
Dissociation of antidepressant-like activity of escitalopram and nortriptyline on behaviour and...
10
Original Paper Dissociation of antidepressant-like activity of escitalopram and nortriptyline on behaviour and hippocampal BDNF expression in female rats Anita C Hansson 1,2,* , Roberto Rimondini 3,* , Markus Heilig 1 , Aleksander A Mathe ´ 4 and Wolfgang H Sommer 1,2 Abstract A major hypothesis of depression postulates that a dysregulation of the neurotrophin systems is directly involved in the pathophysiology of depression, and that restoration of such deficits may underlie the therapeutic efficacy of antidepressant treatment. One key finding supporting this hypothesis is upregulation of brain derived neurotrophic factor (BDNF) in the hippocampus after antidepressant treatment. Here, we further test the hypothesis of BDNF involvement in antidepressant action in a genetic rat model of depression after chronic oral treatment with escitalopram, nortriptyline or placebo. Active treatments had significant behavioural antidepressant-like actions in female rats of the Flinders Sensitive Line (FSL) and non-selected Sprague Dawley (SD) rats, while Flinders Resistant Line (FRL) rats were unaffected. Escitalopram, but not nortriptyline, markedly reduced BDNF mRNA levels in the dentate gyrus of FSL rats. The BDNF downregulation was common to the four major promoters of the gene. Treatments did not affect BDNF expression in FRL or SD strains. We conclude that the antidepressant effects of escitalopram and nortriptyline, two common drugs with different pharmacological profiles, appear to be unrelated to the regulation of hippocampal BDNF expression in female rats. These results indicate that the tropic hypothesis of depression has limitations and emphasize the need for validated disease models of depression to assess potential treatment targets. Keywords Depression, forced swim test, PCR, SSRI Introduction Depression is the most common mental disorder and ranks among the leading causes of disability worldwide (Ezzati et al., 2002), the lifetime prevalence of the disorder being about two times higher in women than in men. The mecha- nism underlying the disease pathology and the mechanism of actions of antidepressant treatments are poorly understood. While a number of clinically proven treatments are available, about 25–40% of patients do not respond or only respond partially to at least one adequately performed medication trial (Fava, 2003). A prominent hypothesis of depression postulates that a dysregulation of the neurotrophin systems, in particular brain derived neurotrophic factor (BDNF), is involved in the pathophysiology of depression, and that reversal of such deficits may underlie the therapeutic efficacy of antidepressant treatments (Duman and Monteggia, 2006). BDNF is widely expressed throughout the brain, is generally produced and released by neurons, and is regulated by neuronal activity. Highest BDNF levels are found within the hippocampus, particularly the dentate gyrus (DG) (Conner et al., 1997; Ernfors et al., 1990; Hansson et al., 2000). The hippocampus is implicated in the pathophysiology of depression; for instance, women suffering from depression show reduced hippocampal volume (Sheline et al., 2003). Moreover, rodents subjected to acute or chronic stress or high levels of glucocor- ticoids show similar effects on hippocampus morphology as well as decreased BDNF expression, while various treatments for depression (including antidepressant drugs and electro- convulsive therapy) increase hippocampal BDNF expression and reverse the morphological changes (Hansson et al., 2003, 2006; Nibuya et al., 1995; Pizarro et al., 2004; Sapolsky, 2000; Schaaf et al., 1998; Smith et al., 1995; Tsankova et al., 2006). 1 Laboratory of Clinical and Translational Studies, NIAAA, National Institutes of Health, Bethesda, MD, USA. 2 Department of Psychopharmacology, Central Institute for Mental Health, Mannheim, Germany. 3 Department of Pharmacology, University of Bologna, Bologna, Italy. 4 Department of Clinical Neuroscience, The Karolinska Institute, Stockholm, Sweden. *These authors contributed equally to the study. Corresponding authors: Wolfgang H Sommer, Central Institute for Mental Health, Square J5, 68159 Mannheim, Germany Email: [email protected] Aleksander A Mathe ´, Karolinska Institute – Huddinge, Department of Psychiatry M56 SE-14186 Stockholm, Sweden Email: [email protected] Journal of Psychopharmacology 25(10) 1378–1387 ! The Author(s) 2011 Reprints and permissions: sagepub.co.uk/journalsPermissions.nav DOI: 10.1177/0269881110393049 jop.sagepub.com at Universitat Heidelberg on September 4, 2014 jop.sagepub.com Downloaded from
Transcript of Dissociation of antidepressant-like activity of escitalopram and nortriptyline on behaviour and...

Original Paper
Dissociation of antidepressant-like activityof escitalopram and nortriptyline onbehaviour and hippocampal BDNFexpression in female rats
Anita C Hansson1,2,*, Roberto Rimondini3,*, Markus Heilig1,Aleksander A Mathe4 and Wolfgang H Sommer1,2
AbstractA major hypothesis of depression postulates that a dysregulation of the neurotrophin systems is directly involved in the pathophysiology of depression,
and that restoration of such deficits may underlie the therapeutic efficacy of antidepressant treatment. One key finding supporting this hypothesis is
upregulation of brain derived neurotrophic factor (BDNF) in the hippocampus after antidepressant treatment. Here, we further test the hypothesis of
BDNF involvement in antidepressant action in a genetic rat model of depression after chronic oral treatment with escitalopram, nortriptyline or placebo.
Active treatments had significant behavioural antidepressant-like actions in female rats of the Flinders Sensitive Line (FSL) and non-selected Sprague
Dawley (SD) rats, while Flinders Resistant Line (FRL) rats were unaffected. Escitalopram, but not nortriptyline, markedly reduced BDNF mRNA levels in
the dentate gyrus of FSL rats. The BDNF downregulation was common to the four major promoters of the gene. Treatments did not affect BDNF
expression in FRL or SD strains. We conclude that the antidepressant effects of escitalopram and nortriptyline, two common drugs with different
pharmacological profiles, appear to be unrelated to the regulation of hippocampal BDNF expression in female rats. These results indicate that the tropic
hypothesis of depression has limitations and emphasize the need for validated disease models of depression to assess potential treatment targets.
KeywordsDepression, forced swim test, PCR, SSRI
Introduction
Depression is the most common mental disorder and ranksamong the leading causes of disability worldwide (Ezzatiet al., 2002), the lifetime prevalence of the disorder being
about two times higher in women than in men. The mecha-nism underlying the disease pathology and the mechanism ofactions of antidepressant treatments are poorly understood.While a number of clinically proven treatments are available,
about 25–40% of patients do not respond or only respondpartially to at least one adequately performed medication trial(Fava, 2003).
A prominent hypothesis of depression postulates that adysregulation of the neurotrophin systems, in particularbrain derived neurotrophic factor (BDNF), is involved in
the pathophysiology of depression, and that reversal of suchdeficits may underlie the therapeutic efficacy of antidepressanttreatments (Duman and Monteggia, 2006). BDNF is widelyexpressed throughout the brain, is generally produced and
released by neurons, and is regulated by neuronal activity.Highest BDNF levels are found within the hippocampus,particularly the dentate gyrus (DG) (Conner et al., 1997;
Ernfors et al., 1990; Hansson et al., 2000). The hippocampusis implicated in the pathophysiology of depression; forinstance, women suffering from depression show reduced
hippocampal volume (Sheline et al., 2003). Moreover, rodents
subjected to acute or chronic stress or high levels of glucocor-
ticoids show similar effects on hippocampus morphology aswell as decreased BDNF expression, while various treatmentsfor depression (including antidepressant drugs and electro-
convulsive therapy) increase hippocampal BDNF expressionand reverse the morphological changes (Hansson et al., 2003,2006; Nibuya et al., 1995; Pizarro et al., 2004; Sapolsky, 2000;Schaaf et al., 1998; Smith et al., 1995; Tsankova et al., 2006).
1Laboratory of Clinical and Translational Studies, NIAAA, National
Institutes of Health, Bethesda, MD, USA.2Department of Psychopharmacology, Central Institute for Mental Health,
Mannheim, Germany.3Department of Pharmacology, University of Bologna, Bologna, Italy.4Department of Clinical Neuroscience, The Karolinska Institute,
Stockholm, Sweden.
