
Distinctive Features of the Gametes and Reproductive Tracts ...
15
BIOLOGY OF REPRODUCTION 50, 820-834 (1994) Distinctive Features of the Gametes and Reproductive Tracts of the Asian Musk Shrew, Suncus murinus J.M. BEDFORD, ' " 2 G.W. COOPER, 3 D.M. PHILLIPS, 4 and G.L. DRYDEN 5 Departments of Obstetrics and Gynecology and of Cell Biology and Anatomy 2 Cornell University Medical College, New York, New York 10021 Fertility Laboratory, 3 North Shore University Hospital, Manhasset, New York 11030 The Population Council, 4 The Rockefeller University, New York, New York 10021 Department of Biology, 5 Slippery Rock University, Slippery Rock, Pennsylvania 16057 ABSTRACT Insectivora are of special interest as the most primitive of the eutherian mammals, but essentially nothing is known of their gamete function. In this respect, the Asian musk shrew (Suncus murinus), investigated in the present study, displays many idiosyncrasies. In the epididymis, the giant acrosome undergoes further stabilization, its unusual resilience being especially evident in a "rim" created by a persistent close alignment of the outer acrosomal and overlying plasma membranes. However, until spermatozoa reached a gland on the vas deferens, no post-testicular change was demonstrable in the sperm head surface, the unusual nature of which was indicated by a dorso-ventral differentiation, by an inability to auto-agglutinate or to bind to the homologous zona pellucida, and by an insensitivity to anti-sperm immunoglobulin IgG in fresh serum. At mating, only about 1 X 106 spermatozoa are inseminated as far as the anterior vagina with plug formation. Within the small (6 mm) fallopian tube, the isthmus and ampulla are sharply delineated by their contractile activity and epithelial character; there is evidence of some sperm entry into isthmic crypts and a tendency for ipsilateral ovarian control of sperm transport to the tubal ampulla. The cumulus oophorus does not undergo preovulatory mucification and expansion, is characterized by persistent intercellular gap junctions, and is insensitive to hyaluronidase and trypsin. It is unclear how the compact cumulus is penetrated at fertilization. The giant acrosome contains acrosin and an unusually temperature-dependent' cumulytic activity; it is intact in motile ampullary sperma- tozoa but appears to be discharged before reaching the zona pellucida. Since eggs were not penetrated in the presence of ampullary spermatozoa until 4-10 h after ovulation, Suncus spermatozoa spend a long period in the female before they can fertilize. The determinants of sperm function, including capacitation and the acrosome reaction (AR), may depend on a different set of controls in Suncus, perhaps as a legacy of the resilient giant acrosome. This possibility could be examined in other Cro- ciduran and Soricine shrews selected according to acrosome size. INTRODUCTION Despite intensive research in selected species, there is much about gamete function in eutherian mammals that has yet to be explained. In addition to the variation in the form of spermatozoa, particularly evident among rodents, there is no consensus as to why the sperm head has evolved an unusual set of new characteristics, why the oocyte has a vestment of such formidable proportions, or why fertil- izing spermatozoa fuse by the equatorial region rather than the inner acrosomal membrane [1]. The exact function of the mammalian acrosome is another point that remains elu- sive, and this issue is complicated by variation in the size and shape of the acrosome. Its most striking development occurs in an insectivore, the Asian musk shrew, Suncus mu- rinus, whose acrosome is intriguing because of its great size [2-6], which outstrips that in related shrews and the few other insectivores studied in this regard [7-12]. Shrews and some other Insectivora are of particular interest as sur- viving links to the progenitors of modern eutherian mam- mals [13]. However, it is difficult to interpret the diversity Accepted November 16, 1993. Received August 31, 1993. 'Correspondence: J.M. Bedford, Box 30, Cornell University Medical College, 1300 York Avenue, New York, NY 10021. FAX: (212) 746-8589. among their acrosomes because there have been few stud- ies of gamete function in any insectivore. It is known that spermatozoa fertilize within the follicle in the tenrec [14- 16] and Northern short-tailed shrew [17]. Otherwise, insec- tivore gamete function and fertilization have been exam- ined only occasionally because of the rarity of species maintained in controlled circumstances. That Suncus mu- rinus is an exception in this respect has made it amenable to study of its reproduction [18-20] and of its unusual re- productive and behavioral endocrinology (e.g., [21, 22]). In an attempt to gain some insight into the significance of the unusually large acrosome in Suncus, we have further examined its spermatozoa: their maturation in the male, their transport in the female, and their interaction with eggs. From the perspective of what is known about the laboratory and domestic mammals, Suncus displays many unusual features in the preliminaries to fertilization. MATERIALS AND METHODS The animals were born in a colony of Suncus murinus from stock obtained originally from Guam [18] and were maintained one to a cage [23]. For mating, a proven stud was introduced into the female's cage until copulation with ejaculation occurred [18]. 820 Downloaded from https://academic.oup.com/biolreprod/article/50/4/820/2762587 by guest on 13 January 2022
-
Upload
khangminh22 -
Category
Documents
-
view
1 -
download
0
Transcript of Distinctive Features of the Gametes and Reproductive Tracts ...

BIOLOGY OF REPRODUCTION 50, 820-834 (1994)
Distinctive Features of the Gametes and Reproductive Tracts of the Asian Musk Shrew,Suncus murinus
J.M. BEDFORD, '"2 G.W. COOPER, 3 D.M. PHILLIPS,4 and G.L. DRYDEN5
Departments of Obstetrics and Gynecology and of Cell Biology and Anatomy2
Cornell University Medical College, New York, New York 10021Fertility Laboratory,3 North Shore University Hospital, Manhasset, New York 11030The Population Council,4 The Rockefeller University, New York, New York 10021
Department of Biology,5 Slippery Rock University, Slippery Rock, Pennsylvania 16057
ABSTRACT
Insectivora are of special interest as the most primitive of the eutherian mammals, but essentially nothing is known of their
gamete function. In this respect, the Asian musk shrew (Suncus murinus), investigated in the present study, displays manyidiosyncrasies. In the epididymis, the giant acrosome undergoes further stabilization, its unusual resilience being especially evident
in a "rim" created by a persistent close alignment of the outer acrosomal and overlying plasma membranes. However, untilspermatozoa reached a gland on the vas deferens, no post-testicular change was demonstrable in the sperm head surface, theunusual nature of which was indicated by a dorso-ventral differentiation, by an inability to auto-agglutinate or to bind to thehomologous zona pellucida, and by an insensitivity to anti-sperm immunoglobulin IgG in fresh serum. At mating, only about 1X 106 spermatozoa are inseminated as far as the anterior vagina with plug formation. Within the small (6 mm) fallopian tube,
the isthmus and ampulla are sharply delineated by their contractile activity and epithelial character; there is evidence of some
sperm entry into isthmic crypts and a tendency for ipsilateral ovarian control of sperm transport to the tubal ampulla. The cumulus
oophorus does not undergo preovulatory mucification and expansion, is characterized by persistent intercellular gap junctions,
and is insensitive to hyaluronidase and trypsin. It is unclear how the compact cumulus is penetrated at fertilization. The giant
acrosome contains acrosin and an unusually temperature-dependent' cumulytic activity; it is intact in motile ampullary sperma-
tozoa but appears to be discharged before reaching the zona pellucida. Since eggs were not penetrated in the presence of
ampullary spermatozoa until 4-10 h after ovulation, Suncus spermatozoa spend a long period in the female before they can
fertilize. The determinants of sperm function, including capacitation and the acrosome reaction (AR), may depend on a differentset of controls in Suncus, perhaps as a legacy of the resilient giant acrosome. This possibility could be examined in other Cro-
ciduran and Soricine shrews selected according to acrosome size.
INTRODUCTION
Despite intensive research in selected species, there ismuch about gamete function in eutherian mammals thathas yet to be explained. In addition to the variation in theform of spermatozoa, particularly evident among rodents,there is no consensus as to why the sperm head has evolvedan unusual set of new characteristics, why the oocyte hasa vestment of such formidable proportions, or why fertil-izing spermatozoa fuse by the equatorial region rather thanthe inner acrosomal membrane [1]. The exact function ofthe mammalian acrosome is another point that remains elu-sive, and this issue is complicated by variation in the sizeand shape of the acrosome. Its most striking developmentoccurs in an insectivore, the Asian musk shrew, Suncus mu-rinus, whose acrosome is intriguing because of its greatsize [2-6], which outstrips that in related shrews and thefew other insectivores studied in this regard [7-12]. Shrewsand some other Insectivora are of particular interest as sur-viving links to the progenitors of modern eutherian mam-mals [13]. However, it is difficult to interpret the diversity
Accepted November 16, 1993.Received August 31, 1993.'Correspondence: J.M. Bedford, Box 30, Cornell University Medical College, 1300
York Avenue, New York, NY 10021. FAX: (212) 746-8589.
among their acrosomes because there have been few stud-ies of gamete function in any insectivore. It is known thatspermatozoa fertilize within the follicle in the tenrec [14-16] and Northern short-tailed shrew [17]. Otherwise, insec-tivore gamete function and fertilization have been exam-ined only occasionally because of the rarity of speciesmaintained in controlled circumstances. That Suncus mu-rinus is an exception in this respect has made it amenableto study of its reproduction [18-20] and of its unusual re-productive and behavioral endocrinology (e.g., [21, 22]).
