
Diversity and abundance of wild host plants of lepidopteran stem borers in two agro-ecological zones...
12
Diversity and abundance of wild host plants of lepidopteran stem borers in two agro-ecological zones of Kenya Nicholas A. Otieno 1 , Bruno P. Le Ru 1 , George O. Ong’amo 1 , Pascal Moyal 2 , Stéphane Dupas 2 , Paul-André Calatayud 1 and Jean-François Silvain 2 1 Noctuid Stem borer Biodiversity Project (NSBB). Institut de Recherche pour le Développement / International Centre of Insect Physiology and Ecology (IRD/ICIPE), Nairobi, Kenya 2 Institut de Recherche pour le Développement (IRD), CNRS, Lab. Populations, Génétique et Evolution. Gif-sur-Yvette cedex, France Key words: Stem borer pest, Busseola fusca, Chilo partellus, cereal crops, uncultivated habitat, refuge area SUMMARY A survey was carried out between 2005 and 2006 in two ecologically different localities, Suam and Mtito Andei, to assess diversity and abundance of wild host plants of lepidopteran stem borers during the cropping and non-cropping seasons. Suam in western Kenya is situated in the moist high tropics and is characterised by an Afromontane vegetation mosaic, while Mtito Andei is located in the dry mid-altitudes characterised by a Somalia Masai vegetation mosaic. In Suam, wild host plants and maize covered 11 and 50%, respectively, of the surveyed area. In Mtito Andei, 27% of the surveyed area was under maize during the cropping season, while wild host plant species covered 13% and 8% during the cropping and non-cropping seasons, respectively. There was no significant variation in the relative abundance and diversity of the wild grasses between the two seasons in either location. The abundance of host plants of B. fusca and C. partellus is low in natural habitats surrounding cereal crops. The abundance of C. partellus was low in its wild host plants and B. fusca was absent. Therefore, the role of wild vegetation surrounding cultivated areas in the carry-over of these pests during the non-cropping season is limited. INTRODUCTION Cereal crops, particularly maize (Zea mays L.) and sorghum (Sorghum bicolor L. (Moench)), constitute the most important sources of food in Africa (FAO 2003). In East and Southern Africa, these two crops are grown by both commercial and resource-poor farmers. Major constraints of crop production include several lepidopteran stem borer species whose distribution and pest status varies with region in Africa and ecozones within a region (Schulthess et al. 1997; Kfir et al. 2002). In eastern Africa, Busseola fusca (Fuller) and Chilo partellus (Swinhoe) are the major pests, whereas Chilo orichalcociliellus Walker and Sesamia calamistis Hampson are of minor importance (Seshu Reddy 1983; Guofa et al. 2002). Distributions of major species vary with alti- tude, with B. fusca dominating high-altitude areas International Journal of Biodiversity Science and Management 4 (2008) 1–12; DOI 10.3843/Biodiv.4.2: Correspondence: Bruno P. Le Ru, Institut de Recherche pour le Développement / International Centre of Insect Physiology and Ecology (IRD/ICIPE), Nairobi. (Kenya). PO Box 30772, Nairobi, Kenya. Email: [email protected] 1
-
Upload
independent -
Category
Documents
-
view
8 -
download
0
Transcript of Diversity and abundance of wild host plants of lepidopteran stem borers in two agro-ecological zones...

Diversity and abundance of wild hostplants of lepidopteran stem borers intwo agro-ecological zones of Kenya
Nicholas A. Otieno1, Bruno P. Le Ru1, George O. Ong’amo1, Pascal Moyal2, Stéphane Dupas2,Paul-André Calatayud1 and Jean-François Silvain2
1Noctuid Stem borer Biodiversity Project (NSBB). Institut de Recherche pour leDéveloppement / International Centre of Insect Physiology and Ecology (IRD/ICIPE),Nairobi, Kenya2Institut de Recherche pour le Développement (IRD), CNRS, Lab. Populations, Génétique etEvolution. Gif-sur-Yvette cedex, France
Key words: Stem borer pest, Busseola fusca, Chilo partellus, cereal crops, uncultivated habitat, refuge area
SUMMARYA survey was carried out between 2005 and 2006 in two ecologically different localities,Suam and Mtito Andei, to assess diversity and abundance of wild host plants oflepidopteran stem borers during the cropping and non-cropping seasons. Suam inwestern Kenya is situated in the moist high tropics and is characterised by an Afromontanevegetation mosaic, while Mtito Andei is located in the dry mid-altitudes characterised by aSomalia Masai vegetation mosaic. In Suam, wild host plants and maize covered 11 and50%, respectively, of the surveyed area. In Mtito Andei, 27% of the surveyed area wasunder maize during the cropping season, while wild host plant species covered 13% and8% during the cropping and non-cropping seasons, respectively. There was no significantvariation in the relative abundance and diversity of the wild grasses between the twoseasons in either location. The abundance of host plants of B. fusca and C. partellus is low innatural habitats surrounding cereal crops. The abundance of C. partellus was low in its wildhost plants and B. fusca was absent. Therefore, the role of wild vegetation surroundingcultivated areas in the carry-over of these pests during the non-cropping season is limited.
INTRODUCTION
Cereal crops, particularly maize (Zea mays L.) andsorghum (Sorghum bicolor L. (Moench)), constitutethe most important sources of food in Africa (FAO2003). In East and Southern Africa, these two cropsare grown by both commercial and resource-poorfarmers. Major constraints of crop productioninclude several lepidopteran stem borer specieswhose distribution and pest status varies with region
in Africa and ecozones within a region (Schulthesset al. 1997; Kfir et al. 2002). In eastern Africa,Busseola fusca (Fuller) and Chilo partellus (Swinhoe)are the major pests, whereas Chilo orichalcociliellusWalker and Sesamia calamistis Hampson are ofminor importance (Seshu Reddy 1983; Guofa et al.2002). Distributions of major species vary with alti-tude, with B. fusca dominating high-altitude areas
International Journal of Biodiversity Science and Management 4 (2008) 1–12; DOI 10.3843/Biodiv.4.2:
Correspondence: Bruno P. Le Ru, Institut de Recherche pour le Développement / International Centre of InsectPhysiology and Ecology (IRD/ICIPE), Nairobi. (Kenya). PO Box 30772, Nairobi, Kenya. Email: [email protected]
1

whereas C. partellus is well established in low- andmid-altitude areas (Nye 1960; Seshu Reddy 1983;Guofa et al. 2002; Kfir et al. 2002; Ong’amo et al.2006).
In the tropics, small-scale cereal fields are sur-rounded by uncultivated habitats that are rich inalternative host plants, which are believed to actas reservoirs for stem borer pests during the non-cropping period. These habitats are consideredresponsible for pest outbreaks in crops during thesubsequent growing season (Ingram 1958; Bowden1976; Khan et al. 1997). However, recent studies inUganda and Cameroon concluded that wild grassesplay a minor role in the seasonal carry-over of stemborer pests (Ndemah et al. 2007; Matama-Kaumaet al. 2008). This is supported by results of surveyscarried out in East and Southern Africa since 2002,which suggested that earlier reports of the commonoccurrence of stem borer pests on wild grasses weredue to misidentification of the specimens (Le Ruet al. 2006a).
