
High-Intensity Extended Swimming Exercise Reduces Pain-Related Behavior in Mice: Involvement of...
10
High-Intensity Extended Swimming Exercise Reduces Pain-Related Behavior in Mice: Involvement of Endogenous Opioids and the Serotonergic System Leidiane Mazzardo-Martins,* Daniel F. Martins,* Rodrigo Marcon, y Ubirajara D. dos Santos,* ,z Breno Speckhann,* Vinı´cius M. Gadotti,* Andre ´ Roberto Sigwalt, x Luiz Guilherme A. Guglielmo, x and Adair Roberto Soares Santos* ,y * Departamento de Cie ˆ ncias Fisiolo ´ gicas, Centro de Cie ˆ ncias Biolo ´ gicas, Universidade, Federal de Santa Catarina, Campus Universita ´rio, Trindade, Floriano ´ polis, SC, Brazil. y Departamento de Farmacologia, Centro de Cie ˆ ncias Biolo ´ gicas, Universidade Federal de Santa Catarina, Campus Universita ´rio, Trindade, Floriano ´ polis, SC, Brazil. z Departamento de Naturologia, Universidade do Sul de Santa Catarina, Pedra Branca, Palhoc ¸a, SC, Brazil. x Departamento de Educac ¸a ˜ o Fı ´sica, Centro de Desportos, Universidade Federal de Santa Catarina, Campus Universita ´rio, Trindade, Floriano ´ polis, SC, Brazil. Abstract: The present study examined the hyponociceptive effect of swimming exercise in a chem- ical behavioral model of nociception and the mechanisms involved in this effect. Male mice were sub- mitted to swimming sessions (30 min/d for 5 days). Twenty-four hours after the last session, we noticed that swimming exercise decreased the number of abdominal constriction responses caused by acetic acid compared with the nonexercised group. The hyponociception caused by exercise in the acetic acid test was significantly attenuated by intraperitoneal (i.p.) pretreatment of mice with naloxone (a nonselective opioid receptor antagonist, 1 mg/kg), r-chlorophenylalanine methyl ester (PCPA, an inhibitor of serotonin synthesis, 100 mg/kg once a day for 4 consecutive days), and by bilateral adrenalectomy. Collectively, the present results provide experimental evidences indicating for the first time that high-intensity extended swimming exercise reduces pain-related behavior in mice. The mechanisms involve an interaction with opioid and serotonin systems. Furthermore, endogenous opioids released by adrenal glands probably are involved in this effect. Perspective: Our results indicate that high-intensity extended exercise endogenously controls acute pain by activation of opioidergic and serotonergic pathways. Furthermore, these results sup- port the use of exercise as a nonpharmacological approach for the management of acute pain. ª 2010 by the American Pain Society Key words: Physical exercise, nociception, opioid, serotonin, mouse. P hysical exercise exerts many benefits for physical and mental health. It reduces disease incidence 6 ; promotes neuroprotection, neuroplasticity, 22,60 cognition improvement 7 ; and has anxiolytic and antide- pressant activities. 6,21 Beyond all these advantages, physical exercise alters the nociceptive threshold. 32,34 Evidence has shown that exercise is a helpful tool in the management of pain conditions. 64 Exercise protocols are therefore used to rehabilitate patients with chronic musculoskeletal conditions such as fibromyalgia, chronic low back pain, and myofascial pain. 20,64 Furthermore, several investigators have reported that acute exercise is able to change pain perception during and after physical exercise. 32,43 Many studies have examined the relationship between exercise and endorphin release, studying the role of these peptides in exercise-induced reduction of pain. 12,17,19,35,46 Endorphins are endogenous opioids released from the pituitary gland that are believed to mediate analgesia. b-Endorphin is abundantly found in Received September 2, 2009; Revised March 11, 2010; Accepted March 21, 2010. Supported by grants from Conselho Nacional de Desenvolvimento Cien- tı ´fico e Tecnolo ´ gico (CNPq), Fundac ¸a ˜ o de Apoio a ` Pesquisa Cientı´fica Tec- nolo ´ gica do Estado de Santa Catarina (FAPESC), Financiadora de Estudos e Projetos [FINEP, Rede Instituto Brasileiro de Neurocie ˆ ncia (IBN-Net)], Brazil. Dr Leidiane Mazzardo-Martins and Dr Daniel F. Martins contributed equally to this work. Address reprint requests to Dr Adair R.S. Santos, Department of Physio- logical Sciences, Federal University of Santa Catarina, Floriano ´ polis 88040-900, SC, Brazil. E-mail: [email protected] 1526-5900/$36.00 ª 2010 by the American Pain Society doi:10.1016/j.jpain.2010.03.015 1 The Journal of Pain, Vol -, No - (-), 2010: pp 1-10 Available online at www.sciencedirect.com ARTICLE IN PRESS
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of High-Intensity Extended Swimming Exercise Reduces Pain-Related Behavior in Mice: Involvement of...

P
The Journal of Pain, Vol -, No - (-), 2010: pp 1-10Available online at www.sciencedirect.com
ARTICLE IN PRESS
High-Intensity Extended Swimming Exercise Reduces Pain-Related
Behavior in Mice: Involvement of Endogenous Opioids and the
Serotonergic System
Leidiane Mazzardo-Martins,* Daniel F. Martins,* Rodrigo Marcon,y Ubirajara D. dosSantos,*,z Breno Speckhann,* Vinıcius M. Gadotti,* Andre Roberto Sigwalt,x
Luiz Guilherme A. Guglielmo,x and Adair Roberto Soares Santos*,y
*Departamento de Ciencias Fisiologicas, Centro de Ciencias Biologicas, Universidade, Federal de Santa Catarina,Campus Universitario, Trindade, Florianopolis, SC, Brazil.yDepartamento de Farmacologia, Centro de Ciencias Biologicas, Universidade Federal de Santa Catarina,Campus Universitario, Trindade, Florianopolis, SC, Brazil.zDepartamento de Naturologia, Universidade do Sul de Santa Catarina, Pedra Branca, Palhoca, SC, Brazil.xDepartamento de Educacao Fısica, Centro de Desportos, Universidade Federal de Santa Catarina,Campus Universitario, Trindade, Florianopolis, SC, Brazil.
Received2010.Supportetıfico e Tenologicae ProjetoBrazil.Dr Leidiaequally tAddresslogical S88040-90
1526-590
ª 2010 b
doi:10.10
Abstract: The present study examined the hyponociceptive effect of swimming exercise in a chem-
ical behavioral model of nociception and the mechanisms involved in this effect. Male mice were sub-
mitted to swimming sessions (30 min/d for 5 days). Twenty-four hours after the last session, we
noticed that swimming exercise decreased the number of abdominal constriction responses caused
by acetic acid compared with the nonexercised group. The hyponociception caused by exercise in
the acetic acid test was significantly attenuated by intraperitoneal (i.p.) pretreatment of mice with
naloxone (a nonselective opioid receptor antagonist, 1 mg/kg), r-chlorophenylalanine methyl ester
(PCPA, an inhibitor of serotonin synthesis, 100 mg/kg once a day for 4 consecutive days), and by
bilateral adrenalectomy. Collectively, the present results provide experimental evidences indicating
for the first time that high-intensity extended swimming exercise reduces pain-related behavior in
mice. The mechanisms involve an interaction with opioid and serotonin systems. Furthermore,
endogenous opioids released by adrenal glands probably are involved in this effect.
Perspective: Our results indicate that high-intensity extended exercise endogenously controls
acute pain by activation of opioidergic and serotonergic pathways. Furthermore, these results sup-
port the use of exercise as a nonpharmacological approach for the management of acute pain.
