
Effect of repeated administration of centhaquin, a centrally acting hypotensive drug, on adrenergic,...
17
Drug Development Research 23:307-323 (1991) Effect of Repeated Administration Centhaquin, a Centrally Acting Hypotensive Drug, on Adrenergic, of Cholinergic (Muscarinic), Dopaminergic, and Serotonergic Receptors in Brain Regions of Rat Anil Gulati, Ghatala Hussain, and Rikhab Chand Srimal Department of Pharmacodynamics (m/c 865), The University of Illinois at Chicago, Chicago, Illinois (A. G.); Division of Pharmacology, Central Drug Research Institute, Lucknow, India (G. H., R.C. S.) ABSTRACT Gulati, A., G. Hussain, and R.C. Srimal: Effect of repeated administration of centhaquin, a centrally acting hypotensive drug, on adrenergic, cholinergic (muscarinic), dopaminergic, and serotonergic receptors in brain regions of rat. Drug Dev. Res. 23:307-323, 1991. Centhaquin, a hypotensive drug, produced a dose dependent decrease in blood pressure and heart rate in urethane anesthetized rats. The hypotensive and bradycardic effects of centhaquin were absent in cervical sectioned rats. There was no effect on blood pressure or heart rate upon intrathecal injection of centhaquin in normal rats. These results indicate that centhaquin is a centrally acting hypotensive drug with supraspinal site of action. Cen- thaquin (0.1 mg/kg) or its vehicle was administered orally to rats for 2 months. The blood pressure was recorded indirectly in unanesthetized rats by the plethysmographictail cuff method. The animals were sacrificed, decapitated, and specific areas of the brain, like corpus striatum, cerebral cortex, hypothalamus,and medulla were dissected out and mem- branes prepared for binding studies. The mean blood pressure of rats before starting the treatment was comparable in control and treated groups. A significant decrease in mean blood pressure was observed in centhaquin treated rats. Chronic administration of cen- Received final version November 21, 1990; accepted February 15, 1991. Address reprint requests to Anil Gulati, M.B., B.S., M.D., Department of Pharmacodynall,.,s (865), The University of Illinois at Chicago, 833, South Wood Street, Chicago, IL 60612. 0 1991 Wiley-Liss, Inc.
Transcript of Effect of repeated administration of centhaquin, a centrally acting hypotensive drug, on adrenergic,...

Drug Development Research 23:307-323 (1991)
Effect of Repeated Administration Centhaquin, a Centrally Acting Hypotensive Drug, on Adrenergic,
of
Cholinergic (Muscarinic), Dopaminergic, and Serotonergic Receptors in Brain Regions of Rat Anil Gulati, Ghatala Hussain, and Rikhab Chand Srimal
Department of Pharmacodynamics (m/c 865), The University of Illinois at Chicago, Chicago, Illinois (A. G.); Division of Pharmacology, Central Drug Research Institute, Lucknow, India (G. H., R. C. S.)
ABSTRACT
Gulati, A., G. Hussain, and R.C. Srimal: Effect of repeated administration of centhaquin, a centrally acting hypotensive drug, on adrenergic, cholinergic (muscarinic), dopaminergic, and serotonergic receptors in brain regions of rat. Drug Dev. Res. 23:307-323, 1991.
Centhaquin, a hypotensive drug, produced a dose dependent decrease in blood pressure and heart rate in urethane anesthetized rats. The hypotensive and bradycardic effects of centhaquin were absent in cervical sectioned rats. There was no effect on blood pressure or heart rate upon intrathecal injection of centhaquin in normal rats. These results indicate that centhaquin is a centrally acting hypotensive drug with supraspinal site of action. Cen- thaquin (0.1 mg/kg) or its vehicle was administered orally to rats for 2 months. The blood pressure was recorded indirectly in unanesthetized rats by the plethysmographic tail cuff method. The animals were sacrificed, decapitated, and specific areas of the brain, like corpus striatum, cerebral cortex, hypothalamus, and medulla were dissected out and mem- branes prepared for binding studies. The mean blood pressure of rats before starting the treatment was comparable in control and treated groups. A significant decrease in mean blood pressure was observed in centhaquin treated rats. Chronic administration of cen-
Received final version November 21, 1990; accepted February 15, 1991.
Address reprint requests to Anil Gulati, M . B . , B.S., M.D., Department of Pharmacodynall,.,s (865), The University of Illinois at Chicago, 833, South Wood Street, Chicago, IL 60612.
0 1991 Wiley-Liss, Inc.

308 Gulati et al.
thaquin in rats produced alterations in central a-adrenergic, dopaminergic, and serotonergic (5-hydroxytryptamine1 [5-HT1]) receptors. In the cerebral cortex and corpus striatum of centhaquin treated rats, B,, and K, values of 3H-dihydroergocryptine binding did not change, while in hypothalamus and medulla, centhaquin treatment increased the B,,, value by 52% and 86%, respectively. There was no change in the K, values. Centhaquin treatment did not affect B,,, and K, values of 3H-dihydroalprenolol binding to P-adrenergic receptors, 3H-quinuclydinyl benzylate binding to cholinergic (muscarinic), and 3H-spiperone binding to 5-hydroxytryptamine’ (5-HTJ receptors in any of the four brain regions. In the cortex, centhaquin treatment decreased the B,, value (25%) but did not change the K, value of 3H-spiroperidol binding to dopaminergic receptors. No change in B,, and K, values was produced in corpus striatum, hypothalamus, and medulla of 3H-spiroperidol binding to dopaminergic receptors. Cerebral cortex, corpus striatum, and hypothalamus of centhaquin treated rats did not show any change in B,,, or K, values of 3H-5-HT binding to 5-HT1 receptors. In medulla, centhaquin treatment increased the B,,, value (41%) of HT, receptors but had no effect on K, values. It can be concluded from the present study that centhaquin, a centrally acting hypotensive drug, acts on several neurotransmitter re- ceptors. The brain regions mainly involved in cardiovascular regulation, hypothalamus and medulla, show up-regulation of a-adrenergic and serotonergic (5-HT1) receptors and may play a role in the mechanism of hypotensive action of centhaquin.
