
Earliest Triassic microbialites in Çürük Dag, southern Turkey: composition, sequences and...
17
Earliest Triassic microbialites in C ¸u ¨ ru ¨ k Dag, southern Turkey: composition, sequences and controls on formation STEVE KERSHAW*, SYLVIE CRASQUIN , MARIE-BE ´ ATRICE FOREL , CARINE RANDON , PIERRE-YVES COLLIN à , ERDAL KOSUN§, SYLVAIN RICHOZ – and AYMON BAUD** *Institute for the Environment, Halsbury Building, Brunel University, Kingston Lane, Uxbridge, Middlesex UB8 3PH, UK (E-mail: [email protected]) CNRS-UMR 7207, CR2P ‘‘Centre de Recherche sur la Pale ´ obiodiversite ´ et les Pale ´ oenvironnements’’, Universite ´ Pierre et Marie Curie–Paris 6, T. 46–56, E. 5, Case 104, 4 Place Jussieu, 75252 Paris Cedex 05, France àCNRS–UMR 7072, IsTEP (Institute of Earth Sciences of Paris), Universite ´ Pierre et Marie Curie–Paris 6 University, T. 56–66, E. 5, Case 117, 4 Place Jussieu, F-75252 Paris Cedex 05, France §Department of Geology, Akdeniz University, 07058 Campus Antalya, Turkey –Commission for the Palaeontological and Stratigraphical Research of Austria, Austrian Academy of Sciences c/o Institute of Earth Sciences, University of Graz, Heinrichstrabe 26, 8010 Graz, Austria **BGC, Rouvraie 28, CH-1018 Lausanne, Switzerland Associate Editor – Daniel Ariztegui ABSTRACT The Permian–Triassic Boundary sequence at C ¸u ¨ru ¨ k Dag, near Antalya, Turkey, begins with a major erosion surface interpreted as being the Late Permian lowstand, on which lies ca 0Æ4 m of grainstone/packstone composed of ooids, peloids and bioclasts. Most ooids are superficial coats on fragments of calcite crystals presumed to be eroded from crystal fans which are no longer present. The erosion surface is smooth and shows no evidence of dissolution; the grainstone/packstone contains intraclasts of the underlying wackestone, proving erosion. Next are 15 m of microbialite comprised of interbedded stromatolites, thrombolites, plus beds of planar limestones with small-scale erosion. The latter comprise a complex interlayering of stromatolitic, thrombolitic and peloidal fabrics and precipitated crystal fans, which form a hybrid of microbialite and inorganic carbonate, together with bioclastic debris and micrite. The C ¸u ¨ru ¨ k Dag microbialite sequence is repetitious; the lower part is more complex, with abundant stromatolites and hybrid microbialites. Some of the stromatolites are themselves hybrids composed of peloids and crystal fans. In the upper part of the sequence stromatolites are missing and the rock is composed mostly of recrystallized thrombolites that develop upwards from tabular to domal form. The domes form directly below small breaks in microbialite growth where very thin shelly micrites and grainstones/ packstones are deposited. Repetition of facies may be controlled by sea-level change; a deepening-up model is consistent with the evidence. Stromatolites (with abundant crystal fans) dominate in shallower water, deepening through hybrid microbialite and interlayered sediments to thrombolite, probably no more than a few tens of metres deep, followed by breaks and renewal of microbialite growth. An interpretation of open marine fully oxygenated waters for microbialite growth is consistent with ongoing parallel work that has identified Bairdioid ostracods in the microbialite, a group known to be open marine. However, other researchers have proposed low oxygen conditions for Sedimentology (2011) 58, 739–755 doi: 10.1111/j.1365-3091.2010.01181.x ȑ 2010 The Authors. Journal compilation ȑ 2010 International Association of Sedimentologists 739
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Earliest Triassic microbialites in Çürük Dag, southern Turkey: composition, sequences and...

Earliest Triassic microbialites in Curuk Dag, southern Turkey:composition, sequences and controls on formation
STEVE KERSHAW*, SYLVIE CRASQUIN� , MARIE-BEATRICE FOREL� ,CARINE RANDON� , PIERRE-YVES COLLIN� , ERDAL KOSUN§, SYLVAIN RICHOZ–
and AYMON BAUD***Institute for the Environment, Halsbury Building, Brunel University, Kingston Lane, Uxbridge,Middlesex UB8 3PH, UK (E-mail: [email protected])�CNRS-UMR 7207, CR2P ‘‘Centre de Recherche sur la Paleobiodiversite et les Paleoenvironnements’’,Universite Pierre et Marie Curie–Paris 6, T. 46–56, E. 5, Case 104, 4 Place Jussieu, 75252 Paris Cedex 05,France�CNRS–UMR 7072, IsTEP (Institute of Earth Sciences of Paris), Universite Pierre et Marie Curie–Paris 6University, T. 56–66, E. 5, Case 117, 4 Place Jussieu, F-75252 Paris Cedex 05, France§Department of Geology, Akdeniz University, 07058 Campus Antalya, Turkey–Commission for the Palaeontological and Stratigraphical Research of Austria, Austrian Academy ofSciences c/o Institute of Earth Sciences, University of Graz, Heinrichstrabe 26, 8010 Graz, Austria**BGC, Rouvraie 28, CH-1018 Lausanne, Switzerland
Associate Editor – Daniel Ariztegui
ABSTRACT
The Permian–Triassic Boundary sequence at Curuk Dag, near Antalya, Turkey,
begins with a major erosion surface interpreted as being the Late Permian
lowstand, on which lies ca 0Æ4 m of grainstone/packstone composed of ooids,
peloids and bioclasts. Most ooids are superficial coats on fragments of calcite
crystals presumed to be eroded from crystal fans which are no longer present.
The erosion surface is smooth and shows no evidence of dissolution; the
grainstone/packstone contains intraclasts of the underlying wackestone,
proving erosion. Next are 15 m of microbialite comprised of interbedded
stromatolites, thrombolites, plus beds of planar limestones with small-scale
erosion. The latter comprise a complex interlayering of stromatolitic,
thrombolitic and peloidal fabrics and precipitated crystal fans, which form a
hybrid of microbialite and inorganic carbonate, together with bioclastic debris
and micrite. The Curuk Dag microbialite sequence is repetitious; the lower part
is more complex, with abundant stromatolites and hybrid microbialites. Some
of the stromatolites are themselves hybrids composed of peloids and crystal
fans. In the upper part of the sequence stromatolites are missing and the rock is
composed mostly of recrystallized thrombolites that develop upwards from
tabular to domal form. The domes form directly below small breaks in
microbialite growth where very thin shelly micrites and grainstones/
packstones are deposited. Repetition of facies may be controlled by sea-level
change; a deepening-up model is consistent with the evidence. Stromatolites
(with abundant crystal fans) dominate in shallower water, deepening through
hybrid microbialite and interlayered sediments to thrombolite, probably no
more than a few tens of metres deep, followed by breaks and renewal of
microbialite growth. An interpretation of open marine fully oxygenated waters
for microbialite growth is consistent with ongoing parallel work that has
identified Bairdioid ostracods in the microbialite, a group known to be open
marine. However, other researchers have proposed low oxygen conditions for
Sedimentology (2011) 58, 739–755 doi: 10.1111/j.1365-3091.2010.01181.x
� 2010 The Authors. Journal compilation � 2010 International Association of Sedimentologists 739

