
Effect of carbon and nitrogen sources on growth and solasodine production in batch suspension...
6
Plant Cell, Tissueand Organ Culture 21: 55-60, 1990. © 1990 KluwerAcademicPublishers.Printedin the Netherlands. Effect of carbon and nitrogen sources on growth and solasodine production in batch suspension cultures of Solanum eleagnifolium Cav. H.M. Nigra, M.A. Alvarez & A.M. Giulietti* Biotecnologia, Facultad de Farmacia y Bioqulmica, Universidad de Buenos Aires, (1113) Junln 956, Buenos Aires, Rep~blica Argentina (*requests for offprints) Received 16 May 1989;acceptedin revised form 15 September 1989 Key words: solasodine, Solanum eleagnifolium Cav., batch suspension culture, carbon source, nitrogen source Abstract The effect of inoculum size, carbon sources (fructose, glucose, maltose, sucrose), nitrate and ammonia on solasodine production by Solanum eleagnifolium Cav. was studied. The specific growth rate was estimated to be 0.15-0.20 d -l with all sugars tested at a concentration of 90 mM. Sucrose (180 mM) produced the highest biomass value (about 2.8 mgDWml 1) while the lowest one was produced by maltose. Although solasodine productivity values after 11 days of culture were similar for all sugars tested, the maximum values of productivity (0.9 mg g-ld-~) were achieved after 6 days of culture with sucrose (180 mM). Solasodine productivity of cultures conducted with a large inoculum (20% w/v fresh material) was double that with a small inoculum (10% w/v fresh material). Introduction Solasodine, the nitrogen analogue of diosgenin, can be used as starting raw material for steroidal drug production [11]. Solasodine was obtained by chemical or microbial hydrolysis of solamargine [19, 20]. Although several species of the genus So- lanum contain solasodine, only a few of them are considered to be important in the pharmaceutical industry [11]. In fact, the use ofS. laciniatum and S. khasianum [11] for commercial production of ste- roidal drugs has been reported. Guerreiro et al. [5] and Kavka et al. [7] reported the presence of glycoalkaloids in unripe fruit of S. eleagnifolium Cav. var. leprosum, a native species from semi-arid regions of Argentina. In addition, Rodriguez et al. [20] reported the synthesis of 16- dehydropregnenolone acetate (16-DHPA) from solasodine obtained from S. eleagnifolium Cav. 16- DHPA is the key compound for the pharmaceutical industry for production of steroidal compounds. We have previously reported the production of solasodine by calli from different explants of seed- lings of S. eleagnifolium Cav. [14] and we have examined the effect of auxins, light and or- ganogenesis on its production [15, 16] but the max- imum yields obtained still remain low compared with those found in fruit of the wild plant. In order to improve the secondary metabolite yield from plant cell culture, several strategies can be carried out, such as selection of environmental conditions, system of culture, and cell lines of high productiv- ity. In the recent years, considerable effort has been directed towards the modifying of the culture media which would optimize the production of secondary metabolites [10]. The medium com- ponents which generally affect the secondary metabolite yields are the concentration of sugar, nitrogen source and phosphate, as well as the growth regulators. In this paper, the effect of carbon sources, relative concentrations of nitrate and ammonia and ino- culum size on solasodine production in batch sus- pension cultures of S. eleagnifolium Cav. is studied.
Transcript of Effect of carbon and nitrogen sources on growth and solasodine production in batch suspension...

Plant Cell, Tissue and Organ Culture 21: 55-60, 1990. © 1990 Kluwer Academic Publishers. Printed in the Netherlands.
Effect of carbon and nitrogen sources on growth and solasodine production in batch suspension cultures of Solanum eleagnifolium Cav.