In an attempt to gain some insight into the significanceof the unusually large acrosome in Suncus, we have furtherexamined its spermatozoa: their maturation in the male, theirtransport in the female, and their interaction with eggs. Fromthe perspective of what is known about the laboratory anddomestic mammals, Suncus displays many unusual featuresin the preliminaries to fertilization.
MATERIALS AND METHODS
The animals were born in a colony of Suncus murinusfrom stock obtained originally from Guam [18] and weremaintained one to a cage [23]. For mating, a proven studwas introduced into the female's cage until copulation withejaculation occurred [18].
820
Dow
nloaded from https://academ
ic.oup.com/biolreprod/article/50/4/820/2762587 by guest on 13 January 2022

UNUSUAL GAMETE FEATURES IN THE ASIAN MUSK SHREW
Male
Adult males were anesthetized with sodium pentobar-bital (0.6 mg/10 g BW, s.c., Nembutal; Abbott, Chicago, IL)or were killed with ether. Testes, epididymides, and vasadeferentia were removed, and pieces were fixed in Bouin'ssolution for light microscopy or in 2% glutaraldehyde in0.1 M phosphate buffer (pH 7.2) containing 0.5% NaCl, fol-lowed by 1% phosphate-buffered osmium tetroxide, fortransmission electron microscopy (TEM).
Spermatozoa were released from different points alongthe epididymis and vas deferens into M199 (Gibco, GrandIsland, NY) or a modified Tyrode's solution (TALP [24]), at370C, and counted with an Improved Neubauer (VWR Sci-entific, Piscataway, NJ) hemacytometer. Movement patternsof spermatozoa from different levels of the tract were stud-ied in TALP on heated glass slides. To test the ability ofsperm extracts to disperse cumulus oophorus, Suncus ep-ididymal spermatozoa and also washed ejaculated (control)rabbit spermatozoa were homogenized and sonicated inM199 at a final concentration of about 1 x 106 in 20 pil ofsonicate solution, followed by centrifugation at 800 x g for10 min. Each supernatant was incubated with cumulus at23 0C or 37 0C (a total of 28 mouse cumuli and 14 rabbitcumuli obtained from gonadotropin-stimulated females).
For studies of their stability, spermatozoa from differentregions of the epididymis were exposed to dithiothreitol(DTT) at 0.31 mg/ml in 0.05 M borate buffer (pH 9.0) and/or to 0.01-0.5% SDS [25]. In one series, a mixture of Sun-cus spermatozoa from the cauda epididymidis and an equalnumber of ejaculated rabbit spermatozoa was suspendedin either 0.01%, 0.025%, or 0.05% SDS for 5 min beforefixation for TEM.
Spermatozoa recovered from the cauda epididymidis inM199 at 37°C were also tested for their fertilizing ability.Females given 50 IU of hCG (Nutritional Biochemicals Corp.,Cleveland, OH) i.p. were inseminated 3-6 h later througha 30-gauge needle via laparotomy into the lower uterus un-der sodium pentobarbital anesthesia (2.0 mg i.p.). Eggs wererecovered from the oviducts about 27-30 h after the hCGinjection and assessed on slides beneath a wax spot-sup-ported cover glass. To examine the role of testicular an-drogen in sperm storage in the cauda epididymidis, underether anesthesia the cauda was ligated with 6-0 silk; thiswas followed by 1) unilateral castration, 2) bilateral castra-tion, or 3) bilateral castration plus testosterone. Sperma-tozoa released from the cauda into Tyrode's solution at 370Cat different times thereafter were assessed immediately fortheir motility.
Female
Ovulation is sporadic at best after one coital episode,particularly in virgin females [24]. However, Suncus ovu-lates in response to a variety of gonadotropins [26], andhere ovulation was induced by i.p. injection of 0.5 mg FSH
+ 25 IU hCG (Nutritional Biochemicals Corp.) or, morecommonly, 50 IU hCG alone.
To examine the number of spermatozoa ejaculated andtheir transport and viability in the female, the vagina, cer-vix, uterus, and tubal isthmus and ampulla were each iso-lated as discrete segments and squeezed into TALP at 37°C;then the epithelium was gently scraped. Regional oviductcontractions were studied on warm slides in TALP beneatha wax spot-supported cover glass. Pieces of oviduct and ovaryas well as whole eggs were also immediately fixed for TEMas described above.
Eggs were extruded from the ampulla or from the peri-ovarian bursa into M199 or TALP at 37 0C at various timesup to 61 h postcoitus (p.c.); and in some instances, eggswere immediately mounted on wax-spot slides and exam-ined with a phase-contrast microscope for the state and ac-tivity of any spermatozoa that were present in the fluid orenmeshed in the cumulus. Those and the other eggs werethen freed from the cumulus with fine needles, assessedfor fertilization, and assessed again after fixation in aceticalcohol and staining with 1% lacmoid in 45% acetic acid.To study the response of the cumulus oophorus to knownenzymes, unfertilized oocytes collected in M199 about 21 hafter hCG were mounted on slides and either 1% bovinetesticular hyaluronidase (type VI; Sigma, St. Louis, MO), 0.5%pancreatic trypsin (type III; Sigma), or both together wereflooded beneath the cover glass at 23°C and 37°C.
RESULTS
Male TractVas deferens gland. The configuration and epithelial
character of the Suncus Wolffian duct are conventional ex-cept that the vas deferens is invested by a large discretegland (Fig. 1), mentioned also by Kitoh et al. [27]. The glandis composed of alveoli (Fig. 1; inset) that open into the vasdeferens by way of narrow apertures (not shown). In 10rested adult Suncus males (including 5 reported by Bed-ford et al. [281), each tract housed between 2.5 and 3.42million spermatozoa, of which about 0.5 million were inthe segment enveloped by the vas gland.
Characteristics of Suncus spermatozoa. Previous stud-ies have revealed in the sperm of this species 1) a giantacrosome (Fig. 2), 2) an unusual coxcomb-like perforato-rium [5], and 3) a dorso-ventral asymmetry of the head sur-face properties [3]; the previous studies 4) have failed todemonstrate any change in that surface during epididymalmaturation [3]. The present observations reveal that theSuncus acrosome has an unusual stability that may favor themaintenance of its erect form, with a particularly stable "rim"revealed in both degenerating (Fig. 3) and detergent-treatedspermatozoa (Figs. 4-6). The degenerating head illustratesthe unusually close relationship between the peri-acroso-mal plasma membrane and the outer acrosomal mem-brane, displayed at all stages after spermiation. The re-
821
Dow
nloaded from https://academ
ic.oup.com/biolreprod/article/50/4/820/2762587 by guest on 13 January 2022

BEDFORD ET AL.
FIG. 1. The male reproductive tract of Suncus. Note the large vas gland (G), beyond which the vas deferenscontinues to join the urethra. x9.5. Inset: histological section of vas deferens lumen (V) and surrounding alveoli (A)within the vas gland. Toludine blue. x50.
spective responses of the acrosome in mature rabbit andSuncus spermatozoa exposed together to 0.01-0.05% SDSalso illustrate well its relative resistance in Suncus. In suchmixed preparations, the rabbit acrosome essentially dis-appears in 0.05% SDS, whereas that of Suncus remains asa distorted but intact structure (Fig. 4).
Measurement of acrosin activity in Suncus spermatozoawas performed by Drs. C.R. Brown and E.F. Hartree (un-published results) [29]. After disruptive vortexing and cen-trifugation of Suncus spermatozoa at 16 000 x g for 15 min,the supernatant was found to contain a soluble inhibitor ofacrosin; noninhibited acrosin activity remained associatedwith the residue. The molar ratio of inhibitor/acrosin was> 1.0. Thus, the acrosome contains an excess of (soluble)inhibitor over acrosin, as in boar spermatozoa (E.F. Har-tree, personal communication). The acrosin value found forSuncus was at least 1.7 mU/107 spermatozoa, significantlymore than in the mouse [30] and equal to that in ram sper-matozoa [29], but small relative to the size of the acrosome.