Permian–Triassic boundary facies globally, so work continues to confirm
whether the Curuk Dag microbialite grew in dysoxic or normally oxygenated
conditions. The principal stimulus for post-extinction microbialites is likely to
be carbonate supersaturation of the oceans. The microbialite sequence is
overlain by a further 25 m of grainstone/packstone (without microbialite),
followed by Early Triassic shales. Overall, microbialites form a thin
aggradational sequence during an overall relative sea-level rise, consistent
with global eustatic rise following the Late Permian lowstand.
Keywords Hybrid microbialite, mass extinction, microbialite, stromatolite,thrombolite, Triassic.
INTRODUCTION
Microbialites in the Permian–Triassic Boundary(PTB) sequence in Curuk Dag, near Antalya,southern Turkish Taurides (Fig. 1), are one of anumber of deposits of microbialites formedworldwide after the end-Permian mass extinc-
tion. These deposits developed in a facies com-plex of changing environmental conditions. Themicrobialites of this study were included togetherwith oolites as ‘calcimicrobial caprock’ by Baudet al. (2005) and illustrated in Crasquin et al.,(2009). Although PTB microbialites are well-known, a continuing problem is precise determi-
AA
Gedelma
10 kmPalaeoTethys
TURKEYPanthalassa
PangaeaU US
SArmutcuk
ANTALYA
KNeoTethys
PangaeaU UU U
S
U
S
U = upwelling
= sinkingS
Uçoluk
Oyacuk
C D
B
Kemer
*CD
Fig. 1. (A) Map of Turkey, showing location of Curuk Dag south-west of Antalya, near Kemer ‘K’; see inset for accessto site (asterisk) via a steep unmade road east of the village of Ucoluk. Detailed geology is provided by Baud et al.(2005) and not repeated here. (B) Palaeogeographic position of Curuk Dag ‘CD’ in western Tethys (map based onCrasquin-Soleau et al., 2001 and Kershaw et al., 2007), with modelled oceanographic characteristics (Kidder &Worsley, 2004). (C) Field photograph of Curuk Dag, highlighting the location of the Permian–Triassic Boundary; thecliff is ca 400 m high. (D) View of Permian–Triassic Boundary transition facies on Curuk Dag, conformable lime-stones [note person for scale (ca 1Æ8 m tall) in lower left hand corner, circled].
740 S. Kershaw et al.
� 2010 The Authors. Journal compilation � 2010 International Association of Sedimentologists, Sedimentology, 58, 739–755
nation of the conditions of growth of any indi-vidual deposit; PTB microbialite sequences varyfrom place to place regionally, even within onenappe in southern Turkey, and variation is clearglobally. Thus, post-mass extinction microbialiteswere under the influence of both global and localenvironmental conditions at any one site (see alsoKershaw et al., 2007). Therefore, it is important toattempt to account for microbialite formation indetail, on a site-by-site basis, to understand theprocesses operating on shallow shelves. A high-resolution approach is probably the best way totest models for shallow ocean change in relationto both local and global effects.
This study describes and interprets the detailedchanges that occurred through the microbialitesequence at Curuk Dag, which is a candidate forthe most complex PTB microbialite sequence
worldwide. The aim is to determine the localenvironmental changes that took place throughthe development of these post-extinction micro-bialites, against the backdrop of global environ-mental changes.
METHODOLOGY
This study was undertaken by high-resolutionfield logging (taking into account palaeontolog-ical content and sedimentary structures) andsampling in May 2008 and September 2009.Examination of about 50 polished hand speci-mens and 120 thin sections through thesequence, from the Late Permian erosion surfacethrough to the top of the microbialite, provideddata for facies analysis.
150
160B
120
13015
14
13
12
11
10
9
8
0
1
2
3
4
5
6
7
m
140
A
100
110
120
70
80
90
40
50
60
20
30
40
0Bioclasticwackestone Intraclasts Late Permian
erosion surface
10cm
Permian-Triassic boundary Shelly micritewith intraclasts
Thromboliteheads and breaks
Tabular beddedthrombolite
Mini-stromatolitedomes
Hybrid microbialite& interbeddedcarbonates
K O
K A
R K
U Y
U F
mP
. Fm
K O
K A
R K
U Y
U F
m
Domal and wavystromatolite
Basal microbialite(thrombolite)
Oolite-bioclasticgrainstone
Late Permianwackestone witherosion surface
microbialite sequence at
Çürük Dag, Antalya, Turkey
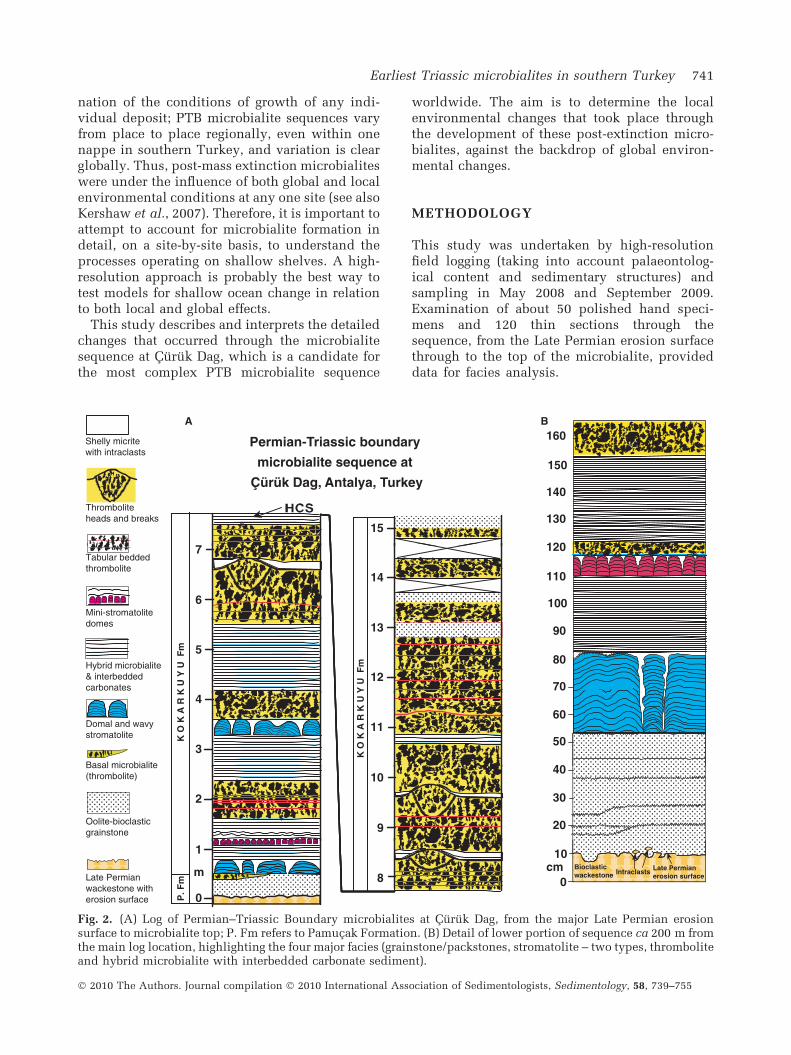
Fig. 2. (A) Log of Permian–Triassic Boundary microbialites at Curuk Dag, from the major Late Permian erosionsurface to microbialite top; P. Fm refers to Pamucak Formation. (B) Detail of lower portion of sequence ca 200 m fromthe main log location, highlighting the four major facies (grainstone/packstones, stromatolite – two types, thromboliteand hybrid microbialite with interbedded carbonate sediment).
Earliest Triassic microbialites in southern Turkey 741
� 2010 The Authors. Journal compilation � 2010 International Association of Sedimentologists, Sedimentology, 58, 739–755