Key words: hypotensive and bradycardic effects, supraspinal site, neurotransmitter receptors
INTRODUCTION
Hypertension is a frequently occurring disease all over the world. The main thrust of antihypertensive therapy consists of treatment with drugs. Many potential sites and mecha- nisms exist through which a drug may exert its antihypertensive effect. The role of the central nervous system (CNS) in regulating blood pressure has been known for a long time. Clonidine was developed and has been widely used as an antihypertensive drug [Srimal et al., 1977; Schmitt and Schmitt, 1969; Houston, 19811. The brain stem has been shown to be an important site of action of clonidine [Schmitt and Schmitt, 19691. It reduces blood pressure by decreasing the sympathetic tone [Houston, 19811 and noradrenaline release [Anden et al., 1970; Ber- thelsen and Pettinger, 19771. It is generally regarded that the main mechanism for hypotensive action of clonidine is by stimulation of central a-adrenergic receptors [Van Zwieten and Timmermans, 1979; Pettinger, 19751. However, Srimal et al. [ 19771 have demonstrated that clonidine produces a fall in blood pressure by acting at cholinergic (muscarinic) receptors at the ventral surface of the medulla in the cat. Criscione et al. [1983] have proposed that cholinergic mechanism in the nucleus tractus solitarius (NTS) tends to lower arterial pressure after clonidine administration and may modulate the baroreceptor reflex without being an integral part of the reflex arc. It is known that centrally administered cholinergic drugs decrease blood pressure [Porsius et al., 1978; Philippu, 19811 and the effect is explained by a simultaneously occurring decrease in sympathetic out flow similar to clonidine. Central serotonergic receptors have also been involved in the hypotensive activity of clonidine. It has been demonstrated that hypotension and bradycardia are produced following intracerebroven- tricular injection of methysergide in dogs and rats [Antonaccio et al., 1975; Antonaccio and Cote, 19761. The effect was associated with a reduced peripheral sympathetic activity. Sim- ilarly, as described for clonidine, methysergide has been shown to enhance vagal reflex bradycardia in cats after injection of the drug in the fourth ventricle [Tadepalli et al., 19791.
Centhaquin, 1-(~-(2-quinolyl)ethyl)-4-m-tolylpiperazine (Fig. 1) structurally different from clonidine, produces a fall in blood pressure and heart rate similar to clonidine, predom- inantly through a cemai mechanism in the cat [Srimal et al., 19881. Intravenous administra- tion of centhaquin to spinal chi produced a much reduced effect on blood pressure and heart

Centhaquin and Central Receptors 309
Fig. 1. Structure of centhaquin.
rate as compared to decerebrate cat [Raghubir et al., 19871. Although centhaquin has been found to be a centrally acting hypotensive agent, there has been no study performed to determine the effect of chronic administration of centhaquin on various central neurotransmit- ter receptors. The aim of the present study is to investigate the effect of chronic administration (2 months) of centhaquin in adrenergic, cholinergic (muscarinic), dopaminergic, and seroto- nergic receptors in brain regions of the rat.
MATERIALS AND METHODS
Albino, male rats of the Charles Foster strain, bred in the animal house of the Central Drug Research Institute, Lucknow were used. Rats weighing 100-150 g were kept in standard laboratory conditions of temperature (23 2 1.5"C), humidity (50 2 lo%), and dark light (5.00-18.00 hr) cycle. The animals were allowed to acclimatize for 4 days before starting the experiment.
Determination of Blood Pressure and Heart Rate in Normal and Cervical Sectioned Rats
Rats were anesthetized with urethane (1.2 gikg, intraperitoneally [i .~.]) . The left fem- oral vein was cannulated (PE 50 tubing) for drug administration. The right femoral artery was cannulated using PE 50 tubing and connected to a Gould P23 ID pressure transducer for recording the blood pressure on a Grass P7D polygraph through a 7PI preamplifier. The heart rate was recorded through a 7P4B Grass tachograph on another channel of the polygraph. The preamplifier was triggered from blood pressure signals. In order to determine the involvement of the brain in the action of centhaquin, the spinal cord was sectioned in some rats at the cervical level and a part of the cervical spinal cord was removed so that there is absolutely no connection with the brain. In another experiment to determine the involvement of the spinal cord in the action of centhaquin, intrathecal administration of drug was done in rats. A PE 10 polyethylene tubing was inserted through a slit in the atlanto-occipital membrane and threaded caudally for 7.0 cm in the subarachnoid space [Yaksh and Rudy, 1976; Tang and Schoenfeld, 19781. Cannula placement was verifed in all the rats at the end of the experiment. The animals were allowed to stabilize after the surgery for at least 1 hr before the administration of drugs.
Drug Administration and Measurement of Blood Pressure by Indirect Method
The hypotensive drug centhaquin (0.1 mg/kg) was dissolved in a drop of ethanol and diluted with water or vehicle and administered orally to rats for 2 months. The dose of this drug was selected on the basis of preliminary work done in our laboratory, indicating the best dose which could be used chronically and effectively. The drug was prepared fresh everyday just before the administration.
The blood pressure was also recorded indirectly in unanesthetized rats, before and after the repeated administration of vehicle or centhaquin, by the plethysmographic tail cuff method

310 Gulati et al.
TABLE 1. Summary of the Various Ligands Used for Radioreceptor Binding to Different Neurotransmitter ReceDtors Receptor a-adrenergic
P-adrenergic
Muscarinic
Dopamine
5 -H ydrox ytry ptamine
5-Hydroxytryptamine, (5-HT 1)
(5-HTZ)
Hot ligand 3H-di hy droergocryptine
0.25-12 nM (Amersham) 3H-dihydroalprenolol
0.12-4 nM (Amersham) 3H-quinuclidiny1 benzylate
0.1-2 nM (NEN) 3H-spiperone
0.1-5 nM (NEN) 3H-5-hydroxytryptamine
0.25-15 nM (NEN) 3H-spiperone
0.15-10 nM (NEN)
Cold ligand Phentolamine
(1 W) Propranolol
(1 W) Atropine
(1 I”M) Haloperidol
(1 CLM) 5-Hydroxytryptamine( 5-HT)
(10 CLW Ketanserin
(1 (LM)
[Friedman and Freed, 19491 using a rat tail manometer system (IITC Model No. 45). The arterial pulses were detected photoelectrically by sensing cuffs and the output was recorded on a dual channel recorder to show the presence of tail pulses on one channel and the matching cuff pressure on the other channel. The IITC method to measure blood pressure determines systolic as well as the mean blood pressure of rodents. The first onset of pulses is taken as the systolic point and the first maximum amplitude is taken as the mean blood pressure point. The blood pressure using this system could be recorded at room temperature without prewarming the rats to increase blood flow to the tail.
