
Novel insight into the role of heterotrophic dinoflagellates in the fate of crude oil in the sea
Upload
independentCategory
view
2download
0
Distribution patterns of isomorphic cold-water dinoflagellates
(Scrippsiella/Woloszynskia complex) causing ‘red tides’ in the Baltic Sea
Andres Jaanus1,*, Susanna Hajdu2, Seppo Kaitala3, Agneta Andersson4, Kaire Kaljurand1,Iveta Ledaine5, Inga Lips1 & Irina Olenina61Estonian Marine Institute, Tartu University, Maealuse 10a, 12618, Tallinn, Estonia2Department of Systems Ecology, Marine and Brackish Water Ecology, Stockholm University, Svante Arrheniusvag 21 A,
SE-106 91, Stockholm, Sweden3Finnish Institute of Marine Research, P.O. Box 33, FIN-00931, Helsinki, Finland4Dept of Ecology and Environmental Science, Umea Marine Sciences Centre, SE-910 20, Hornefors, Sweden5Inst. of Aquatic Ecology, Marine Monitoring Center, University of Latvia, Daugavgrivas str., 8 Riga, LV-1048, Latvia6Centre of Marine Research, Taikos str., 26, LT-5802, Klaipeda, Lithuania(*Author for correspondence: Tel.: +372-671-8974; Fax: +372-671-8973; E-mail: [email protected])
Key words: Baltic Sea, dinoflagellates, Scrippsiella/Woloszynskia complex
Abstract
During the latest years medium-sized (15–30 lm), single-celled dinoflagellates have been reported to formblooms in the northern Baltic Proper and the Gulf of Finland in winter and spring. Recent studies(Kremp et al., 2003. Proceedings of the 7th International conference of Modern and Fossil Dinoflagel-lates, September 21–25, Nagasaki, Japan, 66 pp.) indicate that those blooms are caused by twoisomorphic species – Scrippsiella hangoei (Schiller) Larsen, and a new species, tentatively belonging to thegenus Woloszynskia. Until now there has been no report on how widely distributed these phytoplanktonspecies are in the Baltic Sea. In this study, the occurrence of Scrippsiella/Woloszynskia complex in theentire Baltic Sea was investigated, by using monitoring data from 1997 to 2003. The species occurred in asalinity range from 2 to 8 PSU. Highest concentrations were observed at salinity 4.5–6.5 PSU. Maximumcell densities of Scrippsiella/Woloszynskia complex in the water column were mainly obtained in April orin the beginning of May by the water temperature <3 �C prior to stratification was formed. In thecentral Gulf of Finland, the second maximum was found in 1999 and 2002 by the temperature >6 �C.Bloom formations in the Baltic Proper and in the Gulf of Finland may not only be explained byoptimum temperature and salinity, but also with other factors e.g. high nutrient concentrations and goodseeding conditions from the sediments.
Introduction
During the last decades ‘red tides’ of dinoflagel-lates have become a regular phenomenon duringspring period in the northern Baltic Proper and theGulf of Finland (e.g. Lignell et al., 1993). Theblooms have been attributed to a single-celled,medium-sized (15–30 lm) species with delicatethecal plates called Scrippsiella hangoei (Schiller)
Larsen. Before its re-description by Larsen et al.(1995) S. hangoei was probably recognised underdifferent taxonomical names – Peridinium sp., P.hangoei, Gymnodinium sp., Glenodinium sp. (e.g.Niemi, 1975; Hobro, 1979; HELCOM, 1990, 1996;Heiskanen, 1993; Lignell et al., 1993). However,recent molecular and SEM analyses (Kremp et al.,2003) revealed considerable differences to thespecies description of Larsen et al. (1995), imply-
Hydrobiologia (2006) 554:137–146 � Springer 2006J. Kuparinen, E. Sandberg-Kilpi & J. Mattila (eds), Baltic Sea: A Lost System or a Future TreasuryDOI 10.1007/s10750-005-1014-7

ing that another, hitherto unrecognized speciesco-occurs with S. hangoei during spring bloom.SEM images of this dinoflagellate suggest assign-ment of this species to the genus Woloszynskia.
S. hangoei as well as Woloszynskia sp. are moreor less isomorphic (Kremp et al., 2003) and cannotbe unequivocally identified in Lugol fixed samples.Re-analyzing the historical and even recent sam-ples to specify the taxonomical composition isimpossible or requires different techniques, whichare not used in routine monitoring. Of these rea-sons we use the concept of Scrippsiella/Wol-oszynskia complex further in the text.
The temporal sequence and also the peculiar-ities of bloom onset and development of Scrip-psiella/Woloszynskia on the coastal–open seagradient have been studied thoroughly during thelast decade (Kremp, 2000a, b; Kremp & Heiska-nen, 1999). At the same time we had relativelypoor knowledge about the distribution of thiscomplex on a larger basin-wide scale. S. hangoeisensu Larsen et al. (1995) has been consideredendemic, which is adapted to the specific lowsaline environment of the Baltic Sea. Earlierstudies and laboratory experiments had mostlybeen conducted in a restricted area in the north-ern coastal Baltic. In the present paper, we try tomark out the general features of the spatio-temporal dynamics, supported mainly by themonitoring data from different sub-areas of theBaltic Sea.
We give an account that the taxonomic con-fusions considerably hamper the assessment ofthe distribution and quantitative characteristicsof medium-sized single-celled dinoflagellates. Inorder to avoid a blend of several genus names,which have been probably used for the samecomplex in earlier records (before 1995), weevaluated only recent data (1997–2003) in ourstudy.