The cumulus-dispersing activity of the acrosome is be-lieved to reflect, to a major degree, the action of hyaluron-idase. In order to test for this activity, the ability of the Sun-cus sperm sonicate to disperse the mouse and rabbit cumuluswas compared with that of an equivalent sonicate of rabbitspermatozoa. At 230C, rabbit sperm sonicate dispersed 55%
of mouse cumuli by 6 min and > 90% by 12 min, anddispersal was not detectably faster in replicates conductedat 370C. Similarly, all rabbit cumuli dispersed completelyby 8 min at 230C in the rabbit sperm sonicate. By contrast,the cumulus-dispersing ability of Suncus sperm sonicate wastemperature-dependent. Mouse and rabbit cumuli in Sun-cus sperm sonicate barely changed after 55 min at 230C,whereas at 37 0C a significant dispersal was evident after 12min and dispersal was complete by 20-25 min.
Suncus spermatozoa did not display the surface char-acteristics typical of many other eutherian spermatozoa.Whereas washed spermatozoa of many eutherian speciesauto-agglutinate by the acrosome region in M199 or in Ty-rode's solution containing protein (e.g., BSA or 10% serum),active Suncus spermatozoa from the cauda epididymidis orvas gland displayed no such tendency. More unusual, mo-tile Suncus spermatozoa did not adhere to the Suncus zonapellucida, although washed ejaculated rabbit spermatozoabound promptly to it as they do to other eutherian eggs.
Finally, Suncus epididymal spermatozoa were relativelyinsensitive to the complement-dependent anti-sperm im-munoglobulin IgG in serum [31,32], which disrupts thesperm plasmalemma in other species [36]. They swam vig-orously for some minutes in fresh rabbit serum and in freshSuncus serum, which immediately immobilized rabbitspermatozoa unless heated to 560C for 30 min.
822
Dow
nloaded from https://academ
ic.oup.com/biolreprod/article/50/4/820/2762587 by guest on 13 January 2022

UNUSUAL GAMETE FEATURES IN THE ASIAN MUSK SHREW
Sperm maturation and storage in the epididymis. InSuncus, one face of the caput sperm head differs from thatof the other as reflected in the binding pattern of positivelycharged ferric oxide colloid particles; there is no changein this pattern, or in that created by the lectins FITC-Con Aor FITC-WGA, in spermatozoa from successive regions ofthe epididymis [3]. Neither is the morphology of the Sun-cus acrosome modified as it passes through the epididymis.On the other hand, both the structural quality and capacityof Suncus spermatozoa for motility are enhanced duringepididymal passage.
According to the response to SDS and SDS/DTT [25], theunion of the sperm head and tail, the nucleus, and to alesser degree the Suncus acrosome are all stabilized to avariable extent by the formation of -S-S- bonds during ep-ididymal passage. The caput sperm nucleus was signifi-cantly disrupted by exposure to 0.01% SDS (Fig. 5), whereasthat of cauda spermatozoa was visibly unchanged (Fig. 6)and required the addition of DTT for decondensation. Intesticular and caput spermatozoa subjected to 0.5% SDS,some tails disintegrated, whereas in cauda spermatozoa theirdense fibers and mitochondrial and mainpiece sheaths re-mained intact-changes reminiscent of sperm tail matura-tion in other mammals. In more dilute (0.1%) SDS, the ac-rosomes of caput spermatozoa disappeared or remainedvisible only as faint detached shrouds in the phase-contrastmicroscope, whereas those of cauda spermatozoa usuallyremained as distinct if distorted structures. However, eventhe acrosomes of cauda spermatozoa dispersed if DTT wasadded to the SDS, suggesting that epididymal passage bringssome -S-S- stabilization of the acrosome.
With regard to the maturation of motility, 80-85% of ca-put spermatozoa displayed no motility in TALP at 370C; 10-15% displayed only sluggish movements; and some 3%maintained a vigorous tail beat (3 males). By contrast, about90% of the cauda spermatozoa of each male beat vigorouslyin the same conditions, though most soon adhered to theglass and so did not progress. Some 90% of the sperma-tozoa released from the vas gland (Fig. 1) were similarlymotile but did not adhere to the glass, and so achieved aprogressive space-gaining movement on warm glass slides-an indication that their surface changes in some way afterspermatozoa enter the vas gland.
PLATE .FIG. 2. Suncus spermatozoa released from the vas deferens gland. Note
the enormous fan-shaped acrosome. Phase contrast. x710.FIG. 3. Thin section of the head of a degenerating Suncus sperma-
tozoon lying in the vas deferens. Note the unusually close stable relation-ship (defined by arrowheads) between the outer acrosome membrane andthe peri-acrosomal region of the plasma membrane. This relationship ismaintained despite the swelling and dispersal of the content of the acro-some as well as the loss of plasma membrane posterior to the acrosome.The discrete equatorial segment (E) has moved forward out of the nuclearrecess or "waist" in which it normally resides. TEM. x30 000.
823
Dow
nloaded from https://academ
ic.oup.com/biolreprod/article/50/4/820/2762587 by guest on 13 January 2022

BEDFORD ET AL.
FIG. 4. Suncus spermatozoa from the cauda epididymidis mixed 1:1 with ejaculated rabbit spermatozoa and suspended in 0.05% SDS for 5 min beforeglutaraldehyde/osmium fixation. Whereas the rabbit acrosomes (arrowheads) are destroyed, those of Suncus (S) are only moderately distorted by thistreatment. The dense-fiber configuration in the sperm tail midpiece of the rabbit (r) differs from that of Suncus (s). TEM. x18 000.
To ascertain whether the sperm surface change inferredto occur in the gland segment is important for fertilizingability, in three experiments approximately 50000 caudaspermatozoa were inseminated into each uterine horn 3-6 h after i.p. injection of hCG. About 26 h after insemination(14-15 h after ovulation), 3 of 3 eggs recovered from eachof two females had either a swelling sperm head and a spermtail within the cytoplasm or had two pronuclei. Thus, mi-gration of Suncus spermatozoa from the cauda to the vasgland is not essential for development of their fertilizingability per se.
That a majority of epididymal spermatozoa are housedin a bulbous cauda [28] implies a storage role for this re-gion in Suncus. However, the motility among populationsreleased from the ligated cauda 10 days (52% and 39%),16 days (43%), 19 days (a weaker 39%), and 21 days (aweaker 47%) after castration was not obviously differentfrom that in the ligated cauda of intact males. These limitedexperiments suggest that sperm viability in the cauda maybe less dependent on testicular androgens in Suncus thanin other mammals.
Female Tract
Sperm distribution after mating. Four females exam-ined within 10 min of a single ejaculation by a rested maleeach had a vaginal plug and had, respectively, 1.0, 1.9, 0.6,
and 0.5 million spermatozoa in the anterior vaginal pocket,but none yet in the uterine horns or oviducts. Vaginal plugswere absent in each of four females examined at 4-5 h p.c.,when, according to the individual, 10000-40 000 motilespermatozoa were recovered from the vagina and fewer than5000 were flushed from each uterine horn. In six femalesexamined at or after ovulation, plugs were absent and sper-matozoa (immotile without acrosomes) were observed onlyrarely in the uterine or vaginal flush.
Eggs were collected from 21 hCG-stimulated mated fe-males at various times up to 36 h after ovulation. In twofemales examined before ovulation at 14 h after mating,moving spermatozoa were evident in transmitted light onlywithin the isthmus; but in females examined at various timesafter ovulation, there were also small numbers moving inthe ampulla or in recovered cumuli immediately aftermounting on warm slides. Two mated females analyzed at16.5 h and 18.25 h after hCG, respectively, had each ovu-lated three ova on the left side only, and these were stillunpenetrated even though 25-30 motile spermatozoa wereevident through the wall of each ipsilateral ampulla. Sig-nificantly, there were no spermatozoa in the ampulla of thecontralateral (nonovulating) side, notwithstanding the pres-ence of motile spermatozoa in each associated isthmus. Thisindication that sperm transport to the ampulla is regulatedto some extent by the ovulating ovary was seen in two ad-
824
Dow
nloaded from https://academ
ic.oup.com/biolreprod/article/50/4/820/2762587 by guest on 13 January 2022

UNUSUAL GAMETE FEATURES IN THE ASIAN MUSK SHREW
ditional females, 18-20 h after hCG, that had 6-8 intact mo-tile ampullary spermatozoa on the ovulating side but onlyone in each contralateral ampulla. Among the females re-maining, not more than 6 motile spermatozoa were re-covered per ampulla in the peri-fertilization period, and inmost there were no more than one or two regardless ofthe time after ovulation. Significantly, all motile spermato-zoa had an intact acrosome (cf. spermatozoa within the cu-mulus), whereas those without an acrosome were alwaysimmotile.
