Differential regulation of transport proteins in the periinfarct region following reversible middle...
9
DIFFERENTIAL REGULATION OF TRANSPORT PROTEINS IN THE PERIINFARCT REGION FOLLOWING REVERSIBLE MIDDLE CEREBRAL ARTERY OCCLUSION IN RATS P. DAZERT, a1 Y. SUOFU, b1 M. GRUBE, a A. POPA-WAGNER, b H. K. KROEMER, a * G. JEDLITSCHKY a AND C. KESSLER b a Department of Pharmacology, Research Center of Pharmacology and Experimental Therapeutics, Ernst-Moritz-Arndt University, Greifs- wald, Germany b Department of Neurology, Ernst-Moritz-Arndt University, Greifswald, Germany Abstract—Members of various transport protein families in- cluding ATP-binding cassette transporters and solute carri- ers were shown to be expressed in brain capillaries, choroid plexus, astrocytes or neurons, controlling drug and metabo- lite distribution to and from the brain. However, data are currently very limited on how the expression of these trans- port systems is affected by damage to the brain such as stroke. Therefore we studied the expression of four se- lected transporters, P-glycoprotein (Mdr1a/b; Abcb1a/b), Mrp5 (Abcc5), Bcrp (Abcg2), and Oatp2 (Slc21a5) in a rat model for stroke. Transporter expression was analyzed by real-time polymerase chain reaction in the periinfarcted re- gion and protein localization and cellular phenotyping were done by immunohistochemistry and confocal immunofluo- rescence microscopy. After stroke, P-glycoprotein staining was detected in endothelial cells of disintegrated capillaries and by day 14 in newly generated blood vessels. There was no significant difference, however, in the Mdr1a mRNA amount in the periinfarcted region compared with the con- tralateral site. For Bcrp, a significant mRNA up-regulation was observed from days 3–14. This up-regulation was fol- lowed by the protein as confirmed by quantitative immuno- histochemistry. Oatp2, located in the vascular endothelium, was also up-regulated at day 14. For Mrp5, an up-regulation was observed in neurons in the periinfarcted region (day 14). In conclusion, after stroke the transport proteins were up-regulated with a maximum at day 14, a time point that coincides with behavioral recuperation. The study further suggests Bcrp as a pronounced marker for the regenerative process and a possible functional role of Mrp5 in surviving neurons. © 2006 IBRO. Published by Elsevier Ltd. All rights reserved. Key words: blood– brain barrier, stroke ischemic, disease model animal, drug therapy. The tight junctions between adjacent brain capillary en- dothelial cells restrict the entry of compounds by para- cellular diffusion from the blood to the brain and vice versa, forming the blood-brain barrier (BBB). The tran- scellular transport of compounds through the membrane bilayer of the brain endothelial cells is controlled by transport proteins expressed in these cells, including transmembrane efflux pumps belonging to the ATP- binding cassette protein (ABC) superfamily (De Boer et al., 2003; Sun et al., 2003). P-glycoproteins (P-gps) encoded by MDR1 (ABCB1) in human and mdr1a and mdr1b (abcb1a/abcb1b) genes in rodents have been shown to be physiologically expressed and located at the luminal membrane in capillary blood vessels of the brain (Thiebaut et al., 1987; Croop et al., 1989). P-gps actively expel a wide range of bulky lipophilic neutral or cationic compounds from the brain into the blood, lead- ing to a decrease in their uptake by the brain (Schinkel, 2001; Dagenais et al., 2001). The lack of P-gp in mdr1a/b knock-out mice affects drug transport into the brain more than in any other peripheral organs (Schinkel, 2001). Another member of the ABC super- family, which was first called breast cancer resistance protein (BCRP) or mitoxantrone resistance protein (MXR) and is now designated ABCG2, confers resis- tance to a spectrum of compounds overlapping with P-gp substrates (Haimeur et al., 2004). BCRP/ABCG2 was recently identified in human, porcine, and mice brain capillary endothelial cells (Cooray et al., 2002; Eisenblatter et al., 2003; Zhang et al., 2003; Cisternino et al., 2004) raising the question of how it influences the efflux of drugs at the BBB. Multidrug resistance proteins (MRPs), members of the C-branch of the ABC trans- porter family, mediate the export of organic anions from cells. Conjugates of lipophilic compounds with glutathi- one, glucuronate, or sulfate are preferred substrates of MRP1-3 (Haimeur et al., 2004; Konig et al., 1999), while cyclic purine nucleotides and nucleotide analogs have been identified as substrates for MRP4 and MRP5 (ABCC4 and ABCC5) (Ritter et al., 2005). The physio- logical functions of MRP5 remain still to be defined, however, the finding that it extrudes cGMP from cells, suggests that it can affect the signal transduction role of cGMP by reducing its intracellular content and provide extracellular cGMP for paracrine function. MRP5 protein 1 The first two authors have contributed equally to this work. *Correspondence to: H. K. Kroemer, Institut für Pharmakologie, Ernst- Moritz-Arndt-Universität, Friedrich-Loeffler-Str. 23d, D-17487 Greifs- wald, Germany. Tel: 49-3834-865630; fax: 49-3834-865631. E-mail address: [email protected] (H. K. Kroemer). Abbreviations: ABC, ATP-binding cassette protein; BBB, blood– brain barrier; BCRP, breast cancer resistance protein, ABCG2 (Bcrp: rat ortholog); MRP, multidrug resistance protein, ABCC transporter (Mrp: rat ortholog); OATP, organic anion-transporting polypeptide (Oatp: rat ortholog); PCR, polymerase chain reaction; P-gp(s), P-glycoprotein(s); PXR, pregnane X receptor; RECA, rat endothelial cell-specific anti- body; SLC, solute carrier; TTC, 2,3,5,-triphenyltetrazolium chloride; vWF, von Willebrand factor. Neuroscience 142 (2006) 1071–1079 0306-4522/06$30.000.00 © 2006 IBRO. Published by Elsevier Ltd. All rights reserved. doi:10.1016/j.neuroscience.2006.07.056 1071
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Differential regulation of transport proteins in the periinfarct region following reversible middle...