Lepidopteran stem borers have been associatedwith monocotyledonous host plants belonging tothe families Poaceae, Cyperaceae and Typhaceae(Polaszek and Khan 1998; Guofa et al. 2002; Le Ruet al. 2006a,b) for millions of years (Harris 1962).However, densities of stem borers in wild habitatsrange between 0.001 and 0.03 per tiller, results thatare considerably lower compared to the cultivatedhabitats (Le Ru et al. 2006a, Ndemah et al. 2007;Matama-Kauma et al. 2008). A study carried out inKitale, Kenya, that assessed the potential of usingwild host plants as a refuge for Bt-susceptible pestpopulations, required in resistance managementfor Bt maize (Kanya et al. 2005), showed that theratio of wild host plants to maize was below 10%.However, another study carried out in Kakamegaand Muhaka, representing the moist transitionaland lowland tropical zones, showed that the hostrange of C partellus and B. fusca was limited inboth number and abundance (Otieno et al. 2006),suggesting that Kanya et al. (2005) may have beenoverestimated the populations.
A study was carried out in two locations, Suamand Mtito Andei in Kenya, in an attempt to obtaina wider ecological understanding of the diversityand abundance of wild host plants of the stemborer pests. These locations represent two agro-ecological zones and were chosen for their diversityof habitats and farm management practices. Thestudy was carried out during the cropping and
non-cropping seasons to capture 1) the diversityand abundance of wild host plants of stem borersacross the natural habitat in the different types ofvegetation; and 2) the abundance of stem borerpests in both cultivated and natural habitats.
MATERIALS AND METHODS
Study localities and sampling design
The two study sites, Suam and Mtito Andei, aresituated in two different agro-ecological zones inKenya, dominated by different stem borer pestspecies; Busseola fusca is the dominant borer speciesin Suam while C. partellus dominates in Mtito Andei(Seshu Reddy 1983; Guofa et al. 2002; Ong’amoet al. 2006).
Suam, in western Kenya, is in the moist transi-tional zone and is characterised by an Afromontanemosaic with a lightly wooded grassy formation(Figure 1). It covers an area of 25 km2 and is locatedon the eastern slopes of Mt. Elgon at 1.17°N to1.21°N and 34.77°E to 34.34°E, between 1930 to2150 m above sea level (asl). Approximately 70% ofthe study area is under maize cultivation, 11% iswild habitats, while the remaining 19% is underother vegetation (Goux 2005). Average annualrainfall is 1350 mm and average temperature is20.0°C, with a minimum/maximum temperaturerange of 12.7/26.8°C. The area is characterised bybimodal rainfall that allows only one cropping
Host plants of lepidopteran stem borers Otieno et al.
2 International Journal of Biodiversity Science and Management
Figure 1 Map of Kenya showing the two study loca-tions, Suam and Mtito Andei

season (CS) per annum, which lasts from Aprilto October. The dry season (hereafter referred toas the non-cropping season, NCS) is betweenDecember and March (Corbett and O’Brien 1997).
Mtito Andei is on the western border of theTsavo Park and is composed of thick deciduousshrubby formations with Acacia–Commiphora andother riverine forest species of the Somalia Masaïvegetation mosaic (Figure 1). It covers an area of25 km2 and is located on the border of the TsavoEast National Park, between 2.64°E to 2.69°E, and38.17°N to 38.22°N at 680 to 770 m asl (Goux2005). Annual rainfall is around 660 mm and meantemperature is 25°C, with minimum/maximumtemperatures of 16.8/29.0°C (Corbett and O’Brien1997). The main cropping season falls betweenNovember and January, although maize is grownunder irrigation throughout the year. However, theclimatic conditions are at times irregular, with someyears going without rainfall while others receiverainfall during the known dry period.
In Suam, sampling in the uncultivated habitatswas carried out during the non-cropping season
(NCS) in March and during the cropping season(CS) in July. In Mtito Andei, there was no rain fromNovember to January and sampling was thereforecarried out during the NCS in January 2006, whilethe CS sampling was carried out in April 2006.
Sampling for diversity and abundance ofhost plants in the natural habitat
Sample size
Satellite images were used as basic spatial informa-tion to describe vegetation types of the study sites(Goux 2005). This enabled translation of the vegeta-tion types to the vegetation physiognomic classi-fication by ground truthing. This resulted in adescription of vegetation structural formationclasses that was mainly divided into wild and culti-vated habitats of stem borers (Goux 2005). Samplingwas confined to wild habitats dominated by plantspecies listed by Le Ru et al. (2006a) as host plants ofstem borers. Three vegetation structural formationclasses made up of wild habitats were identified in
Host plants of lepidopteran stem borers Otieno et al.
International Journal of Biodiversity Science and Management 3
Vegetation type Area (km2) %
Suam Forest Woodland (FW)Open Grassland (OG)Savanna Woodland (SW)Agricultural Development Cooperation Maize Farms (ADC)Agricultural Development Cooperation Fallow Farms (ADCF)Local Community Maize Farms (LCM)Wheat Farms (WF)Orange Farms (OF)Other Cultivated Vegetation (OCV)Settlement Area and Infrastructure (SAF)Total Area
1.714.5
101.82.510.080.22.25
25.00
6.8 (9)4 (6)
18 (39)407.2
1040.320.89
100 (54)
Mtito Andei Savanna Grassland (SG)Savanna Woodland (SW)Forest Acacia (FA)Riverine Vegetation (RV)Maize (M)Sorghum (S)Green grams (G)Beans (B)Dam (D)Bare Soil (BS)Total Area
0.829.537.220.226.970.030.510.020.010.16
25.00
3.22 (3)37.39 (28)28.32 (20)0.86 (3)
27.340.1220.080.040.63
100 (54)
Table 1 Surface area (km2) and percentage surface area (relative to total surface area of the study site) of eachvegetation type in Suam and Mtito Andei. In parenthesis is the number of sampling sites in each vegetation type in thewild habitats

Suam and four in Mtito Andei (Table 1). The samplesize n (number of sampling points in areas occupiedby natural vegetation) was determined from theequation of Webster and Oliver (1990). The Geo-graphical Information System (GIS) program, ArcView 3.2 (ESRI 1992) was then used to generate ran-dom sampling points within the vegetation struc-tural formation classes. The number of samplingpoints in the vegetation structures where samplingwas carried out was proportional to the relative areasize of each structure (Table 1). The sample sizeequation was as follows:
n =zI
2
2pq
where n = sample size, I = permitted error (0.1),z = confidence interval (1.64), p = probability ofarea covered by wild host plants, and q = probabilityof area not covered by wild hosts.For Suam: p = 28% and q = 72% where
n =16401
2
2
..
0.28 × 0.72 = 54
For Mtito Andei: p = 69.8% and q = 30.2%, where
n =16401
2
2
..
0.698 × 0.302 = 56.
Hence, 54 points were sampled at both sites.