GEOLOGICAL SETTING
The microbialites crop out in the Tauridestectonic complex and the reader is referred toBaud et al. (1997, 2005) and Crasquin et al.(2009) for details, including a geological map.In Curuk Dag (translated as ‘rotten mountain’due to erosion of shales in its lower part)Permian and Triassic limestones dip to the southat ca 50�, and the PTB is accessed easily from theflat top of the mountain and lies at GPS:36�41¢324¢¢ N, 030�27¢40.1¢¢ E. Rocks directlybelow the microbialite are the topmost part ofthe Pamucak Formation, wackestones and grain-stones/packstones of Late Permian age. Themicrobialite base marks the beginning of theKokarkuyu Formation (see Baud et al., 2005 fordetails), 40 m thick at Curuk Dag. Conodontbiostratigraphy is not well-developed for CurukDag; Isarcicella staeschei has been reported fromthe base of the Korkarkuyu Formation; 55 cmhigher, Hindeodus parvus was found (Richoz,2006). The occurrence of H. parvus aboveI. staeschei suggests that the former is near thetop of the range of H. parvus, since H. parvusfirst appears in the Meishan Global Boundary
Stratotype Section and Point, thereby definingthe base of the Triassic (Erwin, 2006). From thisargument, the base of the microbialite (Kok-arkuyu Formation) is already in the Triassic, butfurther work is needed to define the base of theTriassic in Curuk Dag. Note that Richoz (2006)showed that the prominent negative excursion inthe carbon isotope curve begins at the base of theoolite below the microbialite. In contrast, atsome other sites, such as in Iran (Wang et al.,2007) (the nearest PTB microbialites to Turkeyduring that time), the negative change occurswithin the microbialite, suggesting diachroneityof the beginning of microbialite growth (assum-ing that the negative excursion occurred syn-chronously in Turkey and Iran).
THE PERMIAN–TRIASSIC BOUNDARYSEQUENCE AT CURUK DAG
In Curuk Dag, PTB facies are visible in severalplaces on the west-facing side of the mountain,and show variations laterally and vertically.Several stages of development of the microbialites(Fig. 2) are described below, from bottom to top.
A
B
C
Fig. 3. (A) and (B) Field views of the Late Permian erosion surface below the microbialite sequence, showing itssmooth nature. The large divisions on the scale are centimetres. (C) Vertical thin section view of the erosion surface,showing eroded shelly wackestones and overlying grainstone/packstones, and wackestone intraclast (arrowed) in thegrainstone/packstone. The smooth erosion surface is consistent with physical erosion, without clear evidence ofdissolution. Scale bar is 5 mm.
742 S. Kershaw et al.
� 2010 The Authors. Journal compilation � 2010 International Association of Sedimentologists, Sedimentology, 58, 739–755

The latest Permian limestones contain a prom-inent smooth erosion surface (Figs 2 and 3),probably the Late Permian sea-level lowstand.There is no evidence of dissolution of thissurface, upon which was laid a 0Æ35 to 0Æ45 mthick grainstone/packstone unit composed of amixture of shell fragments, peloids, sphericalcalcitic grains with superficial ooid coats (Figs 3and 4) and uncommon fully formed ooids. Inmost cases, the calcitic grains are single crystalsand cannot be regarded simply as recrystallizedooids; they are interpreted here as reworkedfragments of calcite crystals precipitated afterthe mass extinction. No in-place calcite crystalshave been found in this deposit, or on the LatePermian erosion surface, consistent with erosionof the sea floor in a turbulent shallow waterenvironment.
The grainstone/packstone is overlain, withsharp contact, by microbialites. Baud et al.(2005) described a thin (13 cm thick) laterally
impersistent thrombolite, bound above and belowby distinct breaks that are likely to be erosionsurfaces. However, these surfaces are affected bystylolites directly above the grainstone/pack-stone. In the field, this unit was found only inone small section of the boundary facies in CurukDag, which is not figured here except in Fig. 2.
The first major microbialite is stromatolite,developed on a presumed erosion surface com-prising the thin thrombolite and grainstone/pack-stone described above. The stromatolite hasvarying gross morphology as masses with wavylaminae, prominent domes and narrow columns(Figs 2 and 5). Much of the stromatolite is layeredmicritic material indicative of sediment-trappingby cyanobacterial activity as in many modernstromatolites (Reid et al., 2000) (Fig. 6A to C);some spheroidal fabrics are present (Fig. 6D), buta common additional fabric is composed of layersof sparite (Fig. 7A to C), commonly interlayeredwith micritic stromatolite. Thus, much of the
A
C D
C
B
Fig. 4. Details of grainstone-packstone below the microbialite. (A) and (B) Many superficial ooids have nuclei ofrounded fragments of single crystals. (C) Packstone fabric in parts of the unit. (D) Peloidal sediment and shell(gastropod, arrowed) in parts of the unit. Scale bar for (A), (C) and (D) is 1 mm; for (B) it is 0Æ5 mm.
Earliest Triassic microbialites in southern Turkey 743
� 2010 The Authors. Journal compilation � 2010 International Association of Sedimentologists, Sedimentology, 58, 739–755