DPC
PAGa
awb
G
AceplcpslMmrgdrwanatwlhww
ucspnr
1
*MwEAboroPbv
Neuroscience 142 (2006) 1071–1079
0d
IFFERENTIAL REGULATION OF TRANSPORT PROTEINS IN THEERIINFARCT REGION FOLLOWING REVERSIBLE MIDDLE
EREBRAL ARTERY OCCLUSION IN RATSKm
Tdcvsbttbaemstbaci2mb(fp(tPwbEee(pcoMcb(lhsc
. DAZERT,a1 Y. SUOFU,b1 M. GRUBE,a
. POPA-WAGNER,b H. K. KROEMER,a*
. JEDLITSCHKYa AND C. KESSLERb
Department of Pharmacology, Research Center of Pharmacologynd Experimental Therapeutics, Ernst-Moritz-Arndt University, Greifs-ald, Germany
Department of Neurology, Ernst-Moritz-Arndt University, Greifswald,ermany
bstract—Members of various transport protein families in-luding ATP-binding cassette transporters and solute carri-rs were shown to be expressed in brain capillaries, choroidlexus, astrocytes or neurons, controlling drug and metabo-
ite distribution to and from the brain. However, data areurrently very limited on how the expression of these trans-ort systems is affected by damage to the brain such astroke. Therefore we studied the expression of four se-ected transporters, P-glycoprotein (Mdr1a/b; Abcb1a/b),rp5 (Abcc5), Bcrp (Abcg2), and Oatp2 (Slc21a5) in a ratodel for stroke. Transporter expression was analyzed by
eal-time polymerase chain reaction in the periinfarcted re-ion and protein localization and cellular phenotyping wereone by immunohistochemistry and confocal immunofluo-escence microscopy. After stroke, P-glycoprotein stainingas detected in endothelial cells of disintegrated capillariesnd by day 14 in newly generated blood vessels. There waso significant difference, however, in the Mdr1a mRNAmount in the periinfarcted region compared with the con-ralateral site. For Bcrp, a significant mRNA up-regulationas observed from days 3–14. This up-regulation was fol-
owed by the protein as confirmed by quantitative immuno-istochemistry. Oatp2, located in the vascular endothelium,as also up-regulated at day 14. For Mrp5, an up-regulationas observed in neurons in the periinfarcted region (day 14).
In conclusion, after stroke the transport proteins werep-regulated with a maximum at day 14, a time point thatoincides with behavioral recuperation. The study furtheruggests Bcrp as a pronounced marker for the regenerativerocess and a possible functional role of Mrp5 in survivingeurons. © 2006 IBRO. Published by Elsevier Ltd. All rightseserved.
The first two authors have contributed equally to this work.Correspondence to: H. K. Kroemer, Institut für Pharmakologie, Ernst-oritz-Arndt-Universität, Friedrich-Loeffler-Str. 23d, D-17487 Greifs-ald, Germany. Tel: �49-3834-865630; fax: �49-3834-865631.-mail address: [email protected] (H. K. Kroemer).bbreviations: ABC, ATP-binding cassette protein; BBB, blood–brainarrier; BCRP, breast cancer resistance protein, ABCG2 (Bcrp: ratrtholog); MRP, multidrug resistance protein, ABCC transporter (Mrp:at ortholog); OATP, organic anion-transporting polypeptide (Oatp: ratrtholog); PCR, polymerase chain reaction; P-gp(s), P-glycoprotein(s);XR, pregnane X receptor; RECA, rat endothelial cell-specific anti-
eody; SLC, solute carrier; TTC, 2,3,5,-triphenyltetrazolium chloride;WF, von Willebrand factor.
