
Distribution of Bacterial Proteins in Biofilms Formed by Non-typeable Haemophilus influenzae
15
http://jhc.sagepub.com/ Journal of Histochemistry & Cytochemistry http://jhc.sagepub.com/content/54/7/829 The online version of this article can be found at: DOI: 10.1369/jhc.6A6922.2006 2006 54: 829 J Histochem Cytochem Paul Webster, Siva Wu, Gerardo Gomez, Michael Apicella, Andrew G. Plaut and Joseph W. St. Geme III Haemophilus influenzae Distribution of Bacterial Proteins in Biofilms Formed by Non-typeable Published by: http://www.sagepublications.com On behalf of: Official Journal of The Histochemical Society can be found at: Journal of Histochemistry & Cytochemistry Additional services and information for http://jhc.sagepub.com/cgi/alerts Email Alerts: http://jhc.sagepub.com/subscriptions Subscriptions: http://www.sagepub.com/journalsReprints.nav Reprints: http://www.sagepub.com/journalsPermissions.nav Permissions: What is This? - Jul 1, 2006 Version of Record >> by guest on July 2, 2014 jhc.sagepub.com Downloaded from by guest on July 2, 2014 jhc.sagepub.com Downloaded from
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Distribution of Bacterial Proteins in Biofilms Formed by Non-typeable Haemophilus influenzae

http://jhc.sagepub.com/Journal of Histochemistry & Cytochemistry
http://jhc.sagepub.com/content/54/7/829The online version of this article can be found at:
DOI: 10.1369/jhc.6A6922.2006
2006 54: 829J Histochem CytochemPaul Webster, Siva Wu, Gerardo Gomez, Michael Apicella, Andrew G. Plaut and Joseph W. St. Geme III
Haemophilus influenzaeDistribution of Bacterial Proteins in Biofilms Formed by Non-typeable
Published by:
http://www.sagepublications.com
On behalf of:
Official Journal of The Histochemical Society
can be found at:Journal of Histochemistry & CytochemistryAdditional services and information for
http://jhc.sagepub.com/cgi/alertsEmail Alerts:
http://jhc.sagepub.com/subscriptionsSubscriptions:
http://www.sagepub.com/journalsReprints.navReprints:
http://www.sagepub.com/journalsPermissions.navPermissions:
What is This?
- Jul 1, 2006Version of Record >>
by guest on July 2, 2014jhc.sagepub.comDownloaded from by guest on July 2, 2014jhc.sagepub.comDownloaded from

ARTICLE
Distribution of Bacterial Proteins in Biofilms Formed byNon-typeable Haemophilus influenzae
Paul Webster, Siva Wu, Gerardo Gomez, Michael Apicella, Andrew G. Plaut, andJoseph W. St. Geme III
Ahmanson Advanced Electron Microscopy and Imaging Center, House Ear Institute, Los Angeles, California (PW,SW,GG);Department of Microbiology, University of Iowa, Iowa City, Iowa (MA); Gastroenterology Research, Tufts-New EnglandMedical Center, Boston, Massachusetts (AGP); and Department of Pediatrics, Duke University Medical Center, Durham,North Carolina (JWSG)
SUMMARY The ability to preserve the fragile ultrastructural organization of bacterial bio-films using cryo-preparation methods for electron microscopy has enabled us to probe sec-tions through non-typeable Haemophilus influenzae (NTHi) biofilms and determine thelocalization of NTHi-specific lipooligosaccharide (LOS) and proteins within these structures.Some of the proteins we examined are currently being considered as candidates for vaccinedevelopment, so it is important that their distribution and accessibility within the biofilmsformed by NTHi be determined. We have localized LOS to the extracellular matrix (ECM)of the biofilm and the P6 outer membrane protein to the membrane of what appear to beviable bacteria within the biofilm. The Hap and HWM1/HMW2 adhesive proteins were as-sociated with bacteria within the biofilm and were present in the biofilm ECM. The IgA1protease is a secreted protein that was also associated with NTHi in the biofilm and was in theECM, but was more concentrated in the top region of the biofilm, suggesting a role inprotecting biofilm bacteria from antibody attack. (J Histochem Cytochem 54:829–842, 2006)
KEY WORDS
OMP P6
HMW proteins
Hap protein
IgA1 protease
electron microscopy
extracellular matrix
immunocytochemistry
stereology
NON-TYPEABLEHAEMOPHILUS INFLUENZAE (NTHi) is a com-mon commensal organism in the human nasopharynx(Ingvarsson et al. 1982; Kuklinska and Kilian 1984;Trottier et al. 1989). This organism is also a major otitismedia pathogen (Spinola et al. 1986; Murphy andApicella 1987; Bluestone et al. 1992; Klein 1997; Raoet al. 1999) that accounts for 20 to 30% of all cases ofacute otitis media (Giebink 1989), 40% of cases of otitismedia with effusion (Rayner et al. 1998), and a highpercentage of recurrent cases (Giebink 1989). The tran-sition of NTHi from commensal to pathogen is thoughtto occur as a result of changes in host physiology, per-haps by blockage of the eustachian tube or by loss of themucocilliary elevator. In the compromised host, thebacteria are able to attach to the middle ear mucosa andstimulate an inflammatory response.
Attachment to both inert and living surfaces is acommon property exhibited by many bacteria and isconsidered a first step in biofilm formation. Biofilms havebeen broadly defined as multicellular assemblages ofmicroorganisms and extracellular products attached toan abiotic or biotic surface (Davey and O’Toole 2000).The extracellular products form a matrix, typically richin sugar moieties (Neu et al. 2001), that probably confersprotection from phagocytosis and antibiotic treatment(Donlan and Costerton 2002). Within the biofilm, thebacteria have different growth rates. While some arerapidly growing, others are in a dormant or sessile state, acondition that may be linked with reduced ribosome(rRNA) content (Poulsen et al. 1993).
Otitis media as a biofilm infection was first proposedto explain the observation that 40–60% of patientswith chronic otitis media have sterile middle ear fluids(Rayner et al. 1998), suggesting the absence of cultur-able bacteria (Giebink 1989). The presence of bacterialendotoxin (Dingman et al. 1998), bacterial DNA (Postet al. 1995), and bacterial mRNA (Rayner et al. 1998)in these sterile fluids further supported the hypothesisthat otitis media is a biofilm infection (Dingman et al.
Correspondence to: Paul Webster, PhD, Director, AhmansonAdvanced Electron Microscopy and Imaging Center, House EarInstitute, 2100 West Third Street, Los Angeles, CA 90057. E-mail:[email protected]
Received for publication January 11, 2006; accepted March 3,2006 [DOI: 10.1369/jhc.6A6922.2006].
TheJournal
ofHistoch
emistry&
Cytoch
emistry
C The Histochemical Society, Inc. 0022-1554/06/$3.30 829
Volume 54(7): 829–842, 2006
Journal of Histochemistry & Cytochemistry
http://www.jhc.org
by guest on July 2, 2014jhc.sagepub.comDownloaded from

1998; Costerton 1999). Direct evidence of biofilm for-mation on middle ear mucosa has since been presentedin experimental systems (Post 2001). Bacterial biofilmsin otitis media may help to explain the phenomenon ofpersistent infections that occur in many patients withotitis media (Pichichero 2000; Faden 2001).
Although biofilms have become an accepted conceptin infection, the mechanisms by which they are formedand maintained are still being elucidated. As a con-tribution to the study of biofilms, we have used im-munoelectron microscopy to examine the location ofbacterial adhesins in NTHi biofilms. Specifically, wehave examined the location and distribution of theHMW1/HMW2 proteins, the Hap protein, and IgA1protease. The HMW1/HMW2 proteins (Barenkampand Bodor 1990; Barenkamp and Leininger 1992) in-teract with eukaryotic cell receptors (St Geme et al.1993). The Hap protein (St Geme et al. 1994) and theclosely related IgA1 protease (Kilian et al. 1979,2002)are autotransporters that have both been reported tomediate microcolony formation (Plaut et al. 1992; Finket al. 2003). The Hap protein has also been shown tomediate interactions with human epithelial cells (StGeme et al. 1994; Fink et al. 2003).
The proteinswere selected to explore their distributionwithin a biofilm and because many of them are currentlybeing considered as candidates for vaccine development.We are not aware of earlier reports in which specificantibodies were applied to sections through biofilms thathave been cryo-fixed and freeze substituted.
Materials and Methods
Bacterial Strains, Growth, and Storage
NTHi strain 9274 was originally isolated from the middle earfluid of a patient with acute otitis media (Gu et al. 1995) andwas propagated on chocolate agar at 37C under 5%CO2 and95% humidity. Colonies from the agar plates were trans-ferred to 3.7% brain heart infusion medium (BHI broth;Difco Laboratories, Detroit, MI) containing nicotinamideadenine dinucleotide (NAD, 2 mg/ml) and hemin (10 mg/ml)(both from Sigma Chemical Co., St. Louis, MO). These bac-terial suspensions were incubated overnight at 37C under 5%CO2 and 95% humidity, and the bacteria were then centri-fuged at 15,000 3 g for 5 min. Pellets were resuspended inBHI broth containing 25% glycerol, and the suspension wasstored in small aliquots under liquid nitrogen.
Crystal Violet Assay for Biofilm Formation
The method used to evaluate biofilm formation by NTHistrain 9274 was essentially as previously described (Watnickand Kolter 1999; Murphy and Kirkham 2002). Aliquots of a1:200 dilution of an overnight bacterial culture were placedinto the wells of a 96-well plate and incubated overnight at37C in 5% CO2 and 95% relative humidity. Exactly 24 hrlater, 20 ml of a 1% solution of crystal violet solution wasadded to the wells for 15 min, after which the bacteria were
washed extensively. The wells were air-dried, the dye in thewells was solubilized with DMSO, and the optical density 595(OD595) of each well was read. Sterile BHI was substituted forovernight bacterial cultures in control experiments.
