Dynamic digestive responses to increased energy demands in the leaf-eared mouse (Phyllotis darwini)
6
ORIGINAL PAPER Daniel E. Naya Leonardo D. Bacigalupe Diego M. Bustamante F. Bozinovic Dynamic digestive responses to increased energy demands in the leaf-eared mouse (Phyllotis darwini ) Accepted: 21 September 2004 / Published online: 23 November 2004 Ó Springer-Verlag 2004 Abstract A major area of interest in comparative phys- iology has been to understand how animals cope with changing environmental demands in time and space. The digestive system has been identified as one of the more sensitive systems to changes in environmental condi- tions. However, most research on this topic has evalu- ated these effects during peak energetic demands, which do not allow for evaluation of the dynamics of the digestive response along a more natural continuous gradient of environmental conditions. We examined phenotypic flexibility in digestive responses of the leaf- eared mouse Phyllotis darwini to increments in total energy demands (via sequential exposure to 26, 12 and 0°C). Additionally, we evaluated the effect of a moderate energy demand (12°C) over three different time periods (7, 17 and 27 days) on digestive traits. Moderate in- creases in energy demand were associated with changes in the distribution of digesta in the gut, whereas higher increases in energy demand involved increases in the tissue mass of digestive organs. Time-course analysis showed that at 12°C practically all digestive variables reached stable values within 7 days, which is in agree- ment with empirical data and theoretical deductions from cellular turnover rates. We conclude that although the input of energy and nutrients into the digestive tract is typically periodic, many aspects of digestive physiol- ogy are likely to be flexible in response to environmental variability over both short-term (daily) and long-term (seasonal) time scales. Keywords Digestive system Gastrointestinal tract Gut morphology Phenotypic flexibility Phyllotis darwini Abbreviations DDMI: Digestible dry-matter intake DM: Dry mass DMD: Apparent dry-matter digestibility E: Faecal output HD: High demand I: Food intake MD: Moderate demand N: Nitrogen RCC: Relative amount of digesta in a digestive organ T: Thermoneutrality Ta: Ambient temperature TT: Turnover time V: Total amount of digesta in the gut Introduction Studies of phenotypic flexibility are central to under- standing contemporary evolutionary physiology. Theo- retically, flexibility is heritable and appears to evolve through natural selection (Scheiner 2002). A working example is reversible phenotypic flexibility (sensu Stearns 1989), which is the ability to modulate organis- mal traits in response to environmental conditions (Pi- ersma and Drent 2003). Thus, the environmental modification of an organism’s physiology and mor- phology is often hypothesized to be a response to changing conditions, which increases biological perfor- mance. These adjustments may include changes in the rates of energy acquisition (e.g. foraging, digestion and absorption), and energy expenditure (e.g. metabolic heat production and tissue growth) (Bacigalupe and Bozi- novic 2002). Over the last two decades, digestive phenotypic flex- ibility has been extensively investigated in many verte- brate species (for reviews, see Piersma and Lindstrom 1997; Starck 1999; McWilliams and Karasov 2001; Naya and Bozinovic 2004). These studies demonstrated that: (1) A particular genotype has the potential to produce different digestive phenotypes in response to variation in food quality, quantity and/or to the energetic demands of organisms (Piersma and Lindstrom 1997; Starck Communicated by I.D Hume D. E. Naya L. D. Bacigalupe D. M. Bustamante F. Bozinovic (&) Center for Advanced Studies in Ecology and Biodiversity, and Departamento de Ecologı´a, Facultad de Ciencias Biolo´gicas, Pontificia Universidad Cato´lica de Chile, 6513677 Santiago, Chile E-mail: [email protected] Tel.: +56-2-3542618 Fax: +56-2-3542621 J Comp Physiol B (2005) 175: 31–36 DOI 10.1007/s00360-004-0459-8
-
Upload
independent -
Category
Documents
-
view
6 -
download
0
Transcript of Dynamic digestive responses to increased energy demands in the leaf-eared mouse (Phyllotis darwini)

ORIGINAL PAPER
Daniel E. Naya Æ Leonardo D. Bacigalupe
Diego M. Bustamante Æ F. Bozinovic
Dynamic digestive responses to increased energy demandsin the leaf-eared mouse (Phyllotis darwini )
Accepted: 21 September 2004 / Published online: 23 November 2004� Springer-Verlag 2004
