
Effect of Variation for Major Growth Habit Genes on Maturity and Yield in Five Spring Wheat...
10
RESEARCH CROP SCIENCE, VOL. 49, JULY– AUGUST 2009 WWW.CROPS.ORG 1211 A few major genes control large portions of the variation for growth habit traits in wheat (Triticum aestivum L. em. Thell.). These include genes involved in vernalization, photoperiod response, and plant stature. Several of these major genes con- trolling growth habit have recently been cloned and sequenced (Beales et al., 2007; Ellis et al., 2002; Sherman et al., 2004; Yan et al., 2006), allowing the development of molecular markers useful for assaying alleles present in breeding germplasm. Knowledge of allelic variation at major loci controlling growth habit, and the effects of alternative alleles on important traits, may be useful for directing selection during the breeding process. One of the major growth habit distinctions in wheat relates to response to vernalization, which controls the differentiation of wheat into winter and spring habits. Winter wheat requires a cold period to induce flowering, while spring wheat flowers without cold treatment. Winter and spring habit are controlled by three major loci, referred to as Vrn-A1, Vrn-B1, and Vrn-D1 located on chromosomes 5A, 5B, and 5D, respectively (Stelmakh, 1993; Zhang et al., 2008). The spring habit alleles at these loci are dominant and are epistatic to the winter alleles at other loci. Thus, winter wheat requires the winter-habit allele at all three loci. The spring Vrn-A1 allele provides complete insensitivity to vernalization, while the Effect of Variation for Major Growth Habit Genes on Maturity and Yield in Five Spring Wheat Populations N. K. Blake, S. P. Lanning, J. M. Martin, M. Doyle, J. D. Sherman, Y. Naruoka, and L. E. Talbert* ABSTRACT Segregation at major genes controlling plant height, photoperiod response, and vernaliza- tion response in wheat (Triticum aestivum L. em. Thell.) may have pleiotropic effects on several traits. Allelic variation at these loci can be moni- tored using polymerase chain reaction (PCR) markers. The effect of segregation at these loci on maturity and agronomic traits was measured for sets of recombinant inbred lines (RIL) devel- oped from five crosses between spring wheat lines adapted to Montana and the Northern Great Plains. Results from field trials grown in multiple years in Bozeman, MT, showed that variation at Rht-B1 and Rht-D1 had large effects on most agronomic traits in two populations, including grain yield, grain protein concentra- tion, and test weight. Variation at photoperiod response loci Ppd-B1 and Ppd-D1 influenced both heading date and date of flag leaf senes- cence under field conditions for two of three populations in which they were segregating. The insensitive allele at Ppd-B1 was associated with higher grain yield in one population. Varia- tion at Vrn-B1 had a significant effect on head- ing date in the field for both populations in which it was segregating, with spring allele lines being earlier. Our results suggest that variation at Rht loci impacting plant height had large pleiotropic effects. Variation at photoperiod and vernaliza- tion loci impacted maturity characteristics but had less consistent effects on economic char- acteristics such as grain yield, test weight, and grain protein concentration. Dep. of Plant Sciences and Plant Pathology, Montana State Univ., Bozeman, MT 59717. Received 25 Aug. 2008. *Corresponding author ([email protected]). Abbreviations: CAPS, cleaved amplified polymorphic sequence; PCR, polymerase chain reaction; PI, photoperiod insensitive; PS, pho- toperiod sensitive; RIL, recombinant inbred lines; SNP, single nucle- otide polymorphism. Published in Crop Sci. 49:1211–1220 (2009). doi: 10.2135/cropsci2008.08.0505 © Crop Science Society of America 677 S. Segoe Rd., Madison, WI 53711 USA All rights reserved. No part of this periodical may be reproduced or transmitted in any form or by any means, electronic or mechanical, including photocopying, recording, or any information storage and retrieval system, without permission in writing from the publisher. Permission for printing and for reprinting the material contained herein has been obtained by the publisher.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Effect of Variation for Major Growth Habit Genes on Maturity and Yield in Five Spring Wheat...

RESEARCH
CROP SCIENCE, VOL. 49, JULY–AUGUST 2009 WWW.CROPS.ORG 1211
A few major genes control large portions of the variation for growth habit traits in wheat (Triticum aestivum L. em. Thell.).
These include genes involved in vernalization, photo period response, and plant stature. Several of these major genes con-trolling growth habit have recently been cloned and sequenced (Beales et al., 2007; Ellis et al., 2002; Sherman et al., 2004; Yan et al., 2006), allowing the development of molecular markers useful for assaying alleles present in breeding germplasm. Knowledge of allelic variation at major loci controlling growth habit, and the eff ects of alternative alleles on important traits, may be useful for directing selection during the breeding process.
One of the major growth habit distinctions in wheat relates to response to vernalization, which controls the diff erentiation of wheat into winter and spring habits. Winter wheat requires a cold period to induce fl owering, while spring wheat fl owers without cold treatment. Winter and spring habit are controlled by three major loci, referred to as Vrn-A1, Vrn-B1, and Vrn-D1 located on chromosomes 5A, 5B, and 5D, respectively (Stelmakh, 1993; Zhang et al., 2008). The spring habit alleles at these loci are dominant and are epistatic to the winter alleles at other loci. Thus, winter wheat requires the winter-habit allele at all three loci. The spring Vrn-A1 allele provides complete insensitivity to vernalization, while the
Eff ect of Variation for Major Growth Habit Genes on Maturity and Yield in Five Spring Wheat Populations
N. K. Blake, S. P. Lanning, J. M. Martin, M. Doyle, J. D. Sherman, Y. Naruoka, and L. E. Talbert*
ABSTRACT
Segregation at major genes controlling plant
height, photoperiod response, and vernaliza-
tion response in wheat (Triticum aestivum L. em.
Thell.) may have pleiotropic effects on several
traits. Allelic variation at these loci can be moni-
tored using polymerase chain reaction (PCR)
markers. The effect of segregation at these loci
on maturity and agronomic traits was measured
for sets of recombinant inbred lines (RIL) devel-
oped from fi ve crosses between spring wheat
lines adapted to Montana and the Northern
Great Plains. Results from fi eld trials grown in
multiple years in Bozeman, MT, showed that
variation at Rht-B1 and Rht-D1 had large effects
on most agronomic traits in two populations,
including grain yield, grain protein concentra-
tion, and test weight. Variation at photoperiod
response loci Ppd-B1 and Ppd-D1 infl uenced
both heading date and date of fl ag leaf senes-
cence under fi eld conditions for two of three
populations in which they were segregating.
The insensitive allele at Ppd-B1 was associated
with higher grain yield in one population. Varia-
tion at Vrn-B1 had a signifi cant effect on head-
ing date in the fi eld for both populations in which
it was segregating, with spring allele lines being
earlier. Our results suggest that variation at Rht
loci impacting plant height had large pleiotropic
effects. Variation at photoperiod and vernaliza-
tion loci impacted maturity characteristics but
had less consistent effects on economic char-
acteristics such as grain yield, test weight, and
grain protein concentration.
Dep. of Plant Sciences and Plant Pathology, Montana State Univ.,
Bozeman, MT 59717. Received 25 Aug. 2008. *Corresponding author
Abbreviations: CAPS, cleaved amplifi ed polymorphic sequence;
PCR, polymerase chain reaction; PI, photoperiod insensitive; PS, pho-
toperiod sensitive; RIL, recombinant inbred lines; SNP, single nucle-
otide polymorphism.
Published in Crop Sci. 49:1211–1220 (2009).doi: 10.2135/cropsci2008.08.0505© Crop Science Society of America677 S. Segoe Rd., Madison, WI 53711 USA
All rights reserved. No part of this periodical may be reproduced or transmitted in any form or by any means, electronic or mechanical, including photocopying, recording, or any information storage and retrieval system, without permission in writing from the publisher. Permission for printing and for reprinting the material contained herein has been obtained by the publisher.

1212 WWW.CROPS.ORG CROP SCIENCE, VOL. 49, JULY–AUGUST 2009
spring Vrn-B1 and Vrn-D1 alleles provide a reduced vernal-ization requirement relative to the winter alleles.
