
Effect of a cage divider permitting social stimuli on stress and food intake in rats
7
Effect of a cage divider permitting social stimuli on stress and food intake in rats M.M. Boggiano a, ⁎, S.A. Cavigelli b , J.R. Dorsey a , C.E.P. Kelley a , C.M. Ragan b , P.C. Chandler-Laney a a Department of Psychology, Behavioral Neuroscience Division, University of Alabama at Birmingham, Birmingham, Alabama, 35294, USA b Department of Biobehavioral Health and Integrative Biological Sciences Program, The Pennsylvania State University, State College, PA, 16802, USA ABSTRACT ARTICLE INFO Article history: Received 18 February 2008 Received in revised form 29 April 2008 Accepted 29 April 2008 Keywords: Corticosterone Feces Adrenal Stress hormones HPA Ingestive behavior Feeding Body weight RIA HPLC Glucocorticoids Catecholamines Housing Rodent Enrichment Animal welfare Husbandry The need to obtain data from individual laboratory animals has forced many researchers to singly-house small animals. This is costly to the researcher and isolation can adversely affect animal physiology and behavior which in turn may threaten the validity and generalization of experiment results to humans. We assessed the practical use of a housing device – dubbed “Buddy Barrier” (BB) – that allows social stimulation in a paired- housing situation while at the same time permitting the collection of individual measures that traditionally require individual-housing. To assess stress responses to the BB, adult male rats were single or pair-housed for several days with and without a BB in the cage. Fecal corticosterone metabolites (fCORT), food intake and body weight were monitored daily. Plasma CORT and adrenal catecholamine levels were assessed at the end of the housing manipulation. Stress hormone measures did not differ in paired vs. singly-housed rats and paired rats quickly habituated to introduction and removal of the BB. Barring a trend for paired rats to eat more in the first 4 h of the dark, there was no difference in 24 h intakes or body weight gain between singly and paired-housed rats. While the BB attenuated 24 h intakes in both groups, intakes normalized to non-BB conditions by the third BB reintroduction. A device such as the BB can enhance the welfare of animals by providing social enrichment without compromising the integrity of experimental protocols traditionally requiring single-housing. In times of lagging research funding it can also substantially reduce housing costs. © 2008 Elsevier Inc. All rights reserved. Many experimental protocols that use rodents and other small animals require that the animals be isolated via single-housing for extended periods of time and, often, for the entire lab animals' career. Unfortunately, isolation is not a normal condition in rodents that are, by nature, social creatures. Also, numerous adverse effects of isolation on the behavior and health of laboratory animals have been reported [1–5]. In the lab, such untoward effects have the potential to com- promise the validity and reliability of experimental results. This is particularly important in behavioral rodent studies where it is ex- pected, or actually assumed, that measures obtained from control groups reflect measures obtained from as “normal” a condition or conditions as possible. Additionally, if experimental results are meant to be extrapolated to humans who are also social animals, it makes more sense to allow for social conditions among animals during experimentation. Congruent with the natural social nature of rodents, it is not surprising that rats prefer a location associated with the presence of another rat compared to a similar location paired without a rat [6]. The goal of this study was to assess the practical use of a rodent housing device that we have dubbed a “Buddy Barrier (BB)”, in protocols where individual measures are desired (i.e., where rodents are traditionally singly-housed). Two examples are protocols that call for the collection of fecal samples and of food intake measures from individual rats. Researchers have started to use fecal measures of corticosterone (fCORT) metabolites as a non-invasive method of assessing stress levels in animals [7–11] but rodents must be singly- housed to do this. The same is true of feeding studies where rodents must be isolated to obtain individual recordings. Therefore, we chose to collect fecal samples and record food intake from rats housed in pairs via the intermittent use of a BB in the cage. In this way, the rats did not have to be completely isolated for the obtainment of individual samples. At the same time, by measuring fCORT metabolites, we could determine to what extent, if any, the BB affected stress levels. We also subjected singly-housed rats to the BB to compare their response to that of paired rats. This allowed us to control for any effects due to the Physiology & Behavior 95 (2008) 222–228 ⁎ Corresponding author. Department of Psychology, Behavioral Neuroscience Divi- sion, 415 Campbell Hall, 1300 University Blvd., University of Alabama at Birmingham, Birmingham, AL 35294-1170, USA. Tel.: +1205 934 2439; fax: +1205 975 6110. E-mail address: [email protected] (M.M. Boggiano). 0031-9384/$ – see front matter © 2008 Elsevier Inc. All rights reserved. doi:10.1016/j.physbeh.2008.04.025 Contents lists available at ScienceDirect Physiology & Behavior journal homepage: www.elsevier.com/locate/phb
Transcript of Effect of a cage divider permitting social stimuli on stress and food intake in rats

Physiology & Behavior 95 (2008) 222–228
Contents lists available at ScienceDirect
Physiology & Behavior
j ourna l homepage: www.e lsev ie r.com/ locate /phb
Effect of a cage divider permitting social stimuli on stress and food intake in rats
M.M. Boggiano a,⁎, S.A. Cavigelli b, J.R. Dorsey a, C.E.P. Kelley a, C.M. Ragan b, P.C. Chandler-Laney a
a Department of Psychology, Behavioral Neuroscience Division, University of Alabama at Birmingham, Birmingham, Alabama, 35294, USAb Department of Biobehavioral Health and Integrative Biological Sciences Program, The Pennsylvania State University, State College, PA, 16802, USA
⁎ Corresponding author. Department of Psychology,sion, 415 Campbell Hall, 1300 University Blvd., UniversiBirmingham, AL 35294-1170, USA. Tel.: +1 205 934 2439
E-mail address: [email protected] (M.M. Boggiano)
0031-9384/$ – see front matter © 2008 Elsevier Inc. Aldoi:10.1016/j.physbeh.2008.04.025
A B S T R A C T
A R T I C L E I N F OArticle history:
The need to obtain data from Received 18 February 2008Received in revised form 29 April 2008Accepted 29 April 2008Keywords:CorticosteroneFecesAdrenalStress hormonesHPAIngestive behaviorFeedingBody weightRIAHPLCGlucocorticoidsCatecholaminesHousingRodentEnrichmentAnimal welfareHusbandry
individual laboratory animals has forced many researchers to singly-house smallanimals. This is costly to the researcher and isolation can adversely affect animal physiology and behaviorwhich in turn may threaten the validity and generalization of experiment results to humans. We assessed thepractical use of a housing device – dubbed “Buddy Barrier” (BB) – that allows social stimulation in a paired-housing situation while at the same time permitting the collection of individual measures that traditionallyrequire individual-housing. To assess stress responses to the BB, adult male rats were single or pair-housed forseveral days with andwithout a BB in the cage. Fecal corticosteronemetabolites (fCORT), food intake and bodyweight were monitored daily. Plasma CORT and adrenal catecholamine levels were assessed at the end of thehousing manipulation. Stress hormone measures did not differ in paired vs. singly-housed rats and paired ratsquickly habituated to introduction and removal of the BB. Barring a trend for paired rats to eat more in the first4 h of the dark, there was no difference in 24 h intakes or body weight gain between singly and paired-housedrats.While the BB attenuated 24 h intakes in both groups, intakes normalized to non-BB conditions by the thirdBB reintroduction. A device such as the BB can enhance the welfare of animals by providing social enrichmentwithout compromising the integrity of experimental protocols traditionally requiring single-housing. In timesof lagging research funding it can also substantially reduce housing costs.