Abstract A major area of interest in comparative phys-iology has been to understand how animals cope withchanging environmental demands in time and space. Thedigestive system has been identified as one of the moresensitive systems to changes in environmental condi-tions. However, most research on this topic has evalu-ated these effects during peak energetic demands, whichdo not allow for evaluation of the dynamics of thedigestive response along a more natural continuousgradient of environmental conditions. We examinedphenotypic flexibility in digestive responses of the leaf-eared mouse Phyllotis darwini to increments in totalenergy demands (via sequential exposure to 26, 12 and0�C). Additionally, we evaluated the effect of a moderateenergy demand (12�C) over three different time periods(7, 17 and 27 days) on digestive traits. Moderate in-creases in energy demand were associated with changesin the distribution of digesta in the gut, whereas higherincreases in energy demand involved increases in thetissue mass of digestive organs. Time-course analysisshowed that at 12�C practically all digestive variablesreached stable values within 7 days, which is in agree-ment with empirical data and theoretical deductionsfrom cellular turnover rates. We conclude that althoughthe input of energy and nutrients into the digestive tractis typically periodic, many aspects of digestive physiol-ogy are likely to be flexible in response to environmentalvariability over both short-term (daily) and long-term(seasonal) time scales.
Keywords Digestive system Æ Gastrointestinal tract ÆGut morphology Æ Phenotypic flexibility Æ Phyllotisdarwini
Abbreviations DDMI: Digestible dry-matter intake ÆDM: Dry mass Æ DMD: Apparent dry-matterdigestibility Æ E: Faecal output Æ HD: High demand ÆI: Food intake Æ MD:Moderate demand Æ N: Nitrogen ÆRCC: Relative amount of digesta in a digestiveorgan Æ T: Thermoneutrality Æ Ta: Ambienttemperature Æ TT: Turnover time Æ V: Total amountof digesta in the gut
Introduction
Studies of phenotypic flexibility are central to under-standing contemporary evolutionary physiology. Theo-retically, flexibility is heritable and appears to evolvethrough natural selection (Scheiner 2002). A workingexample is reversible phenotypic flexibility (sensuStearns 1989), which is the ability to modulate organis-mal traits in response to environmental conditions (Pi-ersma and Drent 2003). Thus, the environmentalmodification of an organism’s physiology and mor-phology is often hypothesized to be a response tochanging conditions, which increases biological perfor-mance. These adjustments may include changes in therates of energy acquisition (e.g. foraging, digestion andabsorption), and energy expenditure (e.g. metabolic heatproduction and tissue growth) (Bacigalupe and Bozi-novic 2002).
Over the last two decades, digestive phenotypic flex-ibility has been extensively investigated in many verte-brate species (for reviews, see Piersma and Lindstrom1997; Starck 1999; McWilliams and Karasov 2001; Nayaand Bozinovic 2004). These studies demonstrated that:(1) A particular genotype has the potential to producedifferent digestive phenotypes in response to variation infood quality, quantity and/or to the energetic demandsof organisms (Piersma and Lindstrom 1997; Starck
Communicated by I.D Hume
D. E. Naya Æ L. D. Bacigalupe Æ D. M. BustamanteF. Bozinovic (&)Center for Advanced Studies in Ecology and Biodiversity,and Departamento de Ecologıa, Facultad de Ciencias Biologicas,Pontificia Universidad Catolica de Chile,6513677 Santiago, ChileE-mail: [email protected].: +56-2-3542618Fax: +56-2-3542621
J Comp Physiol B (2005) 175: 31–36DOI 10.1007/s00360-004-0459-8

1999); (2) There is a trade-off between the benefits ofprocessing food through a digestive system with specificattributes (e.g. gut mass or length), and the cost ofmaintaining and carrying it (Moss 1974; Sibly 1981;Diamond and Karasov 1983; Toloza et al. 1991); (3)Digestive phenotypic flexibility can arise at differentorganizational levels (from molecules to organs),namely, hydrolytic enzyme activity (Martınez del Rıoet al. 1995; Sabat et al. 1995; Sabat and Bozinovic 2000),brush-border nutrient transport density and activity(Karasov et al. 1983; Toloza et al. 1991; Konarzewskiand Diamond 1994) and gut surface area, mass andlength (Gross et al. 1985; Green and Millar 1987; Bo-zinovic et al. 1988; Hammond and Wunder 1991; Tolozaet al. 1991; Sabat and Bozinovic 2000). In addition,flexibility can also imply changes in food-retention time(Derting and Bogue 1993; Lopez-Calleja et al. 1997;Levey and Martinez del Rıo 1999) and food distributionin digestive regions (Loeb et al. 1991; Dykstra and Ka-rasov 1992; Levey and Duke 1992); (4) Although re-sponses at different levels can interact in complex ways,current information supports the idea of a sequence ofcompensatory digestive changes (Derting and Bogue1993; Nagy and Negus 1993).