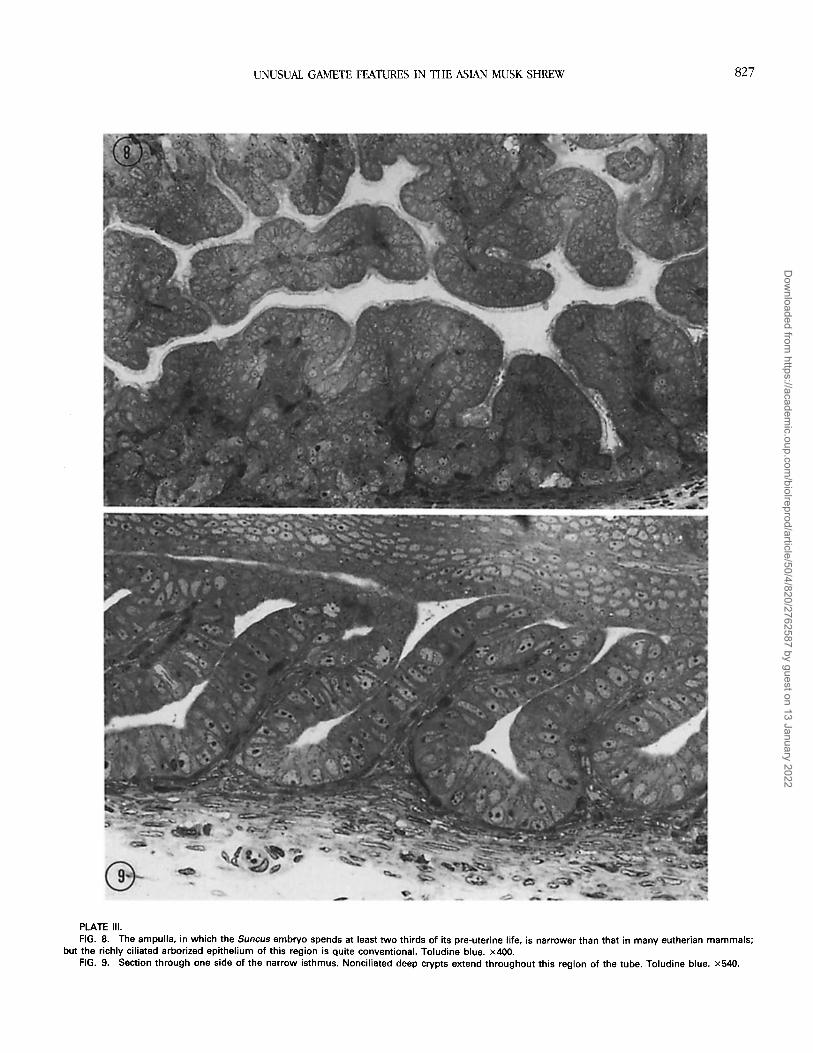
Fallopian tube. The fallopian tube is about 6 mm inlength, the ampulla and isthmus displaying abrupt differ-ences (Figs. 7-9) demarcated sharply at the isthmo-ampul-lary junction. The ampulla has a wider lumen and a richlyciliated epithelium (Fig. 8), and in the postovulatory periodit contracted regularly. The isthmus was distinguished byan absence of visible contractions, by an absence of cilia,and by deep crypts throughout its length (Figs. 7 and 9)that sometimes contained spermatozoa in histological sec-tions.
Ovulation. In accord with observations of Clenendonand Rissman [22], ovulation was very inconsistent after onemating. Among 20 animals mated once and examined be-tween 15.5 and 47 h p.c., only seven had ovulated (71%unilaterally), the earliest at 22 h p.c.; 2.28 + 0.26 (SEM)eggs were released per individual. Ovulation was inducedconsistently, however, after i.p. hCG with or without mat-ing. In that situation, 0 of 5 females had ovulated by 13-14h after hCG and mating, the earliest being observed at 15.75h after hCG. Among 66 additional females examined at 15.5h or more after hCG, with or without mating, 65 that ovu-lated produced 2.75 0.09 eggs per female (62% unilat-erally) with a maximum number of four eggs per ovary.Ova were in the peri-ovarian bursa or in the upper ampullauntil about 36 h after ovulation, when they were found atthe isthmo-ampullary junction.
Cumulus oophorus. In Suncus as in some other in-sectivores [15], no significant follicular antrum is formedand the cumulus fails to expand before ovulation; it is re-leased as a compact ball and remains as a separate entityin the oviduct. Unlike that of other animals, the cumulusin Suncus did not disperse or change within 30 min in M199containing 1% hyaluronidase at 37°C (Fig. 10). After transferto hyaluronidase + 0.5% pancreatic trypsin, the outline ofthe zona pellucida had disappeared after 15 min (Fig. 11),
4PLATE II.FIG. 5. Anterior region of the head of a Suncus spermatozoon from
the caput epididymidis, treated with 0.01% SDS before fixation. The nu-cleus is partially dissociated, but although the acrosome content has dis-persed, a stable rim is evident even in immature spermatozoa. TEM. x28 000.
FIG. 6. Head of a Suncus spermatozoon from the cauda epididymidistreated with 0.01% SDS. In contrast to the reaction of caput spermatozoaas seen in Figure 5, the more stable nucleus is visibly unchanged and muchof the acrosome content is retained. TEM. x28 000.
825
Dow
nloaded from https://academ
ic.oup.com/biolreprod/article/50/4/820/2762587 by guest on 13 January 2022

BEDFORD ET AL.
but again without any visible change in the cumulus ooph-orus.
The hyaluronidase-resistant character of the Suncus cu-mulus appears to be due not only to a paucity of proteo-glycan matrix but, probably more importantly, to multiplejunctions between neighboring cumulus cells (Fig. 12). Theseappear to be gap junctions as evidenced by their appear-ance in the TEM and in freeze-fracture samples studied byDr. Fumie Suzuki of Chiba University (personal commu-nication). As a correlate of its stable character, the cumuluspersisted around fertilized eggs 12 h after ovulation, al-though not around two-cell eggs 20-25 h after ovulation.Since the cumulus remained around 8 of 12 unfertilizedeggs recovered from 5 unmated females some 24 h afterhCG-induced ovulation (although not at 42 h postovulationin other unfertilized eggs), fertilization itself must hastenthe discharge of the Suncus cumulus. Ultimately, the eggemerges from a cumulus mass in which some intercellularjunctions are still evident.
Oocyte. The Suncus oocyte is invested by a zona pel-lucida that displays a subtle radial striation of its matrix inthe unfertilized egg [10] and is ovulated at metaphase II.Soon after ovulation, 18-20 h after the hCG injection, un-fertilized ova had one polar body (PB); somewhat later, de-condensation of the fertilizing sperm head was broadlycoincident with extrusion of a second PB (Fig. 17). How-ever, many unfertilized eggs collected then had an intactspindle but no PB, or one degenerating PB was sometimesevident in these and in recently penetrated eggs (Fig. 16).
Fertilization. The timing of sperm penetration afternatural mating, and especially after intrauterine sperm in-stillation, suggests that epididymal and ejaculated sperma-tozoa of Suncus cannot fertilize for many hours after in-semination. As mentioned earlier, motile spermatozoa fromthe cauda or vas deferens gland displayed no affinity forthe Suncus zona pellucida at 37°C. In addition, althoughspermatozoa were seen in the ampulla 16.5 h after hCG,penetration began only about 4 h or more later (Table 1).Similarly, when intrauterine insemination was performed3-6 h after administration of hCG, eggs at about 14-14 hafter ovulation (or about 24 h after insemination) were onlyrecently penetrated as evidenced by a swelling sperm heador early pronucleus development.
In 11 oviducts examined for spermatozoa between 16.5and 26 h after hCG, most dead spermatozoa had lost theacrosome, whereas the acrosome invariably remained in-tact in motile ampullary spermatozoa and was intact in onemotile cell attached to the cumulus surface. Despite the easewith which the Suncus acrosome can be monitored in thephase-contrast microscope, the small number of sperma-
FIG. 7. Fallopian tube of Suncus. The narrow isthmus (Isth.) is char-acterized by crypts and is abruptly demarcated (arrowheads) from the am-pulla (Amp.) Phase contrast. x40.
826
Dow
nloaded from https://academ
ic.oup.com/biolreprod/article/50/4/820/2762587 by guest on 13 January 2022
UNUSUAL GAMETE FEATURES IN THE ASIAN MUSK SHREW
PLATE 111.FIG. 8. The ampulla, in which the Suncus embryo spends at least two thirds of its pre-uterine life, is narrower than that in many eutherian mammals;
but the richly ciliated arborized epithelium of this region is quite conventional. Toludine blue. x400.FIG. 9. Section through one side of the narrow isthmus. Nonciliated deep crypts extend throughout this region of the tube. Toludine blue. x540.
