
Research review paper Biotechnological significance of toxic marine dinoflagellates
Upload
environcorpCategory
view
1download
0
ORIGINAL PAPER
Paul G. Thomson Æ Andrew McMinn Æ Ilse Kiessling
Mandy Watson Æ Paul M. Goldsworthy
Composition and succession of dinoflagellates and chrysophytesin the upper fast ice of Davis Station, East Antarctica
Received: 7 June 2005 / Revised: 9 August 2005 / Accepted: 9 August 2005 / Published online: 5 October 2005� Springer-Verlag 2005
Abstract Little is known of the wider Antarctic distri-bution of the upper fast ice community now compre-hensively described from McMurdo Sound. Wedetermined the fast ice protist community at DavisStation, East Antarctica and compared it with that ofMcMurdo Sound. As at McMurdo Sound, Davis fast iceis characterised by extreme and transitory salinities (96–2.5 psu) and temperatures (�4.5 to �0.1�C) during thespring/summer transition. Both communities are domi-nated by Polarella glacialis (an autotrophic dinoflagel-late), chrysophytes and their life cycle stages.Furthermore, the physical parameters of brine temper-ature and salinity at which these successions occurredapproximated those of McMurdo Sound. The high de-gree of similarity between the communities from thegeographically disparate locations indicates that thiscommunity type has a circum-Antarctic distribution.Confirming the areal extent and seasonality of thiscommunity type will assist in future predictions of seaice productivity.
Introduction
Upper ice protist communities are known from packand fast ice in the Antarctic (McConville and Weth-erbee 1983; Garrison and Buck 1989; 1991; Archeret al. 1996) and the Artic, where they can contribute upto 40% of the total ice biomass (Gradinger 1999). InMcMurdo Sound, these communities are well knownfrom a succession of studies describing the fast iceenvironment and its community composition (Stoeckeret al. 1992, 1993, 1997, 1998). These studies found thatthe upper fast ice is characterised by extreme andtransitory temperatures and salinities, ranging from�13.8�C and 200 practical salinity units (psu) in earlyspring to �0.2�C and 20 psu by late summer. Theprotist community is dominated by the photosyntheticdinoflagellate Polarella glacialis (Montresor et al.1999), its hypnozygotes (or resting cysts), chrysophytesand their statocysts. Stoecker et al. (1997, 1998) dem-onstrated the excystment and growth of these twodominant groups and proposed life cycles for each,concluding that their resting stages are adapted fordispersal during ice decay and overwintering after beingincorporated back into the reforming ice. Montresoret al. (2003) identified P. glacialis from Arctic ice, thusidentifying its bi-polar distribution. However, the lifecycle successions of P. glacialis have not been reportedfrom Arctic ice.
While the community composition of McMurdoSound fast ice has been well described, the extent anddistribution of this community type in other Antarcticlocations is unknown. P. glacialis has recently beenidentified from fast ice adjacent to Davis Station andappears to have a circum-Antarctic distribution(Thomson et al. 2004). Chrysophyte statocysts havebeen reported from ice, water and sediment samples ofthe Ross and Weddell Seas (Mitchell and Silver 1982)and from fast ice near Syowa Station (69�00¢22¢¢S,39�35¢24¢¢E, Takahashi et al. 1986). Thus, it seems likely
P. G. Thomson (&) Æ M. Watson Æ P. M. GoldsworthyDepartment of the Environment and Heritage,Australian Antarctic Division and the Antarctic Climateand Ecosystems CRC, Channel Highway, Kingston,TAS 7050, AustraliaE-mail: [email protected].: +61-36232-3562
A. McMinnInstitute of Antarctic and Southern Ocean Studiesand the Antarctic Climate and Ecosystems CRC,University of Tasmania, PO Box 252-77,Hobart, TAS 7001, Australia
I. KiesslingSchool of Environmental Research,C- North Australian Research Unit, Charles Darwin University,PO Box 41321, Casuarina,NT 0811, Australia
Polar Biol (2006) 29: 337–345DOI 10.1007/s00300-005-0060-y

that upper fast ice communities, similar to that ofMcMurdo Sound, are widespread in Antarctica.
Confirming a circum-Antarctic distribution of thefast ice community will assist in more accurate estimatesof sea ice productivity, currently thought to comprisearound 20% of the annual primary productivity of theSouthern Ocean (Legendre et al. 1992). This estimate isbased on the partitioning of the sea ice algae into sur-face, freeboard, interior and bottom ice communitiesand calculated using standing stocks, areal extent andtime of production. However, the scarcity of data foreach of these communities is conceded as a limitation ofthe estimate (Legendre et al. 1992). Identifying the ex-tent and standing stocks of a previously unrecognisedcommunity in this estimate will aid future estimates offast ice productivity.
Here we describe the physical nature of the brinechannel environment and its community compositionand succession in Davis Station fast ice. We compare theDavis fast ice environment, its community and com-munity succession with that of McMurdo Sound andexpand on the Antarctic distribution of this communitytype.
Materials and methods
Fieldwork was carried out during the spring summertransitions of 1996 and 1997 in fast ice near Davis Sta-tion in the Vestfold Hills, East Antarctica (68�43.2¢S77�56.4¢E). In 1996, three sampling sites were chosenover an approximate area of 4 km2 and in 1997, five siteswere sampled over approximately 16 km2. Water depthbeneath the sites ranged between 15 and 25 m. Sites werelost sequentially through both seasons as sections of thefast ice broke out along the coast, leaving only one site(O’Gorman Rocks, 1 km offshore from Davis Station)accessible by late December in both years.
Triplicate ice cores were obtained on a weekly basiswhen possible at O’Gorman Rocks to determine thevertical distribution of chl a in the ice. Cores were takenusing a 100 mm SIPRE corer, sectioned immediatelyinto 10-cm sections and placed into individual contain-ers. The sections were then melted in the dark at 4�C inan equal volume of 0.45 lm GF/F filtered seawater. Theentire volume of each section was then filtered through0.45 lm GF/F filters and chl a was extracted in 90%acetone overnight. Chl a concentrations were obtainedusing a GBC 916 UV visible spectrophotometer whereabsorbency was measured at 750, 664, 647 and 630 nm(Parsons et al. 1984). Concentrations were integratedinto three ice horizons of 0–50 cm, 50–100 cm andbottom ice (from 100 cm to the depth of the core).