*These authors contributed equally to the study.
Corresponding authors:Wolfgang H Sommer, Central Institute for Mental Health, Square J5,
68159 Mannheim, Germany
Email: [email protected]
Aleksander A Mathe, Karolinska Institute – Huddinge, Department of
Psychiatry M56 SE-14186 Stockholm, Sweden
Email: [email protected]
Journal of Psychopharmacology
25(10) 1378–1387
! The Author(s) 2011
Reprints and permissions:
sagepub.co.uk/journalsPermissions.nav
DOI: 10.1177/0269881110393049
jop.sagepub.com
at Universitat Heidelberg on September 4, 2014jop.sagepub.comDownloaded from

Furthermore, intra-hippocampal BDNF infusion producesantidepressant-like effects (Shirayama et al., 2002), whilemice with reduced BDNF expression show impaired
antidepressant-like responses (Monteggia et al., 2004).However, the picture of an antidepressant BDNF-
mediated mechanism drawn by these studies is challengedby recent animal and human findings (Groves, 2007). For
example, some mutant mouse lines with lower or no detect-able BDNF expression do not exhibit depressive-like behav-iour (Chan et al., 2006; Chourbaji et al., 2004; MacQueen
et al., 2001; Saarelainen et al., 2003). Also, enhanced hippo-campal BDNF expression as a result of communal nestingincreased depressive-like behaviour in the mouse forced
swim test (FST) (Branchi et al., 2006). In female rats acutestress increased BDNF protein levels in the DG, but chronicstress decreased BDNF in the medial prefrontal cortex
(mPFC) (Lin et al., 2009). In contrast, these regions remainedunaffected in male rats under the same conditions. Moreover,there is evidence that in circuitries other than the hippocam-pus, BDNF actually can increase symptoms of negative affect,
such as social aversion and craving for cocaine (Berton et al.,2006; Lu et al., 2004). Thus, some BDNF effects seem to besex- or brain region-specific, and further clarification of the
role of BDNF in depression is needed.The BDNF gene has a unique, highly conserved architec-
ture. In rodents, it is composed of at least eight 50-exons, eachwith its own promoter, that are spliced to a common 30-term-inal exon containing the coding frame (Aid et al., 2007;Timmusk et al., 1993). Most commonly studied are the pro-moters for exons I and II, which seem to depend on recurrent
protein synthesis for their transcription (Lauterborn et al.,1996), and for exon IV and VI, which have been shown toshare properties with immediate early genes and seem to medi-
ate much of the activity-dependent gene regulation of BDNF(Jiang et al., 2008; Lipsky et al., 2001; Nakayama et al., 1994).
So far, the majority of studies have investigated patho-
physiology of depression and effects of antidepressant treat-ments in healthy mice and rats under baseline conditions orexposed to stress, and more recently selected gene knockouts
in mice. In contrast, in this study we have used an animalmodel of depression, the selectively bred FSL rat that has innumerous studies exhibited a good face, construct and pre-dictive validity (Overstreet et al., 2005). Neurochemical and
pharmacological findings from FSL rats are consistent witha serotonergic and dopaminergic model of depression.Importantly, the FSL rats are a well-validated model for
the study of antidepressant actions (Overstreet, 2002). Liketheir clinical profile, chronic but not acute administration oftricyclic antidepressants and SSRIs reduces immobility in the
FST in this model. Furthermore, despite the fact that womenhave higher prevalence of depression, there is a strong biastowards the use of male animals to study depression-relatedphenotypes. For this reason we used female rats in the present
study.Here, we tested the hypothesis that regulation of BDNF
expression is involved in the actions of antidepressant drugs.
We explored exon-specific regulation of BDNF expression inthe DG of female FSL rats after chronic treatment with esci-talopram, the S-enantiomer of the SSRI citalopram, which is
clinically effective (Cipriani et al., 2009), and has proved to beeffective in preclinical models of depression and anxiety(El Khoury et al., 2006; Petersen et al., 2008; Sanchez et al.,
2003; Uys et al., 2006). We also studied BDNF expression inthe mPFC, another brain region important for the patho-physiology of depression in humans (Andersen and Teicher,2008; Mayberg et al., 2005; Walter et al., 2009). We compared
the effects of the SSRI escitalopram with those of nortripty-line, a tricyclic antidepressant with a more noradrenergic pro-file, on forced swim behaviour and hippocampal BDNF
expression in FSL, their controls, the Flinders ResistantLine (FRL), and non-selected Sprague Dawley (SD) rats.
Experimental procedures
Animals
Female FSL and FRL rats weighing 220–250 g, from the ratcolonies maintained at the animal facility at the KarolinskaInstitute, Sweden, and SD rats weighing 220–240 g (Harlan
Nossan, Italy) were housed in groups of three under con-trolled conditions of light (from 07:00 to 19:00), temperature(22� 2�C) and humidity (65%), and were allowed free accessto the different diets and tap water. The experimental proto-
col was approved by the local Committees for AnimalProtection and the procedures and animal comfort werecontrolled by the Institutional Veterinary Service and are in
accordance with the European Community Council Directive(86/609/EEC). Experiments were carried out under dim light(12 lux), between 09:00 and 16:00. Animals were moved to the
experimental room in their own cages 1 hour before the startof the experiments for habituation to the novel environment.The experiment comparing FSL and FRL rats (experiment 1,
n¼ 10/group) was carried out at the Karolinska Institute, theother experiments using FSL (experiment 2, n¼ 12/group) orSD (experiment 3, n¼ 12/group) rats were performed at theUniversity of Bologna.
Drug treatment
Animals were randomly assigned to a 30-day dietary treatment
with either escitalopram, nortriptyline or vehicle admixed foodpellets (0.35 g/kg pellet) according to amethod developed byHLundbeck A/S (Copenhagen, Denmark) and tested in collab-
oration with A Mørk (El Khoury et al., 2006; Petersen et al.,2009). The administered drug dose was approximately 25mg/kg rat weight/day for escitalopram and 20mg/kg for nortrip-
tyline. The efficacy of the treatments had been determinedbased on their behavioural effects in preliminary experiments.Results in serum concentrations have been established in sev-
eral previous experiments of about 25–35 ng/mL (Bjornebekket al., 2008, 2010; El Khoury et al., 2006).
Behavioural testing
Time line. Animals were allowed to adapt to the facilityfor at least 2 weeks prior to the oral drug treatment. A FST
(2-day protocol) occurred on days 24 and 25 of the oral drugtreatment for all groups. In addition, animals from
Hansson et al. 1379
at Universitat Heidelberg on September 4, 2014jop.sagepub.comDownloaded from

experiment 3 underwent elevated plus maze (EPM) and openfield (OF) testing on days 21 and 22, respectively. On day 30all animals were sacrificed for mRNA analysis. A 5-day
period from the FST was deemed sufficient to recover fromthe acute swim stress, because although acute stress reduceshippocampal BDNF mRNA levels (Arunrut et al., 2009;Ueyama et al., 1997), this effect appears to be transient
(Hansson et al., 2006; Schaaf et al., 1998).
Forced swim test. A 2-day FST was performed as previ-ously described (El Khoury et al., 2006). Animals wereplaced individually in a vertical Plexiglass cylinder (height
45 cm; diameter 19 cm) containing tap water at a depth of20 cm (25�C). The animals were removed from the waterafter 15minutes and dried before being returned to their
home cages. On the next day, the animals were replaced inthe cylinders for 5minutes. The 5-minute swim was videorecorded and the time spent in swimming and immobil-ity were scored via Ethovison� ver. 3.1 (Noldus, the
Netherlands).
Elevated plus maze. The apparatus consisted of twoopen arms (40 cm� 10 cm), two enclosed arms of the samesize, and a central area (10 cm� 10 cm); placement was
50 cm above the floor. Experiments were carried out underdim light (12 lux), between 09:00 and 15:00. Behaviour wasvideotaped and scored via Ethovison� ver. 3.1 (Noldus, theNetherlands). At the beginning of a session, subjects were
placed in the central part of the maze facing one of theopen arms. The numbers of entries made into closed andopen arms and the time spent in closed and open arms were
recorded for 5minutes. An arm entry was defined as all fourpaws into an arm.