827
Dow
nloaded from https://academ
ic.oup.com/biolreprod/article/50/4/820/2762587 by guest on 13 January 2022

828 BEDFORD ET AL.
Dow
nloaded from https://academ
ic.oup.com/biolreprod/article/50/4/820/2762587 by guest on 13 January 2022

UNUSUAL GAMETE FEAqTRES IN THE ASIAN MUSK SHREW
TABLE 1. Fertilization rate in Suncus following one ejaculation, as function of time after ovulation.
Hours afterhCG* Number of animals Eggs fertilized Comment
18-19 h 4 1/12 Sperm in perivitelline space
20-22 h 3 2/11 Swelling sperm head
23-24 h 7 12/20 Decondensing sperm heads-tailsprotrude from zona in 4 eggs
28-30 h 5 16/17 1 swelling head; 1 syngamy; rest 2pronuclei; most eggs with one po-lar body
36 h 2 4/4 2 cell; 1 polar body; no cumulus*Ovulation assumed to occur at approximately 15.5 h after hCG treatment.
tozoa and the density of the cumulus made it difficult todefine precisely the acrosomal discharge that precedes zonapenetration. Spermatozoa were detected by their move-ments within eight separate cumuli, and in two it was pos-sible to see that the acrosome was absent. Among eight otherspermatozoa observed within the cumulus in the TEM, in-cluding two sequestered by cumulus cells and six that hadpassed through the cumulus into a peri-zona space (Figs.13 and 14), all had lost the acrosome. The acrosome wasabsent also from three motile spermatozoa found adheringto the zona of penetrated eggs immediately after dissectionfrom cumulus (Fig. 15), and in no instance were acrosomalremnants seen on the zona of other fertilized eggs. Theselimited observations point to a discharge of the acrosomeafter the Suncus spermatozoon begins to interact with thecumulus but well before it binds to the zona pellucida.
Fertilization itself appears similar to that in other euthe-rian mammals. The spermatozoon creates a conventionalslit as it passes through the zona (Fig. 16), and a sperma-tozoon without an acrosome was seen moving vigorouslywithin such a slit. After sperm/egg fusion, the oocyte in-corporates the head as well as the large tail of the fertilizingspermatozoon (Fig. 17); but the fertilizing sperm tail stillprotruded prominently from the zona surface in four eggsin which incorporated sperm heads were decondensing. Thisimplies some, perhaps rotatory, activity on the part of theegg ultimately in drawing in the whole sperm tail. Two fer-tilized eggs at the two-cell stage presented the unusual pic-
PLATE IV.FIGS. 10 and 11. Suncus oocyte/cumulus complex recovered from the
ampulla 24 h after administration of hCG. Photograph in Figure 10 wastaken about 30 min after incubation in 1% hyaluronidase at pH 7.2 and37°C. In Figure 11, the same egg was photographed 15 min after furtherexposure to 1% hyaluronidase + 1% trypsin, during which the zona pel-lucida (z) then disappeared without visible change in the cumulus oopho-rus. Phase contrast. x275.
FIG. 12. Ultrastructure of midregion of Suncus cumulus oophorus re-covered about 22 h after hCG injection. Slight fixation shrinkage has cre-ated small intercellular spaces, serving to emphasize the multiple intercel-lular junctions, some of which are marked by arrowheads. TEM. x9000.
ture of an incorporated sperm tail extending between theirblastomeres.
DISCUSSION
The preliminaries to fertilization in eutherian mammalsgenerally involve a sequence of sperm plasma membranechanges in the epididymis relating to the fertilizing abilityof spermatozoa and to storage in the cauda; a release fromthat stable state embodied in the process of sperm capac-itation; passage of the fertilizing spermatozoa intact throughcumulus oophorus; binding to the zona via specific ligandson the peri-acrosomal plasma membrane; and finally, anacrosome reaction (AR) characterized by point fusions be-tween plasma membrane and outer acrosomal membrane.The musk shrew presents unusual features at almost everystep of these preliminaries. Anomalies are expressed in thematuration and storage of spermatozoa, low number ofspermatozoa, and surface properties of spermatozoa; in thestructure of the acrosome and its interaction with the eggcoats; and in the highly compartmentalized fallopian tubeand compact cumulus oophorus. However, since shrews arenotable not only as relatively primitive eutherian mammals,but also by their virtual absence from the current literaturein gamete biology, it is difficult to know yet the extent towhich these variant features in Suncus apply to other eu-therians, as well as to understand their significance.
SpermatozoaInsectivore spermatozoa are characterized often by group
features that include an unusually extended acrosome, aserrate perforatorium with an organized matrix, and asym-metrical dense fibers in the tail. In that regard, spermatozoaof elephant shrews [33] and their oocyte/cumulus complexand CL [34] give some credence to the old idea that ele-phant shrews have links to insectivores, as against the re-cent interpretation of their affinity with subprimates [35]. Inthe case of Suncus, however, not only does the sperm headsurface express an unusual dorso-ventral asymmetry andapparently fail to change in the epididymis [3], but (unlike
829
Dow
nloaded from https://academ
ic.oup.com/biolreprod/article/50/4/820/2762587 by guest on 13 January 2022

BEDFORD ET AL.
PLATE V.FIGS. 13 and 14. Longitudinal and transverse sections through the acrosomal region of Suncus sperm heads located between the inner cumulus and
the zona pellucida of recently fertilized eggs. As in all among the few spermatozoa seen within the limits of the cumulus, the acrosome has been discarded.TEM. Figure 13, x36000. Figure 14, x26500.
FIG. 15. Unfixed egg recovered from the ampulla of a mated female 26 h after hCG. Immediate removal of cumulus with fine needles revealed amotile spermatozoon without any acrosomal carapace adherent to the zona pellucida. Phase contrast. x720.
FIG. 16. Sperm penetration slit in the Suncus zona pellucida. Note the fertilizing sperm tail cross section and an already degenerate first PB (1p) withinthis yet nonactivated egg. TEM. x3200.
FIG. 17. Penetrated Suncus egg recovered about 23 h after mating. Decondensation of the sperm head is occurring coincidentally with extrusion ofthe second PB (2p). The PB is still connected to the female chromosomes (c) by spindle fibers that are just out of focus. Acetic alcohol fixation + lacmoidstain. Phase contrast. x740.
that in most Eutheria) it also shows no affinity for the ho-mologous zona pellucida under conditions in which evenrabbit spermatozoa readily adhere to the Suncus zona non-specifically. Nor does the plasma membrane of Suncusspermatozoa seem vulnerable to the spermicidal anti-spermIgG present in fresh serum [31,32], which, in the case ofother spermatozoa, immediately disrupts the plasmalemma[36].
The possibility that these surface properties of Suncusspermatozoa are linked to the unusual stability of the ac-rosome could perhaps be resolved by studying other shrews.The relatively modest proportions of Sorex spermatozoaseem to represent a more general type [6], and the un-precedented dimensions and resilience of the Suncus ac-rosome-especially its rim (Figs. 2-6)-make it the ex-treme example of the insectivore "mold." Thus, the
830
Dow
nloaded from https://academ
ic.oup.com/biolreprod/article/50/4/820/2762587 by guest on 13 January 2022

UNUSUAL GAMETE FEATURES IN THE ASIAN MUSK SHREW
Insectivora may fall into two groups in this respect. Thoseobserved to have modest acrosomes-European mole, Eu-ropean hedgehog, European common shrew, Japanesecommon shrew, Japanese water shrew (and elephantshrews)-all display the loose relationship between the peri-acrosomal plasma membrane and outer acrosomal mem-brane that is typical of most eutherian spermatozoa. Bycontrast, the relatively enormous acrosome of Crociduradsinezumi also manifests the tight relationship between thesemembranes seen in Suncus (T. Mori, personal communi-cation; [12]). Such a grouping may allow experimental ver-ification of the possibility that unusual preliminaries to fer-tilization in Suncus reflect the size and special nature ofthe acrosome.
The Suncus acrosome contains a level of acrosin com-parable only to that in ram spermatozoa [29], and hyal-uronidase may well be responsible for its unusually tem-perature-dependent cumulus-dispersing activity. However,in mammals generally and in Suncus, certainly, it is difficultto grasp the significance of hyaluronidase. The spermato-zoa of one marsupial that has no cumulus possess a higherconcentration of hyaluronidase than rabbit spermatozoa [37];and in the mouse [38], hamster [39], and human [40, 41], atleast, a release of acrosomal hyaluronidase is not requiredfor rapid cumulus penetration. Finally, the cumulus of Sun-cus is not responsive to hyaluronidase (Figs. 10 and 11).
