
Do chitons have a compass? Evidence for magnetic sensitivity in Polyplacophora
14
Do chitons have a compass? Evidence for magnetic sensitivity in Polyplacophora Lauren H. Sumner-Rooney a,b *, James A. Murray c , Shaun D. Cain d and Julia D. Sigwart a,b a 5 Queen’s University Marine Laboratory, Portaferry, Co. Down, Northern Ireland; b School of Biological Sciences, Queen’s University Belfast, Belfast, Northern Ireland; c Department of Biological Sciences, California State University, Hayward, CA, USA; d Department of Biology, Eastern Oregon University, La Grande, OR, USA. (Received 18 November 2013; accepted 21 August 2014) 10 Several animals and microbes have been shown to be sensitive to magnetic fields, though the exact mechanisms of this ability remain unclear in many animals. Chitons are marine molluscs which have high levels of biomineraliz © ed magnetite coating their radulae. This discovery led to persistent anecdotal suggestions that they too may be able to navigationally respond to magnetic fields. Several 15 researchers have attempted to test this, but to date there have been no large- scale controlled empirical trials. In the current study, four chiton species (Katharina tunicata, Mopalia kennerlyi, Mopalia muscosa and Leptochiton rugatus, n = 24 in each) were subjected to natural and artificially rotated magnetic fields while their movement through an arena was recorded over four hours. Field 20 orientation did not influence the position of the chitons at the end of trials, possibly as a result of the primacy of other sensory cues (i.e. thigmotaxis). Under non-rotated magnetic field conditions, the orientation of subjects when they first reached the edge of an arena was clustered around 309–345° ( © north– © north-west) in all four species. However, orientations were random under the 25 rotated magnetic field, which may indicate a disruptive effect of field rotation. This pattern suggests that chitons can detect and respond to magnetism. Keywords: behaviour; magnetoreception; navigation; orientation Introduction Sensitivity AQ1 and behavioural responses to changes in magnetic fields have been demon- 30 strated in such diverse taxa as birds, bacteria and insects, but despite years of study, the precise mechanism or mechanisms remain elusive (Kirschvink et al. 2001; Johnsen and Lohmann 2005; Mouritsen and Hore 2012). The presence of biogenic magnetite in many of these species is a consistent feature, with the movement of grains called magnetosomes in bacteria providing magnetic moment which alters cell orientation 35 when propelled (Schüler 2006). Magnetite is also present in magnetically sensitive animals such as sockeye salmon, honeybees, and several birds (Kirschvink and Gould 1981); however, our understanding of the exact mechanisms of detection is far from complete (Johnsen and Lohmann 2005). However there are also purported mechan- isms which do not centre on magnetite. For example, electromagnetic induction in 40 elasmobranchs would rely only upon the ampullae of Lorenzini, jelly-filled structures *Corresponding author. Email: [email protected] Journal of Natural History, 2014 Vol. 00, No. 00, 1–14, http://dx.doi.org/10.1080/00222933.2014.959574 © 2014 Taylor & Francis
Transcript of Do chitons have a compass? Evidence for magnetic sensitivity in Polyplacophora

Do chitons have a compass? Evidence for magnetic sensitivity inPolyplacophora
Lauren H. Sumner-Rooneya,b*, James A. Murrayc, Shaun D. Caind
and Julia D. Sigwarta,b
a 5Queen’s University Marine Laboratory, Portaferry, Co. Down, Northern Ireland; bSchool ofBiological Sciences, Queen’s University Belfast, Belfast, Northern Ireland; cDepartment ofBiological Sciences, California State University, Hayward, CA, USA; dDepartment of Biology,Eastern Oregon University, La Grande, OR, USA.
(Received 18 November 2013; accepted 21 August 2014)
10Several animals and microbes have been shown to be sensitive to magnetic fields,though the exact mechanisms of this ability remain unclear in many animals.Chitons are marine molluscs which have high levels of biomineraliz©ed magnetitecoating their radulae. This discovery led to persistent anecdotal suggestions thatthey too may be able to navigationally respond to magnetic fields. Several
15researchers have attempted to test this, but to date there have been no large-scale controlled empirical trials. In the current study, four chiton species(Katharina tunicata, Mopalia kennerlyi, Mopalia muscosa and Leptochiton rugatus,n = 24 in each) were subjected to natural and artificially rotated magnetic fieldswhile their movement through an arena was recorded over four hours. Field
20orientation did not influence the position of the chitons at the end of trials,possibly as a result of the primacy of other sensory cues (i.e. thigmotaxis).Under non-rotated magnetic field conditions, the orientation of subjects whenthey first reached the edge of an arena was clustered around 309–345° (©north–©north-west) in all four species. However, orientations were random under the
25rotated magnetic field, which may indicate a disruptive effect of field rotation.This pattern suggests that chitons can detect and respond to magnetism.
Keywords: behaviour; magnetoreception; navigation; orientation
Introduction
Sensitivity
AQ1
and behavioural responses to changes in magnetic fields have been demon-30strated in such diverse taxa as birds, bacteria and insects, but despite years of study,
the precise mechanism or mechanisms remain elusive (Kirschvink et al. 2001; Johnsenand Lohmann 2005; Mouritsen and Hore 2012). The presence of biogenic magnetitein many of these species is a consistent feature, with the movement of grains calledmagnetosomes in bacteria providing magnetic moment which alters cell orientation
35when propelled (Schüler 2006). Magnetite is also present in magnetically sensitiveanimals such as sockeye salmon, honeybees, and several birds (Kirschvink and Gould1981); however, our understanding of the exact mechanisms of detection is far fromcomplete (Johnsen and Lohmann 2005). However there are also purported mechan-isms which do not centre on magnetite. For example, electromagnetic induction in
40elasmobranchs would rely only upon the ampullae of Lorenzini, jelly-filled structures
*Corresponding author. Email: [email protected]
Journal of Natural History, 2014Vol. 00, No. 00, 1–14, http://dx.doi.org/10.1080/00222933.2014.959574
© 2014 Taylor & Francis

on the snout which act as conducting materials which, when moved through amagnetic field, generate an electric current then detected by electrosensitive neuronsin the head (Johnsen and Lohmann 2005). In addition, bird navigation is thought toinvolve the use of cryptochromes and photoreception to maintain magnetic headings
45in migration (Mouritsen and Hore 2012), and not solely relying on magnetite.Magnetoreception is known to exist in molluscs (Lohmann and Willows 1987;
Lohmann et al. 1991) and magnetically sensitive neurons have indeed been identifiedin the nudibranch Tritonia diomedea (Wang et al. 2003; Cain et al. 2006). Workingwith invertebrate systems such as molluscs has helped to identify individual neurons
50due to their relative simplicity (the T. diomedea nervous system contains around 7©000neurons). Like many other invertebrates, they use a polar compass rather thandetecting inclination angles, meaning that they are not affected by differencesbetween magnetic hemispheres (Wiltschko and Wiltschko 1995).