Fast ice brine was collected every 3 days whenpossible. Snow cover at each site was recorded andtriplicate, 50-cm-deep holes were drilled within a 1 mradius by a 30 cm diameter Jiffy drill. The drill holeswere allowed to fill with brine from the surroundingice. Early in the season the brine slowly accumulated
overnight and was collected the following day but asthe season progressed, collection was possible immedi-ately. Brine temperature and salinity were measuredimmediately using a Hanna Checktemp C digital ther-mometer and a Shibuya (Type 147) refractometer. Thebrine was thoroughly mixed and 1 l samples weretransferred to a glass bottle and fixed with acidifiedLugol’s Iodine. A further 1 l of fresh brine was takenfor microscopic identification and isolation of individ-ual cells for culture.
Opportunistic brine samples were collected from fastand pack ice locations to investigate the extent of brinecommunities in Antarctic ice (Fig. 1). Fast ice sampleswere collected from Horseshoe Harbour, Mawson Sta-tion (67�36¢S 62�53¢E) in December 1996 and fromBrowns Bay, Casey Station (66�17¢S 110�32¢E) andNella Fjord, Larsemann Hills (69�23¢S 76�23¢E) inNovember 1997. Four pack ice sites in the East Ant-arctic sector of the Southern Ocean were sampled inOctober 1996 (64�51.79¢S 109�37.32¢E; 63�04.41¢S92�47.53¢E; 62�56.87¢S 92�58.00¢E; 63�46.76¢S74�10.49¢E). All samples were fixed with acidified Lu-gol’s Iodine and examined (as described below) specifi-cally for P. glacialis and chrysophyte motile cells andcysts.
Protist species were identified using both light andelectron microscopy. Subsamples of up to 1 l of freshbrine were filter concentrated (Activon 0.8 lm celluloseacetate membrane filters). Cells were gently resuspendedand a drop of the concentrate was examined using aZeiss Axioscope microscope at 400 and 1,000· magni-fication. Cells were identified as photosynthetic by bluelight epifluorescence microscopy (filter set 487909 with450–490 nm exciter filter, 510 nm chromatic beamsplitter and 520 nm barrier filter). The remaining con-centrate was fixed with glutaraldehyde (2.5% final con-centration).
Species identification was aided by SEM of the glu-taraldehyde and Lugol’s fixed samples, using a PhillipsElectroscan Environmental SEM with EnvironmentalSEM System 2020 Version 3.2 software. Protists werecounted after the Lugol’s fixed brine samples wereconcentrated by sedimentation and settled into 10-mlsettling chambers. Counts were carried out on a ZiessAxiovert inverted microscope under 400 and 1,000·magnification using Nomarski optics. Fifteen replicatefields of view (FOV) at each magnification were countedand the mean cell concentration and standard deviationcalculated. Carbon biomass was calculated for P. gla-cialis and chrysophytes following estimated cell volumesand carbon content used by Stoecker et al. (1998).
Water column samples were obtained near O’Gor-man Rocks during 1996 season. Sub-ice samples weretaken weekly when possible at 0, 5, 10, 15 and 20 mdepths. One-litre samples were fixed with Lugol’s Iodineand triplicate counts of P. glacialis cysts were made aftersettling (as previously described). After fast ice breakout, approximately weekly water samples were taken byboat at the same location.
338

Results
Fast ice thickness at O’Gorman Rocks in both yearsranged between 1.5 m in November and 1.3 m inDecember. Chl a concentrations in the upper 50 cm ofice reached 13.78 mg m�3 in 1996 and 7.17 mg m�3 in1997 (Table 1). Where a bottom ice community waspresent, the upper 50-cm contributed up to 23.3% of thetotal chl a in 1996 and 5% in 1997. In the absence of abottom ice community, the upper ice contributed up to42.3% of the total chl a. The upper 50 cm horizongenerally contained more chl a than the 50–100 cmhorizon over the sampling period in both years.
Brine samples were taken from early November andsampling ceased by late December in both years when
the sea ice broke out or sea ice travel became unsafe.Snow cover on the fast ice in 1996 was light (maximumof 5 cm) and the sites remained largely snow freethroughout the sampling period. In 1997, snow coverwas commonly 20 cm in depth at each site until lateNovember.