Open field. OF activity was measured in a square OF arena(100 cm� 100 cm� 50 cm). At the beginning of a session, the
rat was placed in the central part of the arena. The number ofentries into the centre, corners, walls and periphery (cornersplus walls) and time spent in the same areas were recordedand scored via Ethovison� ver. 3.1 (Noldus, the Netherlands)
for 10minutes.
Gene expression analysis
The effect of escitalopram treatment on BDNF mRNA levelsin DG and mPFC was assessed using quantitative RT-PCR.
Transcriptional activity of BDNF is controlled by differentpromoters located upstream of short untranslated exons ofthe BDNF gene. In experiment 1, the contribution of fourof these promoters to the escitalopram-induced regulation
of BDNF mRNA expression was evaluated using specificprobes for exons I, II, IV and VI. Abundance of the trans-lated part of the BDNF gene was determined using common
exon IXa probes. In experiments 2 and 3 only the latter probewas used because no exon-specific effect on hippocampalBDNF expression was found in experiment 1.
Tissue preparation. Five days following FST the animalswere decapitated between 13:00 and 15:00, brains were frozenin �40�C isopentane and kept at �70�C. Two brain regions
were dissected out: in experiment 1, the dentate gyrus of thehippocampus (DG) and medial prefrontal cortex (mPFC).Samples were obtained under a magnifying lens, using ana-tomical landmarks (Paxinos and Watson, 1998). DG was pre-
pared from a 2-mm thick coronal slice, taken in a Kopf brainslicer by placing the rostral blade on the caudal edge of theoptic chiasm. For preparation of mPFC (containing cingulate
and prelimbic cortex Cg1, Cg2 and PrL according to Paxinosand Watson (1998)), the blades were placed 3mm and 5mmrostral to this landmark, and a second 2-mm coronal slice was
obtained. In experiments 2 and 3 whole hippocampus wasdissected according to the same landmarks used above.Samples were stored at �70�C until RNA was prepared.
RNA extraction. Total RNA was extracted with TRIzolreagent (Gibco BRL Life Technologies, Baltimore, MD,
USA) followed by an RNeasy (Qiagen, Hilden, Germany)clean-up step according to the manufacturer’s instructions.RNA was prepared from individual samples, in parallel and
in balanced order. All RNA samples showed A260/280 ratiosbetween 1.9 and 2.1. RNA integrity was determined using anAgilent 2100 Bioanalyser (Agilent Technologies, Santa Clara,
CA, USA), and only material without signs of degradationwas used.
Quantitative real-time RT-PCR. Reverse transcription(RTþ) was performed in a 100mL reaction mix (AppliedBiosystems, Foster City, CA, USA) using 100 ng total RNA
per animal. To assess contamination of the samples withremaining genomic DNA, parallel control reactions fromeach sample were performed without enzyme (RT–). Using
5mL from either RTþ or RT–, PCR was performed in a 50mLreaction mix containing TaqMan Universal Master Mix(Applied Biosystems) and a gene-specific primer/probe set
(Cybergene AB, Huddinge, Sweden). Primer/probe combina-tions (Table 1) were designed according to the guidelinesgiven by Applied Biosystems and had comparable efficiencyof amplification. Each sample was assayed in triplicate in
parallel with an endogenous control (Cyca or Actx). Allprimer/probe sets had comparable efficiency of amplification.Because of the lack of an exon-specific treatment effect in
experiment 1, material from experiments 2 and 3 were onlyassayed for the common exon IXa together with the endoge-nous controls. The experiments were performed on the ABI
Prism 7700 platform, according to the instructions of themanufacturer (Applied Biosystems), and repeated at leasttwice. Data were normalized for expression of the endoge-nous controls using the ��Ct method as described by the
manufacturer (Applied Biosystems).
Statistical analysis
Behavioural data and expression data were analysed usingfactorial analysis of variance (ANOVA).
1380 Journal of Psychopharmacology 25(10)
at Universitat Heidelberg on September 4, 2014jop.sagepub.comDownloaded from
Results
Escitalopram had antidepressant-like effects
in FSL but not FRL rats
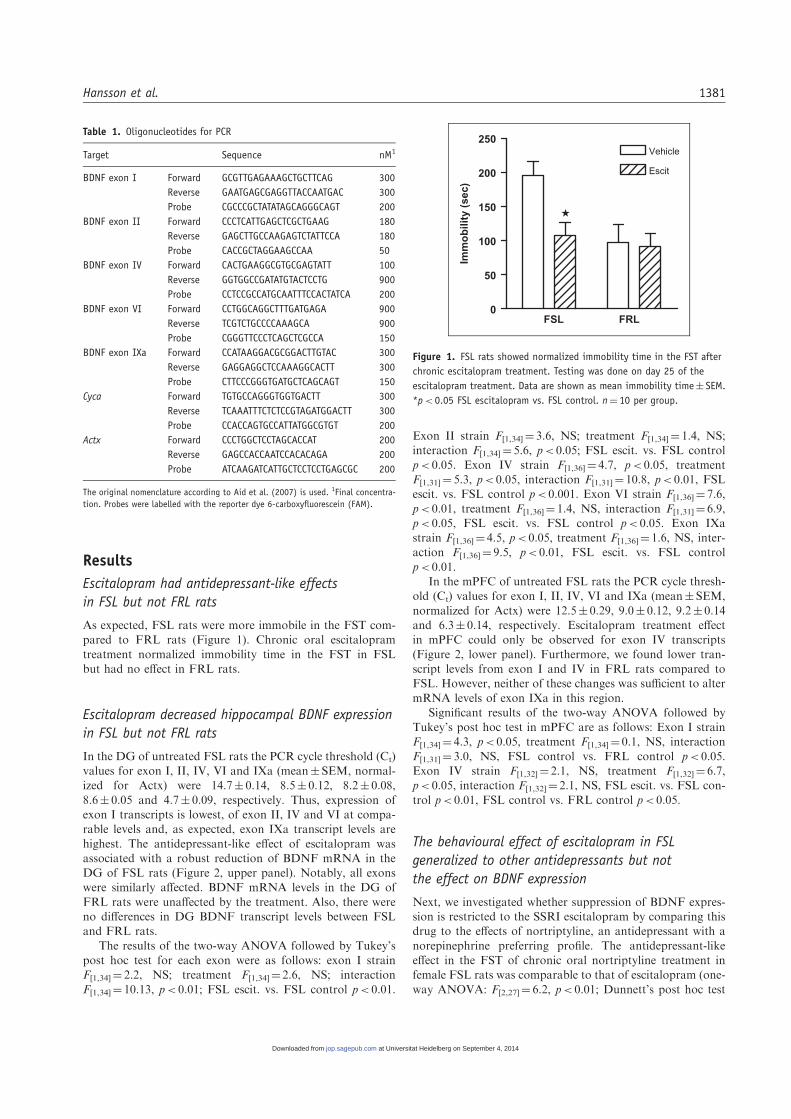
As expected, FSL rats were more immobile in the FST com-pared to FRL rats (Figure 1). Chronic oral escitalopram
treatment normalized immobility time in the FST in FSLbut had no effect in FRL rats.
Escitalopram decreased hippocampal BDNF expression
in FSL but not FRL rats
In the DG of untreated FSL rats the PCR cycle threshold (Ct)values for exon I, II, IV, VI and IXa (mean� SEM, normal-ized for Actx) were 14.7� 0.14, 8.5� 0.12, 8.2� 0.08,
8.6� 0.05 and 4.7� 0.09, respectively. Thus, expression ofexon I transcripts is lowest, of exon II, IV and VI at compa-rable levels and, as expected, exon IXa transcript levels are
highest. The antidepressant-like effect of escitalopram wasassociated with a robust reduction of BDNF mRNA in theDG of FSL rats (Figure 2, upper panel). Notably, all exonswere similarly affected. BDNF mRNA levels in the DG of
FRL rats were unaffected by the treatment. Also, there wereno differences in DG BDNF transcript levels between FSLand FRL rats.