© 2008 Elsevier Inc. All rights reserved.
Many experimental protocols that use rodents and other smallanimals require that the animals be isolated via single-housing forextended periods of time and, often, for the entire lab animals' career.Unfortunately, isolation is not a normal condition in rodents that are,by nature, social creatures. Also, numerous adverse effects of isolationon the behavior and health of laboratory animals have been reported[1–5]. In the lab, such untoward effects have the potential to com-promise the validity and reliability of experimental results. This isparticularly important in behavioral rodent studies where it is ex-pected, or actually assumed, that measures obtained from controlgroups reflect measures obtained from as “normal” a condition orconditions as possible. Additionally, if experimental results are meantto be extrapolated to humans who are also social animals, it makesmore sense to allow for social conditions among animals during
Behavioral Neuroscience Divi-ty of Alabama at Birmingham,; fax: +1 205 975 6110..
l rights reserved.
experimentation. Congruent with the natural social nature of rodents,it is not surprising that rats prefer a location associated with thepresence of another rat compared to a similar location paired withouta rat [6].
The goal of this study was to assess the practical use of a rodenthousing device that we have dubbed a “Buddy Barrier (BB)”, inprotocols where individual measures are desired (i.e., where rodentsare traditionally singly-housed). Two examples are protocols that callfor the collection of fecal samples and of food intake measures fromindividual rats. Researchers have started to use fecal measures ofcorticosterone (fCORT) metabolites as a non-invasive method ofassessing stress levels in animals [7–11] but rodents must be singly-housed to do this. The same is true of feeding studies where rodentsmust be isolated to obtain individual recordings. Therefore, we choseto collect fecal samples and record food intake from rats housed inpairs via the intermittent use of a BB in the cage. In this way, the ratsdid not have to be completely isolated for the obtainment of individualsamples. At the same time, by measuring fCORTmetabolites, we coulddetermine to what extent, if any, the BB affected stress levels. We alsosubjected singly-housed rats to the BB to compare their response tothat of paired rats. This allowed us to control for any effects due to the

223M.M. Boggiano et al. / Physiology & Behavior 95 (2008) 222–228
BB itself and/or the reduction in space produced by having a BB in thecage.We also compared fCORT levels, 4 and 24 h food intake, and bodyweight changes from these rats when the BB was in vs. out of the cageto evaluate habituation to this manipulation.
While there have been other studies examining the effect of singleand group housing on stress and food intake, none have monitoredthese parameters when paired rats are intermittently exposed to adevice that separates them but still allows them some socialstimulation. At the end of the study when all of the rats were keptwith a BB in the cage for several days, we obtained additionalmeasuresof stress including basal plasma CORT (pCORT), adrenal weights, andadrenal levels of catecholamines [11,12]. Ideally, it is our intent that theBB or similar device be used in experimental settings on anintermittent, not permanent basis. However, some protocols mightrequire data to be collected for several consecutive days so wesimulated those conditions here. In addition to improving the well-being of laboratory animals, the BB or similar device should benefit theresearcher by substantially reducing the proportion of funding thatmust be allocated to housing expenses. In essence, costs should be cutin half if per diem charges are based on number of cages utilized.
Fig.1. Panel A) “Buddy Barrier” housing device used to avoid single housing of rats whenindividual measures such as food intake and fecal specimens are needed. Here it wasalso used in rats in the Isolated group to control for space restriction and novelty of theBB when the Paired group was exposed to it. Panel B) The BB allows for visual, audio,olfactory, and limited tactile contact between rats while food and feces are keptseparated.
1. Methods
1.1. Subjects
Male Sprague Dawley rats (n=20; Harlan, IN) weighing 220–250 gat the onset of the study were housed in random pairs in clear259 mm×476 mm×209 mm polypropylene bedded cages with ad libwater and rat chow under a 12-h light/dark phase (lights off from1100–2300). After two weeks of acclimation to the animal colony, therats were weight-matched into one of two groups: an “Isolated”group or a “Paired” group. Those in the Isolated group were housedalone in a cage throughout the experiment. The Paired rats were nowhoused with a different cage mate from the first 2 weeks in the colonyin a cage of the same size as those housing the Isolated rats. EachPaired rat remained with its newly assigned cage mate for the entireduration of the study. The pairs were randomly assigned and one ofeach pair was identified from its cage mate by a black Sharpie® markon its tail.
