
DNA microarrays in the clinic: infectious diseases
10
DNA microarrays in the clinic: infectious diseases Vladimir Mikhailovich, 1 Dmitry Gryadunov, 1 Alexander Kolchinsky, 2 Alexander A. Makarov, 1 * and Alexander Zasedatelev 1 * Summary We argue that the most-promising area of clinical application of microarrays in the foreseeable future is the diagnostics and monitoring of infectious diseases. Microarrays for the detection and characterization of human pathogens have already found their way into clinical practice in some countries. After discussing the persistent, yet often underestimated, importance of infectious diseases for public health, we consider the technologies that are best suited for the detection and clinical investigation of pathogens. Clinical application of microarray technologies for the detection of mycobac- teria, Bacillus anthracis, HIV, hepatitis and influenza viruses, and other major pathogens, as well as the analysis of their drug-resistance patterns, illustrate our main thesis. BioEssays 30:673–682, 2008. ß 2008 Wiley Periodicals, Inc. Introduction The FDA discriminates two types of diagnostics based on the analysis of nucleic acids (nucleic acids testing, NAT): genetic, which ‘‘determines the presence or absence of a particular, targeted DNA sequence already known...to be related to a health outcome; and genomic, which measures gene expres- sion, usually by mRNA as surrogate.’’ (1) In a recent review of the clinical applications of microarrays, Jordan surveyed both approaches for the detection of clinically relevant features in the genes and gene expression patterns of the patients. (2) The author concluded: ‘‘...(T)he adoption of DNA arrays is taking place at a slower rate than expected...’’ This is not surprising, however, when we take into account that most of the reviewed applications target cancer, which remains the highest, yet at the same time the most intractable, target of such translational research. Historically, cancer and cardiovascular diseases only became the major causes of mortality relatively recently, first in developed countries, then gradually in most of the developing world. The radical increase of longevity between 1900 and 1950 was due to the introduction of new chemo- therapies against infectious diseases and, in the following two decades, to the reduction in infant mortality resulting from better maternal care and a decrease in malnutrition. Translational research and fundamentally new approaches have not played significant roles in the overall improvement of cancer treatment, although some relatively rare cancers became curable. One striking example of the failure of cancer-related discoveries to influence clinical progress is the history of p53. (3) Since its discovery almost 30 years ago, at least 4 billion dollars have been spent on p53-related research, (4) because it has been assumed that ‘‘determining the p53 status of cancers would rapidly acquire major clinical significance’’. Yet, ‘‘(a) recent meta-analysis of...studies in colon cancer has...confirmed anecdotal understanding that this is not the case...(T)his can be seen as a massive failure of translational research.’’ (3) Currently, the impact of infectious disease on mortality in developed and some developing countries is low, about 1–3% of all deaths. At the same time, in other developing countries, it remains much higher. For instance, 20% of all deaths in India are caused by infections while, in Zimbabwe, this figure is as high as 74%, with 63% of all deaths caused by HIV/AIDS. (4) Overall, in 1990, 27% of all deaths in the world were due to infectious causes; (5) in 2002, the last year for which the data are available, this number decreased to 20%. (5) However, accurate death statistics collected by WHO, as noted in many of its own documents, are not available. One of the WHO bulletins on the subject reads: ‘‘There are only 23 countries classified as having high-quality (cause of death) data. There are 28 countries classified as having low-quality data; these include some high-income countries, such as 1 Engelhardt Institute of Molecular Biology, Russian Academy of Sciences, Moscow, Russia. 2 Health Front Line, Ltd., Champaign, IL. Funding agencies: This work has been financially supported by the Russian Academy of Sciences’ Program on Molecular and Cellular Biology and by contracts #02.190.11.30, #02.512.11.2089, #02.512.11.2184 and #02.527.11.9006 with the Federal Agency of Sciences and Innovations of the Russian Federation. *Correspondence to: Alexander Zasedatelev and Alexander A. Makarov, Engelhardt Institute of Molecular Biology, Russian Academy of Sciences, 32 Vavilov St, 119991 Moscow, V-334, Russia. E-mail: [email protected] and [email protected] DOI 10.1002/bies.20781 Published online in Wiley InterScience (www.interscience.wiley.com). BioEssays 30:673–682, ß 2008 Wiley Periodicals, Inc. BioEssays 30.7 673 Abbreviations: NAT, nucleic acid testing; FDA, Food and Drug Administration (USA); POCT, point-of-care testing; S/N, signal/noise ratio; QC, qualitycontrol; HPV, human papilloma virus; HBV, hepatitis B virus; HCV, hepatitis C virus. What’s new?
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of DNA microarrays in the clinic: infectious diseases

DNA microarrays in the clinic:infectious diseasesVladimir Mikhailovich,1 Dmitry Gryadunov,1 Alexander Kolchinsky,2
Alexander A. Makarov,1* and Alexander Zasedatelev1*
SummaryWe argue that the most-promising area of clinicalapplication of microarrays in the foreseeable future isthe diagnostics and monitoring of infectious diseases.Microarrays for the detection and characterization ofhuman pathogens have already found their way intoclinical practice in some countries. After discussing thepersistent, yet often underestimated, importance ofinfectious diseases for public health, we consider thetechnologies that are best suited for the detection andclinical investigation of pathogens. Clinical application ofmicroarray technologies for the detection of mycobac-teria, Bacillus anthracis, HIV, hepatitis and influenzaviruses, and other major pathogens, as well as theanalysis of their drug-resistance patterns, illustrate ourmain thesis. BioEssays 30:673–682, 2008.� 2008 Wiley Periodicals, Inc.
Introduction
The FDA discriminates two types of diagnostics based on the
analysis of nucleic acids (nucleic acids testing, NAT): genetic,
which ‘‘determines the presence or absence of a particular,
targeted DNA sequence already known. . .to be related to a
health outcome; and genomic, which measures gene expres-
sion, usually by mRNA as surrogate.’’(1) In a recent review of
the clinical applications of microarrays, Jordan surveyed both
approaches for the detection of clinically relevant features in
the genes and gene expression patterns of the patients.(2) The
author concluded: ‘‘. . .(T)he adoption of DNA arrays is taking
place at a slower rate than expected. . .’’ This is not surprising,
however, when we take into account that most of the reviewed
applications target cancer, which remains the highest, yet at
the same time the most intractable, target of such translational
research.
Historically, cancer and cardiovascular diseases only
became the major causes of mortality relatively recently, first
in developed countries, then gradually in most of the
developing world. The radical increase of longevity between
1900 and 1950 was due to the introduction of new chemo-
therapies against infectious diseases and, in the following two
decades, to the reduction in infant mortality resulting from
better maternal care and a decrease in malnutrition.
