
Promoter DNA Hypermethylation and Gene Repression in Undifferentiated Arabidopsis Cells
Upload
independentCategory
view
1download
0
EXPERIMENTAL NEUROLOGY 131, 124-132 (19%)
Early Axonal Regeneration: Repression by Schwann Cells and a Protease?
MARIO TAPIA, NIBALDO C. INESTROSA, AND JAIME ALVAREZ
Departamento de Biologia Celular y Molecular. Facultad de Ciencias Bioldgicas. Pontificia Universidad Catdlica, Santiago, Chile
We have proposed that mature Schwann cells and an extracellular protease repress the sprouting response of azons. To test this hypothesis, we destroyed all cells by freezing a short span of the rat sciatic nerve or inhibited proteases with subperineurial injections of aprotinin, and a crush was made to induce the sprout- ing response. In unconditioned or in vehicle-injected nerves, 8x0~s began to elongate at a constant rate after a delay of about 1 day. The freezing of the nerve distal to the crush obliterated the delay, but the rate of elongation did not change. A similar pattern was ob- served when the nerve segment was conditioned with aprotinin for 2 days prior to the crush. These effects were abolished when a short untreated segment was left between crush and conditioned region of the nerve. The electron microscopy of the nerve and the immuno- localization of the growth-associated protein (GAP-43) were consistent with the enhanced regrowth observed in conditioned nerves. Our Endings support the notion that Schwann cells repress the onset of regeneration and that a local protease is involved. I 1995 Academic
press Inc.
INTRODUCTION
A number of structural and functional parameters of axons, e.g., microtubular content, diameter, fast trans- port, phosphorylation of proteins, are in a dynamic equilibrium and under the control of local cues ( 12, 20, 21, 30, 38, 40). The terminal arborization of axons is also in a dynamic equilibrium. In fact, partial denerva- tion of a sensory field (48) or a muscle (22, 47) induces the residual axons to extend collaterals which take over the vacant territory. Aguilar et al. (1) suggested that axons possess a natural ability to acquire territory which is normally balanced by competing neighboring nerves. Along their trajectory, intact axons also have the ability to extend sprouts, but in this case the stimulus is other than a vacant territory. Bustos et al. (15) reported that after local administration of actinomycin-D sciatic nerve axons sprouted while Schwann cells were de- stroyed. Alvarez et al. (3) observed that axons sprouted after local administration of aprotinin and the amyloid
precursor protein (APP) with the Kunitz insert, which are inhibitors of serine proteases that remain in the extracellular space (46, 23). The sprouting response of the stem axon is akin to a release behavior and the repression should be provided by the surrounding tis- sue, as proposed already for collateral sprouting.
Langley and Anderson (291 reported that severed axons do not regenerate along a normal nerve which accords with the repressive action of intact nerves. Sectioned axons grow readily in severed nerves, i.e., a peripheral nerve becomes permissive when its axons disappear. Distal to a nerve injury, Schwann cells prolif- erate (11) and eliminate their myelin, i.e., these cells acquire a “denervated phenotype.” Curiously, treat- ment of a nerve with protease inhibitors, in addition to axonal sprouting, induces a similar behavior in Schwann cells since they proliferate and eliminate their myelin (21. In short, protease inhibitors induce the major events that follow a nerve section despite the fact that axons remain uninterrupted. From this body of evidence, we have proposed that normal axons are endowed with a sprouting drive down-regulated by a mechanism that probably involves Schwann cells and extracellular prote- ases (2,3, 15).
After a crush, axons of the central stump begin to elongate with a latency ranging between 1 and 2 days (8, 33). It is attractive to think that this delay reflects the time taken by the ongoing repression to decline in the distal stump until axons can overcome it to start elongating.
We used the nerve regeneration paradigm to get an insight into the proposed repression. Regeneration may be considered a sprouting response to the interruption of axons. Our hypothesis foresees that the experimental derangement of the repressive mechanism, as by destruc- tion of Schwann cells or by inhibition of proteases, should enhance the axonal regeneration. To test these predictions, we froze or conditioned with aprotinin a short nerve segment, crushed the nerve, and assessed the elongation of axons.
METHODS
Freezing ofthe nerve. In 200-g Sprague-Dawley rats under pentobarbitone anesthesia (30 mg/kg), the sciatic
0014-4666/95 $6.00 Copyright G 1995 by Academic Press. Inc. All rights of reproduction in any form reserved.
124

REPRESSION OF EARLY REGENERATION 125
nerve was exposed close to the popliteal fossa. The peroneal branch, at its emergence from the sciatic trunk, was crushed several times with jeweler tweezers over a 0.3-mm span of nerve. (This produces consis- tently a degeneration of all axons, as controlled with
stimulation and electron microscopy of the nerve.) In the first group only a crush was made. In the second group, 7 mm of nerve adjacent to the crush were frozen with a fine silver wire cooled in liquid nitrogen. The freezing was controlled under the dissecting microscope: when the wire touched the nerve, ice cristals formed across the breadth of the nerve in a few seconds, the segment was then allowed to thaw and the cycle re- peated three or more times in less than 90 sec. The freezing was controlled so as to reach the crush. In the third group, an unfrozen span 3 mm long was left between the crush and the frozen segment (Figs. lA, lB, and 1D). A hair stitch was placed in the nearby tissue as a landmark for the crush. The distance from crush to spinal ganglia was estimated in no less than 45 mm. The wound was sutured and the animal was given the standard postoperative care.