Studies conducted in small mammals have demon-strated that cold acclimation results in higher rates offood intake, and hypertrophy of food processing organs(Toloza et al. 1991; McDevitt and Speakman 1994;Hammond and Wunder 1995). However, most researchon this topic has evaluated these effects at two highlycontrasting temperatures, usually under extreme condi-tions (but see Green and Millar 1987; Hammond et al.1999). The use of extreme experimental settings does notallow for the evaluation of the dynamics of the digestiveresponse of animals as they cope with moderate de-mands along a more natural continuous gradient ofenvironmental conditions. In the present work, weexamined phenotypic flexibility in digestive responses ofthe leaf-eared mouse, Phyllotis darwini, to increments intotal energy demands. For this purpose, we sequentiallyexposed individuals to 26, 12 and 0�C, and analysedchanges in food processing (i.e. transit time and thedistribution of digesta along the gastrointestinal tract)and digestive organ tissue mass. We hypothesized, fol-
lowing the economy principle of Sibly (1981), thatmoderate demands can be met by changes in food pro-cessing, while at higher energetic demands additionalchanges in digestive organ tissue mass are necessary.
Materials and methods
Animals and experimental design
The leaf-eared mouse, P. darwini (Sigmodontinae), is anocturnal rodent that feeds primarily on seeds, but alsoconsumes insects and grasses (Meserve 1981). It inhabitssemi-arid and Mediterranean ecosystems of northernand central Chile, from the Pacific Ocean coast to theAndes Mountains (ca. 2,000 m above sea level). Theseenvironments are characterized by marked seasonalvariation (warm dry summers, cold wet winters), andalso by daily fluctuation in abiotic conditions (Nespoloet al. 2001). Thus, P. darwini regularly experienceschanges in food intake and quality on both daily andseasonal time scales, which makes this species a goodchoice for studying how the digestive system of a smallmammal responds to changes in food intake over dif-ferent time scales.
Fifty newborn P. darwini (27 males and 23 females)were selected from a captive population that we hadmaintained for 2 years. All animals were maintained atan ambient temperature (Ta) of 26±2�C, with a pho-toperiod of 14L:10D. Individuals were fed commercialrabbit pellets (Champion) and water ad libitum for3 months (i.e. until adulthood). Diet pellets were anal-ysed at the Institute of Nutrition and Food Technology,Universidad de Chile, following proximate chemicalanalysis methods (Horwitz 1980). The average chemicalcomposition was: dry matter 90.6%, ash 10.8%, crudefibre 16.5%, neutral detergent fibre 37.8%, acid-deter-gent fibre 19.8%, lipid 3.0%, protein (N·6.25) 20.0,available carbohydrate 40.3% and gross energy content18.4±0.5 kJ g�1 (n=3).