Preparation of Membranes
At the end of 2 months of drug treatment the animals were sacrificed by decapitation. Thereafter specific areas of brain, corpus striatum, cerebral cortex, hypothalamus, and me- dulla were dissected out in cold. Each brain area was weighed and then homogenized in 30 volumes of 50 mM Tris-HC1 buffer (pH 7.7 at 25°C) using a motor driven Teflon-glass tissue homogenizer. The tissue homogenate was centrifuged twice at 49,OOOg for 15 min in a refrigerated Sorvall centrifuge after resuspending in fresh Tris-HC1 buffer. The final pellet was suspended in 50 mM Tris buffer (pH 7.4 at 25°C) consisting of 0.1% ascorbic acid, 120 mM NaCl, 5 mM KC1, 2 mM CaC1, and 10 pM pargyline to give a concentration of 10 mg wet tissue per ml of incubation buffer. The binding studies were performed essentially as described by Gulati et al. [ 1988a,b]. Details about the incubation conditions are given in Table 1.
Binding of 3H-Dihydroergocryptine to a-Adrenergic Receptors
The standard assay mixture consisted of 0.2 ml of the homogenate, containing approx- imately 200 pg of tissue, 0.1 ml of 3H-dihydroergocryptine, 0.1 ml of competing agent (phentolamine), and buffer to make up the total volume of 1 ml. Incubation was carried out in triplicate in a shaking water bath maintained at 37°C for 15 min. At the end of the 15 min period, the contents of the incubation tubes were rapidly filered under partial vacuum using Millipore manifold filtration unit and Whatman GF/C filters. It was followed by two 5 ml washes of ice cold 50 mM Tris-HC1 buffer (pH 7.4). The filters were transferred to liquid scintillation vials containing 10 ml of cocktail. After an overnight equilibration period, the radioactivity was determined in a Packard Tricarb liquid scintillation spectrometer. The spe- cific binding of 3H-dihydroergocryptine was defined as the difference in binding observed in the absence and in the presence of 1 pM unlabeled phentolamine.

Centhaquin and Central Receptors 311
Binding of 3H-Dihydroalprenolol to p-Adrenergic Receptors
The standard assay mixture consisted of 0.2 ml of the homogenate, containing approx- imately 200 pg of tissue, 0.1 ml of 3H-dihydroalprenolol, 0.1 ml of competing agent (pro- pranolol), and buffer to make up the total volume of 1 ml. Incubation was carried out in triplicate in a shaking water bath maintained at 37°C for 15 min. The samples were filtered and the radioactivity was determined as described earlier. The specific binding of 3H-dihydroal- prenolol was defined as the difference in binding observed in the absence and in the presence of 1 pM unlabeled propranolol.
Binding of 3H-Quinuclydinyl Benzylate to Cholinergic (Muscarinic) Receptors
The membranes from freshly dissected brain regions were prepared as described earlier. The standard assay mixture consisted of 0.2 ml of the homogenate, containing approximately 200 pg of tissue, 0.1 ml of 'H-quinuclydinyl benzylate, 0.1 ml of competing agent (atropine), and buffer to make up the total volume of 1 ml. Incubation was carried out in triplicate in a shaking water bath maintained at 37°C for 15 min. At the end of the 15 min period, the contents of the incubation tubes were rapidly filtered and the radioactivity was determined. The specific binding of 3H-quinuclydinyl benzylate was defined as the difference in binding observed in the absence and in the presence of 1 p M unlabeled atropine.
Binding of 3H-Spiroperidol to Dopamine Receptors
The 200
The membranes of freshly dissected brain regions were prepared as described earlier. standard assay mixture consisted of 0.2 ml of the homogenate, containing approximately pg of tissue, 0.1 ml of 3H-spiroperidol, 0.1 ml of competing agent (haloperidol), and
buffer to make up the total volume of 1 ml. Incubation and filtration were carried out as described before. The filters were transferred to liquid scintillation vials and the radioactivity was determined in a Packard Tricarb liquid scintillation spectrometer. The specific binding of 'H-spiroperidol was defined as the difference in binding observed in the absence and in the presence of 1 pM unlabeled haloperidol.
Binding of 3H-5-HT to 5-HT, Receptors
The binding assays for 3H-5-HT were carried out as described before. The tissue in each tube was about 300 pg. The incubation was carried out for 20 min in a shaking water bath maintained at 37°C. At the end of the incubation period, the contents of the incubation tubes were filtered rapidly under partial vacuum using Whatman GFIC filter discs on a Millipore manifold filtration unit. The washing and counting of radioactivity was done as described before for other receptor studies. The specific binding was defined as the difference in binding of 'H-5-HT in the absence and presence of 10 pM 5-HT.
Binding of 3H-Spiroperidol to 5-HT2 Receptors
The freshly dissected brain regions were homogenized and neuronal membranes pre- pared as described earlier. The standard assay mixture consisted of 0.2 ml of the homogenate, containing approximately 200 p,g of tissue, 0.1 ml of 3H-spiroperidol, 0.1 ml of competing agent (ketanserin), and buffer to make up the total volume of 1 ml. Incubation and filtration were done and radioactivity determined as described earlier, followed by two 5 ml washes of ice cold 50 mM Tris-HC1 buffer (pH 7.4). The specific binding of 3H-spiroperidol was defined as the difference in binding observed in the absence and the presence of 1 pM unlabeled ketanserin.