ª 2010 by the American Pain Society
Key words: Physical exercise, nociception, opioid, serotonin, mouse.
hysical exercise exerts many benefits for physicaland mental health. It reduces disease incidence6;promotes neuroprotection, neuroplasticity,22,60
cognition improvement7; and has anxiolytic and antide-
September 2, 2009; Revised March 11, 2010; Accepted March 21,
d by grants from Conselho Nacional de Desenvolvimento Cien-cnologico (CNPq), Fundacao de Apoio a Pesquisa Cientıfica Tec-do Estado de Santa Catarina (FAPESC), Financiadora de Estudoss [FINEP, Rede Instituto Brasileiro de Neurociencia (IBN-Net)],
ne Mazzardo-Martins and Dr Daniel F. Martins contributedo this work.reprint requests to Dr Adair R.S. Santos, Department of Physio-ciences, Federal University of Santa Catarina, Florianopolis0, SC, Brazil. E-mail: [email protected]
0/$36.00
y the American Pain Society
16/j.jpain.2010.03.015
pressant activities.6,21 Beyond all these advantages,physical exercise alters the nociceptive threshold.32,34
Evidence has shown that exercise is a helpful tool inthe management of pain conditions.64 Exercise protocolsare therefore used to rehabilitate patients with chronicmusculoskeletal conditions such as fibromyalgia, chroniclow back pain, and myofascial pain.20,64 Furthermore,several investigators have reported that acute exerciseis able to change pain perception during and afterphysical exercise.32,43
Many studies have examined the relationship betweenexercise and endorphin release, studying the role ofthese peptides in exercise-induced reduction ofpain.12,17,19,35,46 Endorphins are endogenous opioidsreleased from the pituitary gland that are believed tomediate analgesia. b-Endorphin is abundantly found in
1

2 The Journal of Pain Exercise Reduces Pain-Related Behavior in Mice
ARTICLE IN PRESS
eyes, heart, kidneys, gastrointestinal tract, spinal cord,and brain.28 In addition, chromaffin cells of the adrenalmedulla have been well characterized as a rich sourceof endogenous opioids that may be useful in pain con-trol.56 Pierce et al46 performed a study measuring plasmaendorphin levels before and after endurance exercise,defined as 45 minutes of high-intensity aerobics. Resultsindicated a significant increase in endorphin levels afterthe exercise as compared with levels before exercise.Such findings support the idea that opioid peptidesmay be released as a result of exercising vigorously fora specific amount of time.
Published studies reveal that incremental graded andshort-term anaerobic exercise leads to an increase inb-endorphin levels. The extent of this increase correlateswith the lactate concentration. During incrementalgraded exercise b-endorphin levels increase when theanaerobic threshold has been exceeded or at the pointof a disproportionate increase in lactate. In enduranceexercise performed at a steady-state between lactateproduction and elimination, blood b-endorphin levelsdo not increase until exercise duration exceeds approxi-mately 1 hour, with the increase being exponentialthereafter.16
Various intensities of exercise have been used in differ-ent studies, and it is currently unclear if there is a specificintensity of exercise that is necessary to produce hypoal-gesia. However, it has been reported that high-intensityexercise produces hypoalgesia.32
Despite the growing amount of experimental dataon physical exercise and nociception, the precisemechanisms through which high-intensity exercisecauses hyponociception remain elusive.33 The specificneurochemistry of nonopioid hyponociception is un-clear, but several neurotransmitters, such as serotoninand norepinephrine, have been implicated.25 Hoff-mann et al25 investigated whether skeletal musclestimulation in the rat would alter pain thresholds be-cause it has been suggested that the analgesic effectof exercise is mediated by activation of group III and/or IV afferents from skeletal muscle. Sixty minutes oflow-frequency muscle stimulation of the hind leg wasfound to increase pain thresholds in male rats. Butwhen the animals were pretreated with r-chlorophe-nylalanine methyl ester (PCPA), a serotonin synthesisblocker, the post-stimulatory analgesia was com-pletely abolished, indicating that serotonin systemsare involved in the analgesic response after musclestimulation.
Thus, the aim of the present study was to investigatehyponociception after a high-intensity extended exer-cise protocol, as well as the potential mechanisms forthis hyponociceptive response in mice. For this pur-pose, we used swimming exercise in the context ofa chemical behavioral model of nociception. To addressthe mechanisms that underlie hyponociception afterhigh-intensity extended swimming exercise, wetreated mice with a nonselective opioid receptorantagonist (naloxone), a tryptophan hydroxylaseenzyme inhibitor (p-chlorophenylalanine methyl ester)and adrenalectomy.
Materials and Methods
AnimalsMale Swiss mice (25 to 35 g) obtained from Federal
University of Santa Catarina (UFSC), were maintained atconstant room temperature of 22o 6 2�C undera 12-hour light/dark cycle (lights on at 6:00 AM), with ac-cess to food and water ad libitum. Mice (n = 8 to 10 ani-mals per group) were used only once and wereacclimatized to the laboratory for at least 1 hour beforetesting, which was conducted during the light cycle. Theexperiments were performed after approval of the pro-tocol by the Institutional Ethics Committee for AnimalResearch of Federal University of Santa Catarina. All ex-periments were conducted in accordance with currentguidelines for the care of laboratory animals and the eth-ical guidelines for investigations of experimental pain inconscious animals.67 The number of animals and intensi-ties of noxious stimuli used were the minimum necessaryto demonstrate a consistent effect of the exercise.
Swimming Exercise ProtocolAnimals were placed in a plastic box divided in eight
compartments (170 � 100 mm), filled with 35 L of waterat 37�C. Liquid soap (8 mL) was added to reduce surfacetension and to abolish the ‘‘floating’’ behavior.34 Aftereach exercise session, animals were gently dried witha cloth towel. Control (nonexercised) mice were allowedto swim for just 30 seconds each day and were thengently dried. Another nonexercised group was exposedto shallow water (3 cm of depth) for 30 minutes eachday, mice could stand normally with their head out ofthe water, did not need to swim.
The swimming exercise protocol was adapted from Ku-phal et al,34 as illustrated in Table 1. Mice were random-ized to nonexercised or exercised groups. Mice in theexercised group were exposed to water for 30 secondstwice on the first day and for 2 minutes twice on the sec-ond day. Mice were thus acclimated to the new environ-ment.31 On the third and fourth days, the animals weresubmitted to intermittent exercise. They swam for 10minutes and had a 5-minute rest on the third day;swam for 15 minutes and had a 5-minute rest on thefourth day. In total, mice performed 30 minutes of swim-ming exercise. On the fifth, sixth and seventh days theanimals swam continuously for 30 minutes. On theeighth day (24 hours after the last session of exercise)mice were submitted to behavioral testing.34
Physical Exercise Intensity (Blood LactateConcentration in Mice)
Blood lactate level was determined from 25 mL of tailcapillary blood during the last session of exercise.62 Theblood lactate concentration was analyzed at rest andthen at 10, 20, and 30 minutes during swimming exercise(Table 2). The separate blood samples were put ontoa glass fiber fleece where the erythrocytes were retainedand placed in 1.5-mL Eppendorf tubes containing 50 mLsodium fluoride (1%). To avoid blood lactate dilutionwith residual water in the tails of the animals, the mice

Table 1. Extended Swimming ProtocolPerformed by Mice Resulted in High-IntensityExercise and Was Performed Above MLSSIntensity
DAY(S)EXERCISE DURATION
(MIN)REST DURATION
(MIN) SESSIONS*TOTAL EXERCISE
(MIN)
1 .5 120 2 1
2 2 120 2 4
3 10 5 3 30
4 15 5 2 30
5 30 0 1 30
6 30 0 1 30
7 30 0 1 30
NOTE. Equivalent to the number of sessions performed (exercise plus rest period)
adapted Kuphal et al.34
Mazzardo-Martins et al The Journal of Pain 3
ARTICLE IN PRESS
were dried with a towel immediately before blood col-lection. The lactate concentrations were determinedwith a lactate analyzer (YSI Model 2700 SPORT; YSI, Yel-low Springs, OH). Before each test, the equipment wascalibrated according to the manufacturer’s instructions.