stromatolite fabric is a mixture of micritic andpeloidal (presumed to be cyanobacterially medi-ated) and cloudy precipitated cements (probablyinorganic but possibly microbially mediated).
Directly overlying the lowest stromatolite is thefirst of several beds of mostly planar laminae(Fig. 8). This deposit is comprised of a varietyof fabrics, including stromatolite, thrombolite,crystal precipitates and peloidal sediment(Fig. 9). Stylolitic contact with underlying stro-matolites prevents determination of the nature ofthe depositional contact. Riding (2008) intro-duced the concept of hybrid microbialitesconsisting of microbial and inorganically precip-itated components within stromatolites; thushybrid microbialite is an appropriate term forthese PTB microbialites in Curuk Dag. In the field,small-scale erosion surfaces are visible (Fig. 8C).In one place the lamination is curved, resembling,
but not proving, hummocky cross-stratification(HCS) (Figs 2 and 8D). However, in some thinsections, erosion surfaces are difficult to prove;part of the fabric appears to be eroded (Fig. 9B),but these features may instead be due to unevenmicrobial micrite. Shelly grainstone/packstonecontaining well-preserved crinoid columnalsoccurs at 2Æ65 m (Fig. 9C and D), but is not foundanywhere else in the section. Sedimentary struc-tures suggest that this facies represents a nor-mally low energy environment subject to episodichigher energy, such as storms.
Between ca 1Æ1 to 1Æ18 m is a unit of smallstromatolite columns occurring at only one hori-zon (Figs 2 and 10); this was found in two sites200 m apart (compare Fig. 2A and B) and there-fore is probably continuous across at least thatdistance. Individual stromatolite columns are upto 3 cm diameter and 10 cm tall. This layer
A
C D
B
Fig. 5. Field photographs of lowermost stromatolites, showing: (A) and (B) domal and wavy and (C) columnar forms.In (A) and (B), the scale bar is 85 mm long; in (C), the large divisions on the scale are 10 mm. (D) Thin section viewshowing stromatolitic lamination. Scale bar is 10 mm.
744 S. Kershaw et al.
� 2010 The Authors. Journal compilation � 2010 International Association of Sedimentologists, Sedimentology, 58, 739–755

indicates unique conditions for a short period inthe development of the microbialite sequence.In detail the stromatolite columns are composedof a mixture of peloidal stromatolite with precip-itated fans in alternating layers and therefore arehybrid stromatolite (Fig. 10C and D).
At several horizons there are tabular thrombo-lites which develop into thrombolitic heads insome cases (Fig. 11). Thrombolites consist of anopen framework of presumed microbial growth(Figs 12 and 13), subsequently infilled withcements and sediments, some of which are geo-petal. Thrombolites are all recrystallized, but themargins of the structure are well-defined, so themeso-architecture and micro-architecture of theclotted fabric is shown clearly (Figs 12 and 13).Thin deposits of non-microbial shelly and intra-clastic wackestones, packstones and grainstones/packstones occur directly above three of thethrombolite units (Figs 2 and 14).
Examination of Fig. 2 reveals that the micro-bialites in Curuk Dag exhibit a repetitive char-acter, with more complex changes in the lowerpart of the sequence, the details of which areaffected by stylolites in some places. Verticalchanges in the lower part terminate sharply atthe top of the lowest thrombolite head, but nomajor erosion has been identified. In places themesoclotted appearance of thrombolites gradesinto a columnar form of thrombolite, similar inexternal form to the digitate dendrolitesdescribed by Kershaw et al. (2007) in Sichuan,south China. Thus, the architecture of thethrombolites in Curuk Dag is complex in detail.The upper part, above 6 m on the log (Fig. 2),consists mostly of thrombolites with some thinplanar hybrid microbialite and deposited sedi-ment. At two further levels there are shelly andintraclastic micrites deposited on the thrombo-lite head top surfaces (Figs 2 and 14). The total
A B
C D
Fig. 6. Thin section views of lowermost stromatolites. (A) to (C) Lamination with interruptions in growth. (B)Stromatolite with possible filaments. (D) Possible spheroids in stromatolite. Scale bar for (A) is 5 mm; for (B), (C) and(D) it is 1 mm.
Earliest Triassic microbialites in southern Turkey 745
� 2010 The Authors. Journal compilation � 2010 International Association of Sedimentologists, Sedimentology, 58, 739–755