H.M. Nigra, M.A. Alvarez & A.M. Giulietti* Biotecnologia, Facultad de Farmacia y Bioqulmica, Universidad de Buenos Aires, (1113) Junln 956, Buenos Aires, Rep~blica Argentina (*requests for offprints)
Received 16 May 1989; accepted in revised form 15 September 1989
Key words: solasodine, Solanum eleagnifolium Cav., batch suspension culture, carbon source, nitrogen source
Abstract
The effect of inoculum size, carbon sources (fructose, glucose, maltose, sucrose), nitrate and ammonia on solasodine production by Solanum eleagnifolium Cav. was studied. The specific growth rate was estimated to be 0.15-0.20 d -l with all sugars tested at a concentration of 90 mM. Sucrose (180 mM) produced the highest biomass value (about 2.8 mgDWml 1) while the lowest one was produced by maltose. Although solasodine productivity values after 11 days of culture were similar for all sugars tested, the maximum values of productivity (0.9 mg g-ld-~) were achieved after 6 days of culture with sucrose (180 mM). Solasodine productivity of cultures conducted with a large inoculum (20% w/v fresh material) was double that with a small inoculum (10% w/v fresh material).
Introduction
Solasodine, the nitrogen analogue of diosgenin, can be used as starting raw material for steroidal drug production [11]. Solasodine was obtained by chemical or microbial hydrolysis of solamargine [19, 20]. Although several species of the genus So- lanum contain solasodine, only a few of them are considered to be important in the pharmaceutical industry [11]. In fact, the use ofS. laciniatum and S. khasianum [11] for commercial production of ste- roidal drugs has been reported.
Guerreiro et al. [5] and Kavka et al. [7] reported the presence of glycoalkaloids in unripe fruit of S. eleagnifolium Cav. var. leprosum, a native species from semi-arid regions of Argentina. In addition, Rodriguez et al. [20] reported the synthesis of 16- dehydropregnenolone acetate (16-DHPA) from solasodine obtained from S. eleagnifolium Cav. 16- DHPA is the key compound for the pharmaceutical industry for production of steroidal compounds.
We have previously reported the production of
solasodine by calli from different explants of seed- lings of S. eleagnifolium Cav. [14] and we have examined the effect of auxins, light and or- ganogenesis on its production [15, 16] but the max- imum yields obtained still remain low compared with those found in fruit of the wild plant. In order to improve the secondary metabolite yield from plant cell culture, several strategies can be carried out, such as selection of environmental conditions, system of culture, and cell lines of high productiv- ity. In the recent years, considerable effort has been directed towards the modifying of the culture media which would optimize the production of secondary metabolites [10]. The medium com- ponents which generally affect the secondary metabolite yields are the concentration of sugar, nitrogen source and phosphate, as well as the growth regulators.
In this paper, the effect of carbon sources, relative concentrations of nitrate and ammonia and ino- culum size on solasodine production in batch sus- pension cultures of S. eleagnifolium Cav. is studied.

56
Materials and methods
Callus cultures were established from hypocotyls of seedlings cultured on a modified mineral nutrient medium [13] with the addition of myo-inositol (100mgl-~), sucrose (30gl ~), RT vitamin com- plex (20 ml 1- ~) [8] and 2.4-dichlorophenoxyacetic acid (2,4-D) (4.5 #M) as growth regulator. The pH was adjusted to 5.6-5.8 and the medium solidified with 8 g 1- l of agar. This medium will be referred to as MSRT medium. The incubation conditions were 25°C _+ 2°C under a 16h photoperiod, using fluorescent lamps at a light intensity of approxi- mately 1.8 W m -2.
For initiation of cell suspensions, pieces of fri- able callus (approximately 20 g fresh weight) were transferred to liquid MSRT medium (40ml in 200 ml Erlenmeyer flasks). The flasks were main- tained on a rotary shaker (100 rpm) under the same culture conditions described above.
The suspensions were subcultured every 24 days and transferred to 40ml of fresh MSRT liquid medium.
For studying the influence of inoculum size on growth and solasodine production, two series of Erlenmeyer flasks containing MSRT medium plus sucrose (90mM) were inoculated with fresh ma- terial at the rate of 10% and 20% (w/v).
