
Effectiveness of exposure to longday followed by melatonin treatment on semen characteristics of...
12
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright
Transcript of Effectiveness of exposure to longday followed by melatonin treatment on semen characteristics of...

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Effectiveness of exposure to longday followed by melatonin
treatment on semen characteristics of Damascus male goats
during breeding and non-breeding seasons
T.A. Ramadan, T.A. Taha *, M.A. Samak, A. Hassan
Animal Production Department, Faculty of Agriculture (El-Shatby), Alexandria University, Alexandria 22545, Egypt
Received 17 December 2007; received in revised form 14 July 2008; accepted 26 July 2008
Abstract
Eight mature Damascus male goats were randomly divided into two equal groups. The first group served as control and the
second group was used to study the effect of exposure to longday (16L/8D) followed by melatonin treatment (2 mg/(animal/day)) on
semen characteristics and blood hormonal levels during breeding and non-breeding seasons. During breeding season, longday
exposure followed by melatonin treatments resulted in significant increases (P < 0.01) in means of sperm motility, ejaculate
volume, sperm concentration, total sperm output, total functional sperm fraction and blood testosterone concentration, while means
of reaction time, dead sperm, abnormal sperm and blood triiodothyronine (T3) concentration were significantly decreased
(P < 0.01). During non-breeding season, longday exposure followed by melatonin treatment exhibited improvements in some
reproductive parameters by reducing (P < 0.05) reaction time and percentage of dead sperm (P < 0.01) and increasing (P < 0.05)
total functional sperm fraction. Furthermore, longday exposure followed by melatonin treatment resulted in significant increases
(P < 0.01) in both of blood testosterone and T3 concentrations. It is concluded that longday exposure followed by melatonin
treatment successfully improve semen characteristics of Damascus male goats during breeding and non-breeding seasons. However,
the beneficial effects were more pronounced during the breeding season.
# 2009 Elsevier Inc. All rights reserved.
Keywords: Longday exposure; Melatonin; Semen characteristics; Blood serum hormones; Goats
1. Introduction
In most species, seasonal reproductive activity is
regulated by an endogenous rhythm. This rhythm is
synchronized with the geophysical year by environ-
mental stimuli, the most important of which is
photoperiod. Unlike temperature, availability of food,
rainfall or other environmental cues, photoperiod
provides information about the season and remains
constant from year to year. Photoperiodic stimuli are
transmitted via neuroendocrine pathways into hormonal
signals that regulate gonadal activity. Under subtropical
conditions in Egypt, ram [1], and goat [2], semen
qualities were found to be better during summer months
than during other months of the year. Sexual activity of
subtropical Mexican Creole bucks can be induced
during the non-breeding season by a longday treatment
followed by melatonin implantation [3]. In Mexican
bucks as in other subtropical breeds including
Damascus bucks, the breeding season takes place in
summer and autumn [4]. In these breeds, breeding
season starts during the natural increasing days, but in
controlled photoperiod conditions, longdays were found
to inhibit testosterone secretion in Creole goats [5], and
www.theriojournal.com
Available online at www.sciencedirect.com
Theriogenology 71 (2009) 458–468
* Corresponding author. Tel.: +20 35921960; fax: +20 35922780.
E-mail address: [email protected] (T.A. Taha).
0093-691X/$ – see front matter # 2009 Elsevier Inc. All rights reserved.
doi:10.1016/j.theriogenology.2008.07.029

Author's personal copy
in Merino and Suffolk rams [6]. In addition, Roca et al.
[7], found that semen quality of Murciano Grandina
goats was highest during summer and fall seasons in
Southern Spain (Mediterranean basin). Recently, the
best semen quality in Damascus bucks was obtained
during increasing photoperiod in spring and summer [8]
In sheep, marked seasonal variations in plasma
thyroid hormone concentrations have been described.
Low and high temperatures, respectively, are stimula-
tory and inhibitory to thyroid activity [9]. Photoperiod
also acts on thyroid activity, as triiodothyronine (T3)
plasma concentration followed the photoperiodic
changes, increasing during long days and decreasing
during short days in Alpine and Saanen male goats [10].
These seasonal cycles of circulating thyroid hormones
have been reported comparatively with testicular
endocrine activity in a broad variety of seasonal
breeders [11]. Plasma testosterone concentrations were
found to be negatively correlated with plasma thyroid
hormone concentrations in goats [12], and sheep [1].
Melatonin (N-acetyl-5-methoxytryptamine) is
synthesized by the pineal gland during the dark phase
of the photoperiod and it is rapidly released to the blood
stream where the highest levels reach nanomolar
concentrations [13]. Melatonin is a signal that conveys
photoperiodic information to synchronize cell physiol-
ogy with the dark–light cycle [13]. In addition, it is a
lipophilic agent that crosses lipid bilayers [14], and acts
as a free radical scavenger, neutralizing hydroxyl and
peroxyl radicals among others, preventing lipid
membrane peroxidation and apoptosis [15], and
protecting the DNA from the damage induced by free
radicals [16]. Moreover, melatonin stimulates gene
expression of antioxidative enzymes including super-
oxide dismutase, glutathione peroxidase, catalase and
glutathione reductase [17]. Studies dealing with the
antioxidative effect of melatonin on spermatozoa have
shown that melatonin significantly reduces the rate of
lipid peroxidation in sperm [18], and it can protect
sperm mitochondria from the damage induced by
reactive oxygen species (ROS) throughout its effective
antioxidative potential [19].
The objectives of the present study were to evaluate
the effect of longday exposure and melatonin treatment
on semen characteristics, and on blood serum con-
centrations of testosterone and T3 of Damascus male
goats during breeding and non-breeding seasons.
2. Materials and methods
The present study was conducted at the Agricultural
Experimental Station (318200N, 308E), Faculty of
Agriculture, Alexandria University, Egypt, on Damas-
cus male goats during the breeding season (from 9 July
to 22 November) and non-breeding season (from 8
January to 27 May).
2.1. Animals and management
Eight Damascus male goats aged 3–4 years and
weighed between 52 and 69 kg were used throughout
the present study. Animals were kept outdoors with
shelter during the daytime and housed in a semi-open
barn at night. They were offered roughage and
concentrate supplement according to their body weight
requirement [20]. Animals were given Egyptian clover
(Trifolium alexandrinum) in winter and spring and
chopped green maize in summer and autumn in addition
to hay. Each animal also received 500 g per day of a
pelleted concentrate mixture that contained 63% total
digestible nutrients (TDN) and 14% crude protein.