Behavioral TestingEach exercise training session was conducted in the
morning and the behavioral testing occurred in themorning of the next day, thus providing a 24-hour restperiod. This time interval minimized the potentially con-founding factor of stress-induced antinociception (SIA),which occurs on the order of minutes, not hours.54
Abdominal Constriction Induced byAcetic Acid and Peritoneal CapillaryPermeability in Mice
The abdominal constrictions were induced accordingto procedures described previously5,52 and resulted inthe contraction of the abdominal muscle together witha stretching of the hind limbs in response to anintraperitoneal (i.p.) injection of acetic acid (.6%, 0.45mL/mouse) at the time of the test. In the beginning ofthe experiment, mice were pretreated intravenouslywith 2.5% Evans blue dye solution (10 mL/kg), used asa marker of peritoneal capillary permeability.36 Onehour later the animals received the acetic acid injectionand were then individually placed in glass cylinders of20-cm diameter. The abdominal constrictions werecounted cumulatively over a period of 20 minutes. Thehyponociceptive activity was expressed as the reduction
Table 2. Blood Lactate Concentration in Mice
TIMES
CONTROL 30SECONDS
EXERCISE 30 MIN/1 SESSION
EXERCISE 30 MIN/5 SESSIONS
Rest 2.42 6 .18 2.37 6 .19 2.31 6 .14
10 min of exercise 5.57 6 .61 3.85 6 .66 3.43 6 .57
20 min of exercise 5.70 6 1.48 4.78 6 1.98 5.46 6 1.18
30 min of exercise 5.74 6 .64 7.13 6 .91 7.84 6 1.40
NOTE. Values are mean 6 SEM (mmol/L).
in the number of abdominal constrictions, for example,comparing control animals (nonexercised 30 secondsand nonexercised 30 minutes, shallow water) with exer-cise animals.
Immediately after the test, mice were killed by cervicaldislocation; the peritoneal cavity was washed with 1.5mL of sterile saline plus heparin (25 IU/mL); peritonealliquid was then collected with automatic pipettes. Totalleukocyte counts were performed using a Neubauerchamber via optical microscopy after diluting a sampleof the peritoneal fluid with Turk solution (1:20). A sampleof the collected fluid (1 mL) was centrifuged at 1000 rpmfor 10 minutes, and the absorbance of the supernatantwas read at 610 nm with an ELISA analyzer. The perito-neal capillary permeability induced by acetic acid isexpressed in terms of dye (mg/mL), which leaked intothe peritoneal cavity according to the standard curve ex-pected for Evans blue dye.36 The procedure describedabove was accomplished in the following groups: 1) non-exercised 30 seconds in water and 2) exercised, exposedto intense swimming exercise protocol.
Measurement of Locomotor ActivityTo exclude any unspecific locomotor effect caused by
the exercise, the locomotor activity was evaluated.Ambulatory behavior was assessed in an open-fieldtest.11,50,51 The apparatus consisted of a wooden boxmeasuring 40 � 60 � 50 cm. The floor of the arena wasdivided in 12 equal squares. At the start of each trial,a mouse was placed in the left corner of the field andwas allowed to freely explore the arena. The numberof squares crossed with all paws (crossing) was countedin a 6-minute session. The apparatus was cleaned witha solution of 10% ethanol between tests to hideanimal clues.
Involvement of Endogenous OpioidsTo investigate the role played by endogenous opioids
in the hyponociceptive effect of exercise, nonexercisedmice were pretreated with naloxone (1 mg/kg, i.p., a non-selective opioid receptor antagonist) or saline (.9% NaClsolution, 10 mL/kg, i.p.). After 20 minutes, the animals re-ceived an intraperitoneal injection of acetic acid. An-other group of control mice received an injection ofsaline (10 mL/kg, i.p.) or naloxone (1 mg/kg, i.p.); after20 minutes, they received morphine (5 mg/kg, subcuta-neous, s.c.), and 30 minutes later, the acetic acid was in-traperitoneally administered. The exercised group waspretreated with saline (10 mL/kg, i.p.) or naloxone (1mg/kg, i.p.); after 20 minutes, the animals received an in-traperitoneal injection of acetic acid. The abdominalconstrictions were counted for 20 minutes.52
Role of Endogenous Opioid Release byAdrenal Glands
To evaluate the involvement of endogenous opioid re-lease from adrenal glands in the hyponociceptive effectcaused by swimming exercise animals were anesthetizedwith xylazine (10 mg/kg, i.p.) and ketamine (80 mg/kg,i.p.); both adrenal glands were removed through a dorsal

4 The Journal of Pain Exercise Reduces Pain-Related Behavior in Mice
ARTICLE IN PRESS
incision, as described previously.53,59 No postoperativeanalgesia was used because it can interfere withexperimental results. After the surgery, the animalswere returned to their cages and had free access tofood and drink. The water was replaced with saline tomaintain a physiological plasma sodium concentration.One week later, the animals were submitted to theswimming exercise protocol and then injected withacetic acid. For this experiment, the following groupswere used: i) nonexercised, 30 seconds of swimming,ii) nonexercised/sham-operated, dorsal incision, iii)sham-operated /exercised group, 30 minutes/5 days ofswimming and dorsal incision, iv) nonexercised/adrenal-ectomy, both adrenal glands removed through dorsalincision, and v) adrenalectomy/exercise.
Involvement of the Serotonergic SystemTo assess the possible contribution of endogenous
serotonin to the hyponociceptive effect of exercise,animals were pretreated with r-chlorophenylalaninemethyl ester (PCPA, 100 mg/kg, i.p., an inhibitor ofserotonin synthesis) or with vehicle, once a day for 4consecutive days. Then, 24 hours after the last PCPA orvehicle injection, animals were tested in the acetic acid–induced abdominal constriction test.52,53 For this experiment,the following groups were used: i) nonexercised group,ii) nonexercised/PCPA group, iii) exercised group,iv) morphine group, v) PCPA/morphine group, andvi) PCPA/exercise group.
DrugsThe following substances were used: morphine hydro-
chloride, acetic acid (Merck, Darmstadt, Germany),naloxone hydrochloride, r-chlorophenylalanine methylester (PCPA), sodium fluoride, xylazine, and ketamine(purchased from Sigma Chemical Company, St Louis,MO, USA). NaCl was supplied by Merck (Haar, Germany).Drugs were dissolved in isotonic saline solution (NaCl.9%) immediately before use. The drugs were adminis-tered by intraperitoneal and subcutaneous routes.
Statistical AnalysisAll experimental results are given as mean 6 SEM.
Comparisons between experimental and control groupswere performed by t test, 1-way ANOVA followed bythe Newman-Keuls test, or 2-way ANOVA followed byTukey HSD test for post hoc comparison when appropri-ate. A value of P < .05 was considered significant.
Figure 1. Effect of swimming exercise on acetic acid-inducedabdominal constrictions. Each column represents the mean of8 animals and error bars indicate SEM. Asterisks denote signifi-cance levels compared with the nonexercised group, **P < .01,1-way ANOVA followed by the Newman-Keuls test.