Material and methods
The samples were collected within the frameof selected national, HELCOM and Alg@linemonitoring programmes in 1997–2003. The num-ber of monitoring stations by different sub-basinswas the following: Gulf of Finland 18 (incl. 10
Alg@line stations), Gulf of Riga 10, Baltic Proper28, Gulf of Bothnia 6 stations and one station inthe Aland Sea (Fig. 1, Table 1). The salinity rangewas between 1.3 in the Stockholm Archipelagoand 7.2 PSU in the southern Baltic Proper. Theselected stations also represent very diversebathymetric structure from shallow bays to deep-est basins in the Baltic Sea and from erosion toactive accumulation areas (Table 1).
In the coastal and open sea monitoring stationsintegrated samples from surface to 10 or 20 mdepth were taken mainly according to the recom-mendations of the BMP (HELCOM, 1988). Somedifferences in sampling methods were caused bydifferent equipment used. Swedish stations weresampled from the photic zone (0–20 m) using ahose (a plastic tube with inside diameter of25 mm), as the samples from Estonian, Latvianand Lithuanian stations were pooled from discretedepths (1, 5 and 10 m). To make counting resultsof 0–10 m comparable with those of 0–20 m, datafrom 0–20 m were multiplied by a factor of 1.36.This factor was derived from intensive verticalstudies during the spring of 1996 at the Swedishoffshore station H3 (Hoglander et al., 2004).Alg@line stations were sampled weekly fromApril to early June from �5 m depth onboardmoving ship by the flow-through system pumps(Rantajarvi & Leppanen, 1994).
Sampling frequency was mostly 1–2 times amonth from March to early June in 1997–2003with some interruptions or reductions dependingon the specific monitoring programmes or on themeteorological conditions (ice-cover). Station H3and H12 were collected weekly during the ongoingspring bloom (mainly April) and biweekly there-after. 1058 samples were analysed in total, 71% ofthose represent April-May, the normal period forphytoplankton spring bloom in the northern andcentral parts of the Baltic Sea (e.g. Hallfors &Niemi, 1981).
Samples were preserved with acetic Lugol’ssolution and counted by the inverted microscopetechnique following the recommendations ofHELCOM (1988). Biomass calculations weredone by multiplying the cell number with the cellvolume, derived from measurements of cell size.We used five categories to characterise thepeak abundances of Scrippsiella/Woloszynskiacomplex:
138

Results
Spatial distribution
Distribution of Scrippsiella/Woloszynskia withindication of relative maximum abundances in
different areas of the Baltic Sea is presented inFigure 1. The map is based on the maximum cellnumber found in all collected samples at eachstation (Table 1). Despite of different samplingstrategies we were able to demonstrate the loca-tions of the strongest bloom potential of the spe-cies. The maximum abundances were highest in theGulf of Finland – up to 10 · 106 cells l)1. At somestations in the northern Baltic Proper and near theLithuanian coast the peak abundance indicatedmoderate blooms (0.5–1 · 106 cells l)1). Cell den-sity decreased gradually both towards the southernBaltic Proper and northward in the Gulf ofBothnia. Scrippsiella/Woloszynskia was also lessabundant in the coastal areas under the influence
.
Maximum cells/l Rank
Seeding banks >5 000 000 5
Strong blooms 1 000 000–5 000 000 4
Moderate blooms 500 000–1 000 000 3
Common 100 000–500 000 2
Present <100 000 1
Figure 1. Sampling locations and distribution of the dinoflagellate Scrippsiella/Woloszinskia complex in the Baltic Sea. Scale bar for