Water was offered in access to animals at all times.
Animals were free of any disease and were clinically
normal with a healthy appearance.
2.2. Photoperiod and melatonin treatment
Eight mature Damascus male goats were randomly
divided into two equal groups. The control group
remained in an open barn under natural day length and
ambient temperature throughout the experimental
period. Day length varied from 13 h 29 min during
the breeding season (summer) to 11 h 24 min during the
non-breeding season (winter). Animals from the
experimental groups were maintained at the same
conditions except for 3 months of photoperiodic
treatment (from 9 July, to 25 September) during the
breeding season. About 3.5 months later, the animals
were subjected to other 3 months of photoperiodic
treatment (from 8 January, to 1 April) during non-
breeding season. Photoperiodic treatment was carried
out in a light-proof building where animals were
exposed to longdays (16 h of light/day, lights on at
06:00 h, lights off at 22:00 h). Photoperiod was
regulated by an electric timer and light intensity was
at least 300 lx, positioned laterally to the eyes of the
animals. At the end of photoperiodic treatment, the light
treatment was stopped, and the males were exposed to
the natural day length until the end of the study.
Treatment with melatonin followed the photoperiodic
treatment in both breeding and non-breeding seasons
and was carried out at 18:00 h on the daytime. It
involved the oral administration of 2 mg melatonin
(Sigma Chemical Co., St. Louis, MO, USA) dissolved
T.A. Ramadan et al. / Theriogenology 71 (2009) 458–468 459

Author's personal copy
in 2 ml 50% ethanol/water (v/v) every day according to
Lincoln and McNeilly [21], during 2 months. The
corresponding control group received an equivalent
dose of the vehicle (50% ethanol/water). The experi-
ment layout is shown in Fig. 1).
2.3. Data collected
The following parameters were measured in all
experimental animals throughout the last 4 weeks of
photoperiodic treatment (prior to melatonin treatment)
and 8 weeks of melatonin administration.
2.3.1. Body weight, relative testes volume and
libido
Live body weight of each animal was recorded
weekly before access to feed and water. Scrotal sac
volume was measured by the water displacement
method described by Evans and Robinson [22], and
was divided by body weight to calculate the relative
testes volume. Sexual behavior of bucks was recorded
weekly by using the reaction time criteria. The time
elapsed between introducing the buck to a female doe at
estrus and semen collection was measured in seconds
using a stop-watch.
2.3.2. Semen evaluation
Semen collection by an artificial vagina was carried
out weekly from all animals throughout the experi-
mental period. The volume of sample was recorded
using a graduated collection tube. Determination of
semen initial fructose was carried out immediately after
collection according to Mann [23]. Ejaculates were
placed in a water bath adjusted to 38 8C. Sperm motility
(%) was subjectively estimated at 400� magnification
using a light microscope equipped with a warm stage.
Sperm concentration was counted by the hemocyt-
ometer method. Total sperm output was calculated by
multiplying semen ejaculate volume and semen con-
centration. The percentages of dead, live and abnormal
spermatozoa were assessed according to Blom [24]. The
total functional sperm fraction (TFSF; X 109) parameter
was calculated as the product of total sperm output by
progressive motility by normal morphology [25,26].
2.4. Blood collection
Blood samples were collected from the jugular vein
of animals every 2 weeks throughout the experimental
period. Blood samples were obtained in the morning
before access to feed and water. Serum was obtained by
centrifugation of samples at 3000 r.p.m for 20 min.
Serum was collected and stored at �20 8C until
analysis.
2.5. Hormonal assays
The concentration of T3 in blood serum samples was
measured using solid-phase enzyme immunoassay kits
obtained from Diatech Diagnostics Inc., Boston, USA.
The lower limit of detection (95% B/B8) was 0.16 ng/ml
serum, and the intra- and inter-assay coefficients of
variation (CV) were 6.4 and 5.1%, respectively based on
the means of low, medium and high quality control
samples measured in 12 assays. The concentration of
testosterone in sera was determined by solid-phase
enzyme immunoassay kits obtained from Bio Source
International, Inc., CA, USA. The lower limit of
detection (95% B/B8) was 0.05 (s.e.0.02) ng/ml serum,
and the intra- and inter-assay CVs were 7.2 and 6.9%,
respectively based on the means of low, medium and
high quality control samples measured in 20 assays.
2.6. Statistical analysis
Data were analyzed using generalized linear model
procedure [27], by two-way (treatments and weeks)
ANOVA. Comparisons among main effects were tested
T.A. Ramadan et al. / Theriogenology 71 (2009) 458–468460
Fig. 1. Experimental layout during breeding and non-breeding seasons.
Author's personal copy
using LSD.05 of SAS [27]. For the significance
interactions difference between the two treatments
within each week was tested using the LSD.05 for the
interaction.