Chitons are marine molluscs that are found from the intertidal into the abyss and55generally feed on algae and diatoms found on rocky substrata. Like many other
molluscs, chitons feed using a radula, a toothed ribbon-like structure with which itrasps against the substratum to remove organic materials. Apparently as an adapta-tion to prevent rapid wear on rocky substrata, the radulae in many taxa are biomi-neraliz©ed, for example in chitons and some limpets (Shaw et al. 2010). Chiton radulae
60are unusual in that the cusps of the large scraping teeth (major laterals) are coatedwith a cap of biomineraliz©ed magnetite, containing up to 29% by weight magnetiteper tooth (Lowenstam 1962; Li et al. 1989). Consequently, chiton radulae exhibitmagnetic properties and radulae can be easily manipulated using magnets (Tomlinson1959; Lowenstam 1962; Carefoot 1965). The ability of chitons to produce biogenic
65magnetite has led to the suggestion that the animals may be equipped to sensemagnetic fields which could be used in navigation (Lowenstam 1962; Carefoot1965), a hypothesis which has been reiterated several times and made its way intoreview literature (Boyle 1977; Kirschvink and Gould 1981; Kirschvink 1997).However, there is no reason to infer any connection between the radula (a feeding
70structure) and a putative magnetic sensory ability in the organism. The ability of theanimals to synthesiz©e magnetite would simply enable them to employ its use outsideof the radula as well.
It is true that many species of chiton display homing behaviour using an unclearmechanism (Crozier 1921; Thorne 1968; Mook 1985; Chelazzi et al. 1988; Kamardin
751989). However, past observations and experiments concerning chiton orientationbehaviour in response to magnetism have given mixed results (Ratner and Jennings1968; Ratner 1976; Tomlinson et al. 1980). Many of these studies have been ham-pered by methodological problems such as sample sizes, length of experiments andstatistical analyses (see discussion). Further, no anatomical evidence for any sensory
80tissues in or around magnetic material (magnetite) has ever been presented in chitons,though these have proved hard to identify in other organisms as well. As a result ofthe poor evidence for magnetic orientation in chitons, many authors have avoidedreferring directly to magnetoreception in chitons, but even so, they are sometimesmentioned in papers reviewing magnetoreception in other organisms (e.g. Kirschvink
85and Gould 1981). Although an intriguing and persistent idea, in fact there is very littlecredible evidence for a magnetic sense in chitons in the literature, and the notion thatit may exist has persisted in the collective scientific conscience as a result of oralhistory and anecdotes.
2 L.H. Sumner-Rooney et al.

In the current study, we used behavioural experiments on four species of chiton to90provide the first robust empirical evidence on whether they can detect and respond to
the orientation of a magnetic field.
Materials and methods
Specimens
Specimens of Leptochiton rugatus (Carpenter in Pilsbry, 1892AQ2 ), Katharina tunicata95(Wood, 1815AQ3 ), Mopalia kennerlyi Carpenter, 1864AQ4 and Mopalia muscosa (Gould,
1846AQ5 ) were collected from multiple sites in the intertidal of San Juan Island (ArgyleCreek, Snug Harbor, Eagle Cove,©WA, USA) and moved to aquarium facilities at theFriday Harbor Laboratories, University of Washington, WA, USAAQ6 . These are fourrelatively common taxa in this region which also represent a decent taxonomic
100spread. Prior to experiments, animals were kept in sea tables supplied with flow-through seawater at local ambient temperature (10–14°C).