Results
Animals Performed a High-IntensityExtended Swimming Exercise Protocol
Endurance capacity can, from a metabolic point ofview, be regarded as the highest steady-state energysupply from oxidative phosphorylation.37 Therefore,another approach to assess aerobic endurance perfor-mance is the determination of the highest constant exer-cise intensity that can be maintained for a longer period
of time without a continuous rise in blood lactate. Themaximal lactate steady state (MLSS) is presumably thehighest blood lactate concentration at which equilib-rium between lactate formation and elimination canbe achieved during prolonged exercise at constantwork load.
An increase in blood of not more than 1 mmol/L be-tween 10 and 30 minutes during the constant-load trialsappears to be the most reasonable procedure for MLSSdetermination.4,58 The MLSS represents the upperborder of constant load endurance training. Intensitiesabove the MLSS have been used to guide intervaltraining sessions in different endurance sports.8,13,40
At the end of the last day of the exercise protocol, theblood lactate level in the swimming exercise group was3.43 6 .57 mmol/L at 10 minutes and 7.84 6 1.40 mmol/L at 30 minutes. We observed an increase in lactate bloodconcentration of more than 1 mmol/L between 10 and 30minutes during the constant-load swimming exercise.This suggests that the mice in our study performed theexercise at above-MLSS intensity.23,45
Abdominal Constriction ResponseCaused by Intraperitoneal Injection ofAcetic Acid and Peritonitis in Mice
The results presented in Fig 1 show that swimming ex-ercise reduced the acetic acid-induced abdominal con-strictions in mice by 44% 6 3% when compared withthe control group (nonexercised). Notably, the 2 controlgroups (nonexercised 30 seconds and nonexercised 30minutes in shallow water) had a similar response. There-fore, subsequent experiments were performed usingonly the nonexercised 30-second control group.
The swimming exercise, in the same conditions thatcaused inhibition of acetic acid-induced nociception inmice, did not alter cell migration (Fig 2A, 2B, and 2C) or
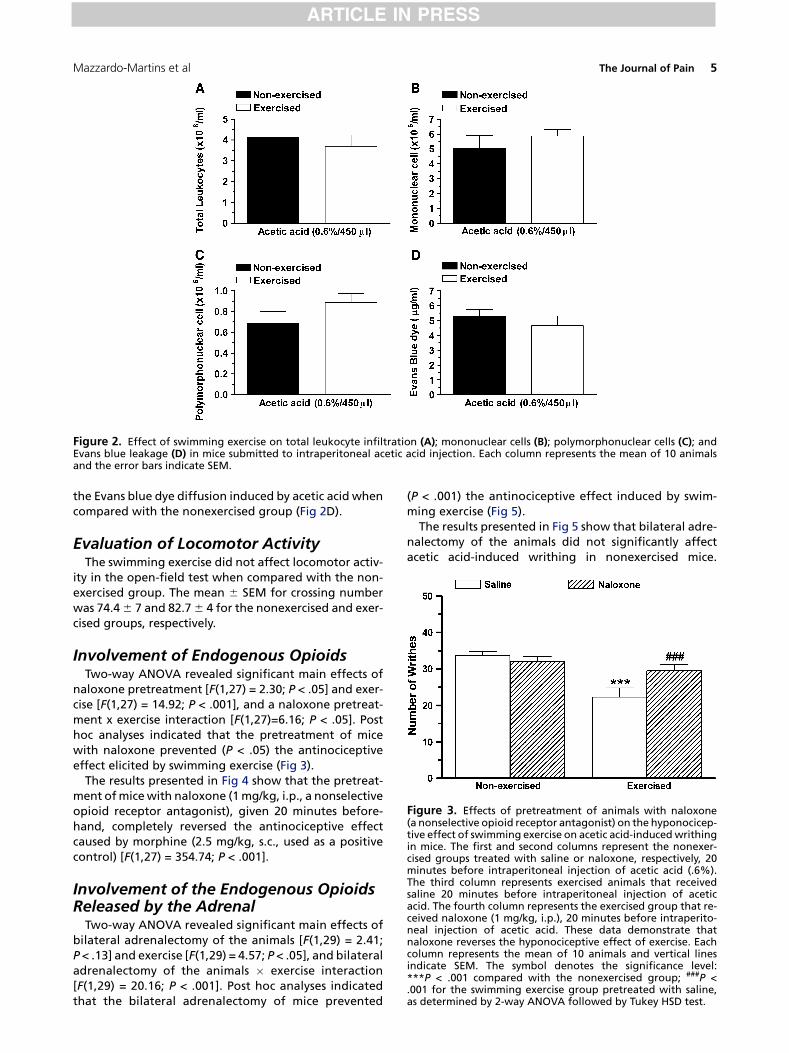
Figure 2. Effect of swimming exercise on total leukocyte infiltration (A); mononuclear cells (B); polymorphonuclear cells (C); andEvans blue leakage (D) in mice submitted to intraperitoneal acetic acid injection. Each column represents the mean of 10 animalsand the error bars indicate SEM.
Figure 3. Effects of pretreatment of animals with naloxone(a nonselective opioid receptor antagonist) on the hyponocicep-tive effect of swimming exercise on acetic acid-induced writhingin mice. The first and second columns represent the nonexer-cised groups treated with saline or naloxone, respectively, 20minutes before intraperitoneal injection of acetic acid (.6%).The third column represents exercised animals that receivedsaline 20 minutes before intraperitoneal injection of aceticacid. The fourth column represents the exercised group that re-ceived naloxone (1 mg/kg, i.p.), 20 minutes before intraperito-neal injection of acetic acid. These data demonstrate thatnaloxone reverses the hyponociceptive effect of exercise. Eachcolumn represents the mean of 10 animals and vertical linesindicate SEM. The symbol denotes the significance level:***P < .001 compared with the nonexercised group; ###P <.001 for the swimming exercise group pretreated with saline,as determined by 2-way ANOVA followed by Tukey HSD test.
Mazzardo-Martins et al The Journal of Pain 5
ARTICLE IN PRESS
the Evans blue dye diffusion induced by acetic acid whencompared with the nonexercised group (Fig 2D).
Evaluation of Locomotor ActivityThe swimming exercise did not affect locomotor activ-
ity in the open-field test when compared with the non-exercised group. The mean 6 SEM for crossing numberwas 74.4 6 7 and 82.7 6 4 for the nonexercised and exer-cised groups, respectively.
Involvement of Endogenous OpioidsTwo-way ANOVA revealed significant main effects of
naloxone pretreatment [F(1,27) = 2.30; P < .05] and exer-cise [F(1,27) = 14.92; P < .001], and a naloxone pretreat-ment x exercise interaction [F(1,27)=6.16; P < .05]. Posthoc analyses indicated that the pretreatment of micewith naloxone prevented (P < .05) the antinociceptiveeffect elicited by swimming exercise (Fig 3).
The results presented in Fig 4 show that the pretreat-ment of mice with naloxone (1 mg/kg, i.p., a nonselectiveopioid receptor antagonist), given 20 minutes before-hand, completely reversed the antinociceptive effectcaused by morphine (2.5 mg/kg, s.c., used as a positivecontrol) [F(1,27) = 354.74; P < .001].
Involvement of the Endogenous OpioidsReleased by the Adrenal
Two-way ANOVA revealed significant main effects ofbilateral adrenalectomy of the animals [F(1,29) = 2.41;P < .13] and exercise [F(1,29) = 4.57; P < .05], and bilateraladrenalectomy of the animals � exercise interaction[F(1,29) = 20.16; P < .001]. Post hoc analyses indicatedthat the bilateral adrenalectomy of mice prevented
(P < .001) the antinociceptive effect induced by swim-ming exercise (Fig 5).
The results presented in Fig 5 show that bilateral adre-nalectomy of the animals did not significantly affectacetic acid-induced writhing in nonexercised mice.