The results of the two-way ANOVA followed by Tukey’spost hoc test for each exon were as follows: exon I strainF[1,34]¼ 2.2, NS; treatment F[1,34]¼ 2.6, NS; interaction
F[1,34]¼ 10.13, p< 0.01; FSL escit. vs. FSL control p< 0.01.
Exon II strain F[1,34]¼ 3.6, NS; treatment F[1,34]¼ 1.4, NS;interaction F[1,34]¼ 5.6, p< 0.05; FSL escit. vs. FSL control
p< 0.05. Exon IV strain F[1,36]¼ 4.7, p< 0.05, treatmentF[1,31]¼ 5.3, p< 0.05, interaction F[1,31]¼ 10.8, p< 0.01, FSLescit. vs. FSL control p< 0.001. Exon VI strain F[1,36]¼ 7.6,
p< 0.01, treatment F[1,36]¼ 1.4, NS, interaction F[1,31]¼ 6.9,p< 0.05, FSL escit. vs. FSL control p< 0.05. Exon IXastrain F[1,36]¼ 4.5, p< 0.05, treatment F[1,36]¼ 1.6, NS, inter-action F[1,36]¼ 9.5, p< 0.01, FSL escit. vs. FSL control
p< 0.01.In the mPFC of untreated FSL rats the PCR cycle thresh-
old (Ct) values for exon I, II, IV, VI and IXa (mean� SEM,
normalized for Actx) were 12.5� 0.29, 9.0� 0.12, 9.2� 0.14and 6.3� 0.14, respectively. Escitalopram treatment effectin mPFC could only be observed for exon IV transcripts
(Figure 2, lower panel). Furthermore, we found lower tran-script levels from exon I and IV in FRL rats compared toFSL. However, neither of these changes was sufficient to alter
mRNA levels of exon IXa in this region.Significant results of the two-way ANOVA followed by
Tukey’s post hoc test in mPFC are as follows: Exon I strainF[1,34]¼ 4.3, p< 0.05, treatment F[1,34]¼ 0.1, NS, interaction
F[1,31]¼ 3.0, NS, FSL control vs. FRL control p< 0.05.Exon IV strain F[1,32]¼ 2.1, NS, treatment F[1,32]¼ 6.7,p< 0.05, interaction F[1,32]¼ 2.1, NS, FSL escit. vs. FSL con-
trol p< 0.01, FSL control vs. FRL control p< 0.05.
The behavioural effect of escitalopram in FSL
generalized to other antidepressants but not
the effect on BDNF expression
Next, we investigated whether suppression of BDNF expres-sion is restricted to the SSRI escitalopram by comparing thisdrug to the effects of nortriptyline, an antidepressant with a
norepinephrine preferring profile. The antidepressant-likeeffect in the FST of chronic oral nortriptyline treatment infemale FSL rats was comparable to that of escitalopram (one-
way ANOVA: F[2,27]¼ 6.2, p< 0.01; Dunnett’s post hoc test
Table 1. Oligonucleotides for PCR
Target Sequence nM1
BDNF exon I Forward GCGTTGAGAAAGCTGCTTCAG 300
Reverse GAATGAGCGAGGTTACCAATGAC 300
Probe CGCCCGCTATATAGCAGGGCAGT 200
BDNF exon II Forward CCCTCATTGAGCTCGCTGAAG 180
Reverse GAGCTTGCCAAGAGTCTATTCCA 180
Probe CACCGCTAGGAAGCCAA 50
BDNF exon IV Forward CACTGAAGGCGTGCGAGTATT 100
Reverse GGTGGCCGATATGTACTCCTG 900
Probe CCTCCGCCATGCAATTTCCACTATCA 200
BDNF exon VI Forward CCTGGCAGGCTTTGATGAGA 900
Reverse TCGTCTGCCCCAAAGCA 900
Probe CGGGTTCCCTCAGCTCGCCA 150
BDNF exon IXa Forward CCATAAGGACGCGGACTTGTAC 300
Reverse GAGGAGGCTCCAAAGGCACTT 300
Probe CTTCCCGGGTGATGCTCAGCAGT 150
Cyca Forward TGTGCCAGGGTGGTGACTT 300
Reverse TCAAATTTCTCTCCGTAGATGGACTT 300
Probe CCACCAGTGCCATTATGGCGTGT 200
Actx Forward CCCTGGCTCCTAGCACCAT 200
Reverse GAGCCACCAATCCACACAGA 200
Probe ATCAAGATCATTGCTCCTCCTGAGCGC 200
The original nomenclature according to Aid et al. (2007) is used. 1Final concentra-
tion. Probes were labelled with the reporter dye 6-carboxyfluorescein (FAM).
0
50
100
150
200
250Vehicle
Escit
FSL
Imm
ob
ility
(se
c)
FRL
Figure 1. FSL rats showed normalized immobility time in the FST after
chronic escitalopram treatment. Testing was done on day 25 of the
escitalopram treatment. Data are shown as mean immobility time� SEM.
*p< 0.05 FSL escitalopram vs. FSL control. n¼ 10 per group.
Hansson et al. 1381
at Universitat Heidelberg on September 4, 2014jop.sagepub.comDownloaded from

p< 0.01 for both treatment groups vs. controls, Figure 3).Neither drug showed any significant effects in the EPM and
OF tests demonstrating behavioural specificity (Table 2).Expression analysis for experiment 2 was restricted to thecommon exon IXa because we did not find any exon-specificeffects in experiment 1. No effect of nortriptyline on hippo-
campal mRNA levels was found, while escitalopram showedthe same treatment effects as in experiment 1 (one-wayANOVA: F[2,26]¼ 9.5, p< 0.01, Dunnett’s post hoc test
p< 0.05, escitalopram vs. control; Figure 3).
Escitalopram had antidepressant effects in SD rats but
did not affect BDNF expression
Finally, to investigate whether the effects of escitalopram andnortriptyline on BDNF expression are a specific characteristicof the FSL model or can be generalized to ‘behaviourallynormal’ rat models, we tested SD rats for their behavioural
and their hippocampal BDNF response to these treatments.In contrast to FSL rats, the treatment effects of escitalopramand nortriptyline were inconsistent, in as much that only esci-
talopram showed a significant effect on immobility time in theFST (one-way ANOVA: F[2,31]¼ 5.9, p< 0.01, Dunnett’s posthoc test p< 0.01, escitalopram vs. control, Figure 4). Also in
this model no contingency of the antidepressant treatmentwith expression of BDNF exon IXa expression in the hippo-campus was found.
Discussion
The most salient finding of the present study, contrary to
several previous reports, is the robust downregulation of hip-pocampal BDNF expression by escitalopram, a clinicallyeffective treatment for major depression (Cipriani et al.,
2009). Moreover, there was a clear dissociation between the
effect of escitalopram on BDNF and the immobility time in
the FST, a finding that was replicated in two independentexperiments, indicating that the antidepressant action is notcontingent on increased BDNF expression. These data are in
line with the growing literature questioning increased BDNFexpression as a requirement for antidepressant effects.Nortriptyline, a classical tricyclic drug that exhibits potenteffects in severely depressed inpatients (Danish University
Antidepressant Group, 1986), also did not modify BDNFexpression despite improving the depressive phenotype inFSL rats, lending further support to the hypothesis that an
increased expression of BDNF is not a prerequisite for anti-depressant effects of drugs.
The neurotrophin hypothesis of depression was postulated
on the basis of clinical observations that depression can be
Exon I
FSL FRL0
25
50
75
100
125
% F
SL
co
ntr
ol
Exon II
FSL FRL0
25
50
75
100
125
Exon IV
FSL FRL0
25
50
75
100
125
Exon VI
FSL FRL0
25
50
75
100
125
Exon IXa
FSL FRL0
25
50
75
100
125VehicleEscit
FSL FRL0
25
50
75
100
125
#
% F
SL
co
ntr
ol
FSL FRL0
25
50
75
100
125
FSL FRL0
25
50
75
100
125
#
FSL FRL0
25
50
75
100
125
FSL FRL0
25
50
75
100
125
DG
mP
FC
Figure 2. Real time RT-PCR for the promoter-specific non-coding exons I, II, IV and VI and the common coding exon IXa mRNA expression in dentate
gyrus (DG) and medial prefrontal cortex (mPFC) of chronically escitalopram (hatched bars) and vehicle (clear bars) treated FSL and FRL rats. Data are
expressed as percentage of the FSL control group (mean� SEM). *p< 0.05, **p< 0.01, and ***p< 0.001 FSL, escitalopram vs. FSL control. #p< 0.05,
FSL control vs. FRL control. n¼ 9–10 per group.