Preparation of NTHi Biofilms on Solid Substrates
Suspensions of NTHi (clone 9274) were removed from stor-age under liquid nitrogen, plated onto chocolate agar plates,and grown overnight at 37C in 5% CO2 and 95% relativehumidity. From these plates, five individual colonies were re-moved and resuspended in BHI broth supplemented withhemin and NAD (Poje and Redford 2003) and incubatedovernight at 37C, 5% CO2, and 95% relative humidity. After20 hr, relative numbers of bacteria in suspension were esti-mated by reading the OD at 600 nm (Poje and Redford 2003).
For electron microscopy, biofilms were prepared on glasscoverslips orMillipore filters. Biofilms on glass coverslipswereprepared by immersing sterile coverslips in a 1:200 dilution ofNTHi suspension in BHI medium and incubating overnight(37C,5%CO2, and95%relative humidity). In some instances,the culture dishes were placed on a slight incline so that thecoverslips were only partially covered with NTHi suspension.
Colony biofilms on filter substrates were prepared using amodification of previously published methods (Anderl et al.2000; Zahller and Stewart 2002; Thurnheer et al. 2003).Sterile Millipore filters (Millipore Corp., Billerica, MA, Cata-log # GSWP 025 00) were placed onto the surface of choco-late agar plates (Hardy Diagnostics, Catalog # E-14) andinoculated with a concentration of NTHi in BHI broth thatproduced a density of approximately 0.3 bacteria/mm2. Thefilters with bacteria were incubated overnight at 37C in 5%CO2 and 95% relative humidity.
Rapid Freezing and Freeze Substitution
Biofilms on glass coverslips and on Millipore filters after 24 hrwere prepared for examination by electron microscopy usingrapid freezing followed by freeze substitution (Webster et al.2004). For scanning electron microscopy (SEM), the coverslipsor filters were rapidly immersed in liquid propane and trans-ferred to dry methanol at 280C for dehydration by freezesubstitution. The frozen specimens were gradually warmed to4C, critical point dried, and given a 6-nm coating of platinumbefore being examined by field emission SEM (using an XL 30SFEG SEM; FEI Company, Hillsboro, OR).
Biofilms on filter substrates were cut into 0.8-mm discsusing a specially manufactured cutter (Morphew andMcIntosh2003) and placed biofilm-face down into aluminum plan-chettes (Ted Pella Inc.; Redwood, CA) containing a thin film ofhexadecene. A second aluminum planchette was placed overeach filter, and the specimen was then immediately frozen at2100 bar in anHPFmachine (HPM010; Bal-Tec, Lichtenstein)using established methods (Sawaguchi et al. 2002,2003).
Frozen filters were placed into polypropylene specimencapsules (Leica Inc.; Deerfield, IL) and either stored underliquid nitrogen for further processing, or immediately trans-ferred to cold, dry ethanol maintained at a temperature of280C in an AFS freeze substitution machine (Leica). Speci-mens were left overnight at280C and were gradually warmedto 260C over a 6-hr period. During the warming period, thespecimens were exposed to two changes of fresh, dry ethanol.
TheJournal
ofHistoch
emistry&
Cytoch
emistry
830 Webster, Wu, Gomez, Apicella, Plaut, St. Geme
by guest on July 2, 2014jhc.sagepub.comDownloaded from

At a temperature of 260C, the specimens were infiltratedwith a 1:1 mixture of ethanol and Lowicryl HM20 resin(Electron Microscopy Sciences; Ft. Washington, PA) for 2 hrand then in a fresh 1:1 ethanol/resin mixture overnight. Thenext day, the specimens were warmed to250C, transferred tofresh Lowicryl HM20 resin, and polymerized under UV light,following protocols provided with the resin kit.
Immunocytochemistry
Thin sections (60–80 nm) through the Lowicryl-embeddedbiofilms were prepared using an Ultracut S ultramicrotome(Leica) with a diamond knife (Diatome USA; Ft. Washington,PA) and collected on metal specimen grids.
The sections were labeled with antibodies using sequentiallabeling protocols (Griffiths 1993) by floating the grids,section side down on small drops of reagent. Polyclonalantibodies to H. influenzae (Difco Laboratories) and to theNTHi IgA1 protease (Plaut et al. 1992) were applied tosections and visualized using 10 nm protein A-gold (PAG;University of Utrecht, The Netherlands). Monoclonal anti-body 3B9 against P6 outer membrane protein (OMP)(Murphy et al. 1985) and monoclonal 6E4 against lipooligo-saccharide (LOS) (Campagnari et al. 1990) were applied tosections and visualized by applying anti-mouse IgG bridgingantibodies produced in rabbits, and then 10 nm PAG. TheHap protein was detected using guinea pig antiserum GP74(Cutter et al. 2002), and the HMW1/HMW2 was detectedusing guinea pig antisera raised against purified HMW1, butwhich recognizes both HMW1 and HMW2 (Buscher et al.2004). The anti-Hap and HMW1/HMW2 were applied tosections and visualized by applying rabbit anti-guinea pig IgGantibodies and 10 nm PAG.
All polyclonal secondary antibodies were diluted in 10%goat serum containing a suspension of heat-killed, homoge-nized NTHi bacteria and centrifuged before use. This treat-ment reduced the nonspecific binding observed with somepolyclonal secondary antibody preparations.
Electron Microscopy
All sectionswere examinedby transmission electronmicroscopy(TEM), using a CM120 BioTwin operating at 80 kV (FEI Com-pany). Imageswere collectedonnegative filmanddigitizedusinga high-resolution digital scanner. Quantification of immuno-gold labeling over NTHi biofilms was performed on imagesobtained using systematic sampling protocols (Lucocq 1993).
Western Blotting
Biofilms of NTHi were grown on Millipore filters as describedabove. The bacteria growing on the filters were solubilizedeither to obtain an enrichment of extracellular matrix (ECM),or to obtain a whole-biofilm homogenate. ECM proteins wereobtained by solubilizing the filters in 1 3 PBS using a probesonicator. The filters were exposed to four rounds of coolingon ice, sonication for 3 min, and cooling for 10 min. Wholebacteria were removed by centrifugation, and the supernatantswere subjected to an SDS-PAGE separation using a NuPAGEsystem and Novex bis-Tris gels (Invitrogen; Carlsbad, CA).
Whole homogenates of biofilm were obtained by solu-bilizing and sonicating the biofilms in lysis buffer (50 mMTris-HCL, 150 mM NaCl, 1% Triton X-100, pH 7.4). The
sonication conditions applied were the same as those used forobtaining biofilm ECM proteins (4 3 3 min sonication and10 min cooling). The supernatants were separated from filterfragments by centrifugation. This solubilization protocol wasalso used to obtain whole-cell lysates from pellets of plank-tonic NTHi that had been grown in suspension.
SDS-PAGE-separated proteins were blotted onto nitrocel-lulose membranes and blocked by incubation with Tris-buffered saline Tween-20 (50 mM Tris-HCL, 150 mM NaCl,0.1% Tween 20, pH 7.5) containing 2% fish skin gelatin. Thebuffer used to dilute secondary and tertiary antibodies alsocontained suspensions of bacterial proteins from heat-killed,lysed NTHi to remove non-relevant bacteria-specific contam-inating antibodies. The blots were washed and then incubatedsequentially with specific antibodies to NTHi proteins, sec-ondary antibodies produced in rabbits, and HRP-conjugatedtertiary antibodies produced in goats. The antibodies werevisualized using a chemiluminescent horseradish peroxidasesubstrate kit (Vector Laboratories; Burlingame, CA).
Results
Formation of NTHi Biofilms on Solid Substrates:Crystal Violet Assay
To establish that NTHi strain 9274 has the ability toform biofilms, we first performed a crystal violet assayon overnight bacterial cultures grown in a 96-well plate.The crystal violet stained a significant amount of ma-terial that had been deposited on the sides of the wells bythe growing bacteria (Figure 1A), compared with crystalviolet staining of wells that contained only culture me-dium (Figure 1B). Measurement of the OD595 of solu-bilized crystal violet stain enabled this observation to beexpressed in relative units (Figure 1C).
SEM of Biofilms on a Glass Substrate and Filters
To examine the biofilms formed by the bacteria, wegrew overnight cultures of NTHi strain 9274 in plasticdishes containing glass coverslips. The material depos-ited on the coverslips stained with crystal violet (datanot shown), and in the SEM was shown to be a thicklayer of amorphous material containing embeddedbacteria (Figure 2A). When coverslips were incubatedhalf submerged in the bacterial suspension, a thickband of bacteria and associated amorphous materialformed at the air–liquid interface (Figures 2A and 2B).The material that was attached to the glass substratewas a thick layer of bacteria covered by a layer ofamorphous material, presumably extracellular materialgenerated by the NTHi (Figure 2C). Large clumps ofbacteria that appeared to be held together by thisextracellular material were observed on the glass thathad been submerged (Figure 2C) as well as on the glasssurface exposed to air (data not shown).
To examine NTHi biofilms in the TEM, it wasnecessary to prepare them on substrates that couldbe thin sectioned. For this reason, we generated bio-films on filters that could be frozen using a high-
TheJournal
ofHistoch
emistry&
Cytoch
emistry
Immunolabeling of NTHi Biofilms 831
by guest on July 2, 2014jhc.sagepub.comDownloaded from

pressure freezer and sectioned. Our first attempts,which consisted of growing NTHi biofilms on Anoporeinsert filters with the sterile side immersed in culturemedium, produced structures with typical biofilm char-acteristics. Large clumps of bacteria were present on thefilter surface, and in the SEM we observed what ap-
peared to be channels running through thick mono-layers of bacteria (data not shown).