The experimental design is shown in Fig. 1. Theexperiment began with 50 individuals, which weresequentially exposed to a decrease in environmentaltemperature (26, 12 and 0�C). On different days (seeFig. 1), ten individuals randomly assigned were sepa-rated from the rest to generate one of the followingexperimental treatments: (1) T (thermoneutrality),7 days at Ta=26±2�C; (2) MD1 (moderate demand),7 days at Ta=12±2�C; (3) MD2 (moderate demand),17 days at Ta=12±2�C; (4) MD3 (moderate demand),
Fig. 1 The experimental design. The experiment began with 50individuals, which were sequentially exposed to lower environmen-tal temperatures. At days 7, 14, 24, 34 and 41, 10 individuals wereseparated from the rest to generate the five experimental groups:26�C (T group), 12�C (three MD groups) and 0�C (HD group)
32

27 days at Ta=12±2�C; and (5) HD (high demand),7 days at Ta=0±2�C after 27 days of acclimation atTa=12±2�C. The photoperiod was kept at 12L:12Dthroughout the study.
Digestive variables
Food intake (I) and faecal output (F) were estimated dailyfor the last 3 days of the experimental period, by placingeach animal individually in a cage with an elevated wirebottom. Faeces and remaining food dropped to the cagebottom and were dried at 60�C for at least 96 h to con-stant mass, and then separated from each other. Thedifference between dry food offered and dry food residuesis dry matter intake. Body mass (bm) of each animal wasmeasured once prior to the I and F estimation (initial bm)and again after the 3-day collection period (final bm). Allmeasurements were taken between 0800 and 0900 hours.Mice were sacrificed by cervical dislocation between 1400and 1800 hours, and their digestive organs (stomach,small intestine, caecum and colon) were removed. Theamount of digesta (the material rinsed out of the intactgut), was dried and weighed separately for each organ onan electronic analytical balance (Chyo JK-180,±0.0001 g), then integrated over the entire gut (V).Digestive organs were washed with Ringer’s solution,dried to constant mass at 80�C for 7 days, then re-weighed. From these data, three kinds of digestive vari-ables were estimated: (1) overall response variables:digestible dry matter intake [DDMI (g day�1)=I(g day�1)�F (g day�1)], and apparent dry-matterdigestibility {DMD (%)=[I (g day�1)�F (g day�1)]/[I(g day�1)]}; (2) digesta processing variables: totalamount of digesta in the gut [V (g)], and the relativeamount of digesta in each digestive organ {RCC i(%)=[V of the organ ‘‘i’’(g)/V (g)]·100}, total turnovertime {TTt (h)=[V (g)/I (g day�1)]·24}, and turnover timeof absorptive organs {TTac (h)=[V of absorptive organs(g)/I (g day�1)]·24}; see Hammond and Wunder (1991)for methodological assumptions; (3) anatomical vari-ables: dry tissue mass of each digestive organ [DM i (g)].
Statistical analysis
Differences in initial and final bm between groups wereevaluated using one-way ANOVA. Changes in alldependent variables were evaluated separately by AN-COVA. We used final bm for overall response anddigesta processing variables, or carcass mass (i.e. vis-cera-free body mass) for anatomical variables, as thecovariates. Prior to all statistical analyses, data wereexamined for assumptions of normality and homoge-neity of variance, using Kolmogorov–Smirnov andLevene tests, respectively. Interactions between covari-ates and factors (parallelism test) were also tested. Whennecessary to meet assumptions, variables weretransformed to the inverse (bm balances), to logarithm
(total turnover time) or to arcsine (dry-matter digest-ibility). We carried out post hoc comparisons throughTukey HSD tests. Statistical analyses were performedusing the statistical package, STATISTICA (2001) ver-sion 6.0 for the Windows operating system.