Translational research and fundamentally new approaches
have not played significant roles in the overall improvement of
cancer treatment, although some relatively rare cancers
became curable. One striking example of the failure of
cancer-related discoveries to influence clinical progress is
the history of p53.(3) Since its discoveryalmost 30 years ago, at
least 4 billion dollars have been spent on p53-related
research,(4) because it has been assumed that ‘‘determining
the p53 status of cancers would rapidly acquire major clinical
significance’’. Yet, ‘‘(a) recent meta-analysis of. . .studies in
colon cancer has. . .confirmed anecdotal understanding that
this is not the case. . .(T)his can be seen as a massive failure of
translational research.’’(3)
Currently, the impact of infectious disease on mortality in
developed and some developing countries is low, about 1–3%
of all deaths. At the same time, in other developing countries, it
remains much higher. For instance, 20% of all deaths in India
are caused by infections while, in Zimbabwe, this figure is as
high as 74%, with 63% of all deaths caused by HIV/AIDS.(4)
Overall, in 1990, 27% of all deaths in the world were due to
infectious causes;(5) in 2002, the last year for which the data
are available, this number decreased to 20%.(5)
However, accurate death statistics collected by WHO, as
noted in many of its own documents, are not available. One of
the WHO bulletins on the subject reads: ‘‘There are only
23 countries classified as having high-quality (cause of death)
data. There are 28 countries classified as having low-quality
data; these include some high-income countries, such as
1Engelhardt Institute of Molecular Biology, Russian Academy of
Sciences, Moscow, Russia.2Health Front Line, Ltd., Champaign, IL.
Funding agencies: This work has been financially supported by the
Russian Academy of Sciences’ Program on Molecular and Cellular
Biology and by contracts #02.190.11.30, #02.512.11.2089,
#02.512.11.2184 and #02.527.11.9006 with the Federal Agency of
Sciences and Innovations of the Russian Federation.
*Correspondence to: Alexander Zasedatelev and Alexander A.
Makarov, Engelhardt Institute of Molecular Biology, Russian Academy
of Sciences, 32 Vavilov St, 119991 Moscow, V-334, Russia.
E-mail: [email protected] and [email protected]
DOI 10.1002/bies.20781
Published online in Wiley InterScience (www.interscience.wiley.com).
BioEssays 30:673–682, � 2008 Wiley Periodicals, Inc. BioEssays 30.7 673
Abbreviations: NAT, nucleic acid testing; FDA, Food and Drug
Administration (USA); POCT, point-of-care testing; S/N, signal/noise
ratio; QC, quality control; HPV, human papilloma virus; HBV, hepatitis
B virus; HCV, hepatitis C virus.
What’s new?

Greece and Portugal.’’ Moreover, death statistics for the most
populous countries, India and China, are based on sampling,
not actual count.(6)
It is quite certain that the available vital statistics diminish
the apparent importance of infectious diseases. First,
infections disproportionately affect the younger part of the
population; therefore its share in the so-called ‘‘years of life
lost’’ is higher.(7) Second, the relatively low mortality caused
by infections in developed countries is not exclusively the
result of their low incidence. To some extent, it is the result of
successful management of infections using antibiotics and
antiviral drugs. Third, infections resulting from underlying
chronic conditions may not be reflected in the overall vital
statistics. For instance, the immediate cause of death of
patients with suppressed immunity may be an opportunistic
infection but, in the death certificate, the listed cause will be
their primary condition—HIV or cancer, if the infection
emerged as the result of chemotherapy treatment. Finally,
dangerous and unusual infections can emerge as the result
of bioterrorism.
In addition to the direct impact of infections on mortality,
many chronic diseases may be initially caused by pathogens.
Thus, human papillomavirus is the cause of most cervical
cancers,(8) hepatitis viruses cause liver cancers.(9,10) and the
bacteria Helicobacter pilori is often responsible for stomach
ulcers and can lead to cancer.(11) Moreover, any chronic
inflammation may be a strong risk factor for malignancy.(12)
Contrary to public perception, therefore, infectious dis-
eases remain at the forefront of modern health care, and the
principal problem is that pathogens are constantly moving
targets. All pathogens sooner or later develop resistance to
new drugs, while viruses rapidly evolve into new strains with
better immune tolerance and more efficient transmission.
Generally speaking, infectious diseases are understood
better than many other common conditions, including cancer.
There are large databases of sequences of pathogens; many
genomes are sequenced fully, including different variants and
strains; the genetic basis of drug resistance and immune
tolerance is often (though not always) known. To address this
field of research and clinical diagnostics, National Institute of
Allergies and Infectious Diseases (NIAID), a member of NIH,
created a network of ‘‘Bioinformatics resource centers for
biodefense and emerging or re-emerging infectious dis-
eases’’(13) and created and regularly updates ‘‘List of Emerg-
ing and Re-Emerging Infectious Diseases’’.(14)
This review will complement the aforementioned paper by
Jordan,(2) which did not mention the diagnostics of infectious
diseases. We outline selected microarray-based technologies
and their clinical applications in this field, emphasizing the
most-recently published work. This outline does not attempt to
provide a comprehensive review, and the works chosen for
citation only illustrate the potential of microarrays to detect and
characterize clinically important pathogens.
Microarray technologies suitable for diagnostics
The demands of clinical diagnostics enforced by national
regulatoryagencies are much stricter than the requirements of
experimental technologies encouraged solely by voluntary
compliance. The quality of data obtained using microarrays, as
well as their inter-laboratory compatibility, is an actively
discussed technical challenge (for review, see Ref. 15). This
is especially true in case of genomic testing, when the results
depend on the preparation, storage and processing of
samples and the synthesis of hybridization probes. The
problems are serious enough that the Microarray Quality
Control (MAQC) project created by FDA in the United
States(16) stopped short of recommending microarray meas-
urements of gene expression (genomic testing) for clinical
diagnostics. In contrast, the combination of genetic testing with
low-density arrays and limited sets of oligonucleotides, which
is most frequently used for the diagnostics of infectious
diseases, can give clear qualitative answers and has much
better standing.
The criteria for quality assessment can be categorized into
those applied to the whole arrays (background variance
across the array and within batches of manufactured arrays,
dynamic range of signal level, precision of the grid) and those
related to individual elements of the arrays (S/N ratio for
positive controls, variance in spot size, variance of signal
intensity across the sets of replicates). While a certain
percentage of ‘‘flagged’’ elements is considered acceptable
in experimental settings, QC procedures must reject arrays
intended for clinical diagnostics if they contain faulty elements.
A good example of careful evaluation of a diagnostic DNA
chip was published by Lee et al.(17) The authors tested DNA
arrays for the detection of human papilloma virus (HPV).
These arrays were approved by Korea Food and Drug
Administration (KFDA) and evaluated by the authors in
accordance with KFDA requirements. Interestingly, these
requirements include ‘‘Clinical effectiveness of the device’’,
in contrast, for instance, to CE (conformite europeenne)
certification adopted by EU and limited to safety of the device
and its manufacture process. In this respect, KFDA require-
ments are similar to FDA policy in the United States.
Technology platforms for clinical diagnostics
It appears that, so far, just a few of the multitude of microarray
technologies have proved suitable for clinical applications.
First, this is the technology of photolithographic synthesis
of probes on the surface of quartz wafers developed by
Affymetrix (GeneChip1
). The resulting two-dimensional micro-
arrays became the first to be approved by the FDA for genetic
testing — clinical diagnostics of mutations in two P450
proteins.
Affymetrix GeneChips carry up to 106 individual elements
(features) and are able to re-sequence by hybridization rather
long segments of DNA and identify large number of variations.
What’s new?