The specific binding was expressed as pmoles of 3H-ligand bound per g tissue. The specific binding of the radioligand was determined at 8-10 concentrations in each case. This was followed by the Scatchard analysis to determine the dissociation constant (Kd) and the receptor density (Bmax) values. Five determinations, one in each rat, were made to compute the
Gulati et al.
2o01
1 Mln L
- ~-
3 0 Mln 4 5 Mln
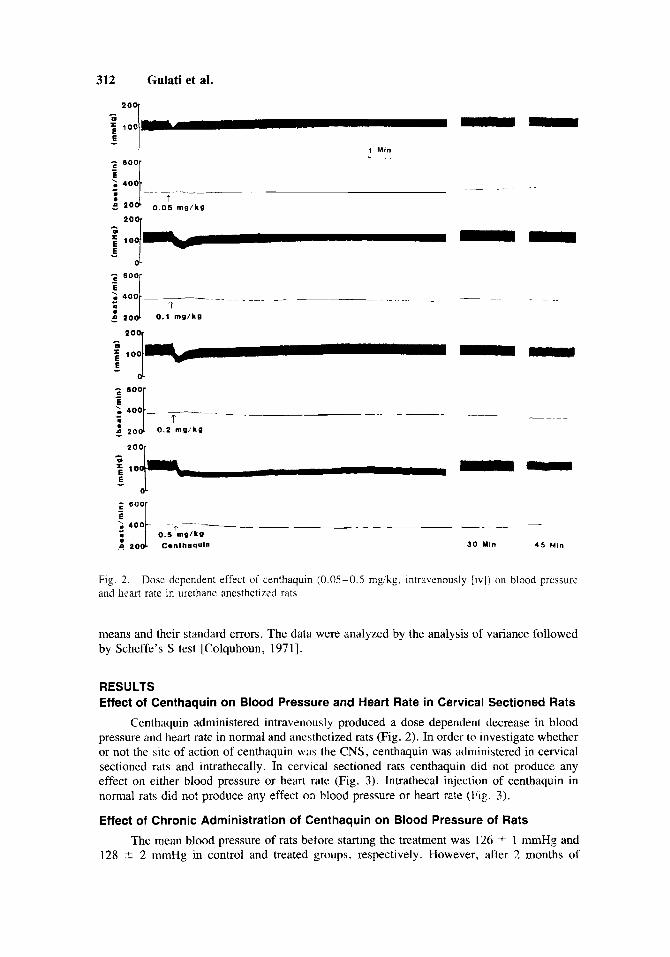
Fig. 2 and heart rate in urethane anesthetized rats
Dose dependent effect of centhaquin (0.05-0.5 mgikg, intravenously [iv]) on blood pressure
means and their standard errors. The data were analyzed by the analysis of variance followed by Scheffe's S test [Colquhoun, 19711.
RESULTS Effect of Centhaquin on Blood Pressure and Heart Rate in Cervical Sectioned Rats
Centhaquin administered intravenously produced a dose dependent decrease in blood pressure and heart rate in normal and anesthetized rats (Fig. 2). In order to investigate whether or not the site of action of centhaquin was the CNS, centhaquin was administered in cervical sectioned rats and intrathecally . In cervical sectioned rats centhaquin did not produce any effect on either blood pressure or heart rate (Fig. 3 ) . Intrathecal injection of centhaquin in normal rats did not produce any effect on blood pressure or heart rate (Fig. 3).
Effect of Chronic Administration of Centhaquin on Blood Pressure of Rats
The mean blood pressure of rats before starting the treatment was 126 2 1 mmHg and 128 t 2 mmHg in control and treated groups, respectively. However, alter 2 months of

Centhaquin and Central Receptors 313
I" E 2oor; loot - = ___
2oo 0.1 mg/kg lv 200,
Cervical Sectioned 0 I E lool --
0 300r
--
2oor
d 600 .- 2 L E 4 0 0 1
0 -- 200 ? n
30MIN 45MIN 1 pg lntrathacal
Centhaquin
Fig. 3. Effect of centhaquin on blood pressure and heart rate in anesthetized rats. The upper and middle panels represent typical blood pressure and heart rate responses induced by 0.1 mgikg, iv centhaquin prior to and 1 hr postcervical sectioning. The lower panel represents typical blood pressure and heart rate tracings obtained from a rat receiving 1 p,g of centhaquin intrathecally.
treatment a significant decrease in mean blood pressure was observed in centhaquin treated rats, and the blood pressure was 130 i 1 mrnHg in control rats and 117 +- 1 mmHg in centhaquin treated rats [F(1,8) = 84.96; P < 0.00051.
Effect of Chronic Administration of Centhaquin on a-Adrenergic Receptors
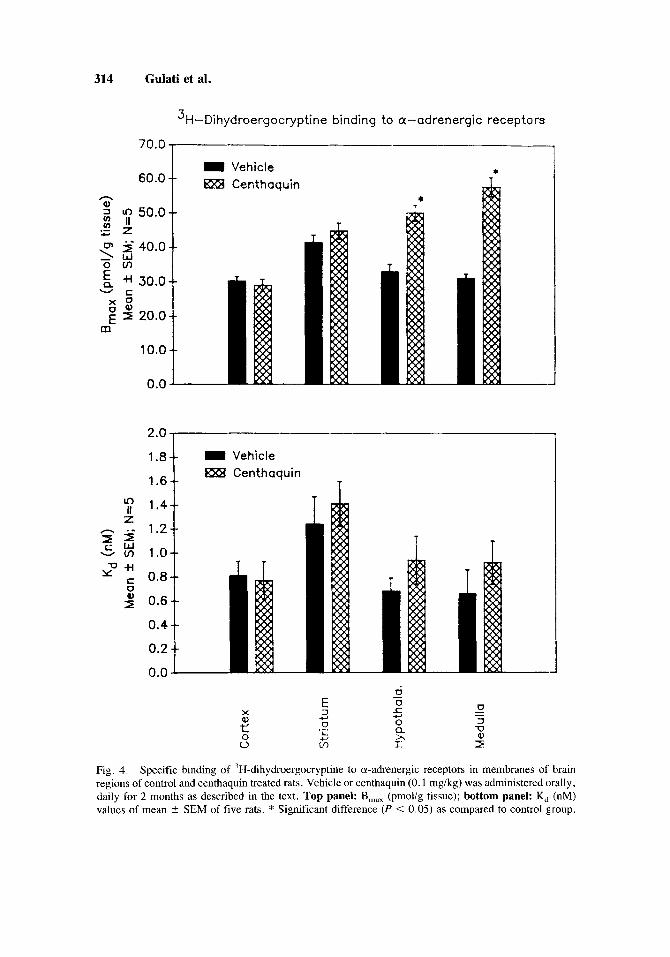
The population (B,J of the adrenergic receptors in normal rat brain was found to be fairly constant in all the four areas. The B,,, values in different regions of the brain ranged from 30.2-41.5 pmol/g tissue, while the K, value ranged from 0.7-1.2 nM. Thus the binding of 'H-dihydroergocryptine was at a single high affinity binding site.
