
Habitat associations of trees and seedlings in a Bornean rain forest
Upload
independentCategory
view
0download
0
Tropical Zoology, 24: 63-86, 2011
Dragonflies (Odonata) in the Bornean rain forest as indicators of changes in biodiversity resulting from forest modification and destruction
A. Dolný 1,5, D. Bárta, S. Lhota 2, 3, Rusdianto 4 and P. Drozd 1
1 Faculty of Science, University of Ostrava, CZ-710 00 Ostrava, Czech Republic2 Department of Zoology, Faculty of Science, University of South Bohemia, CZ-370 05 České Budějovice, Czech Republic3 Ústí nad Labem Zoo, CZ-400 07 Ústí nad Labem, Czech Republic4 Unit Pelaksana Badan Pengelolaan Hutan Lindung Sungai Wain, 76127 Balikpapan, Indonesia
Received 16 November 2010, accepted 1 March 2011
Dragonfly assemblages represent sensitive indicators of en-vironmental conditions including the water environment and forest structure. However, an understanding the ecology of tropical forest odonates remains one of the most significant gaps in our knowledge of the order. We sampled odonates at Sungai Wain Protection Forest, East Kalimantan, Indonesia. Relatively high dragonfly species rich-ness (88 species) was found during 35 survey days divided between two seasons. Seasonal differences in the species diversity were fairly small, in accordance with the fact that the climate in Kalimantan is generally stable. The highest species diversity was observed in intact primary forest: 60% of all recorded species were found there and 32% of all species were exclusive to this habitat. The proportion of bio-tope specialists decreased along the gradient — intact primary forest, slightly degraded primary forest, secondary forest, heavily degraded forest and non-forest. The ordination obtained from detrended cor-respondence analysis showed, along the main axis, a gradient in com-munity similarity corresponding to the degree of forest degradation. The evidence indicates that, within the applied spatial scale, any forest degradation results in fewer species, with a pronounced change in the species composition, and an overall reduction in taxonomic diversity.
key words: dragonflies, Odonata, diversity, primary and secondary dipterocarp forest, Borneo.
5 Corresponding author: Dr Ales Dolný (E-mail: [email protected]).

A. Dolný et alii64
Introduction .................................................................................. 64 Materials and methods ................................................................... 65
Study area .................................................................................. 65Dragonfly sampling ................................................................... 66Localities and habitat types ........................................................ 67Statistical analysis ...................................................................... 67
Results .......................................................................................... 70Discussion .................................................................................... 74Acknowledgements ....................................................................... 79References ..................................................................................... 79Appendix ...................................................................................... 83
INTRODUCTION
Although the tropical rainforest contains by far the greatest species diversi-ty of the planet’s dragonfly fauna, to date these species have been proportionately understudied (Corbet 2006, Kalkman et al. 2008). Many species in tropical forest habitats are poorly studied and often known only from the type specimen (Clausnitzer et al. 2009). An understanding of the ecology and biology of tropi-cal forest dragonfly species is one of the most significant gaps in our knowledge of the order Odonata (Orr 2006), although this is a broad problem, which does not only apply to Odonata (Collen et al. 2008). We have relatively few longitu-dinal studies relating to the species diversity of Odonata in the tropical rainforest, and furthermore most studies derive from brief glimpses that do not embrace the changing seasons (Corbet 2006). Exceptions are the detailed and long-term faunistic or ecological studies of dragonfly species diversity in the tropical rain-forest of Liberia (Lempert 1988), the lower montane rainforest of Papua New Guinea (Oppel 2005) and the Tambopata-Candamo Reserved Zone in south-eastern Peru (Paulson 1985, Butt 1995). Therefore, extensive new field surveys of odonate fauna in tropical forest areas are needed (Clausnitzer et al. 2009).
Tropical forests are the Earth’s most biologically rich ecosystems. Yet, the destruction continues apace, with some thirteen million hectares of forest felled or razed each year. Rapid forest destruction is evident in countries such as Brazil and Indonesia (Butler & Laurance 2008). Monitoring changes in biodiversity resulting from forest modification and destruction requires the study of a wide range of taxa, embracing species with very different ecologies and life histories (Lawton et al. 1998). Dragonflies can be good indicators of forest structure and landscape composition (Clausnitzer 2003) and dragonfly assemblages are high-ly visible and sensitive indicators of long-term environmental conditions of the water environment (Stewart & Samways 1998).
Threatened species are as well clustered in tropical areas, especially in the Indo-Malayan region (Clausnitzer et al. 2009). Thus far, regional or local fau-nistic surveys of dragonflies in Borneo have been very limited, especially those that

Dragonflies of Borneo 65
relate occurrence to habitat (Orr 2006). The northern part of Borneo is by far the best researched part of the island, especially the sultanate of Brunei (Thomp-son & van Tol 1993, Orr 2001, Orr 2006) and to a lesser extent Sarawak (Dow 2008, Dow & Reels 2008, Dow & Reels 2010) and Sabah (Yagi & Kitagawa 2001), including Mount Kinabalu (the highest mountain in Borneo) (Laidlaw 1934, Hämäläinen 1994). Kalimantan (Indonesian Borneo) makes up 2/3 of Borneo, but the odonate fauna of Kalimantan is poorly studied and the distribution and habitat requirements of many dragonfly species are unknown.
Only one recent study, in East Kalimantan, has examined the diversity and community composition of odonates in Indonesian Borneo (Cleary et al. 2004). That research on odonates (and butterflies) took place from January-October 2000 in and around the Balikpapan-Samarinda region and it included Sungai Wain Protection Forest, the focus of our present research. There and at five addi-tional forest sites, extending over an area of 5.2 million ha, 89 species of odonates were recorded in total (in the Results, the article mistakenly states “72 species of odonates“). Only 47 dragonfly species were found in Sungai Wain, as noted after Appendix II in Cleary et al. (2004). However, this relatively small number of dragonfly species may have resulted from a limited sampling effort related to the extensive nature of the survey. In Cleary et al. (2004), 16 sites (32 together in Sungai Wain Protection Forest) were sampled per landscape (primary forest and once-burned forest), which occupied approximately 0.9 ha (along a 300 m transect), and odonates were sampled along the entire transect until at least 20 odonates were caught per site.
This study has the following objectives: (1) to produce a dragonfly species list for a small rainforest reserve, Sungai Wain Protection Forest, in East Kaliman-tan, Indonesia, and describe the patterns of species distribution within this area; (2) to assess the effects of habitat modification on diversity of odonate fauna; (3) to compare the richness and diversity of dragonfly assemblages in natural (pri-mary dipterocarp forest) and human-modified (secondary forest and non-forest habitat) environments in an East Kalimantan landscape.
MATERIAL AND METHODS
Study area
Sungai Wain Protection Forest (SWPF) is a water catchment reserve (first gazetted in 1934) located within the administrative area of Balikpapan, East Kali-mantan, Indonesia (S 01°02’-01°10’, E 116°47’-116°55’). The reserve itself covers an area of less than 100 km2, and the research area covers approximately 20 km2. Average annual rainfall is 2738 mm and the mean daily temperature is 26.5 °C. The climate is relatively stable, although June to September tends to be drier than Octo-ber to May. Most of the terrain is gently undulating, with the northern part of the reserve characterized by steeper slopes and higher ridges, while the southern part of the reserve is relatively flat. The altitude varies from 30 m to 150 m asl. The soil is predominantly sandy ultisol and alluvium with low water retention capacity. There are no big rivers in the reserve and smaller rivers and streams are numerous, running predominantly southwards. Therefore, the southern part of the reserve is moister and