maximum cell densities: 5 – Seeding banks, 4 – Strong blooms, 3 – Moderate blooms, 2 – Common, 1 – Present, 0 – Absent or no
information.
139
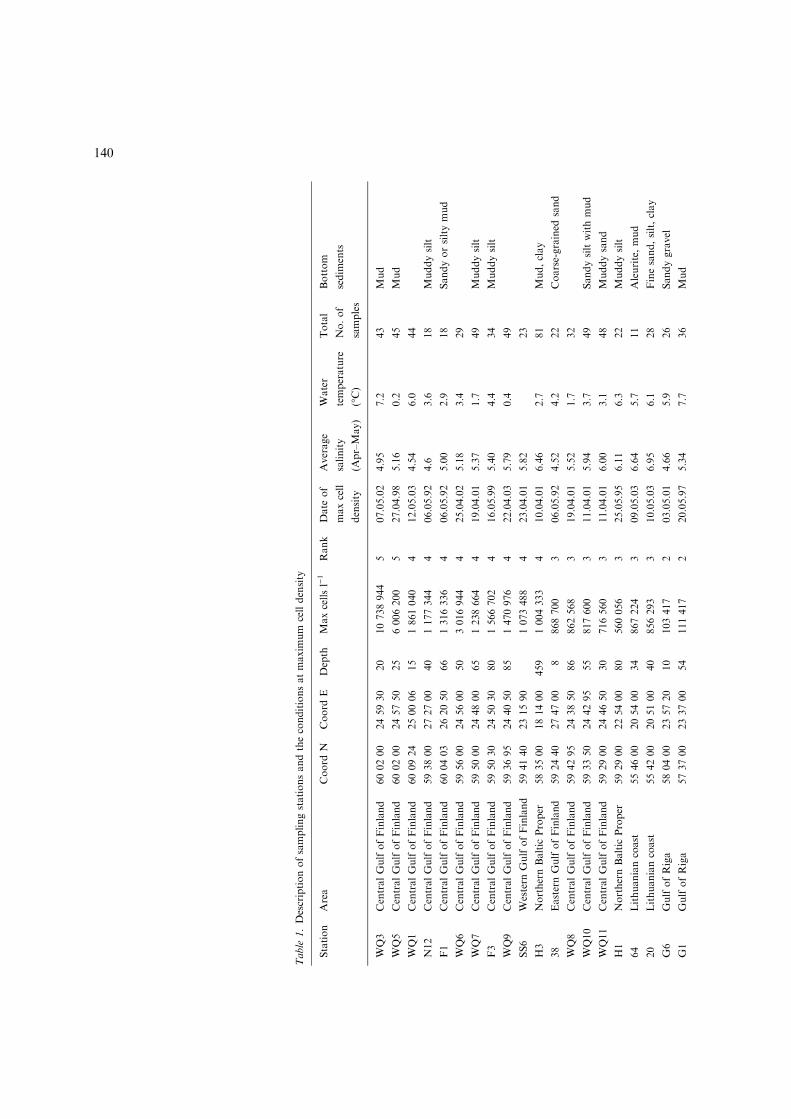
Table
1.Descriptionofsamplingstationsandtheconditionsatmaxim
um
celldensity
Station
Area
Coord
NCoord
EDepth
Maxcellsl)1
Rank
Date
of
maxcell
density
Average
salinity
(Apr–May)
Water
temperature
(�C)
Total
No.of
samples
Bottom
sedim
ents
WQ3
CentralGulfofFinland
600200
245930
20
10738944
507.05.02
4.95
7.2
43
Mud
WQ5
CentralGulfofFinland
600200
245750
25
6006200
527.04.98
5.16
0.2
45
Mud
WQ1
CentralGulfofFinland
600924
250006
15
1861040
412.05.03
4.54
6.0
44
N12
CentralGulfofFinland
593800
272700
40
1177344
406.05.92
4.6
3.6
18
Muddysilt
F1
CentralGulfofFinland
600403
262050
66
1316336
406.05.92
5.00
2.9
18
Sandyorsiltymud
WQ6
CentralGulfofFinland
595600
245600
50
3016944
425.04.02
5.18
3.4
29
WQ7
CentralGulfofFinland
595000
244800
65
1238664
419.04.01
5.37
1.7
49
Muddysilt
F3
CentralGulfofFinland
595030
245030
80
1566702
416.05.99
5.40
4.4
34
Muddysilt
WQ9
CentralGulfofFinland
593695
244050
85
1470976
422.04.03
5.79
0.4
49
SS6
Western
GulfofFinland
594140
231590
1073488
423.04.01
5.82
23
H3
NorthernBaltic
Proper
583500
181400
459
1004333
410.04.01
6.46
2.7
81
Mud,clay
38
EasternGulfofFinland
592440
274700
8868700
306.05.92
4.52
4.2
22
Coarse-grained
sand
WQ8
CentralGulfofFinland
594295
243850
86
862568
319.04.01
5.52
1.7
32
WQ10
CentralGulfofFinland
593350
244295
55
817600
311.04.01
5.94
3.7
49
Sandysiltwithmud
WQ11
CentralGulfofFinland
592900
244650
30
716560
311.04.01
6.00
3.1
48
Muddysand
H1
NorthernBaltic
Proper
592900
225400
80
560056
325.05.95
6.11
6.3
22
Muddysilt
64
Lithuaniancoast
554600
205400
34
867224
309.05.03
6.64
5.7
11
Aleurite,mud
20
Lithuaniancoast
554200
205100
40
856293
310.05.03
6.95
6.1
28
Finesand,silt,clay
G6
GulfofRiga
580400
235720
10
103417
203.05.01
4.66
5.9
26
Sandygravel
G1
GulfofRiga
573700
233700
54
111417
220.05.97
5.34
7.7
36
Mud
140

M3
CentralGulfofFinland
593280
245700
43
186004
220.04.94
5.71
1.3
10
Fine-grained
sand
SS2
ArchipelagoSea
600000
194500
151424
205.05.02
6.00
17
E2
CentralGulfofFinland
593220
244130
44
312732
208.05.03
6.03
5.0
33
Coarse-grained
sand
57a
CentralGulfofFinland
592700
244730
7225784
222.04.02
6.08
3.2
32
Muddysand
H12
Swedisheast
coast
584818
173752
42
220476
226.04.02
6.29
4.3
80
Mud,clay
1B
Lithuaniancoast
560200
205000
27
366202
208.05.03
6.50
4.9
11
Boulders,coarsesand
J57
Lithuaniancoast
555600
205800
18
317011
208.05.03
6.51
6.3
11
Boulders,coarsesand
1Lithuaniancoast
560200
210100
16
307901
208.05.03
6.54
5.9
8Finesand,silt,clay
H2
NorthernBaltic
Proper
590200
210500
180
185469
224.05.95
6.63
5.9
14
Mud
SS4
NorthernBaltic
Proper
592640
212148
203944
203.05.99
6.65
3.7
6
5Lithuaniancoast
554300
210400
15
118424
209.05.03
6.77
6.2
11
Finesand,silt
7Lithuaniancoast
551900
205700
15
428147
209.05.03
6.85
7.5
11
Aleurite
J2Lithuaniancoast
555300
202000
47
329764
209.05.03
7.02
5.0
13
Aleurite,mud
K2
Bornholm
Sea
551500
155900
90
295717
228.03.01
7.10
2.7
15
4Lithuaniancoast
554400
210300
16
109313
209.05.03
7.10
6.2
11
Finesand,silt
J1EasternGotlandBasin
572000
200600
240
274992
227.04.00
7.14
5.8
30
Mud,clay
K4
Bornholm
Sea
550000
140500
47
354960
227.03.99
7.20
2.9
13
C1
BothnianSea
623500
195900
212
113424
204.06.99
5.22
13
Rankingformaxim
um
celldensity:5–Seedingbanks(>
5*106l)1),4–Strongblooms(1–5*106l)1),3–Moderate
blooms(0.5–1*106l)1),2–Common(1–5*105l)1).