Figure 4. Effect of pretreatment of animals with morphine(a nonselective opioid receptor agonist) on acetic acid-inducedwrithing in mice. The first and second columns represent thecontrol groups treated with saline or naloxone, respectively,20 minutes after they received saline (10 mL/kg, s.c.), 30 minutesbefore intraperitoneal injection of acetic acid (.6%). The thirdcolumn represents animals pretreated with saline (i.p.) that 20minutes after received morphine (2.5 mg/kg, s.c.) 30 minutes be-fore intraperitoneal injection of acetic acid. The fourth columnrepresents the animals pretreated with naloxone (1 mg/kg,i.p.) that 20 minutes after received morphine, 30 minutes beforeintraperitoneal injection of acetic acid. These data demonstratethat naloxone reverses the antinociceptive effect of morphine.Each column represents the mean of 10 animals and verticallines indicate SEM. The symbol denotes the significance level:***P < .001 compared with the control group; ###P < .001 com-pared with the group pretreated with morphine. Two-way AN-OVA was followed by Tukey HSD test.
Figure 5. Effect of bilateral adrenalectomy, performed 1 weekbefore the swimming exercise protocol, demonstrating the hy-ponociceptive effect of swimming exercise on acetic acid-induced writhing in mice. The first column represents thesham-operated/nonexercised group; the second: adrenalec-tomy/nonexercised group; the third: sham-operated/exercisedgroup; and the fourth: adrenalectomy/exercised group. Eachcolumn represents the mean of 10 animals and vertical lines in-dicate SEM. The symbol denotes the significance level: *P < .05compared with sham-operated/nonexercised group; ##P < .01compared with the sham-operated/exercised group. Two-wayANOVA was followed by Tukey HSD test.
6 The Journal of Pain Exercise Reduces Pain-Related Behavior in Mice
ARTICLE IN PRESS
Moreover, the nociceptive response caused by acetic aciddid not significantly differ between nonexercised andsham-operated/nonexercised groups (data not shown).
Involvement of the Serotonergic SystemTwo-way ANOVA revealed significant main effects of
PCPA pretreatment [F(1,31) = 11.00; P < .01] and exercise[F(1,31) = 23.52; P < .001], and a PCPA pretreatment� ex-ercise interaction [F(1,31) = 18.23; P < .001]. Post hoc anal-yses indicated that the pretreatment of mice with PCPAprevented (P < .001) the antinociceptive effect causedby swimming exercise (Fig 6).
The results presented in Fig 7 show that the pretreat-ment of animals with PCPA (100 mg/kg, i.p., for 4 consec-utive days) completely reversed the antinociceptiveeffect caused by morphine (2.5 mg/kg, s.c., used as a pos-itive control) [F(1,30) = 154.62; P < .01].
DiscussionThe results presented here show for the first time that
high-intensity exercise produces hyponociceptive effectson chemical models of nociception in mice. The most rel-evant findings of the present work are the following: i)intense swimming exercise reduced the nociceptive re-sponse elicited by acetic acid and ii) the hyponociceptiveeffect of exercise in the acetic acid test was significantlyreversed by intraperitoneal treatment of animals withnaloxone (a nonselective opioid receptor antagonist),PCPA (a tryptophan hydroxylase inhibitor) or bilateral
adrenalectomy. Our data support the hypothesis thathigh-intensity extended swimming exercise reducesacute pain via opioid and serotonergic systems.
Exercise Reduces Acetic Acid-InducedAbdominal Constrictions
The results reported here indicate for the first timethat intense swimming exercise, at an intensity that didnot produce any important motor dysfunction or anydetectable side effects, produced a marked hyponoci-ceptive effect on acetic acid-induced visceral nocicep-tion. Moreover, acetic acid injection in the peritonealcavity of mice promotes the release of many inflamma-tory mediators such as prostaglandin, bradykinin,substance P, tumor necrosis factor-a (TNF-a), interleu-kin-1b, interleukin-8, and others.5,27,49 In addition,these substances stimulate primary afferent neurons,enhancing aspartate and glutamate release in thecerebrospinal fluid.14,66
It has been demonstrated that swimming (particularlyin cold water) produces the well-known phenomenon ofstress-induced antinociception (SIA), which is mediatedby both opioid and nonopioid mechanisms.57 A nonop-ioid mechanism includes the release of endogenouscorticosteroids.26 Additionally, corticosteroids playsa critical role in the regulation of the inflammatoryresponse, such that they can suppress inflammation, inpart, by reducing cellular infiltration and plasma proteinextravasation. The results of the present study also dem-onstrated that intense swimming exercise, at levels thatcause hyponociception, was not capable of reducing leu-kocyte infiltration and plasma protein extravasation(Evans blue dye diffusion) induced by acetic acid in
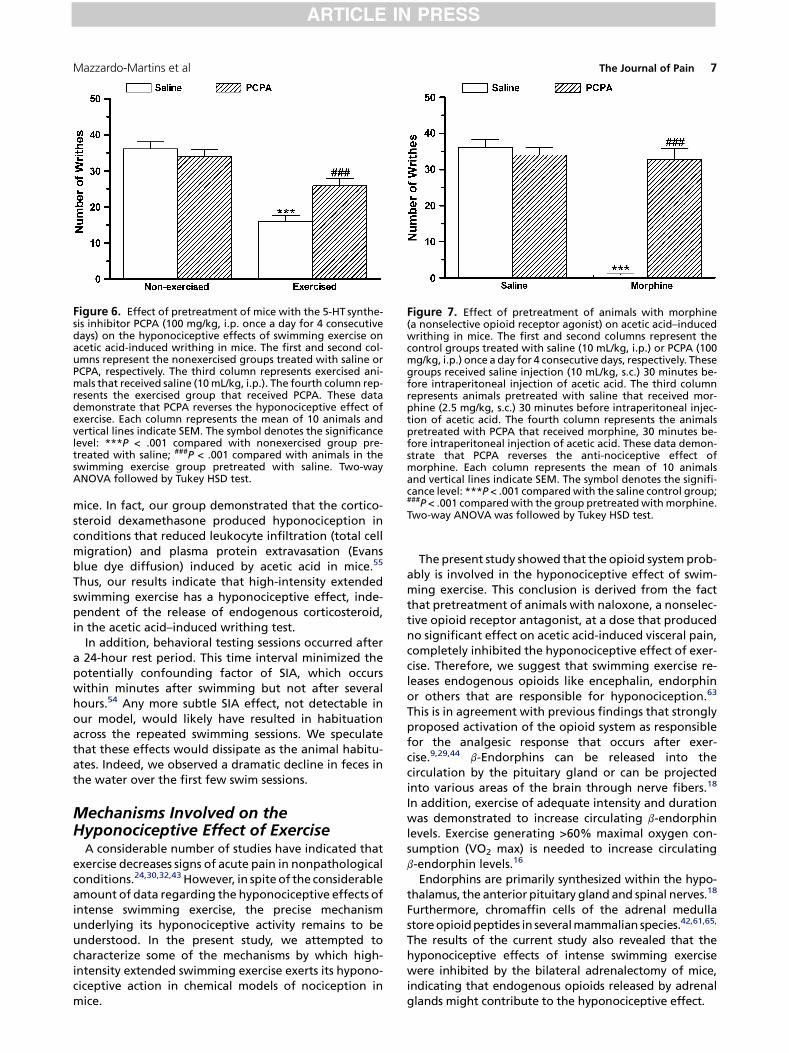
Figure 6. Effect of pretreatment of mice with the 5-HT synthe-sis inhibitor PCPA (100 mg/kg, i.p. once a day for 4 consecutivedays) on the hyponociceptive effects of swimming exercise onacetic acid-induced writhing in mice. The first and second col-umns represent the nonexercised groups treated with saline orPCPA, respectively. The third column represents exercised ani-mals that received saline (10 mL/kg, i.p.). The fourth column rep-resents the exercised group that received PCPA. These datademonstrate that PCPA reverses the hyponociceptive effect ofexercise. Each column represents the mean of 10 animals andvertical lines indicate SEM. The symbol denotes the significancelevel: ***P < .001 compared with nonexercised group pre-treated with saline; ###P < .001 compared with animals in theswimming exercise group pretreated with saline. Two-wayANOVA followed by Tukey HSD test.