In cortex and corpus striatum of centhaquin treated rats the B,,, and K, values of 'H-dihydroergocryptine binding, remained unchanged as compared to control. In hypothala- mus, the B,,, value increased by 52.4% [F( I ,8) = 29.25; P = 0.0011 in centhaquin treated rats while the K, value did not change as compared to control. Similarly, in the medulla of centhaquin treated rats the B,,, value increased [(85.9%) F(1,8) = 75.78; P < 0.0005] and no change occurred in the K, value as compared to control (Fig. 4).
314 Gulati et al.
3H-Dihydroergocryptine binding to a-adrenergic receptors
70*0' 60.0 t
t
I Vehicle * BXX Centhaquin
* t
t
T
= Vehicle 13xB Centhaquin
T 1.6
7 1.4 - * - 1.2
v m 1.0
z
3 5 2 +I 0.8
U 0.6
0.4
0.2
0.0
X
P 0 0
Fig. 4. Specific binding of 3H-dihydroergocryptine to a-adrenergic receptors in membranes of brain regions of control and centhaquin treated rats. Vehicle or centhaquin (0.1 mg/kg) was administered orally, daily for 2 months as described in the text. Top panel: B,,, (pmolig tissue); bottom panel: K, (nM) values of mean * SEM of five rats. * Significant difference (P < 0.05) as compared to control group.

Centhaquin and Central Receptors 315
Effect of Chronic Administration of Centhaquin on p-Adrenergic Receptors
The binding of 'H-dihydroalprenolol to P-adrenergic receptors was at a single high affinity binding site. The B,,, value ranged from 24.8-38.8 pmol/g tissue, while the K, ranged from 0.6-1.8 nM.
In the cortex, corpus striatum, hypothalamus, and medulla of centhaquin treated rats the B,,, and K, values of 3H-dihydroalprenolol binding did not differ as compared to the control rats (Fig. 5) .
Effect of Chronic Administration of Centhaquin on Dopaminergic Receptors
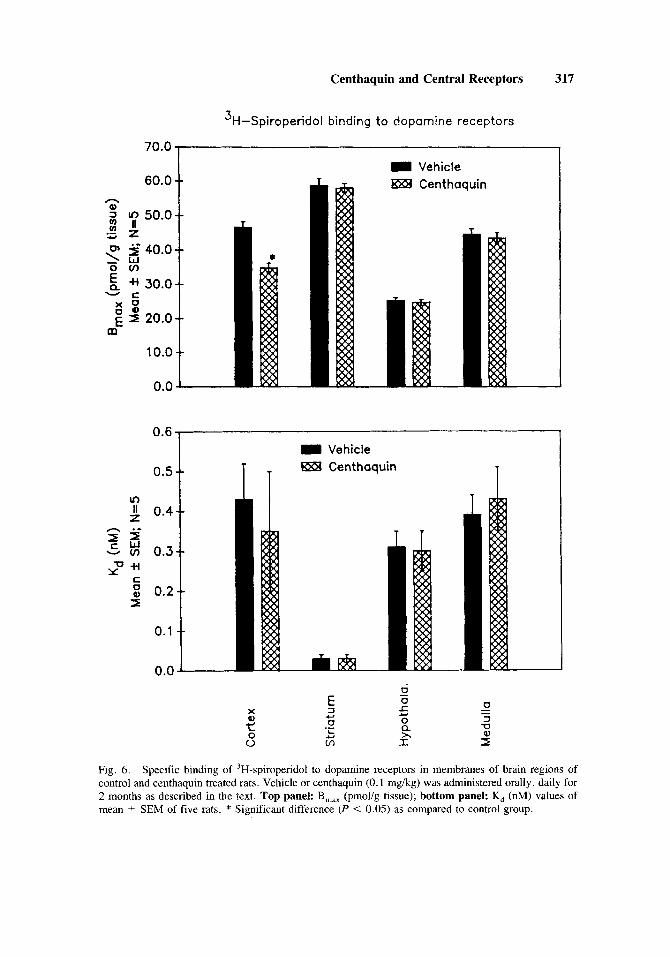
The binding of 'H-spiroperidol to dopaminergic sites in the corpus striatum was found to be at a single high affinity binding site with a B,,, value of 58.7 pmol/g tissue and a K, value of 0.03 nM. In other brain regions the B,,, values ranged from 25.2-46.6 pmol/g tissue, whereas the K, value ranged from 0.3-0.4 nM.
In cortex, centhaquin treatment produced a 25.2% decrease [F(1,8) = 28.73; P = 0.0011 in the B,,, value while no change occurred in the K, value as compared to control. In the corpus striatum, hypothalamus, and medulla of centhaquin treated rats no change occurred in either the B,,, or the K, value of 3H-spiperone binding as compared to control (Fig. 6).
Effect of Chronic Administration of Centhaquin on Cholinergic (Muscarinic) Receptors
The binding of 3H-quinuclidinyl benzylate to muscarinic receptor sites was at a single high affinity binding site. The B,,, value ranged from 36.5-63.1 pmol/g tissue, while the K, value ranged from 0.7-1.4 nM.