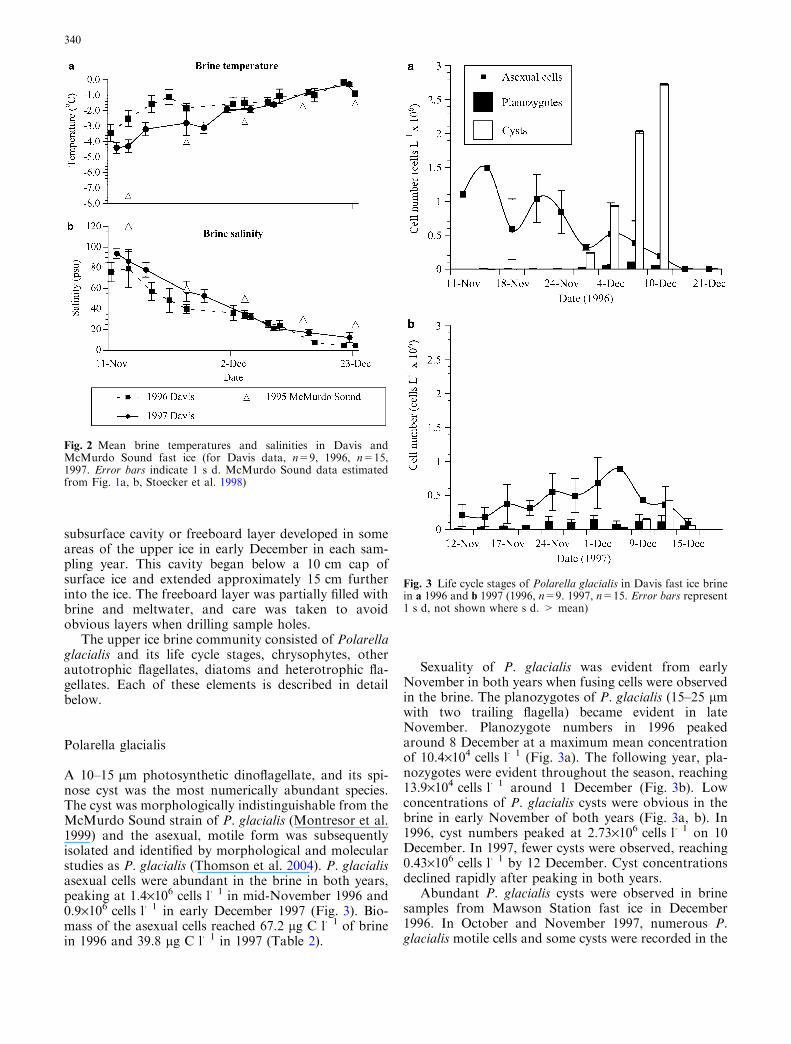
In 1996, mean brine temperatures increased from�3.4�C through to �0.1�C by mid-December (Fig. 2a).Rapid warming by almost 2.5�C occurred between 11and 18 November, followed by a more gradual increaseto �0.1�C by the end of sampling. In 1997, brine tem-peratures increased at a more gradual rate and were atleast 1�C lower than the previous year until earlyDecember. In both years, the brine salinity in earlyNovember ranged between 80 and 94 psu but graduallydeclined to 5 and 10 psu by late December (Fig. 2b). A
Table 1 Mean chlorophyll a concentrations (±1 s d.) from fast ice horizons of cores taken at O’Gorman Rocks, Davis Station in 1996and 1997 (n=3, percentage values of total chl a for horizons in parenthesis, ND not detected)
Depth in ice(cm)
1996 10Nov
17 NovChl a
24 Nov(mg m�3)
2 Dec 1997 7Nov
14 Nov Chl a 21 Nov(mg m�3)
14 Dec
0–50 4.82±1.05(23.3%)
4.99±0.95(6.0%)
8.79±2.22(8.6%)
13.78±3.37(7.4%)
0.40±0.18(7.9%)
5.64±0.65(5.0%)
7.17±1.08(4.8%)
1.75±1.07(42.3%)
50–100 3.01±0.22(14.5%)
2.56±0.63(3.1%)
5.34±0.62(5.2%)
9.13±1.87(4.9%)
4.65±0.52(92.1%)
1.73±0.03(1.6%)
4.16±0.43(2.8%)
2.38±0.87(57.6%)
Bottom ice 12.89±1.09(62.2%)
75.15±5.86(90.9%)
88.04±6.81(86.2%)
163.87±12.56(87.7%)
ND 103.62±20.35(93.4%)
138.88±17.36(92.5)
ND
Total 20.72±2.36 82.7±7.49 102.17±9.65 186.78±17.80 5.05±0.7 110.99±21.73 150.21±18.87 4.13±1.94
Fig. 1 The circumpolardistribution of Polarellaglacialis in Antarctic fast andpack ice [open circle not present;filled circle recorded from thisstudy; filled square recordedfrom other studies at McMurdoSound (Stoecker et al. 1998;Montresor et al. 1999) and theWeddell and Scotia Seas(approximate locations, Bucket al. 1992 and Garrison andBuck 1989, respectively)]
339
subsurface cavity or freeboard layer developed in someareas of the upper ice in early December in each sam-pling year. This cavity began below a 10 cm cap ofsurface ice and extended approximately 15 cm furtherinto the ice. The freeboard layer was partially filled withbrine and meltwater, and care was taken to avoidobvious layers when drilling sample holes.
The upper ice brine community consisted of Polarellaglacialis and its life cycle stages, chrysophytes, otherautotrophic flagellates, diatoms and heterotrophic fla-gellates. Each of these elements is described in detailbelow.
Polarella glacialis
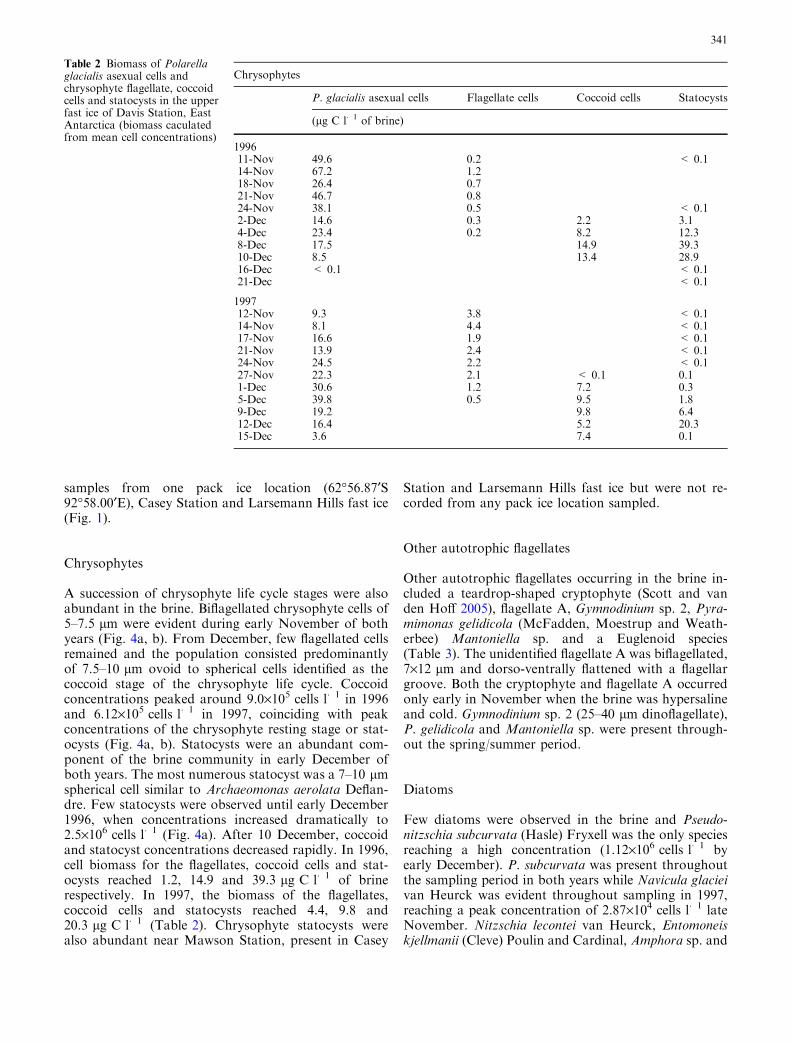
A 10–15 lm photosynthetic dinoflagellate, and its spi-nose cyst was the most numerically abundant species.The cyst was morphologically indistinguishable from theMcMurdo Sound strain of P. glacialis (Montresor et al.1999) and the asexual, motile form was subsequentlyisolated and identified by morphological and molecularstudies as P. glacialis (Thomson et al. 2004). P. glacialisasexual cells were abundant in the brine in both years,peaking at 1.4·106 cells l�1 in mid-November 1996 and0.9·106 cells l�1 in early December 1997 (Fig. 3). Bio-mass of the asexual cells reached 67.2 lg C l�1 of brinein 1996 and 39.8 lg C l�1 in 1997 (Table 2).