Photoperiod sensitive (PS) wheat genotypes require long days for induction of fl owering, while photoperiod insensitive (PI) genotypes fl ower independently of day-length. Three loci control photoperiod response in wheat (Scarth and Law, 1984; Mohler et al., 2004). These include Ppd-D1, Ppd-B1, and Ppd-A1 on chromosomes 2D, 2B, and 2A, respectively. The dominant alleles at these loci confer insensitivity to photoperiod, and the recessive alleles confer sensitivity. Ppd-D1 has been the most well-studied photo-period response locus and is presumed to be the primary determinant of photoperiod insensitivity (Beales et al., 2007; Dyck et al., 2004; Worland et al., 1998). In Europe, the dominant allele conferring an insensitive reaction (Ppd-D1a) was associated with higher yields in dry, hot regions of southern Europe, while in cool, damp environments of England, PI varieties did not have an advantage (Worland et al., 1994; Worland, 1996). In all environments, PI lines tended to head and mature earlier than PS lines. Several studies in the Northern Great Plains have found that PI lines are earlier to head and shorter than their PS counterparts (Busch et al., 1984; Marshall et al.,1989). Dyck et al. (2004) studied sets of near-isogenic lines of Ppd-D1 at 10 locations in the United States and Canada and found that PS lines were later in heading and maturity, taller, and had a signifi -cantly higher grain yield, particularly in the more northerly locations. The dominant Ppd-B1a allele confers an interme-diate level of insensitivity and was found to control from 23 to 45% of the variation for heading date in a doubled haploid population tested in France (Sourdille et al., 2000).
The major semidwarf genes in spring wheat from the Northern Great Plains are found at the Rht-B1 and Rht-D1 loci on chromosomes 4B and 4D, respectively. Presence of either Rht-B1b or Rht-D1b confers semidwarf growth habit, while presence of both alleles confers a dwarf growth habit. Presence of Rht-B1a and Rht-D1a results in tall plants. Several studies have shown that semidwarf cultivars have higher grain yield potential and reduced lodging under high-input growing conditions (Knott, 1986; Hedden, 2003). McNeal et al. (1972) found that semidwarf wheat lines containing either Rht-B1b or Rht-D1b outyielded tall lines in Montana, except in very low yield environments, where tall lines were superior. Butler et al. (2005) showed that in a population of 140 recombinant inbred lines tested in four Colorado environments varying in yield potential, tall genotypes performed equally to or better than semid-warf types. Within the semidwarf types, lines with Rht-B1b yielded more than lines with Rht-D1b.
The recent molecular cloning of the major loci involved in vernalization response, photoperiod response, and plant height allows effi cient characterization of germplasm. In particular, polymerase chain reaction (PCR) markers were developed for Vrn-A1, Vrn-B1, and Vrn-D1 (Fu et al., 2005;
Sherman et al., 2004; Yan et al., 2004). The photoperiod response loci on chromosome 2 have also been cloned (Beales et al., 2007). Primers for PCR that direct amplifi cation of each of the three genes have been developed. Allelic varia-tion at Ppd-D1 has been shown to cosegregate with photo-period response (Beales et al., 2007). Additionally, sequence variation has been observed at the Ppd-B1 and Ppd-A1 loci, although this variation has not been associated with segrega-tion for photoperiod response (Beales et al., 2007). The Rht-B1 and Rht-D1 loci have also been cloned and PCR markers developed for the mutations that distinguish the functional and nonfunctional alleles (Ellis et al., 2002).
The sum of evidence suggests that alleles at Vrn, Ppd, and Rht loci often have pleiotropic eff ects on agronomi-cally important traits. In this report, we investigated the contribution of allelic variation at the Vrn, Ppd, and Rht loci to genetic variation for several traits in fi ve spring wheat populations developed from crosses between key parents for Montana. Our intent was to estimate the degree to which variation for agronomically important characteristics may be explained by segregation at major growth habit loci. In the course of the studies, a PCR-based marker distinguish-ing sensitive from insensitive alleles at Ppd-B1 in several of our populations was developed. Our results have implica-tions for the development of spring wheat genotypes in the Northern Great Plains of the United States and Canada.
MATERIALS AND METHODS
Population DevelopmentRecombinant inbred lines (RIL) were derived by single-seed
descent starting with the F2 generation from fi ve crosses, including
‘McNeal’/’Thatcher’ (CI 10003), ‘Reeder’/’Conan’, ‘Choteau’/
Reeder, McNeal/Reeder, and MTHW9904/Choteau. The par-
ents, McNeal (Lanning et al., 1994), Reeder (PI 613586), Choteau
(Lanning et al., 2004), and Conan (WestBred, LLC) are all widely
grown hard red spring wheat cultivars in Montana (Montana
Agricultural Statistics, 2007), and MTHW9904 is an experimen-
tal hard white spring wheat. Development of the McNeal/Reeder
population and the MTHW9904/Choteau population were pre-
viously described (Blake et al., 2007; Lanning et al., 2006).
The McNeal/Thatcher population consisted of 80 semi-
dwarf and 80 standard height RIL. To eliminate the eff ects
of shading, the RIL were randomized within eight blocks in
groups of 20 for either the tall or semidwarf genotypes. Reeder
and Conan also diff er for mutant Rht alleles, Rht-B1b and Rht-
D1b, respectively. The Reeder/Conan RIL consisted of dwarf,
semidwarf, and standard height genotypes, initially. For this
study, the dwarf genotypes were eliminated and only semid-
warf and standard height RIL were included.
Collection of Phenotypic DataExperimental design for fi eld trials is summarized in Table 1. All
experiments were grown in Bozeman, MT, at the Post Research
Farm (latitude 45.41° N, longitude 111.00° W, elevation 1455 m)
with plots consisting of 3-m rows seeded at 60 seed m–1 for rainfed
trials and 90 seed m−1 for irrigated trials. Data collected in all RIL
CROP SCIENCE, VOL. 49, JULY–AUGUST 2009 WWW.CROPS.ORG 1213
diagnostic markers for Ppd-A1 and Ppd-B1 in our populations,
genome-specifi c primer sets were developed from these two
loci (Ppd-A1: GenBank accession DQ885753; Ppd-B1: GenBank
accession DQ885757). These primers were used to amplify genic
regions from Reeder, Conan, McNeal, and Choteau, which were
then sequenced following the method of Blake et al. (2004). This
allowed development of allele-specifi c primers for both loci.
A single nucleotide polymorphism (SNP) in the Ppd-B1 locus,
distinguishing Reeder from McNeal and Choteau, was used to
develop an allele-specifi c primer (B6-RDR1F: CCCAATATC-
TACTCCTCCGC) paired with a reverse primer (Ppd2B_SNP5_
R1: TCTGAATGATGATACACCATG) published by Beales et
al. (2007). A diff erent polymorphism distinguished the Ppd-B1
locus in Reeder and Conan, allowing the design of a Reeder
allele-specifi c primer set (ppdB-31F: AGGCTCTTTGGCTAT-
GACGT and ppdB8-R2: ATTCCAACGTTACAAGTGGG).
Polymerase chain reaction products were amplifi ed with both sets
of Ppd-B1 markers using an ABI 2720 thermocycler (Applied Bio-
systems, Foster City, CA) with a touchdown program: 94°C for
5min + 10 cycles (94°C for 20 s, 60°C for 20 s, decrease 0.5°C/
cycle, 72°C for 2 min) + 35 cycles (94°C for 20 s, 50°C for 20 s,
72°C for 2 min) + 72°C for 10 min. Reaction products were
visualized using to 0.15 g mL–1 agarose gels.
A similar strategy was followed to develop a Ppd-A1 allele-
specifi c cleaved amplifi ed polymorphic sequence (CAPS) marker
from an SNP between Reeder and Conan. Amplifi cation with
the primers PpdA-F4: GCAGGGAAAGACAAGGCTGAT-
GAAAC and PpdA-R4: CCGCACTTCTACTATGTACTC-
TACG was performed with a touchdown program: 94°C for
5 min + 10 cycles (94°C for 20 s, 63°C for 20 s, decrease 0.5°C/
cycle, 72°C for 2 min) + 35 cycles (94°C for 20 s, 58°C for 20 s,
72°C for 2 min) + 72°C for 10 min.