3. Results
3.1. Effect of longday exposure followed by
melatonin treatment during breeding season
Effects of longday exposure followed by melatonin
treatment on overall mean semen characteristics and
blood hormonal levels of Damascus goats during
breeding season are displayed in Table 1 and their
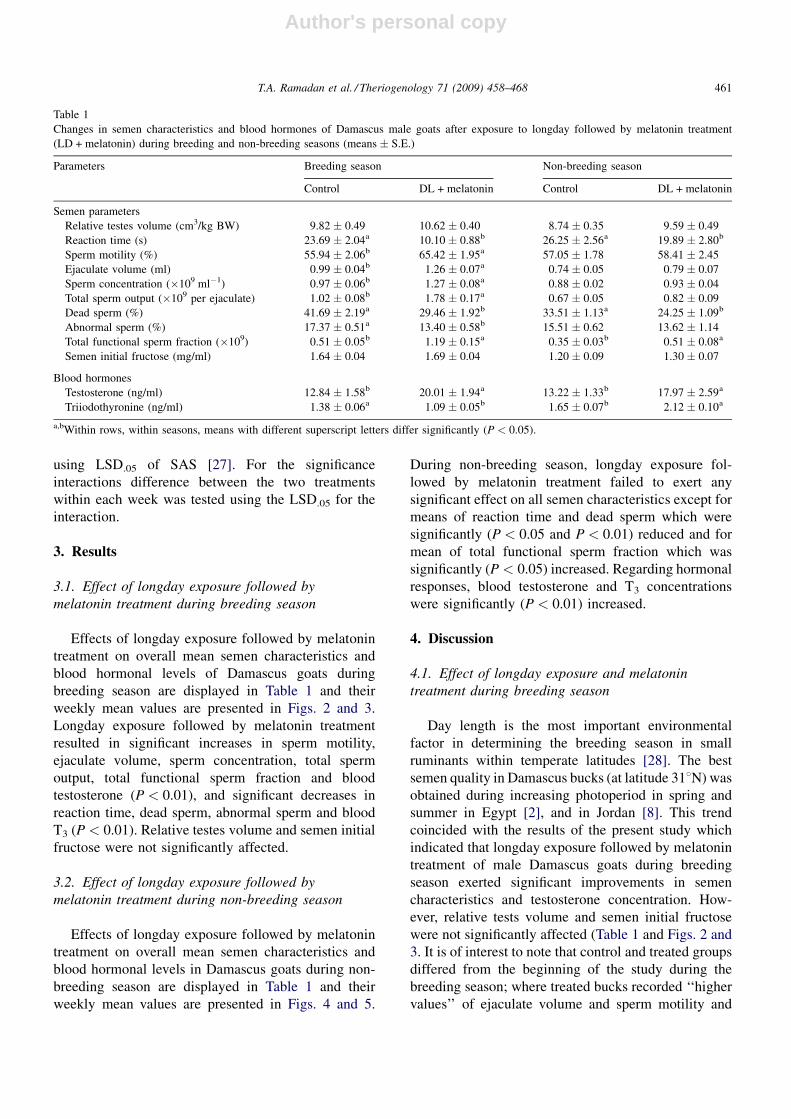
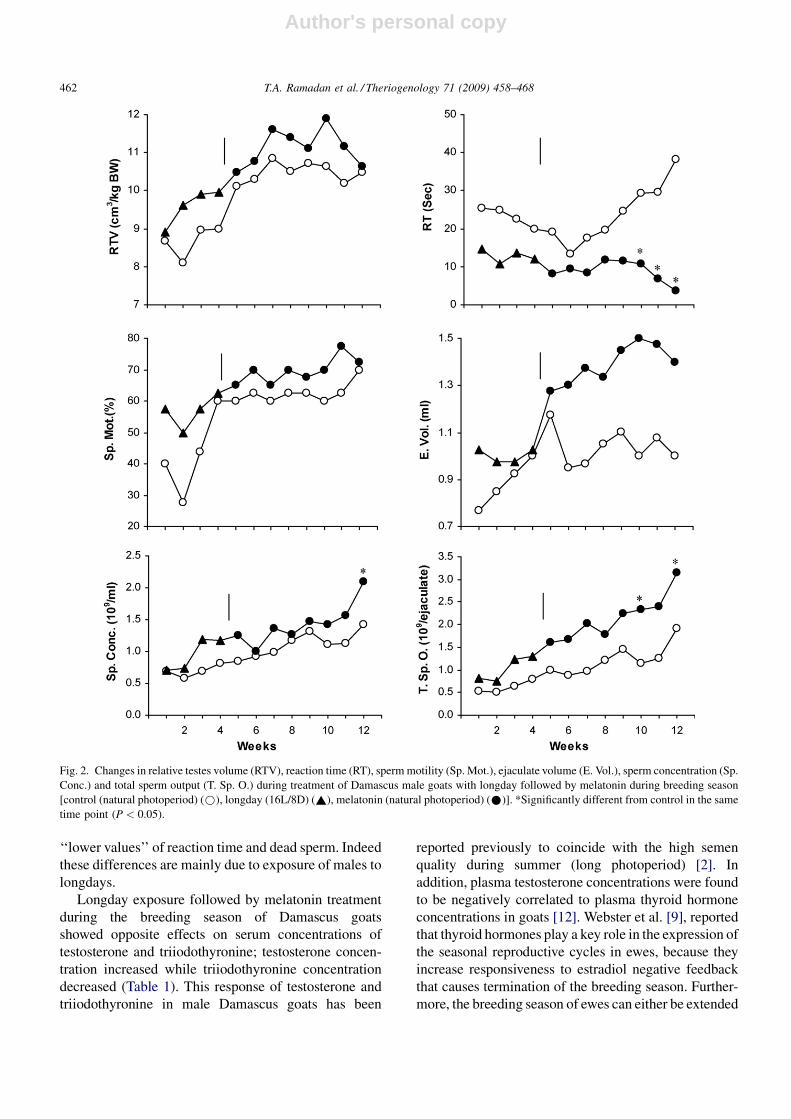
weekly mean values are presented in Figs. 2 and 3.
Longday exposure followed by melatonin treatment
resulted in significant increases in sperm motility,
ejaculate volume, sperm concentration, total sperm
output, total functional sperm fraction and blood
testosterone (P < 0.01), and significant decreases in
reaction time, dead sperm, abnormal sperm and blood
T3 (P < 0.01). Relative testes volume and semen initial
fructose were not significantly affected.