The Anopore filters were too brittle and hard for usein the high-pressure freezer and for sectioning. For thisreason, we turned to growing the NTHi biofilms on
Figure 1 Crystal violet assay. Crystal violet was used to stain biofilmmaterials deposited by NTHi strain 9274. Biofilm material in thewells that contained non-typeable Haemophilus influenzae (NTHi) isstained with crystal violet (Lane A). Wells that only contained culturemedium did not stain (Lane B). The stain was quantified by readingthe optical density 595 (OD595nm) of the solubilized stain (C). Errorbars are standard errors derived from five duplicate experiments.
’
Figure 2 Scanning electron microscopy (SEM) images of NTHi bio-film growing on partially immersed glass substrates. (A) Low-mag-nification image of the zone at the air–liquid interface. The surfaceimmersed in NTHi suspension is on the left of the image (liquid) andshows NTHi attached to the glass. The region exposed to the air is onthe right (air) and also shows NTHi attached to the surface. (B) Athigher magnification, it is possible to observe a layer of amorphousmaterial covering the NTHi attached to the glass on the air-exposedregion of the substrate. (C) Individual NTHi particles not fully em-bedded in the amorphous material are covered with similar amor-phous material. Bars: A 5 50 mm; B 5 1 mm; C 5 0.5 mm.
TheJournal
ofHistoch
emistry&
Cytoch
emistry
832 Webster, Wu, Gomez, Apicella, Plaut, St. Geme
by guest on July 2, 2014jhc.sagepub.comDownloaded from

Millipore filters. Incubating filters overnight that hadbeen inoculated with 0.3 bacteria per square micronproduced an almost uniformly thick layer of bacteriaon the filters (Figure 3). The biofilms were restricted tothe areas of the filters that had been covered by theinitial innoculum of NTHi, and spreading of bacteriabeyond the edges of the biofilm was not observed.However, at the edges of the biofilm zone, the mat ofbacteria tapered down to a layer that was only onebacterium thick (Figure 3A). The region immediatelyoutside the biofilm had a layer of extracellular materialcovering the support filter (Figure 3A) that containedsmall numbers of embedded bacteria (Figure 3A). Thetop of the formed biofilm was uniformly flat and wascovered with a layer of extracellular material over thebacteria and the spaces between them (Figure 3B).Mechanical damage to the bacterial biofilm, especiallyat its edge, revealed the close association of the extra-cellular material with the bacteria (Figure 3C), and thelarge amount of this extracellular material that wasbeing produced by the bacteria (Figure 3C).
These Millipore filters could be easily manipulatedfor high-pressure freezing, resin-embedding, and section-ing. All subsequent sections through biofilms were ofLowicryl-embedded biofilms that had been grown onMillipore filters and prepared using low-temperaturemethods.
Immunocytochemistry on Lowicryl-embeddedBiofilm Sections
When we examined the biofilm sections by TEM, wewere able to easily identify four distinct compartments.The NTHi in the biofilm occupied 68% of the biofilm(Table 1) and consisted of the cell cytoplasm and theouter cell membrane. An electron-opaque, amorphousmaterial that we classified as ECM surrounding the bac-teria occupied 25% of the biofilm, and the remaining7% of the biofilm was occupied by what we designatedas extracellular space (ES), defined as the areas withinthe biofilm that did not contain stainable or electron-opaque material (Table 1).
Anti-NTHi, anti-P6, and anti-LOS Labeling ofBiofilm Sections
To establish that the bacteria in the biofilms wereNTHi, and that the structures being formed on the sup-
’
Figure 3 SEM images of NTHi colony biofilm growing on Milliporefilters. (A) The NTHi grow on the filter substrate and form a thick matof bacteria. At the edge of the NTHi biofilm, the bacterial mat tapersdown to a layer one bacterium thick. However, amorphous materialextends from the biofilm onto the support filter and appears tocover NTHi that are up to 10 mm away from the edge of the biofilm(arrows). (B) The top surface of this biofilm is covered with a filmof amorphous material that hides the structure of the individualbacteria. This film appears to also cover regions with no underlyingbacteria (arrows). (C) At the edge of the biofilm, the amorphousmaterial often undergoes mechanical damage, perhaps as a con-sequence of specimen processing. In this image, the damage appearsto consist of the biofilm edge being pulled away, giving a better viewof the amorphous material covering the NTHi. Other bacteria, inclose proximity to the biofilm, are observed covered with the amor-phous material (arrow). Bars: A 5 10 mm; B 5 1 mm; C 5 2 mm.
TheJournal
ofHistoch
emistry&
Cytoch
emistry
Immunolabeling of NTHi Biofilms 833
by guest on July 2, 2014jhc.sagepub.comDownloaded from

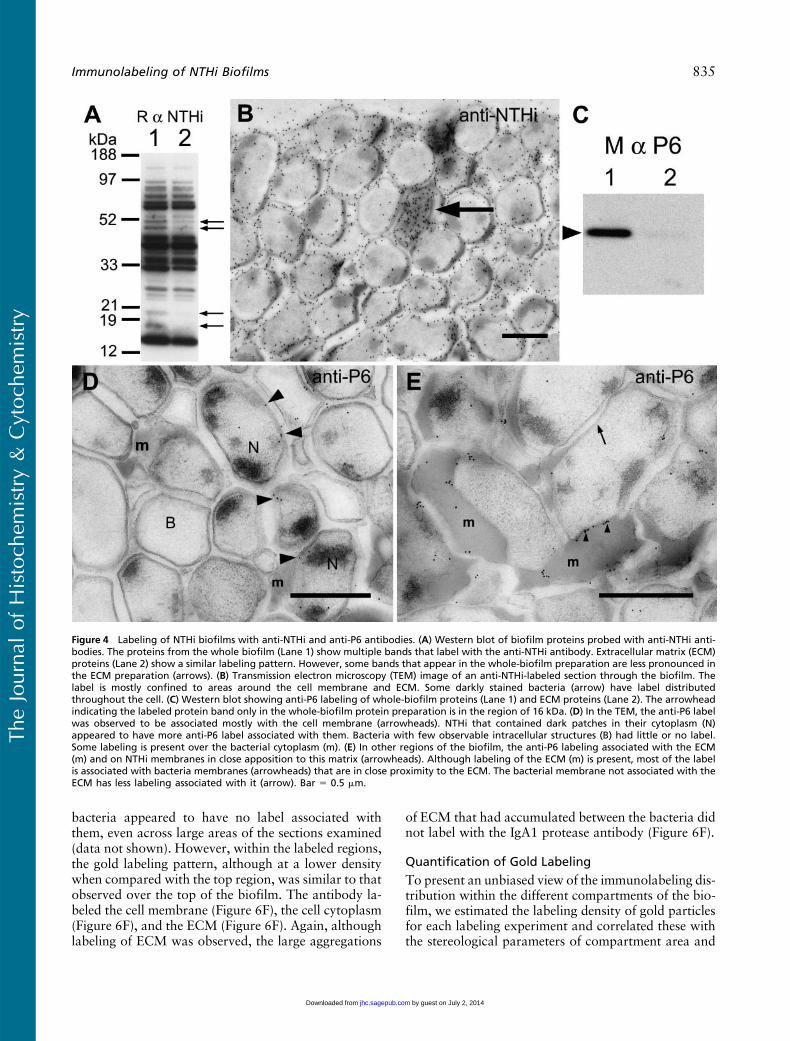
port filters were biofilms, we first labeled the sec-tions with reference antibodies. The first antibody thatwas applied to the sectioned biofilms was a commer-cial polyclonal antibody that recognizes multipleNTHi proteins. This antibody bound to multiple pro-tein bands on Western blots of NTHi whole-cell lysates(Figure 4A) and to protein bands from ECM proteins(Figure 4A). Some protein bands that labeled well inthe whole-biofilm protein lane (Figure 4A, Lane 1)were not so visible in the protein bands from the ECM(Figure 4A, Lane 2). By electron microscopy, the anti-body produced substantial labeling over the biofilmsections (Figure 4B), associating with the bacteria (Fig-ure 4B) and with the ECM (Figure 4B).
We then probed the biofilm with a monoclonal anti-body to the H. influenzae P6 OMP. This protein wasexpressed by planktonic and biofilm forms of NTHi andwas restricted to the outer membrane. OnWestern blots,the anti-P6 antibody bound to a protein band with Mr
approximately 16.6 kDa (Figure 4C), but was confinedto the lane containing proteins from whole biofilms(Figure 4C, Lane 1). No similar protein was labeled inthe lane containing ECM proteins (Figure 4C, Lane 2),suggesting that minimal contamination of bacterial outermembranes in the ECM preparations was occurring.
The anti-P6 labeling of sections through the NTHibiofilm (Figures 4D and 4E) was associated with bacte-ria that appeared to contain more cytoplasmic electron-dense material (Figure 4D). Many of the NTHi inthe biofilm did not label with the anti-P6 antibodies(Figure 4D), and in some regions, the anti-P6 label wasassociated with bacterial membranes close to the ECM(Figure 4E). In some instances, the label over the bacte-rial membrane was concentrated only where the mem-brane was in close apposition to the ECM (Figure 4E).