Polymerase chain reaction products were digested with
two units of restriction enzyme RsaI. The digested reaction
products were distinguished using 12% polyacrylamide gels.
Statistical AnalysisData for each phenotypic trait in each population were analyzed
via analysis of variance (ANOVA) using a model for a random-
ized block combined over environments using PROC MIXED
nurseries included date of heading, plant height, grain yield, test
weight, and grain protein concentration. Heading date was mea-
sured as the number of days from planting when 50% of the heads
were completely emerged. Plant height was the average of two
measurements made from the soil surface to the top of the spikes,
excluding awns. Grain yield was determined from the raw grain
weight of each plot. Test weight was measured from a sample of
cleaned grain on a Seedburo (Chicago, IL) test weight scale. Grain
protein concentration was obtained on whole grain samples using
an Infratec (Tecator, Höganäs, Sweden). Additionally, date of fl ag
leaf senescence was determined for the McNeal/Thatcher, Reeder/
Conan, McNeal/Reeder, and Choteau/Reeder populations as the
number of days from planting when 75% of the plot exhibited fl ag
leaves showing complete loss of green color. The duration of green
leaf period after heading was determined by subtracting heading
date from the date of fl ag leaf senescence. In addition to the fi eld
data, heading date was measured for parents and RIL grown under
12-hour daylength in a growth chamber. For this experiment, two
replications were tested, with each replication consisting of two to
four plants per line. Growth chamber temperatures ranged from 21
to 22°C and photosynthetic radiation was 580 μmol m−2 s−1. Plants
were grown in Cone-tainer pots (Stuey and Sons, Corvallis, OR)
with a soil capacity of 164 mL. Soil was equal parts (by volume) of
loam soil, washed concrete sand, and sphagnum peat moss.
Montana Advanced Spring Wheat Yield TrialThe trial consisted of 64 lines grown in four-row 3.3-m plots
with three replications grown in 10 locations. Phenotypic data
collected from this nursery pertinent for this report included
heading date and plant height. These lines were also tested for
heading under 12-hour days in the growth chamber using the
same conditions as for the RIL populations.
Genotypic DataPrimer sets used to screen the parents of the fi ve populations have
been previously described (Beales et al., 2007; Ellis et al., 2002;
Sherman et al., 2004; Yan et al., 2006). Loci analyzed included
Vrn-B1, Vrn-D1, Ppd-D1, Rht-B1, and Rht-D1. Polymerase chain
reaction conditions followed published protocols. Primer sets that
identifi ed allelic diff erences between parents were used to geno-
type the appropriate RIL population. Reaction
products were visualized using 1.5% agarose gels.
In addition to testing RIL developed from fi ve
spring wheat crosses, we determined genotypes
at the Ppd, Vrn, and Rht loci for lines tested in
the Montana Advanced Spring Wheat Yield
Trial in 2007. Complete genotyping data was
not obtained for six lines from this trial, so these
were excluded from analyses.
Development of Polymerase Chain Reaction Markers for Ppd-B1 and Ppd-A1In the course of the experiments, it became
apparent that an allele conferring photoperiod
insensitivity not detectable by previously pub-
lished primer sets was segregating in populations
derived from the cultivar Reeder. To develop
Table 1. Recombinant inbred line populations tested for association between
variation at growth habit loci and maturity traits in spring wheat (Triticum aes-
tivum L. em. Thell.).
ParentsGeneration of line derivation
Number of RIL† Design‡ Year of
experimentIrrigation regime
McNeal/Thatcher F6
160 RCB§ 2006 Rainfed, irrigated
2007 Rainfed, irrigated
Reeder/Conan F5
92 RCB 2006 Rainfed
2007 Rainfed
MTHW9904/
Choteau
F5
94 RCB 2004 Rainfed, irrigated
2005 Rainfed, irrigated
McNeal/Reeder F4
50 RCB 2004 Rainfed, irrigated
2005 Rainfed, irrigated
Choteau/Reeder F4
46 Lattice 2007 Rainfed
†Recombinant inbred lines.‡All experiments consisted of three replications, with each plot being a single row of 3.3 m. All experiments
were grown at the Arthur Post Research Farm in Bozeman, MT.§RCB, randomized complete block.

1214 WWW.CROPS.ORG CROP SCIENCE, VOL. 49, JULY–AUGUST 2009
in SAS (SAS Institute, 2003). Genotype and replication were con-
sidered random factors, and environment was considered a fi xed
factor. Entry means averaged over environments and replications
were used for analysis of gene eff ects. Data were combined over
environments (years and rainfed versus irrigated) in that the sums
of squares owing to entry by environment interaction were low
relative to sums of squares owing to main eff ects. A single-factor
ANOVA was conducted, where the single factor was the segregat-
ing locus with two allelic classes using PROC GLM in SAS (SAS
Institute, 2003). Diff erences between allele class means were tested
with an F ratio. This was done for each segregating gene for each
population and in the Montana Advanced Spring Wheat Yield
Trial. In populations where more than one locus was segregat-
ing, the model was then extended to include all segregating genes
and their interactions. The proportion of variation attributable to
segregating gene(s) (R2) was computed as the sum of squares for
segregating gene(s) divided by the total sum of squares.
RESULTS
Variation at Growth Habit Loci for Elite Yield Trial LinesTable 2 shows variation among lines for many of the genes important in controlling growth habit in the Montana Advanced Spring Wheat Yield Trial in 2007. Lines within the nursery varied for Vrn-B1, Vrn-D1, Ppd-D1, Ppd-B1, Rht-B1, and Rht-D1. There were 38 and 21 lines with the spring and winter alleles at Vrn-B1, respectively. An F-test showed that the lines with the spring version of Vrn-B1 headed almost 1 d earlier (P = 0.05) than lines with the winter allele (data not shown). There were only eight lines with the spring allele at Vrn-D1, and no signifi cant diff erences were detected among lines with alternate alleles at this locus. Eleven lines contained the insensitive allele at Ppd-D1, and these lines headed 1 d earlier (P = 0.06) and were shorter than lines with the sensitive allele at this locus (data not shown). The frac-tion of lines that were semidwarf was 52/59, with 21 and 31 having reduced height alleles at Rht-B1 and Rht-D1, respec-tively. Comparison of genotype means was not conducted for height, in that the standard height lines were all unimproved cultivars compared with the semidwarf lines.
Development of Polymerase Chain Reaction Markers for Ppd-B1 and Ppd-A1
It was apparent that several PI lines in the Montana Advanced Spring Wheat Yield Trial and in the segregating populations did not contain the insensitive allele at the Ppd-D1 locus. This was particularly the case for populations containing Reeder spring wheat as a parent. Thus, we wished to obtain a PCR marker for the Reeder allele conferring insensitivity to pho-toperiod with which to assay the segregating populations.
McNeal and Choteau are both PS, while Reeder is PI. All three lines contain the sensitive allele at Ppd-D1 as deter-mined by PCR assay. The McNeal/Reeder and Choteau/Reeder populations both segregated in an approximate 1:1 ratio for sensitive versus insensitive types; 20 vs. 30 and 21 vs. 25, respectively. Beales et al. (2007) reported PCR assays for allelic variation at Ppd-A1 and Ppd-B1 amongst a set of European wheat lines. Reeder, McNeal, and Choteau were not polymorphic for any of these PCR assays. One SNP was revealed in 7000 bases of sequence that diff eren-tiated Reeder from both McNeal and Choteau at Ppd-B1. A marker developed from the Reeder SNP was used for analyses of the segregating populations. The product ampli-fi ed from Reeder was approximately 1350 bp.
A similar issue was found in the Reeder/Conan popu-lation. Both Conan and Reeder are PI (data not shown). However, 25 of the 92 RIL developed from the Reeder/Conan cross were PS, suggesting that the lines had insensi-tive alleles at diff erent loci. Screening of the parents showed that Conan had the insensitive allele at Ppd-D1, while Reeder had the sensitive allele. Thus, Reeder was apparently insen-sitive owing to alleles at a diff erent locus. Reeder and Conan did not diff er for the published polymorphisms at Ppd-A1 or Ppd-B1 or for the Ppd-B1 marker we developed to distin-guish Reeder from Choteau and McNeal. One SNP among 13,000 bases was identifi ed between Reeder and Conan for Ppd-B1 that amplifi ed a product of approximately 1500 bp in Reeder and not Conan. This primer set was used to ana-lyze all RIL in the Reeder/Conan population.