FST
Con Cit Nor0
50
100
150
200
250
Imm
ob
ility
[sec
]
Exon IXa mRNA
Con Cit Nor0
25
50
75
100
125
% C
on
tro
l
Figure 3. Immobility time in the FST (left) and exon IXa mRNA expres-
sion in the hippocampus (right) after chronic escitalopram or nortripty-
line treatment in FSL rats. Data are shown as immobility time (sec) or
percentage of the FSL control group (mean� SEM), respectively.
**p< 0.05, escitalopram or nortriptyline vs. controls. n¼ 9–12 per group.
1382 Journal of Psychopharmacology 25(10)
at Universitat Heidelberg on September 4, 2014jop.sagepub.comDownloaded from

precipitated by acute or chronic stress and that in someanimal models administration of physical and emotional
stressors results in downregulation of hippocampal BDNFexpression, which can be reversed by diverse antidepressanttreatments (reviewed in Duman and Monteggia, 2006). Therelationship between stress and depression is, however, com-
plex, and subjects who receive a depression diagnosis arelikely to represent heterogeneous populations of phenocopies,with varying contribution from stress exposure (Kendler
et al., 2001). Also, preclinical evaluation of antidepressant-like activity is commonly carried out in normal animals,and rarely captures a critical feature of antidepressant phar-
macotherapy, a delayed onset of action.The FSL rat model shows stable innate depression-like
phenotype and has a high degree of predictive validity forseveral classes of antidepressants including SSRIs and tricy-
clics in the FST (Overstreet et al., 2005). Several previousstudies in this model have investigated levels of BDNF inthe hippocampus and prefrontal cortex at baseline and
after various antidepressant treatments. Basal BDNF proteinlevels as measured by enzyme-linked immunosorbent assay(ELISA) did not differ in the hippocampus and prefrontal
cortex between male FSL and FRL rats, while in agreementwith the present study, female FSL rats showed increasedBDNF levels in the prefrontal cortex (Angelucci et al.,
2000, 2003a, 2003b). Furthermore, the BDNF levels in maleFSL rats were either not affected or reduced by electrocon-vulsive stimuli or chronic lithium administration, respectively(Angelucci et al., 2000, 2003a, 2003b). More recent studies
also found no differences in cortical and hippocampalBDNF mRNA levels between male FSL and FRL rats(Bjornebekk et al., 2005). Wheel-running increased hippo-
campal BDNF expression, but only in the non-depressedFRL rats, while at the same time exercise improved perfor-mance in the FST and increased hippocampal neurogenesis in
FSL but not FRL rats. In another study, wheel-running butnot chronic escitalopram treatment produced a significantincrease in hippocampal BDNF expression in female FSLrats, however no significant improvement in the FST was
found after either treatment (Bjornebekk et al., 2008). Thesestudies are in part consistent with our results, but no directcomparisons can be made due to the different housing condi-
tions. Thus, social isolation but not group housing, as in thepresent study, profoundly altered neurogenesis in female FSLcompared to SD rats (Bjornebekk et al., 2007a, 2007b).
Further, the PCR assays employed here are more sensitive
than the in situ hybridization method. Our findings are
strengthened by the fact that our behavioural experimentswere performed in two cohorts of animals at two differentsites, showing essentially consistent treatment effects, both
at the behavioural and the gene expression level.Besides the FSL/FRL model only a few animal lines with
innate depressive-like traits exist. Fawn hooded rats, an inbredrat line with high immobility in the FST, when reared in iso-
lation display an anxiety-like phenotype (Rezvani et al., 2002).In another model of genetic susceptibility to develop depres-sion, the selectively bred rat line for sensitivity to acquire
learned helplessness in response to stress (Vollmayr et al.,2001), animals do not respond to antidepressant treatmentand show an uncoupling of hippocampal BDNF expression
from stress-evoked corticosterone levels, a characteristicresponse in normal SD or Wistar lines (Hansson et al., 2003,2006; Sapolsky, 2000; Schaaf et al., 1998; Smith et al., 1995).Also, rats selectively bred for low and high exploratory behav-
iour (LE/HE rats) show increased prefrontal but not hippo-campal BDNF levels in the low exploratory line (Mallo et al.,2008). Collectively, the data from these genetic models of
depression do not support a common BDNF-mediated mech-anism for depression and its treatment.
Limitations of the BDNF hypothesis of depression are
evident also in other rodent models. For example, while
Table 2. Effect of treatment on behaviour in the elevated plus maze and in the open field
Controls Escitalopram Nortriptyline One-way ANOVA
EPM F[2,27]% time in open arms 25�2� 3.25 18�9� 3.82 28�5� 3.11 2.1
% entries in open arm 42�4� 3.95 34�9� 2.66 42�9� 2.64 2.0
Total number of entries 27�6� 1.80 31�8� 2.23 24�9� 2.27 2.7
OF F[2,33]Time in centre (s) 38�1� 5.62 26�8� 4.10 34�5� 5.07 1.4
Entries in centre 16�3� 2.00 12�1� 1.67 14�2� 1.61 1.3
Total duration of movement (s) 500�3� 6.80 500�7� 10.1 482�6� 6.50 1.7
FST
Con Cit Nor0
50
100
150
Imm
ob
ility
[sec
]
Exon IX mRNA
Con Cit Nor0
50
100
150
% C
on
tro
l
Figure 4. Immobility time in the FST (left) and exon IXa mRNA expres-
sion in the hippocampus (right) after chronic escitalopram or nortripty-
line treatment in SD rats. Data are shown as immobility time (sec) or
percentage of the FSL control group (mean� SEM), respectively. For
detailed statistics see Results section. **p< 0.01. n¼ 10–12 per group.
Hansson et al. 1383
at Universitat Heidelberg on September 4, 2014jop.sagepub.comDownloaded from

mice with genetically altered glucocorticoid receptor expres-sion showed both altered sensitivity for stress-induced depres-sive reactions and in agreement with the hypothesis of
reduced hippocampal BDNF levels (Ridder et al., 2005), cen-tral BDNF levels remained unchanged in a mouse modelof learned helplessness (Schulte-Herbruggen et al., 2006).By contrast, the well-established olfactory bulbectomy in
mice led to increased BDNF levels in depression-relatedbrain areas (Hellweg et al., 2007), and in a well-characterizedrat model, in which chronic stress induces depressive-like
behaviour, an increase of cortical BDNF levels could beobserved after stress treatment that was reversed by treatmentwith escitalopram (Schulte-Herbruggen et al., 2009).
Generally, most of the research has focused on male ani-mals. Interestingly, female but not male conditional BDNFknockout mice exhibit depression-like behaviour (Chan et al.,
2006; Monteggia et al., 2007). A recent study compared theresponse to the tricyclic antidepressant clomipramine in maleand female FSL or SD rats, and found similar treatmenteffects for all relevant behavioural parameters in the FST in
both sexes despite some baseline differences (Kokras et al.,2009). In isolated female FSL or SD rats, escitalopram neitheraffected behaviour in the FST nor BDNF mRNA levels
(Bjornebekk et al., 2008). A potential confound in workingwith female rats is the oestrous cycle, which may affect BDNFexpression. In fact, oestradiol was shown to regulate BDNF
levels in the hippocampus (Singh et al., 1995) and this regu-lation may involve serotonergic mechanisms (Cavus andDuman, 2003). However, oestradiol’s effect on BDNFmRNA expression seems to be variable depending on brain
region, age and hormonal status. Thus, in female SD rats,BDNF mRNA levels seem to be about 20% lower duringoestrous compared to proestrus, but oestradiol per se does
not seem to affect the reduction of BDNF in response toacute stress (Cavus and Duman, 2003). However, differencesin cycle are expected to result in increased variance blunting
potential treatment effects, which is not the case for majoroutput parameters in the present study, i.e. FST behaviourand the escitalopram effect on DG BDNF levels in FSL rats.