Results
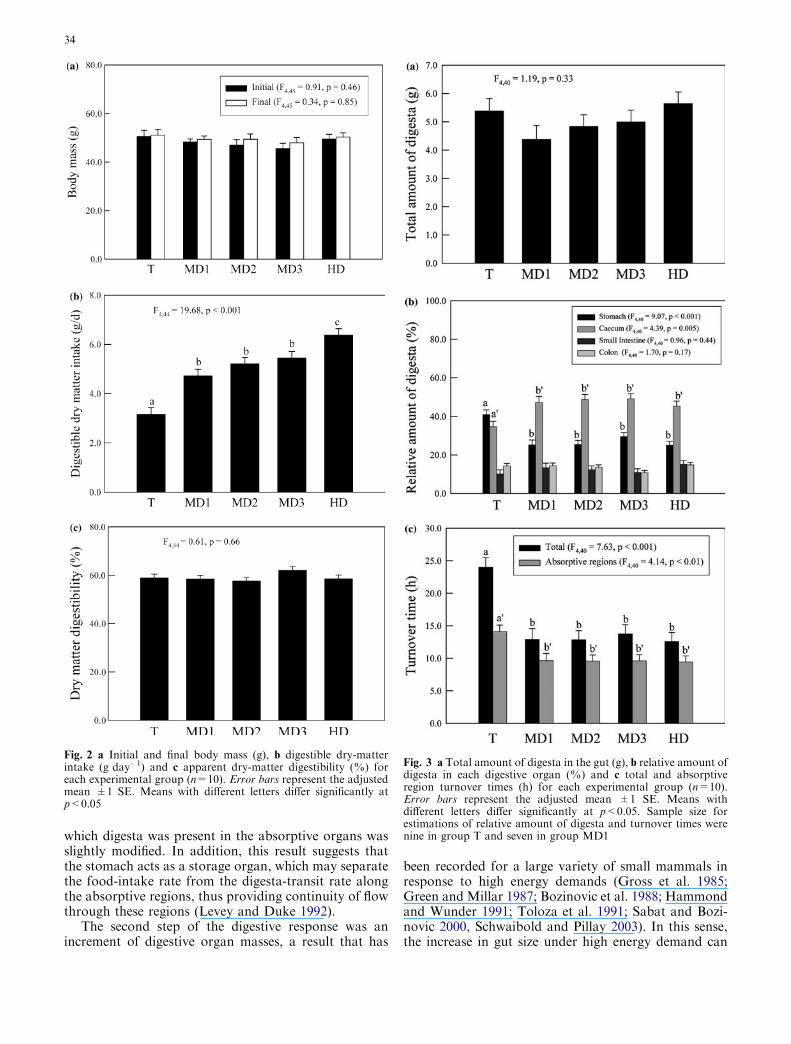
There were no significant differences in initial or final bmbetween groups (Fig. 2a). Digestible dry matter intake(DDMI) increased significantly, but dry matter digest-ibility (DMD) remained constant with decrease inexperimental Ta (Fig. 2b, c). The total amount of di-gesta in the gut did not differ between groups (Fig. 3a).However, individuals exposed to low Ta had the greatestproportion of digesta in the caecum, while at high Ta thegreatest proportion of digesta was in the stomach(Fig. 3b). The proportion of digesta in the small intes-tine and colon was not affected by Ta (Fig. 3b). Bothtotal and absorptive organs turnover times decreased inindividuals moved from high (26�C) to low (12 and 0�C)Ta, but the difference was markedly less for the TT ofabsorptive regions (14 vs. 9 h) than for total gut TT (24vs. 12 h) (Fig. 3c). Digestive organs followed a similarpattern of variation: individuals at 0�C had significantlyhigher digestive organ tissue mass than at the other twoTa treatments, while there were no differences betweenthe latter two groups (Fig. 4). However, tissue mass ofthe caecum and colon tended to increase along theexperimental treatments more gradually than stomachand small intestine tissue mass. In this sense, for the firsttwo digestive organs, animals at moderate energy de-mands (12�C for 27 days) did not differ from animals intheir thermoneutral zone, nor from animals underhigher energy demands (Fig. 4).
Discussion
A major area of interest in comparative physiology hasbeen to understand how animals cope with changingenvironmental demands in time and space. In this study,we found that the leaf-eared mouse displays a progres-sive sequence of compensatory digestive changes toincreasing energy demands. In particular, moderateincrements in energetic demand were met with changesin digesta processing, while higher increments wereassociated with an additional increase in the tissue massof digestive organs (Fig. 5). Adjustments in digestaprocessing included a re-distribution of digesta along thegastrointestinal tract, without changes in total gut con-tent. In this sense, from thermoneutrality to 12�C therelative content of the caecum increased by about 15%,while the relative stomach content decreased by a similarmagnitude. This redistribution means that, despite thetypical reduction in TT concomitant with an increase infood intake (Hammond and Wunder 1991; Toloza et al.1991; Dykstra and Karasov 1992), the period during
33
which digesta was present in the absorptive organs wasslightly modified. In addition, this result suggests thatthe stomach acts as a storage organ, which may separatethe food-intake rate from the digesta-transit rate alongthe absorptive regions, thus providing continuity of flowthrough these regions (Levey and Duke 1992).