Similarly, we developed an allele-specifi c CAPS marker for the Ppd-A1 locus after sequencing 2000 bases from Reeder and Conan. The marker yielded three banding pat-terns that distinguished parents of all populations except McNeal/Thatcher. Thus, the primer set could be used to determine phenotypic associations with sequence varia-tion at the Ppd-A1 locus. Reeder and Choteau had diff erent banding patterns, while McNeal, Conan, and MTHW9904 shared a pattern distinct from the other two cultivars.
Association of Phenotypic Variation with Allelic State for Growth Habit Loci in Five Spring Wheat PopulationsFive spring wheat populations were available with which to analyze the eff ects of alternative alleles at the growth habit loci on maturity and economic traits. Table 3 shows that
Table 2. Allelic variation at major growth habit loci among lines
entered in the 2007 Montana Advanced Spring Wheat (Triti-
cum aestivum L. em. Thell.) Yield Trial.
Locus Allele Number of lines
Vrn-B1Spring (Vrn-B1) 38
Winter (vrn-B1) 21
Vrn-D1Spring (Vrn-D1) 8
Winter (vrn-D1) 51
Ppd-D1Insensitive (Ppd-D1a) 11
Sensitive (Ppd-D1b) 48
Rht-B1Wild-type (Rht-B1a) 38
Mutant (Rht-B1b) 21
Rht-D1Wild-type (Rht-D1a) 28
Mutant (Rht-D1b) 31

CROP SCIENCE, VOL. 49, JULY–AUGUST 2009 WWW.CROPS.ORG 1215
genotype eff ects were signifi cant for all traits in all popula-tions, providing an opportunity to test the phenotypic eff ect of alternative alleles at major growth habit loci. The popula-tions were not all grown in the same years (Table 1), so that comparison between populations is not possible. Heading date under 12-hour daylength was determined for all popu-lations in a growth chamber experiment. Three of the popu-lations showed clear demarcation between lines that fl owered early (photoperiod insensitive) and lines that fl owered late (photoperiod sensitive). Two populations (McNeal/Thatcher and MTHW9904/Choteau) had no lines that headed as early as the PI lines in the other populations, suggesting that RIL in these populations were all PS. The parents were screened for variation at the major growth habit loci, including Vrn-B1, Vrn-D1, Ppd-A1, Ppd-B1, Ppd-D1, Rht-B1, and Rht-D1. The number of loci containing diff erent alleles in the parents ranged from one (Rht-D1) for McNeal and Thatcher to six for Reeder and Conan. Eff ects of alternative alleles on phe-notypic performance of RIL from the fi ve populations are shown in Table 4 and Table 5 and are presented below.
McNeal/Thatcher
McNeal is a semidwarf cultivar owing to Rht-D1b, while Thatcher is standard height. The RIL population consisted of 80 semidwarf and 80 standard height progeny. Alleles at Rht-D1 had a major eff ect on most traits, including fi eld heading date, 12-hour heading date, green leaf duration after heading, plant height, yield, test weight, and grain protein concentration. Allelic state at Rht-D1 accounted for 93% of the height variation in the population, 67% of grain yield variation, 14% of test weight variation, 11% of
heading date variation, and 15% of the variation for green leaf duration after heading (Table 5).
Reeder/Conan
This population was segregating for six of the major growth habit loci. Alternative alleles at the Vrn-B1 locus aff ected maturity traits, with the spring allele conferring earlier head-ing (P = 0.07) and longer green leaf duration after heading (P = 0.01). The population was segregating for all three Ppd loci, based on sequence variation, but only Ppd-B1 and Ppd-D1 impacted maturity traits. Lines with the insensitive Ppd-D1 allele were earlier to head by 3 d in the fi eld (P < 0.001) and had a longer green leaf duration after heading (P < 0.01). These lines headed 13 d earlier under 12-hour daylength (P < 0.001). Test weight was higher for the lines with the insensi-tive Ppd-D1 allele. The eff ect of the Reeder allele at Ppd-B1 similarly decreased both fi eld and 12-hour heading date (P = 0.03 and < 0.001, respectively), indicating it to be the insen-sitive allele. This allele also accelerated fl ag leaf senescence by about a day (P < 0.01). Allelic state at the Ppd-A1 locus had no eff ect on maturity traits in this population but was associated with increased protein concentration (P < 0.01). Cumulatively, alleles at Ppd-B1 and Ppd-D1 controlled 78 and 44% of the variation for 12-hour heading date and fi eld heading date, respectively. There was a signifi cant interac-tion between Ppd-B1 and Ppd-D1 (P < .0001) for 12-hour heading. Lines with both sensitive alleles headed 19 d later than lines with both insensitive alleles. Lines insensitive for Ppd-D1 and sensitive for Ppd-B1 headed 1 d later than lines with both insensitive alleles and almost 6 d earlier than lines sensitive for Ppd-D1 and insensitive for Ppd-B1. Along with
Table 3. Mean and range for maturity traits† in fi ve spring wheat (Triticum aestivum L. em. Thell.) populations.
Poulation HD—12 h HD—fi eld FLS GLDAH Plant height Test weight Grain yield Grain protein
————— d————— d from Jan. 1 d cm kg m−3 kg ha–1 g kg–1
McNeal/Thatcher Mean 76.9 63.4 206.8 27.4 93.5 737 3608 154
Range 61–100‡ 59.2–66.6 204.5–208.9 24.9–30 71.2–114.9 647–737 2439–4770 138–172
P§ <0.0001 <0.0001 <0.0001 <0.0001 <0.0001 <0.0001 <0.0001 <0.0001
Reeder/Conan Mean 58.5 61.3 210.9 32.6 92.3 750 3467 160
Range 47.5–82 56–68 206–216 27.5–36.7 70.5–111.8 696–789 1632–4243 145–179
P <0.0001 <0.0001 <0.0001 <0.0001 <0.0001 <0.0001 <0.0001 <0.0001
MTHW9904/
Choteau
Mean 85.3 70.4 ND¶ ND 96.3 776 5249 155
Range 66.5–99 66.3–74.5 73.5–125.3 726–809 3097–8869 129–176
P <0.0001 <0.0001 <0.0001 <0.0001 <0.0001 <0.0001
McNeal/Reeder Mean 62.4 71.6 218.9 34.3 99.0 773 5776 147
Range 49.5–81.5 68.3–76.6 215.7–222.4 31.8–37.6 72.8–121.7 648–792 4931–6833 137–161
P <0.0001 <0.0001 <0.0001 <0.0001 <0.0001 <0.0001 <0.0001 <0.0001
Reeder/Choteau Mean 62.5 65.9 212.4 31.5 86.8 780 4804 147
Range 47–82 61.7–68.7 209.8–214 28.1–34.1 77.8–92.5 748–811 3837–5778 132–161
P <0.0001 <0.0001 <0.0001 <0.0001 <0.0001 <0.0001 0.01 <0.0001
†HD, heading date; FLS, day of fl ag leaf senescence; GLDAH, green leaf duration after heading.‡Experiment was terminated after 100 d. Two RIL had not headed at that time.§P, genotype P value.¶These traits were not determined in this population.