Furthermore, it is our experience that female rats synchronizetheir cycle spontaneously during long holding times (Jimenez-Vasquez et al., 2000). Also, in line with the present study,increased serotonergic tone at 5HT2A receptors was found
to reduce BDNF expression in the DG of female SD rats.Although independent of oestradiol this effect is differentfrom the males, where acute stress decreased BDNF mRNA
in the hippocampus (Smith et al., 1995; Vaidya et al., 1999).Thus, we cannot presently exclude sexual dimorphism tounderlie the results reported here, although it should be
kept in mind that some antidepressant treatments didreduce central BDNF levels in male FSL rats as well(Angelucci et al., 2003b).
An initial aim of our study was to test the hypothesis of
exon-specific transcriptional regulation of BDNF expressionby escitalopram. Exon-specific upregulation of BDNF wasfound after chronic treatment in SD rats with the SSRI flu-
oxetine, the tricyclic antidepressant desipramine and themonoamine oxidase inhibitor tranylcypromine, which alluse protein synthesis dependent exon I, but not the stimu-
lus-responsive exons IV or VI (Khundakar and Zetterstrom,
2006). Similar studies confirm these results for desipramineand tranylcypromine or phenelzine, but found no effect forfluoxetine (Dias et al., 2003; Dwivedi et al., 2006). Also,
chronic treatment with duloxetine, a serotonin-norepinephr-ine reuptake inhibitor, showed increased expression of BDNFin the frontal cortex which was mainly sustained by exon Iand IV promoters, whereas the expression of exon VI was
reduced (Calabrese et al., 2007). Here, we found a downregu-lation of exon IV transcripts in the mPFC by escitalopram,but there was no effect on the common exon IXa. Together,
these results indicate region-specific and, more importantly,highly pharmacologically distinct recruitment of differentBDNF promoters to regulate transcript levels.
The fact that expression from the four BDNF promoters,which are all differently organized, is similarly downregulatedby escitalopram in the hippocampus makes the involvement
of a specific transcriptional mechanism at this locus unlikely.Instead, the differential BDNF response between FSL andFRL lines may be better explained by global changes in meth-ylation or histone acetylation patterns affecting the entire
locus. Such epigenetic mechanisms have been found to con-tribute to the regulation of BDNF expression (Chen et al.,2003; Tsankova et al., 2006).
In the present study we focused exclusively on BDNF tran-script levels. We and others have demonstrated good correla-tion of BDNFmRNA and protein levels for acute and chronic
challenges (De Foubert et al., 2004; Hansson et al., 2006;Schaaf et al., 1998). However, divergence between mRNAand protein levels has also been observed, in particular afterrepeated drug administration (Fumagalli et al., 2007; Jacobsen
and Mork, 2004). More important than steady state levels ofthe protein, however, appear to be the release properties ofBDNF. The picture emerges that BDNF can be released
in its precursor form, which preferentially binds the pan-neurotrophin receptor p75 (p75NTR), or in its mature formactivates the BDNF-specific TrkB receptor. This is important
as activation of these distinct receptors has opposite effects onsynaptic strength (Lu et al., 2005).Whether processing, storageor release of either BDNF form is differentially regulated
between FSL and FRL rats, and whether such differencescould play a role in the BDNF response to antidepressanttreatment, remains to be determined.
In conclusion, our studies have demonstrated that two
mainstay antidepressants reduce immobility in the FST, avalidated animal model of antidepressant-like activity, bothin FSL, but do not activate hippocampal BDNF expression.
Non-selected SD rats showed an inconsistent behaviouralresponse to the two antidepressants, which for escitalopramwas again not associated with hippocampal BDNF expres-
sion. This demonstrates that increased BDNF expression isunder some conditions not necessary for antidepressantaction and emphasizes the need for validated disease modelsof depression to assess potential treatment targets. To over-
come the present lack of progress in medication developmentit is necessary to evaluate existing animal models for theirplacement within a robust theoretical framework of the dis-
ease process and to select those that show close homology tohuman tests (e.g. biomarkers or endophenotypes of depres-sion) in favour of models that have a loose appearance of
completeness of the general human condition (i.e. face value).
1384 Journal of Psychopharmacology 25(10)
at Universitat Heidelberg on September 4, 2014jop.sagepub.comDownloaded from

Acknowledgement
We thank Asa Sodergren for excellent technical support with the real-
time PCR experiments.
Funding
This work was supported by the intramural research funds of
NIAAA, the Swedish Medical Research Council (grant number
10414 to AAM) and the Karolinska Institute. The sponsors had no
further role in study design; in the collection, analysis and interpre-
tation of data; in the writing of the report; and in the decision to
submit the paper for publication.
Conflict of interest statement
All authors were involved in the design of the study. AAM provided
the animals. RR, ACH and WHS performed experiments and data
analysis. WHS wrote the manuscript; AAM and MH critically revised
it. All authors contributed to and have approved the final manuscript.
All other authors declare that they have no conflicts of interest.
References
Aid T, Kazantseva A, Piirsoo M, Palm K and Timmusk T (2007)
Mouse and rat BDNF gene structure and expression revisited.
J Neurosci Res 85: 525–535.
Andersen SL and Teicher MH (2008) Stress, sensitive periods and
maturational events in adolescent depression. Trends Neurosci 31:
183–191.
Angelucci F, Aloe L, Jimenez-Vasquez P and Mathe AA (2003a)
Electroconvulsive stimuli alter nerve growth factor but not
brain-derived neurotrophic factor concentrations in brains of a
rat model of depression. Neuropeptides 37: 51–56.
Angelucci F, Aloe L, Jimenez-Vasquez P and Mathe AA (2003b)
Lithium treatment alters brain concentrations of nerve growth
factor, brain-derived neurotrophic factor and glial cell line-
derived neurotrophic factor in a rat model of depression. Int J
Neuropsychopharmacol 6: 225–231.
Angelucci F, Aloe L, Vasquez PJ and Mathe AA (2000) Mapping the
differences in the brain concentration of brain-derived neuro-
trophic factor (BDNF) and nerve growth factor (NGF) in an
animal model of depression. Neuroreport 11: 1369–1373.
Arunrut T, Alejandre H, Chen M, Cha J and Russo-Neustadt A
(2009) Differential behavioral and neurochemical effects of exer-
cise, reboxetine and citalopram with the forced swim test. Life Sci
84: 584–589.
Berton O, McClung CA, Dileone RJ, Krishnan V, Renthal W, Russo
SJ, et al. (2006) Essential role of BDNF in the mesolimbic dopa-
mine pathway in social defeat stress. Science 311: 864–868.
Bjornebekk A, Mathe AA and Brene S (2005) The antidepressant
effect of running is associated with increased hippocampal cell
proliferation. Int J Neuropsychopharmacol 8: 357–368.
Bjornebekk A, Mathe AA and Brene S (2007a) Isolated Flinders
Sensitive Line rats have decreased dopamine D2 receptor
mRNA. Neuroreport 18: 1039–1043.
Bjornebekk A, Mathe AA and Brene S (2010) The antidepressant
effects of running and escitalopram are associated with levels of
hippocampal NPY and Y1 receptor but not cell proliferation in a
rat model of depression. Hippocampus 20: 820–828.
Bjornebekk A, Mathe AA, Gruber SH and Brene S (2007b) Social
isolation increases number of newly proliferated cells in hippo-
campus in female flinders sensitive line rats. Hippocampus 17:
1193–1200.
Bjornebekk A, Mathe AA, Gruber SH and Brene S (2008)
Housing conditions modulate escitalopram effects on
antidepressive-like behaviour and brain neurochemistry. Int J
Neuropsychopharmacol 11: 1135–1147.
Branchi I, D’Andrea I, Sietzema J, Fiore M, Di FV, Aloe L, et al.