Behavioural experiments
Chitons (n = 24 for each species) were placed into individual dishes (diameter 10 cm)and kept at constant temperature in a water bath supplied with flow-through sea-
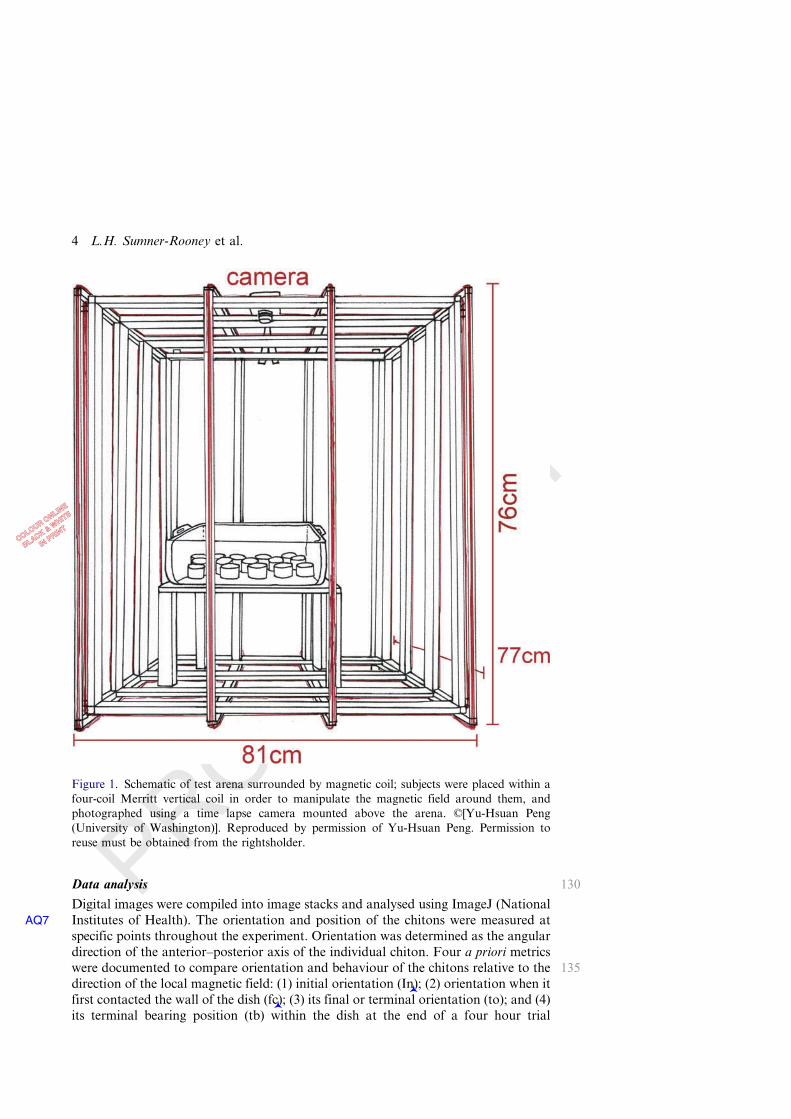
105water (i.e. in the same conditions as the holding tanks). In order to manipulate themagnetic field around the subjects, the water bath was placed within a four-coilMerritt vertical coil (Figure 1) powered by BK Precision 1760 Triple output DCpower supplies (Kirschvink 1992). When the coil was off, the intensity of the magneticfield at the recording site was 53.0 µT and inclination was 76.9°©. When the coil was
110turned on, changes to the total magnetic intensity and inclination angles were mini-mal (the total intensity of the magnetic field was changed by +0.2% and the inclina-tion angle by +0.4%, see Wang et al. 2004). By adjusting the current flowing throughadjacent coils, the direction of the field within could be manipulated. Subjects wereexposed to either the non-rotated ambient magnetic field (‘non-rotated’ hereafter) or
115to a field artificially rotated clockwise by 89–91° (‘rotated’ hereafter); the treatmentwas selected at random by coin flip for each group. Individuals’ positions weredigitally recorded by a camera mounted above the arena at 10 minute intervals,with a compass present in each frame to confirm the magnetic field direction. Eachtrial was run three times. All experiments were performed in identical artificial 40 W
120fluorescent light. As images were captured automatically, it was also unnecessary forthe authors to enter the coil during experiments so there was no mechanical inter-ference with the subjects. The height of the individual dishes was above the surround-ing water height, so there was no mechanical disruption from the flow-through waterwhile it maintained a stable temperature. Experiments lasted for a four-hour period,
125after which subjects were repositioned to the central part in the arena which was thenspun to a new haphazard orientation. On being positioned in the arena, animals wereallowed time to uncurl and then assume normal exploratory behaviour, within thefour-hour experimental window. At the end of three sequential trials, subjects werereturned to a holding tank and not subsequently re-used.
Journal of Natural History 3
130Data analysis
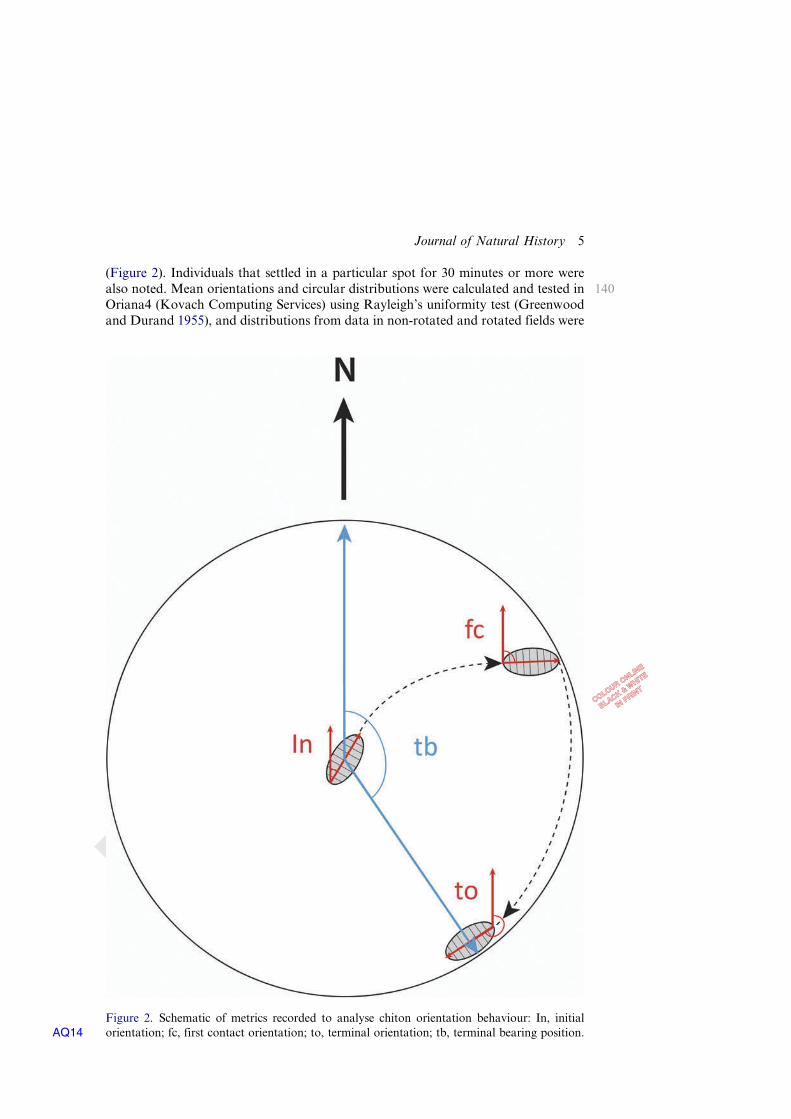
Digital images were compiled into image stacks and analysed using ImageJ (NationalInstitutes of HealthAQ7 ). The orientation and position of the chitons were measured atspecific points throughout the experiment. Orientation was determined as the angulardirection of the anterior–posterior axis of the individual chiton. Four a priori metrics
135were documented to compare orientation and behaviour of the chitons relative to thedirection of the local magnetic field: (1) initial orientation (In©); (2) orientation when itfirst contacted the wall of the dish (fc©); (3) its final or terminal orientation (to); and (4)its terminal bearing position (tb) within the dish at the end of a four hour trial
Figure 1. Schematic of test arena surrounded by magnetic coil; subjects were placed within afour-coil Merritt vertical coil in order to manipulate the magnetic field around them, andphotographed using a time lapse camera mounted above the arena. ©[Yu-Hsuan Peng(University of Washington)]. Reproduced by permission of Yu-Hsuan Peng. Permission toreuse must be obtained from the rightsholder.