3.2. Effect of longday exposure followed by
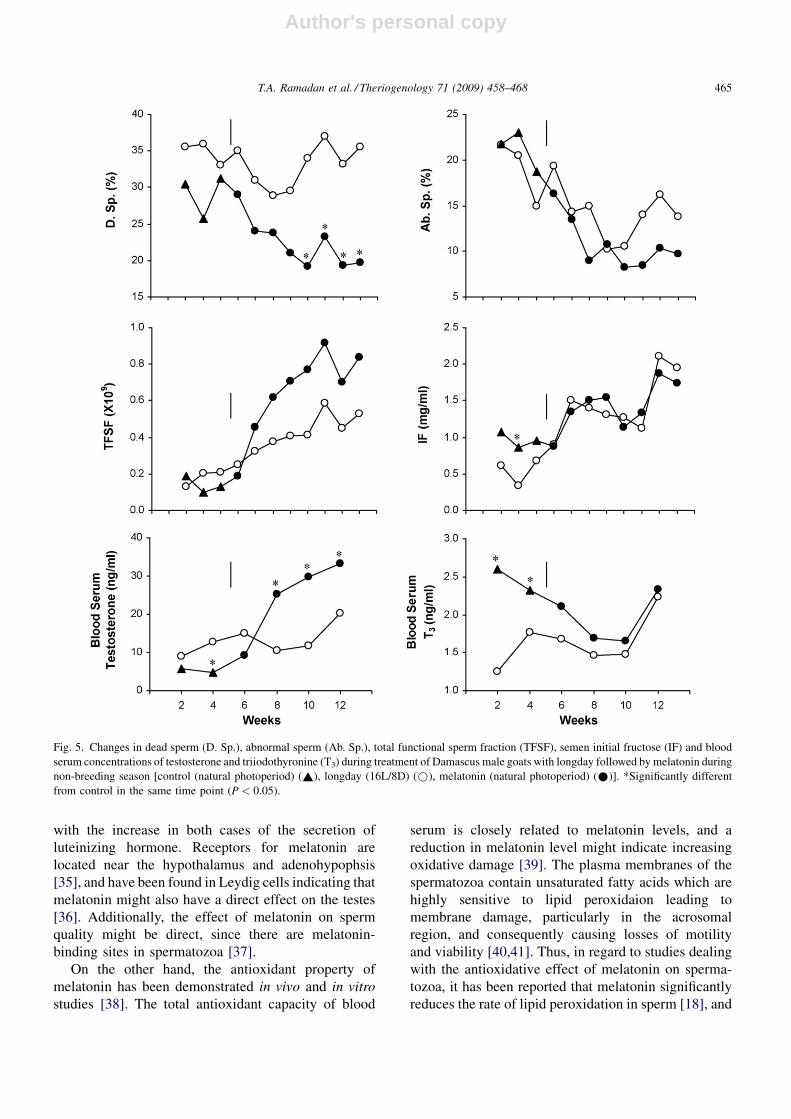
melatonin treatment during non-breeding season
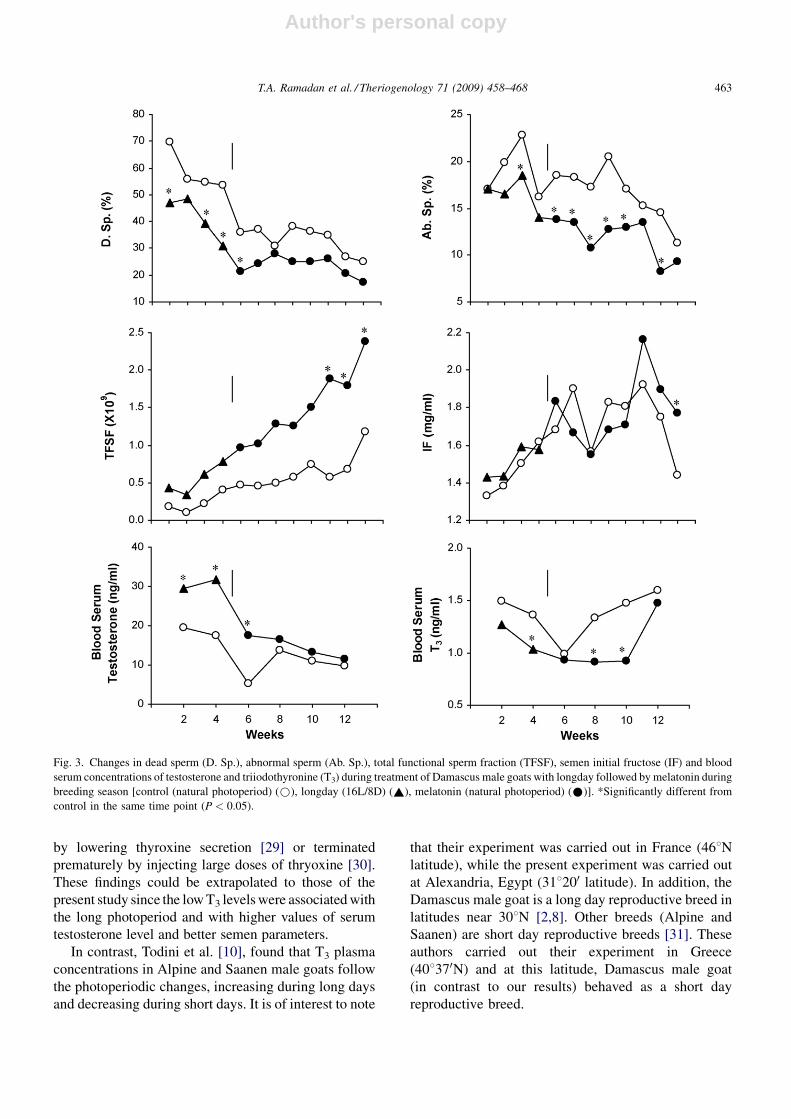
Effects of longday exposure followed by melatonin
treatment on overall mean semen characteristics and
blood hormonal levels in Damascus goats during non-
breeding season are displayed in Table 1 and their
weekly mean values are presented in Figs. 4 and 5.
During non-breeding season, longday exposure fol-
lowed by melatonin treatment failed to exert any
significant effect on all semen characteristics except for
means of reaction time and dead sperm which were
significantly (P < 0.05 and P < 0.01) reduced and for
mean of total functional sperm fraction which was
significantly (P < 0.05) increased. Regarding hormonal
responses, blood testosterone and T3 concentrations
were significantly (P < 0.01) increased.
4. Discussion
4.1. Effect of longday exposure and melatonin
treatment during breeding season
Day length is the most important environmental
factor in determining the breeding season in small
ruminants within temperate latitudes [28]. The best
semen quality in Damascus bucks (at latitude 318N) was
obtained during increasing photoperiod in spring and
summer in Egypt [2], and in Jordan [8]. This trend
coincided with the results of the present study which
indicated that longday exposure followed by melatonin
treatment of male Damascus goats during breeding
season exerted significant improvements in semen
characteristics and testosterone concentration. How-
ever, relative tests volume and semen initial fructose
were not significantly affected (Table 1 and Figs. 2 and
3. It is of interest to note that control and treated groups
differed from the beginning of the study during the
breeding season; where treated bucks recorded ‘‘higher
values’’ of ejaculate volume and sperm motility and
T.A. Ramadan et al. / Theriogenology 71 (2009) 458–468 461
Table 1
Changes in semen characteristics and blood hormones of Damascus male goats after exposure to longday followed by melatonin treatment
(LD + melatonin) during breeding and non-breeding seasons (means � S.E.)