In order to study the effect of the carbohydrate source and its concentration on growth and sol- asodine production, the following sugars were test- ed: glucose, sucrose, fructose and maltose at con- centrations of 90 and 180 mM. The inoculum size was 20% (w/v) of fresh weight of material.
The effect of the relative concentrations of ni- trate and ammonia on solasodine production was tested using six different culture media. The com- position of the media tested was similar to MSRT medium, but the nitrogen sources and NO3 :NH + ratio were modified (see Table 1). The nitrogen sources used were for medium I: KNO3; medium II: NH4C1 and (NHD2SO4; media III, V and VI: (NH4)2SO4, NHaC1 and KNO3; and medium IV: KNO3 and NH4NO3. All media used had the same total nitrogen content as the MSRT medium (30 raM) [13].
All experiments were carried out in 200 ml Erlen- meyer flasks containing 40 ml of the corresponding medium. They were located on a shaker operated at 100rpm and incubated under the culture con-
ditions already described. The estimations of growth and solasodine con-
centrations were carried out as described in a previous paper [14]. The determinations of sucrose, maltose, glucose and fructose were performed ac- cording to Percheron [18].
Results and discussion
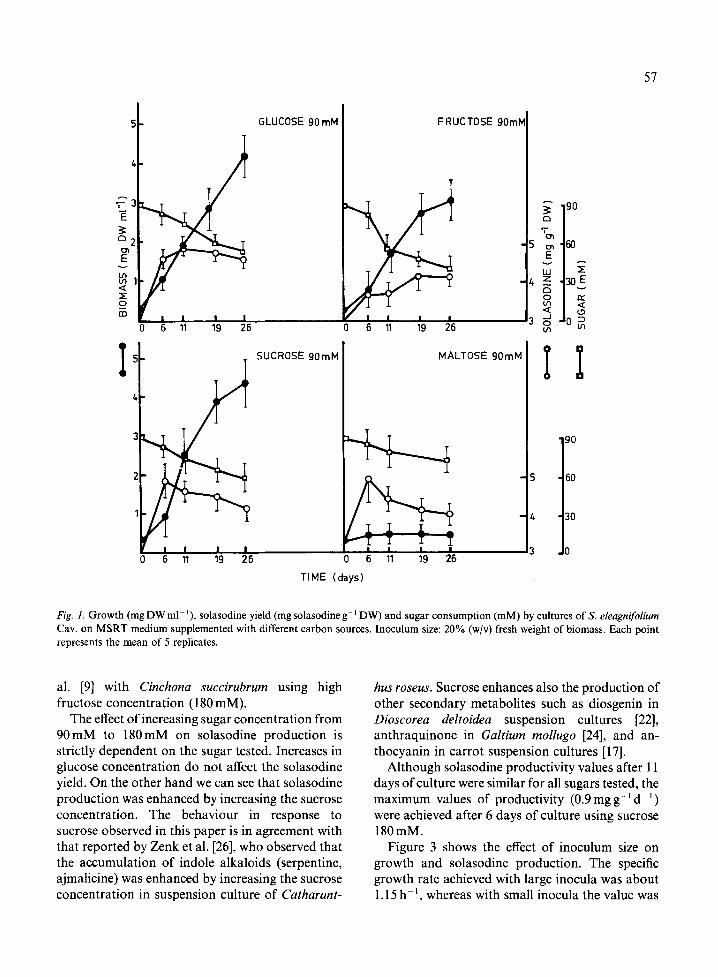
Figure 1 shows the growth and solasodine produc- tion pattern following the use of different car- bohydrates as carbon source. In all cases the specific growth rate (/~) estimated was 0.15-0.20 d 1. These values are within the range obtained for different spe- cies reported by other authors: 0.144 d- 1 for Silybum marianum [3], 0.314d 1 for Nicotiana tabacum [21], and 0.122 d i for Cynara cardunculus [4].