306-4522/06$30.00�0.00 © 2006 IBRO. Published by Elsevier Ltd. All rights reseroi:10.1016/j.neuroscience.2006.07.056
1071
ey words: blood–brain barrier, stroke ischemic, diseaseodel animal, drug therapy.
he tight junctions between adjacent brain capillary en-othelial cells restrict the entry of compounds by para-ellular diffusion from the blood to the brain and viceersa, forming the blood-brain barrier (BBB). The tran-cellular transport of compounds through the membraneilayer of the brain endothelial cells is controlled by
ransport proteins expressed in these cells, includingransmembrane efflux pumps belonging to the ATP-inding cassette protein (ABC) superfamily (De Boer etl., 2003; Sun et al., 2003). P-glycoproteins (P-gps)ncoded by MDR1 (ABCB1) in human and mdr1a anddr1b (abcb1a/abcb1b) genes in rodents have been
hown to be physiologically expressed and located athe luminal membrane in capillary blood vessels of therain (Thiebaut et al., 1987; Croop et al., 1989). P-gpsctively expel a wide range of bulky lipophilic neutral orationic compounds from the brain into the blood, lead-
ng to a decrease in their uptake by the brain (Schinkel,001; Dagenais et al., 2001). The lack of P-gp indr1a/b knock-out mice affects drug transport into therain more than in any other peripheral organsSchinkel, 2001). Another member of the ABC super-amily, which was first called breast cancer resistancerotein (BCRP) or mitoxantrone resistance proteinMXR) and is now designated ABCG2, confers resis-ance to a spectrum of compounds overlapping with-gp substrates (Haimeur et al., 2004). BCRP/ABCG2as recently identified in human, porcine, and micerain capillary endothelial cells (Cooray et al., 2002;isenblatter et al., 2003; Zhang et al., 2003; Cisterninot al., 2004) raising the question of how it influences thefflux of drugs at the BBB. Multidrug resistance proteinsMRPs), members of the C-branch of the ABC trans-orter family, mediate the export of organic anions fromells. Conjugates of lipophilic compounds with glutathi-ne, glucuronate, or sulfate are preferred substrates ofRP1-3 (Haimeur et al., 2004; Konig et al., 1999), while
yclic purine nucleotides and nucleotide analogs haveeen identified as substrates for MRP4 and MRP5ABCC4 and ABCC5) (Ritter et al., 2005). The physio-ogical functions of MRP5 remain still to be defined,owever, the finding that it extrudes cGMP from cells,uggests that it can affect the signal transduction role ofGMP by reducing its intracellular content and provide
xtracellular cGMP for paracrine function. MRP5 proteinved.

comt(MsliV
ttpbpa
dtaaftpBflspOfaifstst
r3pllpSabpMe
A
Eld
ewwws
R
BrawuctctrfTfp�(sts�
I
Scaa6cBspPc(MKb(SSni4(dTrB2dbsm
nr1wT
P. Dazert et al. / Neuroscience 142 (2006) 1071–10791072
ould be detected preferentially in smooth muscle cellsf the genitourinary tract (Nies et al., 2002), in cardio-yocytes and cardiovascular smooth muscle and endo-
helial cells (Dazert et al., 2003) and human placentaMeyer zu Schwabedissen et al., 2005). Furthermore,RP5 protein could be detected in human brain and was
hown to be localized not only in the capillary endothe-ium but also in pyramidal neurons and in astrocytesndicating a role in cell signaling (Nies et al., 2004;ogelgesang et al., 2004).
Among the uptake transporter of the organic anion-ransporting polypeptide (OATPs/solute carrier (SLC) 21A)ransporter family, Oatp2 (Slc21a5), albeit most highly ex-ressed in liver, was also shown to be expressed in the ratrain, on the basolateral membrane (blood side) of choroidlexus epithelium and in brain capillary endothelia (Gao etl., 1999; Sun et al., 2003).
Focal cerebral ischemia of particular intensity anduration may change the expression of transport pro-
eins both in capillary endothelial cells and in neuronsnd glial cells of the affected brain areas and may thuslter the permeability properties of the BBB and neural
unction. Consequences of the expression of drug effluxransporters include limiting the distribution of com-ounds administered to treat the brain damage. (Deoer et al., 2003; Sun et al., 2003) However, so far only
ew studies have addressed changes in the expressionevels and protein localization of transporters aftertroke. An effect of ischemia on P-gp activity was earlyublished by the group of Tsuji (Sakata et al., 1994;hnishi et al., 1995). The authors investigated the ef-
ects of ATP depletion on the transport of cyclosporin And doxorubicin across the BBB using transient brain
schemia in rats to deplete brain ATP content. Theyound the estimated permeability coefficients of cyclo-porin A and doxorubicin increased. However, regardinghe presence of other ABC transporters with similarubstrate specificity as Bcrp at the BBB, it is it is difficulto infer from ATP dependency on P-gp activity.