1.2. Buddy Barriers (BBs) and BB placement procedures
The BBs are constructed from a perforated metal sheet running thelength of the plastic cages and are anchored to narrow rods at the baseof the plate (Fig. 1A). The perforated holes are large enough to allowrats to see, hear, and smell each other as well as to have limited tactilecontact (Fig. 1B). The plate reaches through the cage lid to separatefood placed on the lid indentation. A solid metal strip an inch from thebottom of the divider precluded transfer of feces or food from thebedding. The University of Alabama at Birmingham InstitutionalAnimal Care and Use Committee approved all materials for safety andsanitization in standard commercial cage washers. The reducedhousing space resulting from the temporary placement of the BBinto the cage met AAALAC guidelines [13]. The study took place over atwo-month period during which time the rats had a BB in the cage (BBcondition) for several days followed by a period of consecutive dayswithout a BB in the cage (NoBB condition). This was done for bothgroups, Paired and Isolated, as shown on the x-axis of Figs. 2–4 (onlydays when data were collected are shown and these are labeled “Days1–39” — Saturdays and Sundays were not included in the “Day”count). Placing a BB in the Isolated groups' cages served to control forany effect that decreased space, or the BB itself, might have imposedon the measures. The BBs were always inserted or removed during thehour prior to lights off (1000).
1.3. Fecal collection and CORT metabolite analyses
After a week following assignment of the rats into their groups,fecal collection began on Mondays–Fridays. Cage bedding waschanged daily 1 h prior to lights out so that feces could be collectedfrom fresh bedding over the first 4 h of the dark period (1100–1500 h).We chose to change bedding prior to lights off because rats begineating in the dark andwewanted to prevent eating prior to placementof the pre-measured food in the cage. However and ideally, if onlyfeces and not food intakes were being taken, bedding would havebeen changed in the dark to not disrupt the animals' sleep. Daily feceswere collected for 30 days into individually labeled Whirlpak bags(Nasco, Fort Atkinson, WI) and stored in −20 °C. When feces werecollected from Paired rats without the BB in the cage, the feces fromboth rats was stored in a single bag. Fecal samples were later dried,ground up, and mixed, then a sample of this mixture used for CORTmetabolite analyses. Because there is a 6–9 h lag between circulatingCORT and excretion of fecal CORT metabolites [7] the collected fecalsamples reflected circulating CORT levels secreted between 0200 and0900 h — the approximate time of the daily circulating CORT nadir.Feces were processed and CORT metabolites extracted as previouslydescribed [7] and stored at −80 °C until assayed. CORT metabolite
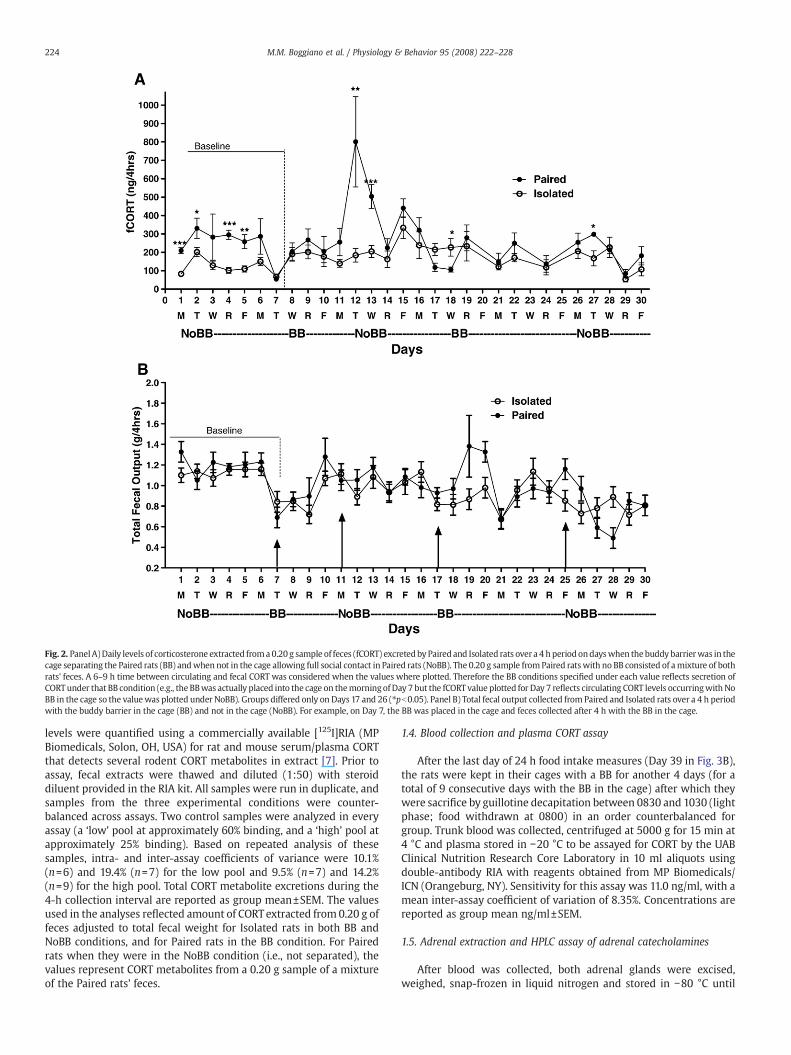
Fig. 2. PanelA)Daily levels of corticosterone extracted froma0.20 g sample of feces (fCORT) excretedby Paired and Isolated rats over a 4 hperiodondayswhen thebuddy barrierwas in thecage separating the Paired rats (BB) andwhen not in the cage allowing full social contact in Paired rats (NoBB). The 0.20 g sample fromPaired ratswith no BB consisted of amixture of bothrats' feces. A 6–9 h time between circulating and fecal CORT was considered when the values where plotted. Therefore the BB conditions specified under each value reflects secretion ofCORTunder that BB condition (e.g., the BBwas actually placed into the cage on themorningof Day 7 but the fCORT value plotted for Day 7 reflects circulating CORT levels occurringwith NoBB in the cage so the valuewas plotted under NoBB). Groups differed only on Days 17 and 26 (⁎pb0.05). Panel B) Total fecal output collected from Paired and Isolated rats over a 4 h periodwith the buddy barrier in the cage (BB) and not in the cage (NoBB). For example, on Day 7, the BB was placed in the cage and feces collected after 4 h with the BB in the cage.