thickness of stromatolites and thrombolites is ca15 m. The uppermost thrombolite is overlain byca 25 m of grainstone/packstone, described byBaud et al. (2005) as oolite; however, the actualthickness of this may be affected by faults visibleon the cliff face, but which are difficult to detectin logged section. No faults were found cuttingthe microbialite.
DISCUSSION
Oolite as a form of microbialite?
Previous work (Baud et al., 2005) interpretedearliest Triassic microbialite limestones at CurukDag to comprise 40 m of limestone, which isthe thickest post-extinction microbialite so fardescribed. However, Baud et al. (2005) grouped
grainstones/packstones (which they described asoolite) with microbialites of the earliest Triassic ofCuruk Dag as ‘calcimicrobial caprock’ on LatePermian limestones, thereby taking into accountthe possible microbial contribution to ooidformation (see Flugel, 2004 for discussion).Although ooids may form under at least partialmicrobial control, perhaps by microbial mediationof carbonate deposition, there are two problemswith the above interpretation in Curuk Dag.Firstly, most ooids are superficial ooids, withconcentric ooids being uncommon; and much ofthe rock is composed of bioclasts, so these depos-its are not simply classified as oolite. Secondly, ifooids are included with microbialites, withoutdistinction from stromatolites and thrombolites(in Curuk Dag), then there are problems indiscriminating facies and making appropriateinterpretations of environmental controls. There
A B
C D
Fig. 7. (A) to (C) Thin section views of precipitated cements; all these were found in stromatolitic layers and forminterlayered cements with stromatolites. Thus stromatolites are hybrid structures containing both cement fans andapparent microbial fabrics. (D) Enlargement of Fig. 6A, showing cement growth on stromatolite laminae, beforemicrite deposition, indicating a hiatus between cessation of stromatolite growth and sediment deposition, possibly ina deepening setting, discussed in the text. Scale bar for (A) is 5 mm; for (B), (C) and (D) it is 1 mm.
746 S. Kershaw et al.
� 2010 The Authors. Journal compilation � 2010 International Association of Sedimentologists, Sedimentology, 58, 739–755

are also numerous well-known large oolite depos-its throughout the rock record which are unrelatedto mass extinctions, yet they are not considered tobe microbialite. In addition, some other post-extinction microbialites are not interbedded withoolites, for example in Sichuan and Guizhou,southern China (Kershaw et al., 1999, 2007).Thus, it is argued that there is no automatic linkbetween oolites and mass extinction, or betweenoolites and microbialites, and the (oolitic) grain-stone/packstone is considered to be separate frommicrobialite in these post-extinction facies.
Nevertheless, such grainstones/packstones arepotentially important as indicators of increasedcarbonate saturation during mass extinctionepisodes, when calcification of metazoans issuppressed due to a lack of shelly organisms(Groves & Calner, 2004), and are important infacies analysis of the sequence. Thus, maintain-ing a distinction from microbialites allows faciescomplexes containing both types of limestones to
be more fully assessed. Therefore, in Curuk Dag,there is only ca 15 m of microbialite, assumingthat the microbialite is not faulted.
Repetition of microbialites and modelsof growth
This study reveals repetitious growth of PTBmicrobialites in the study area. All types ofmicrobialite calcification are probably driven byhigh levels of carbonate saturation (Riding, 2005),which has also been cited as the reason forabundant oolites in earliest Triassic sediments(Groves & Calner, 2004; Calner, 2005). Changes inocean saturation may be expected to take placeover longer time scales than the presumed rapidfacies changes observed at this site. Therefore,because of the dominance of microbialitesthroughout the basal Triassic in Curuk Dag, it isvery unlikely that variations in saturation levelscaused the small-scale microbialite repetitions.
A
CD
B
Fig. 8. Field photographs of hybrid microbialite with interbedded carbonate sediments. (A) and (C) Lowermoststromatolite (55 to 110 cm of log, see Fig. 2). (B) Loose specimen showing cross-lamination in the sediment, indi-cating energetic deposition in at least part of the fabric. (D) In-place example at 7Æ7 m (Fig. 2) showing possible HCS.Scale bar is 85 mm long.
Earliest Triassic microbialites in southern Turkey 747
� 2010 The Authors. Journal compilation � 2010 International Association of Sedimentologists, Sedimentology, 58, 739–755

Also, the microbialites are underlain and overlainby grainstone/packstone, so it is likely that theentire microbialite sequence is associated withshallow water processes, and sea-level change isthe simplest explanation for the cause of theserepetitions. The outcrops currently sit in a nappesequence and, although tectonic activity mayhave affected the region where the microbialitesgrew, sea-level rise was rapid during the EarlyTriassic and was likely to mask any tectonicvertical motion.
Modern microbialites thrive in the brackishconditions of Lake Clifton, the hypersaline con-ditions of Hamelin Pool in Shark Bay, and the
open marine environments of the Bahamas.Microbialites in general apparently are influencedlittle by salinity but instead are stimulated byraised carbonate saturation, a point emphasizedby Burne & Moore (1987) and Riding (2000) intheir descriptions of microbialites from differentenvironments. McNamara (2009) specificallynoted that salinity is unimportant in microbialitegrowth. Therefore, it is difficult to use modernanalogues to provide the precise informationnecessary to interpret the PTB microbialites.However, preliminary evidence from ostracodsin work in progress by Forel et al. shows that thedepositional environments were all open marine.
A B
C D
Fig. 9. (A) and (B) Typical appearance of hybrid microbialite in thin section, at 1Æ4 m in the log in Fig. 2. Detailedlabelling in (B). Note the contrast between the (interpreted) eroded peloidal sediments, stromatolitic peloidal fabrics,cement fans and thrombolite. (C) Shelly grainstone/packstone at 2Æ65 m (Fig. 2). (D) Detail of shelly grainstone/packstone at 2Æ65 m containing several crinoid columnals (with syntaxial cements); note the well-preserved characterof the columnals. Scale bar for (A) is 5 mm, for (B) and (C) it is 1 mm; and for (D) it is 0Æ5 mm.
748 S. Kershaw et al.
� 2010 The Authors. Journal compilation � 2010 International Association of Sedimentologists, Sedimentology, 58, 739–755

Bairdioidea forms, well-known as open marineostracods, are abundant; there is no indication ofrestricted facies based on an ostracod study.Thus, the vertical changes in the microbialitesof Curuk Dag are interpreted as due to environ-mental change in open marine conditions; oxygenlevels are discussed below.
However, whether the repeated microbialitesrepresent shallowing-up or deepening-upchanges is difficult to prove from the currentevidence. Post-extinction stromatolites in theEarly Triassic are known from open marinecarbonate platform systems (for example, in Iran;Baud et al., 1997; Wang et al., 2007) and inHungary there is a strong argument for an outerramp setting (Hips & Haas, 2006), therefore beingin relatively deeper water. However, stromatolitesin general are also common components oflagoonal environments well-known from suchsites as Shark Bay, Australia (McNamara, 2009).Lake Clifton in Western Australia is well-knownfor its large domal thrombolites in very shallowwater (Burne & Moore, 1987), in contrast to
thrombolites in deeper water in other lakes (LakeVan, Turkey, Kempe et al., 1991; Pavilion Lake,British Columbia, Laval et al., 2000). By analogy,in Curuk Dag, in a deepening-up model, stromat-olites may have grown in very shallow water, andthe thrombolites in deeper water; or the reversescenario prevailed in a shallowing-up model.Below is an interpretive outline of these twoalternative pathways, followed by a discussionwhich favours only one of them.
Pathway 1: Shallowing-up modelShallow-marine grainstone/packstone bars devel-oped on top of the Late Permian erosion surface,with a thin thrombolite in shallow water. Rapiddeepening stimulated stromatolite growth inopen marine conditions (however, this does notaccount for the small-scale, sharply boundedchanges in stromatolite morphology; see Fig. 2).Shallowing led to tabular and then domal throm-bolites, with termination at the tops of domalthrombolite heads (but note that no erosion ofthese heads has been observed). A second episode
A
C D
B
Fig. 10. Small stromatolite domesat 110 cm. (A) Field view; scaledivisions 10 mm. (B) Polishedsample; large scale divisions are10 mm. (C) and (D) Thin sectionviews showing that most of thesedomes are composed of cloudyprecipitated cement fans and implya hybrid organic–inorganic fabric.Scale bars are 5 mm.
Earliest Triassic microbialites in southern Turkey 749
� 2010 The Authors. Journal compilation � 2010 International Association of Sedimentologists, Sedimentology, 58, 739–755