Sexuality of P. glacialis was evident from earlyNovember in both years when fusing cells were observedin the brine. The planozygotes of P. glacialis (15–25 lmwith two trailing flagella) became evident in lateNovember. Planozygote numbers in 1996 peakedaround 8 December at a maximum mean concentrationof 10.4·104 cells l�1 (Fig. 3a). The following year, pla-nozygotes were evident throughout the season, reaching13.9·104 cells l�1 around 1 December (Fig. 3b). Lowconcentrations of P. glacialis cysts were obvious in thebrine in early November of both years (Fig. 3a, b). In1996, cyst numbers peaked at 2.73·106 cells l�1 on 10December. In 1997, fewer cysts were observed, reaching0.43·106 cells l�1 by 12 December. Cyst concentrationsdeclined rapidly after peaking in both years.
Abundant P. glacialis cysts were observed in brinesamples from Mawson Station fast ice in December1996. In October and November 1997, numerous P.glacialis motile cells and some cysts were recorded in the
Fig. 2 Mean brine temperatures and salinities in Davis andMcMurdo Sound fast ice (for Davis data, n=9, 1996, n=15,1997. Error bars indicate 1 s d. McMurdo Sound data estimatedfrom Fig. 1a, b, Stoecker et al. 1998)
Fig. 3 Life cycle stages of Polarella glacialis in Davis fast ice brinein a 1996 and b 1997 (1996, n=9. 1997, n=15. Error bars represent1 s d, not shown where s d. > mean)
340
samples from one pack ice location (62�56.87¢S92�58.00¢E), Casey Station and Larsemann Hills fast ice(Fig. 1).
Chrysophytes
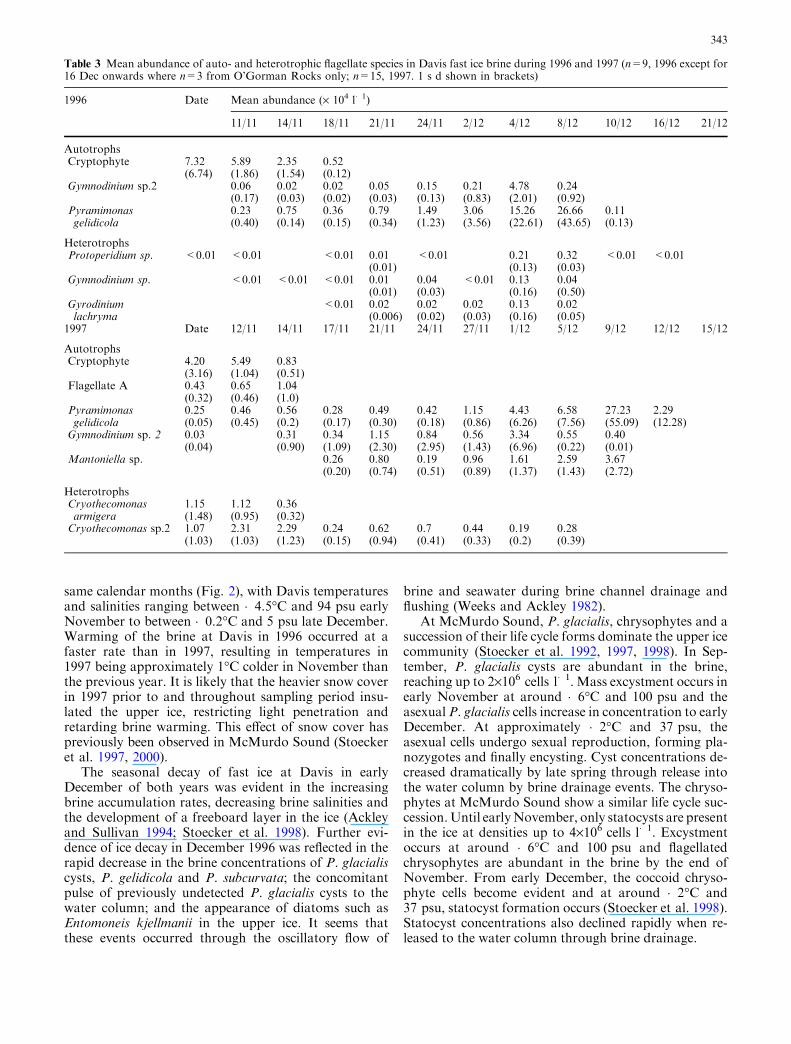
A succession of chrysophyte life cycle stages were alsoabundant in the brine. Biflagellated chrysophyte cells of5–7.5 lm were evident during early November of bothyears (Fig. 4a, b). From December, few flagellated cellsremained and the population consisted predominantlyof 7.5–10 lm ovoid to spherical cells identified as thecoccoid stage of the chrysophyte life cycle. Coccoidconcentrations peaked around 9.0·105 cells l�1 in 1996and 6.12·105 cells l�1 in 1997, coinciding with peakconcentrations of the chrysophyte resting stage or stat-ocysts (Fig. 4a, b). Statocysts were an abundant com-ponent of the brine community in early December ofboth years. The most numerous statocyst was a 7–10 lmspherical cell similar to Archaeomonas aerolata Deflan-dre. Few statocysts were observed until early December1996, when concentrations increased dramatically to2.5·106 cells l�1 (Fig. 4a). After 10 December, coccoidand statocyst concentrations decreased rapidly. In 1996,cell biomass for the flagellates, coccoid cells and stat-ocysts reached 1.2, 14.9 and 39.3 lg C l�1 of brinerespectively. In 1997, the biomass of the flagellates,coccoid cells and statocysts reached 4.4, 9.8 and20.3 lg C l�1 (Table 2). Chrysophyte statocysts werealso abundant near Mawson Station, present in Casey
Station and Larsemann Hills fast ice but were not re-corded from any pack ice location sampled.