674 BioEssays 30.7

Many applications of Affymetrix GeneChips for identification
and typing of pathogens are reviewed at the company web
site(18) and include studies of Streptococcus pyogenes and
other respiratory pathogens, about 200 strains of Staph-
ylococcus, rifampicin resistance in M. tuberculosis, oxacillin
resistance in S. aureus, typing of Neisseria meningitides,
simultaneous detection of a large spectrum of bacterial and
viral pathogens, and a GeneChip with more than 60,000
probes able to identify about 9,000 species of bacteria using
polymorphisms in their 16S ribosomal RNA.
While these data may have outstanding importance for
clinical science, the relative complexity of this approach limits
its usage to advanced laboratories and makes it expensive. For
routine clinical diagnostics of infectious diseases, it is often
more convenient to have limited and reliable information
generated by the use of low-density arrays, in which individual
elements detect specific sequence variations.
Thus, the most-popular microarray technology in clinical
diagnostics today is low-density printing of pre-synthesized
oligonucleotides on chemically treated slides; they often
consist of 96 features to match standard plates. Such slides
are easily prepared, inexpensive and require only the most
basic equipment for printing, hybridization and scanning.
Contrary to the Affymetrix chips, low-density home-printed
arrays can be adapted to new probes or new overall design
without contacting the manufacturer. Such arrays can be
converted into affordable point-of-care testing (POCT) devi-
ces and even field systems.
A versatile and robust method based on the immobilization
of DNA probes in three-dimensional gel drops has been
developed in the laboratory of Andrei Mirzabekov at the
Institute of Molecular Biology in Moscow.(19) The three-
dimensional structure of individual elements and liquid
environment provide significant advantages for clinical diag-
nostics. Namely, it allows using relatively short oligonucleo-
tides for efficient identification of point mutations, not to
mention more extensive sequence variations. Reliable dis-
crimination between matching and mismatching samples,
which can be achieved using gel-based arrays, allows the
number of elements necessary to detect specific sequences to
be minimized. Decreasing the initial concentration of the
probes accelerates the process of discrimination between
perfect and imperfect duplexes upon hybridization, and
optimization of washing (dissociation) can further improve
the discrimination; this occurs in accordance with theoretical
predictions.(20) Direct comparison of hybridization on flat
microarrays and three-dimensional microchips shows that,
although hybridization on flat microarrays is slightly faster,
gel-based microchips discriminate between perfect and
imperfect duplexes more efficiently and generate 5- to 20-fold
stronger signals.(21)
Because of the high water content, high porosity and
complete isolation of gel pads from each other, they can
function as reaction tubes of nanoliter volume. In particular,
this concept has been used in the development of the so-called
‘‘on-chip PCR’’ procedure, which combines PCR amplification
both inside the pads and in overlay solution with hybridization
with immobilized probes (Fig. 1).(22) Pilot experiments were
designed to identify bacteria within the Bacillus cereus group,
which includes B. anthracis. The first unambiguous identifica-
tion of B. anthracis by hybridization of bacillus RNA on
microarrays was reported in 1999,(23) and then using on-chip
PCR in 2001.(24) The same approach was also used for the
detection of mutations in M. tuberculosis conferring drug
resistance.(25) The most-recent application of this technology
is real-time on-chip PCR for identification and quantification of
HIV, HBC and HCV in donor blood.(26)
Another promising technology employs electrochemical
detection of the hybridization events on multiplex microarray
using enzyme-linked assaysor immobilization of the probes on
the surface of silicon wafers.(27) For instance, electrochemical
detection of hybridization on microarrays was successfully
employed for the detection of toxin-encoding genes respon-
sible for pathogenicity in the Bacillus cereus group.(28) The
latter technology does not even require PCR amplification of
target sequences and is therefore very convenient for
conversion into a field-deployed test. In the future, this
microarray format can be further developed into a disposable
cartridge, thus diminishing the possibility of cross-contami-
nation of samples. A similar electrochemical detection of
hybridization eventswithin a disposable cartridge is used in the
eSensor1, a cystic fibrosis carrier detection kit, developed by
Clinical Micro Sensors, Inc. (Pasadena, CA) and approved by
FDA for marketing.
An interesting approach has been proposed by Aragon
et al.(29) During the PCR amplification step, biotinylated
nucleotideswere incorporated in the product. For the detection
of certain resistance mutations, the authors hybridized the
product with immobilized probes, washed the arrays, and
stained them with avidin–peroxidase conjugate and colored
peroxidase substrate. Hybridized sequences were visible as
dark spots without any additional labeling, and the results were
in reasonable concordance with more traditional methods.
Mycobacterium tuberculosis
Traditional evaluation of drug resistance in clinical samples of
M. tuberculosis complex takes at least four weeks, causing
either a delay in treatment, or administration of often
ineffective antimycobacterial drugs. The resistance to two
first-line antibiotics, isoniazid and rifampicin, may be caused
by several different genetic changes, which could be identified
by molecular methods. Besides, NAT allows detecting
resistance to other drugs and to perform strain typing for
epidemiological monitoring (for review, see Ref. 30).
Gryadunov et al. developed a biochip for the detection
of rifampicin-resistant and isoniazid-resistant strains of
What’s new?
BioEssays 30.7 675

M. tuberculosis.(31) The biochip identifies over 95% of
rifampicin-resistant and more than 80% of isoniazid-resistant
M. tuberculosis strains in sputum samples.
The biochip consists of 77 gel elements and detects the
27 most-common mutations in the rpoB gene responsible
for rifampicin resistance, 11 mutations in the katG gene, five
mutations in the promoter region of the inhA gene, and five
mutations in the intergenic regulatory region of the ahpC–
oxyR genes, all of which can cause resistance to isoniazid.
Fig. 2 shows these microarrays after hybridization and
illustrates the interpretation of the results.
The performance of gel-based biochips was successfully
tested by an independent laboratory in the USA.(32) The
system displayed a sensitivity of 80% and a specificity of 100%
relative to conventional testing for rifampicin resistance. All
discrepancies between the microarray test and culture test
were caused by either rare mutations not detectable by the
microarray probes, or by unknown mutations. Gel-based
biochips were also used for large-scale epidemiological
studies of drug resistance in Kyrgyz Republic.(33,34)
The newest generation of TB-biochips identifies mutations
responsible for the emerging resistance of M. tuberculosis to a
highly effective second-line fluoroquinolone antibiotics.(35)
The sensitivity and specificity of the method are 93% and
100%, respectively. Due to the discrimination power of gel-
based hybridization, just 20 individual elements sufficed for
the detection of the majority of known mutations conferring this
resistance. Both TB-biochips for the analysis of rifampicin/
isoniazid or fluoroquinolones susceptibility were among the
first in the world to be certified by a national regulatory agency,
Ministry of Public Health of Russian Federation, for clinical
application.
Staphylococcus aureus
There is a growing threat of staphylococcal infections because
of the rapid spread of drug resistance and the ability of some
strains to produce toxins. Similarly to M. tuberculosis,
molecular tests, in particular microarray-based, provide the
necessary diagnostic tool for fast and reliable characterization
of this pathogen.