In cortex, corpus striatum, hypothalamus, and medulla of centhaquin treated rats the B,,, and K, values of 3H-quinuclidinyl benzylate to muscarinic receptors did not differ as compared to control (Fig. 7).
Effect of Chronic Administration of Centhaquin on 5-HT, Receptors
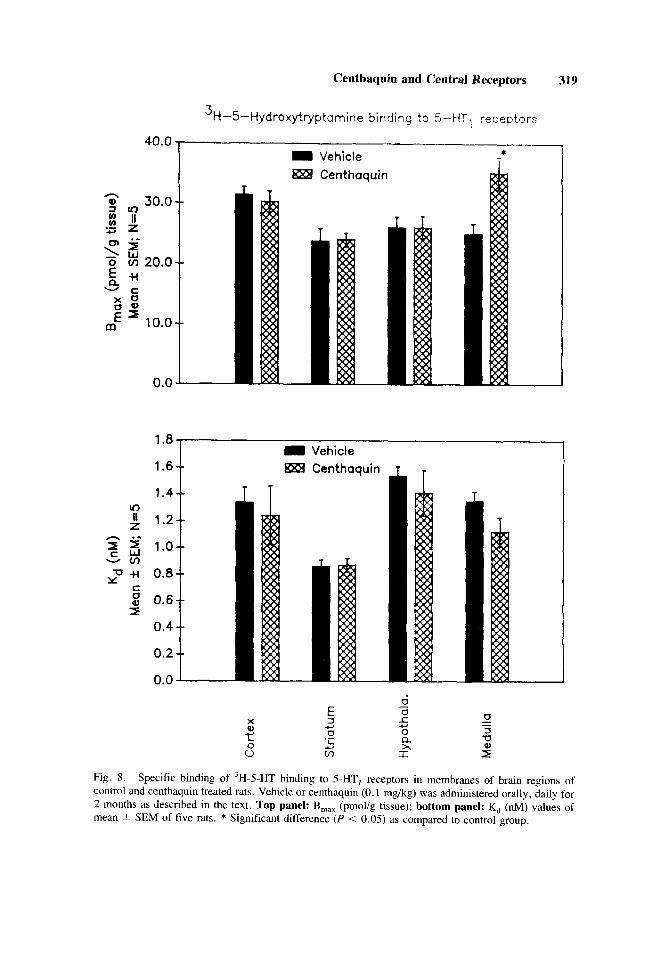
The 3H-5-HT binding to 5-HT, receptors is fairly constant and at a single high affinity binding site. The B,,, value ranged from 23.7-31.5 pmol/g tissue, while the K, value ranged from 0.9-1.5 nM.
The cortex, corpus striatum, and hypothalamus of centhaquin treated rats did not show any change in either the B,,, value or the K, value of 3H-5-HT binding. In the medulla, centhaquin treatment increased [(40.6%) F(1,8) = 10.18; P = 0.0131 the B,,, value while no change occurred in the K, value of 3H-5-HT binding (Fig. 8).
Effect of Chronic Administration of Centhaquin on 5-HT2 Receptors
The binding of 'H-spiperone to 5-HT2 receptors was at a single high affinity binding site. The B,,, value ranged from 23.3-34.9 pmol/g tissue, while the K, value ranged from 0.8-1.4 nM.
In the cortex, corpus striatum, hypothalamus, and medulla of centhaquin treated rats no change in the B,,, and K, values of the 3H-spiperone binding was observed as compared to control (Fig. 9).
DISCUSSION
The results of the present investigation show that chronic administration of centhaquin in rats produced hypotension, bradycardia, and alterations in central a-adrenergic, dopaminer- gic, and serotonergic (5-HT,) receptors. The density of a-adrenergic receptors was increased in the hypothalamus and medulla, and the density of 5-HT, receptors was increased in the

316 Gulati et al.
3H-Dihydroolprenolol binding to 8-adrenergic receptors
50.0
Vehicle
40.0 1
10.0
0.0
1 RSl Centhaquin
2.0 1 1
1.6
f 1.4 z
n .- 1.2 - m 1.0 5 5 y"* 0.8
0 2 0.6
0.4
0.2
0.0
I Vehicle RE4 Centhaquin
Fig. 5. Specific binding of 3H-dihydroalprenolol to P-adrenergic receptors in membranes of brain regions of control and centhaquin treated rats. Vehicle or centhaquin (0.1 mg/kg) was administered orally, daily for 2 months as described in the text. Top panel: B,,, (pmol/g tissue); bottom panel: K, (nM) values of mean * SEM of five rats.
Centhaquin and Central Receptors 317
60.0 = Vehicle --
0, 5 40.0 ' 1 .w
5 +I 30.0
m O m i - c x o ; 2 20.0
'o*ot 0.0
*
0.6 = Vehicle
0.5 1
O-' 0.0 L
$xB Centhaquin
Fig. 6. Specific binding of 3H-spiroperidol to dopamine receptors in membranes of brain regions of control and centhaquin treated rats. Vehicle or centhaquin (0.1 mg/kg) was administered orally, daily for 2 months as described in the text. Top panel: B,,, (pmolig tissue); bottom panel: K, (nM) values of mean +- SEM of five rats. * Significant difference (P < 0.05) as compared to control group.

318 Gulati et al.
3H-Quinuclidinyl benzilate binding to muscarinic (cholinergic) receptors
70.0 1
60.0 - -9i 2 50.0 bY "
0 1'40.0 '1.w
& +1 30.0
.-
o m
- c x o g 2 20.0 in
10.0
0.0
Vehicle El2 Centhaquin
1.6 I
= Vehicle gxB Centhaquin
1.41
1-2 t In
. ._ n a - zz l y"+i - V) 0.8
5 0.6
I 0.4
0.2
0.0
a2
T
Fig. 7. Specific binding of 'H-quinuclidinyl benzylate to muscarinic receptors in membranes of brain regions of control and centhaquin treated rats. Vehicle or centhaquin (0.1 rng/kg) was administered orally, daily for 2 months as described in the text. Top panel: B,,, (pmol/g tissue); bottom panel: K, (nM) values of mean 2 SEM of five rats.