Experimental procedure
Baseline study techniques for sampling vegetationwere applied (Grieg-Smith 1983). Sampling with-out replacement was used, where the baselineformed one of the outer boundaries of the sampl-ing unit. The transect intercept method ofsampling vegetation as described by Grieg-Smith(1983) was employed. At the beginning of eachsampling point, a line (the baseline) stretching200 m and oriented north– south was established.The baseline (200-m long) was divided into tenequal parts (marks), starting from first mark (0 m)at the beginning to the tenth mark (200 m) at theend of the 200 m transect. Three sets of numbersfrom the ten designated marks were randomlygenerated and then allocated to each baseline. Thesets of numbers formed the starting points whereline transects were established from the baseline.The line transects (transects 1, 2 and 3, each mea-suring 50 m) were established starting at themarked points, running perpendicular to the base-line and parallel to one another in a west–eastdirection (Figure 2). The area of a sampling unitwas 200 m × 50 m. Sampling was done along theline transects and the exercise was repeated at allthe sampling points.
All species in the Cyperaceae, Poaceae andTyphaceae that intercepted the transects wererecorded (Phillips 1995; Van Oudtshoorn 2004).However, only plant species listed by Le Ru et al.(2006a) as host plants of stem borers are presented(Tables 2 and 5). Voucher specimen were collec-ted and taken to the East African Herbarium inNairobi, Kenya, when species identification was notpossible in the field. Crown cover (the proportionof the ground occupied by a perpendicular projec-tion of the aerial parts of the individual plantspecies) of each plant intercepting the transectwas recorded (Greig-Smith 1983). The length oftransect intercepted by individual plant species (incentimetres) was recorded. The intercept lengthsfor each plant species in the whole vegetation struc-tural formation classes were summed. This was thendivided by the total length of the transects and con-verted to a percentage to give percentage plantcover of each species in the vegetation structuralformation classes, providing the average coverabundance (%). Relative abundance was achievedby dividing the percentage cover abundance ofeach plant species by the total percentage cover
Host plants of lepidopteran stem borers Otieno et al.
4 International Journal of Biodiversity Science and Management
North
South
Baseline (200 m)
50 m1st transect
2nd transect
3rd transect
Figure 2 Establishment of baseline and transect lineswhere sampling took place

abundance of all the plant species in the respectivevegetation structural formation class. This was thenused to calculate the diversity index.
Sampling for abundance of stem borerpests in both cultivated and naturalhabitats
Cultivated habitat
The total number of maize plants required for eachsite depends on the area under maize relative to thetotal area under study. It was determined usingKarandinos’ (1976) equation:
n =Z
q pa / /22
δ⎛⎝⎜
⎞⎠⎟
{Eq 1}
where n is the number of maize plants required ineach site, δ is the reliability level expressed as a fixedproportion of the mean (0.05), Za/2 is the standardnormal deviate at 95% (1.96), p is the proportion ofland under maize cultivation, and q = 1 – p
The total number of maize plants sampled was942 and 1536 in Suam and Mtito Andei respectivelyThen, the number of maize plants inspected forstem borer infestation in each field was determinedusing the equation described by Zar (1999)
n =Z
Dd
22
24α( ) {Eq 2}
where Za(2) is the standard normal deviate (1.96), dis the permitted error (0.1) resulting in a uniformnumber plants in all farms, and D is the designeffect (1).
n =196
4 1 01
2
2
..× ×
= 96.04 ≈ 100 maize plants per field.
The number of maize fields in the respective locali-ties was determined from the quotient of the totalnumber of plants per site (Eq.1) divided by 100(Eq 2); 10 and 16 maize fields were sampled inSuam and Mtito Andei, respectively In each field,100 randomly selected maize plants were inspectedfor borer infestation symptoms and only infestedstems were dissected for larval recovery.
Natural habitat
Since borer densities on wild hosts plants areconsiderably lower than on cultivated cereals (Nye1960; Schulthess et al. 1997; Ndemah et al. 2007;
Matama-Kauma et al. 2008), selective sampling wasadopted to increase the chances of finding borers.In both localities, all potential hosts belonging tothe Poacaea, Cyperaceae and Typhaceae foundwithin 200–300m from the border of the sampledmaize fields were carefully examined for symptomsof stem borer damage. Damaged plants were cutat the base and dissected for recovery of larvaeand pupae. Plants from which the larvae or pupaewere collected were identified. In cases whereidentities of the infested plants were not known,voucher specimens were taken to the East AfricanHerbarium in Nairobi for identification.
Stem borer species
Plants with infestation symptoms were dissected insitu for recovery of immature borers (i.e. larvae andpupae). Larvae were reared until pupation on anartificial diet according to the method described byOnyango and Ochieng-Odero (1994). Pupae wereremoved from the diet media and kept separatelyin plastic vials until adult emergence. Adultswere identified to species level by the ICIPE Bio-systematic Unit and voucher specimens were depos-ited in the ICIPE Museum (Nairobi, Kenya).
Data analysis
Diversity and abundance of wild host plant speciesat the Suam and Mtito Andei sites were computedusing the Shannon–Weaver diversity index (H)(Magurran 1988):
H = −∑PiInPi where Pi = Ni/N(relative abundance)
H: Shannon diversity index, i: host plant species, N:total crown cover of all wild host plant species in aparticular vegetation structure, Ni: total crowncover of individual wild host plant species, Pi:proportion of N made up of the ith species, In: natu-ral logarithm. The resulting product was multipliedby −1.
A t-test was used to compare the diversity indicesbetween vegetation structural formation classeswithin a location and between seasons within thesame vegetation structural formation classes.
t =H H
VarH VarH1 2
1 21 2
−+( ) /
Host plants of lepidopteran stem borers Otieno et al.
International Journal of Biodiversity Science and Management 5

where H1 is the diversity in structure 1, Var H1 itsvariance and N1 total crown cover.
Var H =pi Inpi piInpi
N
( ) ( )2 2− ∑∑ − SN−1
2 2.
The degrees of freedom were calculated using theequation
df =( )
( ) / ( ) /
VarH VarHVarH N VarH N
1 22
12
1 22
2
++
where S: plant species richness.