A. Dolný et alii66
swampier, while the ridges of the northern area are relatively dry. Swamps represent approximately 5.9% of the primary forest, have no significant peat layer, and are semi permanent, drying out several times per year. Dominant tree families include Dipte-rocarpaceae, Euphorbiaceae, Sapotaceae, and Myrtaceae. Dipterocarpaceae are most dominant in the northern part of the reserve, while Sapotaceae and Euphorbiaceae are more dominant in the south. Undergrowth is relatively dense, frequented with rattan (Arecaceae), Zingiberaceae and Marantaceae.
In 1947, a small water reservoir and pump station was built along the Wain River (Sungai Wain). Today it serves as a source of water for the refineries and house-holds of an oil refinery complex. The water below the reservoir is mildly brackish, with riverbank vegetation dominated by Nypa fruticans. Hence, the reservoir marks a sharp transition between a freshwater and brackish water system. Sungai Bugis, a trib-utary of the Wain River, was later diverted into the same reservoir via a 1.72 km long channel, causing forest flooding above the reservoir and channel and the emergence of swamps. Many of these swamps were initially converted into paddy fields, but follow-ing settlement relocation from the south-western part of the reserve (see below), the paddy fields were left fallow and transformed into open marshes. A smaller portion of the swamp is now covered with almost permanently flooded tall swamp forest.
Only part of SWPF still consists of primary rainforest, as there have been hu-man settlements and farms in the south-western part of the reserve until as recently as the 1970’s and 1980’s. These settlements were gradually relocated to what is today Sungai Wain village and were replaced with a tall secondary forest, containing a large proportion of planted trees such as fruit and rubber. But in the 1960’s and 1970’s, the provincial Samarinda-Balikpapan road was constructed along the eastern border of the Protection Forest, leading to a new wave of encroachment. The north-eastern part of the reserve has been converted into farms and settlements and these conditions still persist today.
Most of the reserve was affected by the prolonged drought of the 1997-1998 ENSO (El Niño Southern Oscillation), which resulted in increased tree mortality in the primary forest and, in March 1998, in forest fires. The origin of the fires was most likely anthropogenic. These fires remained mainly in the undergrowth and caused high tree mortality along dry ridges, but less so along more humid valleys. The fires were eventually stopped by creating firebreaks, although over 50% of the reserve was affected (Fredriksson 2002). The burnt forest has been left to regenerate naturally. It has already developed a closed canopy of pioneer species such as Macaranga gigantea, and regeneration of primary forest species is in process. The area of unburnt forest in the central part of the reserve is approximately 4,000 ha and currently represents the largest fragment of primary rainforest in the Balikpapan-Samarinda area.
Dragonfly sampling
We sampled odonates at SWPF from 29 July to 16 August 2008 (19 days), and from 11 to 26 January 2010 (16 days). Specimens were collected on sunny (non-rainy) days between 10.00 and 18.00 hr. Sampling was carried out at equal intervals in both seasons for four forest types and for all locations. Species accumulation curves (SAC, see e.g. Colwell & Coddington 1994, Magurran 2004) were created using AccuCurve software (Drozd & Novotny 2010) (Fig. 1). All localities were visited on at least two different days each season. Species were identified visually and, when nec-essary, caught with butterfly nets and identified based on the literature. The identifi-cations were later confirmed by Rory A. Dow (National Museum of Natural History Naturalis, Leiden, The Netherlands). Not all specimens could be identified to species level, and some species were new to science and have yet to be formally described.

Dragonflies of Borneo 67
Localities and habitat types
Dragonflies were sampled in four habitats: 1) primary forest (PF): intact, undisturbed forest, including natural swamps, rivers and streams in primary forest; 2) slightly degraded primary forest (SDF): selectively logged forest or forest only marginally affected by human presence or fires; 3) secondary forest (SF): degraded, logged, burnt forest, regeneration of the primary forest, including swamps, rivers and streams in secondary forest; 4) heavily degraded forest and non-forest (HNF): crop-land, dam, etc. Individual research localities have been sketched in the Sungai Wain Protection Forest map (Fig. 2) and a detailed characterisation of habitats at individual research locations is introduced in Table 1.
Statistical analysis
The β-diversity (compositional similarity), i.e. species turnover between habi-tats, was measured using the Cody β-diversity index: βCo = 1 − [a(2a+b+c)/2(a+b)(a+c)], where “a” is the total number of species occurring in any pair of habitats, and “b” and “c” are the number of exclusive species in each habitat. This index is one of several recently used indices of β-diversity (Koleff et al. 2003) and can be used to measure species turnover. The Jaccard similarity index was used to evaluate the qualitative differences in species composition between the four habitat types and was calculated for each pair: J = a/(a+b+c) where a represents the number of species
Fig. 1. — Species accumulation curves for 4 habitat types (○ = 2008, ∆ = 2010; PF = primary forest, SDF = slightly degraded primary forest, SF = secondary forest, HNF = heavily degraded forest and/or non-forest).

A. Dolný et alii68
simultaneously present in these two sampling sites and b and c the number of species found at only one of the sites.
Detrended correspondence analysis (DCA) was chosen as a unimodal in-direct gradient analysis to visualize dissimilarities in species composition. Canonical correspondence analysis was consequently performed to produce a constrained or-dination model with habitat type (PF, SDF, SF, HDF), as a nominal environmental variable (coded as a dummy variable). The significance of the constrained model was tested using a Monte Carlo permutation test. Locality “Cemetery” was omitted for both analyses due to lack of information (only one specimen of one species was re-corded).
Fig. 2. — Map of Sungai Wain Protection Forest (D. Bárta, orig.; landcover interpreted from ALOS AVNIR-2, image scene id. ALAV2A185843620, observed 2/7/2009, processed to level 1B2, (c)JAXA: all rights reserved, distributed by RESTEC).

Dragonflies of Borneo 69
LocalityHabitat
typesEnvironment GPS coordinates
BugiDjam PF River in primary forest S 1.097°, E 116.822°ForeDeva PF Primary forest with closed canopy S 1.103°, E 116.828°LakeNatu PF Natural landslide-dammed lake S 1.102°, E 116.836°StreRhin PF Stream in primary forest S 1.102°, E 116.831°
SwamDjam PF Natural swamp along river source, forming under sun-flacks in a canopy S 1.097°, E 116.822°
SwamNepe PF Natural swamp along river source, forming under sun-flacks in a canopy S 1.089°, E 116.823°
SwamRoot PF Seasonal swamp in primary forest with closed canopy S 1.097°, E 116.822°
SwamTraH PF Natural swamp along river source, forming under sun-flacks in a canopy S 1.097°, E 116.822°
Camp1 SDF Primary forest with small sun-flacks in the canopy as a consequence of human activity S 1.131°, E 116.833°
Camp3 SDF Primary forest with small sun-flacks in a canopy as a consequence of human activity S 1.097°, E 116.822°
Cemetery SDF Primary forest with small sun-flacks in a canopy as a consequence of human activity S 1.149°, E 116.820°
PosUlin SDF Primary forest with small sun-flacks in a canopy as a consequence of human activity S 1.144°, E 116.840°
StreamC1 SDF Stream in slightly degraded primary forest S 1.135°, E 116.832°StreamC2 SDF Stream in slightly degraded primary forest S 1.115°, E 116.830°
SwamBurn SDF Swamp, slightly degraded primary forest surrounded by burnt forest S 1.095°, E 116.808°
SwamGra1 SDF Swamp, slightly degraded primary forest surrounded by burnt forest S 1.098°, E 116.810°
SwamGra2 SDF Swamp, slightly degraded primary forest surrounded by burnt forest S 1.095°, E 116.813°
EcoCircl SDF Primary forest with small sun-flacks in a canopy as a consequence of human activity S 1.138°, E 116.834°
OtterRiv SF Stream in secondary burnt forest S 1.089°, E 116.812°WainBrid SF River in degraded forest by dam reservoir S 1.142°, E 116.837°WainGaze SF Degraded river banks in secondary forest S 1.139°, E 116.841°WainNatu SF River in secondary forest S 1.137°, E 116.842°
WoodPath SFSwamp forest created by a dam with small sun-flacks in the canopy as a consequence of human
activityS 1.140°, E 116.837°
BridCulm HNF Stream, surrounding heavily degraded forest S 1.098°, E 116.870°
BugiChan HNF Channel, dammed stream with slow current in open area with some riverine vegetation S 1.144°, E 116.836°
BugiOutf HNF River outfall into channel S 1.153°, E 116.826°DamReser HNF Dam reservoir in open area S 1.143°, E 116.837°DamWetle HNF Swamps in the alluvial banks of the dam reservoir S 1.145°, E 116.837°
Fielpath HNF Degraded farmlands S 1.091°, E 116.872°SwamFiel HNF Artificial pool in cultivated landscape S 1.097°, E 116.869°
WainUppe HNF Stream, surrounding heavily degraded forest S 1.092°, E 116.866°
Table 1.
Location and selected attributes of sample sites (PF = primary forest, SDF = slightly degraded primary forest, SF = secondary forest, HNF = heavily degraded forest and/or non-forest).