In this study, focal cerebral ischemia was produced byeversible occlusion of the right middle cerebral artery in-month-old male Sprague–Dawley rats. At different timeoints after stroke the mRNA expression levels and protein
ocalization of four selected transport proteins were ana-yzed in the border zone of the infarcted area and com-ared with the contralateral site: the uptake transporter/LC Oatp2, which has been shown to contribute to thective exchange of a variety of amphophilic compoundsetween blood and brain interstitium, and the two mostrominent drug efflux pumps P-gp and Bcrp, as well asrp5, which may mainly function in the transport of endog-nous signaling molecules (Ritter et al., 2005).
EXPERIMENTAL PROCEDURES
nimals
ighteen hours prior to surgery, male 3-month-old Sprague–Daw-ey rats were deprived of food to minimize variability in ischemic
amage that can result from varying plasma glucose levels. The sxperiments reported in this study were conducted in accordanceith the statement regarding the care and use of animals andere approved by a federal animal care committee. Every effortas made to minimize the number of animals used and theiruffering.
eversible occlusion of the middle cerebral artery
lood flow through the middle cerebral artery was transiently inter-upted in deeply anesthetized rats as previously described (Badan etl., 2003; Popa-Wagner et al., 1998). The right middle cerebral arteryas slowly lifted with a tungsten hook attached to a micromanipulatorntil blood flow through the vessel was completely stopped. Bothommon carotid arteries were then occluded by tightening pre-posi-ioned thread loops. After 70 min, the middle cerebral artery and theommon carotid arteries were re-opened, allowing full reperfusion ofhe brain. Subsequent to survival times of 3, 7, 14, or 28 days, theats were deeply anesthetized and perfused with buffered salineollowed by buffered, 4% freshly depolymerized paraformaldehyde.he brain was removed, post-fixed in 4% buffered paraformaldehyde
or 24 h, cryoprotected in 20% sucrose prepared in 10 mmol/Lhosphate-buffered saline, flash-frozen in isopentane and stored at70 °C until sectioning. For real-time polymerase chain reaction
PCR) the brains were perfused with buffered saline, cut into 2 mmlices that were dipped in 2,3,5,-triphenyltetrazolium chloride (TTC)o visualize the infarct core. This procedure allowed us to microdis-ect the periinfarcted area (see Fig. 1a) that was then stored at70 °C until use.
mmunohistochemistry and quantification
ections (25 �m-thick) were cut on a freezing microtome and pro-essed for immunohistochemistry as free-floating material. Using theutomated staining device (PCT, Device assembly for preparing andnalyzing tissue for microscopic examinations; patents: DE 199 4521 A1 and WO 01/22052 A1, 2001), immunohistochemistry wasonducted as previously described (Popa-Wagner et al., 1999).riefly, after incubation with blocking solutions containing 3% donkeyerum/10 mmol/L PBS/0.3% Tween 20, tissue sections were ex-osed overnight at 4 °C to mouse monoclonal antibodies against-gp and Bcrp (C219, 1:200, and BXP-21, 1:500, Alexis Biochemi-als, Gruneberg, Germany), mouse anti NeuN monoclonal antibody1:1000, Chemicon, Pittsburgh, PA, USA), rabbit polyclonal anti-RP5 antibody AMF (kindly provided by Prof. D. Keppler, Deutschesrebsforschungszentrum, Heidelberg, Germany), and characterizedefore (Dazert et al., 2003; Jedlitschky et al., 2000; Nies et al., 2002)1:2000), and rabbit anti-Oatp2 serum (kindly provided by Dr. B.tieger, Division of Clinical Pharmacology and Toxicology, Zürich,witzerland) (Reichel et al., 1999), all diluted in PBS containing 3%ormal donkey serum and 0.3% Tween 20. After extensive washing
n PBS containing 0.3% Tween, sections were incubated overnight at°C with the respective biotinylated donkey secondary antibody
Jackson ImmunoResearch Laboratories, West Grove, PA, USA)iluted 1:400 in PBS containing 1% normal donkey serum and 0.3%ween 20. After washing in PBS, sections were incubated for 2 h at
oom temperature in ABC Elite reagent (Vectastain Elite Kit, Vector,urlingame, CA, USA) diluted 1:100 in PBS containing 0.3% Tween0. The antibody complex was then visualized with 0.025% 3=,3=-iaminobenzidine and 0.005% hydrogen peroxide in 100 mmol/L Trisuffer (pH 7.5) for 5–10 min. Finally, the sections were mounted ontolides, air-dried, and coverslipped using a xylene-based mountingedium.