Other autotrophic flagellates
Other autotrophic flagellates occurring in the brine in-cluded a teardrop-shaped cryptophyte (Scott and vanden Hoff 2005), flagellate A, Gymnodinium sp. 2, Pyra-mimonas gelidicola (McFadden, Moestrup and Weath-erbee) Mantoniella sp. and a Euglenoid species(Table 3). The unidentified flagellate A was biflagellated,7·12 lm and dorso-ventrally flattened with a flagellargroove. Both the cryptophyte and flagellate A occurredonly early in November when the brine was hypersalineand cold. Gymnodinium sp. 2 (25–40 lm dinoflagellate),P. gelidicola and Mantoniella sp. were present through-out the spring/summer period.
Diatoms
Few diatoms were observed in the brine and Pseudo-nitzschia subcurvata (Hasle) Fryxell was the only speciesreaching a high concentration (1.12·106 cells l�1 byearly December). P. subcurvata was present throughoutthe sampling period in both years while Navicula glacieivan Heurck was evident throughout sampling in 1997,reaching a peak concentration of 2.87·104 cells l�1 lateNovember. Nitzschia lecontei van Heurck, Entomoneiskjellmanii (Cleve) Poulin and Cardinal, Amphora sp. and
Table 2 Biomass of Polarellaglacialis asexual cells andchrysophyte flagellate, coccoidcells and statocysts in the upperfast ice of Davis Station, EastAntarctica (biomass caculatedfrom mean cell concentrations)
Chrysophytes
P. glacialis asexual cells Flagellate cells Coccoid cells Statocysts
(lg C l�1 of brine)
199611-Nov 49.6 0.2 < 0.114-Nov 67.2 1.218-Nov 26.4 0.721-Nov 46.7 0.824-Nov 38.1 0.5 < 0.12-Dec 14.6 0.3 2.2 3.14-Dec 23.4 0.2 8.2 12.38-Dec 17.5 14.9 39.310-Dec 8.5 13.4 28.916-Dec < 0.1 < 0.121-Dec < 0.1
199712-Nov 9.3 3.8 < 0.114-Nov 8.1 4.4 < 0.117-Nov 16.6 1.9 < 0.121-Nov 13.9 2.4 < 0.124-Nov 24.5 2.2 < 0.127-Nov 22.3 2.1 < 0.1 0.11-Dec 30.6 1.2 7.2 0.35-Dec 39.8 0.5 9.5 1.89-Dec 19.2 9.8 6.412-Dec 16.4 5.2 20.315-Dec 3.6 7.4 0.1
341
Fragilariopsis kerguelensis (O’Meara) Hust. (Hustedt)were present at low concentrations (<1,000 cells l�1)from mid-December 1996.
Heterotrophs
In 1996, the dinoflagellates Protoperidinium sp. (55 lmin diameter), Gymnodinium sp. (17–20 lm in length) andGyrodinium lachryma (Meunier) Kofoid and Swezydominated the heterotrophic assemblage (Table 3). Fewdinoflagellates were recorded in 1997 when two speciesof Cryothecomonas were evident in the brine (Table 3).Cryothecomonas armigera Thomsen, Buck, Bolt andGarrison was present only under the hypersaline con-ditions early- to mid-November while Cryothecomonassp.2 (10 lm·20 lm), was persistent throughout thesampling period. Strombidium sp. was the only ciliaterecorded in the brine and was present throughout sam-pling in 1996 and was evident from early December in1997. This species reached up to 2.5·103 cells l�1 earlyDecember each year. A 30 lm Euglenoid species, similarto Euglenoid sp.2 observed by Archer et al. (1996) nearDavis Station, was present in both years. This species
occurred at low concentrations (<150 cells l�1) between17 November and 5 December 1996 and 1997.
Water samples
Polarella glacialis cyst concentrations were integratedover the five depths sampled (data not shown). Cystswere not detected in the water column until 8 December(<0.2·104 cysts l�1). On 16 December, just prior to icebreak out, a pulse of cysts of 1.4·104 l�1 was recorded.In the absence of ice cover after 16 December, cystnumbers ranged between 0.08 and 0.32·104 cysts l�1. G.lachryma was a notable component of the under icephytoplankton in November and December.
