
Duplication of the Bx7 high-molecular-weight glutenin subunit gene in bread wheat (Triticum aestivum...
8
Plant Breeding 116. 525 -531 (1997) r 1997 Blackwell Wissenschafts-Verlag. Berlin ISSN 0179-9541 Duplication of the Bx7 high-molecular-weight glutenin subunit gene in bread wheat {Triticum aestivum L.) cultivar 'Red River 68' R- D'OviDio', S. MASCI', E. PORCEDDU' and D. D. KASARDA- 'Dipiirtitnento di Agrobioiogia e Agrochimica. Universita degli Studi della Tuscia, Via S. Camillo de Lellis, I-Ol IOO Viterbo. Italy; ^US Department of Agriculture, Agricultural Research Serviee, Western Regional Research Center, 800 Buchanan Street, Albany, CA 94710, USA Wilh 4 figures ami 2 tables Received November i. I995jAccepted August 6. 1997 Communicaled by C. O. Qualsei Abstract SDS-PAGE analysis of seed proteins ofthe cuitivar 'Red River 68" showed a considerably higher staining intensily of the band cor- responding to HMW-GS H.K? relative to the equividcni band in the ciiliivars "Chinese Spring' and "Cheyenne". Southern blois iiT restriction en/ynie fragmonls Iroin DNA of these three cultivars were analyzed deiisitomelriciilly to reveal that the band corresponding to the S.vZgene of "Red River 68" had a double staining intensity compared to ihc equivalent bands from the other two cultivars, which indicates thai in •Red River 68" a duplication of the B.\7 gene has oecurred. Although the possibility ofthe gene copy being a pseudogene was not ruled out. the greater amount of protein corresponding to B\7 in "Red River 68' most iikely is in accord with an increase in active gene number. SDS- PAGE analysis ofthe proteins showed iilso that the mobility of B.\7 in 'Cheyenne" was slightly different from the mobilities ofthe fi.v7subunils of 'Red River 68" and "Chinese Spring'. The same dilference was observed at the gene level by PCR amplification ofthe genes encoding these subunits. Key words: Triticum uestinim glutenin stibunits — PCR gene duplication — HMW The bread-making properties of hexaploid bread wheat are strongly infltienced by the high-niolecular-weighi glutenin sub- units (HMW-GS), showing both qualitative variations (result- uig from allclic differences: Payne et al. 1987. 1988) and quantitative variations (Huebncr and Bietz 1985. Lawrence et al. 1988. MacRitchie etal. 1991. Kolster et al.1992) in relation to these subunits. HMW-GS are encoded by genes at the three complex Ghi-l loci (Glu-.4I. Ghi-Bt. Glu-Dl), each locus con- taining two different closely linked genes, designated x- and y- types. Thus, there are six different gene types present in hex- aploid wheats, although one, or even both, of the genes at a locus may not be expressed. For example, neither the x- or ihe y-type genes at the Gtu-At locus of 'Chinese Spring' are expressed. The x and y genes were originally classified according to differenees in their protein products (Payne et al. 1981. 1983. 1984. Shewry et al. 1984) and the various expressed proteins corresponding to these two linked gene types are representative of allelic variation at the complex loci in question. Payne and Lawrence (1983) found that allelic variation at the Gtu-I loci in bread wheats, recognized by the corresponding HMW-GS as defined by potyacrylamide gel electrophoresis (SDS-PAGE), could be grouped into 20 different forms, three at the GIu-AI locus. 11 at the Glu-BJ locus, and 6 at the 67//- Dl locus. Since this pioneering paper, additional alleles have been detected for HMW-GS. Among the HMW-GS encoded at Ghi-BJ.fi.v7is qtiite common, being present in many different bread and durum wheat cultivars {see Shewry etal. 1992. Look- hart ct al. 1993). It is usually associated with either oftwo By subunits. ByH or By9. Although it is not possible to separate the quality contributions of B.\7 from its corresponding y-type subunit, botb allelic pairs apparently contribute moderately well to good quality. ll is generally considered that each x and each y HMW-GS gene is present in single copy form, but this was never clearly estabhshed. Earlier workers indicated that there may be 1-3 copies of each gene (Thompson et al. 1983, Eorde et al. 1985. Harberd et al. 19fi6). This is in contrast to other loci coding for gluten proteins, where gene copies are much more numerous. For example, Anderson et al. (1990) estimated that the three Gli-2 loci of the cultivar 'Cheyenne' include 50-60 x-gliadin- type gene copies, although about half of these genes are pseudo- genes (not expressed) and some may correspond to a-type glu- tenin subunits (Lew et al. 1992). Although the Gti-l loci include multiple copies ol'genes coding for ;-glladins (and presumably •/"type glutcnins. Masci et al. 1995). there are apparently fewer ;-type gene copies at 07/-/ (unpublished data) than a-type gene copies at the Gli-2 loci. The situation for lo-gliadins is less clear as the corresponding genes at the Gli~! loci have not been studied to the same degree as for other proteins, but protein studies indicate that there are likely to be multiple copies for these genes as well (Kasarda et al. 1983, Lafiandra et al. 1984). Although in SDS-PAGE subunit B\7 usually stains strongly relative to the other HMW-GS when the patterns of different eultivars arc compared (Kolster et al. 1992. Lookhart ct al. 1993), there are also differences in the intensity of staining of B.x7 among cultivars. Also, small dillerenccs in the nucleotide sequences ofthe genes encoding subunit fi.v7 derived from two different cultivars. "Chinese Spring' and "Cheyenne', have been reported (Thompson et al. 1983. Anderson and Greene 1989). Marchylo et al. (1992) found that different cultivars considered to possess subunit B.\7 differ from one another in SDS-PAGE analysis, indicating that at least two different I B.\ proteins exist in the different cultivars — one was designated subunit 7 and the other was designated subunit 7*. They also reported that subunit 7* generally made up about 27% ofthc total amount of HMW-GS in a cultivar. whereas subunit 7 generally made up about 41%, a substantial difference in expression. Sub- sequently, Lukow et al. (1992) reported that in the cultivar US.CopyriBhl Clearance CenlerC,>deStaicmL.-m 0179-9541/97/1606 0525$ 14.00/0
Transcript of Duplication of the Bx7 high-molecular-weight glutenin subunit gene in bread wheat (Triticum aestivum...