RESULTS
Diversity and abundance of wild hostplant species of stem borers
Suam
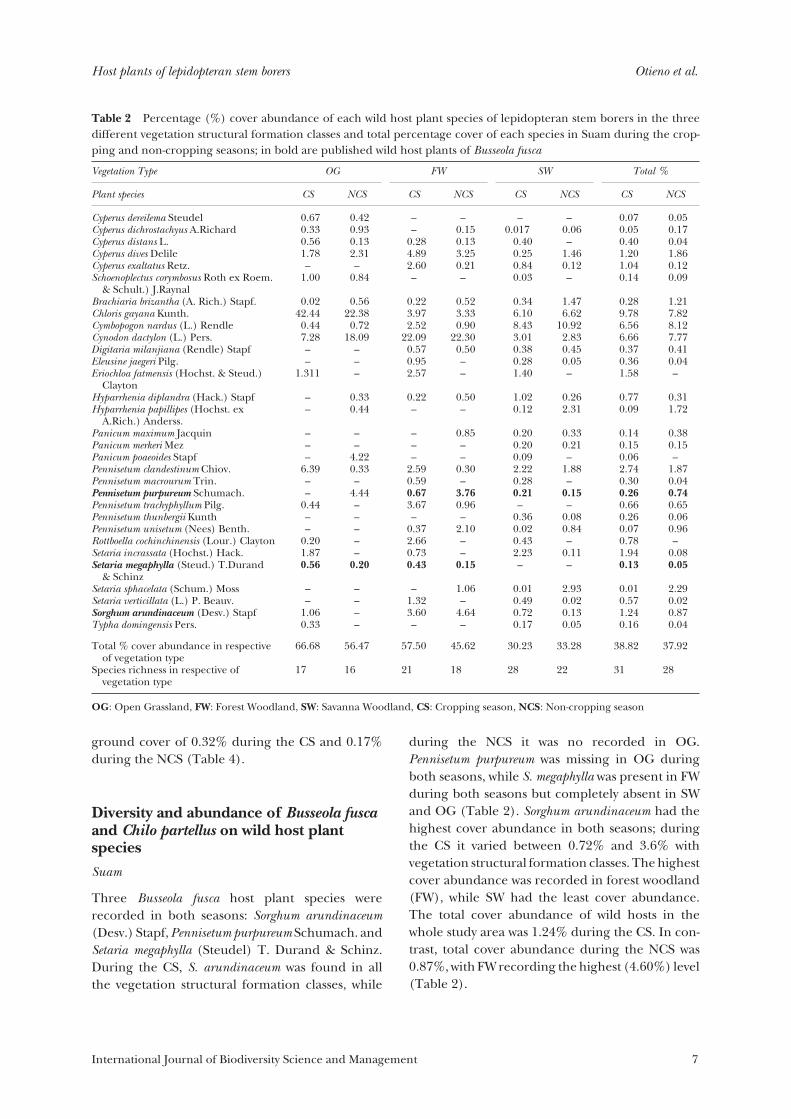
Based on the land-use map described by Goux(2005), three distinct vegetation structural forma-tion classes, namely savanna woodland (SW), forestwoodland (FW) and open grassland (OG), weresurveyed in the Suam location (Table 1). A totalof 31 uncultivated plant species in the Poaceae andCyperaceae, listed as host plants of lepidopteranstem borers by Le Ru et al. (2006a), were recorded.All of them were present during the CS and 28during the NCS (Table 2). During the CS, thenumber of host plant species varied between 17and 28 (17 species recorded in OG and 28 in SW).During the non-cropping season, the highestspecies richness (22) was recorded in SW and thelowest (16) in OG (Table 2). The species richness inthe respective vegetation structural formation classwas proportional to the size of the structure. Thehighest cover abundance of wild host plant specieswas recorded in OG in both seasons (66.7% and56.5% during CS and NCS, respectively), while thelowest cover abundance was recorded in SW(30.23% and 33.29% during CS and NCS, respec-tively) (Table 2). High-cover abundance recordedin OG was attributed to low-woody vegetation pres-ent in this vegetation structural formation class asthe area was mainly used for pasture.
The cover abundance of the wild host plantspecies and the Shannon diversity index (H) didnot vary with season (t76 = 0.98, p = 0.3302 and t76 =0.98; p = 0.4151, respectively); H varied between2.46 and 2.31 for the CS and NCS, respectively. The
highest H of 2.39 and 2.13 were recorded in SWduring the CS and NCS, respectively. Although OGrecorded the lowest H in both seasons, the NCS hada higher H (Table 3).
About 50% of the surface area surveyed wasunder maize cultivation at the time of sampling.This was higher than the surface cover of wild plantspecies (mostly Poaceae) of 11.18% and 10.92%during the CS and NCS, respectively. In general,the surface cover of wild host plants in the differentvegetation structures between the two seasons wassimilar (Table 4).
Mtito Andei
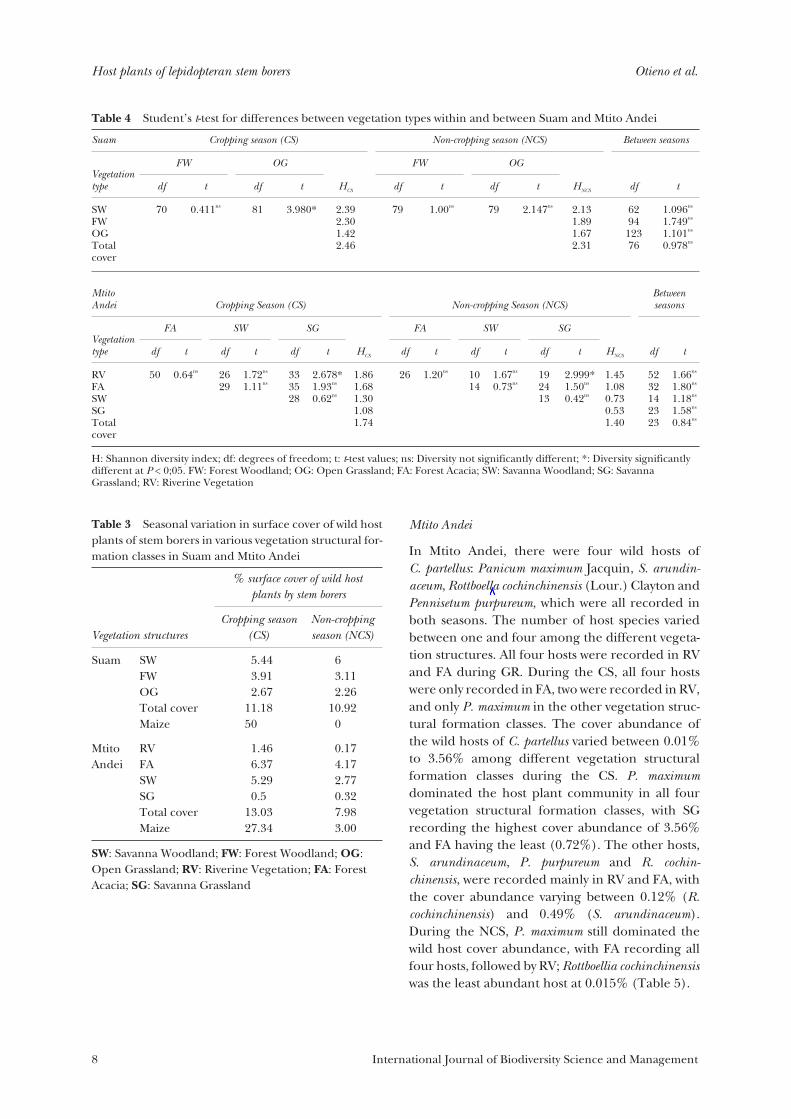
Four vegetation structures were considered forsampling in this location: savanna woodland (SW),savanna grassland (SG), forest acacia (FA) andriverine vegetation (RV). A total of 22 wild hostplant species belonging to the Poaceae and Cyper-aceae were recorded, with all of them presentduring the CS and only 16 during the NCS. Speciesrichness varied between vegetation structural for-mation classes and season. During the CS, thehighest number of species (19) was recorded in RVand the least (6) in SG; while during the NCS, thehighest species richness (11) was recorded inFA with the least (3) in SG (Table 5). Due to thewet microclimate favouring vegetation growththroughout the year, RV registered the highestcover abundance of wild host plant species, with37.3% (CS) and 19.4% (NCS) (Table 5).