A. Dolný et alii70
RESULTS
In total, 88 dragonfly species were found in SWPF (Table 2, Appendix 1). Of these, 72 species were found during July and August 2008 and 78 spe-cies were found in January 2010. Seasonal differences in the species diversity of odonate fauna were relatively small. The Jaccard similarity index was calculated at 0.7045 (Table 2), i.e. 62 species were common to both observed seasons. Ten spe-cies were found only in 2008 (July/August), of which only Tetrathemis flavescens Kirby 1889 occurred in three localities. Other species were found in only one of the localities surveyed, and generally only a few individuals were present. Sixteen species were found only in 2010 (January), most of which occurred at only one locality, a maximum of two localities, and only in very low abundances. Only one species reached high abundance in one season and was entirely absent the following season. It was Lyriothemis biappendiculata Selys 1878, whose popula-tion in 2008 exceeded 100 individuals in one locality but was not found at all at the locality in 2010.
The 88 recorded species belong to 14 families (out of 15 known in Bor-neo): 34 (39%) species belong to the suborder Zygoptera and 54 (61%) species belong to the suborder Anisoptera (Table 3). By far the most plentiful family was Libellulidae, with 38 species, followed by Coenagrionidae, with 15 species. The highest species diversity was observed in intact primary forests. About 60% of all recorded species (from 40 genera and 13 families) could be found in this habitat, and only one of the recorded dragonfly families (Macromiidae) was missing. The proportion of biotope specialists from the total number of species decreased along the disturbance gradient — intact primary forest, slightly degraded primary for-est, secondary forest, to heavily degraded forest and non-forest (Table 3). The intact primary forest also held the greatest number of endemic species. With re-gard to endemism, primary and slightly degraded forests are significantly more important than secondary, heavily degraded and non-forest habitats (Table 3).
About 32% (28 species) were found exclusively in intact primary forest and/or slightly degraded primary forest and only about 11% (10 species) were found exclusively in heavily degraded forest and/or non-forest. Only about 8% (7 species) of all recorded species were observed in all habitat types (Table 2). Of the 53 species recorded at primary forest sites, 39.6% (21 species) were also found in heavily degraded forest and/or non-forest. The highest β-diversity (and the highest species turnover values) was found between heavily degraded/non-forest and slightly degraded primary forest, which shared only 16 odonate species. The highest similarity (β = 0.39, Jaccard’s index = 0.433) was between intact primary forest and secondary forest (Table 4).
Detrended correspondence analysis demonstrates that habitat type was an important factor for the composition of dragonfly assemblages. The species spectrum of odonate assemblages changed significantly in relation to habitat degradation. The ordination diagram showed a gradient in community similar-ity from sites in the intact primary forest to slightly degraded forest, secondary (once-burned) forest to heavily degraded forest and non-forest along the first axis (explaining a total variance of 17.5% of species data). The differentiation of drag-onfly assemblage along the second axis takes into account its water habitat type. The upper part of the graph shows dragonfly assemblages at flowing water, the

Dragonflies of Borneo 71
middle shows dragonfly assemblages at stagnant water or larger open water areas, the bottom shows dragonfly assemblages at forest swamps (Fig. 3). Relationships between species and habitat are also evident from the results of the Monte-Carlo permutation test for constrained ordination models (CCA) where significance of the canonical axes was found (habitat type, F = 1.26, P = 0.011).
The relative representation of Anisoptera and Zygoptera in the odonate community differed by habitat type (Fig. 4). Whereas Anisoptera dominated in secondary forest (24 Anisoptera vs. 19 Zygoptera species) and heavily degraded forest/non-forest habitats (25 vs. 17 species), the two suborders were more equally represented in intact (26 Anisoptera vs. 27 Zygoptera species) and slightly de-graded primary forest (20 vs. 22 species). Within the Zygoptera, Coenagrionidae were the most represented family in all habitat types but relatively more so in the heavily degraded habitats (Fig. 4A). The difference was even more noticeable between forest and non-forest habitats (Fig. 4B). Non-forest habitats had signifi-cantly fewer Zygoptera. Apart from Coenagrionidae (9 species) there were only 2 additional Zygoptera species (vs. 24 Anisoptera species), whereas primary forest localities had 11 coenagrionids plus an additional 16 Zygoptera species.
Fig. 3. — Detrended correspondence analysis (DCA). Ordination diagram represents similarity of the dragonfly assemblages at 31 sites belonging to four different habitat groups. Lines connect samples belonging to the same habitat group (○= intact primary forest, ◊ = slightly degraded forest, □ = secondary forest, = degraded forest and non-forest; for site abbreviations see Table 1).
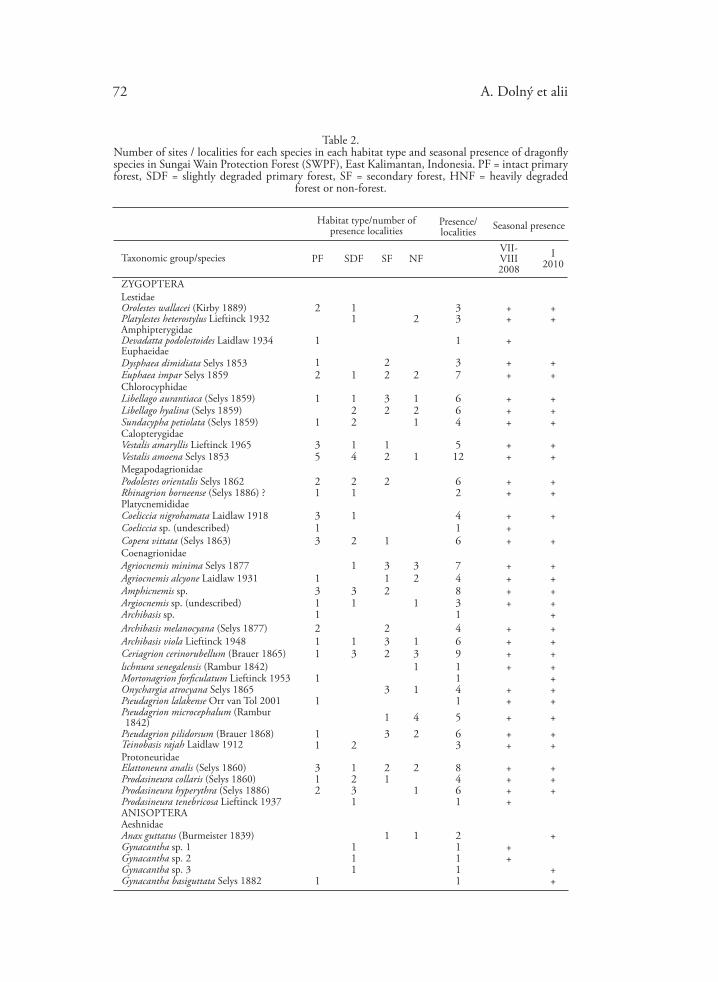
A. Dolný et alii72
Habitat type/number of presence localities
Presence/localities Seasonal presence
Taxonomic group/species PF SDF SF NFVII-VIII2008
I2010
ZYGOPTERALestidaeOrolestes wallacei (Kirby 1889) 2 1 3 + +Platylestes heterostylus Lieftinck 1932 1 2 3 + +AmphipterygidaeDevadatta podolestoides Laidlaw 1934 1 1 +EuphaeidaeDysphaea dimidiata Selys 1853 1 2 3 + +Euphaea impar Selys 1859 2 1 2 2 7 + +ChlorocyphidaeLibellago aurantiaca (Selys 1859) 1 1 3 1 6 + +Libellago hyalina (Selys 1859) 2 2 2 6 + +Sundacypha petiolata (Selys 1859) 1 2 1 4 + +CalopterygidaeVestalis amaryllis Lieftinck 1965 3 1 1 5 + +Vestalis amoena Selys 1853 5 4 2 1 12 + +MegapodagrionidaePodolestes orientalis Selys 1862 2 2 2 6 + +Rhinagrion borneense (Selys 1886) ? 1 1 2 + +PlatycnemididaeCoeliccia nigrohamata Laidlaw 1918 3 1 4 + +Coeliccia sp. (undescribed) 1 1 +Copera vittata (Selys 1863) 3 2 1 6 + +CoenagrionidaeAgriocnemis minima Selys 1877 1 3 3 7 + +Agriocnemis alcyone Laidlaw 1931 1 1 2 4 + +Amphicnemis sp. 3 3 2 8 + +Argiocnemis sp. (undescribed) 1 1 1 3 + +Archibasis sp. 1 1 +Archibasis melanocyana (Selys 1877) 2 2 4 + +Archibasis viola Lieftinck 1948 1 1 3 1 6 + +Ceriagrion cerinorubellum (Brauer 1865) 1 3 2 3 9 + +lschnura senegalensis (Rambur 1842) 1 1 + +Mortonagrion forficulatum Lieftinck 1953 1 1 +Onychargia atrocyana Selys 1865 3 1 4 + +Pseudagrion lalakense Orr van Tol 2001 1 1 + +Pseudagrion microcephalum (Rambur 1842) 1 4 5 + +
Pseudagrion pilidorsum (Brauer 1868) 1 3 2 6 + +Teinobasis rajah Laidlaw 1912 1 2 3 + +ProtoneuridaeElattoneura analis (Selys 1860) 3 1 2 2 8 + +Prodasineura collaris (Selys 1860) 1 2 1 4 + +Prodasineura hyperythra (Selys 1886) 2 3 1 6 + +Prodasineura tenebricosa Lieftinck 1937 1 1 +ANISOPTERAAeshnidaeAnax guttatus (Burmeister 1839) 1 1 2 +Gynacantha sp. 1 1 1 +Gynacantha sp. 2 1 1 +Gynacantha sp. 3 1 1 +Gynacantha basiguttata Selys 1882 1 1 +
Table 2. Number of sites / localities for each species in each habitat type and seasonal presence of dragonfly species in Sungai Wain Protection Forest (SWPF), East Kalimantan, Indonesia. PF = intact primary forest, SDF = slightly degraded primary forest, SF = secondary forest, HNF = heavily degraded
forest or non-forest.