Parameters Breeding season Non-breeding season
Control DL + melatonin Control DL + melatonin
Semen parameters
Relative testes volume (cm3/kg BW) 9.82 � 0.49 10.62 � 0.40 8.74 � 0.35 9.59 � 0.49
Reaction time (s) 23.69 � 2.04a 10.10 � 0.88b 26.25 � 2.56a 19.89 � 2.80b
Sperm motility (%) 55.94 � 2.06b 65.42 � 1.95a 57.05 � 1.78 58.41 � 2.45
Ejaculate volume (ml) 0.99 � 0.04b 1.26 � 0.07a 0.74 � 0.05 0.79 � 0.07
Sperm concentration (�109 ml�1) 0.97 � 0.06b 1.27 � 0.08a 0.88 � 0.02 0.93 � 0.04
Total sperm output (�109 per ejaculate) 1.02 � 0.08b 1.78 � 0.17a 0.67 � 0.05 0.82 � 0.09
Dead sperm (%) 41.69 � 2.19a 29.46 � 1.92b 33.51 � 1.13a 24.25 � 1.09b
Abnormal sperm (%) 17.37 � 0.51a 13.40 � 0.58b 15.51 � 0.62 13.62 � 1.14
Total functional sperm fraction (�109) 0.51 � 0.05b 1.19 � 0.15a 0.35 � 0.03b 0.51 � 0.08a
Semen initial fructose (mg/ml) 1.64 � 0.04 1.69 � 0.04 1.20 � 0.09 1.30 � 0.07
Blood hormones
Testosterone (ng/ml) 12.84 � 1.58b 20.01 � 1.94a 13.22 � 1.33b 17.97 � 2.59a
Triiodothyronine (ng/ml) 1.38 � 0.06a 1.09 � 0.05b 1.65 � 0.07b 2.12 � 0.10a
a,bWithin rows, within seasons, means with different superscript letters differ significantly (P < 0.05).
Author's personal copy
‘‘lower values’’ of reaction time and dead sperm. Indeed
these differences are mainly due to exposure of males to
longdays.
Longday exposure followed by melatonin treatment
during the breeding season of Damascus goats
showed opposite effects on serum concentrations of
testosterone and triiodothyronine; testosterone concen-
tration increased while triiodothyronine concentration
decreased (Table 1). This response of testosterone and
triiodothyronine in male Damascus goats has been
reported previously to coincide with the high semen
quality during summer (long photoperiod) [2]. In
addition, plasma testosterone concentrations were found
to be negatively correlated to plasma thyroid hormone
concentrations in goats [12]. Webster et al. [9], reported
that thyroid hormones play a key role in the expression of
the seasonal reproductive cycles in ewes, because they
increase responsiveness to estradiol negative feedback
that causes termination of the breeding season. Further-
more, the breeding season of ewes can either be extended
T.A. Ramadan et al. / Theriogenology 71 (2009) 458–468462
Fig. 2. Changes in relative testes volume (RTV), reaction time (RT), sperm motility (Sp. Mot.), ejaculate volume (E. Vol.), sperm concentration (Sp.
Conc.) and total sperm output (T. Sp. O.) during treatment of Damascus male goats with longday followed by melatonin during breeding season
[control (natural photoperiod) (*), longday (16L/8D) (~), melatonin (natural photoperiod) (*)]. *Significantly different from control in the same
time point (P < 0.05).
Author's personal copy
by lowering thyroxine secretion [29] or terminated
prematurely by injecting large doses of thryoxine [30].
These findings could be extrapolated to those of the
present study since the low T3 levels were associated with
the long photoperiod and with higher values of serum
testosterone level and better semen parameters.
In contrast, Todini et al. [10], found that T3 plasma
concentrations in Alpine and Saanen male goats follow
the photoperiodic changes, increasing during long days
and decreasing during short days. It is of interest to note
that their experiment was carried out in France (468Nlatitude), while the present experiment was carried out
at Alexandria, Egypt (318200 latitude). In addition, the
Damascus male goat is a long day reproductive breed in
latitudes near 308N [2,8]. Other breeds (Alpine and
Saanen) are short day reproductive breeds [31]. These
authors carried out their experiment in Greece
(408370N) and at this latitude, Damascus male goat
(in contrast to our results) behaved as a short day
reproductive breed.
T.A. Ramadan et al. / Theriogenology 71 (2009) 458–468 463
Fig. 3. Changes in dead sperm (D. Sp.), abnormal sperm (Ab. Sp.), total functional sperm fraction (TFSF), semen initial fructose (IF) and blood
serum concentrations of testosterone and triiodothyronine (T3) during treatment of Damascus male goats with longday followed by melatonin during
breeding season [control (natural photoperiod) (*), longday (16L/8D) (~), melatonin (natural photoperiod) (*)]. *Significantly different from
control in the same time point (P < 0.05).
Author's personal copy
4.2. Effect of longday exposure and melatonin
treatment during non-breeding season
Previous studies indicated that melatonin adminis-
tration to goats during breeding season significantly
improved testes volume, ejaculate volume and sperm
concentration but sperm motility and abnormal sperm
were not affected [32]. Similar studies conducted on
ram by Hanif and Williams [33], indicated that
melatonin administration increased sperm motility, live
sperm, sperm concentration and decreased abnormal
sperm, but relative testes volume was not affected. Such
findings are similar to our results.
Langford et al. [34], reported that melatonin
stimulates spermatogenic activity of ram testes by
increasing the sensitivity of Leydig cells to luteinizing
hormone. The increase in the mean values of
testosterone in blood of rams after administration of
melatonin has also been attributed to the effect of
melatonin on the hypothalamus or hypophysis [35],
T.A. Ramadan et al. / Theriogenology 71 (2009) 458–468464
Fig. 4. Changes in relative testes volume (RTV), reaction time (RT), sperm motility (Sp. Mot.), ejaculate volume (E. Vol.), sperm concentration (Sp.
Conc.) and total sperm output (T. Sp. O.) during treatment of Damascus male goats with longday followed by melatonin during non-breeding season
[control (natural photoperiod) (*), longday (16L/8D) (~), melatonin (natural photoperiod) (*)]. *Significantly different from control in the same
time point (P < 0.05).
Author's personal copy
with the increase in both cases of the secretion of
luteinizing hormone. Receptors for melatonin are
located near the hypothalamus and adenohypophsis
[35], and have been found in Leydig cells indicating that
melatonin might also have a direct effect on the testes
[36]. Additionally, the effect of melatonin on sperm
quality might be direct, since there are melatonin-
binding sites in spermatozoa [37].