Plant Breeding 116. 525 -531 (1997)r 1997 Blackwell Wissenschafts-Verlag. BerlinISSN 0179-9541
Duplication of the Bx7 high-molecular-weight glutenin subunit gene in breadwheat {Triticum aestivum L.) cultivar 'Red River 68'
R- D'OviDio', S. MASCI', E. PORCEDDU' and D. D. KASARDA-
'Dipiirtitnento di Agrobioiogia e Agrochimica. Universita degli Studi della Tuscia, Via S. Camillo de Lellis, I-Ol IOO Viterbo.Italy; ^US Department of Agriculture, Agricultural Research Serviee, Western Regional Research Center,800 Buchanan Street, Albany, CA 94710, USA
Wilh 4 figures ami 2 tables
Received November i. I995jAccepted August 6. 1997Communicaled by C. O. Qualsei
AbstractSDS-PAGE analysis of seed proteins ofthe cuitivar 'Red River 68"showed a considerably higher staining intensily of the band cor-responding to HMW-GS H.K? relative to the equividcni band in theciiliivars "Chinese Spring' and "Cheyenne". Southern blois iiT restrictionen/ynie fragmonls Iroin DNA of these three cultivars were analyzeddeiisitomelriciilly to reveal that the band corresponding to the S.vZgeneof "Red River 68" had a double staining intensity compared to ihcequivalent bands from the other two cultivars, which indicates thai in•Red River 68" a duplication of the B.\7 gene has oecurred. Althoughthe possibility ofthe gene copy being a pseudogene was not ruled out.the greater amount of protein corresponding to B\7 in "Red River 68'most iikely is in accord with an increase in active gene number. SDS-PAGE analysis ofthe proteins showed iilso that the mobility of B.\7 in'Cheyenne" was slightly different from the mobilities ofthe fi.v7subunilsof 'Red River 68" and "Chinese Spring'. The same dilference wasobserved at the gene level by PCR amplification ofthe genes encodingthese subunits.
Key words: Triticum uestinimglutenin stibunits — PCR
gene duplication — HMW
The bread-making properties of hexaploid bread wheat arestrongly infltienced by the high-niolecular-weighi glutenin sub-units (HMW-GS), showing both qualitative variations (result-uig from allclic differences: Payne et al. 1987. 1988) andquantitative variations (Huebncr and Bietz 1985. Lawrence etal. 1988. MacRitchie etal. 1991. Kolster et al.1992) in relationto these subunits. HMW-GS are encoded by genes at the threecomplex Ghi-l loci (Glu-.4I. Ghi-Bt. Glu-Dl), each locus con-taining two different closely linked genes, designated x- and y-types. Thus, there are six different gene types present in hex-aploid wheats, although one, or even both, of the genes at alocus may not be expressed. For example, neither the x- orihe y-type genes at the Gtu-At locus of 'Chinese Spring' areexpressed. The x and y genes were originally classified accordingto differenees in their protein products (Payne et al. 1981. 1983.1984. Shewry et al. 1984) and the various expressed proteinscorresponding to these two linked gene types are representativeof allelic variation at the complex loci in question.
Payne and Lawrence (1983) found that allelic variation atthe Gtu-I loci in bread wheats, recognized by the correspondingHMW-GS as defined by potyacrylamide gel electrophoresis(SDS-PAGE), could be grouped into 20 different forms, threeat the GIu-AI locus. 11 at the Glu-BJ locus, and 6 at the 67//-Dl locus. Since this pioneering paper, additional alleles have
been detected for HMW-GS. Among the HMW-GS encodedat Ghi-BJ. fi.v7 is qtiite common, being present in many differentbread and durum wheat cultivars {see Shewry etal. 1992. Look-hart ct al. 1993). It is usually associated with either oftwo Bysubunits. ByH or By9. Although it is not possible to separatethe quality contributions of B.\7 from its corresponding y-typesubunit, botb allelic pairs apparently contribute moderatelywell to good quality.
ll is generally considered that each x and each y HMW-GSgene is present in single copy form, but this was never clearlyestabhshed. Earlier workers indicated that there may be 1-3copies of each gene (Thompson et al. 1983, Eorde et al. 1985.Harberd et al. 19fi6). This is in contrast to other loci coding forgluten proteins, where gene copies are much more numerous.For example, Anderson et al. (1990) estimated that the threeGli-2 loci of the cultivar 'Cheyenne' include 50-60 x-gliadin-type gene copies, although about half of these genes are pseudo-genes (not expressed) and some may correspond to a-type glu-tenin subunits (Lew et al. 1992). Although the Gti-l loci includemultiple copies ol'genes coding for ;-glladins (and presumably•/"type glutcnins. Masci et al. 1995). there are apparently fewer;-type gene copies at 07/-/ (unpublished data) than a-type genecopies at the Gli-2 loci. The situation for lo-gliadins is less clearas the corresponding genes at the Gli~! loci have not beenstudied to the same degree as for other proteins, but proteinstudies indicate that there are likely to be multiple copies forthese genes as well (Kasarda et al. 1983, Lafiandra et al. 1984).
Although in SDS-PAGE subunit B\7 usually stains stronglyrelative to the other HMW-GS when the patterns of differenteultivars arc compared (Kolster et al. 1992. Lookhart ct al.1993), there are also differences in the intensity of staining ofB.x7 among cultivars. Also, small dillerenccs in the nucleotidesequences ofthe genes encoding subunit fi.v7 derived from twodifferent cultivars. "Chinese Spring' and "Cheyenne', have beenreported (Thompson et al. 1983. Anderson and Greene 1989).Marchylo et al. (1992) found that different cultivars consideredto possess subunit B.\7 differ from one another in SDS-PAGEanalysis, indicating that at least two different I B.\ proteins existin the different cultivars — one was designated subunit 7 andthe other was designated subunit 7*. They also reported thatsubunit 7* generally made up about 27% ofthc total amountof HMW-GS in a cultivar. whereas subunit 7 generally madeup about 41%, a substantial difference in expression. Sub-sequently, Lukow et al. (1992) reported that in the cultivar
US.CopyriBhl Clearance CenlerC,>deStaicmL.-m 0 1 7 9 - 9 5 4 1 / 9 7 / 1 6 0 6 0 5 2 5 $ 14.00/0

526 D'Oviix). MASCI, PoRrppnu and KASARIM
'Glenlea." which showed strong expression of B\7, this subunitwas coded apparently by a single gene dose, pointing towardeither more effective transcription or more efficient translationof the resulting message as responsible for the high level ofexpression, whereas in an Israeli landrace. TAA 36, which alsoshowed strong Bx7 expression, duplication of the Bx7 geneappeared to be responsible.