224 M.M. Boggiano et al. / Physiology & Behavior 95 (2008) 222–228
levels were quantified using a commercially available [125I]RIA (MPBiomedicals, Solon, OH, USA) for rat and mouse serum/plasma CORTthat detects several rodent CORT metabolites in extract [7]. Prior toassay, fecal extracts were thawed and diluted (1:50) with steroiddiluent provided in the RIA kit. All samples were run in duplicate, andsamples from the three experimental conditions were counter-balanced across assays. Two control samples were analyzed in everyassay (a ‘low’ pool at approximately 60% binding, and a ‘high’ pool atapproximately 25% binding). Based on repeated analysis of thesesamples, intra- and inter-assay coefficients of variance were 10.1%(n=6) and 19.4% (n=7) for the low pool and 9.5% (n=7) and 14.2%(n=9) for the high pool. Total CORT metabolite excretions during the4-h collection interval are reported as group mean±SEM. The valuesused in the analyses reflected amount of CORTextracted from 0.20 g offeces adjusted to total fecal weight for Isolated rats in both BB andNoBB conditions, and for Paired rats in the BB condition. For Pairedrats when they were in the NoBB condition (i.e., not separated), thevalues represent CORT metabolites from a 0.20 g sample of a mixtureof the Paired rats' feces.
1.4. Blood collection and plasma CORT assay
After the last day of 24 h food intake measures (Day 39 in Fig. 3B),the rats were kept in their cages with a BB for another 4 days (for atotal of 9 consecutive days with the BB in the cage) after which theywere sacrifice by guillotine decapitation between 0830 and 1030 (lightphase; food withdrawn at 0800) in an order counterbalanced forgroup. Trunk blood was collected, centrifuged at 5000 g for 15 min at4 °C and plasma stored in −20 °C to be assayed for CORT by the UABClinical Nutrition Research Core Laboratory in 10 ml aliquots usingdouble-antibody RIA with reagents obtained from MP Biomedicals/ICN (Orangeburg, NY). Sensitivity for this assay was 11.0 ng/ml, with amean inter-assay coefficient of variation of 8.35%. Concentrations arereported as group mean ng/ml±SEM.
1.5. Adrenal extraction and HPLC assay of adrenal catecholamines
After blood was collected, both adrenal glands were excised,weighed, snap-frozen in liquid nitrogen and stored in −80 °C until

Fig. 3. Panel A) Daily food intake during the first 4 h in the dark of Isolated and Paired rats with and without a BB in the cage. There was no main effect of group across BB conditionsbut individual effect tests revealed increased intake in Paired vs. Isolated rats on some days (⁎pb0.05; ⁎⁎pb0.01, ⁎⁎⁎pb0.001). Panel B) Serial 24-hour chow intake of Isolated vs.Paired rats on days with the buddy barrier in the cage (BB) and without (NoBB). Days omitted (1, 6, 11, 16, 26, and 31) were Mondays, when no 24 h data was obtained (Sunday–Monday). NoBB data for Paired rats were derived from the total intake of both rats in the cage divided by 2.
225M.M. Boggiano et al. / Physiology & Behavior 95 (2008) 222–228
assayed using high-performance liquid chromatography (HPLC) withelectrochemical detection (EC). After thawing, adrenal glands werehomogenized in 0.2 N perchloric acid containing EDTA, sodiummetabisulfite, and dihidroxybenzoacetic acid as the internal standard,then centrifuged at 12,000 g for 20 min at 4 °C. Supernatants werethen diluted in perchloric acid solution and injected into the HPLCsystem (4-channel CoulArray EC detector; ESA, Inc., MA). A C-18reverse-phase column with a 4% methanol-based mobile phase at pH4.3 was used with electrode potentials set at −100 mV, 150, 250, and450 mV to separate norephinephrine, 3-methoxy-4-hydroxyphenyl-glycol (MHPG), epinephrine, and dopamine. Concentration valueswere corrected for dilution and adrenal weight and reported as groupmean ng/ml±SEM.
1.6. Food intake and body weight measures
On the same day that feces began to be collected, we obtained 4 hmeasures of food intake on Monday–Friday for 30 days and 24 hmeasures on the same Tuesday–Fridays for 39 days. Just prior tolights off, a pre-measured amount of Purina rat chow (Harlan TekladGlobal Diets, Indianapolis, IN) was placed on the top of the cage lid.On days when the BBs were in the cage, food was also placed on thecage lid on either side of the BB. Left over food and spillage (whichwas always minimal) was measured after a 4 h period and on thefollowing day for a 24 h recording. Ad lib water in two bottles percage was available at all times (when BB was in and out of the cage).