The doubling time (td) of S. eleagnifolium Cav. cell suspension using sucrose at 90 mM concentra- tion (t d = 3.46 d), is lower than the value obtained for callus culture (td = 8.30d) cultivated in the same medium [14]. This is probably due to facilitat- ed nutrient transport in liquid medium. The maximum value of solasodine was obtained on the 6th day of culture for sucrose and maltose, and on the l l th day for glucose. After achieving the maximum value, in all cases, the solasodine con- centration decreased, probably due to the initiation of a degradation process [14].
The consumption rate of all sugars was similar for the first six days of culture. Over this period, 10-12% of the sugar was consumed, but after 24 days of culture, the consumption was as follows: glucose (38%), fructose (54%), sucrose (27%) and maltose (20%).
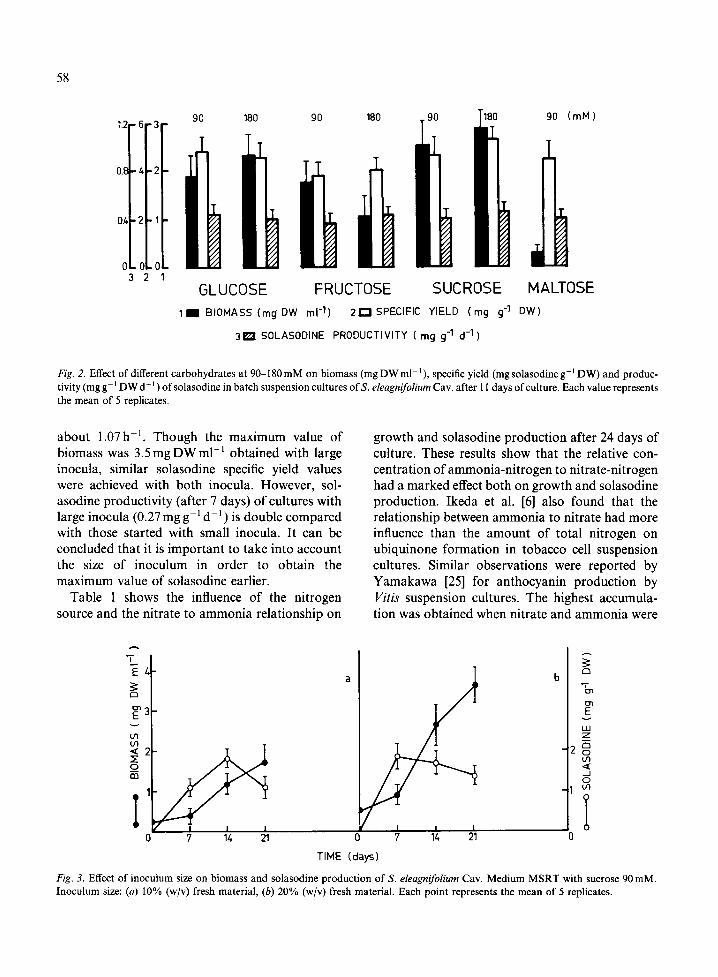
Figure 2 shows the solasodine yield, solasodine productivity and biomass value obtained after I 1 days of culture using MSRT medium with different carbon sources. Sucrose (180mM) produced the highest biomass value while the lowest one corre- sponded to maltose. A possible strategy for increas- ing the biomass production is to carry out a fed batch culture with maltose. For glucose and sucrose the biomass value increases when the sugar concentration was increased from 90mM to 180 mM; fructose appears to have a growth-inhibit- ory effect at the higher concentration. Similar ob- servations were reported by Mantell & Smith [12] working with Nicotiana tabacum and by Khoury et
I I 51 GLUCOSE 90 mM F RUC TOSE 90mM I
I 4
T- 3 ~ 90
2 ~ 60
0 6 11 19 26 Ln u~ Ii I LTOSE 90ram !!I SUCROSE 90rnM I
3
4
! 0 6 11 19 25 6 11 19 26
TIME (days)
57
Fig. 1. Growth (mg DW ml ' ), solasodine yield (mg solasodine g- 1DW) and sugar consumption (mM) by cultures of S. eleagnifolium Cav. on MSRT medium supplemented with different carbon sources. Inoculum size: 20% (w/v) fresh weight of biomass. Each point represents the mean of 5 replicates.
al. [9] with Cinchona succirubrum using high fructose concentration (180mM).