1216 WWW.CROPS.ORG CROP SCIENCE, VOL. 49, JULY–AUGUST 2009
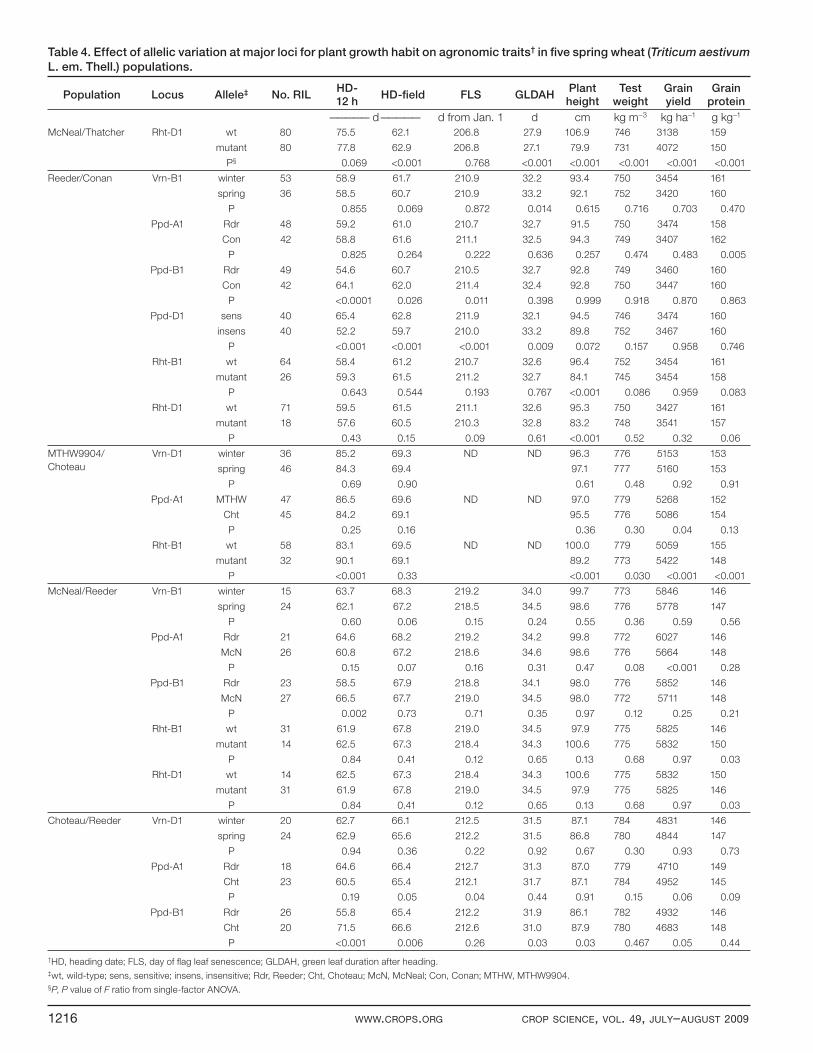
Table 4. Effect of allelic variation at major loci for plant growth habit on agronomic traits† in fi ve spring wheat (Triticum aestivum
L. em. Thell.) populations.
Population Locus Allele‡ No. RILHD-12 h
HD-fi eld FLS GLDAH Plant height
Test weight
Grain yield
Grain protein
————— d————— d from Jan. 1 d cm kg m–3 kg ha–1 g kg–1
McNeal/Thatcher Rht-D1 wt 80 75.5 62.1 206.8 27.9 106.9 746 3138 159
mutant 80 77.8 62.9 206.8 27.1 79.9 731 4072 150
P§ 0.069 <0.001 0.768 <0.001 <0.001 <0.001 <0.001 <0.001
Reeder/Conan Vrn-B1 winter 53 58.9 61.7 210.9 32.2 93.4 750 3454 161
spring 36 58.5 60.7 210.9 33.2 92.1 752 3420 160
P 0.855 0.069 0.872 0.014 0.615 0.716 0.703 0.470
Ppd-A1 Rdr 48 59.2 61.0 210.7 32.7 91.5 750 3474 158
Con 42 58.8 61.6 211.1 32.5 94.3 749 3407 162
P 0.825 0.264 0.222 0.636 0.257 0.474 0.483 0.005
Ppd-B1 Rdr 49 54.6 60.7 210.5 32.7 92.8 749 3460 160
Con 42 64.1 62.0 211.4 32.4 92.8 750 3447 160
P <0.0001 0.026 0.011 0.398 0.999 0.918 0.870 0.863
Ppd-D1 sens 40 65.4 62.8 211.9 32.1 94.5 746 3474 160
insens 40 52.2 59.7 210.0 33.2 89.8 752 3467 160
P <0.001 <0.001 <0.001 0.009 0.072 0.157 0.958 0.746
Rht-B1 wt 64 58.4 61.2 210.7 32.6 96.4 752 3454 161
mutant 26 59.3 61.5 211.2 32.7 84.1 745 3454 158
P 0.643 0.544 0.193 0.767 <0.001 0.086 0.959 0.083
Rht-D1 wt 71 59.5 61.5 211.1 32.6 95.3 750 3427 161
mutant 18 57.6 60.5 210.3 32.8 83.2 748 3541 157
P 0.43 0.15 0.09 0.61 <0.001 0.52 0.32 0.06
MTHW9904/
Choteau
Vrn-D1 winter 36 85.2 69.3 ND ND 96.3 776 5153 153
spring 46 84.3 69.4 97.1 777 5160 153
P 0.69 0.90 0.61 0.48 0.92 0.91
Ppd-A1 MTHW 47 86.5 69.6 ND ND 97.0 779 5268 152
Cht 45 84.2 69.1 95.5 776 5086 154
P 0.25 0.16 0.36 0.30 0.04 0.13
Rht-B1 wt 58 83.1 69.5 ND ND 100.0 779 5059 155
mutant 32 90.1 69.1 89.2 773 5422 148
P <0.001 0.33 <0.001 0.030 <0.001 <0.001
McNeal/Reeder Vrn-B1 winter 15 63.7 68.3 219.2 34.0 99.7 773 5846 146
spring 24 62.1 67.2 218.5 34.5 98.6 776 5778 147
P 0.60 0.06 0.15 0.24 0.55 0.36 0.59 0.56
Ppd-A1 Rdr 21 64.6 68.2 219.2 34.2 99.8 772 6027 146
McN 26 60.8 67.2 218.6 34.6 98.6 776 5664 148
P 0.15 0.07 0.16 0.31 0.47 0.08 <0.001 0.28
Ppd-B1 Rdr 23 58.5 67.9 218.8 34.1 98.0 776 5852 146
McN 27 66.5 67.7 219.0 34.5 98.0 772 5711 148
P 0.002 0.73 0.71 0.35 0.97 0.12 0.25 0.21
Rht-B1 wt 31 61.9 67.8 219.0 34.5 97.9 775 5825 146
mutant 14 62.5 67.3 218.4 34.3 100.6 775 5832 150
P 0.84 0.41 0.12 0.65 0.13 0.68 0.97 0.03
Rht-D1 wt 14 62.5 67.3 218.4 34.3 100.6 775 5832 150
mutant 31 61.9 67.8 219.0 34.5 97.9 775 5825 146
P 0.84 0.41 0.12 0.65 0.13 0.68 0.97 0.03
Choteau/Reeder Vrn-D1 winter 20 62.7 66.1 212.5 31.5 87.1 784 4831 146
spring 24 62.9 65.6 212.2 31.5 86.8 780 4844 147
P 0.94 0.36 0.22 0.92 0.67 0.30 0.93 0.73
Ppd-A1 Rdr 18 64.6 66.4 212.7 31.3 87.0 779 4710 149
Cht 23 60.5 65.4 212.1 31.7 87.1 784 4952 145
P 0.19 0.05 0.04 0.44 0.91 0.15 0.06 0.09
Ppd-B1 Rdr 26 55.8 65.4 212.2 31.9 86.1 782 4932 146
Cht 20 71.5 66.6 212.6 31.0 87.9 780 4683 148
P <0.001 0.006 0.26 0.03 0.03 0.467 0.05 0.44
†HD, heading date; FLS, day of fl ag leaf senescence; GLDAH, green leaf duration after heading.‡wt, wild-type; sens, sensitive; insens, insensitive; Rdr, Reeder; Cht, Choteau; McN, McNeal; Con, Conan; MTHW, MTHW9904.§P, P value of F ratio from single-factor ANOVA.
CROP SCIENCE, VOL. 49, JULY–AUGUST 2009 WWW.CROPS.ORG 1217
variation at Rht-D1, these loci accounted for 39% of the variation for date of fl ag leaf senescence.