4 L.H. Sumner-Rooney et al.
(Figure 2). Individuals that settled in a particular spot for 30 minutes or more were140also noted. Mean orientations and circular distributions were calculated and tested in
Oriana4 (Kovach Computing Services) using Rayleigh’s uniformity test (Greenwoodand Durand 1955), and distributions from data in non-rotated and rotated fields were
Figure 2. Schematic of metrics recorded to analyse chiton orientation behaviour: In, initialorientation; fc, first contact orientation; to, terminal orientation; tb, terminal bearing positionAQ14 .
Journal of Natural History 5

compared using the Mardia–Watson–Wheeler test (Fisher 1993). Animals that didnot move from their initial position at the centre of the dish were excluded from
145analyses.
Results
All following orientations are given relative to magnetic©north (hereafter 0°/360°)within the particular trial’s local magnetic field.
Specimens were placed in the test arena oriented haphazardly. Many started the150trial in a curled posture (a reaction to handling) and were therefore not initially
upright. The initial orientation is primarily a measure of starting placement, but waspartially influenced by some individuals’ movements in uncurling. In general, theinitial orientation of subjects was not significantly different from the expected uni-form circular distribution in almost all cases (Rayleigh test, p > 0.05). However, this
155was not the case in Katharina tunicata under rotated magnetic field (µ = 297.15°,Z = 4.337, p = 0.012) or Mopalia kennerlyi in a non-rotated magnetic field(µ = 340.9°, Z = 5.816, p = 0.003), which both displayed distributions significantlyskewed towards certain orientations.
A minority of subjects remained stationary at the centre of the arena during the160trial (Katharina tunicata, 7.5%; Leptochiton rugatus, 25.3%; Mopalia kennerlyi,
22.3%; Mopalia muscosa, 30.65%) and were therefore excluded from all analyses.The vast majority of non-stationary subjects (96.3%) moved from the centre of theirdish to the edge, making contact with the arena wall.
Under the non-rotated magnetic field, for three out of four species, their orientation165distributions at first contact with the wall were significantly different from a uniform
distribution (Rayleigh test, p ≤ 0.015, see Table 1). That is, although individuals beganthe experiment facing all directions (In, our first separate metric) they turned andclustered around the same mean orientation by the point of contact with the edge ofthe dish. This occurred in K. tunicata (µ = 312°, Z = 4.19, p = 0.015),Mopalia muscosa
170(µ = 337°,Z = 6.378, p = 0.001), andM. kennerlyi (µ = 344°, Z = 15.428, p = 2.0e−7). InLeptochiton rugatus, mean orientation at first contact followed the same trend but wasnot significantly different to random at α = 0.05 (µ = 309°, Z = 2.834, p = 0.059). Themean orientations at contact with the wall were north–north-west (309©–345°) in all fourspecies under the non-rotated field (Figure 3, Table 1).
175Under the experimentally rotated magnetic field, there was no clear preferredheading (i.e. first contact orientation was uniformly distributed; Rayleigh test, p >0.05) with the exception of M. kennerlyi, which also moved toward north-west(µ = 276°, Z = 2.99, p = 0.049).
At the end of all trials in the non-rotated magnetic field, terminal orientation180and terminal bearing of subjects were uniformly distributed in all four species
(Rayleigh test, p > 0.05). Terminal orientations and terminal position bearingswere also uniformly distributed under the artificially rotated field, with the excep-tion of the terminal bearing position of K. tunicata toward west–north-west(µ = 296°, Z = 6.344, p = 0.001).
185There was no significant difference between the distribution of subjects’ orien-tations under non-rotated versus rotated magnetic fields for any of our a priorimetrics, in nearly all of the trials (Mardia–Watson–Wheeler test, p > 0.05). Thesole exception to this is the orientation of M. muscosa at first contact with the dish
6 L.H. Sumner-Rooney et al.
wall which differed significantly in rotated (no significant mean orientation) and190non-rotated (mean north-west) conditions (W = 6.087, p = 0.048, Table 2).
The frequency of participating chitons settling in one place for 30 minutes or longerduring the trial was not affected by field orientation in three of the four species (χ2 tests,p > 0.05) but settling behaviour was found to be significantly more likely to occur inM.kennerlyi in the non-rotated field relative to the rotated field (χ2 = 7.164, df = 1,
195p = 0.0074).