Monecke and Ehricht designed a diagnostic microarray that
identifies the genus and species of the pathogen and detects
more than 20 genes of drug resistance and 10 genes encoding
various toxins simultaneously.(36) An efficient diagnostic
microarray has been developed and tested by Zhu et al.(37)
Their array differentiates S. aureus from other staphylococci
using mutations in the variable region of 16S rRNA and detects
genes encoding resistance to seven antibiotics. The method
was successfully tested on about 400 samples. Another, even
Figure 1. A: On-chip PCR and B: its allele-specific variant. The initial PCR stage (not shown) occurs in reaction solution outside the gel
pad. Reaction mixture contains forward and labeled reverse primers. During the next stage, the immobilized forward primers are involved
in amplification along with free reverse primers from surrounding solution (A1). The immobilized primers are extended enzymatically
(A2), forming new target chains attached to the gel via their 50 termini. These chains serve as targets for further amplification (A3) and
eventually form tethered double-stranded hybridization complexes that either contain a fluorescent label (A4), or acquire the ability to bind
non-specific fluorescent dyes (e.g. SYBR1 green used for on-chip real-time PCR). Since each gel pad contains primers specific to one
target only, the kinetics of increase of fluorescence in each gel pad reflects the accumulation of the corresponding amplification product.
Fluorescence signals in gel pads are measured at the end of each elongation step, when specific double-stranded hybridization complexes
remain stable due to their high melting temperature, in contrast to shorter non-specific products. An oligonucleotide specific to the middle
part of the target product could serve as an additional specificity control (A5). Primer extension occurs only when the terminal nucleotide is
complementary to the tested sequence (B6); otherwise, the allele-specific PCR is not possible (B7).
What’s new?
676 BioEssays 30.7

more general, approach to the detection of drug resistance is
based on a microarray that can identify almost 100 different
genes responsible for drug resistance found in many different
bacteria.(38)
In many clinical situations with limited access to even most-
basic clinical laboratories, there is a need for rapid identi-
fication of pathogens from a broad variety of microorganisms.
Zhu et al. developed a microarray for identification of five
species of bacteria most common in clinical settings, including
Staphylococcus.(39) The method does not require PCR,
making it especially suitable for field applications, and its
specificity is better than 98%. The authors employed the
so-called base stacking hybridization discovered and explored
earlier by Vasiliskov et al.(40)
Malaria
Malaria remains one of the most frequent causes of death
among infectious diseases. According to some estimates, up
to 500 million people in the world are infected, leading to high
morbidity and mortality. The environmental ban on DDT
resulted in dramatic increase in the prevalence of malaria,
while the pathogen acquired resistance to chloroquine,
quinine and other common treatments.
Molecular studies of Plasmodium falciparum began rela-
tively late. NAT of P. falciparum is hindered by the unusually
high AT content of its DNA. Direct testing of the pathogen for
drug resistance is difficult enough. Recent advances in
sequencing and molecular mapping of Plasmodium genome
yielded many SNPs linked to resistance genes or directly
responsible for the resistance. Microarray-based analysis of
the resistance genes already reached better than 94%
specificity, can be performed even on asymptomatic patients
and is highly cost effective.(41) In a recent review, molecular
approaches, in particular microarray-based methods, are
considered the most-promising direction in the diagnostics
and characterization of the malaria pathogen.(42)
Figure 2. Analysis of M. tuberculosis susceptibility to rifampin (RIF) and isoniazid (INH) by hybridization on gel-based biochip. A: The
diagnostic microarray (biochip). The microarray detects 27 most-common mutations in the rpoB gene responsible for RIF
resistance (the corresponding area is marked yellow) and 21 mutations in the katG, inhA, ahpC genes, which cause the resistance to
INH (blue). The biochip also contains probes complementary to the IS6110 element (green) present in most mycobacteria belonging to M.
tuberculosis complex. Gel elements marked ‘R’ serve as negative controls, whereas marker pads ‘M’ are necessary for automatic
processing of fluorescence images. The pads with immobilized oligonucleotides are clustered in 21 groups, which are depicted as boxes.
Each group corresponds to a single variable amino acid position in rpoB and katG genes or variable nucleotide in the promoter regions of
inhA and ahpC genes. One gel pad within each group contains oligonucleotides matching the wild-type sequence (grey circles). Other gel
pads contain immobilized oligonucleotides forming perfect duplexes with different mutant variants of the same codon (white circles). Thus,
only one oligonucleotide in each group could form a perfect duplex with the target DNA. After hybridization, the fluorescence signals are
analyzed within each group separately to decide whether mutation(s) are present in the amino acid position concerned.B:Hybridization of a
wild-type DNA sample on the biochip. The maximum fluorescence intensity within each group is observed in gel pads containing
oligonucleotides corresponding to the wild-type sequence. Positive signals in gel pads with IS6110 probes confirm that the tested strain
belongs to the M. tuberculosis complex. This particular pattern indicates that standard therapy with RIF and INH could be applied.
C: Fluorescence pattern resulting from hybridization of DNA sample containing mutations Ser 531>Leu, TCG>TTG in the rpoB gene and
Ser 315>Thr and AGC>ACC in the katG gene differs from the WT pattern (signal changes are marked by the dashed frames). This TB
strain is considered multidrug-resistant, and therapy with the second-line drugs must be used.
What’s new?
BioEssays 30.7 677

Viruses
The major challenge in the molecular diagnostics of many
viruses,especially influenza,HIVandhepatitisC, is their extreme
variability and mutability. Sequences of viruses of the same
species or even serotype may differ by more than 30%, and
selection of primer sequences for PCR amplification and target
segments for the subsequent hybridization requires extensive
analysis of available databases and phylogenetic trees.
HBV
Impressive results were achieved by Chen et al., who
developed a microarray for detection of resistance of hepatitis
B virus (HBV) to lamivudine, one of the major anti-retroviral
drugs.(43) Expected mutations in HBV gene encoding
DNA polymerase were detected using microarrays in all
388 samples found mutant by direct sequencing. The
sensitivity of the microarray to the presence of HBV was
tested on almost 1000 samples and reached 99.7%. Moreover,
the microarray was able to detect mutant variants against the
background of wild-type virus in patients with mixed infections.
HIV
HIV-infected patients are treated with a mixture of drugs, the
so-called highly active antiretroviral therapy (HAART), which
decreases both morbidity and mortality. Under the pressure of
the drugs, the virus mutates and develops resistance. The
situation is further complicated by pre-existing sequence
polymorphisms and by the complex nature of resistance often
resulting from a combination of mutations.
The complexity of genetic testing of drug resistance in HIV
was reviewed in 2001,(44) and the situation has not changed
much since then. In general, the level of resistance to specific
drugs cannot be easily deduced from the sequence of the
virus. The HIV GeneChip for sequencing of HIV isolates by
hybridization offered in the past by Affymetrix currently is not
even mentioned on its web site. HIV drug resistance is
determined for practical purposes mostly from clinical obser-
vations.
However, certain HIV polymorphisms, including those
associated with drug resistance, can be identified using
dedicated microarrays and employed in epidemiological
studies. For instance, Roudinskii et al.(45) used gel-based
biochips to examine the occurrence of the so-called secondary
protease mutation V77I. The detailed study followed the route
of transmission of this particular strain in Eastern Europe.
Precision screening of more than 100 samples was possible
because of the ability of gel-immobilized oligonucleotides to
discriminate single nucleotide mismatches.