Discussion
The physical environment of the upper fast ice at DavisStation resembled that of McMurdo Sound in terms oftransitory temperatures and salinities and seasonal icedecay. In McMurdo Sound fast ice, brine temperaturesand salinities approximate those found at Davis over the
Fig. 4 Chrysophyte flagellate,coccoid cells and statocysts inDavis fast ice brine in a 1996and b 1997 (1996, n=9. 1997,n=15. Error bars represent 1 sd, not shown where s d >mean)
342
same calendar months (Fig. 2), with Davis temperaturesand salinities ranging between �4.5�C and 94 psu earlyNovember to between �0.2�C and 5 psu late December.Warming of the brine at Davis in 1996 occurred at afaster rate than in 1997, resulting in temperatures in1997 being approximately 1�C colder in November thanthe previous year. It is likely that the heavier snow coverin 1997 prior to and throughout sampling period insu-lated the upper ice, restricting light penetration andretarding brine warming. This effect of snow cover haspreviously been observed in McMurdo Sound (Stoeckeret al. 1997, 2000).
The seasonal decay of fast ice at Davis in earlyDecember of both years was evident in the increasingbrine accumulation rates, decreasing brine salinities andthe development of a freeboard layer in the ice (Ackleyand Sullivan 1994; Stoecker et al. 1998). Further evi-dence of ice decay in December 1996 was reflected in therapid decrease in the brine concentrations of P. glacialiscysts, P. gelidicola and P. subcurvata; the concomitantpulse of previously undetected P. glacialis cysts to thewater column; and the appearance of diatoms such asEntomoneis kjellmanii in the upper ice. It seems thatthese events occurred through the oscillatory flow of
brine and seawater during brine channel drainage andflushing (Weeks and Ackley 1982).
At McMurdo Sound, P. glacialis, chrysophytes and asuccession of their life cycle forms dominate the upper icecommunity (Stoecker et al. 1992, 1997, 1998). In Sep-tember, P. glacialis cysts are abundant in the brine,reaching up to 2·106 cells l�1. Mass excystment occurs inearly November at around �6�C and 100 psu and theasexual P. glacialis cells increase in concentration to earlyDecember. At approximately �2�C and 37 psu, theasexual cells undergo sexual reproduction, forming pla-nozygotes and finally encysting. Cyst concentrations de-creased dramatically by late spring through release intothe water column by brine drainage events. The chryso-phytes at McMurdo Sound show a similar life cycle suc-cession.Until earlyNovember, only statocysts are presentin the ice at densities up to 4·106 cells l�1. Excystmentoccurs at around �6�C and 100 psu and flagellatedchrysophytes are abundant in the brine by the end ofNovember. From early December, the coccoid chryso-phyte cells become evident and at around �2�C and37 psu, statocyst formation occurs (Stoecker et al. 1998).Statocyst concentrations also declined rapidly when re-leased to the water column through brine drainage.
Table 3 Mean abundance of auto- and heterotrophic flagellate species in Davis fast ice brine during 1996 and 1997 (n=9, 1996 except for16 Dec onwards where n=3 from O’Gorman Rocks only; n=15, 1997. 1 s d shown in brackets)
1996 Date Mean abundance (· 104 l�1)
11/11 14/11 18/11 21/11 24/11 2/12 4/12 8/12 10/12 16/12 21/12
AutotrophsCryptophyte 7.32
(6.74)5.89(1.86)
2.35(1.54)
0.52(0.12)
Gymnodinium sp.2 0.06(0.17)
0.02(0.03)
0.02(0.02)
0.05(0.03)
0.15(0.13)
0.21(0.83)
4.78(2.01)
0.24(0.92)
Pyramimonasgelidicola
0.23(0.40)
0.75(0.14)
0.36(0.15)
0.79(0.34)
1.49(1.23)
3.06(3.56)
15.26(22.61)
26.66(43.65)
0.11(0.13)
HeterotrophsProtoperidium sp. <0.01 <0.01 <0.01 0.01
(0.01)<0.01 0.21
(0.13)0.32(0.03)
<0.01 <0.01
Gymnodinium sp. <0.01 <0.01 <0.01 0.01(0.01)
0.04(0.03)
<0.01 0.13(0.16)
0.04(0.50)
Gyrodiniumlachryma
<0.01 0.02(0.006)
0.02(0.02)
0.02(0.03)
0.13(0.16)
0.02(0.05)
1997 Date 12/11 14/11 17/11 21/11 24/11 27/11 1/12 5/12 9/12 12/12 15/12
AutotrophsCryptophyte 4.20
(3.16)5.49(1.04)
0.83(0.51)
Flagellate A 0.43(0.32)
0.65(0.46)
1.04(1.0)
Pyramimonasgelidicola
0.25(0.05)
0.46(0.45)
0.56(0.2)
0.28(0.17)
0.49(0.30)
0.42(0.18)
1.15(0.86)
4.43(6.26)
6.58(7.56)
27.23(55.09)
2.29(12.28)
Gymnodinium sp. 2 0.03(0.04)
0.31(0.90)
0.34(1.09)
1.15(2.30)
0.84(2.95)
0.56(1.43)
3.34(6.96)
0.55(0.22)
0.40(0.01)
Mantoniella sp. 0.26(0.20)
0.80(0.74)
0.19(0.51)
0.96(0.89)
1.61(1.37)
2.59(1.43)
3.67(2.72)
HeterotrophsCryothecomonasarmigera
1.15(1.48)
1.12(0.95)
0.36(0.32)
Cryothecomonas sp.2 1.07(1.03)
2.31(1.03)
2.29(1.23)
0.24(0.15)
0.62(0.94)
0.7(0.41)
0.44(0.33)
0.19(0.2)
0.28(0.39)
343

Similarly to McMurdo Sound, P. glacialis, chryso-phytes and a succession of their life cycle forms werethe dominant and recurrent components of the Davisfast ice community. Few P. glacialis cysts were evidentduring early November in both years at Davis and themass excystment observed in McMurdo Sound hadapparently occurred at an earlier date. Asexual P.glacialis cells reached concentrations of1.4·106 cells l�1 in early November 1996 and0.9·106 cells l�1 in early December 1997. This vari-ability was likely due to the lower light climate and thesubstantially colder brine in 1997, an observationconsistent with the light inhibiting effect of snow coveron biomass production in ice communities (Bunt andLee 1970; Palmisano et al. 1985; Grossi et al. 1987;Stoecker et al. 1998, 2000). Possibly as a consequence,sexual reproduction in P. glacialis varied in timingbetween years. In 1996, planozygotes were apparent ataround �1.5�C and 40 psu and increased rapidly innumber from early December. In contrast, planozygoteabundance in 1997 increased with asexual cell con-centration from mid-November when the brine wasapproximately �3.4�C and 80 psu. Encystment oc-curred by mid-December in both years at around�1.8�C and 33 psu, approximating the conditions of�2�C and 37 psu proposed by Stoecker et al. (1998)for this life cycle stage.