In research on wheat mixing and baking quality, we hadbeen using "Red River 68" as an example of a cultivar withexceptionally strong mixing characteristics and had been im-pressed by the extremely strong staining of Bx7 in this cultivar.In our experience, the strong staining of 6.v7 in 'Red River 68"is rarely seen in the SDS-PAGE patterns of wheat cultivars.Accordingly, we decided to investigate the possibility that oneor more additional gene copies were involved in coding for thisHMW-GS. We provide evidence here that there is a duplicationof the Bx7 gene(s) in "Red River 68" in comparison to thosepresent in 'Cheyenne' and 'Chinese Spring", containing 7* and7, respectively. The presence of an active gene duplication in aregistered cultivar, as opposed to a landrace (Lukow et al.1992), tnakes it more readily useabte in breeding programsaimed at increasing elasticity and mixing strength.
Materials and MethodsWheat samples: "Red River 68" (CI tr ttutiiber 14193) was obtained tromWorld Seeds. Inc. (conipan> no longer in existence). A major hiotypewas exiraeted (Biotype III after SDS-PAGE iin;ilysis of single seeds(unpublished data). 'Cheyenne" (CI tr number K8K5) grain was ohtainedfrom Dr Virgil A, Johnson (USDA. Lincoln, NE) and 'Chinese Spring'(Cl tr numher 14108) grain was ohtatned from Dr B. Lennarl Johnson(University of California, Riverside, CA). Nullisomic lA-tetrasomic IB'Chinese Spring" was ohtained from Dr E. R. Sears (USDA, ARS,Columbia, MO; deceased). Tetrasomic IB "Chinese Spring' wasohtained from Dr Adam Lukaszewski, University of California, River-side. (CA).
(ilutenin subunits and electrophorelic analyi^is: Glutenin suhunits wereprepared according to the procedure of Singh et al. (1991). hut alky-lation was omitted. The same volumes of the residue extracts wereanalyzed hy SDS-PAGE on a 1 mm thick polyacrylamide gel (T = II,C = 1, 0.37M Tris, pH 8.8, and 0.1% SDS in the separating gel andT = 3.75, C = 2.67,0.125 M Tris, pH 6.8. and 0.1 % SDS in the stackinggel) wilh a Hoeler (South San Francisco, CA) SE 600 apparatus. Therunning buffer was 0.3 M glycine. 0.02 M Tris. and 0.1% SDS(pH 8.8).A constant current of 8 mA per gel was applied for 15h and gels werestained overnight according to Neiihoff ct al. (1988), hut without thefixation step, followed by destaining in tap water.
DNA extraction: Genomic DNA was isolated from 5g of leaves fromsitigle plants as previously reported (D'Ovidio et al. 1992).
Polymersise Chain Reaction {PCR) ampliHcatiun: PCR reactions wereperformed in a final reaction volume of IOO/JI using IOO-3OOng ofgenomic DNA. 2.5 units of Taq DNA polymerase (Boehringer,Mannheim, Germany), 10//I of lOx Taq PCR buffer (Boehringer),250 ng each of the two primers, and the solution was 300//M in eachdeoxyribonucleotide. Amplification conditions were those reported inD"Ovidio et al. (1995) using primers, hased on the nucleotide sequenceofthe B,i:7 gene of'Cheyenne' {Anderson and Greene 1989), which hadthe following sequences;(a) 5 ATGGCTAAGCGCCTGGTCCT 3';(h) 5 TGCCTGGTCGACAATGCGTCGCTG 3'
each of dATP. dCTP and dGTP, 45//M in dTTP. 5/iM in Dig-1 l-UTP(Digoxigenin- 1 l-urtdine-5'-triphosphate) and with 20 ng each oftheT3 and T7 primers. The denaturing step was at 94 C for 1 min, theannealing step was at 55 C for 2 min and the extension step was at72 C for 2 min. The pi B9B clone (300 ng) was digoxigenin-Iabelted byrandom priming following the manufacturer's procedure (Boehringer).Tlie pHMWF5 clone contains the N-Ierminal portion of the IDx5*HMW glutenin gene (Lafiandra et al. 1993) and the plB9B clone con-tains the 1.8 Kh Hindlll fragment of the By9 HMW glutenin gene(Halford et al. 1987). The plB9B done was kindly provided hy Dr P.R. Shewry, Long Ashton, U.K. The pUTV1181 and pUTVM09 arewheat RFLP probes isolated at the University of Tuscia (Viterbo, Italy)and localized on homoeologous chromosomes group 1.
Southern biot analysis: Genomic DNA (5-15/ig) was digested withHindlll. fractionated on an agarose gel and transferred to a nylonmembrane (Hyhond-N, Amersham. UK) following standardprocedures. Prehybridization and hybridization reactions were carriedout at 65 C in a solution containing 5X SSC (0.75 M NaCl, 0.075 Msodium citrate pH 7), 0.02% (w/v) sodium dodecylsulphate (SDS), 0.1 %(w,v) N-lauroylsarcosine and 1% (wv) blocking reagent (Boehringer).Filters were prehybridized for 4h and then hyhridized for 16-20 h withdigoxigenin-tabelled clones. After hybridization, filters were washedtwiceat65 Cin2XSSCand0.1%(w/v)SDSfor 15 min, and twice atthe same temperature in 0.1 X SSC plus 0.1% SDS for 30 min each.Immunologieal detection was performed using Anti-Digoxigenin-AP,Fab Fragmenls (Boehringer) following the manufacturer's instructions.
Densitometric measurements: The signal intensity of the hybridizationbands in the Southern analysis was evaluated by using a Hewlett Pack-ard scanner (ScanJet •Jp) and computer software developed by Prof. A.Valentini (University of Tuscia, Viterbo, Italy).