141
of fresh-water inflow (salinity <4 PSU). In morethan 70% of all stations the maximum cell num-bers did not exceed 0.5 · 106 per litre, which weconsidered insufficient to define as bloom-likedensity.
In areas where Scrippsiella/Woloszynskia formedmoderate to strong blooms, the surface layer(0–10 m) salinity varied from 4.52 to 6.46 PSU. Thetwo Lithuanian coastal stations (20 and 64) had,however, higher salinity (6.64–6.95 PSU). Somewhatunexpectedly, in the Gulf of Riga, the area withsalinity from 4 to 6 PSU, Scrippsiella/Woloszynskiacomplex did not belong to the dominant species.
Temporal sequence of the bloom
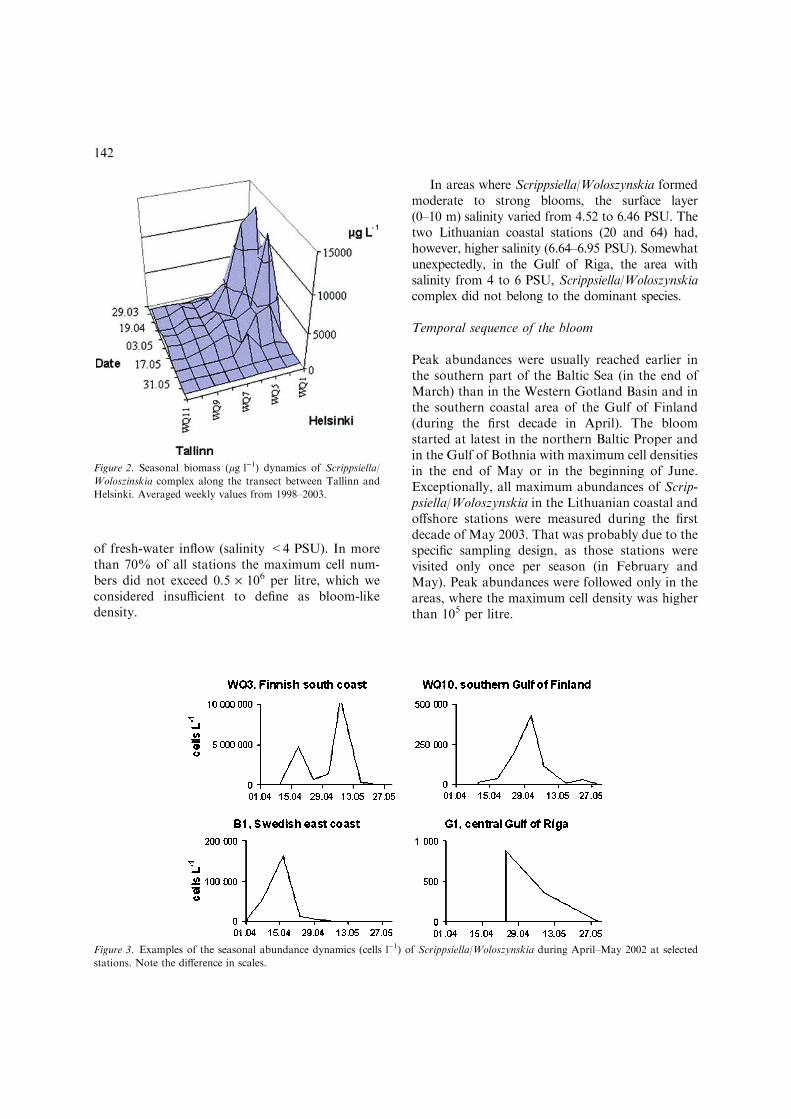
Peak abundances were usually reached earlier inthe southern part of the Baltic Sea (in the end ofMarch) than in the Western Gotland Basin and inthe southern coastal area of the Gulf of Finland(during the first decade in April). The bloomstarted at latest in the northern Baltic Proper andin the Gulf of Bothnia with maximum cell densitiesin the end of May or in the beginning of June.Exceptionally, all maximum abundances of Scrip-psiella/Woloszynskia in the Lithuanian coastal andoffshore stations were measured during the firstdecade of May 2003. That was probably due to thespecific sampling design, as those stations werevisited only once per season (in February andMay). Peak abundances were followed only in theareas, where the maximum cell density was higherthan 105 per litre.
Figure 2. Seasonal biomass (lg l)1) dynamics of Scrippsiella/
Woloszinskia complex along the transect between Tallinn and
Helsinki. Averaged weekly values from 1998–2003.
Figure 3. Examples of the seasonal abundance dynamics (cells l)1) of Scrippsiella/Woloszynskia during April–May 2002 at selected
stations. Note the difference in scales.