of rapid deepening stimulated renewed stromat-olite growth for the lower part of the sequence,while in the upper part the lack of stromatolitessuggests that the deepening was less. The micro-bialite sequence was terminated by a return to theshallow waters of a grainstone/packstone forma-tion that is maintained for ca 25 m above themicrobialites, suggesting steady subsidence orsea-level rise, keeping shallow water for theseoverlying deposits.
Pathway 2: Deepening-up modelShallow water grainstone/packstone bars devel-oped on top of the Late Permian erosion surface.A small sea-level rise in the earliest Triassic led tostromatolite growth in shallow environments.The lowest stromatolite unit consists of severaldifferent architectural sub-units which could beinterpreted as a lagoonal setting, subject to smallscale, rapid sea-level fluctuations. Thus, thestromatolites may be formed with little sea-levelchange in an open marine environment behind
shoals of mobile grainstone/packstone bars, orjust seaward of the bars in shallow sub-tidalconditions. However, in this model the thinimpersistent thrombolite (Fig. 2), described byBaud et al. (2005), may represent a relic of alargely eroded microbialite that formed before thelowest stromatolite, thereby making the sequenceof events at this site more complex. The loweststromatolites may have formed on a hardgroundof pre-existing limestones and, therefore, may nothave been near any active ooid shoals, potentiallyaccounting for the lack of ooids and bioclastsbetween stromatolite columns. Continued sea-level rise led to open water where the hybridmicrobialites and interbedded sediments devel-oped, subject to storm action creating low-anglecross-stratification, including possible HCS. Fur-ther deepening stimulated thrombolite growth inmore open marine and, presumably, quieter con-ditions. At first, thrombolite growth was tabular,then, as accommodation space increased, waterdeepened and light levels fell, these developed
A B
C
Fig. 11. Field photographs of thrombolites. (A) Tabular thrombolite changes upwards to domal head at 8Æ5 m in thelog in Fig. 2. The length of the yellow scale is 0Æ5 m. (B) Typical thrombolitic mesoclot fabric. (C) Transverse sectionof tabular thrombolite showing some organization into a columnar structure. Scale in (B) and (C) is 85 mm long.
750 S. Kershaw et al.
� 2010 The Authors. Journal compilation � 2010 International Association of Sedimentologists, Sedimentology, 58, 739–755

into domal thrombolites. Thrombolite growth wasterminated either by storm action and depositionof sediment on the thrombolites or, alternatively,by deepening water that lowered light levelssufficiently to impede thrombolite growth, afterwhich sediments accumulated. Rapid shallowingand re-establishment of stromatolites in the lowerpart of the sequence followed. As the upper partof the sequence lacks stromatolites, perhaps thelater sea-level fluctuations did not always resultin the shallowest water depositional environ-ments. If this is true, then there was an overalldeepening trend up-section through the micro-bialite. The microbialite sequence is terminatedby a return to the shallow waters of grainstone/packstone formation that is maintained for ca25 m above the microbialites, suggesting contin-ued shallow water through steady subsidence orsea-level rise. The end of microbialite growth maybe due to a fall in carbonate saturation of thewater, higher energy or by grazing pressure fromherbivores.
The deepening-up model is thought to be moreconsistent with the evidence for the followingreasons. Firstly, the lowest part of the micro-bialite contains the most complex components,dominated by stromatolites, so it could beexpected to be a consequence of small changesof sea-level in a very shallow setting. The lowerpart also has abundant precipitated calcite fansassociated with the stromatolites. These fans arecompatible with extreme carbonate saturation inthe shallowest waters of the presumably warm,low-latitude setting of the Taurides in the PTBtransition. Furthermore, the formation of stro-matolites with micritic sediments and no ooids isconsistent with a rising base level where the ooidshoals are likely to have been migrating land-wards and, therefore, are less likely to be avail-able to supply ooids to the area of stromatoliteformation. Nevertheless, the lowest stromatolitesmay have grown on lithified and eroded grain-stones/packstones, as discussed earlier. Whateverthe precise conditions at the base of the first
A B
DC
Fig. 12. (A) Vertical section of a layer of thrombolite showing minor erosion at the top and overlain intraclast-richmicrite. (B) Vertical section of thrombolite with open fabric. (C) Thin section view of thrombolite frame mostlyinfilled with micrite. (D) Transverse section of mesoclot structure. Scale bar for (C) and (D) is 5 mm.
Earliest Triassic microbialites in southern Turkey 751
� 2010 The Authors. Journal compilation � 2010 International Association of Sedimentologists, Sedimentology, 58, 739–755