No significant variation in the species diversityindex (H) was recorded between the seasons (t23 =0.83; p = 0.4151), with the CS having an index of1.73 while the NCS had 1.40. Savanna grassland(SG) showed significant variations in species diver-sity, with RV during the CS (t33 = 2.68; p = 0.01140)and the NCS (t19 = 3.00; p = 0.0074). The othervegetation structural formation classes did notshow significant differences within and between theseasons. There was no variation in cover abundanceof wild host plant species between the seasons (t23 =0.84; p = 0.4151) (Table 3).
During the cropping season, about 27% of thesurface area surveyed was under maize cultivation,while wild host plants accounted for 13% versus 3%and 8%, respectively, during the NCS (Table 4).During the NCS, maize was cultivated under irriga-tion adjacent to RV. However, RV had wild host
Host plants of lepidopteran stem borers Otieno et al.
6 International Journal of Biodiversity Science and Management
ground cover of 0.32% during the CS and 0.17%during the NCS (Table 4).
Diversity and abundance of Busseola fuscaand Chilo partellus on wild host plantspecies
Suam
Three Busseola fusca host plant species wererecorded in both seasons: Sorghum arundinaceum(Desv.) Stapf, Pennisetum purpureum Schumach. andSetaria megaphylla (Steudel) T. Durand & Schinz.During the CS, S. arundinaceum was found in allthe vegetation structural formation classes, while
during the NCS it was no recorded in OG.Pennisetum purpureum was missing in OG duringboth seasons, while S. megaphylla was present in FWduring both seasons but completely absent in SWand OG (Table 2). Sorghum arundinaceum had thehighest cover abundance in both seasons; duringthe CS it varied between 0.72% and 3.6% withvegetation structural formation classes. The highestcover abundance was recorded in forest woodland(FW), while SW had the least cover abundance.The total cover abundance of wild hosts in thewhole study area was 1.24% during the CS. In con-trast, total cover abundance during the NCS was0.87%, with FW recording the highest (4.60%) level(Table 2).
Host plants of lepidopteran stem borers Otieno et al.
International Journal of Biodiversity Science and Management 7
Vegetation Type OG FW SW Total %
Plant species CS NCS CS NCS CS NCS CS NCS
Cyperus dereilema SteudelCyperus dichrostachyus A.RichardCyperus distans L.Cyperus dives DelileCyperus exaltatus Retz.Schoenoplectus corymbosus Roth ex Roem.
& Schult.) J.RaynalBrachiaria brizantha (A. Rich.) Stapf.Chloris gayana Kunth.Cymbopogon nardus (L.) RendleCynodon dactylon (L.) Pers.Digitaria milanjiana (Rendle) StapfEleusine jaegeri Pilg.Eriochloa fatmensis (Hochst. & Steud.)
ClaytonHyparrhenia diplandra (Hack.) StapfHyparrhenia papillipes (Hochst. ex
A.Rich.) Anderss.Panicum maximum JacquinPanicum merkeri MezPanicum poaeoides StapfPennisetum clandestinum Chiov.Pennisetum macrourum Trin.Pennisetum purpureum Schumach.Pennisetum trachyphyllum Pilg.Pennisetum thunbergii KunthPennisetum unisetum (Nees) Benth.Rottboella cochinchinensis (Lour.) ClaytonSetaria incrassata (Hochst.) Hack.Setaria megaphylla (Steud.) T.Durand
& SchinzSetaria sphacelata (Schum.) MossSetaria verticillata (L.) P. Beauv.Sorghum arundinaceum (Desv.) StapfTypha domingensis Pers.
0.670.330.561.78–
1.00
0.0242.440.447.28––
1.311
––
–––
6.39––
0.44––
0.201.870.56
––
1.060.33
0.420.930.132.31–
0.84
0.5622.380.72
18.09–––
0.330.44
––
4.220.33–
4.44–––––
0.20
––––
––
0.284.892.60–
0.223.972.52
22.090.570.952.57
0.22–
–––
2.590.590.673.67–
0.372.660.730.43
–1.323.60–
–0.150.133.250.21–
0.523.330.90
22.300.50––
0.50–
0.85––
0.30–
3.760.96–
2.10––
0.15
1.06–
4.64–
–0.017
0.400.250.840.03
0.346.108.433.010.380.281.40
1.020.12
0.200.200.092.220.280.21–
0.360.020.432.23–
0.010.490.720.17
–0.06–
1.460.12–
1.476.62
10.922.830.450.05–
0.262.31
0.330.21–
1.88–
0.15–
0.080.84–
0.11–
2.930.020.130.05
0.070.050.401.201.040.14
0.289.786.566.660.370.361.58
0.770.09
0.140.150.062.740.300.260.660.260.070.781.940.13
0.010.571.240.16
0.050.170.041.860.120.09
1.217.828.127.770.410.04–
0.311.72
0.380.15–
1.870.040.740.650.060.96–
0.080.05
2.290.020.870.04
Total % cover abundance in respectiveof vegetation type
Species richness in respective ofvegetation type
66.68
17
56.47
16
57.50
21
45.62
18
30.23
28
33.28
22
38.82
31
37.92
28
OG: Open Grassland, FW: Forest Woodland, SW: Savanna Woodland, CS: Cropping season, NCS: Non-cropping season
Table 2 Percentage (%) cover abundance of each wild host plant species of lepidopteran stem borers in the threedifferent vegetation structural formation classes and total percentage cover of each species in Suam during the crop-ping and non-cropping seasons; in bold are published wild host plants of Busseola fusca
brunoleru
Barrer
Mtito Andei