Figure 7. Effect of pretreatment of animals with morphine(a nonselective opioid receptor agonist) on acetic acid–inducedwrithing in mice. The first and second columns represent thecontrol groups treated with saline (10 mL/kg, i.p.) or PCPA (100mg/kg, i.p.) once a day for 4 consecutive days, respectively. Thesegroups received saline injection (10 mL/kg, s.c.) 30 minutes be-fore intraperitoneal injection of acetic acid. The third columnrepresents animals pretreated with saline that received mor-phine (2.5 mg/kg, s.c.) 30 minutes before intraperitoneal injec-tion of acetic acid. The fourth column represents the animalspretreated with PCPA that received morphine, 30 minutes be-fore intraperitoneal injection of acetic acid. These data demon-strate that PCPA reverses the anti-nociceptive effect ofmorphine. Each column represents the mean of 10 animalsand vertical lines indicate SEM. The symbol denotes the signifi-cance level: ***P < .001 compared with the saline control group;###P < .001 compared with the group pretreated with morphine.Two-way ANOVA was followed by Tukey HSD test.
Mazzardo-Martins et al The Journal of Pain 7
ARTICLE IN PRESS
mice. In fact, our group demonstrated that the cortico-steroid dexamethasone produced hyponociception inconditions that reduced leukocyte infiltration (total cellmigration) and plasma protein extravasation (Evansblue dye diffusion) induced by acetic acid in mice.55
Thus, our results indicate that high-intensity extendedswimming exercise has a hyponociceptive effect, inde-pendent of the release of endogenous corticosteroid,in the acetic acid–induced writhing test.
In addition, behavioral testing sessions occurred aftera 24-hour rest period. This time interval minimized thepotentially confounding factor of SIA, which occurswithin minutes after swimming but not after severalhours.54 Any more subtle SIA effect, not detectable inour model, would likely have resulted in habituationacross the repeated swimming sessions. We speculatethat these effects would dissipate as the animal habitu-ates. Indeed, we observed a dramatic decline in feces inthe water over the first few swim sessions.
Mechanisms Involved on theHyponociceptive Effect of Exercise
A considerable number of studies have indicated thatexercise decreases signs of acute pain in nonpathologicalconditions.24,30,32,43 However, in spite of the considerableamount of data regarding the hyponociceptive effects ofintense swimming exercise, the precise mechanismunderlying its hyponociceptive activity remains to beunderstood. In the present study, we attempted tocharacterize some of the mechanisms by which high-intensity extended swimming exercise exerts its hypono-ciceptive action in chemical models of nociception inmice.
The present study showed that the opioid system prob-ably is involved in the hyponociceptive effect of swim-ming exercise. This conclusion is derived from the factthat pretreatment of animals with naloxone, a nonselec-tive opioid receptor antagonist, at a dose that producedno significant effect on acetic acid-induced visceral pain,completely inhibited the hyponociceptive effect of exer-cise. Therefore, we suggest that swimming exercise re-leases endogenous opioids like encephalin, endorphinor others that are responsible for hyponociception.63
This is in agreement with previous findings that stronglyproposed activation of the opioid system as responsiblefor the analgesic response that occurs after exer-cise.9,29,44 b-Endorphins can be released into thecirculation by the pituitary gland or can be projectedinto various areas of the brain through nerve fibers.18
In addition, exercise of adequate intensity and durationwas demonstrated to increase circulating b-endorphinlevels. Exercise generating >60% maximal oxygen con-sumption (VO2 max) is needed to increase circulatingb-endorphin levels.16
Endorphins are primarily synthesized within the hypo-thalamus, the anterior pituitary gland and spinal nerves.18
Furthermore, chromaffin cells of the adrenal medullastore opioid peptides in several mammalian species.42,61,65,
The results of the current study also revealed that thehyponociceptive effects of intense swimming exercisewere inhibited by the bilateral adrenalectomy of mice,indicating that endogenous opioids released by adrenalglands might contribute to the hyponociceptive effect.

8 The Journal of Pain Exercise Reduces Pain-Related Behavior in Mice
ARTICLE IN PRESS
It is well known that serotonin (5-HT) pathways withinthe CNS arise from a series of nuclei situated in themidline of the brain stem, the raphe nuclei, which repre-sent the richest source of neuronal 5-HT synthesized inthe mammalian brain.15,39 In addition, several studieshave shown that the bulbar serotonin nuclei thatproject to the spinal dorsal horn may suppressincoming noxious input to the spinal cord and inhibitpain transmission.2,3 Moreover, multiple 5-HT receptortypes within the spinal cord appear to fulfill differentroles in the control of nociception.
Skeletal muscle tissue consumes branched chainamino acids (BCAAs; ie, leucine, isoleucine, and valine).This consumption of BCAAs is increased during exercise;the BCAA concentration in blood will then tend to de-crease. BCAAs enter the brain via the same carrier astryptophan. Thus, if BCAA concentration goes downwithout a corresponding change in tryptophan level,more tryptophan will enter the brain. Tryptophan isthe precursor of serotonin. Prolonged exercise has twoeffects. First, BCAA concentration decreases, thereby al-tering the ratio of tryptophan-BCAA entering the brainin favor of tryptophan. Second, prolonged exerciseleads to increased levels of fatty acids in the blood.The increase in free fatty acids causes an increase inthe ratio of free versus bound plasma tryptophan,which in turn causes a further increase in the amountof tryptophan entering the brain. The increased levelsof brain tryptophan lead to an increase in the effectsof serotonergic transmission.41
Physical exercise results in an increase in the availabil-ity of brain neurotransmitters (dopamine, noradrenalin,and serotonin) that typically are reduced in depressivepatients.47 Similarly, studies involving animals10 and hu-mans1 have indicated that exercise increases levels of se-rotonin in the brain. Thus, the mechanisms underlyingthe antidepressant-like effect of exercise are similar tothose obtained in the present study. This assertion is sup-ported by the demonstration that the depletion of en-dogenous serotonin using a tryptophan hydroxylaseinhibitor PCPA, at a dose known to decrease corticallevels of serotonin and reverse morphine antinocicep-
tion,38,53 largely antagonized the hyponociceptiveeffect of physical exercise.
Of note is one discrepancy between our results andthose reported by Quintero et al.48 Whereas Quinteroet al48 demonstrated that rats subjected to 10 to 20 min-utes of forced swimming in cool (24� to 26�C) water for 3days presented thermal and chemical hyperalgesia,which according to the authors was an effect mediatedby reduction in central serotonin activity. This might beexplained primarily by the differences in animal species(rats vs mouse) and modalities tested (formalin and hotplate vs acetic acid models), which could be encoded bydifferent nociceptive signaling mechanisms. Further-more, the differences in exercise protocol can alsoexplain this opposite results, such as: the intensity ofstress-induced hyperalgesia (SIH) may inversely correlatewith water temperature. Of fact, swimming in cooler wa-ter may facilitate SIH, whereas swimming in warmerwater facilitates exercise-induced antinociception.34
However, the stress during initial swim sessions of thepresent study might induce SIH; we speculate that thesewould dissipate as the animal habituates. Indeed, we ob-served a decrease in feces in the water over the first fewswim sessions. Finally, the intensity of exercise, in theQuintero et al48 study, the rats did not swim allthe time (20 minutes), whereas in the present study,the soap added prevented floating behavior. However,future studies are expected to determine the exact themechanisms that are involved in exercise-induced hypo-nociception.