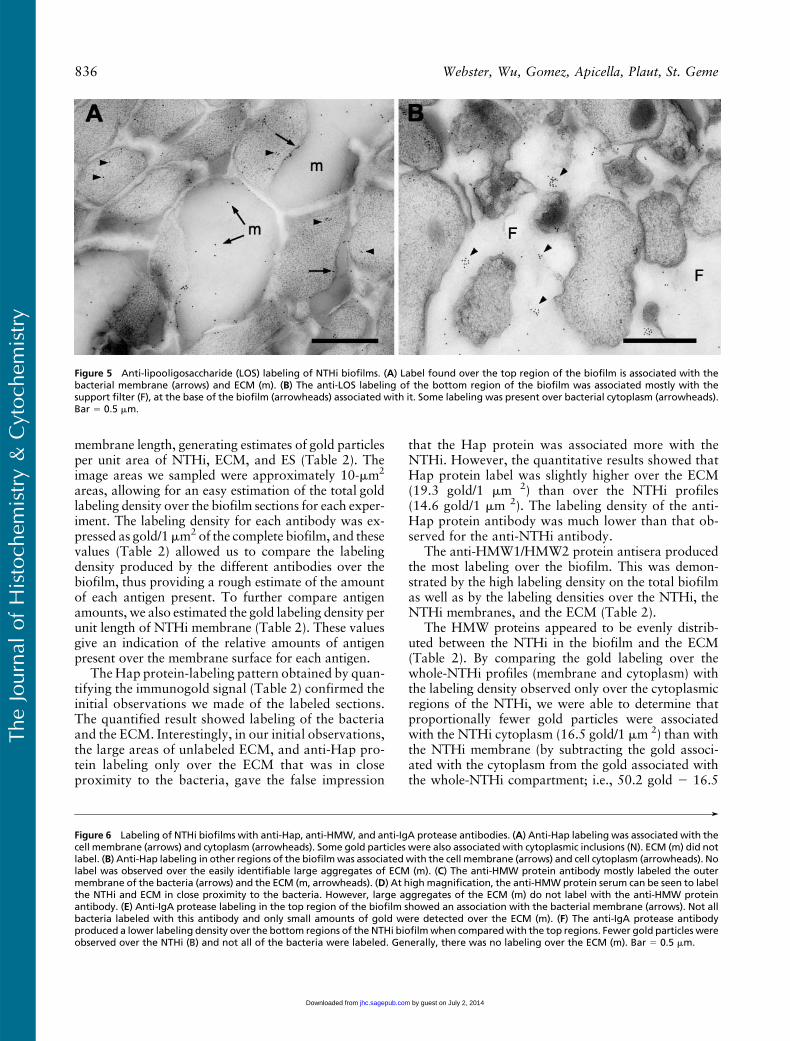
Anti-LOS labeling was also demonstrated on thebiofilm sections, with the label being mostly associatedwith the ECM (Figure 5A). Some label was observedover the cell cytoplasm that could represent newly syn-thesized LOS or nonspecific background label. In thebottom region of the biofilm, the anti-LOS label wasmostly associated the ECM (data not shown), and insome instances, anti-LOS label was observed in regionsof the support filter (Figure 5B), close to bacteria thathad migrated into the support filter (Figure 5B). Fromthese observations it is possible to speculate that the
LOS being shed by the NTHi plays a role in the initialattachment of bacteria to substrates.
Antibodies to Proteins that May Be Involved withBiofilm Formation
Of particular interest to the study of biofilms is the pro-cess of bacterial adhesion. For this reason we treatedbiofilm sections with antibodies to known adhesinsto examine the distribution of these molecules withinthe biofilm.
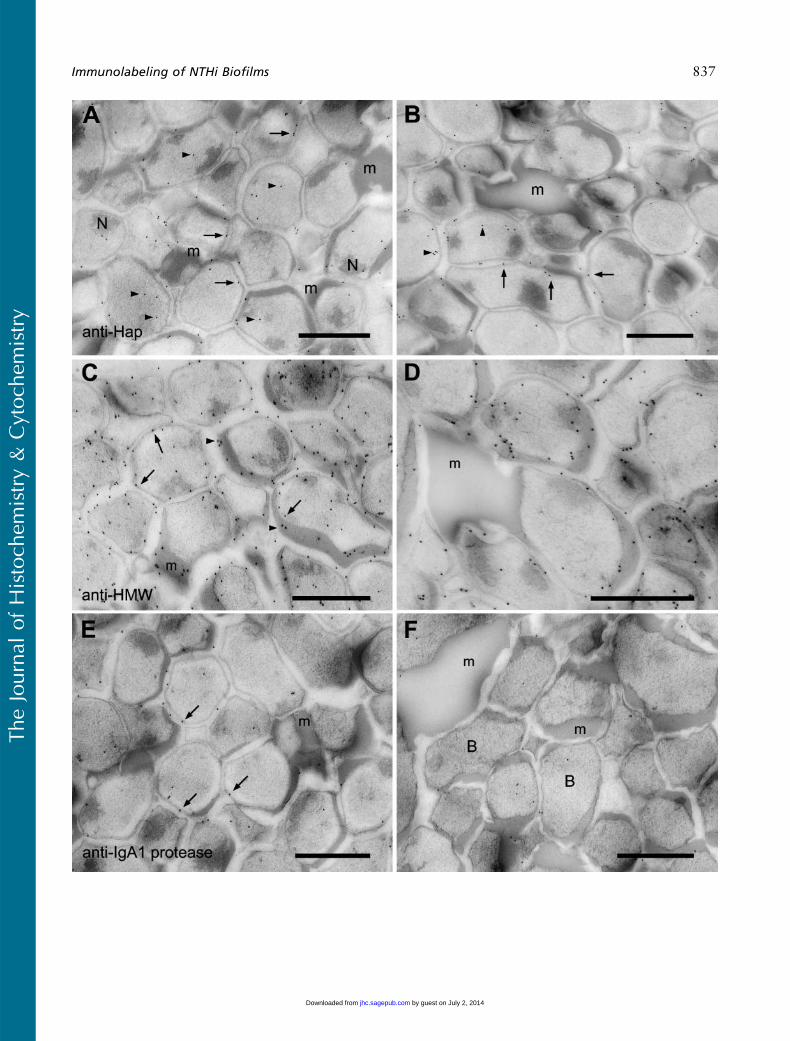
Anti-Hap Protein Labeling
Labeling with antiserum to the Hap protein was asso-ciated with the bacteria in the biofilm (Figures 6A and6B). The label was observed over the bacterial mem-brane and over the cell cytoplasm (Figures 6A and 6B).Although the ECM was labeled with the antiserum,(Figure 6B), the label was confined to material in closeproximity to the bacteria (Figure 6B). The large accu-mulations of ECM in the biofilm did not appear to belabeled (Figures 6A and 6B). In comparison to labelingwith the anti-NTHi antibody, the anti-Hap labelingwas much less, suggesting a lower amount of this anti-gen in the biofilm.
Anti-HMW Protein Labeling
The localization of the anti-HMW1/HMW2 proteinantiserum labeling was also distributed through thewhole of the biofilm (Figures 6C and 6D), and the ex-tensive labeling seen over the biofilm sections suggestedthe presence of a large amount of antigen. The label, al-though concentrated around the surface of the bacteria,was also observed in small amounts over the cell cyto-plasm (Figures 6C and 6D) and over the ECM (Fig-ures 6C and 6D). The large accumulations of ECMin the biofilm did not label with the antiserum (Fig-ure 6D). However, the anti HMW1/HMW2 antiserumdid bind to ECM that was in close association with thebacteria (Figure 6D).
Anti-IgA1 Protease Labeling
Anti-IgA1 protease labeling was present over all regionsof the biofilm, with the majority of the labeling appear-ing to be associated with the cell membrane (Figures 6Eand 6F). What was immediately obvious from initialobservations was that the top region of the biofilm hadmore gold particles over the bacteria than did the bot-tom region. In the top region of the biofilm, labelingwas obvious and mostly associated with the cell mem-brane (Figure 6E), although small amounts of labelwere observed associated with the cell cytoplasm(Figure 6E) and with the ECM (Figure 6E).
The density of the gold label over the bottom regionof the biofilm using anti-IgA1 protease was less thanthat observed for the top region. Many of the biofilm
Table 1 Volume densities of biofilm compartments
Compartment Vv Percentage
Bacteria (NTHi) 0.68 6 0.7 68%Extracellular matrix 0.25 6 0.5 25%Extracellular space 0.07 6 0.3 7%
Stereological estimation of the composition of high-pressure-frozen NTHibiofilms. Relative volumes occupied by each compartment are expressed asvolume densities and as percentages. Vv, volumn densities.
TheJournal
ofHistoch
emistry&
Cytoch
emistry
834 Webster, Wu, Gomez, Apicella, Plaut, St. Geme
by guest on July 2, 2014jhc.sagepub.comDownloaded from
bacteria appeared to have no label associated withthem, even across large areas of the sections examined(data not shown). However, within the labeled regions,the gold labeling pattern, although at a lower densitywhen compared with the top region, was similar to thatobserved over the top of the biofilm. The antibody la-beled the cell membrane (Figure 6F), the cell cytoplasm(Figure 6F), and the ECM (Figure 6F). Again, althoughlabeling of ECM was observed, the large aggregations
of ECM that had accumulated between the bacteria didnot label with the IgA1 protease antibody (Figure 6F).
Quantification of Gold Labeling
To present an unbiased view of the immunolabeling dis-tribution within the different compartments of the bio-film, we estimated the labeling density of gold particlesfor each labeling experiment and correlated these withthe stereological parameters of compartment area and
Figure 4 Labeling of NTHi biofilms with anti-NTHi and anti-P6 antibodies. (A) Western blot of biofilm proteins probed with anti-NTHi anti-bodies. The proteins from the whole biofilm (Lane 1) show multiple bands that label with the anti-NTHi antibody. Extracellular matrix (ECM)proteins (Lane 2) show a similar labeling pattern. However, some bands that appear in the whole-biofilm preparation are less pronounced inthe ECM preparation (arrows). (B) Transmission electron microscopy (TEM) image of an anti-NTHi-labeled section through the biofilm. Thelabel is mostly confined to areas around the cell membrane and ECM. Some darkly stained bacteria (arrow) have label distributedthroughout the cell. (C) Western blot showing anti-P6 labeling of whole-biofilm proteins (Lane 1) and ECM proteins (Lane 2). The arrowheadindicating the labeled protein band only in the whole-biofilm protein preparation is in the region of 16 kDa. (D) In the TEM, the anti-P6 labelwas observed to be associated mostly with the cell membrane (arrowheads). NTHi that contained dark patches in their cytoplasm (N)appeared to have more anti-P6 label associated with them. Bacteria with few observable intracellular structures (B) had little or no label.Some labeling is present over the bacterial cytoplasm (m). (E) In other regions of the biofilm, the anti-P6 labeling associated with the ECM(m) and on NTHi membranes in close apposition to this matrix (arrowheads). Although labeling of the ECM (m) is present, most of the labelis associated with bacteria membranes (arrowheads) that are in close proximity to the ECM. The bacterial membrane not associated with theECM has less labeling associated with it (arrow). Bar 5 0.5 mm.