The data that were analyzed and graphed reflect group mean gramsof intake derived from individual Isolated rats' intakes when the BBwas in and out of the cage. For the Paired rats, the data reflectindividual rats' intake when the BB was in the cage. However, whenthe BB was not in the cage, individual intakes could not be ascer-tained because the rats were not separated. Therefore, the summedamount of intake for each pair was divided in half and these valueswere used to compare intake to the Isolated group. Values werereported as mean kcal±SEM. Body weights were recorded daily in themorning prior to presentation of pre-measured food for 30 days. Weobtained daily body weight changes by subtracting this number fromthe previous day's weight. Values were reported as group meangrams±SEM.
1.7. Statistics
Separate repeated-measures MANOVAs were conducted to deter-mine differences between Isolated and Paired group concentrations offCORT, of 4 and 24-h food intake, of body weight, and body weightfluctuations. BB condition (BB or NoBB) was entered as the withinsubjects factor. Due to the multiple comparisons made (across 30–39 days), the level for significance for each between-group effect wascorrected by dividing alpha of 0.05 by the number of comparisons(e.g., 30–39) to increase the percent confidence level for each analysis.To determine differences in pCORT, adrenal weights, and adrenalcatecholamines, ANOVAs were conducted using group (Isolated vs.
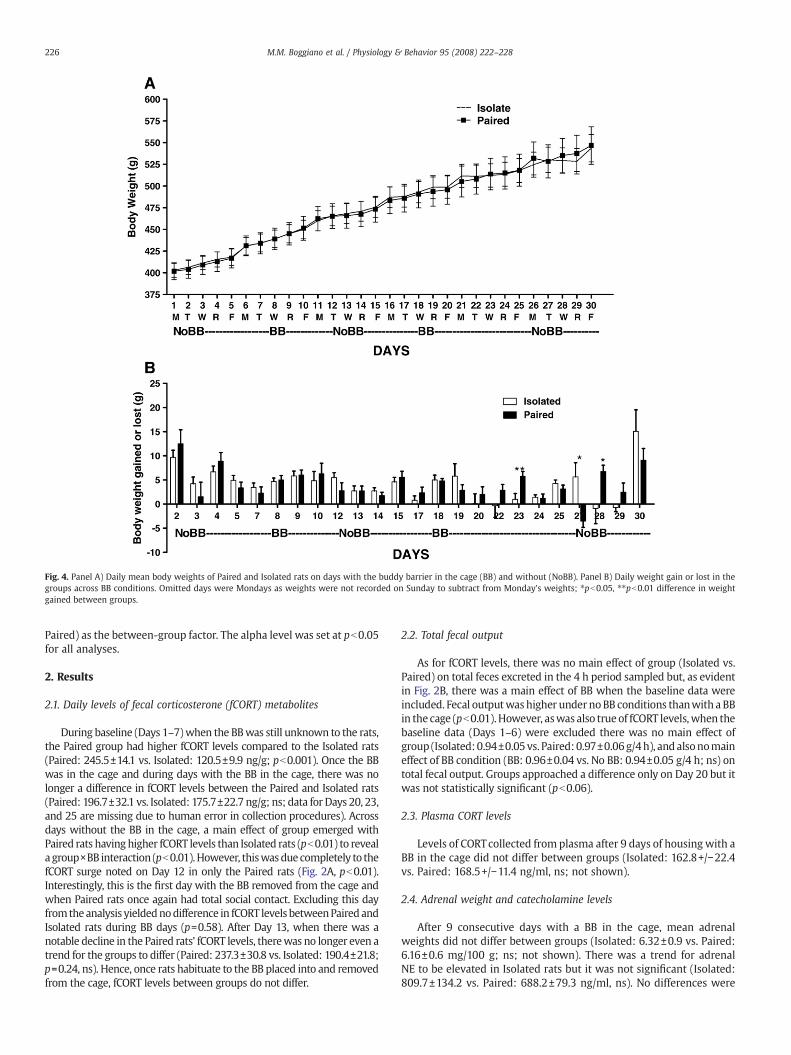
Fig. 4. Panel A) Daily mean body weights of Paired and Isolated rats on days with the buddy barrier in the cage (BB) and without (NoBB). Panel B) Daily weight gain or lost in thegroups across BB conditions. Omitted days were Mondays as weights were not recorded on Sunday to subtract from Monday's weights; ⁎pb0.05, ⁎⁎pb0.01 difference in weightgained between groups.
226 M.M. Boggiano et al. / Physiology & Behavior 95 (2008) 222–228
Paired) as the between-group factor. The alpha level was set at pb0.05for all analyses.
2. Results
2.1. Daily levels of fecal corticosterone (fCORT) metabolites
During baseline (Days 1–7)when the BBwas still unknown to the rats,the Paired group had higher fCORT levels compared to the Isolated rats(Paired: 245.5±14.1 vs. Isolated: 120.5±9.9 ng/g; pb0.001). Once the BBwas in the cage and during days with the BB in the cage, there was nolonger a difference in fCORT levels between the Paired and Isolated rats(Paired: 196.7±32.1 vs. Isolated: 175.7±22.7 ng/g; ns; data for Days 20, 23,and 25 are missing due to human error in collection procedures). Acrossdays without the BB in the cage, a main effect of group emerged withPaired rats having higher fCORT levels than Isolated rats (pb0.01) to reveala group×BB interaction (pb0.01).However, thiswasdue completely to thefCORT surge noted on Day 12 in only the Paired rats (Fig. 2A, pb0.01).Interestingly, this is the first day with the BB removed from the cage andwhen Paired rats once again had total social contact. Excluding this dayfromtheanalysis yieldednodifference in fCORT levels betweenPairedandIsolated rats during BB days (p=0.58). After Day 13, when there was anotable decline in the Paired rats' fCORT levels, therewas no longer even atrend for the groups to differ (Paired: 237.3±30.8 vs. Isolated: 190.4±21.8;p=0.24, ns). Hence, once rats habituate to the BB placed into and removedfrom the cage, fCORT levels between groups do not differ.