Results
Protein analysis
SDS-PAGE separations of the HMW-GS of cultivars 'RedRiver 68', 'Chinese Spring', and 'Cbeyenne' are shown in Fig. I
Sx7
labelling of HMW glutenin clones: The pHMWF5.pUTVl 181 and pUTVl 109 clones (?Ong) were labelled by PCR for 30amplification cycles in 100/d total volume using 2 units of Taq DNApolymerase (Boehringer), IX reaction buffer (Boehringer). 50/(M in
Fig. l;SDS-PAGE patterns of lotal protein extracts from endosperm(flour) of-Cheyenne" (CNN), 'Red River 68' (RR) and 'Chinese Spring"(CS)
Duplication of BA-7 HMW subunit gene in bread wheat 527
Table 1: HMW-GS composition for 'Cheyenne', 'Red River 68' and'Chinese Spring" as deduced from SDS-PAGE
CultivarHMW glutenin subunits
Ax Ay Bx By Dx Dy
'Cheyenne"'Chinese Spring"'Red River 68"
7*77
988
101210
and tbeir respective HMW-GS are listed iti Table 1. The batidcorresponding to subunit Bx7 present in 'Red River 68' was themost intensely stained compared to the otber HMW gluteninsubunits. It may be noted also tbat tbe bands designated as Bx7in 'Chinese Spring" and 'Cheyenne' differ very sligbtly in tbeirrelative mobilities. On the basis of tbe gel pattern sbown inFig. 1, the apparent molecular weight of tbe Bx7 present in'Cbinese Spring" is .sligbtly larger tban its equivalent in 'Cbey-enne". Tbe 5.Y7 subunit of'Red River 68" is close in mobility tothat of 'Chinese Spring". Presumably, the 'Red River 68' and'Chinese Spring" subunits correspond to the subunit 7 desig-nation of Marcbylo et al. (1992) whereas that of 'Cbeyenne"corresponds to tbeir subunit 7* designation. Lookbart et al.(1993) designated the 7 type as 1' and tbe 7* type as 7. Here weshall follow tbe nomenclature of Marcbylo et al. (1992).
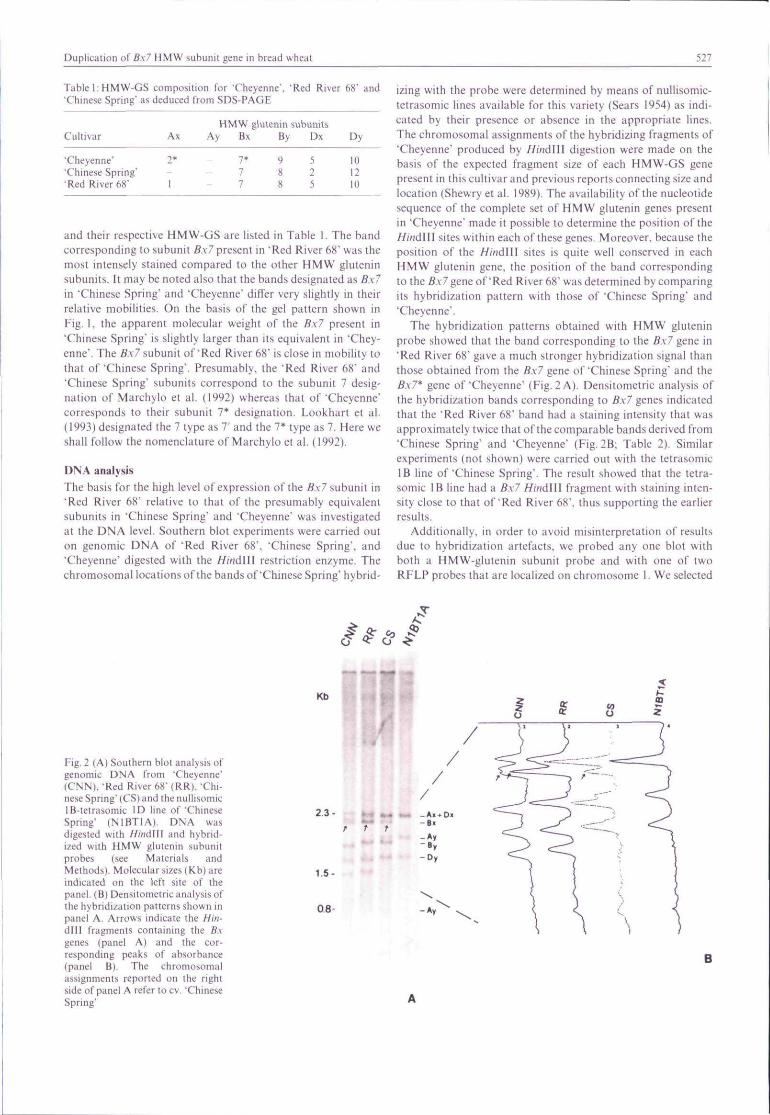
DNA analystsThe basis for tbe bigb level of expression ofthe Bx7 subunit in•Red River 68" relative to that of the presumably equivalentsubunits in 'Chinese Spring" and 'Cbeyenne" was investigatedat the DNA level. Southern blot experiments were carried outon genomic DNA of 'Red River 68", 'Chinese Spring', and'Cheyenne' digested witb the HimMW restriction enzyme. Tbechromosomal locations ofthe bands of'Chinese Spring" hybrid-
izing with the probe were determined by means of nullisomic-tetrasomic lines available for tbis variety {Sears 1954) as indi-cated by their presence or absence in tbe appropriate lines.Tbe cbromosomal assignments ofthe hybridizing fragments of'Cbeyenne' produced by HindWX digestion were made on tbebasis of the expected fragment size of eacb HMW-GS genepresent in tbis cultivar and previous reports connecting size andlocation (Shewry et al. 1989). Tbe availability of tbe nucleotidesequence of the complete set of HMW glutenin genes presentin 'Cbeyenne' made it possible to determine the position of theHindWi sites within eacb ofthese genes. Moreover, because tbeposition of tbe Hind\\\ sites is quite well conserved in eacbHMW glutenin gene, the position of tbe band correspondingto Ihe 6-v7 gene of'Red River 68" was determined by comparingits hybridization pattern witb those of 'Chinese Spring" and"Cheyenne'.
The hybridization patterns obtained with HMW gluteninprobe showed tbat tbe band corresponding to tbe Bx7 gene in•Red River 68' gave a much stronger hybridization signal thanthose obtained from tbe Bx7 gene of 'Chinese Spring' and theB.x7* gene of 'Cheyenne" (Fig. 2 A). Densitometric analysis oftbe bybridization bands corresponding to B.\7 genes indicatedtbat tbe 'Red River 68" band had a staining intensity that wasapproximately twice that ofthe comparable bands derived from'Chinese Spring" and 'Cbeyenne' (Fig.2B: Table 2). Similarexperiments (not shown) were carried out witb tbe tetrasomicIB line of 'Chinese Spring'. The result showed tbat the tetra-somic IB line had a Bx7 HindlU fragment witb staining inten-sity close to tbat of'Red River 68", thus supporting the earlierresults.