In summary, the present results provide convincing ev-idence that high-intensity extended swimming exerciseexerts a pronounced hyponociceptive effect, as revealedby the suppression of nociceptive responses generatedby acetic acid in mice. These experiments are the firstto show that high-intensity extended swimming exerciseactivates several mechanisms to reduce pain-related be-havior in mice. Extended swimming exercise reversedthe nociceptive response to acetic acid in an acute vis-ceral pain model through the activation of opioid and se-rotonin receptors. Endogenous opioid release by adrenalglands is likely to be involved in this effect.
References
1. aan het Rot M, Collins KA, Fitterling HL: Physical exerciseand depression. Mt Sinai J Med 76:204-214, 2009
2. Alhaider AA, Lei SZ, Wilcox GL: Spinal 5-HT3 receptor-mediated antinociception: Possible release of GABA. J Neu-rosci 11:1881-1888, 1991
3. Basbaum AI, Fields HL: Endogenous pain control systems:Brainstem spinal pathways and endorphin circuitry. AnnuRev Neurosci 7:309-338, 1984
4. Beneke R: Methodological aspects of maximal lactatesteady state: Implications for performance testing. Eur JAppl Physiol 89:204-214, 2003
5. Collier HOJ, Dinneen JC, Johnson CA, Schneider C: The ab-dominal constriction response and its suppression by anal-
gesic drugs in the mouse. Br J Pharmacol Chemother 32:295-310, 1968
6. Cotman CW, Berchtold NC, Christie LA: Exercise buildsbrain health: Key roles of grown factor cascades and inflam-mation. Trends Neurosci 30:467-472, 2007
7. Davranche K, McMorris T: Specific effects of acute moder-ate exercise on cognitive control. Brain Cogn 69:565-570,2009
8. Dickhuth HH, Yin L, Niess A, Rocker K, Mayer F,Heitkamp H-C, Horstmann T: Vantilatory, lactate-derivedand catecholamine thresholds during incremental treadmillrunning: Relationship and reproducibility. Int J Sports Med20:122-127, 1999
9. Droste C, Meyer-Blankenburg M, Greenlee MW,Roskamm H: Effect of physical exercise on pain thresh-olds and plasma beta-endorphins in patients with silent

Mazzardo-Martins et al The Journal of Pain 9
ARTICLE IN PRESS
and symptomatic myocardial ischemia. Eur Heart J 9:25-33, 1988
10. Dunn AL, Reigle TG, Youngstedt SD, Armstrong RB,Dishman RK: Brain norepinephrine and metabolites aftertreadmill training and wheel running in rats. Med Sci SportsExerc 28:204-209, 1996
11. Eckeli AL, Dach F, Rodrigues AL: Acute treatments withGMP produces antidepressant-like effects in mice. Neurore-port 11:1839-1843, 2000
12. Farrell PA: Exercise and endorphins: Male responses.Med Sci Sports Exerc 17:89-93, 1985
13. Faude O, Meyer T, Scharhag J, Weins F, Urhausen A,Kindermann W: Volume vs intensity in the training of com-petitive swimmers. Int J Sports Med 29:906-912, 2008
14. Feng Y, Cui M, Willis WD: Effect of anticonvulsant gaba-pentin on visceral nociception and its relationship withamino acid neurotransmitters released from spinal cord. Bei-jing Da Xue Xue Bao 35:307-310, 2003
15. Fields HL, Heinricher MM, Mason P: Neurotransmittersin nociceptive modulatory circuits. Annu Rev Neurosci 14:219-245, 1991
16. Goldfarb AH, Hatfield BD, Armstrong D, Potts J:Plasma beta-endorphin concentration: Response to inten-sity and duration of exercise. Med Sci Sports Exerc 22:241-244, 1990
17. Goldfarb AH, Hatfield BD, Sforzo GA, Flynn MG: Serum[beta]-endorphin levels during a graded exercise test toexhaustion. Med Sci Sports Exerc 19:78-82, 1987
18. Goldfarb AH, Jamurtas AZ: Beta-endorphin response toexercise: An update. Sports Med 24:8-16, 1997
19. Goldfarb AH, Jamurtas AZ, Kamimori GH, Hegde S,Otterstetter R, Brown DA: Gender effect on beta-endorphin response to exercise. Med Sci Sports Exerc 30:1672-1676, 1998
20. Gowans SE, de Hueck A: Effectiveness of exercise inmanagement of fibromyalgia. Curr Opin Rheumatol 16:138-142, 2004
21. Greenwood BN, Fleshner M: Exercise, learned help-lessness and the stress-resistant brain. Neuromol Med10:81-98, 2008
22. Hayes K, Sprague S, Guo M, Davis W, Friedman A,Kumar A, Jimenez DF, Ding Y: Forced, not voluntary, exerciseeffectively induces neuroprotection in stroke. Acta Neuro-pathol 115:289-296, 2008
23. Hill AV, Lupton H: Muscular exercise, lactic acid and thesupply and utilization of oxygen. Q J Med 16:135-1371, 1923
24. Hoffman MD, Shepanski MA, Mackenzie SP, Clifford PS:Experimentally induced pain perception is acutely reducedby aerobic exercise in people with chronic low back pain. JRehabil Res Dev 42:183-190, 2005
25. Hoffmann P, Skarphedinsson JO, Delle M, Thoren P: Elec-trical stimulation of the gastrocnemius muscle in the sponta-neously hypertensive rat increases the pain threshold: Roleof different serotonergic receptors. Acta Physiol Scand138:125-131, 1990
26. Hohmann AG, Suplita RL, Bolton NM, Neely MH,Fegley D, Mangieri R, Krey JF, Walker JM, Holmes PV,Crystal JD, Duranti A, Tontini A, Mor M, Tarzia G,Piomelli D: An endocannabinoid mechanism for stress-induced analgesia. Nature 435:1108-1112, 2005
27. Ikeda Y, Ueno A, Naraba H, Oh-Ishi S: Involvement ofvanilloid receptor VR1 and prostanoids in the acid-induced writhing responses of mice. Life Sci 69:2911-2919, 2001
28. Imura H, Nakai Y: ‘‘Endorphins’’ in pituitary and othertissues. Annu Rev Physiol 43:265-278, 1981
29. Janal MN, Colt EW, Clark WC, Glusman M: Pain sensi-tivity, mood and plasma endocrine levels in man follow-ing long-distance running: Effects of naloxone. Pain 19:13-25, 1984
30. Kemppainen P, Hamalainen O, Kononen M: Different ef-fects of physical exercise on cold pain sensitivity in fighter pi-lots with and without the history of acute in-flight neck painattacks. Med Sci Sports Exerc 30:577-582, 1998
31. Kim J, Arias EB, Cartee GD: Effects of gender and priorswim exercise on glucose uptake in isolated skeletal musclesfrom mice. J Physiol Sci 56:305-312, 2006
32. Koltyn KF: Analgesia following exercise: A review.Sports Med 29:85-98, 2000
33. Koltyn KF, Umeda M: Exercise, hypoalgesia and bloodpressure. Sports Med 36:207-214, 2006
34. Kuphal KE, Fibuch EE, Taylor BK: Extended swimming ex-ercise reduces inflammatory and peripheral neurophaticpain in rodents. J Pain 8:989-997, 2007