The effect of increasing sugar concentration from 90mM to 180mM on solasodine production is strictly dependent on the sugar tested. Increases in glucose concentration do not affect the solasodine yield. On the other hand we can see that solasodine production was enhanced by increasing the sucrose concentration. The behaviour in response to sucrose observed in this paper is in agreement with that reported by Zenk et al. [26], who observed that the accumulation of indole alkaloids (serpentine, ajmalicine) was enhanced by increasing the sucrose concentration in suspension culture of Catharant-
hus roseus. Sucrose enhances also the production of other secondary metabolites such as diosgenin in Dioscorea deltoidea suspension cultures [22], anthraquinone in Galtium mollugo [24], and an- thocyanin in carrot suspension cultures [17].
Although solasodine productivity values after 11 days of culture were similar for all sugars tested, the maximum values of productivity (0.9 mg g-~ d-J ) were achieved after 6 days of culture using sucrose 180mM.
Figure 3 shows the effect of inoculum size on growth and solasodine production. The specific growth rate achieved with large inocula was about 1.15 h ], whereas with small inocula the value was
A q-
tm
E v
U3 U3
:E o ¢n
1.,~ - 6 r 3 r
0.~ - 41- 21,-
0.~ -21- 1 I-
0 - Ok OL 3 2 1
90 180 90 180 9 (mM)
I
GLUCOSE FRUCTOSE SUCROSE MALTOSE 111 BiOMASS (mg DW m1-1) 2[:::1 SPECIFIC YIELD (rag g-1 DW)
3 0 SOLASODINE PRODUCTIVITY (rag g-1 d - l )
Fig. 2. Effect of different carbohydrates at 90-180 mM on biomass (mg DW ml- t ), specific yield (mg solasodine g - t DW) and produc- tivity (mg g - t DW d - ~ ) of solasodine in batch suspension cultures of S. eleagnifolium Cav. after 11 days of culture. Each value represents the mean of 5 replicates.
about 1.07h 1. Though the maximum value of biomass was 3.5 mg DW ml-' obtained with large inocula, similar solasodine specific yield values were achieved with both inocula. However, sol- asodine productivity (after 7 days) of cultures with large inocula (0.27 mg g- l d-1 ) is double compared with those started with small inocula. It can be concluded that it is important to take into account the size of inoculum in order to obtain the maximum value of solasodine earlier.
Table 1 shows the influence of the nitrogen source and the nitrate to ammonia relationship on
growth and solasodine production after 24 days of culture. These results show that the relative con- centration of ammonia-nitrogen to nitrate-nitrogen had a marked effect both on growth and solasodine production. Ikeda et al. [6] also found that the relationship between ammonia to nitrate had more influence than the amount of total nitrogen on ubiquinone formation in tobacco cell suspension cultures. Similar observations were reported by Yamakawa [25] for anthocyanin production by Vitis suspension cultures. The highest accumula- tion was obtained when nitrate and ammonia were
I 0 0 7 14 21
I I I 7 14 21
58
TIME (days)
c'~ b
E tJJ Z
O
< ..A O t/3
Fig. 3. Effect of inoculum size on biomass and solasodine production of S. eleagnifolium Cav. Medium MSRT with sucrose 90 mM. Inoculum size: (a) 10% (w/v) fresh material, (b) 20% (w/v) fresh material. Each point represents the mean of 5 replicates.

Table 1. Influence of the nitrate:ammonia ratio on biomass and solasodine specific yield after 24 days of culture. The inoculum size was 20% (w/v).