Protease inhibition. The sciatic nerve was exposed as before. With a Hamilton syringe, subperineurial injections (2 l~,l of 150 mM NaCl, 0.9% benzyl alcohol, and 0.1% fluorescein with or without aprotinin (1.6 mg/ml, Sigma)) were made in the tibial nerve in a central direction. This infiltrated about 7 mm of the tibial branch ascending onto the sciatic trunk. Four schedules were used: (i) crush made immediately at the central terminus of the infiltrated region; (ii) several injections made at the same site in consecutive days prior to the crush; (iii) crush made 10 mm central to the infiltrated segment, and (iv) crush made in uninjected nerves (Figs. lA, lC, and 1E). As in the case of the peroneal nerve, the distance from crush to the sensory perikarya of the tibial nerve was estimated in no less than 45 mm. The rest of the surgery was as before.
Pinch test. At various times after the lesion, the rats were anesthetized lightly with pentobarbitone and the pinch test was performed, i.e., a reflex response of the abdominal muscles was evoked by pinching the regener-
A B C D E
proximal - distal
FIG. 1. Experimental conditions. Diagrams represent nerves. Proximal and distal to the lesion are indicated. Arrowhead indicates crush. Box, frozen segment. Spindle, infiltrated with one or more injections ofaprotinin or vehicle. The crush is adjacent to (B,C), or at a distance of tD.E), the conditioned segment.
ating nerve with jeweler tweezers in a distal-to-proximal direction 0.3 mm at a time. The distance from the crush to the point that elicited the reflex was considered the elongation attained by axons. This test was performed on the tibial and peroneal branches of the sciatic nerve.
Morphology. The nerves were infiltrated with fixa- tive (4% paraformaldehyde, 1% glutaraldehyde, 0.01% picric acid, 0.1 M buffer phosphate, pH 7.4), removed, and left overnight in the same fixative. After a short rinse, the tissue was immersed in buffered 1% osmium tetroxide for 2 hours followed by 2% uranyl acetate for 2 hours; finally, the tissue was dehydrated with graded acetones and infiltrated with Epon (for details, see 15). For electron microscopy, thin sections were double stained with uranyl acetate and lead citrate.
Detection of GAP-43. The localization of GAP-43 was done with a monoclonal antibody against GAP-43 (clone GAP-7B10, Sigma), following the method of Mieri et al. (34). Briefly, the tissue was fixed in paraformalde- hyde, dehydrated, and embedded in paraplast. Longitu- dinal sections were incubated with milk for 2 h and then, with the antibody 1:250 in 0.01% Triton X-100 milk, at 4°C overnight, rinsed with saline, treated with a goat anti-mouse IgG coupled to biotin 1:50 in milk for 2 h, followed by Extravidin peroxidase (Sigma), and devel- oped with hydrogen peroxide-diaminobenzidine. For positive controls, neonatal brain was used, and for negative controls, the specific antibody was replaced by preimmune serum.
Statistics. Data were subjected first to Bartlett’s test to establish the homogeneity of variance which enables the use of parametric statistics. Student, Newman, Keuls (SNK) test was used for discrete groups; the linear regressions were compared with the likelihood ratio test and ANCOVA. For details, see Mardia et al. (31)
RESULTS
Freezing of the nerve. The morphological pattern distal to the crush in frozen nerves differed in various respects from that of unfrozen nerves. Immediately after freezing, myelin, nuclei, cytoplasm and mem- branes of Schwann cells, and the axoplasm of myelin- ated fibers were clearly altered, and nonmedullated bundles were barely recognizable (Figs. 2A and 2B). Already at Day 1 postlesion (PL), the axoplasm was replaced by an amorphous material The myelin was delaminated extensively without signs of a progressive degeneration (Fig. 2C), even at Day 3 PL. The cytoplasm of Schwann cells was decaying and their nuclei disap- peared. This morphological pattern grants that the freezing of the nerve killed all cells.
Elongation of peroneal nerve axons was assessed at Days 1, 2, and 3 PL. The elongation progressed at a velocity of 2.6 mm/day, after a delay of 19.2 h estimated by extrapolation to zero elongation. The freezing of the

126 TAPLA. INESTROSA, AND ALVAREZ
FIG. 2. Morphology of nerve fibers. (A) Normal nerve, for comparison. (Bl Immediately after freezing; myelin sheaths, nuclei (arrowheads), and myelinated and nonmedullated (nml axons are deeply altered (compare with Al. (Cl Frozen, Day 1 PL; decaying cells are ubiquitous. (D) Frozen, Day 2 PL; 15 axonal profiles lie between basal lamina and myelin; the original axoplasm and its surrounding myelin are decaying. (El Frozen, Day 2 PL; an axonal profile (arrowhead) contacts the basal lamina and debris of Schwann cell cytoplasm and myelin. (F) Treated with aprotinin for 2 days prior to the crush and sacrificed at Day 2 PL; a cluster of axonal profiles, one of which is very large (*I, is surrounded by the Schwann cell; Id, lipid droplet. Calibration: A, B, and C, 2 Frn; D, E, and F, 0.5 pm.

REPRESSION OF EARLY REGENERATION 127
nerve distal to the crush obliterated the delay but the rate of elongation was unchanged (Fig. 3).