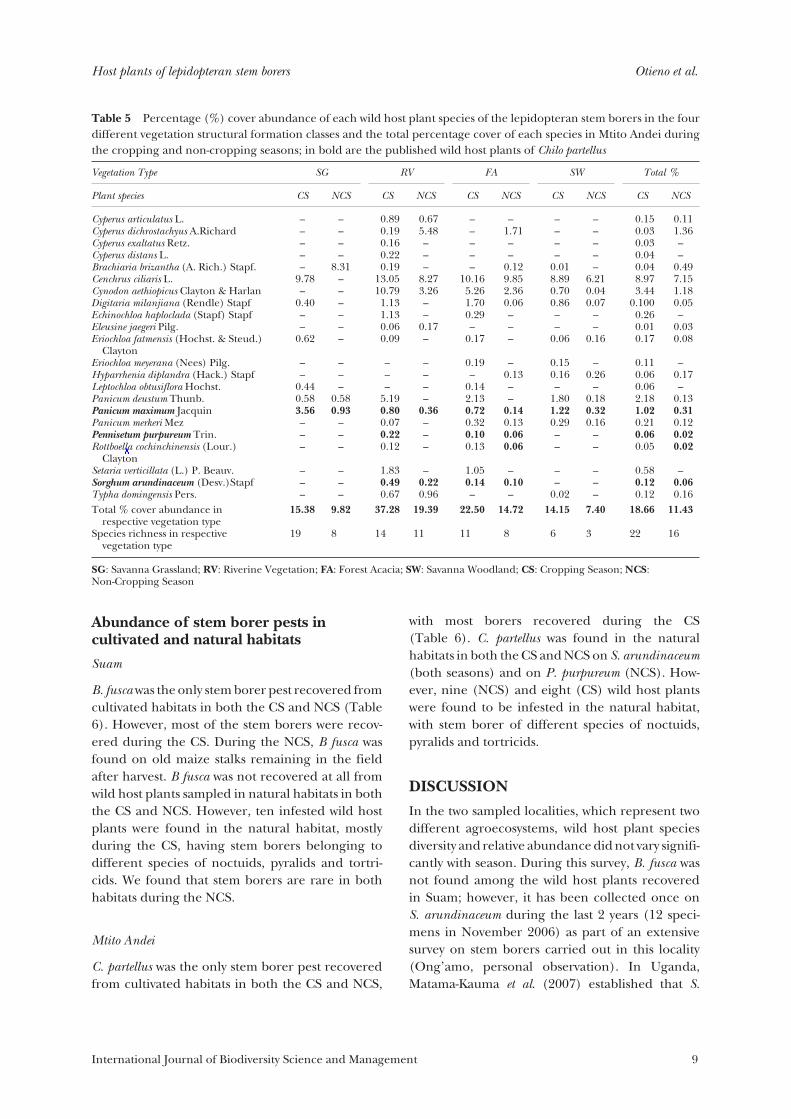
In Mtito Andei, there were four wild hosts ofC. partellus: Panicum maximum Jacquin, S. arundin-aceum, Rottboella cochinchinensis (Lour.) Clayton andPennisetum purpureum, which were all recorded inboth seasons. The number of host species variedbetween one and four among the different vegeta-tion structures. All four hosts were recorded in RVand FA during GR. During the CS, all four hostswere only recorded in FA, two were recorded in RV,and only P. maximum in the other vegetation struc-tural formation classes. The cover abundance ofthe wild hosts of C. partellus varied between 0.01%to 3.56% among different vegetation structuralformation classes during the CS. P. maximumdominated the host plant community in all fourvegetation structural formation classes, with SGrecording the highest cover abundance of 3.56%and FA having the least (0.72%). The other hosts,S. arundinaceum, P. purpureum and R. cochin-chinensis, were recorded mainly in RV and FA, withthe cover abundance varying between 0.12% (R.cochinchinensis) and 0.49% (S. arundinaceum).During the NCS, P. maximum still dominated thewild host cover abundance, with FA recording allfour hosts, followed by RV; Rottboellia cochinchinensiswas the least abundant host at 0.015% (Table 5).
Host plants of lepidopteran stem borers Otieno et al.
8 International Journal of Biodiversity Science and Management
% surface cover of wild hostplants by stem borers
Vegetation structuresCropping season
(CS)Non-croppingseason (NCS)
Suam SWFWOGTotal coverMaize
5.443.912.67
11.1850
63.112.26
10.920
MtitoAndei
RVFASWSGTotal coverMaize
1.466.375.290.5
13.0327.34
0.174.172.770.327.983.00
SW: Savanna Woodland; FW: Forest Woodland; OG:Open Grassland; RV: Riverine Vegetation; FA: ForestAcacia; SG: Savanna Grassland
Table 3 Seasonal variation in surface cover of wild hostplants of stem borers in various vegetation structural for-mation classes in Suam and Mtito Andei
Suam Cropping season (CS) Non-cropping season (NCS) Between seasons
Vegetationtype
FW OG FW OG
df t df t HCS df t df t HNCS df t
SWFWOGTotalcover
70 0.411ns 81 3.980* 2.392.301.422.46
79 1.00ns 79 2.147ns 2.131.891.672.31
629412376
1.096ns
1.749ns
1.101ns
0.978ns
Table 4 Student’s t-test for differences between vegetation types within and between Suam and Mtito Andei
MtitoAndei Cropping Season (CS) Non-cropping Season (NCS)
Betweenseasons
Vegetationtype
FA SW SG FA SW SG
df t df t df t HCS df t df t df t HNCS df t
RVFASWSGTotalcover
50 0.64ns 2629
1.72ns
1.11ns333528
2.678*1.93ns
0.62ns
1.861.681.301.081.74
26 1.20ns 1014
1.67ns
0.73ns192413
2.999*1.50ns
0.42ns
1.451.080.730.531.40
5232142323
1.66ns
1.80ns
1.18ns
1.58ns
0.84ns
H: Shannon diversity index; df: degrees of freedom; t: t-test values; ns: Diversity not significantly different; *: Diversity significantlydifferent at P < 0;05. FW: Forest Woodland; OG: Open Grassland; FA: Forest Acacia; SW: Savanna Woodland; SG: SavannaGrassland; RV: Riverine Vegetation
brunoleru
Texte inséré
i
Abundance of stem borer pests incultivated and natural habitats
Suam
B. fusca was the only stem borer pest recovered fromcultivated habitats in both the CS and NCS (Table6). However, most of the stem borers were recov-ered during the CS. During the NCS, B fusca wasfound on old maize stalks remaining in the fieldafter harvest. B fusca was not recovered at all fromwild host plants sampled in natural habitats in boththe CS and NCS. However, ten infested wild hostplants were found in the natural habitat, mostlyduring the CS, having stem borers belonging todifferent species of noctuids, pyralids and tortri-cids. We found that stem borers are rare in bothhabitats during the NCS.
Mtito Andei
C. partellus was the only stem borer pest recoveredfrom cultivated habitats in both the CS and NCS,
with most borers recovered during the CS(Table 6). C. partellus was found in the naturalhabitats in both the CS and NCS on S. arundinaceum(both seasons) and on P. purpureum (NCS). How-ever, nine (NCS) and eight (CS) wild host plantswere found to be infested in the natural habitat,with stem borer of different species of noctuids,pyralids and tortricids.
DISCUSSION
In the two sampled localities, which represent twodifferent agroecosystems, wild host plant speciesdiversity and relative abundance did not vary signifi-cantly with season. During this survey, B. fusca wasnot found among the wild host plants recoveredin Suam; however, it has been collected once onS. arundinaceum during the last 2 years (12 speci-mens in November 2006) as part of an extensivesurvey on stem borers carried out in this locality(Ong’amo, personal observation). In Uganda,Matama-Kauma et al. (2007) established that S.
Host plants of lepidopteran stem borers Otieno et al.