stromatolite, all the stromatolite deposits in thesequence are associated with micrites, reinforc-ing the view that they developed in low energyenvironments, interpreted as being deep (or atleast quiet) enough to be out of reach of ooidshoals.
Secondly, if the sequence represents a shallow-ing-up process, then there should be more coarse-grained material associated with the thrombo-lites. The thin, shelly, intraclastic wackestonesdeposited above the thrombolites may have beenimported by storm action that terminated thrombo-lite growth.
Thirdly, laminated sediments between stromat-olite and thrombolite beds contain a hybrid ofmicrobialite and deposited sediments, as well asprecipitated crystal fans. The sedimentary struc-tures of small-scale erosion (Fig. 8C) (and possi-ble HCS) are consistent with slightly deeper waterof the shoreface which is subject to episodicstorms. In this view, the thrombolites would haveformed as the deepest deposit.
Fourthly, global sea-level rose after the LatePermian lowstand, consistent with deepening-up; although this is somewhat circumstantial(because local rapid tectonic uplift could theo-retically override a global signal), it is clear thatthe microbialite sequence aggraded to form 15 mof deposit. The sequence must have been laiddown either on a subsiding sea floor or understeady sea-level rise, with deposition to fill theaccommodation space. The deepening-up modelis summarized in Fig. 15.
Oxygenation levels of the microbialitesequence
There is growing evidence of lowered oxygen inshallow water facies associated with upwelling ofdeep ocean waters and elevation of the chemo-cline (Wignall & Twitchett, 1996; Dolenc et al.,2001; Algeo et al., 2007; Kershaw et al., 2007; andreferences therein). However, in Curuk Dag, themicrobialite sequence contains bivalves, gastro-
A B
C D
Fig. 13. (A), (B) and (D) Thin section details of thrombolites, showing primary frame, partially infilled by micrite. (C)Microgastropod encased in thrombolite. Scale bars for (A), (B) and (C) are 1 mm; for (D) the scale bar is 0Æ5 mm.
752 S. Kershaw et al.
� 2010 The Authors. Journal compilation � 2010 International Association of Sedimentologists, Sedimentology, 58, 739–755

A
C D
B
Fig. 14. Sediment layers between microbialites, outlined with white lines in (A) and (B). (A) Shelly and intra-clast micrite on thrombolite head at 8Æ5 m (Fig. 2), overlain by a tabular thrombolite of the next thromboliteunit. (B) Micrite layers between thrombolites at 6Æ8 m (Fig. 2). The yellow rule in (A) and (B) is 0Æ5 m long.(C) Polished sample of shelly and intraclast micrite from sediments shown in (B). The large divisions on thescale bar are 10 mm. (D) Thin section of intraclast wackestone from sample location ‘21’ shown in (B); scale baris 1 mm.
Fig. 15. Model of microbialitefacies at Curuk Dag, showingthrombolite heads in the deepestwater. Note that the contact betweenthe ooid shoal and the stromatolitesis interpreted as an erosion surfacein the field, so that the stromatolitesmay have initiated on a solidsurface. See text for furtherexplanation.
Earliest Triassic microbialites in southern Turkey 753
� 2010 The Authors. Journal compilation � 2010 International Association of Sedimentologists, Sedimentology, 58, 739–755

pods and crinoids, presumed to have been livingnearby because many shells appear to be well-preserved (one gastropod was found encased inthrombolite; Fig. 13C). Beatty et al. (2008) viewedwell-developed ichnofossils in early Triassicdeposits as indicators of areas where recoveryfrom extinction was more rapid, perhaps due tomore favourable conditions, particularly in thehigh latitude location of their study. Whether ornot this argument also applies to the tropicalsetting of central Tethys (where Curuk Dag islocated, see Fig. 1) requires further work, but it ispossible that recovery from mass extinction wasfaster than has been interpreted previously. Thus,the location of Curuk Dag may well have escapedthe worst effects of poor oxygenation (as sup-ported by abundant open marine ostracods);reinforcing the view that carbonate saturation isthe principal cause of microbialite growth afterthe mass extinction. Also, recent work on ostrac-ods from China indicates that microbialites in theGreat Bank of Guizhou did not grow in lowoxygen conditions (Forel et al., 2009), in contrastto those in Sichuan, for which there is evidence oflowered oxygen (Crasquin-Soleau & Kershaw,2005), so there is a more complex situation whichrequires more research to resolve. However, inalmost all cases of microbialites in Curuk Dag, athin isopachous cement is present on the surfacesof microbialite carbonate, deposited prior tosedimentary micrite (for example, Figs 7D, 9Band 13A). Thus, there is a time gap between theend of individual microbialite growth and thedeposition of sediment. It is possible that theoxygen levels of the shallow shelf fluctuated overa short time scale, so that the microbial structuresdeveloped in low oxygen conditions, while theostracods which are deposited between thromb-olite branches and stromatolite heads representmore oxygenated water. More research is requiredto investigate such small-scale changes, whichmay originate in climatically driven ocean-circulation fluctuation below the resolution ofcurrent stratigraphy.
CONCLUSIONS
1 The post-extinction microbialite sequence atCuruk Dag is much more complex than previ-ously recognized.
2 Microbialite form is divided broadly intostromatolite, thrombolite and hybrid microbialite,indicating significant changes in environmentalcontrols as the sequence developed, probably due
to sea-level fluctuations. Some stromatolites arecomposed of peloidal material and precipitatedcrystal fans formed in close association, forminghybrid microbialites.
3 The microbialites show repetitious changeswhich may be interpreted in terms of sea-levelchange after the Late Permian lowstand, whensea-levels rose globally. Vertical changes in themicrobialite facies are more consistent withdeepening-up processes, so that stromatolitesformed in shallow water, and thrombolite headswere the deepest component, probably a few tensof metres below sea-level.
4 This work has permitted exploration of thecontrols on post-extinction microbialite facies ingreater detail than has previously been attainedand emphasizes the need for high-resolutionsampling in the interpretation of the very thinmicrobialite sequences found after the end-Permian mass extinction.
ACKNOWLEDGEMENTS
We are very grateful to Rainer Brandner andCharles Henderson for stimulating field discus-sion. This paper is a contribution to IGCP572‘‘Restoration of marine ecosystems following thePermian-Triassic mass extinction: lessons for thepresent’’. We are grateful to Paul Wignall and ananonymous referee for comments on the manu-script.
REFERENCES
Algeo, T.J., Ellwood, B., Nguyen, T.K.T., Rowe, H. and May-nard, J.B. (2007) The Permian-Triassic boundary at Nhi Tao,
Vietnam: evidence for recurrent influx of sulfidic water-
masses to a shallow-marine carbonate platform. Palaeo-geogr. Palaoeclimatol. Palaeoecol., 252, 304–327.
Baud, A., Cirilli, S. and Marcoux, J. (1997) Biotic response to
mass extinction: the Lowermost Triassic microbialites.
Facies., 36, 238–242.
Baud, A., Richoz, S. and Marcoux, J. (2005) Calcimicrobial
cap rocks from the basal Triassic units: western Taurus
occurrences (SW Turkey). C R Palevol, 4, 569–582.
Beatty, T.W., Zonnefeld, J.-P. and Henderson, C.M. (2008)
Anomalously diverse Early Triassic ichnofossil assemblages
in northwest Pangea: a case for a shallow-marine habitable
zone. Geology, 36, 771–774.
Burne, R.V. and Moore, L.S. (1987) Microbialites: organose-
dimentary deposits of benthic microbial communities. Pal-
aios, 2, 241–254.
Calner, M. (2005) A Late Silurian extinction event and
anachronistic period. Geology, 33, 305–308.
Crasquin, S., Baud, A., Kershaw, S., Richoz, S., Kosun, E. and
Forel, M.B. (2009) The Permian-Triassic transition in the
754 S. Kershaw et al.
� 2010 The Authors. Journal compilation � 2010 International Association of Sedimentologists, Sedimentology, 58, 739–755