The chrysophytes in Davis fast ice showed a similarlytimed life cycle progression in both years. Few statocystswere apparent in early November when the brine wascold and saline (�4.5�C and 94 psu). Flagellate chryso-phyte cells were becoming abundant at this time and itwas apparent that excystment had already occurred. Byearly December, coccoid cells and statocysts were ob-served when the brine temperature and salinity wasaround �1.8�C and 38 psu, similar to �2�C and 37 psufound in McMurdo Sound (Stoecker et al. 1998). By 12December, mass encystment had occurred and statocystconcentrations reached 2.5·106 cells l�1. The most pre-valent statocyst identified at Davis, similar to Archaeo-monas aerolata, is also common in McMurdo Sound(Stoecker et al. 1997).
Total chl a in the upper 50 cm fast ice horizonreached 13.78 and 7.17 mg m�3 in 1996 and 1997,respectively, comparing favourably with brine values ofup to 10 mg chl a m�3 from McMurdo Sound (Stoeckeret al. 2000). The estimated biomass of P. glacialis asex-ual cells at Davis contributed up to 67.2 lg C l�1 ofbrine in 1996 and 39.8 lg C l�1 in 1997, considerablylower than 464.1 lg C l�1 from snow-free, annual fastice at McMurdo Sound (Stoecker et al. 2000). Similarly,the highest biomass of the chrysophyte flagellates andcoccoid cells (4.4 and 14.9 lg C l�1 respectively) in ei-ther year at Davis were substantially lower than themaximums at McMurdo Sound (113.2 l C l�1 for fla-gellates and 62.3 lg C l�1 for the coccoid cells). Thesedifferences give at least some measure of variability inAntarctic fast ice.
The heterotrophs of both locations were remarkablysimilar, with common species including G. lachryma,Protoperidium sp., Gymnodinium sp. [similar to anotherdescribed in Stoecker et al. (1993)], C. armigera and thesmaller Cryothecomonas sp. Autotrophic flagellatesrecorded at Davis but not at McMurdo Sound includedP. gelidicola, the unidentified cryptophyte and flagellateA. Only a small pennate diatom was recorded in theupper ice of McMurdo Sound (Stoecker et al. 1998)whilst at Davis, both P. subcurvata and N. glaciei werepresent in the brine throughout November andDecember. Stoecker et al. (1993) recorded a diverse cil-iate assemblage at McMurdo Sound while at Davis onlyone species of Strombidium was detected.
The similar communities of both locations, and theoccurrence of P. glacialis and chrysophyte cells and cystsalong the East Antarctic coastline indicate a circumpo-lar, fast ice distribution of this community type. In packice, P. glacialis has been recorded from the East Ant-arctic sector (this study), the Scotia Sea (Garrison andBuck 1989) and the Weddell Sea where statocysts havealso been found (Mitchell and Silver 1982; Buck et al.1992). However, the interior communities of pack ice areconsidered to be diatom dominated (Garrison 1991).Reasons for the poor representation of P. glacialis andchrysophytes in pack ice remain unclear but may includethe timing and logistics of sampling pack ice and dif-ferences in the structure of pack and land fast ice (re-viewed in Garrison 1991). Stoecker et al. (1998) suggestthat shipboard sampling when the pack ice is navigableover summer may miss the successional sequence in theflagellate community. We sampled four pack ice sites inOctober 1996 when, judging from the fast ice data,resting stages should have been abundant and onlyfound P. glacialis cysts at one site.
If the pack ice sampling theory is discounted, thelikely explanation may lie in structural differences be-tween fast and pack ice. Fast ice at Davis and McMurdoare reportedly composed of around 80% congelation ice(Gow et al. 1998; Worby et al. 1998), with brine pocketsand channels forming between the boundaries of thevertically orientated, congelation crystals. In contrast,pack ice forms from a random orientation of ice crystalsinto frazil ice (Weeks and Ackley 1982), beneath whichcongelation ice grows. Pack ice thickens by ridging ofconvergent floes and may form complex layers, con-sisting of around 47% frazil ice and 39% congelation ice(Worby et al. 1998). Considering the lack of data on theflagellate community in pack ice and its distribution infast ice, this community may be intimately associatedwith congelation crystal dominated ice. Fast ice adjacentto the McMurdo Ice Shelf can vary in structure, con-sisting of up to 35 and 39% frazil and congelation icerespectively (Gow et al. 1998). Combined protist com-munity and ice structure investigations in a readilyaccessible area such as this may confirm the relationshipbetween flagellate communities and congelation crystaldominated fast ice.
344

Conclusion
The upper fast ice communities of Davis Station andMcMurdo Sound, dominated by P. glacialis andchrysophytes, appears to have a circum-Antarcticdistribution. Furthermore, the life cycles of P. glacialisand the chrysophytes in both regions progressed atsimilar brine temperatures and salinities, despite thegeographical separation of the two locations. Theseemingly conservative and consistent nature of the lifecycles may aid modelling microbial processes in otherfast ice locations where time series studies cannot beperformed. Knowledge of the distribution of thiscommunity, its seasonality and variations in its stand-ing stock will assist future estimates of sea ice pro-ductivity.