A quantitative estimate of the number of transporter immu-opositive cells was obtained by counting the cells in systematicandom series of area units measuring 250�250 �m, in every0th section throughout the infarcted volume using the NIH soft-are PC Image for particle counting (NIH, Bethesda, MD, USA).he diameter of the cell was chosen in a range covering the
mallest and the largest cells. The counting area covered 30% of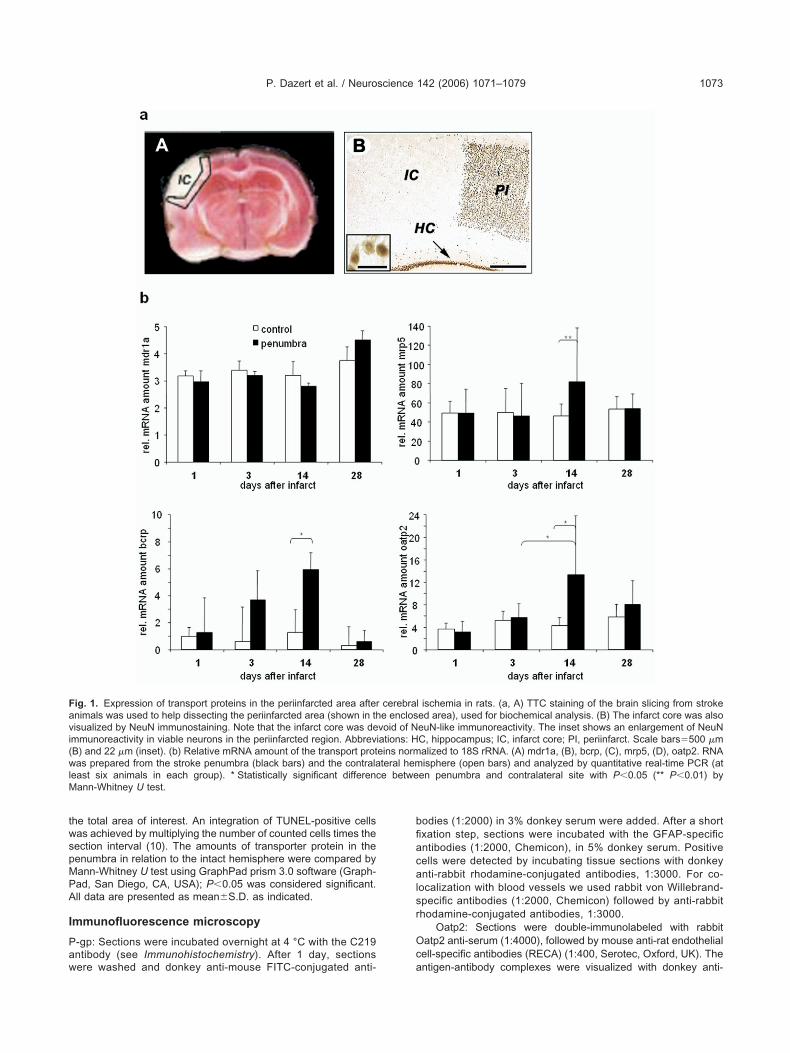
twspMPA
I
Paw
bfiacalsr
Oc
Favi(wlM
P. Dazert et al. / Neuroscience 142 (2006) 1071–1079 1073
he total area of interest. An integration of TUNEL-positive cellsas achieved by multiplying the number of counted cells times theection interval (10). The amounts of transporter protein in theenumbra in relation to the intact hemisphere were compared byann-Whitney U test using GraphPad prism 3.0 software (Graph-ad, San Diego, CA, USA); P�0.05 was considered significant.ll data are presented as mean�S.D. as indicated.
mmunofluorescence microscopy
-gp: Sections were incubated overnight at 4 °C with the C219ntibody (see Immunohistochemistry). After 1 day, sections
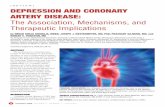
ig. 1. Expression of transport proteins in the periinfarcted area afternimals was used to help dissecting the periinfarcted area (shown in thisualized by NeuN immunostaining. Note that the infarct core was demmunoreactivity in viable neurons in the periinfarcted region. AbbrevB) and 22 �m (inset). (b) Relative mRNA amount of the transport protas prepared from the stroke penumbra (black bars) and the contrala
east six animals in each group). * Statistically significant differencann-Whitney U test.
ere washed and donkey anti-mouse FITC-conjugated anti- a
odies (1:2000) in 3% donkey serum were added. After a shortxation step, sections were incubated with the GFAP-specificntibodies (1:2000, Chemicon), in 5% donkey serum. Positiveells were detected by incubating tissue sections with donkeynti-rabbit rhodamine-conjugated antibodies, 1:3000. For co-
ocalization with blood vessels we used rabbit von Willebrand-pecific antibodies (1:2000, Chemicon) followed by anti-rabbithodamine-conjugated antibodies, 1:3000.