Centhaquin and Central Receptors 319
'H-5-Hydroxytryptamine binding to 5 - H T , receptors
40.0 * T I Vehicle
gxB Centhaquin T
0.0 - 1.8 = Vehicle
gxB Centhaquin
1
Fig. 8. Specific binding of 'H-5-HT binding to 5-HT, receptors in membranes of brain regions of control and centhaquin treated rats. Vehicle or centhaquin (0.1 mg/kg) was administered orally, daily for 2 months as described in the text. Top panel: B,,, (pmol/g tissue); bottom panel: K, (nM) values of mean 2 SEM of five rats. * Significant difference (P < 0.05) as compared to control group.

320 Gulati et al.
3H -S pi ro per1 d 01 binding to 5 - HT rece D t o rs
n
5 v
U Y
Vehicle Centhaquin
1.6- = Vehicle 1.4-- ggS Centhaquin
1.2 --
1 .o - -
Fig. 9. Specific binding of 'H-spiroperidol binding to 5-HT2 receptors in membranes of brain regions of control and centhaquin treated rats. Vehicle or centhaquin (0.1 mg/kg) was administered orally, daily for 2 months as described in the text. Top panel: B,,, (pmolig tissue); bottom panel: K, (nM) values of mean t SEM of five rats.

Centhaquin and Central Receptors 321
medulla, while the density of dopaminergic receptors was decreased in the cortex of cen- thaquin treated rats.
The up-regulation of a-adrenergic receptors in the hypothalamus and medulla supports the presynaptic action of centhaquin. Clonidine like centhaquin also produced up-regulation of a-adrenergic receptors in the hypothalamus and medulla upon chronic administration in rats [Gulati et al., 19911. The affinity of clonidine for presynaptic a,-adrenergic receptors is at least 10 times higher than its affinity for postsynaptic a,-adrenergic receptors [Starke, 19771. It would be possible that repeated administration of centhaquin decreases the release of nor- epinephrine in the hypothalamus and medulla by acting on presynaptic receptors similar to clonidine. The decreased concentration of norepinephrine in the synapse will result in an increase in the density of a-adrenergic receptors.
The involvement of cholinergic (muscarinic) receptors in the mechanism of action of clonidine has been demonstrated. Srimal et al. [1977] suggested that an intact cholinergic link in the brain stem is essential for the hypotensive action of clonidine. Criscione et al. [1983] have proposed that cholinergic mechanism in the NTS tends tonically to lower arterial pressure after clonidine administration and may modulate the baroreflex without being an integral part of the reflex arc. The up-regulation of cholinergic (muscarinic) receptors in the hypothalamus and medulla was found in rats when clonidine was repeatedly administered for 2 months [Gulati et al., 19911. Although cholinergic (muscarinic) receptors may be playing an important role in the mechanism of action of clonidine, no change was observed in muscarinic receptors upon chronic administration of centhaquin .
The results of the present study show that centhaquin had no effect on dopaminergic receptors in the corpus striatum, hypothalamus, and medulla. It is known that clonidine does not stimulate the dopamine receptors in the corpus striatum of the rat [Anden et al., 19701. The decrease in the density of cortical dopamine receptors observed by chronic centhaquin treat- ment is possibly not related to the cardiovascular actions of centhaquin, since cortical dopa- minergic receptors have not been implicated in regulation of cardiovascular functions. It could be possible that some behavioral effects may be produced due to the effect of centhaquin on cortical dopaminergic receptors. It is interesting to note that centhaquin produces a decrease in the density of cortical dopamine receptors, whereas clonidine produces an increase in the density of these receptors [Gulati et al., 19911. This difference should be further investigated from the point of view of the behavioral effect of these drugs.
Centhaquin was found to produce an up-regulation of 5-HT1 receptors in the medulla similar to that observed with clonidine [Gulati et al., 19911. Clonidine has been found to decrease serotonin (5-HT) turnover [Anden et al., 1970; Rochette et al., 19741, and to reduce the concentration of 5-HT metabolites 5-hydroxy-3-indoleacetic acid (5-HIAA) in the rat brain [Scheel-Kruger and Hasselager, 19741. Maura et al. [1985] have shown that the presynaptic action of clonidine at a-adrenergic receptors mediates inhibition of 3H-noradrenaline and 3H-5-HT release. Since the changes in 5-HT receptor produced by centhaquin are similar to those produced by clonidine, it could be possible that centhaquin also decreases the release of 5-HT which will result in up-regulation of 5-HT receptors as observed in the present study.
It is well known that major cardiovascular regulatory centers are located in the hypo- thalamus and medulla and we have observed selective changes induced by centhaquin on the a-adrenergic and serotonergic neurotransmitter receptors in the hypothalamus and medulla similar to clonidine. However, centhaquin did not produce any change in cholinergic (mus- carinic) receptors a5 observed with clonidine [Gulati et al., 19911. It could be possible that these changes in receptors might be important in the hypotensive action of clonidine. On the other hand, changes in cerebral cortical dopaminergic receptors by centhaquin could be re- sponsible for the behavioral effects.
Finally, it can be concluded from the present study that centhaquin like clonidine acts on several neurotransmitter receptors. The brain regions mainly involved in cardiovascular

322 Gulati et al.
regulation, hypothalamus and medulla, show up-regulation of a-adrenergic and serotonergic (SHT,) receptors and may play a role in the mechanism of hypotensive action of centhaquin.
ACKNOWLEDGMENTS
The financial support from the Dean, College of Pharmacy, The University of Illinois at Chicago, is gratefully acknowledged. We are grateful to Dr. Eugene F. Woods for taking a keen interest in the study.
REFERENCES
Anden, N.E., Corrodi, H., Fuxe, K., Hokfelt, B., Hokfelt, T. , Rydin, C., and Svensson, T.: Evidence for a central noradrenaline receptor stimulation by clonidine. Life Sci. 9:5 13-523, 1970.