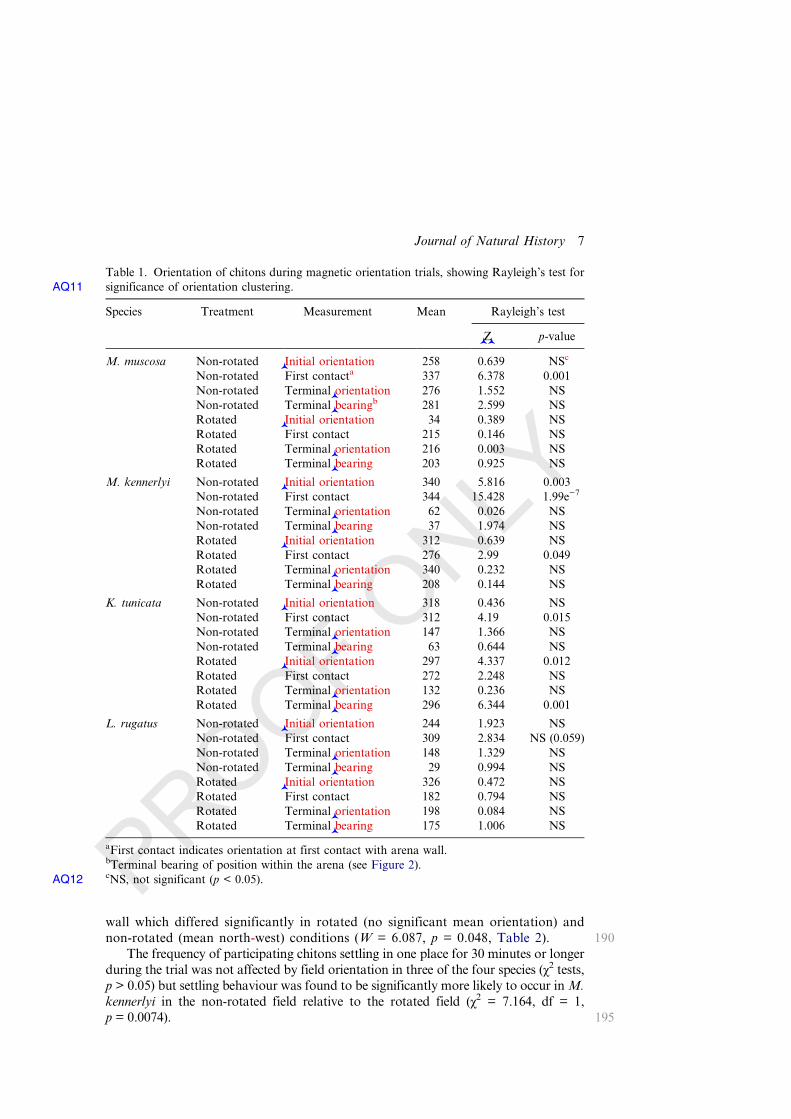
Table 1. Orientation of chitons during magnetic orientation trials, showing Rayleigh’s test forsignificance of orientation clusteringAQ11 .
Species Treatment Measurement Mean Rayleigh’s test
©Z© p-value
M. muscosa Non-rotated ©Initial orientation 258 0.639 NSc
Non-rotated First contacta 337 6.378 0.001Non-rotated Terminal©orientation 276 1.552 NSNon-rotated Terminal©bearing
b 281 2.599 NSRotated ©Initial orientation 34 0.389 NSRotated First contact 215 0.146 NSRotated Terminal©orientation 216 0.003 NSRotated Terminal©bearing 203 0.925 NS
M. kennerlyi Non-rotated ©Initial orientation 340 5.816 0.003Non-rotated First contact 344 15.428 1.99e−7
Non-rotated Terminal©orientation 62 0.026 NSNon-rotated Terminal©bearing 37 1.974 NSRotated ©Initial orientation 312 0.639 NSRotated First contact 276 2.99 0.049Rotated Terminal©orientation 340 0.232 NSRotated Terminal©bearing 208 0.144 NS
K. tunicata Non-rotated ©Initial orientation 318 0.436 NSNon-rotated First contact 312 4.19 0.015Non-rotated Terminal©orientation 147 1.366 NSNon-rotated Terminal©bearing 63 0.644 NSRotated ©Initial orientation 297 4.337 0.012Rotated First contact 272 2.248 NSRotated Terminal©orientation 132 0.236 NSRotated Terminal©bearing 296 6.344 0.001
L. rugatus Non-rotated ©Initial orientation 244 1.923 NSNon-rotated First contact 309 2.834 NS (0.059)Non-rotated Terminal©orientation 148 1.329 NSNon-rotated Terminal©bearing 29 0.994 NSRotated ©Initial orientation 326 0.472 NSRotated First contact 182 0.794 NSRotated Terminal©orientation 198 0.084 NSRotated Terminal©bearing 175 1.006 NS
aFirst contact indicates orientation at first contact with arena wall.bTerminal bearing of position within the arena (see Figure 2).cNS, not significant (p < 0.05AQ12 ).
Journal of Natural History 7

Discussion
The evidence here strongly suggests that at least some species of chitons are sensitiveto the orientation of local magnetic fields; their heading in the absence of otherstimuli tends toward north-west, yet this is disrupted in a rotated magnetic field.
Figure 3. Orientation of chitons at first contact with the arena wall in non-rotated and rotatedmagnetic fields. Vector indicates mean orientation at significance of α = 0.05 or less (indicatedby inner circle). Compass directions are those as experienced within a given experimental arena.
Table 2. Comparison of orientation distribution metrics between rotated and non-rotatedmagnetic fields using Mardia–Watson–Wheeler tests.
Species Metric (nnon-rotated and nrotated) Wa p
M. muscosa Initial (41 and 45) 1.851 NSb
First contact (41 and 45) 6.087 0.048Terminal orientation (35 and 40) 1.574 NSTerminal bearing (35 and 40) 0.361 NS
M. kennerlyi Initial (78 and 43) 2.889 NSFirst contact (78 and 43) 1.271 NSTerminal orientation (71 and 48) 0.172 NSTerminal bearing (71 and 48) 1.819 NS
K. tunicata Initial (67 and 45) 3.658 NSFirst contact (67 and 45) 2.782 NSTerminal orientation (36 and 27) 1.137 NSTerminal bearing (36 and 27) 5.503 NS
L. rugatus Initial (49 and 54) 6.9 0.032First contact (50 and 42) 0.953 NSTerminal orientation (50 and 54) 1.53 NSTerminal bearing (45 and 47) 3.87 NS
aW, test statisticAQ13 .bNS, not significant (p > 0.05).
8 L.H. Sumner-Rooney et al.

200Under a non-rotated magnetic field, chitons travelled in a non-random pre-ferred direction (generally north-west) until they reached the wall of the dish. Inthe large majority of cases, chitons moved directly from their starting position atthe centre of the dish to the edge, without noticeably changing direction inbetween. Their direction of travel is also independent of their initial orientation.