Influenza
Influenza causes high morbidity and mortality throughout the
world. Its diagnosis based on clinical symptoms is unreliable,
but timely administration of antiviral drugs is considered highly
cost effective and curative. Massive implementation of
affordable molecular diagnostics of influenza will serve many
purposes: improving discrimination of influenza from other
respiratory diseases, facilitating prophylaxis and treatment,
monitoring the spread of specific types and subtypes of the
virus, and helping to prevent pandemic of new highly
pathogenic and contagious strains.
Influenza viruses are divided into groups A, B and C, and
each of them subdivided into several subtypes based on
serological characteristics of two proteins, hemagglutinin (H)
and neuraminidase (N). The C virus is mostly asymptomatic;
B virus usually causes minor illness, but can be dangerous in
the elderly; Aviruses are the most dangerous, and some of the
A subtypes can cause pandemics. High mutability of the
influenza viruses is the major challenge in the design of
diagnostic microarrays.
Microarrays for identification of influenza A recognizing all
16 forms of hemagglutinins and 9 neuraminidases were
designed and successfully tested.(46) These microarrays
(commercial name GreenChips) consist of a relatively high-
density set of 15,000 60-mers and require rather advanced
technology for the manufacture of the chips, preparation of
probes and analysis of the results, so the authors do not
consider their approach ‘‘a stand-alone diagnostic platform’’.
Dawson et al. developed another technology for identifica-
tion of both modern(47) and historical(48) samples of the Avirus
(commercial name MChips). MChips employs low-density
arrays and identify polymorphisms in the so-called M gene of
influenza virus, which strongly correlate with the subtypes of
the agglutinin and neuraminidase genes. MChip targets
primarily clinical applications. Its strength stems from novel
software used for the analysis of sequence databases,
selection of highly informative sets of diagnostic oligonucleo-
tides and sophisticated analysis of the relative intensities of
fluorescence after hybridization of the probes.(49) The MChip
was successfully used for identification of influenza B in both
historical and modern clinical samples(50) and for identification
of subtypes of B virus and subtypes of A virus currently
circulating in humans (A/H3N2 and A/H1N1) and birds (A/
H5N1).(51) When referenced to traditional cultural typing of the
viruses and to typing using RT-PCR, the performance of
MChip equaled or exceeded that of commercial immunoassay
QuickVue (Quidel Corp., San Diego, CA).
Fesenko et al.(52) developed a gel-based biochip, which
detects 15 subtypes of HA and two most significant subtypes of
NA in influenza virus. The microchip was validated using a panel
of 21 reference strains and successfully tested on 41 samples
isolated during the poultry epizootics in Novosibirsk (Russia,
2005). The sensitivity and specificity of the method were 76%
and 100%, respectively, compared with standard techniques of
virus isolation followed by immunoassay. The whole procedure
can be performed in 10 hours—significantly less than conven-
tional cultivation-based identification.
What’s new?
678 BioEssays 30.7
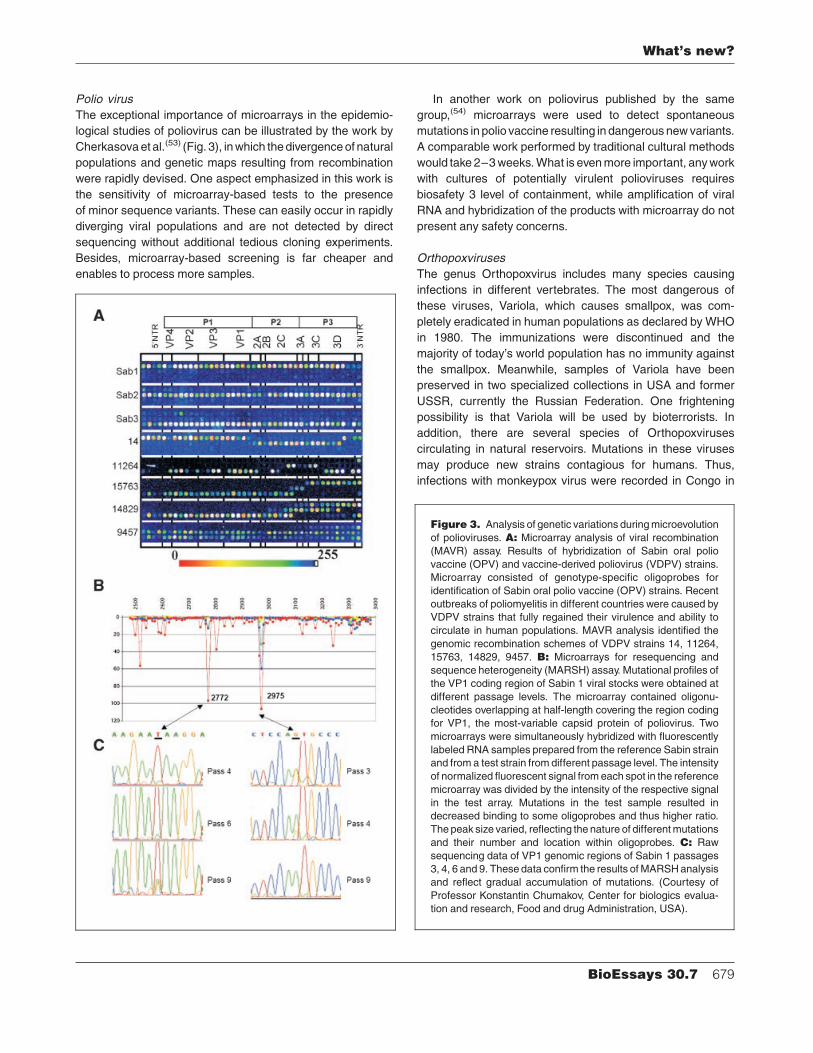
Polio virus
The exceptional importance of microarrays in the epidemio-
logical studies of poliovirus can be illustrated by the work by
Cherkasova et al.(53) (Fig. 3), in which the divergence of natural
populations and genetic maps resulting from recombination
were rapidly devised. One aspect emphasized in this work is
the sensitivity of microarray-based tests to the presence
of minor sequence variants. These can easily occur in rapidly
diverging viral populations and are not detected by direct
sequencing without additional tedious cloning experiments.
Besides, microarray-based screening is far cheaper and
enables to process more samples.
In another work on poliovirus published by the same
group,(54) microarrays were used to detect spontaneous
mutations in polio vaccine resulting in dangerous new variants.
A comparable work performed by traditional cultural methods
would take 2–3 weeks. What is even more important, any work
with cultures of potentially virulent polioviruses requires
biosafety 3 level of containment, while amplification of viral
RNA and hybridization of the products with microarray do not
present any safety concerns.
Orthopoxviruses
The genus Orthopoxvirus includes many species causing
infections in different vertebrates. The most dangerous of
these viruses, Variola, which causes smallpox, was com-
pletely eradicated in human populations as declared by WHO
in 1980. The immunizations were discontinued and the
majority of today’s world population has no immunity against
the smallpox. Meanwhile, samples of Variola have been
preserved in two specialized collections in USA and former
USSR, currently the Russian Federation. One frightening
possibility is that Variola will be used by bioterrorists. In
addition, there are several species of Orthopoxviruses
circulating in natural reservoirs. Mutations in these viruses
may produce new strains contagious for humans. Thus,
infections with monkeypox virus were recorded in Congo in
Figure 3. Analysis of genetic variations during microevolution
of polioviruses. A: Microarray analysis of viral recombination
(MAVR) assay. Results of hybridization of Sabin oral polio
vaccine (OPV) and vaccine-derived poliovirus (VDPV) strains.