142
Due to the scarcity of intensive monitoring data,the variation in cell maxima and the duration ofmedium-sized dinoflagellate blooms on interannualbasis was only possible to evaluate in the samplesfrom the central Gulf of Finland and for Swedishstations (B1 and H3). The bloom peaked almostsimultaneously in both locations, but in generalbetween 10th and 30th April. The peak values werefound during a relatively short period (1–2 weeks).Only after the severe winter in 2003, the bloompersisted from the mid of April to the mid of May,i.e. almost a month. A second maximum of Scrip-psiella/Woloszynskia was, however, found at sta-tionsWQ5 andWQ3near the Finnish south coast inMay 1999 and 2002, respectively (Figs. 2 and 3).The water temperature measured during the inten-sive growth of Scrippsiella/Woloszynskia wasmainly below 3 �C, indicating unstable conditionswhen the stratification was not yet developed.During the second maximum and also at theLithuanian stations, the temperature was >6 �C.
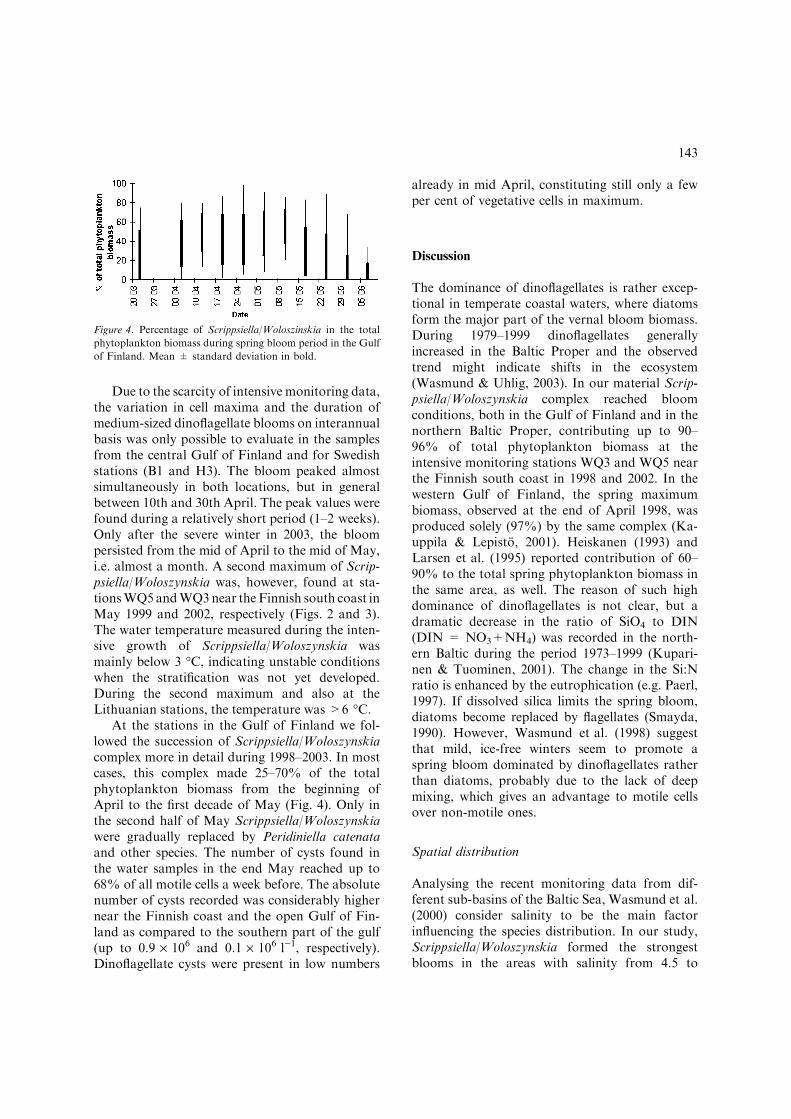
At the stations in the Gulf of Finland we fol-lowed the succession of Scrippsiella/Woloszynskiacomplex more in detail during 1998–2003. In mostcases, this complex made 25–70% of the totalphytoplankton biomass from the beginning ofApril to the first decade of May (Fig. 4). Only inthe second half of May Scrippsiella/Woloszynskiawere gradually replaced by Peridiniella catenataand other species. The number of cysts found inthe water samples in the end May reached up to68% of all motile cells a week before. The absolutenumber of cysts recorded was considerably highernear the Finnish coast and the open Gulf of Fin-land as compared to the southern part of the gulf(up to 0.9 · 106 and 0.1 · 106 l)1, respectively).Dinoflagellate cysts were present in low numbers
already in mid April, constituting still only a fewper cent of vegetative cells in maximum.
Discussion
The dominance of dinoflagellates is rather excep-tional in temperate coastal waters, where diatomsform the major part of the vernal bloom biomass.During 1979–1999 dinoflagellates generallyincreased in the Baltic Proper and the observedtrend might indicate shifts in the ecosystem(Wasmund & Uhlig, 2003). In our material Scrip-psiella/Woloszynskia complex reached bloomconditions, both in the Gulf of Finland and in thenorthern Baltic Proper, contributing up to 90–96% of total phytoplankton biomass at theintensive monitoring stations WQ3 and WQ5 nearthe Finnish south coast in 1998 and 2002. In thewestern Gulf of Finland, the spring maximumbiomass, observed at the end of April 1998, wasproduced solely (97%) by the same complex (Ka-uppila & Lepisto, 2001). Heiskanen (1993) andLarsen et al. (1995) reported contribution of 60–90% to the total spring phytoplankton biomass inthe same area, as well. The reason of such highdominance of dinoflagellates is not clear, but adramatic decrease in the ratio of SiO4 to DIN(DIN = NO3+NH4) was recorded in the north-ern Baltic during the period 1973–1999 (Kupari-nen & Tuominen, 2001). The change in the Si:Nratio is enhanced by the eutrophication (e.g. Paerl,1997). If dissolved silica limits the spring bloom,diatoms become replaced by flagellates (Smayda,1990). However, Wasmund et al. (1998) suggestthat mild, ice-free winters seem to promote aspring bloom dominated by dinoflagellates ratherthan diatoms, probably due to the lack of deepmixing, which gives an advantage to motile cellsover non-motile ones.