2.2. Total fecal output
As for fCORT levels, there was no main effect of group (Isolated vs.Paired) on total feces excreted in the 4 h period sampled but, as evidentin Fig. 2B, there was a main effect of BB when the baseline data wereincluded. Fecal outputwashigher undernoBB conditions thanwith a BBin the cage (pb0.01).However, aswas also trueof fCORT levels,when thebaseline data (Days 1–6) were excluded there was no main effect ofgroup (Isolated:0.94±0.05 vs. Paired: 0.97±0.06g/4h), and alsonomaineffect of BB condition (BB: 0.96±0.04 vs. No BB: 0.94±0.05 g/4 h; ns) ontotal fecal output. Groups approached a difference only on Day 20 but itwas not statistically significant (pb0.06).
2.3. Plasma CORT levels
Levels of CORTcollected from plasma after 9 days of housing with aBB in the cage did not differ between groups (Isolated: 162.8+/−22.4vs. Paired: 168.5+/−11.4 ng/ml, ns; not shown).
2.4. Adrenal weight and catecholamine levels
After 9 consecutive days with a BB in the cage, mean adrenalweights did not differ between groups (Isolated: 6.32±0.9 vs. Paired:6.16±0.6 mg/100 g; ns; not shown). There was a trend for adrenalNE to be elevated in Isolated rats but it was not significant (Isolated:809.7±134.2 vs. Paired: 688.2±79.3 ng/ml, ns). No differences were

227M.M. Boggiano et al. / Physiology & Behavior 95 (2008) 222–228
detected between Isolated and Paired rats in amount of adrenal con-tent of MHPG (Isolated: 4.6±0.2 vs. Paired: 4.4±0.3 ng/ml, ns), nor ofepinephrine (Isolated: 8260.3±1186 vs. Paired: 8356.8±1087 ng/ml,ns) or dopamine (Isolated: 28±2.5 vs. Paired: 22.5±2.5 ng/ml, ns; notshown).
2.5. 4 h food intake
When the baseline data (Days 1–6) were included, there was nomain effect of group or of BB condition. Without the baseline data inthe analyses, therewas still nomain effect of group (Isolated: 8.77±0.5vs. Paired: 10.33±0.7 kcal; ns) on 4 h food intake. Although notsignificant, inspection of Fig. 3A suggests a trend for Paired rats to eatmore than Isolated rats. The significant increases occurred as fre-quently on BB days than on NoBB days, explaining the lack of agroup×BB interaction.
2.6. 24-hour food intake
As evident in Fig. 3B, the trend to increase intake in the first 4 hobserved in the Paired rats on some days (Fig. 3A) disappeared by 24 hso that there was no group difference in intake or interaction with BBconditions. This was also true if baseline data was excluded from theanalysis (Isolated: 25.3±0.7 vs. Paired: 25.4±0.9 kcal; ns). Only on Day12, when fCORT levels where elevated in the Paired rats, did foodintake differ between groups; it was lower in Paired vs. Isolated rats(pb0.01; Fig. 3B). While there was no group difference, there was amain effect of BB condition such that 24 h intakes were lower in bothgroups on days when the BB was in the cage vs. when it was not in thecage (BB: 23.2±0.8 vs. NoBB: 27.5±0.6 kcal; pb0.001). This was truewhether baseline intake was included in the analysis or not. However,as evident by inspection of Fig. 3B, by the 3rd reintroduction of the BB(Day 34), 24 h intakes were the same as during NoBB conditions (BB:28.4±0.6 vs. NoBB: 28.05±0.5 kcal; ns). In fact by Day 27, 24 h foodintakes under both NoBB conditions (Days 27–33) and BB conditions(Days 34–39) were more uniform compared to fluctuations underboth conditions noted prior to this day (Fig. 3B).
2.7. Body weight
As shown in Fig. 4A, mean body weights of Isolated and Paired ratsdid not differ throughout the study (Isolated: 561.2±15.5 vs. Paired:558.1±16.9 g, ns, at time of sacrifice). Therewas also no effect of groupor BB condition on daily body weight fluctuations (Fig. 4B). Significantweight change differences between groups occurred only on Days 23,27, and 28 but there was no group or BB condition pattern to thesechanges.
3. Discussion
This study explored the practical use of a simple perforatedhousing unit that temporarily separates rats within a cage – dubbed“Buddy Barrier” – in an effort to avoid single-housing animals inresearch. It was used in a protocol that allowed us to measure stresshormone metabolites in feces and to record food intake fromindividual rats without isolating them. The study was driven by thetenet that “stressed animals do not make good research subjects”([14], pg.18). It was our wish to test a device that aimed at improvingthe welfare of lab animals that are, by nature, social animals.
We found that fCORT levels revolved around a slightly higheramount in the Paired rats once the BBs started being used, however,this elevationwas the result of one day of elevated fCORT levels on theday the BB was first removed from the pair-housed males. After thisfirst exposure to the BB, having the device in or out of the cage did notdifferentially affect fCORT levels. One might argue that fCORTmeasures are not as sensitive as plasma CORT measures. However,
this may only be the case in measuring short-lived changes inglucocorticoid production (e.g. 1–2 h changes) which are not readilyidentified in fecal samples that provide an integrated index ofcirculating steroid levels over many hours. When measuring longer-term changes in glucocorticoid production (e.g. several hours) there isample evidence that fCORT measures can identify stress-relatedelevations in glucocorticoid production [8,15,16]. The early differencein fCORT levels of the Paired and Isolated rats during the NoBBbaseline period may have resulted from decreased physical/socialactivity in the newly isolated males. Long-term voluntary exercise canelevate circulating glucocorticoid levels in rodents [17] and loss of asocial partner for the single-housed rats may have led to less physicalactivity and thus lowered glucocorticoid production for a short-period. Alternatively, elevated fCORT metabolite levels in the pair-housed rats may reflect increased activity or stress created byinteraction with an unfamiliar adult male during the baseline period.Importantly however, after the first exposure to the BB, fCORT levelsbetween Isolated and Paired rats become the same. No differences intotal fecal output were noted between the BB and NoBB conditions, foreither the pair- or single-housedmales, suggesting that metabolic rateis not highly affected by insertion of the BB.
