensation. In: Handbook of Sen- 145, 1963.sory Physiology. Vestibular System. Psychophysics, Applied Aspects and SCHULTHEIS, L. W. AND ROBINSON, D. A. Directional plasticity of the ves-General Interpretations , edited by H. H. Kornhuber. New York: tibulo-ocular reflex in the cat. Ann. NY Acad. Sci. 374: 504–512, 1981.Springer-Verlag, 1974, vol. 6(2) , p. 3–154. SIROTA, M. G., BABAEV, B. M., BELOOZEROVA, I. N., NYROVA, A. N., YA-
HALMAGYI, G. M. AND CURTHOYS, I. S. A clinical sign of canal paresis. KUSHIN, S. B. AND KOZLOVSKAYA, I. B. Neuronal activity of vestibularArch. Neurol. 45: 737–739, 1988. nuclei during coordinated movement of eyes and head in microgravita-
HIGHSTEIN, S. M., RABBITT, R. D., AND BOYLE, R. Determinants of semicir- tion. Physiologist 31: 8–9, 1988.cular canal afferent response dynamics in the toadfish, Opsanus tau. J. STEINHAUSEN. Uber die Beobachtung der Cupula in den Bogengansampul-Neurophysiol 75: 575–596, 1996. len des Labyrinths des lebenden Hechts. Arch. Ges. Physiol. 232: 500–
HARRISON, R.E.W., BAKER, J.F., ISU, N., WICKLAND, C.R., AND PETERSON, 512, 1933.B.W. Dynamics of adaptive changes in vestibulo-ocular reflex directions. SUZUKI, J.-I. AND COHEN, B. Integration of semicircular canal activity. J.I. Rotations in the horizontal plane. Brain Res. 371: 162–165, 1986. Neurophysiol. 29: 981–995, 1966.
HENN, V., STRAUMANN, D., HESS, B.J.M., HASLWANTER, T., AND IIDA, N. SUZUKI, J., COHEN, B., AND BENDER, M. B. Compensatory eye movementsThree dimensional transformation from vestibular and visual input to induced by vertical semicircular canal stimulation. Exp. Neurol. 9: 137–oculomotor output. Ann. NY Acad. Sci. 656: 166–180, 1992. 160, 1964.
JUDGE, S. J., RICHMOND, B. J., AND CHU, F. C. Implantation of magnetic SUZUKI, J.-I., KODAMA, A., COHEN, B., AND HENN, V. Canal-plugging insearch coils for measurement of eye position: an improved method. Vision the rhesus monkey: a tool to study the contribution of individual canals
to nystagmus generation. Acta Otolaryngol. (Suppl.) 481: 91–93, 1991.Res . 20: 535–538, 1980.TABAK, S. AND COLLEWIJN, H. Evaluation of the human vestibulo-ocularKEPPEL, G. Design and Analysis: A Researcher’s Handbook . Englewood
reflex at high frequencies with a helmet, driven by reactive torque. ActaCliffs, NJ: Prentice Hall, 1991.Otolaryngol. (Stockh.) 520: 4–8, 1995.LANDOLT, J. P. AND CORREIA, M. J. Neurodynamic response analysis of
TELFORD, L., SEIDMAN, S. H., AND PAGE, G. D. Canal-otolith interactionsanterior semicircular canal afferents in the pigeon. J. Neurophysiol. 43:driving vertical and horizontal eye movements in the squirrel monkey.1746–1770, 1980.Exp. Brain Res. 109: 407–418, 1996.MINOR, L. B. AND GOLDBERG, J. M. Influence of static head position on
WEARNE, S., RAPHAN, T., AND COHEN, B. Nodulo-uvular control of thethe horizontal nystagmus evoked by caloric, rotational and optokineticcentral vestibular dynamics determines spatial orientation of the angularstimulation in the squirrel monkey. Exp. Brain Res. 82: 1–13,vestibulo-ocular reflex. Ann. NY Acad. Sci. 781: 364–384, 1996.1990.
WEARNE, S., RAPHAN, T., AND COHEN, B. Contribution of vestibular com-MONEY, K. E. AND SCOTT, J. W. Functions of separate sensory receptorsmissural pathways to spatial orientation of the angular vestibulo-ocularof non-auditory labyrinth of the cat. Am. J. Physiol. 202: 1211–1220,reflex. J. Neurophysiol. 78: 1193–1197, 1997.1962.
WEARNE, S., RAPHAN, T., AND COHEN, B. Control of spatial orientation ofNATIONAL RESEARCH COUNCIL. Guide for the Care and Use of Laboratorythe angular vestibulo-ocular reflex by the nodulus and uvula. J. Neuro-Animals . Washington, DC: National Academy Press, 1996.physiol. 79: 2690–2715, 1998.
PENG, G.C.Y., BAKER, J. F., AND PETERSON, B. W. Dynamics of directional YAKUSHIN, S. B., DAI, M., RAPHAN, T., SUZUKI, J.-I., AND COHEN, B. Eyeplasticity in the human vertical vestibulo-ocular reflex. J. Vestib. Res. 4: velocity induced by steps of angular velocity after canal plugging. In:453–460, 1994. Three-Dimentional Kinematics of Eye-, Head-, and Limb Movements ,
PETERSON, B. W., BAKER, J. F., AND HOUK, J. C. A model of adaptive edited by M. Fetter, H. Misslisch, and D. Tweed. Switzerland: Harwoodcontrol of vestibuloocular reflex based on properties of cross-axis adapta- Academic Publishers GMBH, 1997, p. 209–215.tion. Ann. NY Acad. Sci. 627: 319–337, 1991. YAKUSHIN, S. B., DAI, M., SUZUKI, J.-I., RAPHAN, T., AND COHEN, B. Semi-
RABBITT, R. D., BOYLE, R., YAMAUCHI, A., AND HIGHSTEIN, S. M. A resid- circular canal contributions to the three-dimensional vestibuloocular re-ual and organ response persists following semicircular canal plugging in flex: a model-based approach. J. Neurophysiol. 74: 2722–2738, 1995.the toadfish, Opsanus tau . Soc. Neurosci. Abstr. 22: 1819, 1996. YAKUSHIN, S. B., SUZUKI, J.-I., DAI, M., RAPHAN, T., AND COHEN, B. Nor-
RAPHAN, T. AND COHEN, B. The role of integration in oculomotor control. malization effects of vision on the compensatoty VOR after canal plug-In: Models of Oculomotor Behavior and Control , edited by B. Zuber. ging. Ann. NY Acad. Sci. 781: 713–717, 1996.West Palm Beach, FL: CRC, 1981, p. 91–109. YUE, Q., STRAUMANN, D., AND HENN, V. Three-dimensional characteristics
RAPHAN, T. AND COHEN, B. Multidimensional organization of the vestibulo- of rhesus monkey vestibular nystagmus after velocity steps. J. Vestib.Res. 4: 313–323, 1994.ocular reflex (VOR). In: Adaptive Processes in Visual and Oculomotor
J1032-7/ 9k2e$$no07 12-09-98 07:00:21 neupa LP-Neurophys
by guest on June 11, 2013http://jn.physiology.org/
Dow
nloaded from




