Spatial distribution
Analysing the recent monitoring data from dif-ferent sub-basins of the Baltic Sea, Wasmund et al.(2000) consider salinity to be the main factorinfluencing the species distribution. In our study,Scrippsiella/Woloszynskia formed the strongestblooms in the areas with salinity from 4.5 to
Figure 4. Percentage of Scrippsiella/Woloszinskia in the total
phytoplankton biomass during spring bloom period in the Gulf
of Finland. Mean ± standard deviation in bold.
143

6.5 PSU. However, it concerns only the Gulf ofFinland and the northern Baltic Proper and eventhere the variation was extremely large in a rela-tively short distance (Fig. 2). High abundance(1–3 · 106 l)1) was also obtained in some deeperareas in the Gulf of Finland and the northernBaltic Proper. Kremp (2000a) suggested that theannual blooms of medium-sized dinoflagellates inthe northern Baltic Sea are seeded by large coastalresting cyst populations and the offshore bloomslikely originate from those coastal inocula trans-ported to the central parts of the Baltic Sea byoutflowing surface water. On the other hand, inthe deeper basins of the Baltic Proper and the Gulfof Finland anoxia, particularly if coupled to thepresence of H2S, may prevent germination ofdinoflagellate cysts from sediments (Kremp &Anderson, 2000).
As the station grid for regular monitoring inthe Baltic Sea is rather uneven, the real distribu-tion pattern may differ from that derived from ourstudy. We probably miss information on severalpotential seeding/bloom areas, especially in theBaltic Proper and the Gulf of Bothnia, but also onsome coastal sites in the northern Gulf of Finland.Scrippsiella and/or Woloszynskia cysts have beenfound in the low-saline coastal areas (Kremp,2000a), however, the maximum number of livingcells at the stations with average salinity<4.5 PSU remained below 105 l)1 in our data set.Due to the lack of recent phytoplankton data fromthe SE Finnish coastal area and the Bothnian Bay,we have rather poor knowledge on the lowestsalinity tolerance of Scrippsiella/Woloszynskia.The species was sporadically found in the Stock-holm archipelago (station SS1, average salinity1.34 PSU), but not observed or recognized inNeva Estuary, the easternmost part of the Gulf ofFinland (salinity 0.3–1.7 PSU) (Nikulina, 2003).One or another species of the complex has beenpresent in the plankton of Mecklenburg Bight withsalinity 8–9 PSU (Alg@line, 2001). Scrippsiella/Woloszynskia have probably not spread outsidethe Baltic Sea, however, dinoflagellate cysts maysurvive transport with ballast water and can bemoved to areas where they have not recordedpreviously. ‘S. hangoei’ cysts have been found inthe sediment samples from English and Welshports (McCollin et al., 1999), but without anyrecord of living cells.
Although the maximum cell number decreasedboth towards the higher and lower salinity areas,salinity is probably not the main factor governingthe distribution of these species as they were notdominating in the Gulf of Riga with the samesalinity range. It may also be possible that thedinoflagellates found in the southern and northernparts of the Baltic Sea are not con-specific.Moreover, Kremp et al. (2003) found thatWoloszynskia sp. thrives in salinities that are muchhigher than what the species experience in thenorthern Baltic Proper, whereas salinities lowerthan 6 PSU seemed to inhibit growth.
Analysing the distribution of dinoflagellatecysts along an estuarine salinity gradient(2–7 PSU), Kremp (2000a) also took into consid-eration the character of bottom sediments.Although no correlation was found between cystconcentration and water depth of the samplingsites, almost all sampling locations with high cystabundances were muddy with a high percentage oforganic content. The Finnish coastline mainlyconsists of bedrock, but the sediments in the outerarchipelago and open sea zone are mostly mud-covered. This is also characteristic for the stationsWQ3 and WQ5 near the Finnish south coast wherethe observed number of living cells of Scrippsiella/Woloszynskia was the highest – up to 107 l)1 (seealso Fig. 2).
The Gulf of Riga has no permanent haloclineand it is mixed down to the bottom in winter. Thecharacter of bottom sediments is variable althoughorganic-rich soft bottom are prevailing only in themiddle deepest (40–55 m) part of the gulf (Stiebrins& Valing, 1996). Theoretically, a seed populationcan be permanently available. Monitoring obser-vations made in the Gulf of Riga and in theadjacent areas to the central Baltic Proper inMarch–April still indicate the clear dominance ofdiatoms, mainly Thalassiosira baltica and Pauliellataeniata. The percentage of dinoflagellates hasincreased in the spring blooms at the end of 1980sand the beginning of the 1990s, but it is mainly dueto the increase in Peridiniella catenata biomass(Yurkovskis et al., 1999).