Acknowledgements Special thanks to Vanessa Noble for collectingsamples at Mawson Station, to Andrew Davidson for advice andto the expeditionary staff of Davis Station for their support. Theauthors appreciate the comments of the editor and the reviewerswho greatly improved the manuscript. This research was sup-ported by the Institute of Antarctic and Southern Studies(University of Tasmania), ASAC Project 40 and the AustralianAntarctic Division.
References
Ackley SF, Sullivan CW (1994) Physical controls on the develop-ment and characteristics of Antarctic sea ice biological com-munities-a review and synthesis. Deep Sea Res 41:1583–1604
Archer SD, Laekey RJG, Burkill PH, Sleigh MA, Appleby CJ(1996) Microbial ecology of sea ice at a coastal Antarctic site:community composition, biomass and temporal structure. MarEcol Prog Ser 135:179–195
Buck KR, Bolt PA, Bentham WN, Garrison DL (1992) A dino-flagellate cyst from Antarctic sea ice. J Phycol 28:15–18
Bunt JS, Lee CC (1970) Seasonal primary production in Antarcticsea ice at McMurdo Sound in 1967. J Mar Res 28:304
Garrison DL (1991) Antarctic sea ice biota. Am Zool 31:17–33Garrison DL, Buck KR (1989) The biota of Antarctic pack ice in
the Weddell Sea and Antarctic Peninsula Regions. Polar Biol10:211–219
Garrison DL, Buck KR (1991) Surface-layer sea ice assemblages inAntarctic pack ice during the austral spring: environmentalconditions, primary production and community structure. MarEcol Prog Ser 75:161–172
Gow AJ, Ackley SF, Govoni JW, Weeks WF (1998) Physical andstructural properties of land-fast sea ice in McMurdo Sound,Antarctica. In: Jeffries MO (ed) Antarctic sea ice: physicalprocesses, interactions and variability. American GeophysicalUnion, Washington DC, pp 355–374
Gradinger R (1999) Vertical fine structure of the biomass andcomposition of algal communities in Arctic pack ice. Mar Biol133:745–754
Grossi SM, Kottmeier ST, Moe RL, Taylor GT, Sullivan CW(1987) Sea ice microbial communities. IV. Growth and primaryproduction in bottom ice under graded snow cover. Mar EcolProg Ser 35:153–164
Legendre L, Ackley SF, Dieckmann GS, Gulliksen B, Horner R,Hoshiai T, Melnikov IA, Reeburgh WS, Spindler M, SullivanCW (1992) Ecology of sea ice biota. Polar Biol 12:429–444
McConville MJ, Wetherbee R (1983) The bottom-ice microalgalcommunity from annual ice in the inshore waters of east Ant-arctica. J Phycol 19:431–439
Mitchell JG, Silver MW (1982) Modern archaeomonads indicatesea-ice environments. Nature 296:437–439
Montresor M, Procaccini G, Stoecker DK (1999) Polarella gla-cialis, Gen. Nov., sp. Nov (Dinophyceae): Suessiaceae are stillalive. J Phycol 35:186–197
Montresor M, Lovejoy C, Orsini L, Procaccini G, Roy S (2003)Bipolar distribution of the cyst-forming dinoflagellate Polarellaglacialis. Polar Biol 26:186–194
Palmisano AC, Kottmeier ST, Moe RL, Sullivan CW (1985) Seaice Microbial Communities. VI. The effect of light perturbationon microalage at the ice–seawater interface in McMurdoSound, Antarctica. Mar Ecol Prog Ser 21:37–45
Parsons TR, Maita Y, Lalli CM (1984) Determination of chloro-phylls and total carotenoids: spectrophotometric method. In: Amanual of chemical and biological methods for seawater anal-ysis. Pergamon Press, Oxford, pp 101–104
Scott FJ, van den Hoff J (2005) Cryptophytes. In: Scott F, Mar-chant HJ (eds) Antarctic marine protists. Australian BiologicalResources Studies, Canberra, pp 317–318
Stoecker DK, Buck KR, Putt M (1992) Changes in the sea-ice brinecommunity during the spring-summer transition, McMurdoSound, Antarctica. 1. Photosynthetic protists. Mar Ecol ProgSer 84:265–278
Stoecker DK, Buck KR, Putt M (1993) Changes in the sea-ice brinecommunity during the spring -summer transition, McMurdoSound, Antarctica. II. Phagotrophic protists. Mar Ecol ProgSer 95:103–113
Stoecker DK, Gustafson DE, Merrell JR, Black MMD, Baier CT(1997) Excystment and growth of chrysophytes and dinofla-gellates at low temperatures and salinities in Antarctic sea ice.J Phycol 33:585–595
Stoecker D, Gustafson DE, Black MMD, Baier CT (1998) Popu-lation dynamics of microalgae in the upper land-fast sea ice at asnow free location. J Phycol 34:60–69
Stoecker DK, Gustafson DE, Baier T, Black MMD (2000) Primaryproduction in the upper sea ice. Aquat Microb Ecol 21:275–287
Takahashi E, Watanabe K, Satoh H (1986) Siliceous cysts fromKita-no-seto Strait, north of Syowa Station, Antarctica. MemNatl Inst Polar Res 40:84–95
Thomson PG, Wright SW, Bolch CJS, Nichols PD, Skerratt JH,McMinn A (2004) Antarctic distribution, pigment and lipidcomposition, and molecular identification of the brine dinofla-gellate Polarella glacialis (Dinophyceae). J Phycol 40:867–873
Weeks WF, Ackley SF (1982) The growth, structure, and proper-ties of sea ice. United States Army Cold Regions Research andEngineering Laboratory, Hanover
Worby AP, Massom RA, Allison I, Lytle VI, Heil P (1998) EastAntarctic sea ice: a review of its structure, properties and drift.In: Jeffries MO (ed) Antarctic sea ice: physical processes,interactions and variability. American Geophysical Union,Washington, pp 41–67
345
Copyright © 2022 FDOKUMEN




