In terms of the practical use of the BB in experiments, it may benecessary for rat pairs to grow accustomed to each other for at least1 week before introducing the BB or similar device. Housing maleswith a novel cagemate and immediately exposing them to the BBmayintroduce an elevated level of stress that could slow habituation to theBB manipulation. Other neurochemical indices of stress includingpCORT, and adrenal catecholamines did not differ between isolatedand paired rats housed with a BB in their cage for 9 consecutive days.Unfortunately the limited number of subjects precluded us frommeasuring these parameters in a group of rats without a BB in the cageprior to sacrifice. Nonetheless, we are primarily concerned with the“real life” application of the BB or similar device in research settings.Since samples such as feces or food intakewould be collected with theBB in the cage, we conclude that an extended period of time with a BBin the cage will neither increase nor decrease stress compared tosingle-housed animals. While our data revealed no effect of the BB orof single vs. paired housing on basal stress hormone levels during thecircadian nadir, it is important not to rule out the possibility thatisolation may predispose rodents to hyper-respond to an aversivestressor or conversely, that social contact protects them from con-sequences of isolation. This was not tested here but several studiessupport this possibility [18–24].
Our results with food intake revealed that during the first 4 h offeeding in the dark, when rats tend to consume their largermeal, therewas a non-significant trend for paired rats to consume more thanisolated rats regardless of whether the BB was in or out of the cage. By24 h, however, this trend disappeared so there was no difference inintake between groups. Regarding any effect of the BB itself, 4 hintakes were greater in both Isolated and Paired groups but this had tobe followed by a decrease in intake at later hours since 24 h intakeswhen the BB was in the cage was lower than on days without the BB.By the 3rd time that the BB was re-introduced, 24 h intakes becamequite stable across days and did not differ from intakes when the BBwas not in the cage. A conservative conclusion for the practical use of aBB or similar device in food intake studies using pair-housed rats isthat intakes should be expected to be somewhat lower than the firstfew times that the BB is in the cage. Hence, it is recommended that therats be habituated to having the BB in and out of the cage for at least 3times to habituate the animals and arrive at food intake levelscomparable to those seen without the BB present. Food intake shouldbemeasured on days when the BB is in the cage to establish a baseline.If body weight, and not food intake, is the main dependent variable ofinterest, there should be no concern that using the BB or similar devicewill affect these parameters. One caveat that should be considered,however, is that pairing rats may not be the same as group-housing

228 M.M. Boggiano et al. / Physiology & Behavior 95 (2008) 222–228
rats (3+per cage). Scalera et al. found that housing more than two ratstogether resulted in decreased food intake and body weight gaincompared to isolating or pairing rats [25].
Besides the measures taken here, the BB or similar device can beused to decrease physical contact between aggressive male rodents.Gradual habituation of two male rats to each other via the BB mayreduce other psychosocial aspects of aggression but this would needto be empirically determined. The BB can be used to allow stud malessome social contact when they must be isolated. In post-surgicalanimals the BB can protect the animals and implanted apparatus whileallowing some social stimuli. In transgenic mouse and other studiestypically requiring that a large number of animals to be housedindividually, BBs or similar devices could mean substantial grant-dollar savings. This is important given the flat growth rate of federalfunding for biomedical research [26,27]. There are some caveats thatshould be considered when extrapolating from our stress and feedingdata. For one, we did not test female rats and greater sensitivity tostress in female vs. male rats have been reported [21]. However, iffemales are more susceptible to isolation-induced stress, use of the BBor similar device may prove more beneficial as the rats would be ableto be kept in pairs. Secondly, our hormone and feeding results may notdirectly generalize to mice or certain mouse species as species-typicalsocial and stress responses have been observed [28]. Thirdly and inkeeping with the goal of improving animal welfare, although thereduced space left by placing a BB in the cage meets IACUC guidelines,we condone the intermittent, not long-term use of the BBs in the cage.We cannot rule adverse effects due to restricted space for a longerperiod of consecutive days than was conferred on the present rats.
In a “call to arms” for more sensitive treatment of laboratoryanimals, Reinhardt points out that many researchers ignore the statusof housing conditions despite the fact that stress evoked by suchconditions can jeopardize the value of their data [29]. We posit thatthere aremany of us who have great sensitivity for our animal subjectsbut are forced nonetheless to isolate them in order to obtain the datawe need. Use of the BB or similar cage devices is only one, but asignificant way of improving our subjects' welfare.
Acknowledgements
We thank Dr. Sam Cartner, Jim Crawford, and Shellie Hyde of theUAB Animal Resources Program for their assistance with thedevelopment of the Buddy Barriers and assuring that they adhere toIACUC regulations. We thank Harry Stubblefield and Jerry Sewell forthe construction and modifications of the BBs, respectively, andappreciate Dr. Bob Baker and Matt Giddings for their assistance withadrenal extractions. In addition, we thank Krishna Mehta, Cole Mateo,Janey Quinn, Collen Kovacsics, Jenna Smith, Alex Bruscke, RachelAnolik and Kerry Michael for assistance in fecal sample preparationand extraction. The study was supported by the following grants:P30DK056336 (UAB CNRC); DK066007 and UAB-Support for Devel-opment and Application of Research Using Animal Models Grant(Boggiano); and internal Pennsylvania State University Funding(Cavigelli).
References
[1] Van Loo PL, Van de Weerd HA, Van Zutphen LF, Baumans V. Preference for socialcontact versus environmental enrichment in male laboratory mice. Lab Anim2004;38:178–88.