(2006) Early social enrichment augments adult hippocampal
BDNF levels and survival of BrdU-positive cells while increasing
anxiety- and ‘‘depression’’-like behavior. J Neurosci Res 83:
965–973.
Calabrese F, Molteni R, Maj PF, Cattaneo A, Gennarelli M, Racagni
G, et al. (2007) Chronic duloxetine treatment induces
specific changes in the expression of BDNF transcripts and in
the subcellular localization of the neurotrophin protein.
Neuropsychopharmacology 32: 2351–2359.
Cavus I and Duman RS (2003) Influence of estradiol, stress, and
5-HT2A agonist treatment on brain-derived neurotrophic factor
expression in female rats. Biol Psychiatry 54: 59–69.
Chan JP, Unger TJ, Byrnes J and Rios M (2006) Examination of
behavioral deficits triggered by targeting Bdnf in fetal or postna-
tal brains of mice. Neuroscience 142: 49–58.
Chen WG, Chang Q, Lin Y, Meissner A, West AE, Griffith EC, et al.
(2003) Derepression of BDNF transcription involves calcium-
dependent phosphorylation of MeCP2. Science 302: 885–889.
Chourbaji S, Hellweg R, Brandis D, Zorner B, Zacher C, Lang UE,
et al. (2004) Mice with reduced brain-derived neurotrophic factor
expression show decreased choline acetyltransferase activity, but
regular brain monoamine levels and unaltered emotional behav-
ior. Brain Res Mol Brain Res 121: 28–36.
Cipriani A, Santilli C, Furukawa TA, Signoretti A, Nakagawa A,
McGuire H, et al. (2009) Escitalopram versus other antidepressive
agents for depression. Cochrane Database Syst Rev CD006532.
Conner JM, Lauterborn JC, Yan Q, Gall CM and Varon S (1997)
Distribution of brain-derived neurotrophic factor (BDNF) pro-
tein and mRNA in the normal adult rat CNS: evidence for anter-
ograde axonal transport. J Neurosci 17: 2295–2313.
Danish University Antidepressant Group Citalopram: clinical effect
profile in comparison with clomipramine. A controlled multicen-
ter study. Psychopharmacology (Berl) 90: 131–138.
De Foubert G, Carney SL, Robinson CS, Destexhe EJ, Tomlinson R,
Hicks CA, et al. (2004) Fluoxetine-induced change in rat brain
expression of brain-derived neurotrophic factor varies depending
on length of treatment. Neuroscience 128: 597–604.
Dias BG, Banerjee SB, Duman RS and Vaidya VA (2003)
Differential regulation of brain derived neurotrophic factor tran-
scripts by antidepressant treatments in the adult rat brain.
Neuropharmacology 45: 553–563.
Duman RS and Monteggia LM (2006) A neurotrophic model for
stress-related mood disorders. Biol Psychiatry 59: 1116–1127.
Dwivedi Y, Rizavi HS and Pandey GN (2006) Antidepressants
reverse corticosterone-mediated decrease in brain-derived neuro-
trophic factor expression: differential regulation of specific exons
by antidepressants and corticosterone. Neuroscience 139:
1017–1029.
El Khoury A, Gruber SH, Mork A and Mathe AA (2006) Adult life
behavioral consequences of early maternal separation are allevi-
ated by escitalopram treatment in a rat model of depression. Prog
Neuropsychopharmacol Biol Psychiatry 30: 535–540.
Ernfors P, Wetmore C, Olson L and Persson H (1990) Identification
of cells in rat brain and peripheral tissues expressing mRNA for
members of the nerve growth factor family. Neuron 5: 511–526.
Ezzati M, Lopez AD, Rodgers A, van den Horn S and Murray CJ
(2002) Selected major risk factors and global and regional burden
of disease. Lancet 360: 1347–1360.
Fava M (2003) Diagnosis and definition of treatment-resistant
depression. Biol Psychiatry 53: 649–659.
Fumagalli F, Di PL, Caffino L, Racagni G and Riva MA (2007)
Repeated exposure to cocaine differently modulates BDNF
Hansson et al. 1385
at Universitat Heidelberg on September 4, 2014jop.sagepub.comDownloaded from

mRNA and protein levels in rat striatum and prefrontal cortex.
Eur J Neurosci 26: 2756–2763.
Groves JO (2007) Is it time to reassess the BDNF hypothesis of
depression? Mol Psychiatry 12: 1079–1088.
Hansson AC, Cintra A, Belluardo N, Sommer W, Bhatnagar M,
Bader M, et al. (2000) Gluco- and mineralocorticoid receptor-
mediated regulation of neurotrophic factor gene expression in
the dorsal hippocampus and the neocortex of the rat. Eur J
Neurosci 12: 2918–2934.
Hansson AC, Sommer WH, Metsis M, Stromberg I, Agnati LF and
Fuxe K (2006) Corticosterone actions on the hippocampal brain-
derived neurotrophic factor expression are mediated by exon IV
promoter. J Neuroendocrinol 18: 104–114.
Hansson AC, Sommer W, Rimondini R, Andbjer B, Stromberg I and
Fuxe K (2003) c-fos reduces corticosterone-mediated effects on
neurotrophic factor expression in the rat hippocampal CA1
region. J Neurosci 23: 6013–6022.
Hellweg R, Zueger M, Fink K, Hortnagl H and Gass P (2007)
Olfactory bulbectomy in mice leads to increased BDNF levels
and decreased serotonin turnover in depression-related brain
areas. Neurobiol Dis 25: 1–7.
Jacobsen JP and Mork A (2004) The effect of escitalopram, desipra-
mine, electroconvulsive seizures and lithium on brain-derived neu-
rotrophic factor mRNA and protein expression in the rat brain
and the correlation to 5-HT and 5-HIAA levels. Brain Res 1024:
183–192.
Jiang X, Tian F, Du Y, Copeland NG, Jenkins NA, Tessarollo L,
et al. (2008) BHLHB2 controls Bdnf promoter 4 activity and neu-
ronal excitability. J Neurosci 28: 1118–1130.
Jimenez-Vasquez PA, Overstreet DH and Mathe AA (2000)
Neuropeptide Y in male and female brains of Flinders Sensitive
Line, a rat model of depression. Effects of electroconvulsive stim-
uli. J Psychiatr Res 34: 405–412.
Kendler KS, Thornton LM and Gardner CO (2001) Genetic risk,
number of previous depressive episodes, and stressful life events
in predicting onset of major depression. Am J Psychiatry 158:
582–586.
Khundakar AA and Zetterstrom TS (2006) Biphasic change in
BDNF gene expression following antidepressant drug treatment
explained by differential transcript regulation. Brain Res 1106:
12–20.
Kokras N, Antoniou K, Dalla C, Bekris S, Xagoraris M, Overstreet D,
et al. (2009) Sex-related differential response to clomipramine treat-
ment in a ratmodel of depression. JPsychopharmacol 23(8): 945–956.
Lauterborn JC, Rivera S, Stinis CT, Hayes VY, Isackson PJ and Gall
CM (1996) Differential effects of protein synthesis inhibition on
the activity-dependent expression of BDNF transcripts: evidence
for immediate-early gene responses from specific promoters.
J Neurosci 16: 7428–7436.
Lin Y, Ter Horst GJ, Wichmann R, Bakker P, Liu A, Li X, et al.
(2009) Sex differences in the effects of acute and chronic stress and
recovery after long-term stress on stress-related brain regions of
rats. Cereb Cortex 19(9): 1978–1989.
Lipsky RH, Xu K, Zhu D, Kelly C, Terhakopian A, Novelli A, et al.
(2001) Nuclear factor kappaB is a critical determinant in N-
methyl-D-aspartate receptor-mediated neuroprotection.
J Neurochem 78: 254–264.
Lu L, Dempsey J, Liu SY, Bossert JM and Shaham Y (2004) A single
infusion of brain-derived neurotrophic factor into the ventral teg-
mental area induces long-lasting potentiation of cocaine seeking
after withdrawal. J Neurosci 24: 1604–1611.