When the frozen segment was separated from the crush by 3 mm of unfrozen nerve (see Fig. ID), the elongation at Day 2 PL was similar to that of control nerves (Fig. 4). That is, a short undisturbed degenerat- ing segment interposed between crush and the frozen nerve region annulled the effect of freezing.
Under the electron microscope, we searched for re- growing axons 3-4 mm distal to the crush, at Days 1, 2, and 3 PL. In unfrozen nerves, these were not observed at Day 1 PL and seldom observed at later times. In frozen nerves, regrowing axons were seen regularly, usually associated with former myelinated fibers. They appeared as single or multiple profiles laying between the basal lamina and the remnants of Schwann cell cytoplasm or myelin; they had microtubules and fila- ments with side arms, which are the hallmark of neurofilaments (Figs. 2D and 2E). Macrophages were infrequent and they were not digesting myelin debris at this stage.
GAP-43 is a protein present in the front of axons that elongate actively (25). To visualize the extent of re- growth, GAP-43 was immunolocalized at Day 2 PL. In conditioned regenerating nerves, the positive profiles were more numerous, invaded a longer span of nerve, and were coarser, compared to those of unfrozen nerves (Fig. 5). Incubation with preimmune serum resulted in no reaction (not shown). This accords with the abun-
Freezing Aprotinm 10
a 6 1./-: 4
2
A A A days
FIG. 3. Time course of axonal elongation after a crush (mm + SEMI. Frozen (0) and unfrozen (0) peroneal nerves (see Figs. 1A and 1B). Aprotinin CO)- and saline (.)-injected tibial nerves (see Fig. 1C) for 2 days prior to the crush (arrowheads). Each time point includes 4-9 individuals. Regression lines and parameters (?SEM) are provided; r, correlation coefficient; n, sample size. Both treatments significantly shift the intercept (P < 0.0001) but do not affect the slope tANOVA, Bartlett, ANCOVA, and likelihood ratio tests).
Treatment Intercept Slope r R
Freez. 0 0
Aprot. 0 0
0.02 2 0.96 2.38 + 0.26 0.90 21 -2.06 t 0.52 2.57 e 0.13 0.97 24
-0.68 k 0.64 3.42 -c 0.23 0.98 12 -1.93 2 0.44 2.81 2 0.16 0.99 12
8
P
7
E6 T .P 5
p 7E3
2
1
0 1: co3 frozen
(3daY.5)
* li c 0 l( aprot.
Pdays)
-
FIG. 4. Undisturbed degenerating nerve segment interposed be- tween crush and conditioned region (mm ? SEM). C, control. Figures indicate gap between crush and treated segment. Frozen, peroneal nerve. Aprotinin, tibial nerve conditioned for 2 days before the crush. Survival time is indicated below the columns. Sample size, four individuals each. *, different from other groups (ANOVA, Bartlett, and SNK test). The effect of either freezing or aprotinin is abolished when a few millimeters of nerve undergoing a regular degeneration are interposed between crush and conditioned segment.
dance and size of axonal profiles as observed under the electron microscope and provides an independent evi- dence that the freezing of the nerve distal to the crush enhances regeneration.
Inhibition of proteases. When the tibial nerve was infiltrated with saline or aprotinin, but not crushed, the rat did not exhibit motor alterations after recovery from anesthesia or in the ensueing days, i.e., vehicle and aprotinin did not affect overtly the function of the nerve. One subperineurial injection of vehicle, or several in consecutive days, prior to the crush resulted in an axonal elongation similar to that of uninjected nerves (Table 1). Therefore, the infiltration itself does not affect the regenerative response.
Conditioning of the tibial nerve with aprotinin at the time of the crush determined an axonal elongation comparable to that of controls, as assessed at Day 3 PL (Fig. 6). To improve the effect of aprotinin, we condi- tioned the tibial nerve with several doses for up to 3 days prior to the crush. The elongation of axons was not affected by conditioning with aprotinin for up to 1.5 days prior to the crush. In contrast, the elongation was enhanced substantially after 2, and even more after 3 days of preconditioning (Fig. 6). Hence, the exposure to aprotinin seems to require a certain duration below which the effect is not discernible.
We assessed whether the infiltration with aprotinin of a sciatic nerve fascicle affected a contiguous one. The subperineurial injection of 2 ~1 into the tibial nerve was sufficient to displace the solution up into the tibial fascicle of the sciatic trunk, where it ran flanked by the peroneal fascicle. Following a preconditioning for 2 days, the regrowth of tibial nerve axons was enhanced, as already mentioned, but not that of peroneal nerve axons

128 TAPIA, INESTROSA, AND ALVAREZ
P I I
#
1 2mm 2mm 1 5mm
+
FIG. 5. Immunolocalization of GAP-43 in regrowing axons. Treatment and the distance to the crush are indicated below the micrograph. Survival, 2 days PL. Reaction products (arrows) are more abundant (center) and extend further distal in the frozen nerve (right) than in the unfrozen nerve (left), where no deposits were seen beyond 2 mm distal to the crush. Cal., 50 pm.
(Fig. 7). This suggests that aprotinin remains within the perineurial limits of the conditioned nerve branch.
The time course of regeneration of tibial nerve axons pretreated with vehicle alone reveals a rate of regrowth of 2.8 mm/day, after an estimated delay of 16.7 h (Fig. 3). Pretreatment of the nerve with aprotinin for 2 days prior to the crush reduced the delay to 4.7 h, which amounts to 28% of the control value. The elongation rate, though, did not change significantly (Fig. 3). That is, a nerve conditioned either with aprotinin or by freezing produces a similar pattern of axonal regenera- tion.