The second step of the digestive response was anincrement of digestive organ masses, a result that has
been recorded for a large variety of small mammals inresponse to high energy demands (Gross et al. 1985;Green and Millar 1987; Bozinovic et al. 1988; Hammondand Wunder 1991; Toloza et al. 1991; Sabat and Bozi-novic 2000, Schwaibold and Pillay 2003). In this sense,the increase in gut size under high energy demand can
Fig. 2 a Initial and final body mass (g), b digestible dry-matterintake (g day�1) and c apparent dry-matter digestibility (%) foreach experimental group (n=10). Error bars represent the adjustedmean ±1 SE. Means with different letters differ significantly atp<0.05
Fig. 3 a Total amount of digesta in the gut (g), b relative amount ofdigesta in each digestive organ (%) and c total and absorptiveregion turnover times (h) for each experimental group (n=10).Error bars represent the adjusted mean ±1 SE. Means withdifferent letters differ significantly at p<0.05. Sample size forestimations of relative amount of digesta and turnover times werenine in group T and seven in group MD1
34

yield several benefits to an animal, manifested as anincrease in the amount of energy ingested in any givenperiod of time (by increasing the total number oftransporters in the gut lining) or by allowing food-retention time to remain constant with increased foodintake (Hammond and Wunder 1991).
In small endotherms, daily, seasonal and spatialvariations in temperature may set limits on the use ofboth space and time (Bacigalupe et al. 2003). In itsnatural habitat, the leaf-eared mouse faces changes inenergy demands (i.e. Ta) at two different time scales:moderate daily fluctuations, and high seasonal fluctua-tions. We suggest that adjustments in digestive pro-cessing are sufficient to allow these rodents to cope withshort-term fluctuations, but increases in digestive organmass are necessary to cope with seasonal changes inenvironmental thermal demands (Fig. 5). However, it
should be noted that intermediate levels of flexibilitythat were not analysed in the present work, such asdigestive enzymes and brush-border transporter activity,may also play an important role in the observed overallresponse variables (Nagy and Negus 1993).
Digestive adjustments in the leaf-eared mouse, atboth moderate and high energetic demands includedchanges in both content and tissue mass of the caecum,suggesting an increase in fermentative activity. Thiscompensatory mechanism has been observed when sev-eral small mammal species were changed to poor-qualitydiets (Gross et al. 1985; Bozinovic et al. 1997; Pei et al.2001) or to low environmental temperatures (Hammondand Wunder 1995; Nespolo et al. 2002). It would beinteresting to test whether this potential increase offermentative activity is followed by any incidence ofcoprophagy, a behaviour not yet described in P. darwini.
Finally, time-course analyses indicate that, at 12�C,all digestive variables reached stable values within7 days and then remained constant. This observation,together with the increase in the tissue mass of somedigestive organs when animals were exposed to 0�C for7 days, indicate that 1 week is long enough for a re-sponse in most of the variables under study. These re-sults are in agreement with both empirical data (e.g.Konarzewski and Diamond 1994) and theoreticaldeductions from cellular turnover rates (Starck 1999).
The basic structural and functional digestive designof an animal, as well as its behaviour and the environ-mental demands it faces, determine the limits to the foodresources that can be utilized. Although the input ofenergy and nutrients into the digestive tract is typicallyperiodic, it is clear that many aspects of digestivephysiology are likely to be flexible as a function of theenvironment. Timing and efficiency of digestion aretherefore dependent on a dynamic balance between foodingestion and digesta distribution among regions of the
Fig. 4 Dry tissue mass of stomach, small intestine, caecum andcolon for each experimental group (n=10). Error bars represent theadjusted mean ± 1 SE. Means with different letters differsignificantly at p<0.05
Fig. 5 Sequence ofcompensatory digestive changesdisplayed by Phyllotis darwiniwhen individuals weresequentially exposed to 26, 12and 0�C. Moderate incrementsin energy demand (e.g. naturaldaily fluctuations intemperature) were met bychanges in digesta processing,while higher increments inenergy demand (e.g. naturalseasonal fluctuations intemperature) were met byincreases in the tissue mass ofdigestive organs
35

gut. As we show here, digestive variables are likely to beflexible, allowing animals to match their performance tochanges in energy demand on both a daily and a sea-sonal basis.