On the other hand, the antioxidant property of
melatonin has been demonstrated in vivo and in vitro
studies [38]. The total antioxidant capacity of blood
serum is closely related to melatonin levels, and a
reduction in melatonin level might indicate increasing
oxidative damage [39]. The plasma membranes of the
spermatozoa contain unsaturated fatty acids which are
highly sensitive to lipid peroxidaion leading to
membrane damage, particularly in the acrosomal
region, and consequently causing losses of motility
and viability [40,41]. Thus, in regard to studies dealing
with the antioxidative effect of melatonin on sperma-
tozoa, it has been reported that melatonin significantly
reduces the rate of lipid peroxidation in sperm [18], and
T.A. Ramadan et al. / Theriogenology 71 (2009) 458–468 465
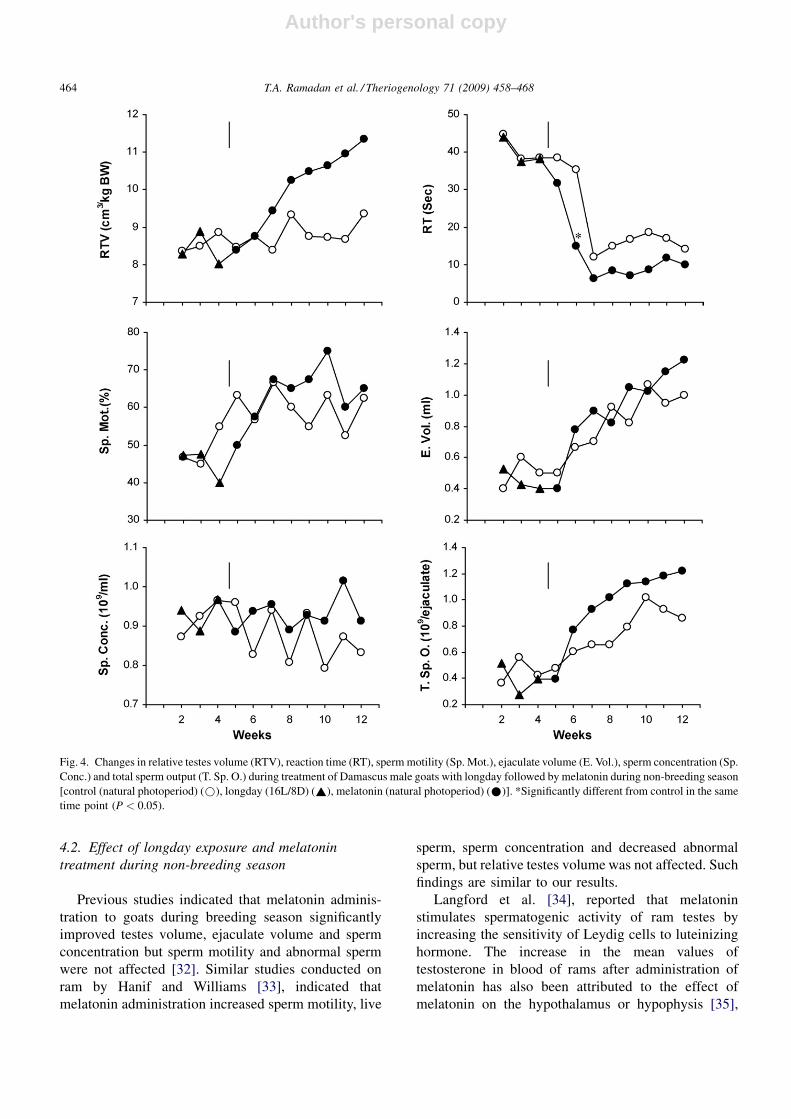
Fig. 5. Changes in dead sperm (D. Sp.), abnormal sperm (Ab. Sp.), total functional sperm fraction (TFSF), semen initial fructose (IF) and blood
serum concentrations of testosterone and triiodothyronine (T3) during treatment of Damascus male goats with longday followed by melatonin during
non-breeding season [control (natural photoperiod) (~), longday (16L/8D) (*), melatonin (natural photoperiod) (*)]. *Significantly different
from control in the same time point (P < 0.05).

Author's personal copy
it can protect sperm mitochondria from the damage
induced by reactive oxygen species (ROS) throughout
its effective antioxidative potential [19]. Furthermore,
melatonin implantation has been reported to improve
freezability of ram semen due to its antioxidant property
and protective effect against oxidative damage [42].
The beneficial effect of photoperiodic treatment
and melatonin administration on semen characteristics
during non-breeding season was not as obvious as it
was in the breeding season. This treatment exhibited
its improvement in semen characteristics as significant
decreases in reaction time and in the percentage of
dead sperm and significant increases in TFSF, while
other semen characteristics were not affected (Table 1
and Figs. 4 and 5). In agreement with the present
results, Rosa et al. [43], reported that melatonin
administration improved ram’s libido during non-
breeding season. However, previous studies dealing
with melatonin administration during non-breeding
season in goat and sheep yielded inconsistent results
where it ranged between no response to slight
improvement in some semen characteristics. Discre-
pancies between breeding and non-breeding seasons
in improving semen quality by melatonin treatment
could be explained on the basis of seasonal profile of
melatonin. Sheikheldin et al. [44], reported that
melatonin concentrations during summer and autumn
(breeding season) were significantly higher than those
in winter and spring (non-breeding season). Thus,
melatonin treatment during breeding season, where
the endogenous melatonin reached the maximum
level, represent an additive effect resulting in superior
improvement in semen quality compared to that of
non-breeding season where the endogenous melatonin
recorded the lowest level.
The percentages of dead sperm were ‘‘lower’’ in the
treated animals due to the beneficial effects of ‘‘longday
and/or melatonin’’ treatments. This was observed
during both breeding and non-breeding seasons
(Figs. 3 and 5). However, the percentages of dead
sperm were lower in non-breeding season than in
breeding season in both treated and control bucks. This
could be explained by the fact that after long sexual
arrest (during the non-breeding season), low semen
quality was noted at the beginning of breeding season,
hence it improved as ‘‘breeding season’’ advanced.
Thus, higher percentages of dead sperm were observed
at the beginning of the breeding season in both treated
and control bucks, but it tented to decrease sharply
throughout the breeding season (Fig. 3). On the other
hand, the percentages of dead sperm were lower at the
beginning of the non-breeding season in both control
and treated bucks than those of breeding season.
However, it failed to improve throughout the non-
breeding season in the control group, while dead sperm
decreased in the treated bucks as the season advanced
(Fig. 5).
Melatonin administration during non-breeding sea-
son also resulted in increases in serum concentrations of
testosterone and T3 hormones (Table 1). This finding
agrees with those reported in goats [3], and in sheep
[43]. The increase in mean values of blood testosterone
after administration of melatonin has been attributed to
the effect of melatonin on the hypothalamus or
hypophysis [35], which increases the secretion of
luteinizing hormone in both cases.