Microarray consisted of genotype-specific oligoprobes for
identification of Sabin oral polio vaccine (OPV) strains. Recent
outbreaks of poliomyelitis in different countries were caused by
VDPV strains that fully regained their virulence and ability to
circulate in human populations. MAVR analysis identified the
genomic recombination schemes of VDPV strains 14, 11264,
15763, 14829, 9457. B: Microarrays for resequencing and
sequence heterogeneity (MARSH) assay. Mutational profiles of
the VP1 coding region of Sabin 1 viral stocks were obtained at
different passage levels. The microarray contained oligonu-
cleotides overlapping at half-length covering the region coding
for VP1, the most-variable capsid protein of poliovirus. Two
microarrays were simultaneously hybridized with fluorescently
labeled RNA samples prepared from the reference Sabin strain
and from a test strain from different passage level. The intensity
of normalized fluorescent signal from each spot in the reference
microarray was divided by the intensity of the respective signal
in the test array. Mutations in the test sample resulted in
decreased binding to some oligoprobes and thus higher ratio.
The peak size varied, reflecting the nature of different mutations
and their number and location within oligoprobes. C: Raw
sequencing data of VP1 genomic regions of Sabin 1 passages
3, 4, 6 and 9. These data confirm the results of MARSH analysis
and reflect gradual accumulation of mutations. (Courtesy of
Professor Konstantin Chumakov, Center for biologics evalua-
tion and research, Food and drug Administration, USA).
What’s new?
BioEssays 30.7 679

1996–2001 and later in the United States, and cowpox
infections were reported in humans in Europe and Brazil (for
references, see Refs 55,56).
Traditional methods of detection and characterization of
Orthopoxviruses are slow and laborous, and cannot be
performed in the field. The advantages of microarray-based
technologies for this application are especially striking (for
more detailed discussion see Ref. 57). Besides Orthopoxvi-
ruses, the microarrays carry specific probes for several strains
of herpes viruses, which during the early phase of infection can
cause a clinical pattern similar to Orthopoxviruses. Successful
detection and identification of these viruses has been achiev-
ed most recently using both traditional two-dimensional
arrays(56,58) and gel-based three-dimensional biochips.(57)
Identification of pathogens in complex mixtures
The need to identify pathogens in complex environmental or
clinical samples arises in the course of counter-terrorism
measures and many other situations. Contrary to traditional
immunological methods, microarrays can carry a large
number of probes directed against many diverse viruses and
bacteria. Identification of bacteria is usually achieved by
hybridization of segments of ribosomal genes, which are
relatively easy to amplify with universal primers, with variable
internal probes. In some cases, when the variability of
the ribosomal genes is insufficient, the sequence of the gyr
gene is used as an additional source of polymorphisms. Other
probes may be placed on the array to detect genes encoding
toxins and antibiotic resistance. The bottleneck in the design of
microarray-based tests for broad spectrum of pathogens is
often the choice of sequences to be amplified and correspond-
ing primers for multiplex PCR.
One application of broad NAT designed to detect viruses is
monitoring of safety of donor blood supplies. Since antibodies
against viruses may not appear in circulation immediately after
the infection, detection of viral nucleic acids is a more-reliable
alternative. Microarrays were developed to detect the most-
important viruses—HIV and hepatitis B and C—in donor
blood, and the sensitivity of the pilot tests was sufficient for
further development of the method.(58) In particular, gel-based
biochips developed by Khodakov et al.(26) allow blood-borne
pathogens to be quantified.
A microarray for the detection of food-born pathogens was
designed and successfully tested by Wang et al.(59) The
authors amplified the variable region of the 16S RNA gene and
hybridized it to oligonucleotides corresponding to unique
sequences in 115 different bacteria; during pilot experiments,
113 of these were identified correctly. The sensitivity of the
array was 102 CFU. The same group developed a microarray
for identification of fungal pathogens carrying specific probes
for 122 species.(60)
One of the most challenging clinical tasks is the correct
diagnosis of respiratory infections. A microarray-based test with
electrochemical enzyme-linked registration was developed by
Lodes et al.(61) The array detects four major bacterial pathogens
and nine viruses. In a similar work, Lin et al. screened more than
400 nasal wash specimens from patients with febrile respiratory
disease.(62) The microarray detection showed better than 98%
agreement with other detection methods with the sensitivity
101–103 genomic copies. The scale of this study enabled
researchers to demonstrate the presence of multiple pathogens
in 13% of samples. Interestingly, the high sensitivity of micro-
array-based detection of pathogens causing generic respiratory
infections calls for a complete consideration of clinical pattern,
since such pathogens are known to reside in healthy individuals,
especially children(63) and elderly.(64)
Conclusion
So far, clinical applications of oligonucleotide microarrays for
detection, identification, characterization and monitoring of
pathogens proved to be more successful than microarrays
intended for diagnosis, prognosis and management of non-
infectious diseases. As far as the growth of commercial
applications of NAT is concerned, it is anticipated to increase
from 8% of all molecular diagnostics in 2005 to 14% in 2010
(the total number include all immunological testing).(65) It is
quite likely that a significant share of this growth will take place
in mass applications of the diagnostics of infectious diseases
displacing or verifying some immunological tests.
Judging by publications, microarray-based diagnostics is
being developed in many countries, but the status of its actual
clinical implementation is mostly unknown. Because of the
dominating size of American diagnostic market and trans-
parency of its regulatory environment, the approval of micro-
arrays for clinical use in the USA is usually immediately made
public. The situation in EU countries is less clear, because
there is no explicit requirement of clinical relevance for new
tests, and information about the rest of the world is almost
impossible to find.
Nevertheless, we believe that microarray technologies for
the detection of pathogens hold great promise. They are
affordable, require small capital investment and are easily
converted into POCT and field applications. Detection and
identification of pathogens usually employ low-density arrays,
which are designed for straightforward answers without the
need for sophisticated equipment and data processing. Such
arrays better satisfy regulatory requirements. Last, but not
least, a significant proportion of humankind is in dire need of
effective diagnostics and treatment of infectious diseases,
and, even with the highest standards of health care, we will
always face the threat of emerging infections.
References1. http://www.fda.gov/cdrh/osel/programareas/genomic.html
2. Jordan BR. 2007. DNA microarrays in the clinic: how soon, how
extensively? BioEssays 29:699–705.
What’s new?
680 BioEssays 30.7

3. Lane DP. Exploiting the p53 Pathway for the Diagnosis and Therapy of
Human Cancer. Cold Spring Harbor Sym Quant Biol LX:489–497.
4. http://www.who.int/healthinfo/statistics/bodgbddeathdalyestimates.xls
5. Michaud CM. 2000. Global burden of infectious disease. In: Lederberg J,
editor. Encyclopedia of Microbiology, v. 2. San Diego: Academic Press.