Medium a NO[ :NH~- Biomass Solasodine DW (mgg IDW) (mgml -I)
I 1:0 4.69 _+ 0.12 1.03 _+ 0.03 II 0:1 0.21 _+ 0.03 0.41 _ 0.01 III 1:1 4.89 _+ 0.25 2.50 _+ 0.1 IV 2:1 6.88 _+ 0.35 4.08 _+ 0.25 V 1:2 5.56 + 0.30 3.00 _ 0.18 VI 2:1 6.54 _+_ 0.40 3.50 _+ 0.20
a For details of nitrogen-containing compounds see text
used together (media III, IV, V, VI). When nitrate and ammonia were supplied separately (media I and II), both growth and solasodine production decreased. However, Nicotiana tabacum cultures produced more ubiquinone when ammonia was the sole nitrogen source [6]. On the other hand, for diosgenin production, the use of both nitrogen sources together are essential [23].
As was to be expected, the relationship between nitrate and ammonia used in media composition has a marked effect on growth and solasodine pro- duction. A nitrate:ammonia ratio of 2:1 (media IV, VI) produced the highest biomass, while solasodine values were highest in medium IV, where the nitrogen sources were NHgNO 3 and KNO 3. These results suggest that NH4NO3 was a better nitrogen source than NH4C1 and (NH4)2SO 4 (medium VI).
In general the effect of nitrogen on production of secondary metabolites should be viewed as a delicate interaction of different nitrogen sources with the carbon source available to the cell, where even a small alteration of the C:N ratio may have a marked influence on growth and solasodine pro- duction.
The yields of solasodine obtained in our cultures are higher than those reported for S. dulcamara [2] (0.1 mggDW) and S. nigrum [1] (1.31 mggDW) in tissue cultures but are lower than those reported in fruits of plants of S. eleagnifolium Cav. [15].
Acknowledgements
This work was supported by the Secretaria de Cien- cia y Tecnologia (SECyT), the Consejo Nacional de Investigaciones Cientificas y T6cnicas (CONICET)
59
and Laboratorio Cientifico Paul Hnos, Castelar (Argentina).
References
1. Bhatt PN, Bhatt DP, Sussex I (1983) Studies on some factors affecting solasodine contents in tissue cultures of Solanum nigrum. Physiol Plant 57:159-162
2. Emke A, Eilert U (1986) Steroidal alkaloids in tissue cul- tures and regenerated plants of Solanum dulcamara. Plant Cell Rep 5:31-34
3. Fevereiro P, Cabral JMS, Fonseca MNR, Novais JM, Pais MSM (1986) Callus and suspension culture of Silybum marianum: biosynthesis of proteins with clotting activity. Biotechnol Lett 8:' 19-24
4. Figuereido AC, Fevereiro P, Cabral JMS, Novais JM, Pais MSM (1987) Callus and suspension cultures for biomass production of Cynara cardunculus (Compositae). Biotech- nol Lett 9:213-218
5. Guerreiro E, Giordano OS, Kavka J, D'Arcangelo AT (1971) Aislamiento de compuestos esteroidales de Solanum eleagnifolium Cav. Anales Quimica 67:789-794
6. Ikeda T, Matsumoto T, Noguchi M (1977) Studies on the culture conditions of higher plant cells in suspension cul- ture. Part IX. Effects of inorganic nitrogen source and physical factors on the formation of ubiquinone by tobac- co plant cells in suspension culture. Agric Biol Chem 41: 1197-1201
7. Kavka J, Guerreiro E, Gianello JC, Giordano OS, D'Arcangelo AT (1973) Glicoalcaloides esteroidales en cuatro especies del g6nero Solanum (Solanaceae). Anales Real Sociedad Espaflola Fisica Quimica 69:929-932
8. Khana P, Staba J (1968) Antimicrobials from plant tissue culture. Lloydia 3h 180-189
9. Khouri HE, Ibrahim R, Rideau M (1986) Effects of nutri- tional and hormonal factors on growth and production of anthraquinone glucosides in cell suspension cultures of Cinchona succirubia. Plant Cell Rep 5:423-426