Additionally, in order to avoid misinterpretation of resultsdue to bybridization artefacts, we probed any one blot witbboth a HMW-giutenin subunit probe and with one of twoRFLP probes tbat are localized on chromosome 1. We selected
Kb
Fig. 2 (A) Soulhern blot analysis ofgenomic DNA from 'Cheyenne'(CNN), -Red River 68" (RR), 'Chi-nese Spring' (CS) and the nuliisomicIB-tetrasomic ID line of'ChineseSprtng" (N1 BT 1 A). DNA wasdigested with liindlW and hybrid-ized with HMW glutenin subunilprobes (see Materials andMethods). Molecular sizes (Kb) areindicated on the left site of thepanel. (B) Densitometric analysis ofthe hybridization patterns shown inpanel A. Arrows indicate the Hin-dlll fragments containing the fi.vgenes (panel A) and the cor-responding peaks of absorbance(panel B). The chromosomalassignments reported on the rightside of panel A refer to cv. 'ChineseSpring"
/
t f' 4
r•• - A x * OK
- B K
- A y- B y
1.5-
0.8-
- D y
-Ay
B

528 DOviDO, MASCI, PuRt'EDDU and KASARDA
Table2: Intensity of the HiiidlU fragments corresponding to the B.\7genes as obtained by densitometry ofthe blot pattern (as in Fig. 2). Thevalues are expressed as percentage ofthe total area and correspond tothe mean of triplicate determinations carried out with varying amountof genomic DNA
Cultivar
"Cheyenne"'Red River 68"'Chinese Spring"
14.5%27.9%14.6%
Kb
6 5 -
f *
UTV1181
2.3-
T f r
1.5-
HMW-GS
reaction between tbe two probes. Tbe resttlts for tbe pUTVi 109probe are not shown, but they duplicated tbe results obtainedwith Ihe pUTV1181 probe. Tbe blot was first bybridized witbtbe tiMW glutcnin subiinit probe and (hen re-hybridizcd withone of the RFLP probes. Tbe result obtained witb ibepUTVl 181 probe indicated RFLP fragments of similar si^e andintensity in the patterns of "Red River 68" and 'Chinese Spring',whereas tbe reaction of the HMW glutcnin subunit probe wasmucb stronger wiib the Bx7 fragment of'Red River 68' tbanfor the corresponding Bx7 fragment of "Chinese Spring' (Fig. 3).The cultivar 'Cheyenne' gave a somewhat different bybrid-ization pattern both in number and inten.sity of tbe bands ascompared to tbe 'Red River 6^" and 'Chinese Spring' patternsas sbown in Fig. 3. Still, tbe B.\7 Hind\\\ fragment gave a signalcomparable in intensity to tbe signal observed for the 'ChineseSpring" Bx7 band, but weaker tban tbe equivalent 'Red River68" band. This confirms the results ^hown in Fig. 2 where adouble staining intensity was observed for the 'Red River 68"B.\7 fragment relative to tbe corresponding bands in "ChineseSpring* and 'Cbeyenne".
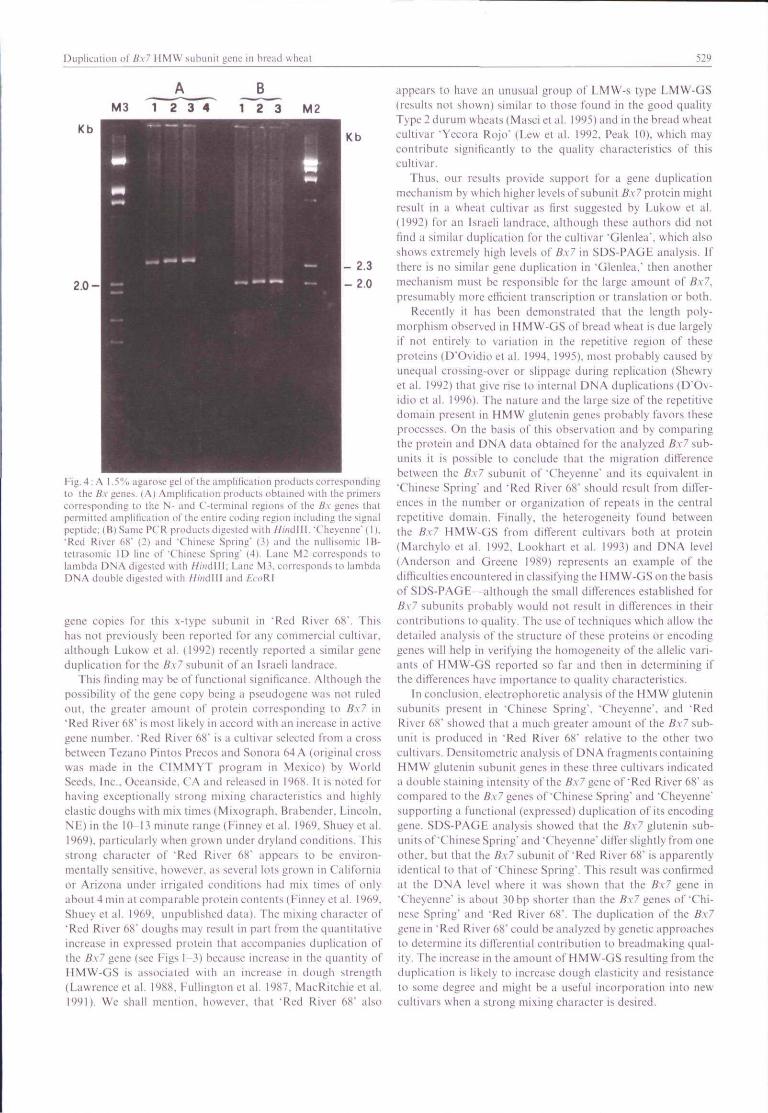
Southern analysis also indicated tbat tbe Hin<\\\\ fragmentcorresponding to tbe B.\7 gene of 'Red River 68" bas a sizesimilar to tbat of tbe equivalent gene present in 'Chinese Spring",but is sligbtly larger tban tbe 7* gene present in 'Cheyenne'. Inorder to determine tbe exact size of tbe complete Bx7 gene in'Red River 68" and 'Cbinese Spring' as compared to tbe onepreviously characterized in 'Cbeyenne', PCR analyses usingprimers specific for tbe complete coding region of this gene(D"Ovidio et al. 1995) were carried out. The amplification pro-ducts obtained were found to be of similar size in all tbreecultivars (Fig. 4) but, as already observed at protein level, theBx7 gene from 'Cheyenne' is slightly shorter (about 30 basepairs) than tbat of "Chinese Spring' and 'Red River 68'. Theestimated lengths of tbe amplification products was 2380 bp for"Chinese Spring" and 'Red River 68" and 2350bp for "Cbey-enne'. This latter value is very close to 2373 bp reported for tbe6.v7gene of tbis cultivar as determined by nucleotide sequencing(Anderson and Greene 1989). To verify tbe occurrence oftheobserved difference between the Bx7 gene of 'Cheyenne' andthose from 'Chinese Spring" and 'Red River 68", //mdlli diges-tions were carried out on their Bx7 amplification products. Asobserved for the entire coding region, tbe ////?dni digested PCRproduct from 'Cheyenne" was slightly shorter tban the one from'Chinese Spring" and 'Red River 68" {Fig. 4). Altbougb tbedifference in mobilities of the fragments eompared was small.It was consistently observed.