The Reeder/Conan RIL had two height phenotypes, semidwarf and standard height. The semidwarf lines resulted from the mutant allele at either Rht-B1 or Rht-D1, while the standard height lines were wild-type at both loci. Compar-isons of semidwarf lines (either Rht-B1b or Rht-D1b) with standard height lines showed that semidwarf lines were sig-nifi cantly shorter, with lower protein concentration and test weight (data not shown). This is also refl ected in the single locus comparisons in Table 4, as lines with the mutant allele at either Rht-B1 or Rht-D1 were shorter and had lower grain protein concentration than lines with the respective wild-type allele. In combination with variation at Ppd-D1, 65% of height variation was explained by segregation at these loci.
MTHW9904/Choteau
The MTHW9904/Choteau population was segregating at Vrn-D1 and Rht-B1. There was no measurable eff ect of the alternative alleles at Vrn-D1. The mutant allele at Rht-B1 (Rht-B1b) resulted in signifi cantly shorter plants (P < 0.001), lower test weight (P = 0.03), higher grain yield (P < 0.001), lower grain protein concentration (P < 0.001), and later heading under 12-hour days (P < 0.001). This population did not vary for either Ppd-D1 or Ppd-B1, as is refl ected in the late heading date for all RIL in this population under
12-hour days. The MTHW9904 allele for Ppd-A1 increased yield (P = 0.04) but did not impact other traits. Cumula-tively, variation at Rht-B1 and Ppd-A1 explained 22% of the variation for yield in this population.
McNeal/Reeder
The McNeal/Reeder population varied at Vrn-B1, Ppd-A1, Ppd-B1, Rht-B1, and Rht-D1 (Table 4). In contrast to the Reeder/Conan population, all RIL were semidwarf with no standard height lines tested in the population. As with the Reeder/Conan RIL, the Vrn-B1 spring allele tended to cause earlier heading in the fi eld (P = 0.06). The insen-sitive Ppd-B1 allele contributed by Reeder had a major eff ect only on heading date under 12-hour days (P < 0.01). The alternative alleles at Ppd-A1 had a measurable eff ect, with lines containing the Reeder allele heading 1 d later (P = 0.07), yielding signifi cantly more (P < 0.001) with lower test weight (P = 0.08) than lines with the McNeal allele (Table 4). Allelic state at Ppd-A1 accounted for 7% of the variation for test weight and 23% of the variation for grain yield (Table 5). The cumulative eff ects of alleles at Vrn-B1 and Ppd-A1 described 18% of the variation for heading date in the fi eld. Eff ects of the Rht alleles were modest, although the Rht-B1b lines had signifi cantly higher grain protein concentration (P = 0.03), accounting for 9% of variance for this trait.
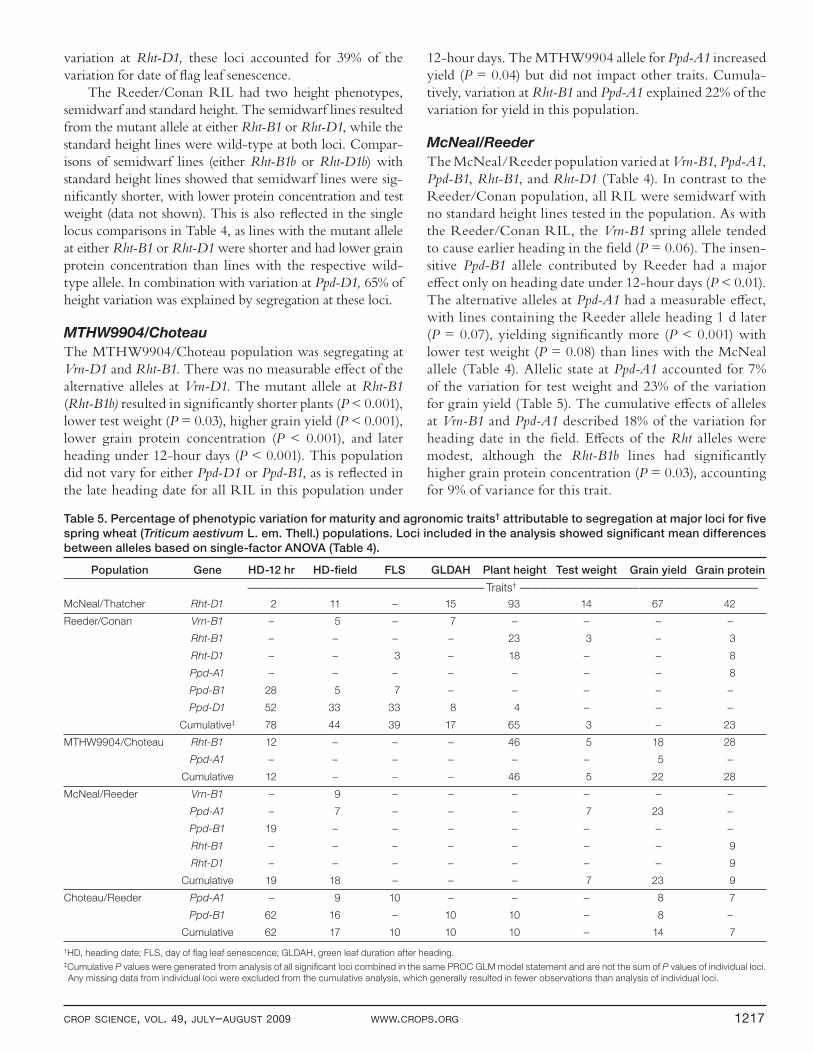
Table 5. Percentage of phenotypic variation for maturity and agronomic traits† attributable to segregation at major loci for fi ve
spring wheat (Triticum aestivum L. em. Thell.) populations. Loci included in the analysis showed signifi cant mean differences
between alleles based on single-factor ANOVA (Table 4).
Population Gene HD-12 hr HD-fi eld FLS GLDAH Plant height Test weight Grain yield Grain protein
—————————————————————————————— Traits† ——————————————— ———————————————
McNeal/Thatcher Rht-D1 2 11 – 15 93 14 67 42
Reeder/Conan Vrn-B1 – 5 – 7 – – – –
Rht-B1 – – – – 23 3 – 3
Rht-D1 – – 3 – 18 – – 8
Ppd-A1 – – – – – – – 8
Ppd-B1 28 5 7 – – – – –
Ppd-D1 52 33 33 8 4 – – –
Cumulative‡ 78 44 39 17 65 3 – 23
MTHW9904/Choteau Rht-B1 12 – – – 46 5 18 28
Ppd-A1 – – – – – – 5 –
Cumulative 12 – – – 46 5 22 28
McNeal/Reeder Vrn-B1 – 9 – – – – – –
Ppd-A1 – 7 – – – 7 23 –
Ppd-B1 19 – – – – – – –
Rht-B1 – – – – – – – 9
Rht-D1 – – – – – – – 9
Cumulative 19 18 – – – 7 23 9
Choteau/Reeder Ppd-A1 – 9 10 – – – 8 7
Ppd-B1 62 16 – 10 10 – 8 –
Cumulative 62 17 10 10 10 – 14 7
†HD, heading date; FLS, day of fl ag leaf senescence; GLDAH, green leaf duration after heading.‡Cumulative P values were generated from analysis of all signifi cant loci combined in the same PROC GLM model statement and are not the sum of P values of individual loci.
Any missing data from individual loci were excluded from the cumulative analysis, which generally resulted in fewer observations than analysis of individual loci.

1218 WWW.CROPS.ORG CROP SCIENCE, VOL. 49, JULY–AUGUST 2009
Choteau/Reeder
The Choteau/Reeder population was segregating at Vrn-D1, Ppd-A1, and Ppd-B1 (Table 4). Alternative alleles at Vrn-D1 had no measurable eff ect on phenotype. Alterna-tive alleles at Ppd-A1 impacted heading date, as lines with the Reeder allele headed 1 d later in the fi eld (P = 0.05) and 4 d later under 12-hour conditions. The Ppd-A1 Reeder allele was associated with lower yield, in contrast to its eff ect in the McNeal/Reeder population. However, Choteau and McNeal have diff erent alleles at this locus. Variation at Ppd-B1 had a major eff ect on most maturity traits. Heading date in the fi eld was 1 d earlier (P < 0.01) and almost 16 d earlier under 12-hour days (P < 0.001) in lines with the Reeder allele. Lines with the Reeder allele were shorter (P = 0.03), had longer green leaf dura-tion after heading (P = 0.03), and greater yield (P = 0.05). The cumulative eff ect of variation at Ppd-A1 and Ppd-B1 accounted for 17% and 14% of the variation for heading date and grain yield, respectively (Table 5).