In the experiments described above, the crush was
TABLE 1
Elongation of Regenerating Axons after Injection of Saline into the Nerve (mm f SEM)
Iniections” Ree-rowthb nc
None 6.6 + 0.2 4 1 6.5 + 0.1 6 4 6.4 + 0.3 4
Note. A crush was made in the proximal region of the infiltrated segment. The injection of vehicle prior to the crush does not affect the elongation of regenerating axons.
D 1, one injection at the time of the crush. 4, four injections over a 3-day period; the last was simultaneous with the crush.
b assessed at Day 3 PL. c sample size.
made in the conditioned segment. To assess whether aprotinin conditioned the neuron as a whole or only the exposed region of the axon, the crush was performed away from the conditioned segment. Nerves pretreated with aprotinin for 2 days (three doses) were crushed about 10 mm central to the estimated limit of the conditioned region (see Fig. 1E). Despite the strong aprotinin treatment, the axonal elongation was similar to that of nerves injected with vehicle (Fig. 4). Thus,
c 0 1 1.5
** II 4
3 hyS
FIG. 6. Duration of aprotinin pretreatment and axonal growth (mm k SEM). Elongation was evaluated at Day 3 PL. Sample size is given inside the column. * and **, different between them and from all other groups; others are not statistically different between them (ANOVA and SNK test). The elongation is enhanced when the pretreatment lasts more than 1.5 days.

REPRESSION OF EARLY REGENERATION 129
tl bial
: : I’. 1,‘. ‘,
Iii .’ ., ~._ \, ,, . ‘;\
peroneal
FIG. 7. Restricted effect of aprotinin. Diagram: S, sciatic; T, tibia]; and P. peroneal nerves; dotted area, infiltrated segment of the tibia] nerve; arrowhead, crush. The tibia1 nerve was injected three times over a 2-day period before the crush. Elongation (mm ? SEM). evaluated at Day 3 PL. Empty columns. nerves corresponding to conditioning of the tibia1 nerve with vehicle; hatched columns. nerves corresponding to conditioning of the tibia1 nerve with aprotinin. Each experimental group, four individuals. *, statistically different from other groups IANOVA. Bartlett, and NSK tests). Aprotinin enhances the regrowth of the injected tibia1 fascicle but not that of the contiguous peroneal fascicle.
regeneration is affected only when the crush is made in the conditioned segment, as was also observed after freezing.
The cytological picture of intact nerve injected with aprotinin is normal at Day 3 postinjection (3). In crushed nerves, the tissue was studied under the elec- tron microscope at 3-4 mm distal to the crush. Condition- ing with aprotinin at the time of the crush resulted in a morphological pattern of degeneration similar to that of nerves pretreated with one or several injections of vehicle or uninjected degenerating nerves. At Day 2 PL, sprouts were rare. In contrast, after a 2-day pretreat- ment with aprotinin, axonal sprouts were seen regularly at Day 2 PL, some of them were large (Fig. 2F).
DISCUSSION
Our main finding is that local freezing and inhibition of proteases next to a nerve crush enhance regeneration as assessed with functional, morphological, and molecu- lar indicators. Both treatments reduce substantially the latency of regrowth; the rate of elongation notwithstand- ing is not affected. The freezing of the nerve segment distal to a crush has been reported to produce a discrete shortening of the delay, no effect, or a mild reduction of the elongation rate (42,43,44), as assessed from the Day 3 PL onward. In these reports, the surgical procedure was more traumatic than ours since a longer nerve segment (15 mm or more) was frozen with chilled forceps or with dry ice and thawed, a cycle repeated several times during more than 5 min. Our technique was more gentle (see Methods) and probably our cryole- sion resulted in a lesser disruption of the noncellular components or surrounding tissue. On the other hand,
we assessed the early axonal elongation, up to Day 3 PL, whereas their first time point was Day 3 PL.
Local mechanism. It has been generally considered that the early axonal regrowth does not involve the perikaryon (cf. 6). Notwithstanding, Shaw and Bray (41) showed that neurites detached from their cell bodies extend new growth cones. Rotshenker (37) in toad, and Masson and Muller (32) in leech showed that isolated axonal domains sprout before degeneration comes in. These observations indicate that the perikaryon is not necessary for the initiation of regrowth and that axons have all the elements of a sprouting program. Early morphological studies of Cajal (17), confirmed later by Wyrwicka (49) and Zelena et al. (50) indicate that axons sprout within hours of the lesion, occurrence that has been largely overlooked. Recently, the early elongation of axons was reevaluated and the latency was estimated in 2.8 h; this latency was deemed too short for the perikaryon to participate in the initiation of regrowth (42). In present experiments, the involvement of the perikaryon in the early regenerative events can be reasonably excluded. A systemic signalling can be ruled out because the treatment is of no consequence for a neighboring crushed nerve. Intraaxonal signalling can also be excluded; in the case of freezing because no latency was observed and in the case of aprotinin the latency of 4.5 h is insufficient for the 90-mm round trip of signals, even assuming the upper limit for the fast anterograde and retrograde transport velocities, 17 mm/h (410 mm/day) and 11 mm/h (264 mm/day), respectively (26).