Southwestern Taurus Mountains (South Turkey). In: IGCP
572 annual Meeting & Field Workshop in southern Turkey,
Antalya, Sept. 2–6, 2009. (Ed. A. Baud), Field Guide Book.
48 pp.
Crasquin-Soleau, S. and Kershaw, S. (2005) Ostracod fauna
from the Permian-Triassic boundary interval of South China
(Huaying Mountains, eastern Sichuan Province): palaeo-
environmental significance. Palaeogeogr. Palaeoclimatol.Palaeoecol., 217, 131–141.
Crasquin-Soleau, S., Broutin, J., Besse, J. and Berthelin, M.(2001) Ostracodes and palaeobotany from the Middle
Permian of Oman: implications on Pangea reconstruction.
Terra Nova, 13, 38–43.
Dolenc, T., Lojen, S. and Ramovs, A. (2001) The Permian-
Triassic boundary in western Slovenia (Idrijca Valley sec-
tion): magnetostratigraphy, stable isotopes and elemental
variations. Chem. Geol., 175, 175–190.
Erwin, D.H. (2006) Extinction. Princeton University Press,
Princeton and Oxford, 296 pp.
Flugel, E. (2004) Microfacies of Carbonate Rocks: Analysis
Interpretation and Application. Springer, Berlin, 976 pp.
Forel, M.-B., Crasquin, S., Kershaw, S., Feng, Q.L. and Collin,P.-Y. (2009) Ostracods (Crustacea) and water oxygenation in
the earliest Triassic of South China: implications for oceanic
events at the end-Permian mass extinction. Aust. J. Earth
Sci., 56, 815–823.
Groves, J.R. and Calner, M. (2004) Lower Triassic oolites in
Tethys: a sedimentologic response to the end-Permian mass
extinction. Geol. Soc. America Abstracts with Programs,
36, 336.
Hips, K. and Haas, J. (2006) Calcimicrobial stromatolites at the
Permian-Triassic boundary in a western Tethyan section,
Bukk Mountains, Hungary. Sed. Geol., 185, 239–253.
Kempe, S., Kazmierczak, J., Landmann, G., Konuk, T., Rei-mer, A. and Lipp, A. (1991) Largest known microbialites
discovered in Lake Van, Turkey. Nature, 349, 605–608.
Kershaw, S., Zhang, T. and Lan, G. (1999) A? microbialite
crust at the Permian-Triassic boundary in south China, and
its palaeoenvironmental significance. Palaeogeogr. Palaeo-
climatol. Palaeoecol., 146, 1–18.
Kershaw, S., Li, Y., Crasquin-Soleau, S., Feng, Q., Mu, X.,Collin,P.-Y., Reynolds, A. and Guo, L. (2007) EarliestTriassic
microbialites in the South China block and other areas: con-
trols on their growth and distribution. Facies, 53, 409–425.
Kidder, D.L. and Worsley, T.R. (2004) Causes and conse-
quences of extreme Permo-Triassic warming to globally
equable climate and relation to the Permo-Triassic extinc-
tion and recovery. Palaeogeogr. Palaeoclimatol. Palaeoecol.,203, 207–237.
Laval, B., Cady, S.L., Pollack, J.C., McKay, C.P., Bird, J.S.,Grotzinger, J.P., Ford, D.C. and Bohm, H.R. (2000) Modern
freshwater microbialite analogues for ancient dendritic reef
structures. Nature, 407, 626–629.
McNamara, K. (2009) Stromatolites. Geoscientist, 19, 16–22.
Reid, R.P., Visscher, P.T., Decho, A.W., Stolz, J.F., Bebout,B.M., Dupraz, C., MacIntyre, I.G., Paerl, H.W., Pinckney,J.L., Prufert-Bebout, L., Steppe, T.F. and DesMarais, D.J.(2000) The role of microbes in accretion, lamination, and
early lithification of modern marine stromatolites. Nature,
406, 989–992.
Richoz, S. (2006) Stratigraphie et variations isotopiques du
Carbon dans le Permien superieur et le Trias inferieur de la
Neotethys (Turquie, Oman et Iran). Memoires de Geologie(Lausanne), 46, 264.
Riding, R. (2000) Microbial carbonates: the geological record
of calcified bacterial-algal mats and biofilms. Sedimentol-ogy, 47(Suppl. 1), 179–214.
Riding, R. (2005) Phanerozoic reefal microbial carbonate
abundance: comparisons with metazoan diversity, mass
extinction events, and seawater saturation state. Rev. Esp.Micropaleontol., 37, 23–39.
Riding, R. (2008) Abiogenic, microbial and hybrid authigenic
carbonate crusts: components of Precambrian stromatolites.
Geologia Croatica, 61, 73–103.
Wang, W., Kano, A., Okumura, T., Ma, Y., Matsumoto, R.,Matsuda, N., Ueno, K., Chen, X., Kakuwa, Y., Gharaie,M.H.M. and Ilkhchi, M.R. (2007) Isotopic chemostratigra-
phy of the microbialite-bearing Permian–Triassic boundary
section in the Zagros Mountains, Iran. Chem. Geol., 244,708–714.
Wignall, P.B. and Twitchett, R.J. (1996) Ocean anoxia and the
end-Permian mass extinction. Science, 272, 1155–1158.
Manuscript received 21 October 2009; revisionaccepted 11 June 2010
Earliest Triassic microbialites in southern Turkey 755
� 2010 The Authors. Journal compilation � 2010 International Association of Sedimentologists, Sedimentology, 58, 739–755




