Sperm Maturation and StorageAlthough Suncus spermatozoa develop the capacity for
progressive motility, and an -S-S- mediated stabilization ofsperm head and tail organelles in the epididymis, in somerespects their maturation is unusual. The failure to dem-onstrate changes in the Suncus sperm surface using tech-niques that consistently show these changes in other Euth-eria [3] does not necessarily mean that the sperm surfaceundergoes no modification during maturation, since thissurface change is demonstrable in nonpasserine bird andreptile spermatozoa only with immunocytochemical mark-ers for Wolffian duct secretory proteins [42-44]. However,not only do mature Suncus epididymal spermatozoa fail toexpress the usual changes and the affinities displayed typ-ically by eutherian spermatozoa; they also evidence a sig-nificant surface charge at pH 1.8 before entering the epi-didymis [3]. There, the acrosome is unusual in becomingmore resistant to disruptive agents, and this resilience mayhelp to maintain its erect form. While the evident stabilityof the acrosomal "rim" especially could be a link to thevariant character of Suncus sperm surface properties, it isnot clear why the behavior of the Suncus sperm surfaceshould differ from that of other mammals. However, sincethe epididymal surface factors seen in other species mayrelate to the ability of the peri-acrosomal region to bind tothe zona pellucida [45], and later to storage in the caudaepididymidis [46], it is interesting that in Suncus such bind-ing does not occur and that these spermatozoa seem less
acutely dependent on testicular androgen for storage thanthose of some other mammals. This could reflect some in-put of a nontesticular source of androgen in Suncus [47]and/or the fact that mature Suncus spermatozoa are rela-tively resilient cells.
Vas Deferens GlandVariation in the male accessory glands is very common
in eutherian mammals, but the tract of Suncus possesses a"pre" accessory gland on the vas deferens itself. A similarstructure is situated terminally in some other insectivores[48-51], but in Suncus the gland region continues again asvas deferens (Fig. 1). Since the number of spermatozoa inboth gland regions is close to that inseminated at coitus,this local portion of vas deferens and not the caudaepididymidis is probably the primary source of spermato-zoa in a first ejaculate. Although the secretion of the glandseems to change the sperm head surface, as judged by itsaffinity for glass, cauda spermatozoa were able to fertilizenormally. Thus, like the accessory gland secretions of otheranimals, the gland product is not essential for fertilizingability per se. The difficult approach of microsurgical ad-enectomy with natural mating may be the only way to es-tablish the underlying physiological role of the gland.
Spermatozoa in the Female
Vaginal plug formation and penile "locking" appear asalternative strategies for avoidance of sperm loss from thevagina in the peri-copulation period in muroid rodents [52],and these alternatives seem to be utilized in shrews. InBlarina, there is no vaginal plug and the mating pairs ex-hibit prolonged locking [17], whereas Suncus males do notlock but leave a prominent vaginal plug. While its loss by4-5 h p.c. here may have been spontaneous, females havebeen observed to pull out the plug and consume it (G.L.D;personal observation).
In terms of sperm production, Suncus lies at the lowend of the eutherian spectrum. Although Suncus femaleswill copulate repeatedly in captivity, the number of spermavailable in the male tract is not much greater than that ina first ejaculate. The one million or so ejaculated is some50 times less than in the mouse and also much less thancan be inferred from sperm-filled ducts of smaller shrewssuch as Sorex and Blarina [17, 48, 50]. However, in accordwith the findings of Clenendon and Rissman on fecundity[22], Table 1 shows that one ejaculate is quite sufficient foroptimal fertilization. Thus the relatively few spermatozoainseminated finally operate with efficiency in the quest tofertilize eggs.
In the effort to understand the frequent profligacy ofsperm production in mammals, male-male competition hasbeen invoked as a selective factor, particularly in polyan-drous species [53]. The situation in shrews suggests that thedeterminants of sperm production may be more compli-
831
Dow
nloaded from https://academ
ic.oup.com/biolreprod/article/50/4/820/2762587 by guest on 13 January 2022

BEDFORD ET AL.
cated. The social behavior and organization of various typesof shrews seem rather similar in that most have a shortreceptive period and most are asocial and solitary [54].However, the apparent multitude of spermatozoa in the fe-male tract in Blarina and Sorex [17,48], and also in thetenrec [16], certainly contrasts with the small population inmated Suncus females. Very low sperm numbers are as-sociated with storage in isthmic crypts in some marsupials[55]; but although some Suncus spermatozoa enter the cryptsin the fallopian tube, these structures occur also in Sorexaraneus [56], which appears to produce many more sper-matozoa. The success of small numbers of spermatozoa inSuncus may depend primarily on the particular characterof the sperm cell itself (e.g., its stable acrosome).
Fallopian Tube
A naive observer of the situation in Suncus might con-clude that its female tract has four compartments, i.e., va-gina, uterus, isthmus, and ampulla-so abruptly delineatedare the two segments of the oviduct (Fig. 7). This degreeof compartmentation distinguishes the Suncus fallopian tubeeven from that of a related shrew, Crocidura russula, inwhich there is a gradual decline in the concentration oftubal cilia from the bursa along the oviduct [57]. Our fewobservations suggest that ipsilateral ovulating follicles havesome regulatory influence on the eventual release of sper-matozoa from the isthmus to the ampulla in Suncus. Thehigh incidence of unilateral ovulation in this shew, the de-fined isthmus and ampulla, and the fact that spermatozoacan be distinguished through the oviduct wall in transmit-ted light may make Suncus a useful model in which to ex-plore the mechanisms of this final step of sperm transport.
Cumulus Oophorus
The cumulus oophorus of Suncus is unusual in two re-spects. It does not undergo the mucification and expansionthat normally parallel the development of a sensitivity tohyaluronidase before ovulation [58, 59]. It is also com-pletely resistant to hyaluronidase, presumably because ofits myriad junctions (Fig. 12) as well as a relative absenceof intercellular matrix. Further study of the cumulus oo-phorus in other shrews, whose acrosomes range from amodest to an exaggerated form approaching that in Suncus[12], may indicate to what extent the cumulus has been adeterminant of acrosomal morphology in this group.
Recent experimental evidence [60] supports the earlysuggestion [61] that the cumulus functions to sequester thefew spermatozoa within the ampulla as eggs are being pen-etrated. If the cumulus does act to promote early contactbetween gametes, how is such contact achieved in vivipa-rous species whose eggs are devoid of cumulus, and whydoes the cumulus expand before ovulation in rodents, rab-bits, and humans, among others, yet remain as a compactstructure in Suncus and many other insectivores? With re-
gard to the first point, the small eggs of many viviparousfish are fertilized within the confines of the ovarian follicle;and in other subtherians that practice internal fertilization,large eggs present an easy target for spermatozoa. In mar-supials, the eggs are very small but are fertilized within anarrow upper oviduct whose inner dimension equals thediameter of the egg. Shrews represent an intermediate sit-uation in which the modest space of the ampulla comparesto the size of the compact cumulus. Thus, whereas the sizeof the egg matches the dimensions of the fertilization sitein many viviparous species, it is only by preovulatory ma-trix secretion and cumulus expansion that this result isachieved in the spacious amnpulla typical of most eutherianmammals. This view does not ignore experimental evi-dence that cumulus expansion facilitates sperm passagethrough the cumulus [62-64]. However, the comparativepicture suggests that preovulatory expansion of the cu-mulus evolved as a means of enhancing the meeting of smalleggs and a few spermatozoa within a relatively large am-pullary space.
Sperm/Egg InteractionInoue et al. [65] stated that Suncus eggs are fertilized
about 7 h after ovulation, and in the present study pene-tration was not complete until about 10 h after ovulation(Table 1) despite the earlier presence of motile sperma-tozoa in the ampulla. These limited observations and thedelay of fertilization after direct intrauterine inseminationpoint to a need for Suncus spermatozoa to spend a pro-longed period in the female before they can fertilize. How-ever, this delay may not necessarily reflect a process similarto sperm capacitation in other Eutheria. Whereas capaci-tation seems to be linked to sperm surface modificationsrelated to storage in the cauda [1, 46, 66], it has not beenpossible to demonstrate these in the Suncus spermatozoon;and whether Suncus sperm undergo a comparable AR (seebelow) also seems questionable.