205Under controlled experimental conditions (i.e. in the absence of other stimuli),chitons are apparently able to use magnetic fields to determine their orientation.We cannot totally exclude the possibility that chitons in these experiments wereresponding to some other uncontrolled stimulus, but the disruption of this strongorientation pattern under an artificial change in the magnetic field indicates a
210sensitivity to magnetism.Leptochiton rugatus followed the same trend as the other three test species, but
did not demonstrate statistically significant clustering to any particular orienta-tion; however, practical constraints may be to blame in that adult L. rugatus aremuch smaller than the other three species, but in our experiments©the arenas used
215were the same size©in all trials. As a result, fewer of the L. rugatus subjectsreached the edge of the arena during the trial period, resulting in smaller qualify-ing datasets and an intrinsically lower likelihood that any behavioural responsewould reach a level of statistical significance. This is one possibility that couldexplain why the distribution of orientation at first contact with the wall under
220non-rotated magnetic field for L. rugatus was clustered around a mean north-westerly bearing but fell short of statistical significance (p = 0.059) at the 0.05level. Interestingly, L. rugatus is notably different from the other test subjects inthat it is within a phylogenetically separate order (Lepidopleurida). The two livingorders within Polyplacophora (Lepidopleurida and Chitonida) possess markedly
225different sensory organs (reviewed in Sigwart et al. 2014). It is therefore alsospeculatively plausible that they may employ different sensory cells or pathwaysto determine orientation behaviour.
The other three species used in our experiments (M. muscosa, M. kennerlyi©and K.tunicata) are within a single taxonomic family, Mopalidae. Differences in magnetic
230sensitivity between species were reported previously by Tomlinson et al. (1980), whonoted that M. muscosa and K. tunicata demonstrated particularly strong apparentresponses to magnetism compared to Nuttallina californica. Nuttallina also belongs tothe superfamily Mopalioidea but is more phylogenetically distant than the other twogenera (Sirenko et al. 2013). There is therefore room for speculation about species-
235level differences in orientation behaviour and magnetic sensitivity, but on balance theevidence of the current experiments suggests that across a broad taxonomic samplingfrom one biogeographic region, and in absence of other stimuli, chitons tend towardnorth-west in this test.
Within an artificially rotated magnetic field, one might intuitively expect the240animals to immediately adjust their orientation according to the subjective field
direction. Aside from field orientation, there were no differences between the twoexperimental conditions; thus we would expect that any response should be preservedand the mean orientation rotated accordingly. However,©we did not observe thisresponse in any of the species tested.
245There are several biological reasons why animals with genuine magnetic sensitiv-ity may not immediately follow a clear orientation under an experimentally manipu-lated magnetic field. First, subjects are likely to use a combination of orientation
Journal of Natural History 9

cues, of which magnetic field orientation is only one component (Able 1991;Wiltschko and Wiltschko 1994). Within the experimental framework, when the field
250is rotated using the coil, magnetic cues become misaligned with any other cues presentin the arena environment and therefore the animals’ overall response may be con-founded by this conflict of information (Wiltschko et al. 1998). In addition, therotation of the field by the coil is instantaneous and occurred after the animalswere placed inside the arena. Such a rapid shift would never occur within an animal’s
255natural experience (Bullard et al. 1950). The rotated field in these experiments was infact marginally (0.2%) stronger than the Earth’s (ambient) magnetic field (around 53µT); some magnetosensitive organisms have decreased activity in stronger thanambient magnetic fields, and move and rotate more in weakened magnetic fields(Wiltschko and Wiltschko 1995). Combined, the conflicting cues and sudden change
260in field orientation and strength could cause a chaotic response, thus explaining therandom distribution of first-contact orientation found under rotated conditions (inthree species, all but M. kennerlyi). The actual change in strength is considered to bevery minimal and compares well to other experimental magnetic studies (Wiltschkoand Wiltschko 1995). Nevertheless, that chitons do behave differently in non-rotated
265and rotated magnetic fields strongly indicates that they could be influenced by thechange.
Although the magnetic field affected the direction of first contact (fc) with thearena wall,©subsequent orientation was random. No species showed significant clus-tering in terminal orientation under either ambient or experimental magnetic fields
270(Rayleigh test, p >0.05). Katharina tunicata was the only species that had a significantclustering in mean terminal bearing (i.e. position within the arena, see Figure 2), asanimals were generally positioned in the (experimental) north-west side of the dishunder the artificially rotated field. Once the chitons encountered the tactile cuesprovided by the wall of the dish they generally no longer displayed any clustered
275orientation or differences between magnetic field treatments. When they reached theedge of the dish, and they usually did so head-on, they were effectively forced to actby turning either left or right. From this point onward, the edge itself provides asecond, tactile, sensory cue and the chitons behaved thigmotactically, often circlinground the edge for the remainder of the trial. In the natural world many other more
280complex sensory cues would be available and it seems likely that these other cues maybe dominant sensory inputs.
Contrary to the findings of some previous studies, we did not observe chitonsdisplaying any preferred orientation relative to magnetic n©orth at the end of trials(contra Tomlinson et al. 1980). If magnetic orientation was used in navigation, we
285would expect to see an innate preference for settling facing a particular direction,which might indicate a role in homing or settling behaviours, as has been found inother organisms which use magnetic navigation (Wiltschko and Wiltschko 2005).Additionally, there were no significant differences in the nature of subjects’ finaldistributions under the two different fields.