Acknowledgements Ian Hume provided valuable comments. DENand DMB acknowledge fellowships from the Direccion General dePosgrado de la Pontificia Universidad Catolica de Chile (DIPUC,Chile) and Comision Nacional de Investigacion Cientıfica y Tec-nologica (CONICYT, Chile). LDB is a post-doc fellow from theCenter for Advances Studies in Ecology and Biodiversity (CASEB,Program 1). This work was funded by Fondo Nacional de Cienciay Tecnologıa FONDAP 1501-0001 (Program 1). All experimentswere conducted according to current Chilean law, and under permitSAG 698. This paper is dedicated to the memory of Dr MarioRosenmann.
References
Bacigalupe LD, Bozinovic F (2002) Design, limitation and sus-tained metabolic rate: lessons from small mammals. J Exp Biol205:2963–2970
Bacigalupe LD, Rezende EL, Kenagy GJ, Bozinovic F (2003)Activity and space use by degus: a trade-off between thermalconditions and food availability? J Mammal 84:311–318
Bozinovic F, Veloso C, Rosenmann M (1988) Cambios del tractodigestivo de Abrothrix andinus (Cricetidae): efecto de la calidadde dieta y requerimientos de energıa. Rev Chil Hist Nat 61:245–251
Bozinovic F, Novoa F, Sabat P (1997) Feeding and digesting fiberand tannins by the Herbivorous rodent, Octodon degus (Rod-entia: Caviomorpha). Comp Biochem Physiol 118A:625–630
Derting TL, Bogue BA (1993) Responses of the gut to moderateenergy demands in a small herbivore (Microtus pennsylvanicus).J Mammal 74:59–68
Diamond JM, Karasov WH (1983) Trophic control of the intesti-nal mucosa. Nature 304:18
Dykstra CR, Karasov WH (1992) Changes in gut structure andfunction of house wrens (Troglodytes aedon) in response to in-creased energy demands. Physiol Zool 65:422–442
Green DA, Millar JS (1987) Changes in gut dimensions andcapacity of Peromyscus maniculatus relative to diet quality andenergy needs. Can J Zool 65:2159–2162
Gross JE, Wang Z, Wunder BA (1985) Effects of food quality andenergy needs: changes in gut morphology and capacity of Mi-crotus ochrogaster. J Mammal 66:661–667
Hammond KA, Wunder BA (1991) The role of diet quality andenergy need in the nutritional ecology of a small herbivore,Microtus ochrogaster. Physiol Zool 64:541–567
Hammond KA., Wunder BA (1995) Effect of cold temperatures onthe morphology of gastrointestinal tracts of two microtine ro-dents. J Mammal 76:232–239
Hammond KA, Roth J, Janes DN, Dohm MR (1999) Morpho-logical and physiological responses to altitude in deer micePeromyscus maniculatus. Physiol Biochem Zool 72:613–622
Horwitz W (1980) Official methods of analysis. Association ofOfficial Analytical Chemists, Washington, DC
Karasov WH, Pond RS III, Solberg DH, Diamond J (1983) Reg-ulation of proline and glucose transport in mouse intestine bydietary substrate levels. Proc Natl Acad Sci USA 80:7674–7677
Konarzewski M, Diamond J (1994) Peak sustained metabolic rateand its individual variation in cold-stressed mice. Physiol Zool67:1186–1212
Levey DJ, Duke GE (1992) How do frugivores process fruit?Gastrointestinal transit and glucose absorption in cedar wax-wings (Bombycilla cedrorum). Auk 109:722–730
Levey DJ, Martınez del Rıo C (1999) Test, rejection, and refor-mulation of a chemical reactor-based model of gut function in afruit-eating bird. Physiol Biochem Zool 72:369–383
Loeb SC, Schwab RG, Demment MW (1991) Responses of pocketghophers (Thomomys bottae) to changes in diet quality. Oeco-logia 86:542–551