Despite the fact that homogenates of Suncus spermato-zoa display some temperature-dependent ability to dis-perse the mouse and rabbit cumulus, it is not clear howthey penetrate the Suncus cumulus. Although the huge ac-rosome appears to be discharged in passage through it (Figs.13-15), the cumulus is unresponsive to hyaluronidase aswell as to trypsin. A related question concerns the stimulusfor the physiological reaction/loss of the acrosome in Sun-cus, which our observations suggest is provided by the cu-mulus, not the zona pellucida. The acrosome was intact inall free motile spermatozoa in the ampulla and in one seenattached to the cumulus surface, yet spermatozoa within thecumulus (Figs. 13 and 14) and motile spermatozoa attachedto the zona (Fig. 15) had no acrosome. As expected, thiswas true of a spermatozoon moving within the zona, but,again, acrosomal "ghosts" were never evident on the zonasurface of penetrated eggs-in contrast to observations ina variety of laboratory and domestic mammals. On the other
832
Dow
nloaded from https://academ
ic.oup.com/biolreprod/article/50/4/820/2762587 by guest on 13 January 2022

UNUSUAL GAMETE FEATURES IN THE ASIAN MUSK SHREW
hand, the evidence that Suncus spermatozoa bind to thezona by the inner acrosome membrane is consistent withthe observation that intact spermatozoa failed to adhere tothe zona pellucida. This situation is reminiscent of that inthe guinea pig, which displays the unusual surface-relatedphenomenon of sperm rouleaux formation in the epidid-ymis [67].
We suspect that Suncus may be unusual not only in los-ing the acrosome before zona binding, but also in the modeof this loss. In view of the failure to detect conventionalreactions characterized by focal fusion between outer ac-rosomal and peri-acrosomal plasma membranes [68], it isinteresting that the loose or "wavy" peri-acrosomal plas-malemma-seen commonly in intact spermatozoa of othermammals-is substituted in Suncus by a stable acrosomal"rim" and a close relationship between these membranes(Figs. 3-6). Thus, it is possible that the reaction of the ac-rosome in Suncus does not mirror the AR in other mam-mals and that it is discharged as a whole cap from the an-terior border of the equatorial segment. The large variationin acrosome dimensions among other shrews may providethe means to examine further not only the extent to whichthe character of the Suncus acrosome is a legacy of its bi-zarre dimensions, but also the degree to which this featurehas determined other unusual aspects of conception dis-cussed here.
The final steps leading to syngamy in Suncus appearedto be relatively conventional. The egg was ovulated at meta-phase II, the fertilizing spermatozoon created a penetrationslit (Fig. 16), and decondensation of the sperm nucleus co-incided with extrusion of a second PB (Fig. 17). Althoughthe tail was fully incorporated, this step was often delayedwith much of the tail still protruding outside the zona ofeggs within which the fertilizing sperm head was alreadydecondensing. That the first PB tends to degenerate ac-counts for a report [19] that over half of the unfertilizedeggs recovered from the Suncus oviduct had no polar body.We have not yet captured the fusion and incorporation ofthe Suncus sperm head in TEM, but given its typical eu-therian design (equatorial segment, perforatorium, andkeratinoid flat nucleus), it is likely that this follows the uniquemode seen in other Eutheria.
Concluding Comment
While it has appeared that a given set of special gametecharacteristics constitutes one defining feature of eutherianmammals [1], wider examination of unconventional speciesbegins to suggest a greater diversity than anticipated in thisrespect. Recently, Breed [69] has described in the rodentgenus, Bandicota, a fundamentally different organization ofthe sperm head that complements an unusually trypsin-sen-sitive fragile zona pellucida in this genus (Bedford and Breed,unpublished results). From the current viewpoint of whatis normal in eutherian mammals, the preliminaries to fer-tilization seem highly unusual in Suncus also, although it
is not clear why this is so. Whatever the reason, it appearsthat some determinants of sperm function that operate inmost eutherian mammals are represented by another set ofcontrols in this insectivore; and that the major anomalies/distinctions in this respect are, domino-like, consequencesof others. In Suncus at least, much may really stem fromthe unprecedented size of the acrosome and a related sta-bility of its rim that may preclude the usual multi-point fu-sion AR and favor a dehiscence of the whole cap from theequatorial segment border. Since this appears to occur be-fore zona binding, it is consistent perhaps that the acro-some has no affinity for the zona and that the epididymalsurface change that usually creates this affinity in the mouse,rabbit, etc. [45], could not be detected in Suncus.
ACKNOWLEDGMENTS
We thank Drs. E.F. Hartree and C.R. Brown for their assay of acrosin in Suncusspermatozoa and Dr. F. Suzuki for her examination of the cumulus oophorus usingfreeze-fracture techniques.
REFERENCES1. Bedford JM. The co-evolution of mammalian gametes. In: Dunbar BS, O'Rand
MG (eds.), A Comparative Overview of Mammalian Fertilization. New York: PlenumPress; 1991: 3-35.
2. Green JA, Dryden GL. Ultrastructure of epididymal spermatozoa of the AsiaticMusk Shrew, Suncus murinus. Biol Reprod 1976; 14:327-331.
3. Cooper GW, Bedford JM. Asymmetry of spermiation and sperm surface chargepatterns over the giant acrosome in the musk shrew, Suncus murinus. J Cell Biol1976; 69:415-428.
4. Koehler JK. Fine structure of spermatozoa of the Asiatic musk shrew, Suncusmurinus. AmJ Anat 1977; 149:135-152.
5. Phillips DM, Bedford JM. Unusual features of sperm ultrastructure in the muskshrew Suncus murinus. J Exp Zool 1985; 235:119-126.
6. Mori T, Uchida AT. Fine structure of the spermatozoon of the house musk shrew,Suncus murinus. In: Oda S, KitohJ, Ota K, Isomura G. (eds.), Suncus murinus-Biology of the Laboratory Shrew. Tokyo: Japanese Scientific Societies Press SSP);1985: 335-351 (in Japanese with English summary).
7. Suzuki F, Racey PA Fine structural changes in the epididymal epithelium of moles(Talpa europaea) throughout the year. J Reprod Fertil 1976; 47:47-54.
8. Castellani-Ceresa L, Cotelli F, Lora-Lamia C, Berruti G. The spermatozoa of Talpaewopaea with particular reference to the acrosome region. J Ultrastruct Res 1980;72:112-118.
9. Tripepi S, Saita A, Carelli A. Ultrastructural peculiarities and movement of thespermatozoa of Erinaceus europaeus. Biol Cell 1982; 45:157.
10. Phillips DM, Dryden GL. Comparative morphology of mammalian gametes. In:Dunbar BS, O'Rand MG (eds.), A Comparative Overview of Mammalian Fertil-ization. New York: Plenum Press; 1991: 37-50.
11. Ploen L, Ekwall H, Afzelius B. Spermiogenesis and the spermatozoa of the Eu-ropean common shrew (Sorex araneus L). J Ultrastruct Res 1979; 68:149-159.
12. Mori T, Arai S, Shiraishi S, Uchida TA Ultrastructural observations on sperma-tozoa of the soricidae, with special attention to a sub-family revision of the Jap-anese water shrew, Chimarrogale himalayica. J Mammal Soc Jpn 1991; 16:1-12.
13. Carroll RL. Vertebrate Paleontology and Evolution. New York: Freeman & Co.;1988: 460.
14. Strauss F. Ripe follicles without antra and fertilization within the follicle: a normalsituation in a mammal. Anat Rec 1950; 106:251-252.
15. Mossman H, Duke KL. Comparative Morphology of the Mammalian Ovary. Mad-ison, WI: Univ. of Wisconsin Press; 1973.
16. Nicoll ME, Racey PA. Follicular development, ovulation, fertilization, and fetaldevelopment in tenrecs (Tenrec ecaudatus). J Reprod Fertil 1985; 74:47-55.
17. Pearson OP. Reproduction in the shrew (Blarina brevicauda Say). Am J Anat1944; 75:39-93.
18. Dryden GL. Reproduction in Suncus murinus. J Reprod Fertil Suppl 1969; 6:377-396.
833
Dow
nloaded from https://academ
ic.oup.com/biolreprod/article/50/4/820/2762587 by guest on 13 January 2022

BEDFORD ET AL.
19. Hasler MJ, Nalbandov AV. Ovulation, ovum maturation and changes in plasmaand adrenal progesterone concentration in the musk shrew (Suncus murinus).Biol Reprod 1980; 22:377-381.
20. Oda S, Kitoh J, Ota K, Isomura G. Suncus murinus-Biology of the LaboratoryShrew. Tokyo: Japan Sci Soc Press SSP); 1985: (in Japanese).
21. Rissman EF. Behavioral endocrinology of the female musk shrew. Horm Behav1991; 25:125-127.
22. Clenendon AL, Rissman EF. Prolonged copulatory behavior facilitates pregnancysuccess in the musk shrew. Physiol Behav 1990; 47:831-835.