Oatp2: Sections were double-immunolabeled with rabbitatp2 anti-serum (1:4000), followed by mouse anti-rat endothelialell-specific antibodies (RECA) (1:400, Serotec, Oxford, UK). The
l ischemia in rats. (a, A) TTC staining of the brain slicing from strokeed area), used for biochemical analysis. (B) The infarct core was alsoeuN-like immunoreactivity. The inset shows an enlargement of NeuNC, hippocampus; IC, infarct core; PI, periinfarct. Scale bars�500 �malized to 18S rRNA. (A) mdr1a, (B), bcrp, (C), mrp5, (D), oatp2. RNAisphere (open bars) and analyzed by quantitative real-time PCR (at
en penumbra and contralateral site with P�0.05 (** P�0.01) by
cerebrae enclos
void of Niations: Heins normteral heme betwe
ntigen-antibody complexes were visualized with donkey anti-

rm
sNbcc
1gCc
(
cgbGssE
R
TRPhB8(eFPGA(gtpp(a7wmOl(ao
dd1tt
T
Fcrr
a1gspf1Pmipa
cuPpcfibcasvdfw
T
WmpfBabsFaAtplBtddip
T
Tt2(lcb
P. Dazert et al. / Neuroscience 142 (2006) 1071–10791074
abbit Cy3-conjugated antibodies (1:2000) and donkey anti-ouse Cy2-conjugated antibodies (1:1000), respectively.
Mrp5: Sections were double-immunolabeled with the MRP5-pecific polyclonal antibody AMF (1:2000) followed by mouseeuN-specific antibodies (1:1000, Chemicon). The antigen–anti-ody complexes were visualized with donkey anti-rabbit Cy3-onjugated antibodies (1:2000) and donkey anti-mouse FITC-onjugated antibodies (1:1000), respectively.
Bcrp: Sections were double-immunolabeled with BXP-21 (1:000) and rat anti-BrdU antibodies (1:2000; Serotec). The anti-en–antibody complexes were visualized with donkey anti-mousey3-conjugated antibodies (1:3000) and donkey anti-rat FITC-onjugated antibodies (1:2000) respectively.
All secondary antibodies were from the Jackson LaboratoriesDianova, Hamburg, Germany).
Fluorescence signals were detected using a Nikon E800 mi-roscope at excitation/emission wavelengths 490/508 nm (FITC,reen), 570/590 nm (rhodamine, red), and 650/668 nm (Cy5,lue). Results were recorded with a Visitech Systems (Munich,ermany) digital camera. For a group of micrographs, the cameraetting for exposure, gain, and contrast enhancement was theame. Confocal analysis of sections was done using a Nikonclipse microscope equipped with a laser device from Visitech.
eal-time PCR
otal RNA was isolated from frozen tissue homogenate using aNeasy Mini extraction kit (Qiagen, Hilden, Germany). For real-timeCR, 200 ng of total RNA was reverse-transcribed using randomexamers and the TaqMan reverse transcription reagents (Appliediosystems, Weiterstadt, Germany). Real-time PCR were set up withng of reverse-transcribed RNA for mdr1a, mrp5 (Abcc5), bcrp
Abcg2), oatp2 (Slc21a5), vWF and 18S rRNA. Intron-spanning prim-rs and the FAM-labeled probes were as follows: Mdr1a (AF257746)P-CCCATGGCCGGAACAGT; RP-ATGGGCTCCTGGGACACA;robe-TGGCTCCGCGCCCACCTG; Mrp5 (AB020209) FP-ATG-TCCTCTTCATGCTCAATGT; RPGTCCCTCATGCTGTCACTC-CA; Probe-TGATCCAGTAGCTTAGCCACCAGGT-GCT; oatp2
Slc21a5) (NM 131906) FP-ggctctgcctgtctgagtacctt; RP-atagttggt-ctgaaccccttc; Probe-aattttccagtggcaggcttaacaac-ctctta. Amplifica-ion reactions of 18SrRNA and Abcg2 were performed using theredeveloped TaqMan assay reagent kit (Applied Biosystems). PCRroducts were amplified with the TaqMan universal PCR mastermix50 °C, 2 min; 95 °C, 10 min; followed by 40 cycles of 95 °C for 15 snd 60 °C for 1 min) and analyzed on a real-time PCR cycler (SDS700, Applied Biosystems). For vWF mRNA analysis, PCR reactionas set up by mixing 10 ng of cDNA, primers (Superarray), Masterix (Superarray), and SYBR Green I (Molecular Probes, Eugene,R, USA) and real-time PCR amplification was performed as fol-
ows: one cycle of 15 min at 95 °C and 45 cycles in three steps each95 °C for 30 s, 55 °C for 30 s, 72 °C for 30 s). At the end ofmplification cycles, primer specificity was checked by appearancef single bands of PCR products in a 1% agarose gel.
A standard curve was generated by plotting the log10 [targetilution] of template on the x axis against the Ct value from serialilutions of target DNA on the y axis. Copy numbers of vWF and8S rRNA were determined based on the standard curve equa-ions. The relative copy number of vWF transcript was normalizedo 18S rRNA.