Lopez-Calleja MV, Bozinovic F, Martınez del Rıo C (1997) Effectsof sugar concentration on hummingbird feeding and energy use.Comp Biochem Physiol 118A:1291–1299
Martınez del Rıo C, Brugger KE, Rıos JL, Vergara ME, Witmer M(1995) An experimental and comparative study of dietarymodulation of intestinal enzymes in European starlings (Stur-nus vulgaris). Physiol Zool 68:490–511
McDevitt RM, Speakman JR (1994) Central limits to sustainablemetabolic rate have no role in a cold acclimation of the short-tailed field vole (Microtus agrestis). Physiol Zool 67:1117–1139
McWilliams SR, Karasov WH (2001) Phenotypic flexibility indigestive system structure and function in migratory birds andits ecological significance. Comp Biochem Physiol 128A:579–593
Meserve PL (1981) Trophic relationship among small mammals ina Chilean semiarid thorn scrub community. J Mammal 62:304–314
Moss R (1974) Winter diets, gut lengths, and interespecific com-petition in Alaskan Ptarmigan. Auk 91:737–746
Nagy TR, Negus NC (1993) Energy acquisition and allocation inmale collared lemmings (Dicrostonyx groenlandicus): effects ofphotoperiod, temperature, and diet quality. Physiol Zool66:537–560
Naya DE, Bozinovic F (2004) Digestive phenotypic flexibility inpost-metamorphic amphibians: studies on a model organism.Biol Res 37:365–370
Nespolo RF, Opazo JC, Bozinovic F (2001) Thermal acclimationand non-shivering thermogenesis in three species of SouthAmerican rodents: a comparison between arid and mesic hab-itats. J Arid Environ 48:581–590
Nespolo RF, Bacigalupe LD, Sabat P, Bozinovic F (2002) Inter-play among energy metabolism, organ mass and digestive en-zyme activity in the mouse-opossum Thylamys elegans: the roleof thermal acclimatation. J Exp Biol 205:2697–2703
Pei Y-X, Wang D-H, Hume ID (2001) Selective digesta retentionand coprophagy in Brandt’s vole (Microtus brandti). J CompPhysiol B 171:457–464
Piersma T, Drent J (2003) Phenotypic flexibility and the evolutionof organismal design. Trends Ecol Evol 18:228–233
Piersma T, Lindstrom A (1997) Rapid reversible changes in organsize as a component of adaptative behaviour. Trends Ecol Evol12:134–138
Sabat P, Bozinovic F (2000) Digestive plasticity and the cost ofacclimation to dietary chemistry in the omnivorous leaf-earedmouse Phyllotis darwini. J Comp Physiol B 170:411–417
Sabat P, Bozinovic F, Zambrano F (1995) Role of dietary sub-strates on intestinal disaccharidases, digestibility and energeticsin the insectivorous mouse-opossum (Thylamys elegans).J Mammal 76:603–611
Scheiner SM (2002) Selection experiments and the study of phe-notypic plasticity. J Evol Biol 15:889–898
Schwaibold U, Pillay N (2003) The gut morphology of the Africanice rat, Otomys sloggetti robertsi, shows adaptation to coldenvironments and sex-specific season variation. J Comp PhysiolB 173:653–659
Sibly RM (1981) Strategies of digestion and defecation. In: Tow-send CR, Calow P (eds) Physiological ecology: an evolutionaryapproach to resource use. Blackwell, Oxford, pp 109–139
Starck JM (1999) Structural flexibility of the gastro-intestinal tractof vertebrates—implications for evolutionary morphology.Zool Anz 238:87–101
Stearns C (1989) The evolutionary significance of phenotypicplasticity. Bioscience 39:436–445
Toloza E, Lam M, Diamond J (1991) Nutrient extraction by cold-exposed mice: a test of digestive safety margins. Am J Physiol261:G608–G620
36