23. Dryden GL. Establishment and maintenance of shrew colonies. Int Zoo Yearb1975; 15:12-18.
24. Yanagimachi R. In vitro sperm capacitation and fertilization of Golden Hamstereggs incubated in a chemically defined medium. In: Hafez ESE, Semm K (eds.),In Vitro Fertilization and Embryo Transfer. Lancaster, PA: MTP Press; 1982: 65-76.
25. Calvin HI, Bedford JM. Formation of disulphide bonds in the nucleus and ac-cessory structures of mammalian spermatozoa during maturation in the epidid-ymis. J Reprod Fertil Suppl 1971; 13:65-75.
26. Dryden GL, Pucek Z. Insectivores in reproduction studies, with emphasis on ovu-lation in American, Asian and European shrews. In: Antikatzides S, Erichsen S,Spiegel A (eds.), The Laboratory Animal in the Study of Reproduction. Stuttgart:Gustav Fischer Verlag; 1976: 39-50.
27. Kitoh J, Ota K, Yamashita K, Sugiura Y, Hirunagi K, Orita S, Yokoyama A Theurogenital organs. In: Oda S, KitohJ, Ota K, Isomura G (eds.), Suncus murinus-Biology of the Laboratory Shrew. Tokyo: Japan Sci Soc Press SSP); 1985: 239-258 (in Japanese).
28. Bedford JM, Berrios M, Dryden GL. Biology of the scrotum. IV. Testis locationand temperature sensitivity. J Exp Zool 1982; 224:379-388.
29. Brown CR, Andani Z, Hartree EF. Studies on ram acrosin. Isolation from sper-matozoa, activation by cations and organic solvents, and influence of cations onits reaction with inhibitors. Biochem J 1975; 149:133-146.
30. Brown CR, Hartree EF. Identification of acrosin in mouse spermatozoa. J ReprodFertil 1976; 46:249-251.
31. Chang MC. The effects of serum on spermatozoa. J Gen Physiol 1947; 30:321-335.
32. Hancock RJT. Spermotoxic properties of IgG fractions from normal rabbit sera.Arch Androl 1979; 2:171-177.
33. Woodall PF. An ultrastructural study of the spermatozoa of elephant shrews(Mammalia.Macroscelidea) and its phylogenetic implications. J SubmicroscopCytol Pathol 1991; 23:47-58.
34. Tripp HRH. Reproduction in elephant shrews (Macroscelididae) with special ref-erence to ovulation and implantation. J Reprod Fertil 1971; 26:149-159.
35. Thenius E. Die Evolution der Saugetiere. Stuttgart.: Fischer; 1979.36. Bedford JM. Sperm capacitation and fertilization in mammals. Biol Reprod 1970;
2: (suppl. 2):128-158.37. Rodger JC, Young RJ. Glycosidase and cumulus dispersal activities of acrosomal
extracts from opossum (marsupial) and rabbit (eutherian) spermatozoa. GameteRes 1981; 4:507-514.
38. Ward C, Storey BT. Determination of the time course of capacitation in mousespermatozoa using a chlortetracycline fluorescence assay. Dev Biol 1984; 104:287-296.
39. Corselli J, Talbot P. In vitro penetration of hamster oocyte-cumulus complexesusing physiological numbers of sperm. Dev Biol 1987; 122:227-242.
40. Chen C, Sathananthan AH. Early penetration of human sperm through the vest-ments of human eggs in vitro. Archs Androl 1986; 16:183-197.
41. White DR, Phillips DM, Bedford JM. Factors affecting the acrosome reaction inhuman spermatozoa. J Reprod Fertil 1990; 90:71-80.
42. Depeiges A, Dufaure J-P. Binding to spermatozoa of a major soluble protein se-creted by the epididymis of the lizard, Lacerta vivipara. Gamete Res 1983; 7:401-406.
43. Esponda P, Bedford JM. The rooster sperm surface changes in passing throughthe Wolffian duct. J Exp Zool 1985; 234:441-449.
44. Esponda P, Bedford JM. Post-testicular changes in the reptile sperm surface withparticular reference to the snake, Natrixfasciata. J Exp Zool 1987; 242:189-198.
45. Saling PM. Development of the ability to bind to zona pellucida during epidid-ymal maturation; reversible immobilization of mouse spermatozoa by lan-thanum. Biol Reprod 1982; 26:429-436.
46. Bedford JM, Yanagimachi R. Epididymal storage at abdominal temperature re-duces the time required for capacitation of hamster spermatozoa. J Reprod Fertil1991; 91:403-410.
47. Rissman EF, Crews D. Effect of castration on epididymal sperm storage in malemusk shrews (Suncus murinus) and mice (Mus musculus). J Reprod Fertil 1989;86:219-222.
48. Eadie WR. Homologies of the male accessory reproductive glands in Sorex andBlarina. Anat Rec 1947; 98:347-360.
49. Kowalska-Dyrcz A A comparative study of the male genital tract in some speciesof shrews. Acta Theriol 1966; 11:71-127.
50. Suzuki F, Racey PA. Light and electron microscopical observations on the maleexcurrent duct system of the common shrew (Sorex araneus). J Reprod Fertil1984; 70:419-428.
51. Brambell FWR. Reproduction in the common shrew (Sorex araneus Linnaeus):seasonal changes in the reproductive organs of the male. Philos Trans R SocLond B 1935; 225:51-62.
52. Hartung TC, Dewsbury DA A comparative analysis of copulatory plugs in muroidrodents and their relationship to copulatory behavior. J Mammal 1978; 59:717-723.
53. Gomendio M, Roldan ERS. Mechanisms of sperm competition: linking physiologyand behavioural ecology. TREE 1993; 8:95-100.
54. Churchfield S. Shrews. Ithaca, NY: Comstock Publishers; 1990.55. Bedford JM, Rodger JC, Breed WG. Why so many mammalian spermatozoa-a
clue from marsupials? Proc R Soc Lond B 1984; 221:221-233.56. Brambell FWR. Reproduction in the common shrew (Sorex araneus Linnaeus)
1. The oestrous cycle of the female. Philos Trans R Soc Lond B 1935; 225:2-49.57. Kress A, Milian J. The female genital tract of the shrew, Crocidura russula. Adv
Anat Embryol Cell Biol 1987; 101:1-76.58. Dekel N, Hillensjo T, Kraicer PF. Maturational effects of gonadotrophins on the
cumulus-oocyte complex of the rat. Biol Reprod 1979; 20:191-197.59. Dekel N, Phillips DM. Maturation of rat cumulus oophorus-a scanning electron
microscope study. Biol Reprod 1979; 21:9-18.60. Bedford JM, Kim HH. Cumulus oophorus as a sperm sequestering device. J Exp
Zool 1993; 265:321-328.61. Austin CR, Braden AWH. Passage of the sperm and the penetration of the egg in
mammals. Nature (Lond) 1952; 170:919-921.62. Ball GD, Liebfried ML, Lenz RW, Ax RL, Bavister BD, First NL. Factors affecting
successful in vitro fertilization of bovine follicular oocytes. Biol Reprod 1983;28:717-725.
63. Cuasnicu PS, Bedford JM. Hamster oocyte penetrability during preovulatory mat-uration. Mol Reprod Dev 1991; 29:72-76.
64. Chen L, Russell PT, Larsen WJ. Functional significance of cumulus expansion inthe mouse: roles for the preovulatory synthesis of hyaluronic acid within thecumulus mass. Mol Reprod Dev 1993; 34:87-93.
65. Inoue M, Orita S, Shimamura K, Kameyama Y. Development and abnormal de-velopment. In: Oda S, Kitoh J, Ota K, Isomura G (eds.), Suncus murinus-Bi-ology of the Laboratory Shrew. Tokyo: Japan Sci Soc Press SSP); 1985: 127-149(in Japanese).
66. Thomas TS, Wilson S, Reynolds AB, Oliphant G. Chemical and physical charac-teristics of the rabbit sperm acrosome stabilizing factor. Biol Reprod 1986; 30:693-703.
67. Huang ITF, Fleming AD, Yanagimachi R Acrosome-reacted spermatozoa can bindto and penetrate the zona pellucida: a study using guinea pig spermatozoa. J ExpZool 1981; 217:287-290.
68. Barros C, Bedford JM, Franklin LE, Austin CR. Membrane vesiculation as a featureof the mammalian acrosome reaction. J Cell Biol 1967; 34:C1-C5.
69. Breed WG. Novel organization of the spermatozoon in two species of muridrodents from Southern Asia. J Reprod Fertil 1993; 99:149-158.
834
Dow
nloaded from https://academ
ic.oup.com/biolreprod/article/50/4/820/2762587 by guest on 13 January 2022




