DISCUSSIONMajor genes controlling growth habit response in wheat have been well characterized genetically. These include Vrn genes that infl uence vernalization response, Ppd genes that control photoperiod response, and Rht genes that infl uence plant height. Allelic variants for these loci may have pleiotropic eff ects that infl uence a range of economic traits (Butler et al., 2005; Dyck et al., 2004; Iqbal et al., 2007; Sourdille et al., 2000; Zhang et al., 2008). Deter-mination of alleles at Vrn, Ppd, and Rht loci is possible owing to the recent development of PCR-based markers for major genes controlling wheat development and matu-rity. Many of these markers are diagnostic across a range of germplasm, such as the Rht genes that confer semidwarf habit, the Vrn genes, and the Ppd-D1 gene determining photoperiod sensitivity. The Ppd-A1 and Ppd-B1 genes have also been cloned, and PCR primers exist to pref-erentially amplify these genes. However, the published polymorphisms at these loci were not present in our ger-mplasm. Our results showed that a photoperiod response gene in addition to Ppd-D1 was present in our material, especially in lines derived from the cultivar Reeder. Thus an initial objective was to develop PCR-based markers to allow us to distinguish Reeder from other parents in RIL populations. Two diff erent primer sets were developed for Ppd-B1 in this regard, one that identifi ed an SNP distin-guishing Reeder from Choteau and McNeal and one that distinguished Reeder from Conan. Additionally, an SNP-based primer set was developed to diff erentiate Ppd-A1 in parents of four of the populations.
Screening of the germplasm in the Montana Advanced Spring Wheat Yield Trial showed variation for most of the major growth-related loci. In particular, the nursery contained both semidwarf and standard height lines, with
semidwarf lines resulting from either Rht-B1b or Rht-D1b. In the McNeal/Thatcher population, lines with semidwarf growth habit resulting from Rht-D1b yielded approximately 30% more than standard height lines. In the Reeder/Conan populations, semidwarf lines showed no signifi cant yield advantage (data not shown). The yield advantage for semid-warf lines was 7% in the MTHW9904/Choteau population. Thus, with the genotypes and environments in this experi-ment, semidwarf growth appears to be favorable. Higher grain yield potential has been observed under high-input growing conditions (Knott, 1986; Hedden, 2003; McNeal et al., 1972) for semidwarf lines in previous experiments. However, McNeal et al. (1972) found that tall lines were superior in low-yield environments. Butler et al. (2005) found no advantage to semidwarf genotypes in a popula-tion of 140 recombinant inbred lines tested in four Colo-rado environments varying in yield potential. Both rainfed and irrigated environments used for the present study rep-resent high water inputs relative to spring wheat production in most of the Great Plains.
There was also variation at photoperiod response loci among lines in the Montana Advanced Spring Wheat Yield Trial. Eleven lines had the insensitive allele at Ppd-D1. Sev-eral additional lines were apparently insensitive to photo-period based on 12-hour heading date (data not shown), although there was not a clear phenotypic demarcation, as observed in the RIL populations. However, a preponder-ance of the lines that headed relatively early under 12-hour days had the Reeder nucleotide for the Ppd-B1 SNP marker, suggesting that they contained the insensitive allele at Ppd-B1. This conjecture is supported by pedigree data, as over one-half of these lines had Reeder as a parent or grandpar-ent. However, the Reeder SNP is not completely diagnos-tic for the insensitive allele, as some sensitive alleles (as in Thatcher) had the Reeder nucleotide at this SNP site. This indicates that the Reeder SNP is unlikely to be the causal mutation for insensitive photoperiod response.
The insensitive allele at Ppd-D1 was segregating only in the Reeder/Conan population and had a major eff ect on heading date under 12-hour days. The insensitive allele at Ppd-D1 was associated with earlier heading under fi eld conditions and shorter stature similar to eff ects shown in separate populations (Dyck et al., 2004). Earlier heading of insensitive genotypes under fi eld conditions refl ects their ability to initiate fl owering before the onset of long days in the summer. No signifi cant eff ects on grain yield were observed in this population.
The insensitive allele at Ppd-B1 had major phenotypic eff ects in our populations. This allele was associated with earlier heading both in the fi eld and under 12-hour days in Reeder/Conan and Choteau/Reeder (Table 4). It also decreased 12-hour day heading in McNeal/Reeder by 8 d, though it had no eff ect on heading date in the fi eld. Flag leaf senescence was 1 d earlier in Reeder/Conan lines with

CROP SCIENCE, VOL. 49, JULY–AUGUST 2009 WWW.CROPS.ORG 1219
the insensitive Reeder allele, though not signifi cantly earlier in McNeal/Reeder and Choteau/Reeder lines. Green leaf duration after heading was signifi cantly increased by slightly less than a day in Choteau/Reeder lines with the insensitive Reeder allele. Interestingly, the insensitive allele at Ppd-B1 introduced from Reeder appears to be the prevalent cause of photoperiod insensitivity in the Montana germplasm pool.
An interesting eff ect was seen with variation at the Ppd-A1 allele identifi ed by sequence variation between parents involved in the crosses. Sequence diff erences allowed distinc-tion between alleles for the Reeder/Conan, MTHW9904/Choteau, McNeal/Reeder, and Choteau/Reeder RIL. Although fi eld heading date was impacted in two popula-tions, in no case was variation at Ppd-A1 signifi cantly associ-ated with heading date under 12-hour days. This may refl ect previous observations that Ppd-A1 has a weak eff ect on pho-toperiod response relative to Ppd-B1 and Ppd-D1 (Worland et al., 1994). However, in two of the crosses, allelic variation at Ppd-A1 was associated with grain yield. Whether this is owing to an eff ect of a Ppd-A1 mutation linked to the SNP marker or to linkage to another gene is not known.
All Montana spring wheat lines have the spring allele at Vrn-A1 (Sherman et al., 2004). Recombinant inbred line populations segregating for Vrn-D1 were Choteau/Reeder and MTHW9904/Choteau. There was no eff ect of the alternative alleles on any trait in either population. The Reeder/Conan and McNeal/Reeder populations were both segregating at Vrn-B1. In both populations, the spring allele resulted in earlier heading. Similarly, Iqbal et al. (2007) found that Canadian lines with Vrn-B1 tended to be earlier, and Stelmakh (1993) reported that wheat geno-types with spring alleles at all three of the group 5 Vrn loci were early to head in Ukraine. This is also consistent with data from our Montana Spring Wheat Advanced Yield Trial, where lines with the spring allele at Vrn-B1 headed earlier than those with the winter allele.
CONCLUSIONSAllelic variation was observed for Vrn-B1, Vrn-D1, Ppd-B1, Ppd-D1, Rht-B1, and Rht-D1 among lines tested in the Montana Spring Wheat Advanced Yield Trial. Evaluation of fi ve RIL populations, each segregating for subsets of the loci, showed that variation at these major genes had mea-surable eff ects on several traits. As expected, allelic varia-tion at Rht loci explained the predominant proportion of height variation in populations that were segregating at these loci. The spring allele at the Vrn-B1 locus resulted in earlier heading under fi eld conditions in two populations. Alleles for photoperiod insensitivity at either Ppd-B1 or Ppd-D1 caused earlier heading in the fi eld for two of three populations showing segregation at these loci.
The importance of segregation at major genes on phe-notypic variation varied depending on the population. For instance, the Reeder/Conan RIL were notable for showing
segregation for six of the loci being investigated. Segrega-tion at these loci explained a large percentage of variation for heading date (44%), date of fl ag leaf senescence (39%), and green leaf duration after heading (17%). The McNeal/Thatcher RIL only diff ered for alleles at Rht-D1. However, a large percentage of variation in grain yield (67%) and grain protein concentration (42%) was explained by segregation at this locus. In sum, our results suggest that variation at genes for vernalization, plant height, and photoperiod response contribute signifi cantly to phenotypic variation for economi-cally important traits in the sampled germplasm pool.