Dragonflies of Borneo 73
Gynacantha limbalis Karsch 1892 1 1 +Gynacantha dohrni Kruger 1899 1 1 + +Heliaeschna idae (Brauer 1865) 1 1 +Oligoaeschna mutata Lieftinck 1940 ? 1 1 +Tetracanthagyna degorsi Martin 1895 1 1 + +Tetracanthagyna plagiata (Waterhouse 1877) 1 1 1 3 + +
GomphidaeIctinogomphus decoratus (Selys 1854) 1 2 3 + +Leptogomphus coomansi Laidlaw 1936 1 1 +MacromiidaeEpophthalmia vittigera (Rambur 1842) 1 1 2 4 + +Macromia cincta Rambur 1842 2 2 +CorduliidaeHemicordulia tenera Lieftinck 1930 1 1 +LibellulidaeAcisoma panorpoides Rambur 1842 1 1 +Aethriamanta gracilis (Brauer 1878) 1 1 +Agrionoptera insignis (Rambur 1842) 1 1 + +Agrionoptera sexlineata Selys 1879 1 1 +Brachydiplax chalybea Brauer 1868 1 1 2 4 + +Brachydiplax farinosa Kruger 1902 1 2 3 + +Brachygonia oculata (Brauer 1878) 1 3 1 5 + +Cratilla metallica (Brauer 1878) 1 1 2 + +Diplacodes trivialis (Rambur 1842) 3 3 + +Hydrobasileus croceus (Brauer 1867) 1 1 + +Lyriothemis biappendiculata (Selys 1878) 1 1 +Lyriothemis cleis Brauer 1868 1 1 +Nannophya pygmaea Rambur 1842 1 1 2 4 + +Nesoxenia lineata (Selys 1868) 1 1 +Neurothemis fluctuans (Fabricius 1793) 1 3 4 8 + +Neurothemis ramburii (Kaup in Brauer 1866) 1 1 + +
Neurothemis terminata Ris 1911 1 1 2 2 6 + +Onychothemis coccinea Lieftinck 1953 2 1 3 + +Onychothemis culminicola Förster 1904 1 1 2 4 + +Orchithemis pulcherrima Brauer 1878 1 2 3 + +Orthetrum chrysis (Selys 189]) 2 1 1 4 + +Orthetrum sabina (Drury 1770) 1 3 4 + +Orthetrum testaceum (Burmeister 1839) 1 1 2 4 + +Pantala flavescens (Fabricius 1798) 1 1 + +Pornothemis serrata Krüger 1902 1 1 2 +Rhodothemis rufa (Rambur 1842) 1 1 1 3 + +Rhyothemis obsolescens Kirby 1889 2 2 4 + +Rhyothemis phyllis (Sulzer 1776) 1 1 + +Rhyothemis triangularis Kirby 1889 1 1 1 3 + +Tetrathemis flavescens Kirby 1889 1 1 1 3 +Tetrathemis irregularis Brauer 1868 2 2 +Tholymis tillarga (Fabricius 1798) 2 2 + +Tramea transmarina Brauer 1867 1 1 +Trithemis aurora (Burmeister 1839) 1 2 3 + +Tyriobapta laidlawi Ris 1919 3 1 4 + +Tyriobapta torrida Kirby 1889 3 8 3 14 + +Urothemis signata (Rambur 1842) 1 2 3 + +Zyxomma petiolatum Rambur 1842 1 1 + +Species aggregate 53 42 43 42 72 78Jaccard index 70,45
Total number of species 88
Table 2 (continued)Habitat type/number of
presence localitiesPresence/localities Seasonal presence
Taxonomic group/species PF SDF SF NFVII-VIII2008
I2010

A. Dolný et alii74
DISCUSSION
Our research shows that deforestation will reduce species richness, with an especially significant change in the identity of species. Endemics, more special-ized species and species tending toward smaller ranges (predominantly Zygop-tera) were more vulnerable to forest degradation than commonly caught species, whereas anthropogenic disturbance in a locality resulted in widespread species colonisation or persistence at that site (Table 3). The qualitative aspect of the changes in dragonfly diversity in relation to tropical rain forest degradation is em-phasized in several similar studies. A comparison between the natural and modi-fied forest in Papua New Guinea showed that the modified forest clearly holds more species that are widespread (Oppel 2006). During a ten-month study, 78 species were found, of which 61 occurred in a natural rain forest area and only 37 in a modified rain forest. The primary forest (including the swamps and spring sources) in coastal habitats of Kenya contained a very unique and highly special-ised dragonfly community (Clausnitzer 2003).
It is obvious, however, that the biodiversity of intact primary forest and of habitats resulting from modification of tropical forests showed taxonomic group — and guild-specific differences (Lawton et al. 1998). Inventories of fruit-feed-ing butterflies and dung beetles in Sulawesi (Indonesia) showed that some human land-use systems supported relatively high numbers of species and might play a significant role in tropical biodiversity conservation (Schulze et al. 2004). Some studies in Indonesia indicated that species diversity was significantly higher in secondary or degraded forest than in intact or less disturbed forest (Hamer et al.
PF SDF SF HNF Total
Number of families 13 (93%) 10 (71%) 11 (79%) 10 (71%) 14
Number of genera 40 (67%) 33 (55%) 31 (52%) 34 (57%) 60Number of species 53 (60%) 42 (48%) 43 (49%) 42 (48%) 88Biotope specialist species 38 (69%) 35 (64%) 27 (49%) 14 (25%) 55Aggregate of Zygoptera 27 (79%) 22 (65%) 19 (56%) 17 (50%) 34Aggregate of Anisoptera 26 (48%) 20 (37%) 24 (44%) 25 (46%) 54Habitat-specific species 12 (34%) 9 (26%) 4 (11%) 10 (29%) 35Habitat-specific Zygoptera 5 (71%) 1 (14%) 0 1 (14%) 7Habitat-specific Anisoptera 7 (25%) 8 (29%) 4 (14%) 9 (32%) 28Endemic species 8 (73%) 7 (64%) 1 (9%) 2 (18%) 11Localities with endemics 15 (50%) 11 (37%) 2 (7%) 2 (7%) 30
Table 3.
Inventory data of the odonate assemblages of all habitat types in Sungai Wain Protection Forest (SWPF), East Kalimantan, Indonesia, in 2008 and 2010.