529–540.
6. http://www.who.int/bulletin/volumes/83/3/171.pdf
7. Meltzer MI. Economic consequences of infectious diseases. In: Leder-
berg J, editor. Encyclopedia of Microbiology, v. 2. San Diego: Academic
Press. 137–155.
8. Bosch FX, de Sanjose S. 2007. The epidemiology of human papilloma-
virus infection and cervical cancer. Dis Markers. 23:213–227.
9. Kremsdorf D, Soussan P, Paterlini-Brechot P, Brechot C. 2006. Hepatitis
B virus-related hepatocellular carcinoma: paradigms for viral-related
human carcinogenesis. Oncogene 25:3823–3833.
10. Levrero M. 2006. Viral hepatitis and liver cancer: the case of hepatitis C.
Oncogene 25:3834–3847.
11. Malfertheiner P, Sipponen P, Naumann M, Moayyedi P, Megraud F, et al.
2005. Helicobacter pylori eradication has the potential to prevent gastric
cancer: a state-of-the-art critique. Am J Gastroenterol 100:2100–
2115.
12. Moss SF, Blaser MJ. 2005. Mechanisms of disease: Inflammation and the
origins of cancer. Nat Clin Pract Oncol 2:90–97.
13. http://www.niaid.nih.gov/dmid/genomes/brc/
14. http://www3.niaid.nih.gov/research/topics/emerging/list.htm
15. Wilkes T, Laux H, Foy CA. 2007. Microarray data quality review of current
developments. OMICS A J Integrative Biol 11:1–13.
16. http://www.fda.gov/nctr/science/centers/toxicoinformatics/maqc/
17. Lee HK, Myoungho-Lee, Roh HW, Lee N, Cho YH, et al. 2005. DNA
chip evaluation as a diagnostic device. Curr App Physics 5:433–437.
18. http://www.affymetrix.com/products/fos/infectious_disease_identification.
affx
19. Rubina AYu, Pan_kov SV, Dementieva EI, Pen’kov DN, Butygin AV, et al.
2004. Hydrogel drop microchips with immobilized DNA: properties and
methods for large-scale production. Anal Biochem 325:92–106.
20. Sorokin NV, Chechetkin VR, Livshits MA, Pan’kov SV, Donnikov MY, et al.
2005. Discrimination between perfect and mismatched duplexes with
oligonucleotide gel microchips: Role of thermodynamic and kinetic
effects during hybridization. J. Biomol Struct Dyn 22:725–734.
21. Sorokin NV, Chechetkin VR, Pan’kov SV, Somova OG, Livshits MA, et al.
2006. Kinetics of hybridization on surface oligonucleotide microchips:
Theory, experiment, and comparison with hybridization on gel-based
microchips. J. Biomol Struct Dyn 24:57–66.
22. Strizhkov BN, Drobyshev AL, Mikhailovich VM, Mirzabekov AD. 2000.
PCR amplification on a microarray of gel-immobilized oligonucleotides:
detection of bacterial toxin- and drug-resistant genes and their
mutations. Biotechniques 29:844–854.
23. Bavykin SG, Mikhailovich V, Zakharyev V, Lysov YP, Chernyi A, et al.
1999. Biological microchip technology for discrimination of Bacillus
anthracis and closely related species on the basis of genetic criteria. In:
Second international workshop on the molecular biology of Bacillus
cereus, Bacillus anthracis and Bacillus thuringensis. Taos, New Mexico,
August 11–13.
24. Gryadunov DA, Mikhailovich VM, Noskov AN, Lapa SA, Sobolev AYu,
et al. 2001. Detection of Bacillus anthracis using multiplex PCR on the
oligonucleotide biochip. Dokl Biochem Bioph 381:384–386.
25. Mikhailovich V, Lapa S, Gryadunov D, Sobolev A, Strizhkov B, et al.
2001. Identification of rifampin-resistant Mycobacterium tuberculosis
strains by hybridization, PCR, and ligase detection reaction on
oligonucleotide microchips. J. Clin Microbiol 39:2531–2540.
26. Khodakov DA, Zakharova NV, Gryadunov DA, Filatov FP, Zasedatelev
AS, Mikhailovich VM. 2008. An oligonucleotide microarray for multiplex
real-time PCR identification of HIV-1, HBV, and HCV et al. Biotechinques
44:241–248.
27. Cai W, Peck JR, van der Weide DW, Hamers RJ. 2004. Direct electrical
detection of hybridization at DNA-modified silicon surfaces. Biosens
Bioelectron 19:1013–1019.
28. Liu Y, Elsholz B, Enfors SO, Gabig-Ciminska M. 2007. Confirmative
electric DNA array-based test for food poisoning Bacillus cereus. J.
Microbiol Methods 70:55–64.
29. Aragon LM, Navarro F, Heiser V, Garrigo M, Espanol M, et al. 2006.
Rapid detection of specific gene mutations associated with isoniazid or
rifampicin resistance in Mycobacterium tuberculosis clinical isolates
using non-fluorescent low-density DNA microarrays. J. Antimicrob
Chemother 57:825–831.
30. Cheng VCC, Yew WW, Yuen KY. 2005. Molecular diagnostics in
tuberculosis. Eur J Clin Microbiol Infect Dis 24:711–720.
31. Gryadunov D, Mikhailovich V, Lapa S, Roudinskii N, Donnikov M, et al.
2005. Evaluation of hybridisation on oligonucleotide microarrays for
analysis of drug-resistant Mycobacterium tuberculosis. Clin Microbiol
Infect 11:531–539.
32. Caoili JC, Mayorova A, Sikes D, Hickman L, Plikaytis BB, et al. 2006.
Evaluation of the TB-Biochip oligonucleotide microarray system for rapid
detection of rifampin resistance in Mycobacterium tuberculosis. J. Clin
Microbiol 44:2378–2381.
33. Isakova ZhT, Pak OA, Iusupova EIu, Myrzaliev BB, Chubakov TCh, et al.
2005. [Use of biological microchips in the determination of drug-
resistance of M. tuberculosis to rifampicin. Probl Tuberk Bolezn Legk
8:50–53.
34. Isakova ZhT, Goncharova ZK, Iusupova EU, Tumashova AF, Kozhomku-
lov MD, et al. 2007. [Analysis of mutations of multidrug-resistant M.
tuberculosis strains in patients with tuberculosis in the Kyrghyz Republic.
Probl Tuberk Bolezn Legk 417–421.
35. Antonova O, Gryadunov D, Lapa S, Kuz’min A, Nosova E. 2008.
Identification of mutations responsible for fluoroquinolone resistance of
Mycobacterium tuberculosis using hybridization on the biological
microchips. Bull Exp Biol Med 145:115–120.
36. Monecke S, Ehricht R. 2005. Rapid genotyping of methicillin-resistant
Staphylococcus aureus (MRSA) isolates using miniaturised oligonucleo-
tide arrays. Clin Microbiol Infect 11:825–833.
37. Zhu LX, Zhang ZW, Wang C, Yang HW, Jiang D, et al. 2007.
Simultaneous Detection of Antibiotic Resistance Genes among Staph-
ylococcal Clinical Isolates with a DNA Microarray. J. Clin Microbiol Aug
29; [Epub ahead of print]
38. Frye JG, Jesse T, Long F, Rondeau G, Porwollik S, et al. 2006. DNA
microarray detection of antimicrobial resistance genes in diverse
bacteria. Int J Antimicrob Agents 27:138–151.