Antonaccio, M.A.J., and Cote, D.: Centrally mediated antihypertensive and bradycardic effects of methysergide in spontaneously hypertensive rats. Eur. J. Pharmacol. 36:45 1-454, 1976.
Antonaccio, M.A.J., Kelly, E., and Halley, J.: Centrally mediated hypotension and bradycardia by methysergide in anaesthetized dogs. Eur. J. Pharmacol. 33: 107-1 17, 1975.
Berthelsen, S., and Pettinger, W.A.: A functional basis for classification of a adrenergic receptors. Life Sci. 21:595-606, 1977.
Colquhoun, D.: The analysis of variance. How to deal with two or more samples. In Colquhoun, D. (ed.): “Lectures on Biostatistics.” London: Oxford University Press, 1971, pp. 171-207.
Criscione, L., Reis, D.J., and Talman, W.T.: Cholinergic mechanisms in the nucleus tractus solitarii and cardiovascular regulation in the rat. Eur. J. Pharmacol. 88:47-55, 1983.
Friedman, M., and Freed, S.C.: Microphonic manometer for indirect determination of systolic blood pressure in the rat. Proc. SOC. Exp. Biol. Med. 70:670-672, 1949.
Gulati, A,, Hussain, G., and Srimal, R.C.: Effect of repeated administration of clonidine on adrenergic, cholinergic (muscarinic), dopaminergic and serotonergic receptors in brain regions of rat. Drug Dev. Res. 22:141-152, 1991.
Gulati, A., Hussain. G., Srimal, R.C., and Dhawan, B.N.: Comparison of cortical adrenergic, cholin- ergic and benzodiazepine receptors between albino rat and desert rat (Mastomys Natalensis) using radioreceptor binding. Pharmacology 36:325-330, 1988b.
Gulati, A., Srimal, R.C., and Dhawan, B.N.: Differential alteration in striatal dopaminergic and cortical serotonergic receptors induced by repeated administration of haloperidol or centbutindole in rats. Pharmacology 36:396-404, 1988a.
Hokfelt, B., Hedeland, H., and Dymling, J.F.: Studies on catecholamines, renin and aldosterone fol- lowing catapresan (2-(2,6-dichloro-phenyamino)-2-imidazoline hydrochloride) in hypertensive pa- tients. Eur. J. Pharmacol. 10:389-397, 1970.
Houston, M .C. : Clonidine hydrochloride: review of pharmacologic and clinical aspects. Prog. Cardio- vasc. Dis. 23:337-350, 1981.
Maura, G., Bonanno, G., and Raiteri, M.: Chronic clonidine induces functional down-regulation of presynaptic a2 adrenoceptors regulating [3H] noradrenaline and [3H] 5-hydroxytryptamine release in the rat brain. Eur. J. Pharmacol. 112:105-110, 1985.
Pettinger, W.A.: Clonidine, a new antihypertensive drug. N. Engl. J. Med. 293:1179-1180, 1975. Philippu, A.: Involvement of cholinergic systems of the brain in the central regulation of cardiovascular
functions. J . Auton. Pharmacol. 1:321-330, 1981. Porsius, A.J., Mutschler, E., and Van Zwieten, P.A.: The central action of various arecaidine esters
(arecoline derivatives) on blood pressure and heart rate in the cat. Arzneimittelforschung 28:
Raghubir, R., Srimal, R.C., and Dhawan, B.N.: Enkephalinergic modulation of cardiovascular effects of clonidine from ventral surface of medulla of cat. In Tangri, K.K., Vrat, S., and Saxena, A. (eds): ‘‘Brain Neurotransmitter Mechanisms and Hypertension. ” Lucknow, India: Kamla Printers, 1987,
Rochette, L. , Bralet, A.M., and Bralet, J.: Influence de la clonidine sur la synthese et la liberation de la noradrenaline dans differentes structures cerebrales du rat. J. Pharmacol. (Paris) 5:209-220, 1974.
Scheel-Kmger, J . , and Hasselager, E.: Studies of various amphetamines, apomorphine and clonidine on
1373-1 376, 1978.
pp. 109-115.

Centhaquin and Central Receptors 323
body temperature and brain 5-hydroxytryptamine metabolism in rats. Psychopharmacology (Berl.) 36:189-202, 1974.
Schmitt, H., and Schmitt, H.: Interaction between 2-(2,6-dichlorophenylamino)-2-imidazoline hydro- chloride (ST 155 Catapresan) and a adrenergic blocking drugs. Eur. J. Pharmacol. 9:7-13, 1969.
Srimal, R.C., Gulati, K., and Dhawan, B.N.: On the mechanism of central hypotensive action of clonidine. Can. J. Physiol. Pharmacol. 55: 1007-1014, 1977.
Srimal, R.C., Raghubir, R., Gulati, K., Agarwal, S.K., and Dhawan, B.N.: Possible involvement of endogenous opioids in the mediation of cardiovascular effects of centhaquin. In Dhawan, B.N., and Rapaka, R.S. (eds.): “Recent Progress in Chemistry and Biology of Centrally Acting Pep- tides.” Lucknow, India: Central Drug Research Institute Press, 1988, pp. 191-199.
Starke, K .: Regulation of noradrenaline release by presynaptic receptor systems. Rev. Physiol. Biochem. Pharmacol. 77: 1-124, 1977.
Tadepalli, A, , Ho, K.A.W., and Buckley, J.P.: Enhancement of reflex vagal bradycardia following intracerebroventricular administration of methysergide in cats. Eur. J. Pharmacol. 59:85-93, 1979.
Tang, A.H., and Schoenfeld, M.J.: Comparison of subcutaneous and spinal subarachnoid injections of morphine and naloxoc- on analgesic tests in the rat. Eur. J. Pharmacol. 52:215-223, 1978.
Van Zwieten, P.A., and Timmermans, P.B.M.W.M.: The role of central a adrenoceptors in the mode of action of hypotensive drugs. Trends Pharmacol. Sci. 1:39-41, 1979.
Yaksh, T.L., and Rudy, T.A.: Chronic catheterization of the spinal subarachnoid space. Physiol. Behav. 17:1031-1036, 1976.




