TheJournal
ofHistoch
emistry&
Cytoch
emistry
Immunolabeling of NTHi Biofilms 835
by guest on July 2, 2014jhc.sagepub.comDownloaded from
membrane length, generating estimates of gold particlesper unit area of NTHi, ECM, and ES (Table 2). Theimage areas we sampled were approximately 10-mm2
areas, allowing for an easy estimation of the total goldlabeling density over the biofilm sections for each exper-iment. The labeling density for each antibody was ex-pressed as gold/1mm2 of the complete biofilm, and thesevalues (Table 2) allowed us to compare the labelingdensity produced by the different antibodies over thebiofilm, thus providing a rough estimate of the amountof each antigen present. To further compare antigenamounts, we also estimated the gold labeling density perunit length of NTHi membrane (Table 2). These valuesgive an indication of the relative amounts of antigenpresent over the membrane surface for each antigen.
The Hap protein-labeling pattern obtained by quan-tifying the immunogold signal (Table 2) confirmed theinitial observations we made of the labeled sections.The quantified result showed labeling of the bacteriaand the ECM. Interestingly, in our initial observations,the large areas of unlabeled ECM, and anti-Hap pro-tein labeling only over the ECM that was in closeproximity to the bacteria, gave the false impression
that the Hap protein was associated more with theNTHi. However, the quantitative results showed thatHap protein label was slightly higher over the ECM(19.3 gold/1 mm 2) than over the NTHi profiles(14.6 gold/1 mm 2). The labeling density of the anti-Hap protein antibody was much lower than that ob-served for the anti-NTHi antibody.
The anti-HMW1/HMW2 protein antisera producedthe most labeling over the biofilm. This was demon-strated by the high labeling density on the total biofilmas well as by the labeling densities over the NTHi, theNTHi membranes, and the ECM (Table 2).
The HMW proteins appeared to be evenly distrib-uted between the NTHi in the biofilm and the ECM(Table 2). By comparing the gold labeling over thewhole-NTHi profiles (membrane and cytoplasm) withthe labeling density observed only over the cytoplasmicregions of the NTHi, we were able to determine thatproportionally fewer gold particles were associatedwith the NTHi cytoplasm (16.5 gold/1 mm 2) than withthe NTHi membrane (by subtracting the gold associ-ated with the cytoplasm from the gold associated withthe whole-NTHi compartment; i.e., 50.2 gold 2 16.5
Figure 5 Anti-lipooligosaccharide (LOS) labeling of NTHi biofilms. (A) Label found over the top region of the biofilm is associated with thebacterial membrane (arrows) and ECM (m). (B) The anti-LOS labeling of the bottom region of the biofilm was associated mostly with thesupport filter (F), at the base of the biofilm (arrowheads) associated with it. Some labeling was present over bacterial cytoplasm (arrowheads).Bar 5 0.5 mm.
’
Figure 6 Labeling of NTHi biofilms with anti-Hap, anti-HMW, and anti-IgA protease antibodies. (A) Anti-Hap labeling was associated with thecell membrane (arrows) and cytoplasm (arrowheads). Some gold particles were also associated with cytoplasmic inclusions (N). ECM (m) did notlabel. (B) Anti-Hap labeling in other regions of the biofilmwas associated with the cell membrane (arrows) and cell cytoplasm (arrowheads). Nolabel was observed over the easily identifiable large aggregates of ECM (m). (C) The anti-HMW protein antibody mostly labeled the outermembrane of the bacteria (arrows) and the ECM (m, arrowheads). (D) At high magnification, the anti-HMW protein serum can be seen to labelthe NTHi and ECM in close proximity to the bacteria. However, large aggregates of the ECM (m) do not label with the anti-HMW proteinantibody. (E) Anti-IgA protease labeling in the top region of the biofilm showed an association with the bacterial membrane (arrows). Not allbacteria labeled with this antibody and only small amounts of gold were detected over the ECM (m). (F) The anti-IgA protease antibodyproduced a lower labeling density over the bottom regions of the NTHi biofilmwhen comparedwith the top regions. Fewer gold particles wereobserved over the NTHi (B) and not all of the bacteria were labeled. Generally, there was no labeling over the ECM (m). Bar 5 0.5 mm.
TheJournal
ofHistoch
emistry&
Cytoch
emistry
836 Webster, Wu, Gomez, Apicella, Plaut, St. Geme
by guest on July 2, 2014jhc.sagepub.comDownloaded from
TheJournal
ofHistoch
emistry&
Cytoch
emistry
Immunolabeling of NTHi Biofilms 837
by guest on July 2, 2014jhc.sagepub.comDownloaded from
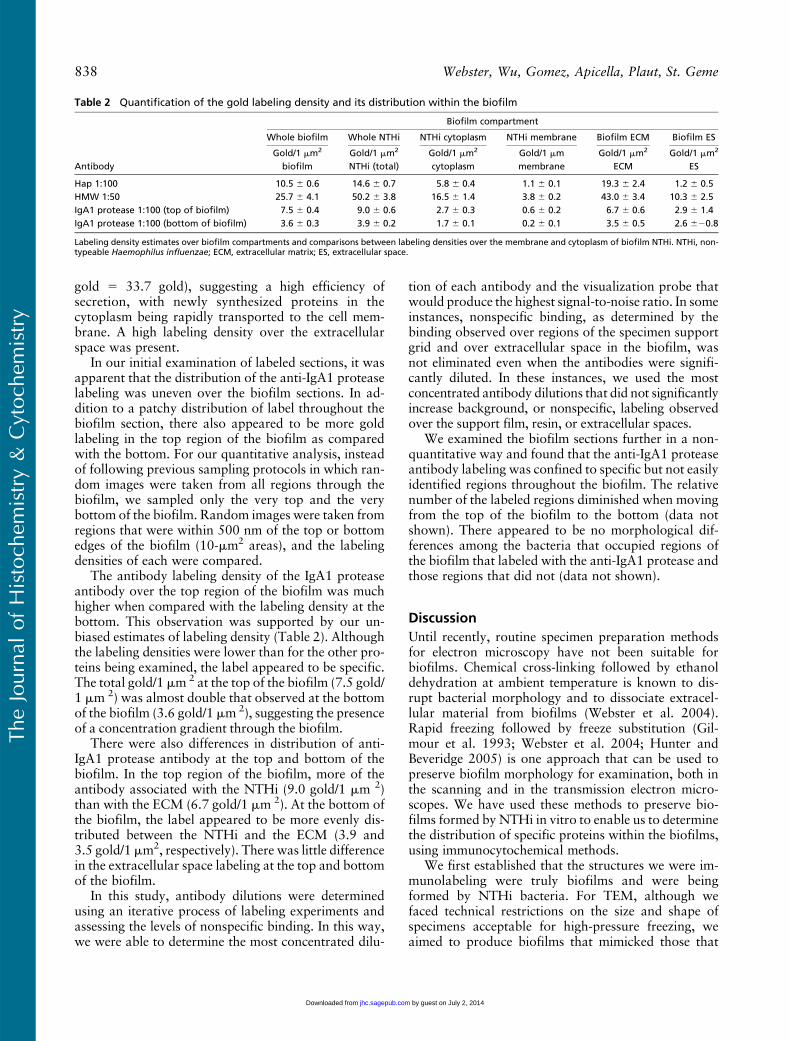
gold 5 33.7 gold), suggesting a high efficiency ofsecretion, with newly synthesized proteins in thecytoplasm being rapidly transported to the cell mem-brane. A high labeling density over the extracellularspace was present.
In our initial examination of labeled sections, it wasapparent that the distribution of the anti-IgA1 proteaselabeling was uneven over the biofilm sections. In ad-dition to a patchy distribution of label throughout thebiofilm section, there also appeared to be more goldlabeling in the top region of the biofilm as comparedwith the bottom. For our quantitative analysis, insteadof following previous sampling protocols in which ran-dom images were taken from all regions through thebiofilm, we sampled only the very top and the verybottom of the biofilm. Random images were taken fromregions that were within 500 nm of the top or bottomedges of the biofilm (10-mm2 areas), and the labelingdensities of each were compared.
The antibody labeling density of the IgA1 proteaseantibody over the top region of the biofilm was muchhigher when compared with the labeling density at thebottom. This observation was supported by our un-biased estimates of labeling density (Table 2). Althoughthe labeling densities were lower than for the other pro-teins being examined, the label appeared to be specific.The total gold/1 mm 2 at the top of the biofilm (7.5 gold/1 mm 2) was almost double that observed at the bottomof the biofilm (3.6 gold/1 mm 2), suggesting the presenceof a concentration gradient through the biofilm.
There were also differences in distribution of anti-IgA1 protease antibody at the top and bottom of thebiofilm. In the top region of the biofilm, more of theantibody associated with the NTHi (9.0 gold/1 mm 2)than with the ECM (6.7 gold/1 mm 2). At the bottom ofthe biofilm, the label appeared to be more evenly dis-tributed between the NTHi and the ECM (3.9 and3.5 gold/1 mm2, respectively). There was little differencein the extracellular space labeling at the top and bottomof the biofilm.
In this study, antibody dilutions were determinedusing an iterative process of labeling experiments andassessing the levels of nonspecific binding. In this way,we were able to determine the most concentrated dilu-
tion of each antibody and the visualization probe thatwould produce the highest signal-to-noise ratio. In someinstances, nonspecific binding, as determined by thebinding observed over regions of the specimen supportgrid and over extracellular space in the biofilm, wasnot eliminated even when the antibodies were signifi-cantly diluted. In these instances, we used the mostconcentrated antibody dilutions that did not significantlyincrease background, or nonspecific, labeling observedover the support film, resin, or extracellular spaces.