RESULTS
ime course of the P-gp expression
ocal cerebral ischemia was produced by reversible oc-lusion of the right middle cerebral artery in 3-month-oldats. After 1, 3, 14 and 28 days of reperfusion, brains were
emoved, cut into 2 mm thick slices and the periinfarcted 2rea was dissected after TTC staining as indicated in Fig.a (A). Using NeuN, a sensitive marker of neuronal de-eneration, we found that the areas most affected bytroke were the parietal cortex and, to a lesser extent, theosterior frontal cortex (Fig. 1a (B)). RNA was preparedrom the stroke penumbra and the contralateral site (Fig.b). The expression of mdr1a was analyzed by real-timeCR and normalized to 18S rRNA (Fig. 1b-A). mdr1aRNA was detectable in all samples. However, no signif-
cant difference was detected for the mRNA amount in theeriinfarcted region compared with the contralateral sitend throughout the time course.
Protein localization was analyzed by immunohisto-hemistry and confocal immunofluorescence microscopysing the monoclonal antibody C219, detecting all Mdr/-gp isoforms (Fig. 2A–C). In control animals, P-gp ex-ression was restricted to the relatively compact and intactapillaries of the contralateral, control side (Fig. 2A). In therst week after the ischemic event, a disintegration oflood vessels, concomitant with an invasion of the infarctore by astrocytes, was noted (Badan et al., 2003). Thus,t 1 day post-stroke, blood vessels were fragmented, andome P-gp immunoreactivity that co-localized with that ofon Willebrand factor (vWF), staining blood vessels, wasetected (Fig. 2B). By day 14, P-gp immunoreactivity wasound in newly generated brain capillaries in close contactith reactive astrocytes (Fig. 2C).
ime course of Bcrp/Abcg2 expression
e found a significant up-regulation of the Bcrp (Abcg2)RNA at days 3 and 14 in the periinfarcted region com-ared with the contralateral hemisphere and day 1 (4.6-old and 5.0-fold, respectively, Fig. 1b-B). By day 28, thecrp expression returned to control levels. The proteinmount followed the increased transcription as indicatedy the quantitative evaluation of the immunohistochemicaltaining (5.7-fold enhanced staining on day 14; P�0.004;ig. 2M–P; Fig. 3A;). In control animals, Bcrp was localizedlong the wall of the lateral and 3rd ventricle (Fig. 2M).lthough an ischemic insult does not significantly change
he expression of Bcrp in the ventricular wall, some Bcrp-ositive cells are in close contact with BrdU-positive, pro-
iferating cells (Fig. 2N, green color) suggesting that newcrp-positive cells (Fig. 2N, red color) may have arisen. In
he lesioned hemisphere, moderate Bcrp expression wasetected in surviving neurons in the periinfarcted area atays 3–7 (Fig. 2O). Bcrp expression in recovering neurons
n the periinfarcted region reached a maximum at day 14ost-stroke (Fig. 2P, and the inset).
ime course of Mrp5 expression
he mrp5 mRNA expression did not change significantly inhe first week post-stroke. However, on day 14 we noted a.4-fold increased transcription compared with the control2.0-fold compared with day 1; P�0.05; Fig. 1b-C). Mrp5ocalization was analyzed in the immunofluorescence mi-roscopy using the polyclonal antibody AMF, which haseen characterized before (Dazert et al., 2003; Nies et al.,
002) (Fig. 2D–G). Mrp5 was mainly detected in neurons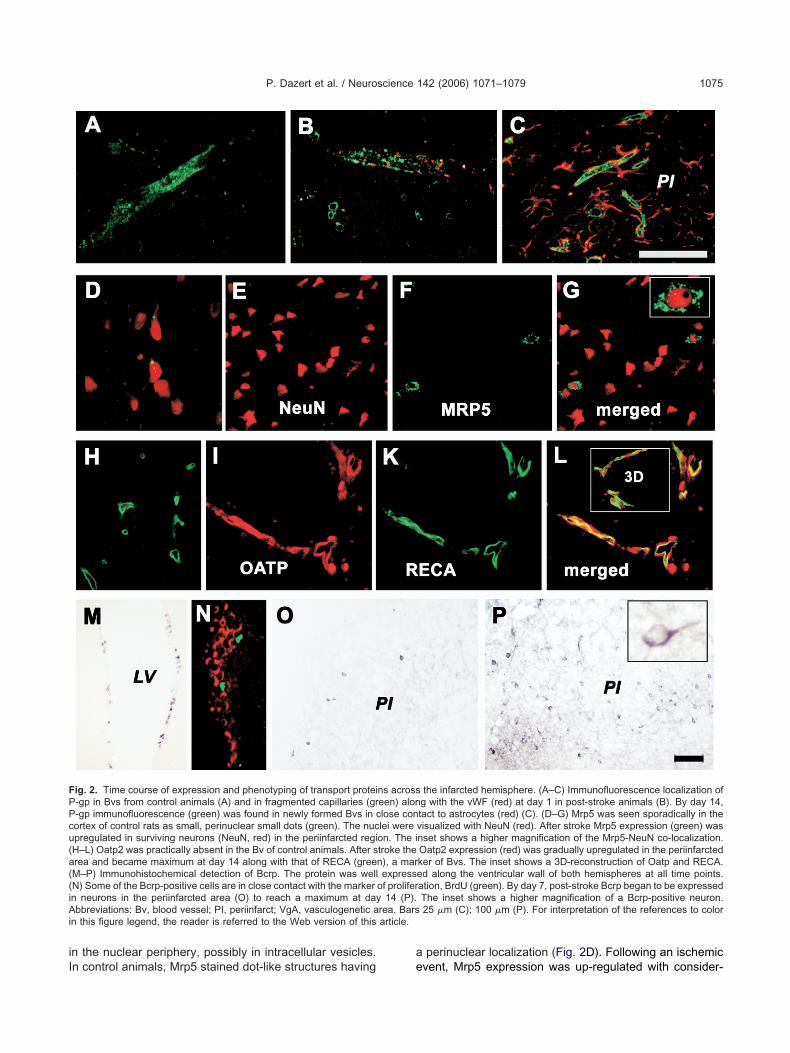
iI
a
FPPcu(a((iAi article.