Temporal distribution
A maximum cell density of Scrippsiella/Wol-oszynskia in the water column was mainly obtained
144

in March–April or in the beginning of May by thewater temperature <3 �C, under unstable condi-tions prior to stratification was formed. Krempet al. (2003) found Woloszynskia sp. to be a trulystenothermic organism, with a temperature windowbetween 0 and 6 �C. As S. hangoei has optimumtemperatures a few centigrade higher, they sug-gested that temperature may be a potential factorseparating niches of the two species. Here is acontradiction with our data, as the salinity opti-mum for Woloszynskia sp. is in general higher(>6 PSU) than in the areas of strongest ‘red tide’dinoflagellate blooms in the Baltic Sea. The bloomsprobably appear to be a mixture of several specieswith unknown proportions, involving both, Scrip-psiella and Woloszynskia.
According to Kremp et al. (2003) also theresting cysts, which had previously assigned toS. hangoei were formed in clonal cultures of theWoloszynskia isolates and the species also differ intheir mode of sexual reproduction. In the experi-ment, dinoflagellate (probably Woloszynskia)cysts, formed during the spring bloom terminationin the northern Baltic Sea, remained dormant for6 months (Kremp, 2000a). Thus a new populationcan be seeded even under unfavourable conditionsin early winter when the light conditions are poor.On the other hand, such an early inoculum allowsthe species to immediately exploit favourablegrowth conditions provided. In our material,Scrippsiella/Woloszynskia made up to 75% of thetotal phytoplankton biomass already in March(Fig. 4), and these medium-sized dinoflagellateswere often the first dominants in spring bloomdevelopment. In the Gulf of Finland, these speciespersisted dominant until mid of May. We do notstill understand, in which proportion each speciescontribute to the total dinoflagellate biomass.There are also some opinions that environmentalchanges can influence the further bloom initiationand alter the respective proportions of the specieslater during the spring bloom (Kremp & Heiska-nen, 1999; Kremp, 2000b). Woloszynskia sp. mayhave a benefit in the earlier stage of the bloomaccording to its temperature optimum, whereasScrippsiella hangoei can be the major componentbefore the bloom terminates. The second maximarecorded in May 1999 and 2002, developed by thesurface water temperature >6 �C, thus can beattributed to S. hangoei.
Mild winters without ice cover seem promotethe dominance of dinoflagellates in the springbloom of the Baltic Sea (Heiskanen, 1998; Was-mund et al., 1998; Hajdu, 2002). Extremelystrong blooms of Scrippsiella/Woloszynskia in thecentral Gulf of Finland were observed in 1998and 1999, but especially after mild winters in2000 and 2002. We cannot still agree that med-ium-sized single-celled dinoflagellates are theminor component of the vernal phytoplanktoncommunity after harsh winters (Kremp &Heiskanen, 1999). In 2003, these species made upto 92–94% of the dinoflagellate and 70–88% ofthe total phytoplankton biomass in the same areaduring the spring bloom peak. It allows us toconclude that Scrippsiella/Woloszynskia bloom israther a yearly phenomenon at least in the Gulfof Finland. The observed dominance of thiscomplex in the gulf might be a sign for ecosystemchange due to decreasing inorganic Si:N ratio or/and climatic changes (e.g. Paerl, 1997).
The available monitoring data did not clarifythe ecology of both species mainly due to theidentification problems. These data may still servea basis for the further investigations, including theeffect of eutrophication. On the other hand, any ofsuch basin-wide research needs also a betterintercalibration of methods and carefully plannedsampling strategy of phytoplankton.
Acknowledgements
This work was partly financed by the EstonianMinistry of Environment, the Finnish Ministry ofEnvironment and the EU project FerryBox(EVK2-CT-2002–00144).
References
Alg@line, 2001. Baltic Sea Phytoplankton Sheet. http://jolly.
fimr.fi/Checklist3.nsf/Main?OpenFrameSet.
Hajdu, S., 2002. Phytoplankton of Baltic Environmental Gra-
dients: Observations on potentially Toxic Species. PhD
thesis, Stockholm University, Sweden, 42 pp.
Heiskanen, A.-S., 1993. Mass encystment and sinking of dino-
flagellates during a spring bloom.Marine Biology 116: 161–168.
Heiskanen, A.-S., 1998. Factors Governing Sedimentation and
Pelagic Nutrient Cycles in the Northern Baltic Sea. Mono-
graphs of the Boreal Environment Research 8, 80 pp.
145

HELCOM, 1988. Guidelines for the Baltic Monitoring Pro-
gramme for the third stage; Part D. Biological Determin-
ands. Baltic Sea Environment Proceedings 27D.
HELCOM, 1990. Second periodic assessment of the state of the
marine environment of the Baltic Sea, 1984–1988; Back-
ground document. Baltic Sea Environment Proceedings 35
B: 1–432.
HELCOM, 1996. Third periodic assessment of the state of the
marine environment of the Baltic Sea, 1989–1993; Background
document. Baltic Sea Environment Proceedings 64 B: 1–252.
Hobro, R., 1979. Annual phytoplankton successions in a
coastal area in the northern Baltic. In Naylor, E. & R. G.
Hartnoll (eds.) Cyclic Phenomena in Marine Plants and
Animals. Pergamon, Oxford: 3–10.
Hallfors, G. & A. Niemi, 1981. Vegetation and primary pro-
duction. In Voipio, A. (ed.) The Baltic Sea. Elsevier,
Amsterdam: 220–237.