International Journal of Biodiversity Science and Management 9
Vegetation Type SG RV FA SW Total %
Plant species CS NCS CS NCS CS NCS CS NCS CS NCS
Cyperus articulatus L.Cyperus dichrostachyus A.RichardCyperus exaltatus Retz.Cyperus distans L.Brachiaria brizantha (A. Rich.) Stapf.Cenchrus ciliaris L.Cynodon aethiopicus Clayton & HarlanDigitaria milanjiana (Rendle) StapfEchinochloa haploclada (Stapf) StapfEleusine jaegeri Pilg.Eriochloa fatmensis (Hochst. & Steud.)
ClaytonEriochloa meyerana (Nees) Pilg.Hyparrhenia diplandra (Hack.) StapfLeptochloa obtusiflora Hochst.Panicum deustum Thunb.Panicum maximum JacquinPanicum merkeri MezPennisetum purpureum Trin.Rottboella cochinchinensis (Lour.)
ClaytonSetaria verticillata (L.) P. Beauv.Sorghum arundinaceum (Desv.)StapfTypha domingensis Pers.
–––––
9.78–
0.40––
0.62
––
0.440.583.56–––
–––
––––
8.31––––––
–––
0.580.93
–––
–––
0.890.190.160.220.19
13.0510.791.131.130.060.09
–––
5.190.800.070.220.12
1.830.490.67
0.675.48–––
8.273.26––
0.17–
––––
0.36–––
–0.220.96
–––––
10.165.261.700.29–
0.17
0.19–
0.142.130.720.320.100.13
1.050.14–
–1.71––
0.129.852.360.06–––
–0.13––
0.140.130.060.06
–0.10–
––––
0.018.890.700.86––
0.06
0.150.16–
1.801.220.29––
––
0.02
–––––
6.210.040.07
––
0.16
–0.26
–0.180.320.16
––
–––
0.150.030.030.040.048.973.44
0.1000.260.010.17
0.110.060.062.181.020.210.060.05
0.580.120.12
0.111.36––
0.497.151.180.05–
0.030.08
–0.17–
0.130.310.120.020.02
–0.060.16
Total % cover abundance inrespective vegetation type
Species richness in respectivevegetation type
15.38
19
9.82
8
37.28
14
19.39
11
22.50
11
14.72
8
14.15
6
7.40
3
18.66
22
11.43
16
SG: Savanna Grassland; RV: Riverine Vegetation; FA: Forest Acacia; SW: Savanna Woodland; CS: Cropping Season; NCS:Non-Cropping Season
Table 5 Percentage (%) cover abundance of each wild host plant species of the lepidopteran stem borers in the fourdifferent vegetation structural formation classes and the total percentage cover of each species in Mtito Andei duringthe cropping and non-cropping seasons; in bold are the published wild host plants of Chilo partellus
brunoleru
Texte inséré
i

arundinaceum was the principal host of B. fusca,while Ingram (1958) hypothesised that grassesoccurring next to cultivated fields, may constitute alarge reservoir of stem borers. The low-surfacecover (0.5%) of S. arundinaceum, in either seasoncorroborates results from work done earlier in thesame district (Trans-Nzoia) (Kanya et al. 2005). Thisis an indication that natural habitats do not serveas a major reservoir for B. fusca during the non-cropping season. Moreover, in Suam, it is a normalpractice for maize stalks to be cleared by burning,which normally extends to patches of the wild habi-tats surrounding the maize field, thereby destroy-ing also the wild host plants of B. fusca. Tscharntkeand Brandl (2003) noted that destruction of thewild habitat of the borers might disrupt theplant–herbivore interaction, thereby affecting thepest population in the wild habitat. However,diapausing B. fusca larvae have been recoveredfrom old maize stalks that survive burning during
land preparation (Ong’amo, personal observa-tions), suggesting that the majority of B. fusca larvaepass the NCS on maize residues in Suam.
Prior to the present surveys, a prolongeddrought of more than 3 years occurred in MtitoAndei (Le Ru, personal observation). Since then,green vegetation has been confined to the river-banks, leaving the rest of the study area with drytillers of wild host plants. This explains, in part, whythere was no variation in both species richness andrelative abundance between the seasons despite theprolonged drought. Although not very abundant,C. partellus was found among the wild host plantsrecovered in Mtito Andei during this survey.According to the surveys done by Le Ru et al.(2006b), C. partellus was present all year round inthis area, but was minimal during the severedrought. This suggests that the wild host plantspresent along the riverbank could support largeC. partellus population. In addition, some farmers
Host plants of lepidopteran stem borers Otieno et al.
10 International Journal of Biodiversity Science and Management
CS NCS
Infested plant species B. fusca C. partellus Other borers B. fusca C. partellus Other borers
Suam C. nardusC. dactylonC. divesP. maximumP. macrourumP. purpureumP. trachyphyllumS. confususS. arundinaceumS. incrassataS. bicolor*Zea mays*
–––––––––––
251
––––––––––––
132
10761
71311––
––––––––––1
14
––––––––––––
–––––––8––––
Mtito Andei C. aethiopicusC. distansC. involucratusE. jaegeriP. deustumP. maximumP. purpureumR. cochinchinensisS. verticillataS. arundinaceumT. domingensisS. bicolor*Z. mays*
–––––––––––––
––––––2––
46–
63148
410531
288––
16285
36
–––––––––––––
–––––––––
11––
22
16–
22–1
13–111
16–
17
Table 6 Number of stem borer larvae recovered in Suam and Mtito Andei from various host plants during differentseasons. Asterisks indicate the cereal plants found infested
brunoleru
Barrer
brunoleru
Texte de remplacement
corymbosus
brunoleru
Barrer
brunoleru
Texte de remplacement
insect

were able to grow maize under irrigation in fieldsadjacent to the riverbanks during the dry season(Otieno, personal observation), thus enhancingthe survival of the pest.
Recently, Bt maize resistant to C. partellus hasbeen introduced into Kenya (Mugo et al. 2005). Inorder to prevent development of insect resistanceto the Bt toxin, it is generally recommended thatnot less than 20% of the cropping area should beplanted with non-transgenic maize (i.e. refuge areastrategy, sensu Brousseau et al. 1999), which wouldsustain a susceptible pest population (Fitt et al.2004). It was considered that, in Africa, naturalhosts of stem borer pests growing in natural habitatsmay serve as refuges (Khan et al. 1997). However,according to the results presented here, less than5% of the area of both localities is occupied by wildhost plants of stem borer pests, and even less duringthe non-cropping season. Furthermore, our resultsshow that even if C. partellus was found in the natu-ral habitat surrounding cereal plots, B fusca wasabsent. Thus, it can be concluded that wild habitats
surrounding cereal plots will play little if any role inthe resistance management of Bt maize targetingeither C. partellus or B. fusca. Additional refuges areneeded to complement the wild host plants to delayresistance development in stem borer pests to trans-genic crops. However, further research on adultstem borer dispersal is required, since movement ofB. fusca from distant (up to several km) wild hostsor crops to the maize crop cannot be ruled out.