We examined the biofilm sections further in a non-quantitative way and found that the anti-IgA1 proteaseantibody labeling was confined to specific but not easilyidentified regions throughout the biofilm. The relativenumber of the labeled regions diminished when movingfrom the top of the biofilm to the bottom (data notshown). There appeared to be no morphological dif-ferences among the bacteria that occupied regions ofthe biofilm that labeled with the anti-IgA1 protease andthose regions that did not (data not shown).
Discussion
Until recently, routine specimen preparation methodsfor electron microscopy have not been suitable forbiofilms. Chemical cross-linking followed by ethanoldehydration at ambient temperature is known to dis-rupt bacterial morphology and to dissociate extracel-lular material from biofilms (Webster et al. 2004).Rapid freezing followed by freeze substitution (Gil-mour et al. 1993; Webster et al. 2004; Hunter andBeveridge 2005) is one approach that can be used topreserve biofilm morphology for examination, both inthe scanning and in the transmission electron micro-scopes. We have used these methods to preserve bio-films formed by NTHi in vitro to enable us to determinethe distribution of specific proteins within the biofilms,using immunocytochemical methods.
We first established that the structures we were im-munolabeling were truly biofilms and were beingformed by NTHi bacteria. For TEM, although wefaced technical restrictions on the size and shape ofspecimens acceptable for high-pressure freezing, weaimed to produce biofilms that mimicked those that
Table 2 Quantification of the gold labeling density and its distribution within the biofilm
Biofilm compartment
Whole biofilm Whole NTHi NTHi cytoplasm NTHi membrane Biofilm ECM Biofilm ES
AntibodyGold/1 mm2
biofilmGold/1 mm2
NTHi (total)Gold/1 mm2
cytoplasmGold/1 mmmembrane
Gold/1 mm2
ECMGold/1 mm2
ES
Hap 1:100 10.5 6 0.6 14.6 6 0.7 5.8 6 0.4 1.1 6 0.1 19.3 6 2.4 1.2 6 0.5HMW 1:50 25.7 6 4.1 50.2 6 3.8 16.5 6 1.4 3.8 6 0.2 43.0 6 3.4 10.3 6 2.5IgA1 protease 1:100 (top of biofilm) 7.5 6 0.4 9.0 6 0.6 2.7 6 0.3 0.6 6 0.2 6.7 6 0.6 2.9 6 1.4IgA1 protease 1:100 (bottom of biofilm) 3.6 6 0.3 3.9 6 0.2 1.7 6 0.1 0.2 6 0.1 3.5 6 0.5 2.6 620.8
Labeling density estimates over biofilm compartments and comparisons between labeling densities over the membrane and cytoplasm of biofilm NTHi. NTHi, non-typeable Haemophilus influenzae; ECM, extracellular matrix; ES, extracellular space.
TheJournal
ofHistoch
emistry&
Cytoch
emistry
838 Webster, Wu, Gomez, Apicella, Plaut, St. Geme
by guest on July 2, 2014jhc.sagepub.comDownloaded from

may form in the middle ear. During an infection, suchbiofilms colonize biotic surfaces within or on top of amucous layer that covers epithelial cells. Newly form-ing biofilms have the possibility of being exposed onone surface to a non-liquid environment of humid air.Watnick and Kolter (1999) have previously shown thatthe morphology of biofilms formed in static culture(colony biofilms) is similar to that of biofilms formed inflow cells. Colony biofilms have thus become a usefultool for antibiotic testing (Anderl et al. 2000,2003).
The crystal violet assay established that the NTHistrain used in this study has the ability to form biofilms.An SEM examination of the biofilm material formed byNTHi on solid substrates revealed layers of bacteriaembedded in ECM. The morphology of the biofilmsformed on the glass surfaces was similar to that of thoseformed on Millipore filters, and those that had beenembedded and sectioned in Lowicryl.
Applying NTHi-specific antibodies to the sectionsconfirmed that NTHi, and not contaminating bacteria,were forming the biofilms. The anti-NTHi antibodieslabeled all the bacteria we examined, confirming thepresence of an axenic population of NTHi. The anti-body also detected NTHi proteins in Western blots,enabling the protein profiles reacting with the antibodyto be compared between those in the whole biofilmwith those in the ECM. Although slight differencesbetween the two protein preparations were present, theWestern blots confirmed the presence of a large numberof proteins that reacted with the anti-NTHi antibodiesin both the whole biofilm and in the ECM.
Anti-P6 OMP and anti-LOS antibodies both labeledthe NTHi biofilm sections. Expression of P6 OMP ispreserved in NTHi biofilms, and monoclonal anti-LOS6E4 labeling is increased when compared with plank-tonically grown NTHi (Murphy and Kirkham 2002).
The importance of LOS in biofilm formation byNTHi has been established in previous work (Greineret al. 2004; Swords et al. 2004). Examination of thedistribution of LOS in NTHi biofilms is therefore animportant step in understanding how biofilms areformed. From our labeling experiments, it appears thatthe bacteria secrete LOS over the substrate on which thebacteria have attached. At the base of the biofilm, theLOS is mostly extracellular and accumulates in the ECMat the base of the biofilm. LOS was also observed topenetrate into the supporting filter at the biofilm base.Throughout the biofilm, the LOS was also associatedwith NTHi membranes. Although LOS is important formaintaining biofilms as they develop, perhaps by main-taining biofilm architecture, the accumulation of LOS atthe biofilm base may suggest that it also plays a role inthe initial attachment of bacteria to substrates. LOS bind-ing to ear tissue may be an important pathogenesis factor(Demaria 1999). The possibility of LOS-based vaccinesbeing used in otitis media treatments (Gu et al. 2003)
suggests that the localization of LOS within NTHi bio-films will be important information for understandinghow such vaccines may work.
The presence and distribution of the P6 OMP, ahighly conserved protein in H. influenzae (Murphyet al. 1985,1986), is important because it is currentlybeing considered as a basis for a vaccine (Murphy et al.1986; Wu et al. 2005) and is able to stimulate mac-rophage and epithelial cells (Chen et al. 2004; Berensonet al. 2005). In the labeled biofilm sections, the anti-P6antibody was only associated with the membranes ofbacteria containing observable amounts of intracellularparticulate electron-opaque material. Previous studiesof cryo-fixed bacteria have demonstrated that electron-opaque particles in the cell cytoplasm are mostly ribo-somes (Hobot et al. 1985; Bohrmann and Kellenberger2001). Thus, a large number of these particles withinthe bacterial cell would indicate a high level of proteinsynthesis. The P6 OMP is known to be associated withviable bacteria, so it is possible that the non-labeledcells in the NTHi biofilms could represent a populationof either dormant or dead cells. Reduced amounts ofelectron-opaque material in the cell cytoplasm of non-labeled cells further suggests reduced metabolic levels.If dormant cells are present in NTHi biofilms and theydo not express the P6 OMP on their surface, thenvaccines targeting this molecule may not be effectiveagainst some of the bacteria present in the biofilm. Theycould then emerge from their dormant state ready to re-establish an infection.
Although the anti-P6 antibody label was confined tothe bacterial membranes, there was an association be-tween labeled membranes and the ECM. The anti-P6label was often associated with parts of the bacterialmembrane that were also in close apposition to theECM. The exact meaning of this association is cur-rently unknown. Immunization of animal models withpurified P6 OMP resulted in the production of NTHi-specific bactericidal antibodies but did not reduce theincidence of NTHi-induced otitis media (DeMaria et al.1996). The meaning of this result in the context ofNTHi biofilm infections is not clear, but it may suggestthat the P6 OMP may not play an important role in theestablishment of NTHi biofilms.
Of particular interest to uswhenwe first demonstratedan ability to preserve biofilm ultrastructure for sectioningand immunolabeling for electron microscopy was thelocalization of proteins involved in bacterial adhesion.Proteins such as the Hap protein and the HMW1/HMW2 proteins are adhesins known to play a role inbacterial attachment. We also examined the location ofthe NTHi IgA1 protease, because this protein has alsobeen implicated in bacterial microcolony formation.
The Hap protein belongs to the autotransporter fam-ily of proteins and is synthesized as a large precursorprotein with three functional domains. The N-terminal
TheJournal
ofHistoch
emistry&
Cytoch
emistry
Immunolabeling of NTHi Biofilms 839
by guest on July 2, 2014jhc.sagepub.comDownloaded from

signal sequence directs the protein from the cytoplasminto the periplasm, a C-terminal 45-kDa domain insertsinto theoutermembrane to formapore, and through thispore the 110-kDa N-terminal domain is translocated tothe cell surface. Through a final step called autoprote-olysis, the N-terminal domain, which includes a serineprotease active site, cleaves the precursor to release the110 kDa fragment from the cell surface (St Geme et al.1994; Hendrixson et al. 1997; Henderson et al. 1998).
The Hap protein has also been proposed as a po-tential candidate vaccine (Liu et al. 2004). It is involvedwith binding to human epithelial cells in vitro (St Gemeet al. 1994), binds to extracellular mammalian proteins(Fink et al. 2002), and promotes microcolony forma-tion of the bacteria (Hendrixson and St Geme 1998;Fink et al. 2003). These properties suggest that the Happrotein may play an important role in the process ofbiofilm formation.
In the TEM, the amount of specific anti-Hap proteinlabel we detected was low, considering the possible rolethis protein has in adherence. The specific signal wasdistributed between the bacteria and the ECM, withmore Hap protein present in the ECM than over theNTHi. Inasmuch as the passenger domain of Hap isreleased extracellularly by autoproteolysis, it is notsurprising to find that it is present in the ECM of thebiofilm. However, the ability of the Hap protein to bindbacteria to each other to form microcolonies dependson the passenger domain being present on the NTHimembrane surface (Fink et al. 2001).