Lu B, Pang PT and Woo NH (2005) The yin and yang of neurotro-
phin action. Nat Rev Neurosci 6: 603–614.
MacQueen GM, Ramakrishnan K, Croll SD, Siuciak JA, Yu G,
Young LT, et al. (2001) Performance of heterozygous brain-
derived neurotrophic factor knockout mice on behavioral
analogues of anxiety, nociception, and depression. Behav
Neurosci 115: 1145–1153.
Mallo T, Koiv K, Koppel I, Raudkivi K, Uustare A, Rinken A, et al.
(2008) Regulation of extracellular serotonin levels and brain-
derived neurotrophic factor in rats with high and low exploratory
activity. Brain Res 1194: 110–117.
Mayberg HS, Lozano AM, Voon V, McNeely HE, Seminowicz D,
Hamani C, et al. (2005) Deep brain stimulation for treatment-
resistant depression. Neuron 45: 651–660.
Monteggia LM, Barrot M, Powell CM, Berton O, Galanis V, Gemelli
T, et al. (2004) Essential role of brain-derived neurotrophic factor
in adult hippocampal function. Proc Natl Acad Sci U S A 101:
10827–10832.
Monteggia LM, Luikart B, Barrot M, Theobold D, Malkovska I, Nef
S, et al. (2007) Brain-derived neurotrophic factor conditional
knockouts show gender differences in depression-related behav-
iors. Biol Psychiatry 61: 187–197.
Nakayama M, Gahara Y, Kitamura T and Ohara O (1994)
Distinctive four promoters collectively direct expression of
brain-derived neurotrophic factor gene. Brain Res Mol Brain
Res 21: 206–218.
Nibuya M, Morinobu S and Duman RS (1995) Regulation of BDNF
and trkB mRNA in rat brain by chronic electroconvulsive seizure
and antidepressant drug treatments. J Neurosci 15: 7539–7547.
Overstreet DH (2002) Behavioral characteristics of rat lines selected
for differential hypothermic responses to cholinergic or serotoner-
gic agonists. Behav Genet 32: 335–348.
Overstreet DH, Friedman E, Mathe AA and Yadid G (2005) The
Flinders Sensitive Line rat: a selectively bred putative animal
model of depression. Neurosci Biobehav Rev 29: 739–759.
Paxinos G and Watson C (1998) The Rat Brain in Stereotaxic
Coordinates. San Diego: Academic Press.
Petersen A, Wortwein G, Gruber SH, El-Khoury A and Mathe AA
(2009) Nortriptyline mediates behavioral effects without affecting
hippocampal cytogenesis in a genetic rat depression model.
Neurosci Lett 451: 148–151.
Petersen A, Wortwein G, Gruber SH and Mathe AA (2008)
Escitalopram reduces increased hippocampal cytogenesis in a
genetic rat depression model. Neurosci Lett 436: 305–308.
Pizarro JM, Lumley LA, Medina W, Robison CL, Chang WE,
Alagappan A, et al. (2004) Acute social defeat reduces neurotro-
phin expression in brain cortical and subcortical areas in mice.
Brain Res 1025: 10–20.
Rezvani AH, Parsian A and Overstreet DH (2002) The Fawn-
Hooded (FH/Wjd) rat: a genetic animal model of comorbid
depression and alcoholism. Psychiatr Genet 12: 1–16.
Ridder S, Chourbaji S, Hellweg R, Urani A, Zacher C, Schmid W,
et al. (2005) Mice with genetically altered glucocorticoid receptor
expression show altered sensitivity for stress-induced depressive
reactions. J Neurosci 25: 6243–6250.
Saarelainen T, Hendolin P, Lucas G, Koponen E, Sairanen M,
MacDonald E, et al. (2003) Activation of the TrkB neurotrophin
receptor is induced by antidepressant drugs and is required for
antidepressant-induced behavioral effects. J Neurosci 23: 349–357.
Sanchez C, Bergqvist PB, Brennum LT, Gupta S, Hogg S, Larsen A,
et al. (2003) Escitalopram, the S-(þ)-enantiomer of citalopram, is
a selective serotonin reuptake inhibitor with potent effects in
animal models predictive of antidepressant and anxiolytic activi-
ties. Psychopharmacology (Berl) 167: 353–362.
Sapolsky RM (2000) Glucocorticoids and hippocampal atrophy in
neuropsychiatric disorders. Arch Gen Psychiatry 57: 925–935.
Schaaf MJM, de Jong J, De Kloet ER and Vreugdenhil E (1998)
Downregulation of BDNF mRNA and protein in the rat hippo-
campus by corticosterone. Brain Res 813: 112–120.
Schulte-Herbruggen O, Chourbaji S, Muller H, Danker-Hopfe H,
Brandwein C, Gass P, et al. (2006) Differential regulation of
1386 Journal of Psychopharmacology 25(10)
at Universitat Heidelberg on September 4, 2014jop.sagepub.comDownloaded from

nerve growth factor and brain-derived neurotrophic factor in a
mouse model of learned helplessness. Exp Neurol 202: 404–409.
Schulte-Herbruggen O, Fuchs E, Abumaria N, Ziegler A, Danker-
Hopfe H, Hiemke C, et al. (2009) Effects of escitalopram on the
regulation of brain-derived neurotrophic factor and nerve growth
factor protein levels in a rat model of chronic stress. J Neurosci
Res 87: 2551–2560.
Sheline YI, Gado MH and Kraemer HC (2003) Untreated depression
and hippocampal volume loss. Am J Psychiatry 160: 1516–1518.
Shirayama Y, Chen AC, Nakagawa S, Russell DS and Duman RS
(2002) Brain-derived neurotrophic factor produces antidepressant
effects in behavioral models of depression. J Neurosci 22:
3251–3261.
Singh M, Meyer EM and Simpkins JW (1995) The effect of ovariec-
tomy and estradiol replacement on brain-derived neurotrophic
factor messenger ribonucleic acid expression in cortical and hip-
pocampal brain regions of female Sprague-Dawley rats.
Endocrinology 136: 2320–2324.
Smith MA, Makino S, Kvetnansky R and Post RM (1995) Stress and
glucocorticoids affect the expression of brain-derived neuro-
trophic factor and neurotrophin-3 messenger-RNAs in the hippo-
campus. J Neurosci 15: 1768–1777.
Timmusk T, Palm K, Metsis M, Reintam T, Paalme V, Saarma M,
et al. (1993) Multiple promoters direct tissue-specific expression of
the rat BDNF-gene. Neuron 10: 475–489.
Tsankova NM, Berton O, Renthal W, Kumar A, Neve RL and
Nestler EJ (2006) Sustained hippocampal chromatin regulation
in a mouse model of depression and antidepressant action. Nat
Neurosci 9: 519–525.
Ueyama T, Kawai Y, Nemoto K, Sekimoto M, Tone S and Senba E
(1997) Immobilization stress reduced the expression of neurotro-
phins and their receptors in the rat brain. Neurosci Res 28:
103–110.
Uys JD, Muller CJ, Marais L, Harvey BH, Stein DJ and Daniels WM
(2006) Early life trauma decreases glucocorticoid receptors in rat
dentate gyrus upon adult re-stress: reversal by escitalopram.
Neuroscience 137: 619–625.
Vaidya VA, Terwilliger RM and Duman RS (1999) Role of 5-HT2A
receptors in the stress-induced down-regulation of brain-derived
neurotrophic factor expression in rat hippocampus. Neurosci Lett
262: 1–4.
Vollmayr B, Faust H, Lewicka S and Henn FA (2001) Brain-derived-
neurotrophic-factor (BDNF) stress response in rats bred for
learned helplessness. Mol Psychiatry 6: 471–474, 358.
Walter M, Henning A, Grimm S, Schulte RF, Beck J, Dydak U, et al.
(2009) The relationship between aberrant neuronal activation in
the pregenual anterior cingulate, altered glutamatergic metabo-
lism, and anhedonia in major depression. Arch Gen Psychiatry
66: 478–486.
Hansson et al. 1387
at Universitat Heidelberg on September 4, 2014jop.sagepub.comDownloaded from




