35. Langenfeld ME, Hart LS, Kao PC: Plasma [beta]-endorphin responses to one hour bicycling and running at60% [latin capital V with dot above] O2 max. Med Sci SportsMed 19:83-86, 1987
36. Lucena GM, Gadotti VM, Maffi LC, Silva GS, Azevedo MS,Santos ARS: Antinociceptive and anti-inflammatory proper-ties from the bulbs of Cipura paludosa Aubl. J Ethnopharma-col 112:19-25, 2007
37. Mader A, Heck H: A theory of metabolic origin of the‘‘anaerobic threshold.’’ Int J Sports Med 7:45-65, 1986
38. Mendes GL, Santos ARS, Malheiros A, Filho VC,Yunes RA, Calixto JB: Assessment of mechanisms involvedin antinociception caused by sesquiterpene polygodial. JPharmacol Exp Ther 292:164-172, 2000
39. Millan MJ: Descending control of pain. Prog Neurobiol66:355-474, 2002
40. Mujika I, Padilla S: Cardiorespiratory and metaboliccharacteristics of detraining in humans. Med Sci Sports Exerc33:413-421, 2001
41. Newsholme EA, Blomslrand E: Tryptophan 5-hydroxy-tryptamine and a possible explanation for central fatigue,in Gandevia SC, Enoka HM, McComas AJ, et al (eds): Fatigue:Neural and Muscular Mechanisms: Advances in Medicineand Biology, Vol. 20. New York & London, Plenum Press,1995, pp 384-315
42. North RA, Egan TM: Actions and distributions of opioidpeptides in peripheral tissues. Br Med Bull 39:71-75, 1983
43. O’Connor PJ, Cook DB: Exercise and pain: The neuro-biology, measurement, and laboratory study of pain inrelation to exercise in humans. Exerc Sport Sci Rev 27:119-166, 1999
44. Olausson B, Eriksson E, Ellmarker L, Rydenhag B,Shyu BC, Andersson SA: Effects of naloxone on dental pulppain threshold following muscle exercise and low frequencytranscutaneous nerve stimulation: A comparative study inman. Acta Physiol Scand 126:299-305, 1986

10 The Journal of Pain Exercise Reduces Pain-Related Behavior in Mice
ARTICLE IN PRESS
45. Peronnet F, Aguilaniu B: Lactic acid buffering, non-metabolic CO2 and exercise hyperventilation: A critical reap-praisal. Respir Physiol Neurobiol 150:4-18, 2006
46. Pierce EF, Eastman NW, Tripathi HL, Olson KG,Dewey WL: b-Endorphin response to endurance exercise: Re-lationship to exercise dependence. Percept Mot Skills 77:167-770, 1993
47. Pierce D, Kuppart I, Harry D: Urinary epinephrine andnorepinephrine levels in women athletes during trainingand competition. Eur J Appl Occup Physiol 36:1-6, 1979
48. Quintero L, Moreno M, Avila C, Arcaya J, Maixner W,Suarez-Roca H: Long-lasting delayed hyperalgesia after sub-chronic swim stress. Pharmacol Biochem Behav 67:449-458,2000
49. Ribeiro RA, Vale MV, Thomazzi SM, Paschoalato ABP,Poole S, Ferreira SH, Cunha FQ: Involvement of resident mac-rophages and mast cells in the writhing nociceptive re-sponse induced by zymosan and acetic acid in mice. Eur JPharmacol 387:111-118, 2000
50. Rodrigues AL, da Silva GL, Mateussi AS, Fernandes ES,Miguel OG, Yunes RA, Calixto JB, Santos ARS: Involvementof monoaminergic system in the antidepressant-like effectof the hydroalcoholic extract of Siphocampylus verticillatus.Life Sci 70:1347-1358, 2002
51. Rosa AO, Lin J, Calixto JB, Santos ARS, Rodrigues AL: In-volvement of NMDA receptors and L-arginine-nitric oxidepathway in the antidepressant-like effects of zinc in mice.Behav Brain Res 144:87-93, 2003
52. Santos ARS, Gadotti VM, Oliveira G, Tibola D, Paszuk AF,Neto A, Spindola HM, Souza MM, Rodrigues AL, Calixto JB:Mechanisms involved in the antinociception caused by ag-matine in mice. Neuropharmacol 48:1021-1034, 2005
53. Santos ARS, Miguel OG, Yunes OG, Calixto JB: Antinoci-ceptive properties of the new alkaloid, cis-8, 10-di-Npropyllo-belidiolhydrochloridedihydrate isolated fromSiphocampylusverticillatus: Evidence for the mechanism of action. J Pharma-col Exp Ther 289:417-426, 1999
54. Schwarz L, Kindermann W: Changes in beta-endorphinlevels in response to aerobic and anaerobic exercise. SportsMed 13:25-36, 1992
55. Smiderle FR, Olsen LM, Carbonero ER, Baggio CH,Freitas CS, Marcon R, Santos ARS, Gorin PA, Iacomini M:Anti-inflammatory and analgesic properties in a rodent
model of a (1-3), (1-6)- linked beta-glucan isolated fromPleurotus pulmonarius. Eur J Pharmacol 597:86-91, 2008
56. Stein C, ed: Opioids in Pain Control: Basic and Clinical As-pects. Cambridge, United Kingdom, Cambridge UniversityPress, 1999
57. Terman GW, Morgan MJ, Liebeskind JC: Opioid and non-opioid stress analgesia from cold water swim: Importance ofstress severity. Brain Res 372:167-171, 1986
58. Urhausen A, Coen B, Weiler B, Kindermann W: Individ-ual anaerobic threshold and maximum lactate steady state.Int J Sports Med 14:134-139, 1993
59. Vaz ZR, Filho VC, Yunes RA, Calixto JB: Antinociceptiveaction of 2- (4-bromobenzoyl)-3-methyl-4,6-dimethoxy ben-zofuran, a novel Xanthoxyline derivative on chemical andthermal models of nociception in mice. J Phamacol ExpTher 278:304-312, 1996
60. Vega SR, Struder HK, Wahrmann BV, Schmidt A, Bloch W,Hollmann W: Acute BDNF and cortisol response to low inten-sity exercise and following ramp incremental exercise toexhaustion in humans. Brain Res 1121:59-65, 2006
61. Viveros OH, Diliberto EJ Jr., Hazum E, Chang KJ: Opiate-like materials in the adrenal medulla: Evidence for storageand secretion with catecholamines. Mol Pharmacol 16:1101-1108, 1979
62. Voltarelli FA, Gobatto CA, Mello MAR: Determination ofanaerobic threshold in rats using the lactate minimum test.Braz J Med Biol Res 35:1389-1394, 2002
63. Willow M, Carmody J, Carroll P: The effects of swimmingin mice on pain perception and sleeping time in response tohypnotic drugs. Life Sci 26:219-224, 1980
64. Wright A, Sluka KA: Nonpharmacological treatments formusculoskeletal pain. Clin J Pain 17:33-46, 2001
65. Yoshimasa T, Nakao K, Ohtsuki H, Li S, Imura H: Methio-nine-enkephalin and leucine-enkephalin in human sympa-thoadrenal system and pheochromocytoma. J Clin Invest69:643-650, 1982
66. Zhu CZ, Wilson SG, Mikusa JP, Wismer CT, Gauvin DM,Lynch JJ 3rd, Wade CL, Decker MW, Honore P: Assessingthe role of metabotropic glutamate receptor 5 in multiplenociceptive modalities. Eur J Pharmacol 506:107-118, 2004
67. Zimmermann M: Ethical guidelines for investigations ofexperimental pain in conscious animals. Pain 16:109-110, 1983




