Fig. 3: Soulhern blot analysis of genomic DNA from 'Cheyenne'(CNN).'Red River ftf:!" ( RR'),'Chinese Spring" (CS). DNA was digestedwith Hind\\\ and hybridized with the pUTVl 181 probe and the HMWsubunit probes as in Fig. 2. Molecular weight sizes (Kb) are indicatedon the left site of ihf panel. Arrows indicate the HindlW fragmentscontaining the B.\ genes. The fragments hybridizing to the pUTVI IHlprobe fell iti the mobility range of Ihe upper bracket and the fragmentshybridizing with the HMW glutenin suhunii probes fell in Ihe mobilityrange ofthe lower bracket with no cross-hybridi/ation
two RFLP probes (pUTVl 181 and pUTVl 109) that hybridizedto ////?dlll fragments baving sizes very different from thosetbat were recognized by tbe HMW glutenin probe. The RFLPprobes reacted witb a group of slower moving bands on tbeblot whereas tbe HMW glutenin subunit probe reacted witb agroup of faster moving bands (Fig. 3). Tbere was no cross
DiscussionThe notably bigb intensity of tbe BA 7 HMW-GS of'Red River68" in SDS-PAGF analysis provided tbe incentive for our inves-tigation o\^ tbe possibility tbat tbis bigh intensity migbt resultfrom a gene duplication in tbis particular cultivar. As we baveindicated, it is frequently assumed tbat each HMW-GS is codedfor by a single gene, but tbis is not clearly establisbed. Exper-itncntal work bas mainly indicated that tbe copy number islikely to be between 1 and 3 (Tbompson et al. 1983, Forde etal. 1985). Accordingly, it would be more rigorous to considertbat the initial number of genes might be more than one. Ourresult indicating Ihat the //f>/dlll fragment of B.x7 from "RedRiver 68" gave a band intensity in Soutbern analysis appro.xi-mately twice tbat for the equivalent bands from "Cbeyenne" and'Chinese Spring" provides strong evidence for duplication in
Duplication of 5.v7 HMW subunit gene in bread wheat
M3
2.0-
Fig.4: A l..'>% agarose gel ofthc ampliticatioji products corresponding!c> the B.\ genes. (A) Amplification products oblained with the primerscorresponding to tlie N- and C'-terminal regions ofthe B.\ genes thatpennittcd amplification ofthe entire coding region including the signalpeplide; (B) Same PCR products digested with HiiiiMW. Cheyenne' (I),'Red River (iS' (2) and "Chinese Spring' (3) and the nullisomic 1 li-tetrasomic ID line of 'Chinese Spring' (4). Lane M2 corresponds tolambdii DNA digested with ///iidlll; l..;ine M.̂ . corresponds to lambdaDNA double digested with //;>idlll and IAOKI
gene copies for tbis x-type subunit in 'Red River 68". Tbishas not previously been reporled for any commercial cultivar.allboLigb Ltikow et al. (1^92) recently reported a similar geneduplication for tbe fi.v7 subunit of an Israeli landrace.
Tbis linding may be of functional significance. Altbough thepossibility ofthe gene copy being a pseudogene was not ruledout, tbe greater amount of protein corresponding to Bx7 in•Red River 6S' is most likely in accord with an increase in activegene nutnber. 'Red River 68" is a cultivar selected from a crossbetween Tezano Pintos Precos and Sonora 64 A (original crosswas made in the CIMMYT program in Mexico) by WorldSeeds. Inc.. Oceanside, CA and released in 1968. It is noted forhaving exceptionally strong mixing characteristics and highlyelastic dougbs with mix times (Mixograph, Brabcnder, Lincoln,NE) in tbe 10-13 minute range {Finney et al. 1969, Sbuey et al.1969). particularly when grown under dryland conditions. Tbisstrong cburacter oi' "Red River 6S' appears to be environ-mentally sensitive, however, as several lots grown in Californiaor Arizona under irrigated conditions had mix times of onlyabout 4 min at comparable protein contents (Finney et al. 1969.Sbuey el al. 1969, unpublished data). Tbe mixing cbaracter of•Red River 68" doughs may result in part frotn (he quantitativeincrease in expressed protein tbat accompanies duplication ofthe Bx7 gene {see Figs 1-3) because increase in tbe quantity ofHMW-GS is associated witb an increase in dougb strengtb{Lawrence et a l 19S8, Fullinglon et al. 1987, MacRitchie el al.1991). We shall mention, however, that 'Red River 68" also
appears to bave an unusual group of LMW-s type LMW-GS(results not sbown) similar to those found in the good qualityType 2 durum wheats {Masci etal. 1995) and in tbe bread wheatcultivar 'Yecora Rojo' {Lew et al. 1992, Peak 10), which maycontribute significantly to tbe quality characteristics of tbiscultivar.
Tbus. our results provide support for a gene duplicationmecbanism by wbich bigber levels of subunit Bx7 protein mightresult in a wbeat cultivar as iirst suggested by Lukow et al.(1992) for an Israeli landrace. although these authors did notfind a similar duplication for the cuitivar 'Glenlea", which alsoshows extremely high levels of Bx7 in SDS-PAGE analysis. Iftbere is no similar gene duplication in •Glenlea," then anothermecbanism must be responsible for the large amount of S.v7.presumably more efficient transcription or translation or both.