[2] Gonder JC, Laber K. A renewed look at laboratory rodent housing and manage-ment. ILAR J 2007;48:29–36.
[3] Fox MW. Laboratory animal husbandry: ethology, welfare and experimentalvariables. Albany, N.Y.: State University of New York Press; 1986.
[4] Russell WMS. Shooting the clock: timeless lessons of the past still guide today'srefinement initiatives. Sci Animal Care WARDS Newsl 1997;8:1–2.
[5] Claasen V. Neglected factors in pharmacology and neuroscience research. Elsevier:Amsterdam, Netherlands; 1994.
[6] Crowder WF, Hutto CWJ. Operant place conditioning measures examined usingtwo nondrug reinforcers. Pharmacol Biochem Behav 1992;41:817–24.
[7] Cavigelli SA, Monfort SL, Whitney TK, Mechref YS, Novotny M, McClintock MK.Frequent serial fecal corticoid measures from rats reflect circadian and ovariancorticosterone rhythms. J Endocrin 2005;184:153–63.
[8] Cavigelli SA, Guhad FA, Ceballos RM, Whetzel CA, Nevalainen T, Lang CM, et al.Fecal corticoid metabolites in aged male and female rats after husbandry-relateddisturbances in the colony room. J Am Assoc Lab Anim Sci 2006;45:17–21.
[9] Good T, Khan MZ, Lynch JW. Biochemical and physiological validation of acorticosteroid radioimmunoassay for plasma and fecal samples in oldfield mice(Peromyscus polionotus). Physio Behav 2003;80:405–11.
[10] Wasser SK, Hunt KE, Brown JL, Cooper K, Crockett CM, Bechert U, et al. Ageneralized fecal glucocorticoid assay for use in a diverse array of nondomesticmammalian and avian species. Gen Comp Endocrinol 2000;120:260–75.
[11] Möstl E, Palme R. Hormones as indicators of stress. Domest Anim Endocrinol2002;23:67–74.
[12] Vollmer RR, Baruchin A, Kolibal-Pegher SS, Corey SP, Stricker EM, Kaplan BB.Selective activation of norepinephrine-and epinephrine-secreting chromaffin cellsin rat adrenal medulla. Am J Physiol 1992;263:R716–721.
[13] Institute of Laboratory Animal Resources Commission on Life Sciences;Council,N.R.. Guide for the care and use of laboratory animals. Washington, D.C.:National Academy Press; 1996
[14] American Medical Association. Use of animals in biomedical research – thechallenge and response – an American Medical Association White Paper. Chicago:Group on Science and Technology, AMA; 1992.
[15] Harper JM, Austad SN. Effect of capture and season on fecal glucocorticoid levels indeer mice (Peromyscus maniculatus) and red-backed voles (Clethrionomys gapperi).Gen Comp Endocrinol 2001;123:337–44.
[16] Royo F, Björk N, Carlsson HE, Mayo S, Hau J. Impact of chronic catheterization andautomated blood sampling (Accusampler) on serum corticosterone and fecalimmunoreactive corticosterone metabolites and immunoglobulin A in male rats.J Endocrinol 2004;180:145–53.
[17] Borer KT, Bestervelt LL, Mannheim M, Brosamer MB, Thompson M, Swamy U, et al.Stimulation by voluntary exercise of adrenal glucocorticoid secretion in maturefemale hamsters. Physio Behav 1992;51:713–8.
[18] Bartolomucci A, Palanza P, Sacerdote P, Ceresini G, Chirieleison A, Panerai AE, et al.Individual housing induces altered immuno-endocrine responses to psychologicalstress in male mice. Psychoneuroendocrin 2003;28:540–58.
[19] Stranahan AM, Khalil D, Gould E. Social isolation delays the positive effects ofrunning on adult neurogenesis. Nat Neurosc 2006;9:526–33.
[20] Nunes Mamede Rosa ML, Nobre MJ, Ribeiro Oliveira A, Brandao ML. Isolation-induced changes in ultrasonic vocalization, fear-potentiated startle and prepulseinhibition in rats. Neuropsychobio 2005;51:248–55.
[21] Westenbroek C, Snijders TA, den Boer JA, Gerrits M, Fokkema DS, Ter Horst GJ. Pair-housing of male and female rats during chronic stress exposure results in gender-specific behavioral responses. Horm Behav 2005;47:620–8.
[22] Gavrilovic L, Dronjak S. Activation of rat pituitary-adrenocortical and sympatho-adrenomedullary system in response to different stressors. Neuro Endocrin Lett2005;26:515–20.
[23] Andrade CS, Guimarães FS. Anxiolytic-like effect of group housing on stress-induced behavior in rats. Depress Anxiety 2003;18:149–52.
[24] Weiss IC, Pryce CR, Jongen-Rêlo AL, Nanz-Bahr NI, Feldon J. Effect of social isolationon stress-related behavioural and neuroendocrine state in the rat. Behav Brain Res2004;152:279–95.
[25] Scalera G. Taste preferences, body weight gain, food and fluid intake in singly orgroup-housed rats. Physiol Behav 1992;52:935–43.
[26] Loscalzo J. The NIH budget and the future of biomedical research. NEJM2006;354:1665–7.
[27] Dove A. Biomedical research faces flat budget for 2008. Nat Med 2007;13:228–9.[28] Parmigiani S, Palanza P, Rogers J, Ferrari PF. Selection, evolution of behavior and
animal models in behavioral neuroscience. Neurosci Biobehav Rev1999;23:957–69.
[29] Reinhardt V. Compassion for animals in the laboratory: impairment or refinementof research methodology? J Appl Anim Welf Sci 2003;6:123–30.




