Naturally, seasonal variations in serum concentration
of T3 showed an increase in its level during winter and
spring (non-breeding time) in Damascus male goats [2].
As shown in the present study, melatonin treatment
caused a significant increase (P < 0.01) in T3 con-
centration (2.12 ng/ml) than that of control group
(1.65 ng/ml) (Table 1). It is known that thyroid
hormones accelerate the basal metabolic rate [45],
and may increase free radicals production and lipid
peroxide level [46]. Melatonin stimulates the activities
of some enzymes that metabolize reactive species and
also alters cell membrane fluidity, thereby reducing the
ability of damaging species to attack polyunsaturated
fatty acids in this structure [47]. The present study
suggests a protective role of melatonin in maintaining
normal sperm parameters, shown as significant reduc-
tions in reaction time and in the percentage of dead
sperm and significant increase in TFSF. However, the
melatonin-induced increase (P < 0.05) in T3 concen-
tration during non-breeding season did not reduce
sperm quality parameters due to the antioxidative effect
of melatonin on free radicals and on spermatozoal
membranes during that time.
5. Conclusions
The present study indicated that photoperiodic
treatment followed by melatonin administration
improved semen quality in Damascus male goats
during both of breeding and non-breading seasons.
However, this improvement was more pronounced
during breeding season compared with non-breeding
season and was suggested to be due to the antioxidative
effect of melatonin on spermatozoa. The divergent
responses of improvement of semen quality between
breeding and non-breeding seasons suggest that
Damascus goats are influenced by several environ-
mental factors in addition to photoperiod.
T.A. Ramadan et al. / Theriogenology 71 (2009) 458–468466

Author's personal copy
Acknowledgements
This research was partially funded by a research
grant awarded by the British Council, Egypt. The
authors wish to thank Dr. Mamdouh El-Rouby, Prof. of
Statistics, Faculty of Agriculture, Alexandria University
for his advice on the statistical analysis of data.
References
[1] Taha TA, Abdel-Gawad EI, Ayoub MA. Monthly variations in
some reproductive parameters of Barki and Awassi rams
throughout 1 year under subtropical conditions. 1. Semen char-
acteristics and hormonal levels. Anim Sci 2000;71:317–24.
[2] Ayoub MA, Taha TA, Abdel-Gawad EI. Monthly and seasonal
variations in semen characteristics and serum testosterone and
triiodothyronine levels in Barki, Damascus male goats and their
crosses under subtropical conditions. In: Proceedings of the third
all Africa conference on animal agriculture and 11th conference
of the Egyptian Society of animal production; 2000. p. 459–68.
[3] Delgadillo JA, Carrillo E, Moran J, Duarte G, Chemineau P,
Malpaux B. Induction of sexual activity of male Creole goats in
subtropical northern Mexico using long days and melatonin. J
Anim Sci 2001;79:2245–52.
[4] Delgadillo JA, Canedo GA, Chemineau P, Guillaume D, Mal-
paux B. Evidence for an annual reproductive rhythm indepen-
dent of food availability in male Creole goats in subtropical
northern Mexico. Theriogenology 1999;52:727–37.
[5] Delgadillo JA, Cortez ME, Duarte G, Chemineau P, Malpaux B.
Evidence that the photoperiod controls the annual changes in
testosterone secretion, testicular and body weight in subtropical
male goats. Reprod Nutr Dev 2004;44:183–93.
[6] Martin GB, Hotzel MJ, Blache D, Walkden-Brown SW, Black-
berry MA, Boukhliq R, et al. Determinants of the annual pattern
of reproduction in mature male Merino and Suffolk sheep:
modification of responses to photoperiod by an annual cycle
in food supply. Reprod Fert Dev 2002;14:165–75.
[7] Roca T, Martinez E, Vazquez JM, Coy P. Characteristics and
seasonal variations in the semen of Murciano-Granadina goats in
the Mediterranean area. Anim Reprod Sci 1992;29:255–62.
[8] Al-Ghalban AM, Tabbaa MJ, Kridli RT. Factors affecting semen
characteristics and scrotal circumference in Damascus bucks.
Small Rumin Res 2004;53:141–9.
[9] Webster JR, Moenter SM, Woodfill CIJ, Karsch FJ. Role of the
thyroid gland in seasonal reproduction. II. Thyroxine allows a
season-specific suppression of gonadotropin secretion in sheep.
Endocrinology 1991;129:176–83.
[10] Todini L, Delgadillo JA, Debenedetti A, Chemineau P. Plasma
total T3 and T4 concentrations in bucks as affected by photo-
period. Small Rumin Res 2006;65:8–13.
[11] Karsch FJ, Dahl GE, Hachigian TM, Thrun LA. Involvement of
thyroid hormones in seasonal reproduction. J Reprod Fertil
Suppl 1995;49:409–22.
[12] Todini L, Lucaroni A, Malfatti A, Debenedetti A, Costarelli S.
Andamento ormonale della concentrazione ematica delgli
ormoni tiroidei nella capra. Differenze fra masch e femmine
(male–female differences in the annual profiles of the thyroid
hormones blood level by the goat). Atti Soc Ital Sci Vet
1992;46:169–73.
[13] Reiter RJ. The melatonin rhythm: both a clock and a calendar.
Experientia 1993;49:654–64.
[14] Bongiorno D,CerauloL,Ferrugia M.Localization and interactions
of melatonin in dry cholesterol/lecithin mixed reversed micelles
used as cell membrane models. J Pineal Res 2005;38:292–8.
[15] Jou MJ, Peng TI, Reiter RJ. Visualization of the antioxidative
effects of melatonin at the mitochondrial level during oxidative
stress induced apoptosis in rat brain astrocytes. J Pineal Res
2004;37:55–70.
[16] Majsterek I, Gloc E, Blasiak J. A comparison of the action of
amifostine and melatonin DNA-damaging effects and apoptosis
induced by idarubicin in normal and caner cells. J Pineal Res
2005;38:254–63.
[17] Rodriguez C, Mayo JC, Sainz RM, Antolin I, Herrera F, Martin
V, et al. Regulation of antioxidative enzymes: a significant role
for melatonin. J Pineal Res 2004;36:1–9.
[18] Gavella M, Lipovac V. Antioxidative effect of melatonin on
human spermatozoa. Arch Androl 2000;44:23–7.