The existence of diversity at the major growth habit loci may refl ect the fact that the selection environment changes every year, and alternative alleles may be favored depending on conditions. For instance, the recent tran-sition to higher temperatures at heading may favor PI lines that head earlier to avoid heat stress. Additionally, the diversity refl ects continual introduction of new germ-plasm into the breeding program. An example indicated by this experiment is the prevalence of the Ppd-B1 allele from Reeder in the advanced breeding germplasm. Main-tenance of variation for major genes controlling maturity and growth habit may allow rapid response to changing environments through breeding and selection and may also allow area growers to be responsive to shifting condi-tions through appropriate cultivar selection.
AcknowledgmentsThis work was supported in part by grants from USDA/
CSREES-NRICAP (2006-55606-16629) and the Montana
Board of Research and Commercialization.
ReferencesBeales, J., A. Turner, S. Griffi ths, J.W. Snape, and D.A. Laurie.
2007. A Pseudo-Response Regulator is misexpressed in the pho-
toperiod insensitive Ppd-D1a mutant of wheat (Triticum aesti-
vum L.). Theor. Appl. Genet. 115:721–733.
Blake, N.K., S.P. Lanning, J.M. Martin, J.D. Sherman, and L.E.
Talbert. 2007. Relationship of fl ag leaf characteristics to eco-
nomically important traits in two spring wheat crosses. Crop
Sci. 47:489–494.
Blake, N.K., J.D. Sherman, J. Dvor̆ák, and L.E. Talbert. 2004.
Genome-specifi c primer sets for starch biosynthesis genes in
wheat. Theor. Appl. Genet. 109:1295–1302.
Busch, R.H., F.A. Elsayed, and R.E. Heiner. 1984. Eff ect of day-
length insensitivity on agronomic performance in spring
wheat. Crop Sci. 21:57–60.
Butler, J.D., P.F. Byrne, V. Mohammadi, P.L. Chapman, and
S.D. Haley. 2005. Agronomic performance of Rht alleles in
a spring wheat population across a range of moisture levels.
Crop Sci. 45:939–947.
Dyck, J.A., M.A. Matus-Cadiz, P. Hucl, L. Talbert, T. Hunt, J.P.
Dubuc, H. Nass, G. Clayton, J. Dobb, and J. Quick. 2004.
Agronomic performance of hard red spring wheat isolines sen-
sitive and insensitive to photoperiod. Crop Sci. 44:1976–1981.
Ellis, M.H., W. Spielmeyer, K.R. Gale, G.J. Rebetzke, and R.A.
Richards. 2002. ‘Perfect’ markers for the Rht-B1b and Rht-D1b
dwarfi ng genes in wheat. Theor. Appl. Genet. 105:1038–1042.

1220 WWW.CROPS.ORG CROP SCIENCE, VOL. 49, JULY–AUGUST 2009
Fu, D., P. Szucs, L. Yan, M. Helguera, J.S. Skinner, J.V. Zitzewitz,
P.M. Hayes, and J. Dubcovsky. 2005. Large deletions within
the fi rst intron in VRN-1 are associated with spring growth
habit in barley and wheat. Mol. Genet. Genomics 273:54–65.
Hedden, P. 2003. The genes of the green revolution. Trends Genet.
19:5–9.
Iqbal, M., A. Navabi, R.-C. Yang, D.F. Salmon, and D. Spaner.
2007. The eff ect of vernalization genes on earliness and
related agronomic traits of spring wheat in northern growing
regions. Crop Sci. 47:1031–1039.
Knott, D.R. 1986. Eff ects of genes for photoperiodism, semi-
dwarfi sm, and awns on agronomic characters in a spring
wheat cross. Crop Sci. 26:1158–1162.
Lanning, S.P., G.R. Carlson, D. Nash, D.M. Wichman, K.D.
Kephart, R.N. Stougaard, G.D. Kushnak, J.L. Eckhoff , W.E.
Grey, and L.E. Talbert. 2004. Registration of ‘Choteau’
wheat. Crop Sci. 44:2264–2265.
Lanning, S.P., P. Fox, J. Elser, J.M. Martin, N.K. Blake, and L.E.
Talbert. 2006. Microsatellite markers associated with a second-
ary stem solidness locus in wheat. Crop Sci. 46:1701–1793.
Lanning, S.P., L.E. Talbert, C.F. McGuire, H.F. Bowman, G.R.
Carlson, G.D. Jackson, J.L. Eckhoff , G.D. Kushnak, R.N.
Stougaard, and G.F. Stallknecht. 1994. Registration of
‘McNeal’ wheat. Crop Sci. 34:1126–1127.
Marshall, L., R. Busch, F. Cholick, I. Edwards, and R. Frohberg.
1989. Agronomic performance of spring wheat isolines diff er-
ing for daylength response. Crop Sci. 29:752–757.
McNeal, F.H., M.A. Berg, V.R. Stewart, and D.E. Baldridge.
1972. Agronomic response of three height classes of spring
wheat, Triticum aestivum L., compared at diff erent yield levels.
Agron. J. 64:362–364.
Montana Agricultural Statistics. Montana Dep. of Agriculture
and U.S. Dep. of Agriculture National Agricultural Statistics
Service. vol. XLIV. October 2007. (www.nass.usda.gov/mt/;
verifi ed 11 May 2009).
Mohler, V., R. Lukman, S. Ortiz-Islas, M. William, A.J. Wor-
land, J. van Beem, and G. Wenzel. 2004. Genetic and physical
mapping of photoperiod insensitive gene Ppd-B1 in common
wheat. Euphytica 138:33–40.
SAS Institute. 2003. SAS/STAT 9.1 user’s guide. SAS Inst., Cary, NC.
Scarth, R., and C.N. Law. 1984. The control of day-length
response in wheat by the group 2 chromosomes. Z. Pfl anzen-
zuchtg. 92:140–150.
Sherman, J.D., L. Yan, L.E. Talbert, and J. Dubcovsky. 2004. A
PCR marker for growth habit in common wheat based on
allelic variation at the Vrn-A1 gene. Crop Sci. 44:1832–1838.
Sourdille, P., J.W. Snape, T. Cadalen, G. Charmet, N. Nakata, S.
Bernard, and M. Bernard. 2000. Detection of QTL for head-
ing time and photoperiod response in wheat using a doubled-
haploid population. Genome 43:487–494.
Stelmakh, A.F. 1993. The genetic eff ect of Vrn genes on heading date
and agronomic traits in spring wheat. Euphytica 65:53–60.
Worland, A.J. 1996. The infl uence of fl owering time on environmen-
tal adaptability in winter European wheats. Euphytica 89:49–57.
Worland, A.J., M.L. Appendino, and E.J. Sayers. 1994. The dis-
tribution in European winter wheats of genes that infl uence
ecoclimatic adaptability whilst determining photoperiod
insensitivity and plant height. Euphytica 80:219–228.
Worland, A.J., A. Borner, V. Korzun, W.M. Li, S. Petrovic, and
E.J. Sayers. 1998. The infl uence of photoperiod genes on the
adaptability of European winter wheat populations. Euphyt-
ica 100:385–394.
Yan, L., D. Fu, C. Li, A. Blechl, G. Tranquilli, M. Bonafede, A.
Sanchez, M. Valarik, S. Yasuda, and J. Dubcovsky. 2006. The
wheat and barley vernalization gene VRN3 is an orthologue
of FT. Proc. Natl. Acad. Sci. USA 103:19581–19586.
Yan, L., M. Helguera, K. Kato, S. Fukuyama, J. Sherman, and J.
Dubcovsky. 2004. Allelic variation at the Vrn-1 promoter region
in polyploid wheat. Theor. Appl. Genet. 109:1677–1686.
Zhang, X.K., Y.G. Xiao, Y. Zhang, X.C. Xia, J. Dubcovsky, and
Z.H. He. 2008. Allelic variation at the vernalization genes Vrn-
A1, Vrn-B1, Vrn-D1, and Vrn-B3 in Chinese wheat cultivars and
their association with growth habit. Crop Sci. 48:458–470.




