39. Zhu LX, Wang D, Zhang GB, Jiang D, Zhang ZW, et al. 2007.
Development of a base stacking hybridization-based microarray method
for rapid identification of clinical isolates. Diagn Microbiol Infect Dis
59:149–156.
40. Vasiliskov VA, Prokopenko DV, Mirzabekov AD. 2001. Parallel multiplex
thermodynamic analysis of coaxial base stacking in DNA duplexes by
oligodeoxyribonucleotide microchips. Nucleic Acids Res 29:2303–2313.
41. Crameri A, Marfurt J, Mugittu K, Maire N, Regos A, et al. 2007. A rapid
microarray-based method for monitoring of all currently known single
nucleotide polymorphisms associated with parasite resistance to
antimalaria drugs. J. Clin Microbiol 45:3685–3691.
42. Ekland EH, Fidock DA. 2007. Advances in understanding the genetic
basis of antimalarial drug resistance. Curr Opin Microbiol 10:363–370.
43. Chen LY, Huang J, Zhang XP, Qiao P, Zhang W. 2005. Clinical evaluation
of oligonucleotide microarrays for the detection of HBV mutants
associated with lamivudine resistance. Pharmacogenomics 6:721–730.
44. Hanna GJ, D’Aquila RT. 2001. Clinical use of genotypic and phenotypic
drug resistance testing to monitor antiretroviral chemotherapy. Clin Infect
Dis 32:774–782.
45. Roudinskii NI, Sukhanova AL, Kazennova EV, Jonathan N, Weber JN,
et al. 2004. Diversity of human immunodeficiency virus type 1 subtype A
and CRF03_AB protease in Eastern Europe: Selection of the V77I variant
and its rapid spread in injecting drug user populations. J. Virol
78:11276–11287.
46. Quan PL, Palacios G, Jabado OJ, Conlan S, Hirschberg DL, et al. 2007.
Detection of respiratory viruses and subtype identification of influenza A
viruses by GreeneChipResp oligonucleotide microarray. J. Clin Microbiol
45:2359–2364.
47. Dawson ED, Moore CL, Smagala JA, Dankbar DM, Mehlmann M, et al.
2006. MChip: a tool for influenza surveillance. Anal Chem 78:7610–7615.
48. Moore CL, Smagala JA, Smith CB, Dawson ED, Cox NJ, et al. 2007.
Evaluation of MChip with historic A/H1N1 influenza viruses including the
1918 ‘‘Spanish Flu’’. J. Clin Microbiol 45:3805–3810.
What’s new?
BioEssays 30.7 681

49. Mehlmann M, Dawson ED, Townsend MB, Smagala JA, Moore CL, et al.
2006. Robust sequence selection method used to develop the FluChip
diagnostic microarray for influenza virus. J. Clin Microbiol 44:2857–
2862.
50. Dankbar DM, Dawson ED, Mehlmann M, Moore CL, Smagala JA, et al.
2007. Diagnostic microarray for influenza B viruses. Anal Chem
79:2084–2090.
51. Mehlmann M, Bonner AB, Williams JV, Dankbar DM, Moore CL, et al.
2007. Comparison of the MChip to viral culture, reverse transcription-
PCR, and the QuickVue influenza AþB test for rapid diagnosis of
influenza. J. Clin Microbiol 45:1234–1237.
52. Fesenko EE, KireyevDE, Gryadunov DA, Mikhailovich VM, Grebennikova
TV, et al. 2007. Oligonucleotide microchip for subtyping of influenza A
Virus. Influenza Other Respir Viruses 1:121–129.
53. Cherkasova E, Laassri M, Chizhikov V, Korotkova E, Dragunsky E, et al.
2003. Microarray analysis of evolution of RNA viruses: evidence of
circulation of virulent highly divergent vaccine-derived polioviruses. Proc
Natl Acad Sci USA 100:9398–9403.
54. Cherkasova EA, Yakovenko ML, Rezapkin GV, Korotkova EA, Ivanova
OE, et al. 2005. Spread of vaccine-derived poliovirus from a paralytic
case in an immunodeficient child: an insight into the natural evolution of
oral polio vaccine. J. Virol 79:1062–1070.
55. Laassri M, Chizhikov V, Mikheev M, Shchelkunov S, Chumakov K.
2003. Detection and discrimination of orthopoxviruses using micro-
arrays of immobilized oligonucleotides. J. Virol Methods 112:67–
78.
56. Myznikova AI, Lapa SA, Griadunov DA, Khodakov DA, Zasedatelev AS,
et al. 2007. [Biological microchip for identification of orthopoxviruses and
agents causing the same clinical picture as in smallpox] [Article in
Russian]. Vopr Virusol 52:41–45.
57. Ryabinin VA, Shundrin LA, Kostina EB, Laassri M, Chizhikov V, et al.
2006. Microarray assay for detection and discrimination of Orthopoxvirus
species. J. Med Virol 78:1325–1340.
58. Hsia CC, Chizhikov VE, Yang AX, Selvapandiyan A, Hewlett I, et al. 2007.
Microarray multiplex assay for the simultaneous detection and discrim-
ination of hepatitis B, hepatitis C, and human immunodeficiency type-1
viruses in human blood samples. Biochem Biophys Res Commun
356:1017–1023.
59. Wang XW, Zhang L, Jin LQ, Jin M, Shen ZQ, et al. 2007. Development
and application of an oligonucleotide microarray for the detection of
food-borne bacterial pathogens. Appl Microbiol Biotechnol 76:225–233.
60. Huang A, Li JW, Shen ZQ, Wang XW, Jin M. 2006. High-throughput
identification of clinical pathogenic fungi by hybridization to an
oligonucleotide microarray. J. Clin Microbiol 44:3299–3305.
61. Lodes MJ, Suciu D, Wilmoth JL, Ross M, Munro S, et al. 2007.
Identification of upper respiratory tract pathogens using electrochemical
detection on an oligonucleotide microarray. PLoS ONE 2:e924.
62. Lin B, Malanoski AP, Wang Z, Blaney KM, Ligler AG, et al. 2007.
Application of broad-spectrum, sequence-based pathogen identification
in an urban population. PLoS ONE 2:e419.
63. Lin B, Wang Z, Vora GJ, Thornton JA, Schnur JM, et al. 2006. Broad-
spectrum respiratory tract pathogen identification using resequencing
DNA microarrays. Genome Res 16:527–535.
64. Konno M, Baba S, Mikawa H, Hara K, Matsumoto F, et al. 2006. Study of
upper respiratory tract bacterial flora: first report. Variations in upper
respiratory tract bacterial flora in patients with acute upper respiratory
tract infection and healthy subjects and variations by subject age. J.
Infect Chemother 12:83–96.
65. Doig AP. 2007. Molecular diagnostics market assessment. Genet Eng
News 27:No. 3 (http://www.genengnews.com/articles/chitem.aspx?aid¼ 2006).
What’s new?
682 BioEssays 30.7




