290The magnetic properties of chiton radulae were first observed by Thorpe (fideTomlinson 1959) and confirmed by Tomlinson (1959) and others (Carefoot 1965;Eibschütz et al. 1967); although not studied herein, the radula is central to the originsof interest in chitons’ magnetic sensitivity. The composition of the tooth cappingmaterials was determined by Lowenstam (1962) who suggested that the presence of
295large quantities of magnetite could facilitate magnetic orientation behaviour. The
10 L.H. Sumner-Rooney et al.

basis of this supposition is flawed, owing to the absence of single-domain magnetite inthe chiton radula. However, magnetoreception is widespread in molluscs and it wouldbe a plausible suggestion that chitons may be included also, irrespective of the radulacomposition. Since then, several authors described apparent magnetic responsive
300behaviours (Ratner and Jennings 1968; Ratner 1976; Tomlinson et al. 1980), butthese studies provided mixed results and varied in their quality and scientific rigour.Some of the multiple experiments within these studies used very low sample sizes ofseven to 14 animals (Ratner and Jennings 1968; Ratner 1976), or inappropriatelysimplified statistics, such as categorically comparing ‘northerly’ versus ‘southerly’
305facing subjects (Tomlinson et al. 1980). The use of field observations of chitonorientation (Tomlinson et al. 1980), which have been the subject of many unpublishedstudent reports, may also be unreliable due to the presence of many other stimuli. Forexample, it has been demonstrated that chitons and other molluscs may orientaterelative to the direction of the shore (Focardi and Chelazzi 1990; Minchin 1992). In
310the case of Tomlinson et al. (1980), fieldAQ8 observations were made in a locality incentral California, where ‘northerly’ oriented chitons on the shore were possibly alsomore likely to be facing inland.
Some of the methods historically used for testing sensitivity to magnetic fieldshave also been inappropriate; Tomlinson et al. (1980) placed groups of Mopalia
315muscosa and Katharina tunicata into saucepans made of either aluminium or ironand steel, covered the lids and observed their orientation some hours later. Althoughthey reported significant orientation in aluminium pans and not in the others, theauthors’ reasoning that the iron and steel pans should block out the Earth’s magneticfield was fundamentally flawed (Kirschvink et al. 1987). In other studies, which have
320used supplementary magnets and electromagnetic coils, no statistically significantorientation to field direction was reported; however, as described above, the samplesizes for these experiments were probably too small to have appropriate statisticalpower (Ratner and Jennings 1968; Ratner 1976). Finally, several observations con-cerning magnetic orientation have not been tested statistically, for example that
325responses to magnetism vary seasonally, with ‘summer being the best, and winterthe worst for orientation’ (Tomlinson et al. 1980). The©suggestion that chitonsfloating on pieces of wood in water (in the summer months) will naturally spin topoint north (Tomlinson et al. 1980)© has not been replicated (JDS, personal©observation©2013). Overall, work on magnetic orientation in chitons has produced
330mixed results, with some authors observing statistically significant northerly-facingbehaviour and others failing to do so. Our aim has been to provide further empiricalevidence but avoiding some of the methodological limitations in previous tests ofchitons’ orientation in magnetic fields.
Chitons apparently orientate in a non-random manner under a natural mag-335netic field until they reach an obstacle. Magnetoreception in chitons was first
proposed more than 50 years ago but until now had not been robustly tested.However, the importance of the ability to sense magnetic field orientation may besubstantially less than anecdotal evidence suggests. Although these four specieshave an apparently generally consistent preference toward north-west, as soon as
340other tactile stimuli are available (i.e. the edge of the dish) their orientation andposition within our experimental set-up were no longer magnetically determined.The initial response to magnetism in natural conditions is disrupted under anartificially rotated magnetic field, which further supports the conclusion that the
Journal of Natural History 11

non-random innate tendency toward a north-westerly orientation is inoperable345under experimental manipulation of the magnetic field. Yet in two exceptional
cases under the rotated field – M. kennerlyi at first contact with the wall and theterminal bearing of K. tunicata – local (experimental) north-west is still predomi-nant. The results we have seen from the terminal orientation and position ofchitons would suggest that although it appears that they can detect and respond
350to magnetic field characteristics, the actual importance of this ability may be oflittle significance in realistic natural situations given the size of chitons’ ranges,and that magnetic sensitivity, while clearly present, is potentially still quite low inthe hierarchy of natural environmental cues (e.g. thigmotaxis) used by chitons innavigation.
355Acknowledgements
This project was inspired by a student summer research project by Rosemary Murphy and Yu-Hsuan (Mary) Peng conducted in July 2012 at Friday Harbor Laboratories, WA©, U©S©A. Theauthors also thank the staff at the Friday Harbor Laboratories, and the California StateUniversity East Bay for their support. Finally, thank you to two anonymous reviewers who
360gave much constructive feedback on an original version of this manuscript. This is a contribu-tion to the collection©Who are the ‘Aculifera’? in memory of Christoffer Schander.
References
Able KP. 1991. Common themes and variations in animal orientation systems. Am Zool.31:157–167.
365Boyle PR. 1977. The physiology and behaviour of chitons (Mollusca: Polyplacophora).Oceanogr Mar Biol Annu Rev. 15:461–509.
Bullard EC, Freedman C, Gellman H, Nixon J. 1950. The westward drift of the Earth’smagnetic field. Philos Trans R Soc A Math Phys Eng Sci. 243:67–92. doi:10.1109/20.42443
370Cain SD, Wang JH, Lohmann KJ. 2006. Immunochemical and electrophysiological analyses ofmagnetically responsive neurons in the mollusc Tritonia diomedea. J Comp Physiol ANeuroethol Sens Neural Behav Physiol. 192:235–245. doi:10.1007/s00359-005-0063-8
Carefoot TH. 1965. Magentite in the radula of the Polyplacophora. Proc Malacol Soc London.36:203–213.
375Chelazzi G, Focardi S, Deneubourg J-L. 1988. Analysis of movement patterns and orientationmechanisms in intertidal chitons and gastropods. In: Chelazzi G, Vannini M, editors.Behavioral adaptation to intertidal life. New York: Plenum Press; p. 173–184.
Crozier WJ. 1921. “Homing” behavior in chiton. Am Nat. 55:276–281. doi:10.1086/279812Eibschütz M, Frei EH, Gorodetsky G, Steinitz H. 1967. Nature of the magnetic radula in
380chitons from Eilat on the Red Sea. Nature. 216:1138–1139. doi:10.1038/2161138a0AQ9Fisher NI. 1993. Statistical analysis of circular data. Cambridge: Cambridge University Press.Focardi S, Chelazzi G. 1990. Ecological determinants of bioeconomics in three intertidal
chitons (Acanthopleura spp. J Anim Ecol. 59:347–362. doi:10.2307/5177Greenwood JA, Durand D. 1955. The distribution of the length and components of the sum of
385n random unit vectors. Ann Math Stat. 26:233–246. doi:10.1214/aoms/1177728540Johnsen S, Lohmann KJ. 2005. The physics and neurobiology of magnetoreception. Nat Rev
Neurosci. 6:703–712. doi:10.1038/nrn1745Kamardin NN. 1989. Study of homing in Acanthopleura gemmata (Polyplacophora, Mollusca).