Recently it bas been demonstrated that the length poly-morphism observed in HMW-GS of bread wbeat is due largelyif not entirely to variation in the repetitive region of theseproteins (D'Ovidio et al. 1994. 1995), most probably caused byunequal crossing-over or slippage during replication (Sbewryet al. 1992) that give rise to internal DNA duplications (D'Ov-idio et al. 1996). The nature and tbe large size of tbe repetitivedomain present in HMW glutcnin genes probably favors tbeseprocesses. On tbe basis of tbis observation and by comparingtbe protein and DNA data obtained for the analyzed Bx7 sub-units it is possible to conclude tbat tbe migration differencebetween tbe Bx7 subunit of "Cheyenne" and its equivalent in•Cbinese Spring" and 'Red River 68" sbould result from differ-ences in tbe number or organization of repeats in tbe centralrepetitive domain. Finally, tbe beterogeneity found betweentbe Bx7 HMW-GS from different cultivars botb at protein{Marcbylo et al. 1992. Lookbart et al. 1993) and DNA level{Anderson and Greene 1989) represents an example of tbedifficulties encounlered in classifying the HMW-GS on tbe basisof SDS-PAGE —although tbe small dilferences establisbed forBx7 subunits probably would not result in differences in theireontributions to quality. The use of tecbniques which allow tbedetailed analysis of the structure of these proteins or encodinggenes will belp in verifying the homogeneity of the allelic vari-ants of HMW-GS reported so far and then in determining ifthe differenees have importance to quality characteristics.
In conclusion, electrophoretic analysis of the HMW gluteninsubunits present in •Chinese Spring", 'Cheyenne", and 'RedRiver 68" sbowed tbat a mucb greater amount of the Bx7 sub-unit is produced in "Red River 68" relative to the otber twocultivars. Densitometric analysis of DNA fragments containingHMW glutenin subunit genes in tbese tbree cultivars indicateda double staining intensity of tbe S.v7gene of'Red River 68" ascompared to the Rv7 genes of'Cbinese Spring" and •Cheyenne"supporting a functional (expressed) duplication of its encodinggene. SDS-PAGE analysis sbowed that the B.x7 glutenin sub-units of'Cbinese Spring" and •Cbeyenne" differ slighliy from oneotber, but that the Bx7 subunit of •Red River 68" is apparentlyidentical to tbat of 'Chinese Spring'. This result was confirmedat the DNA level wbere it was sbown that tbe B.v7 gene in'Cbeyenne' is about 30 bp sborter than the Bx7 genes of 'Chi-nese Spring' and 'Red River 68". Tbe duplication of tbe Bx7gene in •Red River 68" could be analyzed by genetic approacbesto determine its differential contribution to breadmaking qual-ity. Tbe increase in tbe amount of HMW-GS resulting from tbeduplication is likely to increase dougb elasticity and resistanceto some degree and might bc a useful incorporation into newcultivars wben a strong mixing cbaracter is desired.

530 D'Oviuo, MASCI, PORCHDDU and KASARDA
Acknowledgements
The authors thank Dr P. R. Shewry for providing the p i B9B clone andProf. A. Valentini for providing soflware and assistance for densi-totnetric analysis. They also thank DrC. O. Qualsetand MrH. Vogtforincreasing wheat stocks and helpful discussion, and George Lookhart(USDA-ARS, Manhattan, K.S» and Gary Hareland (USDA-ARS.Fargo. NU) for finding and supplying the 1969 Crop Quality Reports.The research was :iupported primarily hy the Italian Miiiistero delleRisorse Agricole, Alimentari e Forestall, Nationai Research Project'Plant Biotechnology'.
ReferencesAnderson. O. D., F. C. Greene, and J. C. Litts. 1990: Structure of the
j-gliadm gene family from the bread wheat eultivar 'Cheyenne".In: W. Bushuk, and R. Tkachuk (eds) Gluten Proteins, 640—645.American Association of Cerea! Chemists, St. Paul. MN.
Anderson, O. D., and F. C. Greene, 1989: The characterization andcotnparative analysis of high-molecular-weight genes frotii genomesA and B ofa hexaploid bread wheat. Theor. Appl. Genet. 77, 689—700.
D'Ovidio, R., O. A. Tanzerella. and H. Porceddu. 1992: Isolation ofan alpha-type gliadin gene from Triticum durum Desf and geneticpolymorphism at the (i!i-2 loci. J. Genet., Breed. 46, 41 ^ 8 .
, E. Porceddu. and D. Lafiandra, 1994: PCR analysis of genesencoding allelic variants of high-molecular-weight glutenin subunitsat the Glu-Dl locus. Theor. Appl. Genet. 88, 175—180.
— —, S. Masei, and E. Poreeddu, 1995: Development o f a set ofoligonudeotide primers specific for genes at the Glu-1 locus. Theor.Appl. Genet. 91. 189—194.
. D. Lafiandra. and E. Porceddu. 1996: Identilication and molec-ular characterization ofa large high-molecular-weight glutenin subu-nit gene from hexaploid wheat. Theoret. Appl. Genet. 93, 1048—1053.
Finney. K. F., M. D. Shogren, L. C. Bolce, R. C. Hoseney. J. D.Hubbard, B. M. Eichman, and W. J. Hoover, 1969: Quality charac-teristics of hard red winter wheat varieties grown in the Southern.Central, and Northern Great Plains ofthe United States, 1968 Crop.Progress report ofthe US Department of Agriculture, AgriculturalResearch Service Crops Research Division and tbe Kansas Agri-cultural Experimeni Station. Manhattan. KS.
Forde, J., J.-M. Malpica. N. G. Halford, P. R. Shewry. O. D. Anderson,F. C Greene and B. J. Miflins, 1985: The nucleotide sequence ofaHMW glutenin subunit gene located on chromosome I A of wheat(Trilicum aeslivum L.). Nucl. Acids Res. 13, 6817 -68.12.
Fullington, J. G.. D. M. Miskelly, C. W. Wrigley, and D. D. Kasarda,1987: Quality-related endosperm proleins in sulfur-deticien! and nor-mal wheat grain. J. Cereal Sci. 5, 2.13 245.
Halford. N. G.. J. Forde, O. D. Anderson, F. C. Greene, and P. R.Shewry. 1987: The nutleotide and deduced amino acid sequences ofan HMW glutenin subunit gene from chromosome 1B of bread wheat{Trilicum aesliium L.) and comparison with ihose of genes fromchromosomes 1 A and ID. Theor. Appl. Genet. 75, 117—126.
Harberd, N. P., D. Bartels. and R. D. Tbompson, 1986: DNA restric-tion-fragment variation in the gene family encoding high molecular(HMW) gluteninsubunits of wheat. Biochem. Genet. 24, 579—595.
Huebner. F. R.. and J. A. Bietz. 1985: Detection of quality differencesamong wheats by high performance liquid chromatography. J. Chro-matogr. 327. 333—342.