To affect the latency, the conditioned nerve segment has to be apposed to the blind end of axons. This emphasizes the role of the environment in the early steps of regeneration and suggests that the treatment modifies primarily the receiving tissue leading to novel interactions with the invading axons.
Repression, Schwann cells, and proteases. A natural ability to acquire territory has been attributed to axons whose balance implies a mutual repression between fibers (see Introduction). In principle, any of the compo- nents of the distal nerve tissue may be responsible for the repressive action, namely, Schwann cells, extracellu- lar matrix, perineurial cells, fibroblasts, resident macro- phages, endothelial cells. Among these, Schwann cells stand out as the major cellular component of the peripheral nerve and seem a good candidate to partici- pate in the repression insofar as they are in close association with axons. In the peripheral nervous sys- tem, Schwann cells are the equivalent of CNS oligoden- drocytes, which repress the outgrowth of axons in viva and in vitro (16). Upon nerve injury, Schwann cells express different genes (5, 6, 18, 19, 28, 36, 39, 45), proliferate (111, and eliminate their myelin; at the same time, nerves become permissive for invading axons. As mentioned, treatment with inhibitor of proteases mim-
130 TAPIA. INESTROSA. AND ALVAREZ
its a nerve section despite the fact that axons remain intact; axons sprout (3), i.e., the tissue has become permissive, and Schwann cells proliferate and eliminate their myelin (2). It is suggestive the association between repression and mature Schwann cells on the one hand, and on the other, between the proliferating condition (which implies dedifferentiation) and permissiveness. Interestingly, this accords with in vitro observations, namely, proliferating Schwann cells support the re- growth of axons while quiescent cells do so poorly (4).
tioned nerves begin to elongate sooner but they do so at a normal rate; this accords with a release mechanism which leaves the processes working at their own rate. Present results admit interpretations unrelated to our hypothesis, e.g., aprotinin, or early degradation prod- ucts of the frozen tissue, may stimulate directly the severed axons to regrow sooner than usual. These possibilities, though, await independent supporting evi- dence.
Aprotinin, a potent inhibitor of serine proteases, is a 6.5kDa polypeptide found in the extracellular space (46). Nexin-1, a serine protease inhibitor, promotes neurite extension of cultured neurons by inhibiting thrombin (271. Leupeptin, aprotinin, and APP with the Kunitz insert, all of which are inhibitors of serine proteases, induce a sprouting response of intact axons while saline, serum albumin, APP without the Kunitz insert, and heated APP with the Kunitz insert, do not (3). Thus, a parsimonious interpretation of present results is that aprotinin reduced the delay by inhibiting an extracellular serine protease.
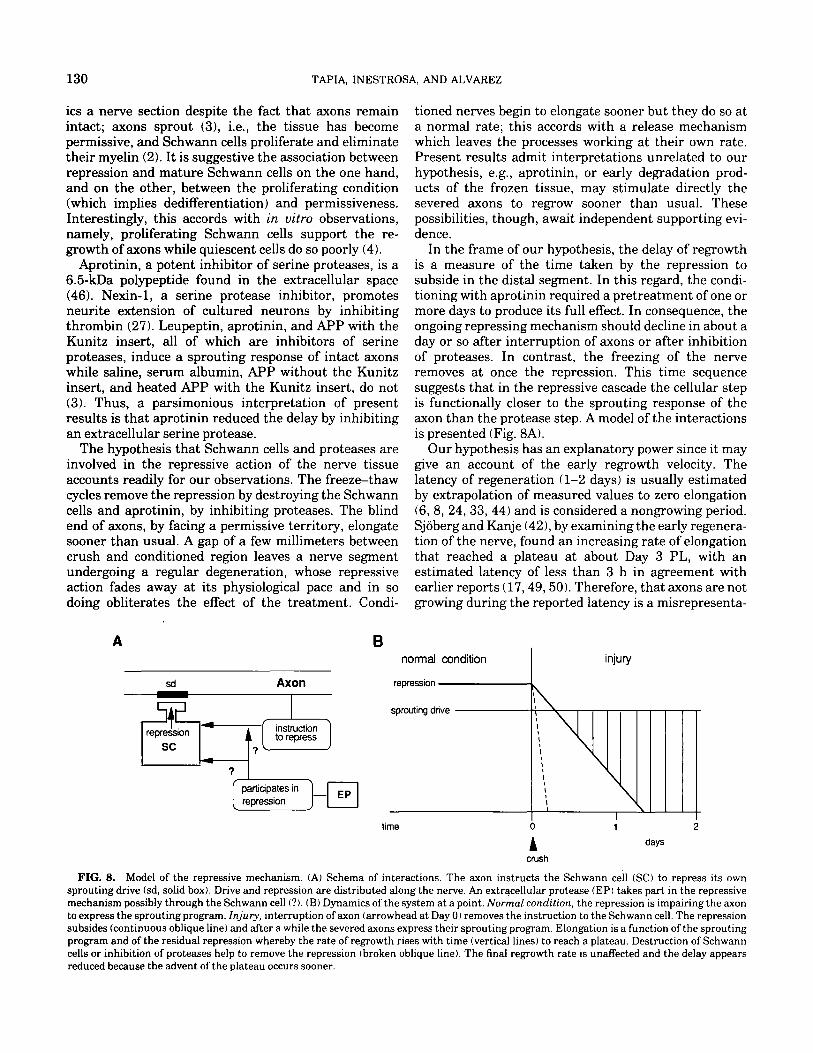
In the frame of our hypothesis, the delay of regrowth is a measure of the time taken by the repression to subside in the distal segment. In this regard, the condi- tioning with aprotinin required a pretreatment of one or more days to produce its full effect. In consequence, the ongoing repressing mechanism should decline in about a day or so after interruption of axons or after inhibition of proteases. In contrast, the freezing of the nerve removes at once the repression. This time sequence suggests that in the repressive cascade the cellular step is functionally closer to the sprouting response of the axon than the protease step. A model of the interactions is presented (Fig. 8A).