Vestn Leningr Univ Biol. 3:58–62.
12 L.H. Sumner-Rooney et al.

390Kirschvink JL. 1992. Uniform magnetic fields and double-wrapped coil systems: Improvedtechniques for the design of bioelectromagnetic experiments. Bioelectromagnetics.13:401–411. doi:10.1002/bem.2250130507
Kirschvink JL. 1997. Magnetoreception: homing in on vertebrates. Nature. 390:339–340.doi:10.1038/36986
395Kirschvink JL, Gould JL. 1981. Biogenic magnetite as a basis for magnetic field detection inanimals. BioSystems. 13:181–201. doi:10.1016/0303-2647(81)90060-5
Kirschvink JL, Jones DS, MacFadden BJ, editors. 1987. Magnetite biomineralization andmagnetoreception: a new biomagnetism. New York: Plenum Press.
Kirschvink JL, Walker MM, Diebel CE. 2001. Magnetite-based magnetoreception. Curr Opin400Neurobiol. 11:462–467. doi:10.1016/S0959-4388(00)00235-X
Li C-W, Chin TS, Li JS, Huang SH. 1989. Growth of chiton teeth evidenced from magneticmeasurement and microstructure characterization. IEEE Trans Magn. 25:3818–3820.doi:10.1109/20.42443
Lohmann KJ, Willows AO. 1987. Lunar-modulated geomagnetic orientation by a marine405mollusk. Science. (80-). 235:331–334. doi:10.1126/science.3798115AQ10
Lohmann KJ, Willows AO, Pinter RB. 1991. An identifiable molluscan neuron responds tochanges in earth-strength magnetic fields. J Exp Biol. 161:1–24.
Lowenstam HA. 1962. Magnetite in denticle capping in Recent chitons (Polyplacophora). GeolSoc Am Bull. 73:435–438. doi:10.1130/0016-7606(1962)73[435:MIDCIR]2.0.CO;2
410Minchin D. 1992. Multiple species, mass spawning events in an Irish sea lough: the effect oftemperatures on spawning and recruitment of invertebrates. Invertebr Reprod Dev.22:229–238. doi:10.1080/07924259.1992.9672275
Mook D. 1985. Some preliminary observations on the homing of the West Indian chitonAcanthopleura granulata. The Veliger. 28:115–118.
415Mouritsen H, Hore PJ. 2012. The magnetic retina: light-dependent and trigeminal magnetor-eception in migratory birds. Curr Opin Neurobiol. 22:343–352. doi:10.1016/j.conb.2012.01.005
Ratner SC. 1976. Kinetic movements in magnetic fields of chitons with ferro-magnetic struc-tures. Behav Biol. 17:573–578. doi:10.1016/S0091-6773(76)91045-2
420Ratner SC, Jennings JW. 1968. Magnetic fields and orienting movements in mollusks. J CompPhysiol Psychol. 65:365–368. doi:10.1037/h0025543
Schüler D. 2006. Magnetoreception and magnetosomes in bacteria. Volume 3. Berlin: Springer.Shaw JA, Macey DJ, Brooker LR, Clode PL. 2010. Tooth use and wear in three iron-
biomineralizing mollusc species. Biol Bull. 218:132–144.425Sigwart JD, Sumner-Rooney LH, Schwabe E, Hess M, Brennan GP, Schrödl M. 2014. A new
sensory organ in “primitive” molluscs (Polyplacophora: Lepidopleurida), and its contextin the nervous system of chitons. Front Zool. 11:7. doi:10.1186/1742-9994-11-7
Sirenko BI, Abramson NI, Vagapov AI. 2013. Schizoplax brandtii (Middendorff, 1847)(Mollusca: Polyplacophora) – an example of “explosive speciation”? Bull Russ Far East
430Malacol Soc. 17:151–166.Thorne MJ. 1968. Studies on homing in the chiton Acanthozostera gemmata. Aust J Mar
Freshw Res. 19:151–160. doi:10.1071/MF9680151Tomlinson JT. 1959. Magnetic properties of chiton radulae. The Veliger. 2:36.Tomlinson J, Reilly D, Ballering R. 1980. Magnetic radular teeth and geomagnetic responses in
435chitons. The Veliger. 23:167–170.Wang JH, Cain SD, Lohmann KJ. 2003. Identification of magnetically responsive neurons in
the marine mollusc Tritonia diomedea. J Exp Biol. 206:381–388. doi:10.1242/jeb.00076Wang JH, Cain SD, Lohmann KJ. 2004. Indentifiable neurons inhibited by Earth-strength
magnetic stimuli in the mollusc Tritonia diomedea. J Exp Biol. 207:1043–1049.440doi:10.1242/jeb.00864
Journal of Natural History 13

Wiltschko R, Wiltschko W. 1994. Avian orientation: multiple sensory cues and the advantageof redundancy. In: Davies MO, Green P, editors. Percept ot control birds SE - 6. Berlin:Springer; p. 95–119.
Wiltschko R, Wiltschko W. 1995. Magnetic orientation in animals. Berlin: Springer-Verlag.445Wiltschko W, Wiltschko R. 2005. Magnetic orientation and magnetoreception in birds and
other animals. J Comp Physiol A. 191:675–693. doi:10.1007/s00359-005-0627-7Wiltschko W, Weindler P, Wiltschko R. 1998. Interaction of magnetic and celestial cues in the
migratory orientation of passerines. J Avian Biol. 29:606–617. doi:10.2307/3677181
14 L.H. Sumner-Rooney et al.




