The HMW proteins also mediate binding of NTHito macrophages and human epithelial cells (St Gemeet al. 1993; Noel et al. 1994). These proteins are lo-calized to the NTHi cell surface (Bakaletz and Baren-kamp 1994), and their interaction with glycosylatedproteins suggests that carbohydrate moieties within thebiofilm ECM may have an important role in biofilmformation. The HMW proteins on the cell surface mayhave the ability to bind to extracellular glycosylatedproteins to form complex cross-linking bridges withproteins on adjacent bacteria. The distribution of HMWproteins labeling over the NTHi and the ECM makes itpossible to speculate any number of roles for these pro-teins within the biofilm. Examination of HMW protein-deficient mutants for their biofilm-forming abilities willbe important if we are to assign any links between theHMW proteins and biofilms.
All human isolates of NTHi secrete IgA1 prote-ases (Plaut et al. 1992). They are autotransporters withsignificant sequence homology to the Hap protein (StGeme et al. 1994), and like the Hap protein, IgA1 pro-tease has been reported to mediate microcolony for-mation (Plaut et al. 1992). The patchy labeling of theanti-IgA1 protease antibody over the biofilm sectionssuggests that some parts of the biofilm, where the IgA1protease is present, may have properties that differ from
those where there is no labeling. Previous observationshave already suggested that some parts of the NTHibiofilms grown on support filters are more susceptibleto removal when exposed to aqueous medium (Websteret al. 2004). As yet, little is known about the charac-teristics of the parts of the biofilm that are not washedaway. However, it is possible that increased adhesionresulting from concentrations of adhesins may help toexplain the local differences found within the biofilms.
Although we examined the distribution of the IgA1protease within biofilms looking at the possibility thatthe protease had a role in bacterial adhesion, we arealso aware of the proposed role of this enzyme as aninactivator of serum and secretory forms of mammalianIgA antibodies (Kilian et al. 1979; Plaut et al. 1992).With this in mind, it is reasonable to speculate that theincreased levels of IgA1 protease in the top region of theNTHi biofilm may form a useful part of a defensemechanism for protecting against antibody attack.
In this study, we have demonstrated that fragile bio-film structures can be easily preserved and embedded inresin for subsequent sectioning and immunolabeling.Wehave examined biofilms formed in vitro by NTHi, anexclusively human pathogen that is a common cause ofotitis media, sinusitis, pneumonia, and chronic bron-chitis (Turk 1984). The increasing reports of biofilmformation by NTHi and the role of biofilms in diseasesuggest an important need to understand how bacteriaare able to establish biofilms on epithelial cell surfaces. Ifwe are to develop vaccines against bacterial proteins, wealso need to know the composition of biofilms, as well asthe subcellular distribution of antigens that are beingused in vaccine development. Only then will we be ableto fully appreciate the efficacy of potential vaccines.
Because this may be the first time that antibodies havebeen applied to sections through biofilms prepared usingcryo-methods, we believe that we have revealed newinformation on the distribution of NTHi proteins onbacteria in a biofilm and the association of these proteinswith ECM that has been shed by the bacteria.
The goals of our study were to determine whether itwas possible to preserve NTHi biofilms so that theycould be probed with antibodies at the ultrastructurallevel and to examine the distribution of antibodiesdirected to NTHi-specific proteins. In achieving thesegoals, we have obtained information on the distributionof NTHi proteins that are being considered as suitablecandidates for vaccine development and on their possibleaccessibility for antibody attack. We have also obtainedresults that may reveal some of the early stages of NTHibiofilm formation, and these may stimulate questionsabout how the bacteria are able to maintain the biofilmphenotype. These methods for preserving biofilmultrastructure for subsequent immunochemical analysiswill enable us to continue examining the composition ofNTHi biofilms. In addition, these methods may also be
TheJournal
ofHistoch
emistry&
Cytoch
emistry
840 Webster, Wu, Gomez, Apicella, Plaut, St. Geme
by guest on July 2, 2014jhc.sagepub.comDownloaded from

of value for preserving the ultrastructure of biofilmsformed by other bacteria, revealing important structuraland immunochemical information on biofilm formationin general.
Acknowledgments
This work was funded in part by the National Organiza-tion for Hearing Research (NOHR) and the DeafnessResearch Foundation (PW); National Institutes of HealthGrant NIH R01 DE-015844, and the core facilities of theCenter for Gastroenterology Research on Absorptive andSecretory Processes (GRASP), NIH Grant P30 DK-034928(AGP); and NIH Grant RO1AI AJ-44322 to J.W.S.G. G.G.was supported by a scholarship from the Hope for HearingFoundation of Los Angeles.
Literature Cited
Anderl JN, Franklin MJ, Stewart PS (2000) Role of antibioticpenetration limitation in Klebsiella pneumoniae biofilm resistanceto ampicillin and ciprofloxacin. Antimicrob Agents Chemother44:1818–1824
Anderl JN, Zahller J, Roe F, Stewart PS (2003) Role of nutrientlimitation and stationary-phase existence in Klebsiella pneumoniaebiofilm resistance to ampicillin and ciprofloxacin. AntimicrobAgents Chemother 47:1251–1256
Bakaletz LO, Barenkamp SJ (1994) Localization of high-molecular-weight adhesion proteins of nontypeable Haemophilus influenzaeby immunoelectron microscopy. Infect Immun 62:4460–4468
Barenkamp SJ, Bodor FF (1990) Development of serum bactericidalactivity following nontypable Haemophilus influenzae acute otitismedia. Pediatr Infect Dis J 9:333–339
Barenkamp SJ, Leininger E (1992) Cloning, expression, and DNAsequence analysis of genes encoding nontypeable Haemophilusinfluenzae high-molecular-weight surface-exposed proteins relatedto filamentous hemagglutinin of Bordetella pertussis. Infect Immun60:1302–1313
Berenson CS, Murphy TF, Wrona CT, Sethi S (2005) Outer mem-brane protein P6 of nontypeable Haemophilus influenzae is apotent and selective inducer of human macrophage proinflamma-tory cytokines. Infect Immun 73:2728–2735
Bluestone CD, Stephenson JS, Martin LM (1992) Ten-year review ofotitis media pathogens. Pediatr Infect Dis J 11:S7–S11
Bohrmann B, Kellenberger E (2001) Cryosubstitution of frozenbiological specimens in electron microscopy: use and applicationas an alternative to chemical fixation. Micron 32:11–19
Buscher AZ, Burmeister K, Barenkamp SJ, St Geme JW 3rd (2004)Evolutionary and functional relationships among the nontypeableHaemophilus influenzae HMW family of adhesins. J Bacteriol186:4209–4217
Campagnari AA, Spinola SM, Lesse AJ, Kwaik YA, Mandrell RE,Apicella MA (1990) Lipooligosaccharide epitopes shared amonggram-negative non-enteric mucosal pathogens. Microb Pathog 8:353–362
Chen R, Lim JH, Jono H, Gu XX, Kim YS, BasbaumCB,Murphy TF,et al. (2004) Nontypeable Haemophilus influenzae lipoprotein P6inducesMUC5ACmucin transcriptionviaTLR2–TAK1-dependentp38MAPK-AP1 and IKKbeta-IkappaBalpha-NF-kappaB signalingpathways. Biochem Biophys Res Commun 324:1087–1094
Costerton JW (1999) Introduction to biofilm. Int J Antimicrob Agents11:217–221
Cutter D, Mason KW, Howell AP, Fink DL, Green BA, St Geme JW3rd (2002) Immunization with Haemophilus influenzae Hap ad-hesin protects against nasopharyngeal colonization in experimen-tal mice. J Infect Dis 186:1115–1121
Davey ME, O’Toole GA (2000) Microbial biofilms: from ecology tomolecular genetics. Microbiol Mol Biol Rev 64:847–867
Demaria TF (1999) Localization of nontypeable Haemophilusinfluenzae endotoxin in the middle and inner ear during ex-perimental otitis media. Acta Otolaryngol 119:583–587
DeMaria TF, Murwin DM, Leake ER (1996) Immunization withouter membrane protein P6 from nontypeable Haemophilus in-fluenzae induces bactericidal antibody and affords protection inthe chinchilla model of otitis media. Infect Immun 64:5187–5192
Dingman JR, Rayner MG, Mishra S, Zhang Y, Ehrlich MD, Post JC,Ehrlich GD (1998) Correlation between presence of viable bacteriaand presence of endotoxin in middle-ear effusions. J Clin Micro-biol 36:3417–3419
Donlan RM, Costerton JW (2002) Biofilms: survival mechanismsof clinically relevant microorganisms. Clin Microbiol Rev 15:167–193
Faden H (2001) The microbiologic and immunologic basis for re-current otitis media in children. Eur J Pediatr 160:407–413
Fink DL, Buscher AZ, Green B, Fernsten P, St Geme JW 3rd (2003)The Haemophilus influenzae Hap autotransporter mediates mi-crocolony formation and adherence to epithelial cells and extra-cellular matrix via binding regions in the C-terminal end of thepassenger domain. Cell Microbiol 5:175–186