The hypothesis that Schwann cells and proteases are Our hypothesis has an explanatory power since it may involved in the repressive action of the nerve tissue give an account of the early regrowth velocity. The accounts readily for our observations. The freeze-thaw latency of regeneration (l-2 days) is usually estimated cycles remove the repression by destroying the Schwann by extrapolation of measured values to zero elongation cells and aprotinin, by inhibiting proteases. The blind (6, 8, 24,33,44) and is considered a nongrowing period. end of axons, by facing a permissive territory, elongate Sjoberg and Kanje (42), by examining the early regenera- sooner than usual A gap of a few millimeters between tion of the nerve, found an increasing rate of elongation crush and conditioned region leaves a nerve segment that reached a plateau at about Day 3 PL, with an undergoing a regular degeneration, whose repressive estimated latency of less than 3 h in agreement with action fades away at its physiological pace and in so earlier reports (17,49,50). Therefore, that axons are not doing obliterates the effect of the treatment. Condi- growing during the reported latency is a misrepresenta-
A 6
sd Axon
normal condition I
injury
time
crush
1 2
days
FIG. 8. Model of the repressive mechanism. (A) Schema of interactions. The axon instructs the Schwann cell (SC) to repress its own sprouting drive (sd, solid box). Drive and repression are distributed along the nerve. An extracellular protease (EP) takes part in the repressive mechanism possibly through the Schwann cell (?l. (B) Dynamics of the system at a point. Normal condition, the repression is impairing the axon to express the sprouting program. Injury, interruption of axon (arrowhead at Day 0) removes the instruction to the Schwann cell. The repression subsides (continuous oblique line) and after a while the severed axons express their sprouting program. Elongation is a function of the sprouting program and of the residual repression whereby the rate of regrowth rises with time (vertical lines) to reach a plateau. Destruction of Schwann cells or inhibition of proteases help to remove the repression (broken oblique line). The final regrowth rate is unaffected and the delay appears reduced because the advent of the plateau occurs sooner.

REPRESSION OF EARLY REGENERATION 131
tion of reality. The observed increasing rate of elonga- tion can be readily accounted for by the notion of a repressive action. After nerve injury, as the repression declines, the rate of elongation is expected to rise until it reaches a plateau when the tissue becomes fully permis- sive (Fig. 8B ). Our model also has room for an enhancing but delayed influence of perikaryon and supporting tissue on the regrowing axons.
Finally, we want to mention the abnormal regenera- tion reported for the C57BL/Ola mouse strain. For several weeks after a crush, degeneration of the distal nerve domain is considerably delayed, and proximal axons do not regenerate (7, 13, 35). Recently, Brown et al. (14) reported that after crushing a span of nerve, axons do regrow but they come to a halt upon reaching the uncrushed domain. They proposed that either the normal nerve fails to encourage, or actively inhibits, axonal regrowth. We think that the genetic disorder impairs the normal decline of the repression in the distal domain after nerve injury.
In conclusion, local freezing of a nerve segment or administration of an inhibitor of serine proteases re- duces the apparent latency of axonal regeneration. These findings support the notion that the sprouting response of axons is repressed by a local mechanism that presumably involves the mature Schwann cell and an extracellular serine protease.
ACKNOWLEDGMENTS
We thank M. P&ez. G. MQndez, and J. Morillas for technical assistance. This work was supported by FONDECYT 660192, the Stiftung Volkswagenwerk, and Deutsche Gesellschaft fiir Technische Zusammenarbeit GmbH.
8.
9.
10.
11.
12.
13.
14.
15.
16.
17.
18.
1.
2.
3.
4.
5.
6.
7.
REFERENCES 19.
AGCILI\R, C. E.. M. A. BISBY. E. COOPER, AND J. DI~\MOND. 1973. Evidence that axoplasmic transport of trophic factors is involved in the regulation of peripheral nerve fields in salamanders. J. 20. Phyysiol. iLondon) 234: 449-464. ALVAREZ, J., R. D. MORENO, AND N. C. INESTROSA. 1994. Mitosis of Schwann cells and demyelination are induced by the amyloid precursor protein and other protease inhibitors in the rat sciatic 21. nerve. Eur. J. Neurosci. 7: 152-159.
ALVAREZ, J., R. D. MORENO, 0. L~~h’os, N. C. INESTROSA. E. BR&ND.,XN, T. COLBY, AND F. S. ESCH. 1992. Axonal sprouting induced in the sciatic nerve by the amyloid precursor protein 22. (APP) and other antiproteases. Neurosci. Lelf. 144: 130-134. BAEHR, M., AND R. P. BUNG!?. 1989. Functional status influences the ability of Schwann cells to support adult rat retinal ganglion 23. cell survival and axonal regrowth. Exp. Neurol. 106: 2740.