ACKNOWLEDGEMENTS
This work was supported by the Institut deRecherche pour le Développement (IRD) throughthe insect–plant interaction unit (R072) in collabo-ration with the International Centre of InsectPhysiology and Ecology (ICIPE). The authors grate-fully acknowledge staff of the Noctuid Stem BorerBiodiversity Project (NSBB), especially LeonardNgala for technical support and Simon Mathenge(Botany Department, University of Nairobi) foridentification of plant specimens.
REFERENCESBowden J. Stem borer ecology and strategy for control.
Annals of Applied Biology 1976;84:107–11Brousseau R, Masson L and Hegedus D. Insecticidal
transgenic plants: are they irresistible. AdbiotechNet1999;I(July): ABN 022
Corbett JD and O’Brien RF. The Spatial CharacterizationTool – African version 1.0. Report No. 97–03,CDROM. Texas Agricultural Experiment Station,Texas A & M University, Blackland ResearchCenter, Texas; 1997
FAO (Food and Agriculture Organization of theUnited Nations). Production Yearbook for 2002.Rome, Italy: FAO; 2003
Fitt GP, Andow DA, Carrière Y, Moar WJ, Schuler TH,Omoto C, Kanya J, Okech MA, Arama P andManiania NK. Resistance risks and managementassociated with Bt Maize in Kenya. In Hilbeck Aand Andow DA (eds), Environmental risk assessmentof genetically modified organisms. Oxford: CABI;2004;209–50
Guofa Z, Overholt WA and Mochiah MB. Changes inthe distribution of lepidopteran maize stem borersin Kenya from 1950s to 1990s. Insect Science and itsApplication 2002;21:395–402
Goux I. Characterization of wild and cultivated habitatsof insect pests in maize crops (Zea mays L.) in twolocalities, Mtito Andei and Suam in Kenya. MSc in
Management of Agro-Systems in Tropical Areas,Université de Paris XII, Paris; 2005
Grieg-Smith P. Quantitative Plant Ecology. 3rd ed.Oxford: Blackwell; 1983
Harris KM. Lepidopteran stem borers of cereals inNigeria. Bulletin of Entomological Research 1962;53:139–71
Ingram WR. The lepidopterous stalk borers associatedwith Gramineae in Uganda. Bulletin of EntomologicalResearch 1958;49:367–83
Kanya JI, Ngi-song AJ, Setamou MF, Overholt W,Ochora J and Osir EO. Diversity of alternative hostsof maize stem borers in Trans-Nzoia district ofKenya. Environmental Biosafety Research 2005;3:159–68
Karandinos MG. Optimum sample size and commentson some published formulae. Bulletin of the Entomo-logical Society of America 1976;22:417–21
Kfir R, Overholt WA, Khan ZR and Polaszek A. Biologyand management of economically importantlepidopteran cereal stem borers in Africa. AnnualReview of Entomology 2002;47:701–31
Khan ZR, Chiliswa P, Ampong-Nyarko K, Smart LE,Polaszek A, Wandera J and Mulaa MA. Utilisationof wild gramineous plants for management ofcereal stem borers in Africa. Insect Science and itsApplication 1997;17:143–50
Host plants of lepidopteran stem borers Otieno et al.
International Journal of Biodiversity Science and Management 11

Le Ru, BP, Ong’amo GO, Moyal P, Ngala L, MusyokaB, Abdullah Z, Cugala D, Defabachew B, Haile TA,Kauma Matama T, Lada VY, Negassi B, PallangyoK, Ravololonandrianina J, Sidumo A, Omwega C,Schulthess F, Calatayud P-A and Silvain, JF. Diver-sity of lepidopteran stem borers on monocotyle-donous plants in eastern Africa and the islandsof Madagascar and Zanzibar revisited. Bulletin ofEntomological Research 2006a;96:555–63
Le Ru BP, Ong’amo GO, Moyal S, Muchungu E, NgalaL, Musyoka B, Abdulla Z, Matama-Kauma T, LadaVY, Pallangyo B, Omwega CO, Schulthess F,Calatayud P-A and Silvain JF. Geographic distribu-tion and host plant ranges of East Africa noctuidstem borers. Annales de la Société Entomologique deFrance 2006b;42:353–61
Magurran AE. Ecological diversity and its measurement.Princeton: Princeton University Press; 1988
Matama-Kauma T, Schulthess F, Le Ru BP, Mueke JM,Ogwang JA and Omwega CO. Abundance anddiversity of lepidopteran stem borers and theirparasitoids on selected wild grasses in Uganda.Crop Protection 2008;27:505–13
Mugo S, De Groote H, Bergvinson D, Mulaa M, Songa Jand Gichuki S. Developing Bt maize for resource-poor farmers – Recent advances in the IRMAproject. African Journal of Biotechnology 2005;4:1490–504
Ndemah R, Schulthess F, Le Ru B and Bame I.Lepidopteran cereal stem borers and associatednatural enemies in maize and wild grass hosts inCameroon. Journal of Applied Entomology 2007;131(9–10):658–68
Nye IWR. The insect pests of graminaceous crops inEast Africa. Colonial Research Studies 1960;31:1–48
Ong’amo GO, Le Ru B, Dupas S, Moyal P, CalatayudP-A and Silvain JF. Distribution, pest status and
agroclimatic preferences of lepidopteran stemborers of maize and sorghum in Kenya. Annales dela Société Entomologique de la France 2006;42:171–7
Onyango FO and Ochieng’Odero JPR. Continuousrearing of the maize stem borer Busseola fusca on anartificial diet. Entomologia experimentalis et Applicata1994;73:139–44
Otieno NA, Le Ru BP, Ong’amo GO, Dupas S,Calatayud P-A, Makobe M, Ochora J and Silvain JF.Diversity and abundance of wild host plants oflepidopteran stem borers in two different agro-ecological zones of Kenya. Annales de la SociétéEntomologique de France 2006;42:371–80
Phillips S. Flora of Ethiopia and Eritrea Volume 7. Poaceae(Gramineae). Addis Ababa, Ethiopia; Uppsala,Sweden; 1995
Polaszek A and Khan ZR. Host plants. In Polaszek A(ed.), African cereal stem borers: economic importance,taxonomy, natural enemies and control. Cambridge:CAB International, University Press; 1998:5–23
Schulthess F, Bosque-Pérez NA, Chabi-Olaye A,Gounou S, Ndemah R and Goergen G. Exchangeof natural enemies of lepidopteran cereal stemborers between African regions. Insect Science andits Application 1997;17:97–108
Seshu Reddy KV. Study of the stem borer complex ofsorghum in Kenya. Insect Science and its Application1983;4:3–10
Tscharntke T and Brandl R. Plant–insect interactionsin fragmented landscapes. Annual Review ofEntomology 2003;49:405–30
Van Oudtshoorn F. Guide to Grasses of Southern Africa.2nd edition. Pretoria, SA: Briza; 2004
Webster R and Oliver MA. Statistical methods in soil andland resource survey. New York: Oxford UniversityPress; 1990
Zar JH. Biostatistical Analysis. 4th edition. New Jersey:Prentice Hall; 1999:40–44
Host plants of lepidopteran stem borers Otieno et al.
12 International Journal of Biodiversity Science and Management




