Dragonflies of Borneo 75
Fig. 4. — Taxonomic composition of dragonflies in four different habitats (A) and the difference between forest and non-forest habitats (B). PF = intact primary forest, SDF = slightly degraded primary forest, SF = secondary forest, HNF = heavily degraded forest and non-forest; DF = degraded forest, range from slightly to heavily, NF = non-forest.
1997 for butterflies, Shahabuddin et al. 2010 for dung beetles). The same group of bio-indicators (dung beetles) in a different geographic area (Colombian Andes) yielded completely different results, showing greater numbers of species and in-dividuals in primary forest than in secondary forest and pasture (Escobar 2004). Another example of the negative effects of deforestation and forest degradation was found with ants in Cameroon (Watt et al. 2002). These differences may re-

A. Dolný et alii76
sult not only from differences between taxonomical groups but also from different methodologies used in the research. The effects of disturbance in tropical forests have been studied at a wide range of spatial scales, and species diversity within landscapes depends on the scale at which communities are sampled (Hamer & Hill 2000). Tropical rainforest is the planet’s most diverse terrestrial ecosystem in terms of species and habitats. It provides habitat for dragonfly species throughout some or all of their life-history stages (Corbet 2006). Of 164 Neotropical genera, 144 (87.8%) include forest species (Paulson 2006). Orr (2006) estimated that at least 70% of the Bornean odonate fauna is presently confined to forest habitats and probably depends on the forest for its survival.
We found 88 species in an area covering approximately 20 km2 in SWPF, which is a relatively high diversity of dragonflies. However, there are sites on earth that are much more species-rich. The most species-rich site in the world known so far is probably Tambopata-Candamo Reserved Zone in south-eastern Peru, at the upper edge of the Amazon basin (Paulson & von Ellenrieder 2005). This locality, only about 5.3 km2 in area, is comprised of large and small oxbow lakes, marshes and water courses. Between June 1977 and June 1984, 151 species of Odonata were collected or observed there (Paulson 1985) and, at present, its list of species totals 186 (Paulson & von Ellenrieder 2005). However, our sam-pling effort is still much less than in the Tambopata-Candamo study site; future research in Sungai Wain would likely document additional species (see Fig. 1).
There are a few detailed analyses of dragonfly diversity from smaller iso-lated natural areas on the island of Borneo. However, almost all of them are from the northern part of the island (Sarawak, Brunei). Dow & Reels (2008) found 106 dragonfly species from 13 families at Gunung Mulu National Park in north-eastern Sarawak, Malaysian Borneo in 2005 (April) and 2006 (February). This study site is much larger and has a greater habitat diversity than SWPF. It is the largest national park in Sarawak, with an area of 544 km2, and contains almost all of the main vegetation types found in Sarawak (not only mixed dipterocarp forest). Two years later, the same authors (Dow & Reels 2010) published species lists for three of Sarawak’s most easily accessible national parks (generally 5-year sampling period from 2005 through 2009): Kubah National Park (22 km2), 84 species from 13 families, Similajau National Park (71 km2), 51 species from 13 families; Lambir Hills National Park (70 km2), 73 species from 14 families. These
Table 4. Variation of the species composition of assemblages of all habitat types. Values of species turnover between habitats is indicated by Cody’s β-diversity index (upper diagonal). Faunistic similarity among of assemblages of all habitat types is calculated using classic Jaccard similarity index (lower
diagonal).
Habitat PF SDF SF HNFPF – 0.424 0.389 0.551
SDF 0.397 – 0.529 0.619SF 0.433 0.308 – 0.411
HNF 0.284 0.235 0.417 –

Dragonflies of Borneo 77
sites are located approximately 600-700 km north-east of SWPF, but have gener-ally the same climate and predominant habitat (mixed dipterocarp forest).
Our results differ from the only comparable study on the diversity of drag-onflies in Indonesian Borneo (Cleary et al. 2004). Cleary et al.’s list identi-fies only three species not included in our list, all of which were found in very low abundance: Euphaea subcostalis Selys 1873 (1 specimen), Lathrecista asiatica (Fabricius 1798) (3 specimens) and Vestalis atropha Lieftinck 1965 (1 specimen). However, we found a further 43 species in SWPF (including 5 endemic to Bor-neo): 8 of them occurred in purely non-forest habitat, which was not surveyed by Cleary et al. (2004), but we found a further 29 species of dragonfly new for SWPF in primary forest areas and 18 species in secondary, once-burned forest. These differences were most likely caused by different sampling objectives, plot design and sampling effort. Our objective was finding the maximum number of species at each of the 23 forest sites, without following a transect. Cleary et al. (2004) sampled along a randomly placed transect until at least 20 odonates were caught per site, which may lowered the chances of finding rarities. Equally im-portant may be temporary changes in the Sungai Wain odonate fauna following the 1997-98 prolonged drought and 1998 forest fire. Cleary et al.’s (2004) sam-pling took place in 2000. At that time, the burnt forest was colonized by dense underbrush of ferns and sedges, with only moderate recovery of the canopy. In this succession stage, the forest interior had been markedly dry and hot. At the time of our survey, a new closed canopy of fast-growing pioneer tree species had already developed. Ferns and sedges receded from most of the area where natural regeneration took place, air humidity increased and temperature decreased. These conditions are likely more hospitable for the forest odonate species.
Seasonal changes in dragonfly diversity in SPWF were relatively small, which is in accordance with the fact that the climate in the Kalimantan region is relatively aseasonal. Most species in south-eastern Kalimantan are probably facultatively multivoltine (unregulated life-cycle category according to Corbet 1999). Their larvae inhabit aquatic habitats that are continuously available for occupancy. Such dragonfly species may occur in a given area all year round. The more significant interseasonal change is absence of Lyriothemis biappen-diculata Selys 1878 and Lyriothemis cleis Brauer 1868 in January 2010. Inter-seasonal changes of these species would relate to their specific autoecology. Orr (1994) reported the breeding of Lyriothemis cleis in phytotelmata in the understorey of lowland mixed dipterocarp rainforest in Borneo. However, the use of phytotelmata as larval habitats for Lyriothemis species is facultative, tree holes being favoured when there is a paucity of standing water, e.g. on slop-ing ground. Dragonfly species typical for phytotelmata (temporary habitats) in lowland forest have a larval stage duration of about 1-3 months, and they survive the dry season as pre- or non-reproductive adults or perhaps in the egg stage (Corbet 1999).
Of particular interest are the records of Pseudagrion pilidorsum Brauer 1868, Pseudagrion lalakense Orr & van Tol 2001, Devadatta podolestoides Laid-law 1834, Onychothemis coccinea Lieftinck 1953 and Onychothemis culminicola Förster 1904. Pseudagrion pilidorsum had never before been recorded in Borneo (Orr 2003, 2006). The related species Pseudagrion lalakense had until recently been known from only a few blackwater lagoons in Brunei and northern Sarawak

A. Dolný et alii78
(Orr 2003), as a species found breeding in strongly acidic water rather than in more neutral water. But P. lalakense is likely a rather common species in Bor-neo (e.g. Dow & Reels 2008, 2010). At least two of the sampled dragonflies are still undescribed taxa (at least at the subspecies level); one representative of the genus Argiocnemis and one representative of Rhinagrion. Three dragonflies, Devadatta podolestoides, Onychothemis coccinea and Onychothemis culminicola are among priority species [based on recommendations of the IUCN/SSC, Odonata Specialist Group (Moore 1997)]. In addition, they are taxonomically isolated species from the Oriental Region (groups of species found nowhere else outside the Oriental Region).
A high species number (>15) was found in localities of all four main habitat types in the SPWF area according to the degree of environmental degradation by anthropogenic disturbances. The most species (27) were found at a locality (Wood Path) where a swamp forest had formed due to the building of a dam and the subsequent increase of the subterranean water level; further development of habitat had been spontaneous. The second highest number of species was found at a natural landslide-dammed lake (Lake Natu), without human impact, but where the formation and character of the habitat had been significantly affected by natural disturbances (landslide and damming of a stream). The other three most species-rich localities were significantly affected by human activities, specifi-cally the building and operation of a dam; the richest of them was a river above the inlet into the dam, bordered by a forest where vegetation on the banks was regularly cut (Wain Brid, 22 species).
From the analysis of community similarity (Fig. 3, Table 4), it follows that dragonfly assemblages from a natural landslide-dammed lake (Lake Natu) were very close in species composition to dragonfly assemblages from degrad-ed habitats. The odonate fauna of this locality has a total of 12 shared species (Agriocnemis alcyone Laidlaw 1931, Argiocnemis spA, Brachydiplax chalybea Brauer 1868, Ceriagrion cerinorubellum (Brauer 1865), Epophthalmia vittigera (Rambur 1842), Ictinogompus decoratus (Selys 1854), Nannophya pygmaea Rambur 1842, Neurothemis terminata Ris 1911, Neurothemis ramburii (Kaup in Brauer 1866), Orthetrum testaceum (Burmeister 1839), Rhodothemis rufa (Rambur 1842), Rhyo-themis triangularis Kirby 1889), which did not occur in any other natural habitat (PF). It is possible to conclude that a natural disturbance, which conditioned the formation of this aquatic bio-type, had in many respects a similar impact on dragonflies as the impact of human activities. This fact had a profound in-fluence on the values of species turnover between habitats indicated by Cody’s β-diversity index (Table 4), where the highest β-diversity values (and the highest species turnover values) were found between heavily degraded forest or non-forest (HNF) and slightly degraded primary forest (SDF), and not between HNF and primary forest (PF), as had been expected. If we exclude the dragonfly assemblage of “Lake Natu” from the evaluation of β-diversity indices between the groups of habitat types, we obtain different results. In this case, the values of β-diversity index increase in an expected sequence: the lowest value of species turnover is between PF and SDF (0.424), the middle value is between PF and SF (0.566) and the highest is between PF and HNF (0.742).
The differences in taxonomic composition of the odonate fauna in individ-ual groups of habitats usually agree with relevant literature data. The distribution