[19] Shang X, Huang Y, Ye Z, Yu X, Gu W. Protection of melatonin
against damage of sperm mitochondrial function induced by
reactive oxygen species. Zhonghua Nan Ke Xue 2004;10:604–7.
[20] NRC. Nutrient requirement of domestic animals, fourth revised
ed., Washington, D.C., USA: National Academy of Sciences;
1974.
[21] Lincoln GA, McNeilly AS. Inhibin concentrations in the per-
ipheral blood of rams during a cycle in testicular activity induced
by changes in photoperiod or treatment with melatonin. J
Endocrinol 1989;120:R9–13.
[22] Evans G, Robinson TJ. Reproductive potential and endocrino-
logical responses of sheep kept under controlled lighting. II.
Pituitary and gonadal responses of ewes and rams to a six-
monthly light cycle. Anim Reprod Sci 1980;3:39–56.
[23] Mann T. Fructose content and fructolysis in semen: practical
application in the evaluation of semen quality. J Agri Sci Camb
1948;38:323–31.
[24] Blom E. Pathological conditions in the genital organs and in the
semen as grounds for rejection of breeding bulls for import or
export to or from Denmark 1958–1982. Nordisk veterinarme-
dicin 1983;35:105–30.
[25] Zavos PM, Centola GM. Selection of sperm from oligozoosperic
men for ARTA: comparison between swin-up vs. sperm perpTM
fraction. J Ass Reprod Technol Androl 1991;1:338–45.
[26] Correa JR, Zavos PM. Preparation and recovery of frozen-
thawed bovine spermatozoa via sperm selection techniques
employed in assisted reproductive technologies. Theriogenology
1996;46:1225–32.
[27] SAS. SAS user’s guide: statistics, version 8 edition. Cary, NC:
SAS Inst, Inc.; 1999.
[28] Folch J. The influence of age, photoperiodism and temperature
on semen production of rams. In: Courot M, editor. The male in
farm animal reproduction. Amsterdam: Martinus Nijhoff; 1984.
p. 141–60.
[29] Follett BK, Potts C. Hypothyroidism affects reproductive refrac-
toriness and seasonal oestrous period in Welsh Mountain ewes. J
Endocrinol 1990;127:103–9.
[30] O’Callaghan D, Wendling A, Karsch FJ, Roche JF. The effect of
exogenous thyroxine on timing of seasonal reproductive transi-
tions in ewe. Biol Reprod 1993;49:311–5.
[31] Karagiannidis A, Varsakeli S, Karatzas G. Characteristics and
seasonal variations in the semen of Alpine. Saanen and Damas-
cus goat bucks born and raised in Greece. Theriogenology
2000;53:1285–93.
T.A. Ramadan et al. / Theriogenology 71 (2009) 458–468 467

Author's personal copy
[32] Trejo GA, Ponce LC, Vidal GMA. Effects of melatonin and
GnRH treatments on semen quality in young male goats. In:
Proceedings of seventh international conference on goats; 2000.
[33] Hanif M, Williams HL. The effect of melatonin and light
treatment on the reproductive performance of yearling Suffolk
rams. Br Vet J 1991;147:49–56.
[34] Langford GA, Ainsworth L, Marcus G, Shrestha JNB. Photo-
period entrainment of testosterone, lutenizing hormone, follicle
stimulating hormone and prolactin cycles in rams in relation to
testis size and semen quality. Biol Reprod 1987;37:489–99.
[35] Arendt J. Physiology of pineal: role in photoperiodic seasonal
functions. In: Arendt J, editor. Melatonin and mammalian pineal
gland. London: Champan and Hall; 1995. p. 110–58.
[36] Shiu S, Yu Z, Chow P, Pang S. Putative melatonin receptors in the
male reproductive tissues. In: Tang P, Pang S, Reiter R, editors.
Melatonin a universal photoperiodic signal with diverse actions.
AG, Hong Kong: Karger; 1996. p. 90–100.
[37] Van Vuuren RJ, Pitout MJ, Van Aswegen CH, Theron JJ. Putative
melatonin receptor in human spermatozoa. Clin Biochem
1992;25:125–7.
[38] Reiter RJ, Melchiorri D, Sewerynek E, Poeggeler B, Barlow-
Walden L, Chuang J, et al. A review of the evidence supporting
melatonin’s role as an antioxidant. J Pineal Res 1995;18:1–11.
[39] Reiter RJ, Guerrero JM, Garcia JJ, Acuna-Castroviejo D. Reac-
tive oxygen intermediates, molecular damage and aging. Rela-
tion to melatonin. Ann NY Acad Sci 1998;20:410–24.
[40] Jones R, Mann T, Sherins R. Peroxidative breakdown of phos-
pholipids in human spermatozoa, spermicidal properties of fatty
acid peroxides and protective action of seminal plasma. Fertil
Steril 1979;31:531–7.
[41] Lewis SEM, Sterling ESL, Young IS, Thompson W. Comparison
of individual antioxidant of sperm and seminal plasma in fertile
and infertile men. Fertil Steril 1997;67:142–7.
[42] Kaya A, Aksoy M, Baspinar N, Yildrz C, Ataman MB. Effect of
melatonin implantation to sperm donor rams on post-thaw
viability and acrosomal integrity of sperm cells in the breeding
and non-breeding season. Reprod Dom Anim 2001;36:211–5.
[43] Rosa HJD, Juniper DT, Bryant MJ. Effects of recent sexual
experience and melatonin treatment of rams on plasma testos-
terone concentration, sexual behaviour and ability to induce
ovulation in seasonally anoestrous ewes. J Reprod Fertil
2000;120:169–76.
[44] Sheikheldin MA, Howland BE, Palmer WM. Seasonal profiles of
melatonin in adult rams. J Pineal Res 1992;12:58–63.
[45] Schwartz HL, Oppenheimer JH. Physiologic and biochemical
actions of thyroid hormone. Pharmacol Ther 1978;3:349–76.
[46] Venditti P, Balestrieri M, Di Meo S, De Leo T. Effect of thyroid
state on lipid peroxidant, antioxidant defenses, and susceptibility
to oxidative stress in rat tissues. J Endocrinol 1997;155:151–7.
[47] Garcia JJ, Reiter RJ, Ortiz GG. Melatonin enhances tamoxifen’s
ability to prevent the reduction in microsomal membrane fluidity
induced by lipid peroxidation. J Membr Biol 1998;162:59–65.
T.A. Ramadan et al. / Theriogenology 71 (2009) 458–468468




