Hoglander, H., U. Larsson & S. Hajdu, 2004. Vertical distribu-
tion and settling of spring phytoplankton in the offshore NW
Baltic Sea proper.Marine Ecology Progress Series 283: 15–27.
Kauppila, P. & L. Lepisto, 2001. Changes in phytoplankton. In
Kauppila, P. & S. Back (eds.) The State of Finnish Coastal
Waters in the 1990s. Finnish Environment Institute, Hel-
sinki: 61–70.
Kremp, A., 2000a. Distribution, dynamics and in situ seeding
potential of Scrippsiella hangoei (Dinophyceae) cyst popu-
lations from the Baltic Sea. Journal of Plankton Research
22: 2155–2169.
Kremp, A., 2000b. The role of life cycle in the population
dynamics of the bloom forming dinoflagellates Scrippsiella
hangoei and Peridiniella catenata in the Baltic Sea. Walter
and Andree de Nottbeck Foundation Scientific Reports 22.
Kremp, A. & D. M. Anderson, 2000. Factors regulating ger-
mination of resting cysts of the spring bloom dinoflagellate
Scrippsiella hangoei from the northern Baltic Sea. Journal of
Plankton Research 22: 1311–1327.
Kremp, A. & A.-S. Heiskanen, 1999. Sexuality and cyst for-
mation of the spring-bloom dinoflagellate Scrippsiella han-
goei in the coastal northern Baltic Sea. Marine Biology 134:
771–777.
Kremp, A., M. Elbrachter, J. Wolny & A. Shurtleff, 2003. A
new cyst forming spring bloom dinoflagellate co-occuring
with Scrippsiella hangoei in the northern Baltic Sea. Pro-
ceedings of the 7th International Conference of Modern and
Fossil Dinoflagellates. September 21–25, Nagasaki, Japan,
p. 66 (poster abstract).
Kuparinen, J. & L. Tuominen, 2001. Eutrophication and Self-
purification: counteractions forced by large -scale cycles and
hydrodynamic processes. Ambio 30: 190–194.
Larsen, J., H. Kuosa, J. Ikavalko, K. Kivi & S. Hallfors, 1995.
A redescription of Scrippsiella hangoei (Schiller) comb.
nov. – a ‘red tide’ dinoflagellate from the northern Baltic.
Phycologia 34(2): 135–144.
Lignell, R., A.-S. Heiskanen, H. Kuosa, K. Gundersen, P.
Kuuppo-Leinikki, R. Pajuniemi & A. Uitto, 1993. Fate of a
phytoplankton spring bloom: sedimentation and carbon flow
in the planktonic food web in the northern Baltic. Marine
Ecology Progress Series 94: 239–252.
McCollin, T. A., J. P. Hamer & I. A. N. Lucas, 1999. Transport
of Marine Organisms via Ships’ Ballast into Ports around
England and Wales. National Conference on Marine Bio-
invasions. Boston, Massachusetts, 1999.
Niemi, A., 1975. Ecology of phytoplankton in the Tvarminne
area SW coast of Finland. II. Primary production and
environmental conditions in the archipelago and the sea
zone. Acta Botanica Fennica 105: 1–73.
Nikulina, V., 2003. Seasonal dynamics of phytoplankton in the
inner Neva Estuary in the 1980s and 1990s. Oceanologia
45(1): 25–39.
Paerl, H. W., 1997. Coastal eutrophication and harmful algal
blooms: importance of atmospheric deposition and
groundwater as ‘‘new’’ nitrogen and other nutrient sources.
Limnology and Oceanography 42: 1154–1165.
Rantajarvi, E. & J.-M. Leppanen, 1994. Unattended Algal
Monitoring in Merchant Ships in the Baltic Sea. Tema Nord
546, 60 pp.
Smayda, T. J., 1990. Novel and nuisance phytoplankton
blooms in the sea: evidence for a global epidemic. In Graneli,
E., B. Sundstrom, L. Edler & D. M. Anderson (eds), Toxic
Marine Phytoplankton. Proc. Fourth. Int. Conf. on Toxic
Marine Phytoplankton. Elsevier, New York: 29–40.
Stiebrins, O. & P. Valing, 1996. Bottom sediments of the Gulf
of Riga. Geological Survey of Latvia. Geological Survey of
Estonia. Riga, 53 pp. and map.
Wasmund, N. & S. Uhlig, 2003. Phytoplankton trends in
the Baltic Sea. ICES Journal of Marine Science 60: 177–
186.
Wasmund, N., G. Nausch & W. Matthaus, 1998. Phytoplank-
ton spring bloom in the southern Baltic Sea – spatio-tem-
poral development and long-term trends. Journal of
Plankton Research 20: 1099–1117.
Wasmund, N., G. Nausch, L. Postel, Z. Witek, M. Zalewski,
S. Gromisz, E. ysiak-Pastuszak, I. Olenina, R. Kavolyte,
A. Jasinskaite, B. Muller-Karulis, A. Ikauniece, A. Andru-
shaitis, H. Ojaveer, K. Kallaste & A. Jaanus, 2000. Trophic
status of coastal and open areas of the south-eastern Baltic
Sea based on nutrient and phytoplankton data from 1993–
1997. Marine Science Reports 38, 83 pp.
Yurkovskis, A., E. Kostrichkina & A. Ikauniece, 1999. Sea-
sonal succession and growth in the plankton communities of
the Gulf of Riga in relation to long-term nutrient dynamics.
Hydrobiologia 393: 83–94.
146
Copyright © 2022 FDOKUMEN




