BAIT-WALTER, I., C. Due, L. L. SARLIEVE, J. PL~MIRAT, J. H. DUSSALII.T, ,WD B. DROZ. 1992. The expression of nuclear 3,5.3’ triiodothyronine receptors is induced in Schwann cells by nerve 24. transection. Exp. Neural. 116: 189-197. BEREMBERC, R. A.. D. S. FORMAN, D. K. WOOD, A. DE SILVA. AND J. DEMAREE. 1977. Recovery of peripheral nerve function after axotomy: Effect of triiodothyramine. Exp. Neural. 57: 349-363. 25. BISBY, M. A., AND S. CHEN. 1990. Delayed waIlerian degeneration
in sciatic nerves of C57B110la mice is associated with impaired regeneration of sensory axons. Brain Res. 530: 117-120. BISRY. M. A., AND P. KEEN. 1985. The effect of a conditioning lesion on the regeneration rate of peripheral nerve axons contain- ing substance P. Brain Res. 336: 201-206. BISBY, M. A., AND W. TETZLAFF. 1992. Changes in cytoskeletal protein synthesis following axon injury and during axon regenera- tion. Mol. Neurobiol. 6: 107-123.
BOSCH, E. P., W. ZHONG, AND R. LIN. 1989. AxonaI signals regulate expression of glia maturation factor-beta in Schwann cells: An immunochemical study of injured sciatic nerves and cultured Schwann cells. J. Neurosci. 9: 3690-3698. BRADL.EY, W. G., AND A. K. ASBURY. 1970. Duration of synthesis phase in neurilemma cells in mouse sciatic nerve duringdegenera- tion. Exper. Neural. 26: 275-282.
BR,!Y. G. M., M. RASMINSKY, &VD A. J. ACUAYO. 1981. Interactions between axons and their sheath cells. Annu. Reu. Neurosci. 4: 127-162. BRONX, M. C., E. R. LYNN, ANDV. H. PERRY. 1992. Consequences of slow waIlerian degeneration for regenerating motor and sensory axons. J. Neurobiol. 23: 521-536. BROWN, M. C., V. H. PERRY, S. P. HUNT, .~ND S. R. LAPER. 1994. Further studies on motor and sensory nerve regeneration in mice with delayed WaIlerian degeneration. Eur. J. Neurosci. 6: 420- 428. BUSTOS, J.. J. D. VIAL. V. FA~~NDEZ, AND J. ALVAREZ. 1991. Axons sprout and microtubules increase after local inhibition of RNA synthesis, and microtubules decrease after inhibition of protein synthesis: A morphometric study of rat sural nerves. Eur. J. Neurosci. 3: 1123-1133. CADELLI, D. S., C. E. BANDTLOW, AND M. E. SCHWAB. 1992. Oligodendrocyte- and myelin-associated inhibitors of neurite outgrowth: Their involvement in the lack of CNS regeneration. Exp. Neural. 115: 189-192.
CIWAL, S. R. (1928) Degeneration and Regeneration of the Nervous Sys/em. Oxford Univ. Press, Oxford. COHEN, J. A., A. T. YACHNIS, M. ARAB. J. G. DAVIS, AND S. S. SCHERER. 1992. Expression of the neu proto-oncogene by Schwann cells during peripheral nerve development and WaIlerian degen- eration. J. Neurosci. Res. 31: 622-634. DE LEON, M.. A. A. WELCHER, V. SUTER, AND E. M. SHOOTER. 1991. Identification of transcriptionally regulated genes after sciatic nerve transection. J. Neurosci. Res. 29: 437448. DE WAECH, S., AND S. T. BRADY. 1990. Altered slow axonal transport and regeneration in a myelin-deficient mutant mouse: the trembler as an in uivo model for Schwann cell-axon interac- tions. J. Neurosci. 10: 1855-1865. DE WAEGH, S., V.M.-Y LEE, AND S. T. BRADY. 1992. Local modulation of neurofilament phosphorylation, axonal caliber, and slow axonal transport by myelinating Schwann cells. Cell 68: 451-463.
EDDS, M. V. 1950. Collateral regeneration of residual motor axons in partially denervated muscles. J. Exp. Zool. 113: 517- 532. ESCH. F. S., P. S. KEIM, E. C. BEATTIE, A. R. BLXHER, R. C. CULWELL, T. OLTERSDORF, D. MCCLURE, AND J. P. WARD. 1990. Cleavage of amyloid p peptide during constitutive processing of its precursor. Science 248: 1122-l 124. FORMAN, D. S., I. G. MCQUARRIE, F. W. LABARE, D. K. WOOD, L. S. STONE, C. H. BRADDOCK AND D. A. FUCHS. 1980. Time course of the conditioning lesion effect on axonal regeneration. Brain Res. 182: 180-185. GISPEN, W. H., H. B. NIELANDER, P. N. E. DE GRAAN, A. B. OESI’REICHER. L. H. SCHRAMA, MD P. SCHOTMAN. 1992. Role of the

132 TAPIA, INESTROSA, AND ALVAREZ
26.
21.
28.
29.
30.
31.
32.
33.
34.
35.
36.
37.
growth-associated protein B-50/GAP-43 in neuronal plasticity. Mol. Neurobiol. 5: 61-78.