Kasarda. D. D., J.-C. Autran, F. J.-L. Lew, C. C. Nimmo, and P. R.Shewry, 1983: N-terminal atnino acid sequences ofoj-gliadinsandiu-secalins. Implications for the evoiution of prolamin genes. Biochem.Biophys. Acta 747, 138—150.
Kolster, P.. C. F. Krechting and W. M. J. van Gdder, 1992: Quanti-fication of individual high molecular weight subunits of giuteninusing SDS-PAGF and scanning densitometry. J. Cereal Sci. 15, 4961.
Latiandra, D.. R. D'Ovidio. F. Porceddu, B. Margiotta, and G. Colap-nco, 1993: New data supporting High Mr glutenin subunit 5 as the
determinant of quality differences among the pairs 5 -(- 10 vs. 2 + 12.J. Cerea! Sci. 18, 197 205.
, D. D. Kasarda, and R. Morris, 1984: Chromosomal assignmentof genes coding for the wheat gliadin components of the cultivars'Cheyenne" and 'Chinese Spring' by two-dimensional (two-pH) eiec-trophoresis. Theor. Appi. Genet. 68, 531—539.
Lawrence, G. J.. F. MacRitchie. and C. W. Wrigley, 1988: Dough andbaking quality of wheat lines deficient in glutenin subunits controlledby theOVw-^/, 67(J-.e/. andG/»-£)/lod. J. Cerea!Sci.7, 109—112.
Lew, E. J.-L., D. D. Kuzmicky, and D. D. Kasarda. 1992: Charac-terization of LMW-glutenin subunits by RP-HPLC, SDS-PAGE,and N-terminal amino acid sequencing. Cereal Chemistry 69, 508—515.
Lookhart, G. L.. K. Hagman, and D. D. Kasarda, 1993: High-molec-ular-weight glutenin subunits of the most commonly grown wheatcultivars in the U.S. in 1984. Plant Breeding 110, 48 -62.
Lukow, O. M., S. A. Forsyth, atid P. I. Payne, 1992: Over-productionof HMW glutenin subunits coded on chromosome IB in commonwheat. Triticum aestivum. J. Genet. Breed. 46, 187—192.
MacRitchie, F., D. D. Kasarda, and D. D. Kuzmicky, 1991: Charac-terization of wheat protein fractions differing in contribution tobreadmaking quality. Cereal Chem. 68, 122—130.
Marchylo, B. A., O. M. Lukow, and J. E. Kruger, 1992: Quantitativevartation in high molecular weight glutenin subunit 7 in some Can-adian wheats. J. Cereal Sc. 15, 29—37.
Masci, S., E. J.-L. Lew, D. Lafiandra, E. Porceddu, and D. D. Kasarda,1995: Characterization of low molecular weight glutenin subunits indurum wheat by reversed-phase high-performance liquid chro-matrography and N-terminal sequencing. Cereal Chem. 12. IOO—104.
Neuhoff, V.. N. Arold, D. Taube, W. Fhrhardt, 1988: Improved stainingof proteins in polyacrylamide gels including isoelectric focusing gelswitb clear background at nanogram sensitivity using Coomassie Bril-liant Blue G-250 and R-250. Electrophoresis 9, 255—262.
Payne. P. L, and G. J. Lawrence. 1983: Catalogue of alleles for thecomplex gene loci Giii-Al. Glu-Bl and Glu-Dl which code for high-molecular-weight subunits of glutenin in hexaploid wheat. CerealRes. Commun. 11,29 -35.
- —. L. M. Holt, and C. N. Law, 1981: Structural and geneticalstudies on high-molecular-weight subunits of wheat glutenin. I: allelicvariation in subunits amongst varieties of wheat (Trilicum aeslivum).Theor. Appl. Genetic 60, 229 236.
. .R. D. Thompsoti, D. Bartels, N. P. Harberd. P. A. Harris.and C. N. Law. 19K3: The high-molecular-weight glutenins: classicalgenetics, molecular genetics and the relationship to bread makingquality. Proc. 6th Int. Wheat Genet. Symp. 827—834.
, , F. A. Jackson, and C. N. Law. 1984: Wheat storageproteins: their genetics and their potential for manipulation by plantbreeding. Phil Trans. R. Soc. Food Agric. 32, 51 -60.
, M. A. Nightingale, A. F. Krattiger, and L. M. Holt, 1987:The relationship between HMW subunit composition and the bread-making quality of British-grown wheat varieties. J. Sc. Food Agric.40,51—65.
, L. M. Holt, A. T. Krattiger. and J. M. Carrillo, 1988: Reiation-ship between HMW glutenin subunit composition and measures ofthe bread-makingquality of wheat varieties grown in Spain. J. CerealSci.7, 229—236.
Sears, E. R. 1954: The aneuploids of common wheat. Res. Bull. No.572, Missouri Agricultural Experiment Station Bulletin. Columbia,MO.
Shewry, P. R., J. M. Field, A. J. Faulks. S. Parmar. B. J. Miflin, M. D.Dietlers, E. Lew. and D. D. Kasarda. 1984: The purification and N-terminal amino acid sequence analysis ofthe high-molecular-weightgluten polypeptides of wheat. Biochem. Biophys. Acta 788, 23—34.
, N. G. Halford, and A. S. Tatham. 1989: The high molecularweight subunits of wheat, barley and rye: genetics, molecular biology,chemistry and role in wheat gluten structure and functionality.Oxford Surv. of Plant Mol. Cell Biol. 6, 163—219.

Duplication of fi.v7 HMW subunit gene in bread wheat 531
. . . 1992: High-molecular-weight subunits of wheat Singh. N. K., K. W. Shepherd, and G. B. Cornish, 1991: A simplitiedglutenin. J. Cereal Sci. 15, 105—120, SDS-PAGE procedure for separating LMW subunits of glutenin. J.
Shuey, W. C . V. L. Youngs, K. J. Sprick, R. D. Crawford. R. D. Cereal Sci. 14,203 208.Maneval. D. J. Thompson. N. B. Lofthus, and K. A. Gilles, 1969: Thompson, R. D., D. Bartels, N. P. Harberd, and R. B. Flavell, 1983:Annual Report for 1968, Hard Red Spring and Durum Wheat Qual- Charaeterization of the multigene family coding for HMW gluteninity Laboratory, U.S. Department of Agriculture, Agriculture subunits in wheat using cDNA clones. Theor. Appl. Genet. 67, 87—Research Service and Crops research Division, Cereal Crops 96.Research Branch, North Dakota State University AgriculturalExperimental Station. Fargo, North Dakota.





