Fink DL, Cope LD, Hansen EJ, Geme JW 3rd (2001) TheHemophilusinfluenzae Hap autotransporter is a chymotrypsin clan serine pro-tease and undergoes autoproteolysis via an intermolecular mech-anism. J Biol Chem 276:39492–39500
Fink DL, Green BA, St Geme JW 3rd (2002) The Haemophilusinfluenzae Hap autotransporter binds to fibronectin, laminin, andcollagen IV. Infect Immun 70:4902–4907
Giebink GS (1989) The microbiology of otitis media. Pediatr InfectDis J 8(suppl 1):18–20
Gilmour A, Wilson AB, Fraser TW (1993) Microbial adherence tofood contact surfaces. In Denyer SP, Gorman SP, Sussman M, eds.Microbial Biofilms: Formation and Control. Oxford, UK, Black-well Scientific Publications, 293–313
Greiner LL, Watanabe H, Phillips NJ, Shao J, Morgan A, Zaleski A,Gibson BW, et al. (2004) Nontypeable Haemophilus influenzaestrain 2019 produces a biofilm containing N-acetylneuraminicacid that may mimic sialylated O-linked glycans. Infect Immun72:4249–4260
Griffiths G (1993) Fine Structure Immunocytochemistry. Heidelberg,Springer-Verlag
Gu XX, Rudy SF, Chu C, McCullagh L, Kim HN, Chen J, Li J, et al.(2003) Phase I study of a lipooligosaccharide-based conjugatevaccine against nontypeable Haemophilus influenzae. Vaccine 21:2107–2114
Gu XX, Tsai CM, Apicella MA, Lim DJ (1995) Quantitation andbiological properties of released and cell-bound lipooligosaccha-rides from nontypeable Haemophilus influenzae. Infect Immun 63:4115–4120
Henderson IR, Navarro-Garcia F, Nataro JP (1998) The great escape:structure and function of the autotransporter proteins. TrendsMicrobiol 6:370–378
Hendrixson DR, de la Morena ML, Stathopoulos C, St Geme JW3rd (1997) Structural determinants of processing and secretionof the Haemophilus influenzae hap protein. Mol Microbiol 26:505–518
HendrixsonDR, St Geme JW 3rd (1998) TheHaemophilus influenzaeHap serine protease promotes adherence and microcolony forma-tion, potentiated by a soluble host protein.Mol Cell 2:841–850
Hobot JA, Villiger W, Escaig J, Maeder M, Ryter A, Kellenberger E(1985) Shape and fine structure of nucleoids observed on sectionsof ultrarapidly frozen and cryosubstituted bacteria. J Bacteriol162:960–971
Hunter RC, Beveridge TJ (2005) Application of a pH-sensitivefluoroprobe (C-SNARF-4) for pH microenvironment analysis inPseudomonas aeruginosa biofilms. Appl Environ Microbiol 71:2501–2510
Ingvarsson L, Lundgren K, Ursing J (1982) The bacterial flora inthe nasopharynx in healthy children. Acta Otolaryngol Suppl 386:94–96
KilianM,Mestecky J, Schrohenloher RE (1979) Pathogenic species of
TheJournal
ofHistoch
emistry&
Cytoch
emistry
Immunolabeling of NTHi Biofilms 841
by guest on July 2, 2014jhc.sagepub.comDownloaded from

the genus Haemophilus and Streptococcus pneumoniae produceimmunoglobulin A1 protease. Infect Immun 26:143–149
Kilian M, Poulsen K, Lomholt H (2002) Evolution of the paralogoushap and iga genes in Haemophilus influenzae: evidence for aconserved hap pseudogene associated with microcolony formationin the recently diverged Haemophilus aegyptius and H. influenzaebiogroup aegyptius. Mol Microbiol 46:1367–1380
Klein JO (1997) Role of nontypeable Haemophilus influenzae in pe-diatric respiratory tract infections. Pediatr Infect Dis J 16(suppl 2):5–8
Kuklinska D, Kilian M (1984) Relative proportions of Haemophilusspecies in the throat of healthy children and adults. Eur J ClinMicrobiol 3:249–252
Liu DF, Mason KW, Mastri M, Pazirandeh M, Cutter D, Fink DL, StGeme JW 3rd, et al. (2004) The C-terminal fragment of the internal110-kilodalton passenger domain of the Hap protein of nontype-able Haemophilus influenzae is a potential vaccine candidate.Infect Immun 72:6961–6968
Lucocq J (1993) Unbiased 3-D quantitation of ultrastructure in cellbiology. Trends Cell Biol 3:354–358
Morphew MK, McIntosh JR (2003) The use of filter membranes forhigh-pressure freezing of cell monolayers. J Microsc 212:21–25
Murphy TF, Apicella MA (1987) Nontypable Haemophilus influen-zae: a review of clinical aspects, surface antigens, and the humanimmune response to infection. Rev Infect Dis 9:1–15
Murphy TF, Bartos LC, Rice PA, Nelson MB, Dudas KC, ApicellaMA (1986) Identification of a 16,600-dalton outer membrane pro-tein on nontypeable Haemophilus influenzae as a target for humanserum bactericidal antibody. J Clin Invest 78:1020–1027
Murphy TF, Kirkham C (2002) Biofilm formation by nontypeableHaemophilus influenzae: strain variability, outer membrane an-tigen expression and role of pili. BMC Microbiol 2:7
Murphy TF, Nelson MB, Dudas KC, Mylotte JM, Apicella MA(1985) Identification of a specific epitope of Haemophilus in-fluenzae on a 16,600-dalton outer membrane protein. J Infect Dis152:1300–1307
Neu T, Swerhone GD, Lawrence JR (2001) Assessment of lectin-binding analysis for in situ detection of glycoconjugates in biofilmsystems. Microbiol 147:299–313
Noel GJ, Barenkamp SJ, St Geme JW 3rd, Haining WN, Mosser DM(1994) High-molecular-weight surface-exposed proteins of Hae-mophilus influenzae mediate binding to macrophages. J Infect Dis169:425–429
Pichichero ME (2000) Recurrent and persistent otitis media. PediatrInfect Dis J 19:911–916
Plaut AG, Qiu J, Grundy F,Wright A (1992) Growth of Haemophilusinfluenzae in human milk: synthesis, distribution, and activity ofIgA protease as determined by study of iga1 andmutant iga2 cells.J Infect Dis 166:43–52
Poje G, Redford RJ (2003) General methods for culturing Haemo-philus influenzae. In Herbert MA, Hood DW, Moxon ER, eds.Haemophilus Influenzae Protocols. Totowa, New Jersey, HumanaPress, 51–56
Post JC (2001) Direct evidence of bacterial biofilms in otitis media.Laryngoscope 111:2083–2094
Post JC, Preston RA, Aul JJ, Larkins-Pettigrew M, Rydquist-White J,
Anderson KW, Wadowsky RM, et al. (1995) Molecular analysisof bacterial pathogens in otitis media with effusion. JAMA 273:1598–1604
Poulsen LK, Ballard G, Stahl DA (1993) Use of rRNA fluorescencein situ hybridization for measuring the activity of single cells inyoung and established biofilms. Appl Environ Microbiol 59:1354–1360
Rao VK, Krasan GP, Hendrixson DR, Dawid S, St Geme JW 3rd(1999) Molecular determinants of the pathogenesis of disease dueto non-typable Haemophilus influenzae. FEMS Microbiol Rev23:99–129
Rayner MG, Zhang Y, Gorry MC, Chen Y, Post JC, Ehrlich GD(1998) Evidence of bacterial metabolic activity in culture-negativeotitis media with effusion. JAMA 279:296–299
Sawaguchi A, McDonald KL, Karvar S, Forte JG (2002) A newapproach for high-pressure freezing of primary culture cells: thefine structure and stimulation-associated transformation of cul-tured rabbit gastric parietal cells. J Microsc 208:158–166
Sawaguchi A, Yao X, Forte JG, McDonald KL (2003) Direct attach-ment of cell suspensions to high-pressure freezing specimen plan-chettes. J Microsc 212:13–20
Spinola SM, Peacock J, Denny FW, Smith DL, Cannon JG (1986)Epidemiology of colonization by nontypable Haemophilus in-fluenzae in children: a longitudinal study. J Infect Dis 154:100–109
St Geme JW 3rd, de la Morena ML, Falkow S (1994) A Haemophilusinfluenzae IgA protease-like protein promotes intimate interactionwith human epithelial cells. Mol Microbiol 14:217–233
St Geme JW 3rd, Falkow S, Barenkamp SJ (1993) High-molecular-weight proteins of nontypable Haemophilus influenzae mediateattachment to human epithelial cells. Proc Natl Acad Sci USA90:2875–2879
Swords WE, Moore ML, Godzicki L, Bukofzer G, Mitten MJ,VonCannon J (2004) Sialylation of lipooligosaccharides promotesbiofilm formation by nontypeable Haemophilus influenzae. InfectImmun 72:106–113
Thurnheer T, Gmur R, Shapiro S, Guggenheim B (2003) Mass trans-port of macromolecules within an in vitro model of supragingivalplaque. Appl Environ Microbiol 69:1702–1709
Trottier S, Stenberg K, Svanborg-Eden C (1989) Turnover of non-typable Haemophilus influenzae in the nasopharynges of healthychildren. J Clin Microbiol 27:2175–2179
Turk DC (1984) The pathogenicity of Haemophilus influenzae. JMedMicrobiol 18:1–16
Watnick PI, Kolter R (1999) Steps in the development of a Vibriocholerae El Tor biofilm. Mol Microbiol 34:586–595
Webster P, Wu S, Webster S, Rich KA, McDonald K (2004) Ultra-structural preservation of biofilms formed by non-typeable Hemo-philus influenzae. Biofilms 1:165–182. Published online January24, 2005 (DOI: 10.1017/S1479050504001425)
Wu T, Chen J, Murphy TF, Green BA, Gu XX (2005) Investigationof nontypeable Haemophilus influenzae outer membrane proteinP6 as a new carrier for lipooligosaccharide conjugate vaccines.Vaccine 23:5177–5185
Zahller J, Stewart PS (2002) Transmission electron microscopic studyof antibiotic action on Klebsiella pneumoniae biofilm. AntimicrobAgents Chemother 46:2679–2683
TheJournal
ofHistoch
emistry&
Cytoch
emistry
842 Webster, Wu, Gomez, Apicella, Plaut, St. Geme
by guest on July 2, 2014jhc.sagepub.comDownloaded from




