GRAFSTEIN, B., AND D. S. FORMAN. 1980. Intracellular transport in neurons. Physiol. Reu. 60: 1167-1283. GIJRWITZ, D., AND D. D. CUNNINGHAM. 1990. Neurite outgrowth activity of protease nexin-1 on neuroblastoma cells requires thrombin inhibition. J. Cell. Physiol. 142: 155-162. IGNATIUS, M. J., P. J. GEBICKE-H~~RTER, J. H. P. SKENE, J. W. SCHILLING, K. H. WEISGRABER, R. W. MAHLEY, AND E. M. SCHOOTER. 1986. Expression of apolipoprotein E during nerve degeneration and regeneration. Proc. Natl. Acad. Sci. USA 83: 1125-1129. LANGLEY, J. N., AND H. K. ANDERSON. 1904. The union of different kinds of nerve fibres. J. Physiol. 31: 365-391. L~)PEz, J. M. A., AND J. ALVAREZ. 1990. The microtubular pattern changes at the spinal cord-root junction and reverts at the root-peripheral nerve junction in sensory and motor fibres of the rat. Eur. J. Neurosci. 2: 873-878. ~DIA, K. V., J. T. KENT, AND J. M. BIBBY. 1979. Mulliuariale Analysis. Academic Press, London. WON, A., AND K. J. MULLER. 1982. Axon segments sprout at both ends: Tracking growth with fluorescent D-peptides. Nature 296: 655-657. MCQLJARRIE, I. G. AND J. M. JACOB. 1991. Conditioning nerve crush accelerates cytoskeletal protein transport in spronts that form after a subsequent crush. J. Camp. Neurol. 305: 139-147. MEIRI, K. F., L. E. BICKERSTAFF. AND J. E. SCHWOD. 1991. Monoclonal antibodies show that kinase C phosphorylation of GAP-43 during axonogenesis is both spatially and temporally restricted in vivo. J. Cell Biol. 112: 991-1005. PERRY, V. H., M. C. BROWN, E. R. LUNN, P. TREE, AND S. GORDON. 1990. Evidence that very slow waIlerian degeneration in C57BLi Ola mice is an intrinsic property of the peripheral nerve. Eur. J. Neurosci. 2: 802-808. RABINOVSKY, E. D., G. M. SMITH, D. P. BROWDER, H. D. SHINE, AND J. L. MCMANAMAN. 1992. Peripheral nerve injury down-regulates CNTF expression in adult rat sciatic nerves. J. Neurosci. Res. 31: 188-192. ROTSHENKER, S. 1981. Sprouting and synapse formation by motor axons separated from their cell bodies. Brain Res. 223: 141-145.
38.
39.
40.
41.
42.
43.
44.
45.
46.
47.
48.
49.
50.
SAITUA, F., AND J. ALVAREZ. 1989. Microtubular packing varies along the course of motor and sensory axons: Possible regulation of microtubules by environmental cues. Neurosci. Let/. 104: 249-252.
SENDI‘NER. M., K. A. ST&XLI, AND H. THOENEN. 1992. Synthesis and localization of ciliary neurotrofic factor in the sciatic nerve of the adult rat after lesion and during regeneration. J. Cell Biol. 118: 139-148.
SERB, M.. AUD J. ALVAREZ. 1989. On the asymmetry of the primary branching of vagal sensory axons: Possible role of the supporting tissue. J. Camp. Neural. 284: 108-l 18.
SHAW. G., AND D. BRAY. 1977. Movement and extension of isolated growth-cones. Exp. Cell Res. 104: 55-62.
SJOUER(;, J., AND M. KANJE. 1990. The initial period of peripheral nerve regeneration and the importance of local environment for the conditioning lesion effect. Brain Rex 529: 79-84.
SJ~BERC., J.. M. KANJE, AND A. EDSTROM. 1988. Influence of non-neuronal cells on regeneration of the rat sciatic nerve. Brain Res. 453: 221-226.
SKETELJ, J., M. BRESJANAC’, AND M. POPOVIC. 1989. Rapid growth of regenerating axons across the segments of sciatic nerve devoid of Schwann cells. J. Neurosci. Res. 24: 153-162. SPREYER, P., G. KUHN, C. 0. HI\NEMANN, C. GILLEN, H. SCHAAL, R. KUHN, G. LEMKE, AND H. W. MULLER. 1991. Axon-regulated expression of a Schwann cell transcript that is homologous of a ‘growth arrest-specific’ gene. EMBO J. 10: 3661-3668.
TIUUTSCHOLD, I., E. WERLE, AND G. ZICKGRAF-RUDEI.. 1967. Trasylol. Biochem. PharmacoI. 16: 59-72. VAN HARREVELD, A. (1945) RE-INNERVATION OF DENERVATED hluscLE FIBRES BY ADJACENT FUNCTIONING MOTOR UNITS. z&f. J. PHKSIOL.
144: 477493.
WEDDELL, G., L. GUT-I-MANN, AND E. GUTMANN. 1941. The local extension of nerve fibres into denervated areas ofskin. J. Neural. Neurosurg. Psychiatr. 5: 206225.
WYRWICKA, W. 1950. On the rate of regeneration of the rat sciatic nerve in the white mouse. Acta Biol. Exp. 15: 147-153. ZELENA, J., L. LUBINSKA. AND E. GLITMANN. 1968. Accumulation of organelles at the ends of interrupted axons. 2. Zellforsch. 91: 200-2 19.
Copyright © 2022 FDOKUMEN




