Dragonflies of Borneo 79
of odonate species, including their family profiles, in a mosaic of complex tropical forest formations in Brunei was studied by Orr (2006). The summary results of that paper as well as the results of our study point to a significant reduction in the proportional representation of Zygoptera and the increase of Anisoptera (espe-cially the family Libellulidae) in relation to the increasing degree of forest degrada-tion and deforestation. The odonate fauna of mixed dipterocarp forest in Brunei and SWPF is formed by approximately 50% Zygoptera species and 50% Anisop-tera species. It concerns especially the family Libellulidae, 25% (Brunei) vs. 39% (SPWF), other Anisoptera, 29% (Brunei) vs. 9% (SPWF), and also Coenagrioni-dae, 8% (Brunei) vs. 21% (SPWF). The family profiles of odonate species from sites in non-forest are, however, quite similar in both areas, Libellulidae represent 64% in Brunei and 60% in SWPF, other Anisoptera, 15 % in Brunei and 9% in SWPF, Coenagrionidae, 19% in Brunei and 26% in SWPF, and other Zygoptera, 2% in Brunei and 6% in SWPF. In a pristine lower mountain rainforest study area in Papua New Guinea, Oppel (2005) found a very high ratio of Zygoptera to Anisoptera and he concluded that the habitat requirements of most closed-forest Zygoptera are unlikely to be met in degenerated forest or in a largely deforested landscape; therefore, he proposed the use of these groups as indicators of an un-disturbed rainforest. Therefore, our study confirms previous findings that a low ratio of Zygoptera to Anisoptera is an indication for highly disturbed rivers and other water bodies and particularly of disturbance caused by commercial forestry (Stewart & Samways 1998, Samways & Steytler 1996, Clausnitzer 2003).
ACKNOWLEDGEMENTS
This paper was written in connection with an Institute of Environmental Technologies project, reg. no. CZ.1.05/2.1.00/03.0100, supported by Research and Development for Innovations Operational Programme financed by Structural Founds of Europe Union. The fieldwork was supported by Grant No. SM/6/104/05 from the Ministry of Environment of Czech Republic and Grant No. 206/07/0811 from the Grant Agency of Czech Republic. We would also like to thank Pilsen, a candidate city for European Capital of Culture 2015, for its support. While preparing the manuscript, Stanislav Lhota was supported by Grant No. MSMT 6007665801 from the Ministry of Education, Youth and Sports of Czech Republic. We would like to thank Muhammad Fitriadi and Muhalir for field assistance, and Agusdin, Purwanto and Gabriella Fredriksson for additional facilitation of the fieldwork, and Sheryl Gruber for language correction of the manuscript, and Rory A. Dow for help with determination.
REFERENCES
Butler R.A. & Laurance W.F. 2008. New strategies for conserving tropical forests. Trends in Ecology & Evolution 23: 469-472.
Butt M. 1995. Odonata collected from the Tambopata-Candamo Reserved Zone, southeastern Peru, August 1992-January 1993. Notulae Odonatologicae 4: 93-97.

A. Dolný et alii80
Clausnitzer V. 2003. Dragonfly communities in coastal habitats of Kenya: indica-tion of biotope quality and the need of conservation measures. Biodiversity and Conservation 12: 333-356.
Clausnitzer V., Kalkman V.J., Ram M., Collen B., Baillie J.E.M., Bedjanič M., Darwall W.R.T., Dijkstra K.-D.B., Dow R., Hawking J., Karube H., Malikova E., Paulson D., Schütte K., Suhling F., Villanueva R., von Ellenrieder N. & Wilson K. 2009. Odonata enter the biodiversity crisis debate: the first global assessment of an insect group. Biological Conservation 142: 1864-1869.
Cleary, D.F.R., Mooers A.Ø., Eichhorn K.A.O., van Tol J., de Jong R. & Menken S.B.J. 2004. Diversity and community composition of butterflies and odonates in an ENSO-induced fire affected habitat mosaic: a case study from East Kalimantan, Indonesia. Oikos 105: 426-446.
Collen B., Ram M., Zamin T. & McRae L. 2008. The tropical biodiversity data gap: addressing disparity in global monitoring. Tropical Conservation Science 1: 75-88.
Colwell R.K. & Coddington J.A. 1994. Estimating terrestrial biodiversity through extrapolation. Philosophical Transactions of the Royal Society of Lon-don B 345: 101-118.
Corbet P.S. 1999. Dragonflies: Behaviour and Ecology of Odonata. Colchester UK: Harley Books, 829 pp.
Corbet P.S. 2006: Forests as habitats for dragonflies (Odonata), pp. 13-36. In: Rivera A.C., Edit. Forest and Dragonflies. Fourth WDA International Symposium of Odonatology, Pontevedra (Spain), July 2005. Sofia-Moscow: Pensoft Publishers, 299 pp.
Dow R.A. 2008. Odonata of the “Glen Forest” at Samarakan, Bintulu Division, Sarawak, Malaysian Borneo. Agrion 12(2): 46-48.
Dow R.A. & Reels G.T. 2008. List of species recorded at Gunung Mulu national park, Sarawak, Malaysian Borneo in 2005-2006. Echo 5: 2-3 (published in Agrion 12(1)).
Dow R.A. & Reels G.T. 2010. The Odonata of three National Parks in Sarawak. Agrion 14(1): 14-19.
Drozd P. & Novotny V. 2010. AccuCurve. Version 1. (Available from: http://prf.osu.cz/kbe/index.php?id=8072).
Escobar F. 2004. Diversity and composition of dung beetle (Scarabaeinae) as-semblages in heterogeneous Andean landscape, Colombia. Tropical Zoology 17: 123-136.
Fredriksson G.M. 2002. Extinguishing the 1998 forest fires and subsequent coal fires in the Sungai Wain Protection Forest, East Kalimantan, Indonesia, pp. 74-81. In: Moore P. et al., Edits. Communities in flames. Proceedings of an in-ternational conference on community involvement in fire management, Bangkok: FAO and Fire-Fight SE Asia, 133 pp.
Hämäläinen M. 1994. Dragonflies of Mount Kinabalu (the highest mountain in Borneo). Malangpo 11: 77-81.
Hamer, K.C., Hill J.K., Lace L.A. & Longan A.M. 1997. Ecological and biogeo-graphical effects of forest disturbance on tropical butterflies of Sumba, Indo-nesia. Journal of Biogeography 24: 67-75.
Hamer, K.C. & Hill J.K. 2000. Scale-dependent effects of habitat disturbance on species richness in tropical forests. Conservation Biology 14: 1435-1440.
Kalkman V.J., Clausnitzer V., Dijkstra K.D.B., Orr A.G., Paulson D.R. & van Tol J. 2008. Global diversity of dragonflies (Odonata) in freshwater. Hydro-biologia 595: 351-363.
Koleff P., Gaston K.J. & Lennon J.J. 2003. Measuring beta diversity for presence-