10. Kurz WGW, Constabel F (1985) Aspects affecting biosyn- thesis and biotransformation of secondary metabolites in plant cell culture. CRC Critical Reviews Biotechnology 2: 105-118
11. Mann JB (1978) Production of solasodine for the pharma- ceutical industry. Adv Agronomy 30:207-245
12. Mantell AH, Smith H (1983) Cultural factors that influ- ence secondary metabolism accumulation in plant cell and tissue cultures. In: Mantell A, Smith H (Eds) Plant Biotechnology (pp75-110) Cambridge University Press, Cambridge
13. Murashige T, Skoog F (1962) A revised medium for rapid growth and bioassays with tobacco tissue culture. Physiol Plant 15:473-479
14. Nigra HM, Caso O, Giulietti AM (1987) Production of solasodine by calli from different parts of Solanum eleag- nifolium Cav. plants. Plant Cell Rep 6:135-137
15. Nigra HM, Caso O, Giulietti AM 0985) Producci6n de solasodina en callos de Solanum eleagnifolium Cav. Re- sfimenes XVI Reuni6n Nacional de Fisiologia Vegetal, La Plata, Argentina (p 79)

60
16. Nigra HM, Alvarez MA, Giulietti AM (I 987) Influencia de la fuente de carbono y del tamafio del inoculo en la produc- ci6n de solasodina en suspensiones celulares de Solanum eleagnifolium. Libro de Resfimenes, I Congreso Latino- americano de Biotecnologia, Tucum~n, Argentina, D-2-2
17. Ozeki Y, Komamine A (1985) Changes in activities of enzymes involved in general phenylpropanosoid metabol- ism during the induction and reduction of anthocyanin synthesis in a carrot suspension culture as regulated by 2,4-D. Plant Cell Tissue Organ Culture 5:45-53
18. Percheron F (1962) Colorimetric determination of fructose and fuctufuranosides by the thiobarbituric acid reaction. Comptes Rendus 225:2521-2522
19. Rodriguez J (1984) Producci6n de solasodina por hidrfli- sis microbiana de los glicoalcaloides de Solanum eleag- nifolium Cav. Tesis Doctoral, Universidad Nacional de San Luis, Argentina
20. Rodriguez J, Segovia R, Guerreiro E, Sosa GZ de, Ferretti F, Ertola R (1979) Production of solasodine by microbial hydrolysis of glycoalkaloids of Solanum eleagnifolium Cav. and its application onto the synthesis of 16-dehydropreg- nelone acetate. J Chem Technol Biotechnol 29:525-530
21. Sahai OP, Shuler ML (1984) Environmental parameters influencing phenolic production by batch cultures of Nico- tiana tabacum. Biotechnol Bioeng 26:111-120
22. Tal B, Goldberg I (1982) Growth and diosgenin produc- tion by Deltoidea cells in batch and continuous cultures. Planta Medica 44:107-110
23. Tal B, Gressel J, Goldberg I (1982) The effects of medium constituents on growth and diosgenin production by Dios- corea deltoidea cells grown in batch cultures. Planta Medi- ca 44:111-113
24. Wilson G, Galagu6 C (1985) Biosynthesis of anthraqui- none by cells of Galium mollugo L. grown in a chemostat with limiting sucrose or phosphate. J Exp Bot 36:485-493
25. Yamakawa T, Kato S, Ishida K, Kodama T, Minoda Y (1983) Production ofanthocyanins by Vitis cells in suspen- sion culture. Agric Biol Chem 47:2185-2191
26. Zenk MH, El-Shagi H, Arens H, Stockigt J, Weiler EW, Deus B (1977) Formation of the indole alkaloids serpen- tine and ajmalicine in cell suspension cultures of Cathar- anthus roseus. In: Barz W, Reinhard E, Zenk MH (Eds) Plant Tissue Culture and its Biotechnological Applications (pp 27-43), Springer-Verlag, Berlin




