Dragonflies of Borneo 81
absence data. Journal of Animal Ecology 72: 367-382. Laidlaw F.F. 1934. A note on the dragonfly fauna (Odonata) of Mount Kinabalu
and some other mountain areas of Malaysia: with a description of some new or little known species. Journal Federated Malay States Museums 17 (3): 549-561.
Lawton J.H., Bignell D.E., Bolton B., Bloermers G.F., Eggleton P., Hammond P.M., Hodda M., Holt R.D., Larsen T.B., Mawdsley N.A., Stork N.E., Srivastava D.S. & Watt A.D. 1998. Biodiversity inventories, indicator taxa and effects of habitat modification in tropical forest. Nature 391: 72-76.
Lempert J. 1988. Untersuchungen zur Fauna, Ökologie und zum Fortpflanzungsver-halten von Libellen (Odonata) an Gewässern des tropischen Regenwaldes in Liberia, Westafrika. Diplomarbeit, Bonn, Germany: Friedrich-Wilhelms Uni-versität, 238 pp.
Magurran A.E. 2004. Measuring Biological Diversity. Malden, Massachusetts: Black-well Publishing Company, 256 pp.
Moore N.W. (compiler) 1997. Dragonflies - Status Survey and Conservation Ac-tion Plan. IUCN/SSC Odonata Specialist Group. Gland, Switzerland and Cambridge, UK: IUCN, V + 28 pp.
Oppel S. 2005. Habitat associations of an Odonata community in a lower mon-tane rainforest in Papua New Guinea. International Journal of Odonatology 8: 243-257.
Oppel S. 2006. Comparison of two Odonata communities from a natural and a modified rainforest in Papua New Guinea. International Journal of Odonatol-ogy 9: 89-102.
Orr A.G. 1994. Life histories and ecology of Odonata breeding in phytotelmata in Bornean rainforest. Odonatologica 23 (4): 365-377.
Orr A.G. 2001. An annotated checklist of the Odonata of Brunei with ecological notes and descriptions of hitherto unknown males and larvae. International Journal of Odonatology 4: 167-220.
Orr A.G. 2003. A guide to the dragonflies of Borneo: their identification and biol-ogy. Kota Kinabalu, Sabah, Borneo: Natural History Publications, 195 pp.
Orr A.G. 2006. Odonata in Bornean tropical rain forest formations: diversity, ende-micity and implications for conservation management, pp. 51-78. In: Rivera A.C., Edit. Forest and Dragonflies. Fourth WDA International Symposium of Odonatology, Pontevedra (Spain), July 2005. Sofia-Moscow: Pensoft Publishers, 299 pp.
Paulson D.R. 1985. Odonata of the Tambopata Reserved Zone, Madre de Dios, Peru. Revista Peruana de Entomología 27: 9-14.
Paulson D.R. 2006. The importance of forests to neotropical dragonflies. pp. 79-101. In: Rivera A.C., Edit. Forest and Dragonflies. Fourth WDA International Symposium of Odonatology, Pontevedra (Spain), July 2005. Sofia-Moscow: Pensoft Publishers, 299 pp.
Paulson D.R. & von Ellenrieder N. 2005. Synonymy of Subaeschna Martin, 1908, with Gynacantha Rambur, 1842, and a new species of Gynacantha from Peru (Anisoptera: Aeshnidae). Odonatologica 34: 65-72.
Samways M.J. & Steytler N.S. 1996. Dragonfly (Odonata) distribution patterns in urban and forest landscapes, and recommendations for riparian management. Biological Conservation 78: 279-288.
Schulze C.H., Waltert M., Kessler P.J.A., Pitopang R., Shahabuddin, Ved-deler D., Muhlenberg M., Gradstein S.R., Leuschner C., Steffan-De-wenter I. & Tscharntke T. 2004. Biodiversity indicator groups of tropical land-use systems: comparing plants, birds, and insects. Ecological Applications 14: 1321-1333.
Shahabuddin, Hidayat P., Manuwoto S., Noerdjito W.A., Tscharntke T. &

A. Dolný et alii82
Schulze C.H. 2010. Diversity and body size of dung beetles attracted to dif-ferent dung types along a tropical land-use gradient in Sulawesi, Indonesia. Journal of Tropical Ecology 26: 53-65.
Stewart D.A.B. & Samways M.J. 1998. Conserving dragonfly (Odonata) assem-blages relative to river dynamics in an African savanna game reserve. Conserva-tion Biology 12: 683-692.
Thompson D.J. & van Tol J. 1993. Damselflies and dragonflies from four forest types in Brunei. Brunei Museum Journal 8: 57-72.
Watt A.D., Stork N.E. & Bolton B. 2002. The diversity and abundance of ants in relation to forest disturbance and plantation establishment in southern Cam-eroon. Journal of Applied Ecology 39: 18-30.
Yagi T. & Kitagawa K. 2001. A survey of the dragonflies in the Klias and Binsulok Forest Reserves, Sabah, Malaysia. Nature and Human Activities 6: 31-39.
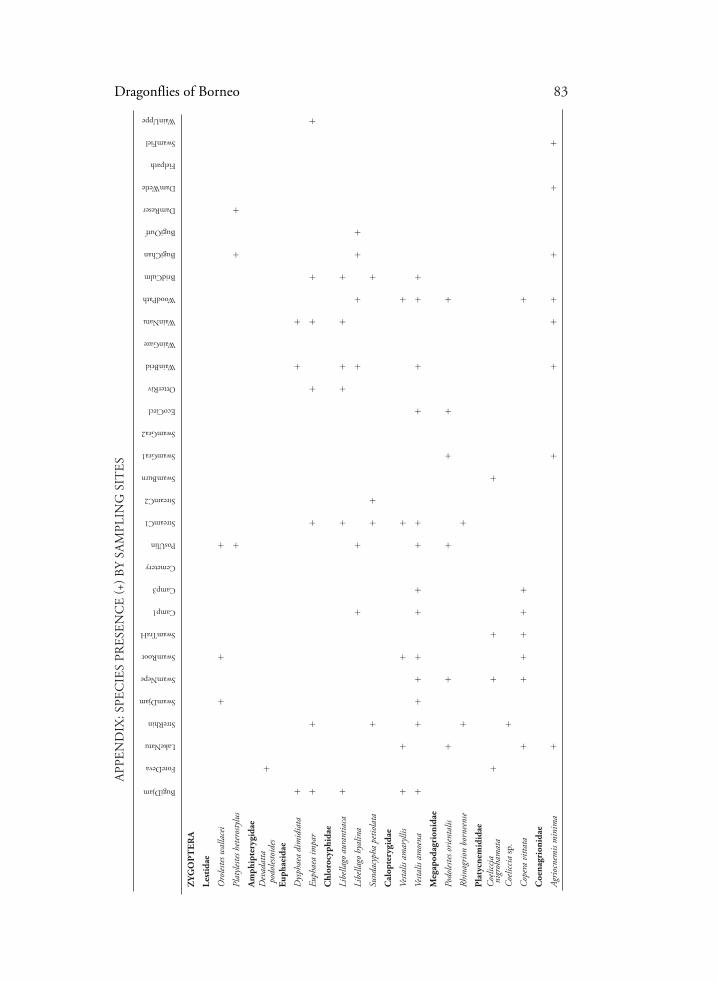
Dragonflies of Borneo 83
APP
END
IX: S
PEC
IES
PRES
ENC
E (+
) BY
SAM
PLIN
G S
ITES
BugiDjam
ForeDeva
LakeNatu
StreRhin
SwamDjam
SwamNepe
SwamRoot
SwamTraH
Camp1
Camp3
Cemetery
PosUlin
StreamC1
StreamC2
SwamBurn
SwamGra1
SwamGra2
EcoCircl
OtterRiv
WainBrid
WainGaze
WainNatu
WoodPath
BridCulm
BugiChan
BugiOutf
DamReser
DamWetle
Fielpath
SwamFiel
WainUppe
Zyg
Op
te
ra
Lest
idae
Oro
leste
s wal
lace
i +
++
Plat
yleste
s het
eros
tylu
s +
++
am
phip
tery
gida
eD
evad
atta
po
doles
toid
es +
euph
aeid
ae
Dys
phae
a di
mid
iata
+
++
Euph
aea
impa
r +
++
++
++
Chl
oroc
yphi
dae
Libe
llago
aur
antia
ca
++
++
++
Libe
llago
hya
lina
++
++
++
Sund
acyp
ha p
etio
lata
+
++
+C
alop
tery
gida
e
Vesta
lis a
mar
yllis
++
++
+Ve
stalis
am
oena
+
++
++
++
++
++
++
Meg
apod
agri
onid
ae
Podo
leste
s orie
ntal
is +
++
++
+Rh
inag
rion
born
eens
e +
+pl
atyc
nem
idid
aeC
oelic
cia
nigr
oham
ata
++
++
Coe
liccia
sp.
+C
oper
a vi
ttata
+
++
++
++
Coe
nagr
ioni
dae
Agrio
cnem
is m
inim
a +
++
++
++
+

A. Dolný et alii84
Agrio
cnem
is al
cyon
e +
++
+Am
phicn
emis
sp.
++
++
++
++
Argi
ocne
mis
sp.
++
+Ar
chib
asis
sp.
+Ar
chib
asis
mela
nocy
ana
++
++
Arch
ibas
is vi
ola
++
++
++
Cer
iagr
ion
cerin
orub
ellum
+
++
++
++
++
lschn
ura
seneg
alen
sis
+M
orto
nagr
ion
forfi
cula
tum
+
Ony
char
gia
atro
cyan
a +
++
+Ps
euda
grio
n la
lake
nse
+Ps
euda
grio
n m
icroc
epha
lum
+
++
++
Pseu
dagr
ion
pilid
orsu
m
++
++
++
+Te
inob
asis
raja
h +
++
prot
oneu
rida
e
Elat
tone
ura
anal
is +
++
++
++
+Pr
odas
ineu
ra co
llaris
+
++
+Pr
odas
ineu
ra
hype
ryth
ra
++
++
++
Prod
asin
eura
te
nebr
icosa
+
an
isO
pt
er
a
aes
hnid
ae
Anax
gut
tatu
s +
+G
ynac
anth
a sp
. 1
+G
ynac
anth
a sp
. 2
+G
ynac
anth
a sp
. 3+
Gyn
acan
tha
basig
utta
ta
+
BugiDjam
ForeDeva
LakeNatu
StreRhin
SwamDjam
SwamNepe
SwamRoot
SwamTraH
Camp1
Camp3
Cemetery
PosUlin
StreamC1
StreamC2
SwamBurn
SwamGra1
SwamGra2
EcoCircl
OtterRiv
WainBrid
WainGaze
WainNatu
WoodPath
BridCulm
BugiChan
BugiOutf
DamReser
DamWetle
Fielpath
SwamFiel
WainUppe
APP
END
IX (c
ontin
ued)
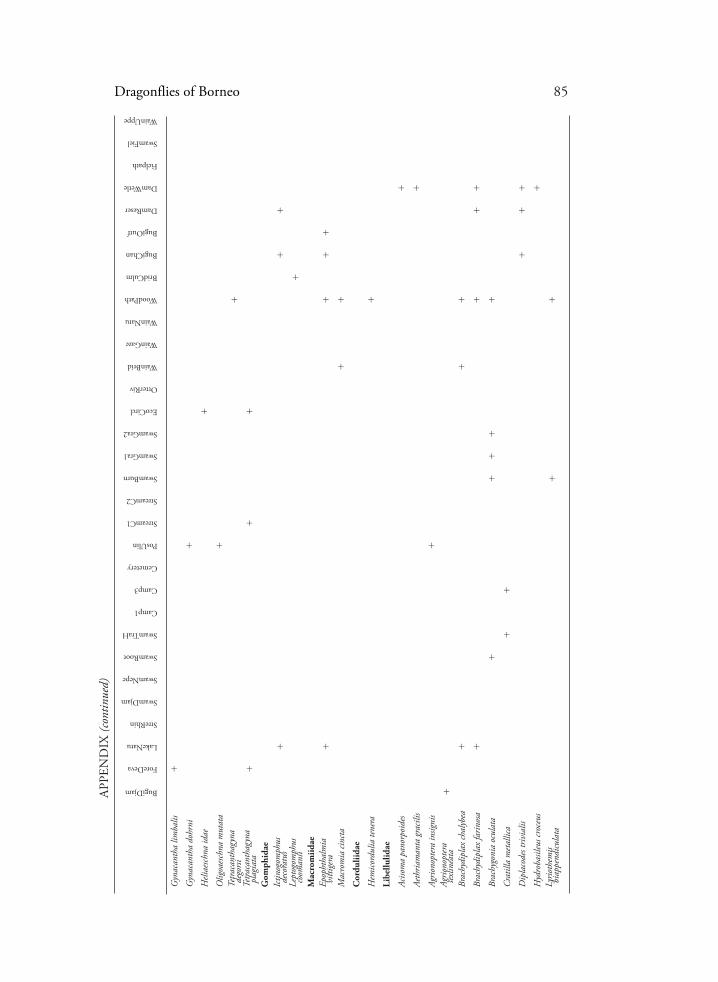
Dragonflies of Borneo 85
Gyn
acan
tha
limba
lis
+G
ynac
anth
a do
hrni
+
Heli
aesch
na id
ae
+O
ligoa
eschn
a m
utat
a +
Tetra
cant
hagy
na
dego
rsi
+Te
traca
ntha
gyna
pl
agia
ta
++
+g
omph
idae
Ictin
ogom
phus
de
cora
tus
++
+Le
ptog
omph
us
coom
ansi
+M
acro
miid
aeEp
opht
halm
ia
vitti
gera
+
++
+M
acro
mia
cinc
ta
++
Cor
dulii
dae
Hem
icord
ulia
tene
ra
+Li
bellu
lidae
Aciso
ma
pano
rpoi
des
+Ae
thria
man
ta g
racil
is +
Agrio
nopt
era
insig
nis
+Ag
riono
pter
a sex
linea
ta
+Br
achy
dipl
ax ch
alyb
ea
++
+Br
achy
dipl
ax fa
rinos
a +
++
+Br
achy
goni
a oc
ulat
a +
++
++
Cra
tilla
met
allic
a +
+D
ipla
code
s triv
ialis
+
++
Hyd
roba
sileu
s cro
ceus
+
Lyrio
them
is bi
appe
ndicu
lata
+
+
BugiDjam
ForeDeva
LakeNatu
StreRhin
SwamDjam
SwamNepe
SwamRoot
SwamTraH
Camp1
Camp3
Cemetery
PosUlin
StreamC1
StreamC2
SwamBurn
SwamGra1
SwamGra2
EcoCircl
OtterRiv
WainBrid
WainGaze
WainNatu
WoodPath
BridCulm
BugiChan
BugiOutf
DamReser
DamWetle
Fielpath
SwamFiel
WainUppe
APP
END
IX (c
ontin
ued)

A. Dolný et alii86
BugiDjam
ForeDeva
LakeNatu
StreRhin
SwamDjam
SwamNepe
SwamRoot
SwamTraH
Camp1
Camp3
Cemetery
PosUlin
StreamC1
StreamC2
SwamBurn
SwamGra1
SwamGra2
EcoCircl
OtterRiv
WainBrid
WainGaze
WainNatu
WoodPath
BridCulm
BugiChan
BugiOutf
DamReser
DamWetle
Fielpath
SwamFiel
WainUppe
Lyrio
them
is cle
is +
+N
anno
phya
pyg
mae
a +
++
+N
esoxe
nia
linea
ta
+ N
euro
them
is flu
ctuan
s +
++
++
+N
euro
them
is ra
mbu
rii
++
++
++
++
Neu
roth
emis
term
inat
a +
Ony
chot
hem
is co
ccin
ea
++
+O
nych
othe
mis
culm
inico
la
++
++
Orc
hith
emis
pulch
errim
a +
++
Ort
hetr
um ch
rysis
+
++
+O
rthe
trum
sabi
na
++
++
Ort
hetr
um te
stace
um
++
++
Pant
ala
flave
scens
+
Porn
othe
mis
serra
ta
++
Rhod
othe
mis
rufa
+
++
Rhyo
them
is ob
soles
cens
+
++
+Rh
yoth
emis
phyll
is +
Rhyo
them
is tri
angu
laris
+
++
Tetra
them
is fla
vesce
ns
++
+Te
trath
emis
irreg
ular
is +
+Th
olym
is til
larg
a+
+Tr
amea
tran
smar
ina
+Tr
ithem
is au
rora
++
+Ty
rioba
pta
laid
law
i+
++
+Ty
rioba
pta
torr
ida
++
++
++
++
++
++
++
Uro
them
is sig
nata
++
+Zy
xom
ma
petio
latu
m+
APP
END
IX (c
ontin
ued)
Copyright © 2022 FDOKUMEN




