P. Dazert et al. / Neuroscience 142 (2006) 1071–1079 1075
n the nuclear periphery, possibly in intracellular vesicles.
ig. 2. Time course of expression and phenotyping of transport protei-gp in Bvs from control animals (A) and in fragmented capillaries (gr-gp immunofluorescence (green) was found in newly formed Bvs in cortex of control rats as small, perinuclear small dots (green). The nucpregulated in surviving neurons (NeuN, red) in the periinfarcted regiH–L) Oatp2 was practically absent in the Bv of control animals. After srea and became maximum at day 14 along with that of RECA (greeM–P) Immunohistochemical detection of Bcrp. The protein was wellN) Some of the Bcrp-positive cells are in close contact with the markern neurons in the periinfarcted area (O) to reach a maximum at daybbreviations: Bv, blood vessel; PI, periinfarct; VgA, vasculogenetic a
n this figure legend, the reader is referred to the Web version of this
n control animals, Mrp5 stained dot-like structures having e
perinuclear localization (Fig. 2D). Following an ischemicthe infarcted hemisphere. (A–C) Immunofluorescence localization ofg with the vWF (red) at day 1 in post-stroke animals (B). By day 14,tact to astrocytes (red) (C). (D–G) Mrp5 was seen sporadically in thevisualized with NeuN (red). After stroke Mrp5 expression (green) wasnset shows a higher magnification of the Mrp5-NeuN co-localization.Oatp2 expression (red) was gradually upregulated in the periinfarctedker of Bvs. The inset shows a 3D-reconstruction of Oatp and RECA.ed along the ventricular wall of both hemispheres at all time points.ation, BrdU (green). By day 7, post-stroke Bcrp began to be expressedThe inset shows a higher magnification of a Bcrp-positive neuron.25 �m (C); 100 �m (P). For interpretation of the references to color
ns acrosseen) alonlose conlei wereon. The itroke then), a marexpress
of prolifer14 (P).
rea. Bars
vent, Mrp5 expression was up-regulated with consider-
am(tie1
T
Tfiibaittr1esct
T
TpPsv(b
Tp
fssfomOlocl2spsces
Foc ned areaa ce with P
Fp(a
P. Dazert et al. / Neuroscience 142 (2006) 1071–10791076
ble delay (day 14) and co-localized with the neuronalarker NeuN, in surviving neurons in the infarct periphery
Fig. 2E–G, and the inset). Semi-quantitative evaluation ofhe AMF staining in the immunohistochemistry (Fig. 3B)ndicated also a trend to a overall higher Mrp5 proteinxpression on day 14 compared with the control and to day, which was, however, statistically not significant.
ime course of Oatp2 (Slc21A5) expression
he oatp2 mRNA levels did not change significantly in therst 7 days post-stroke. Thereafter, a significant differencen the amount of oatp2 mRNA was detected in the penum-ra at day 14 as compared with the control hemispherend day 1 (2.3-fold and 3.6-fold, respectively, P�0.01). By
mmunohistochemistry, Oatp2 protein expression in con-rol animals was virtually absent (Fig. 2H). However, afterhe ischemic event Oatp2 expression began to be up-egulated by the time vasculogenesis was initiated, i.e. day4. By immunofluorescence, Oatp2 co-localized with thendothelial cell marker, RECA, at 14 and 28 days post-troke, shown both in two-dimensional confocal micros-opy (Fig. 2I–L) and by 3D-reconstruction of sectionsaken serially along the z axis (Fig. 2L, inset).
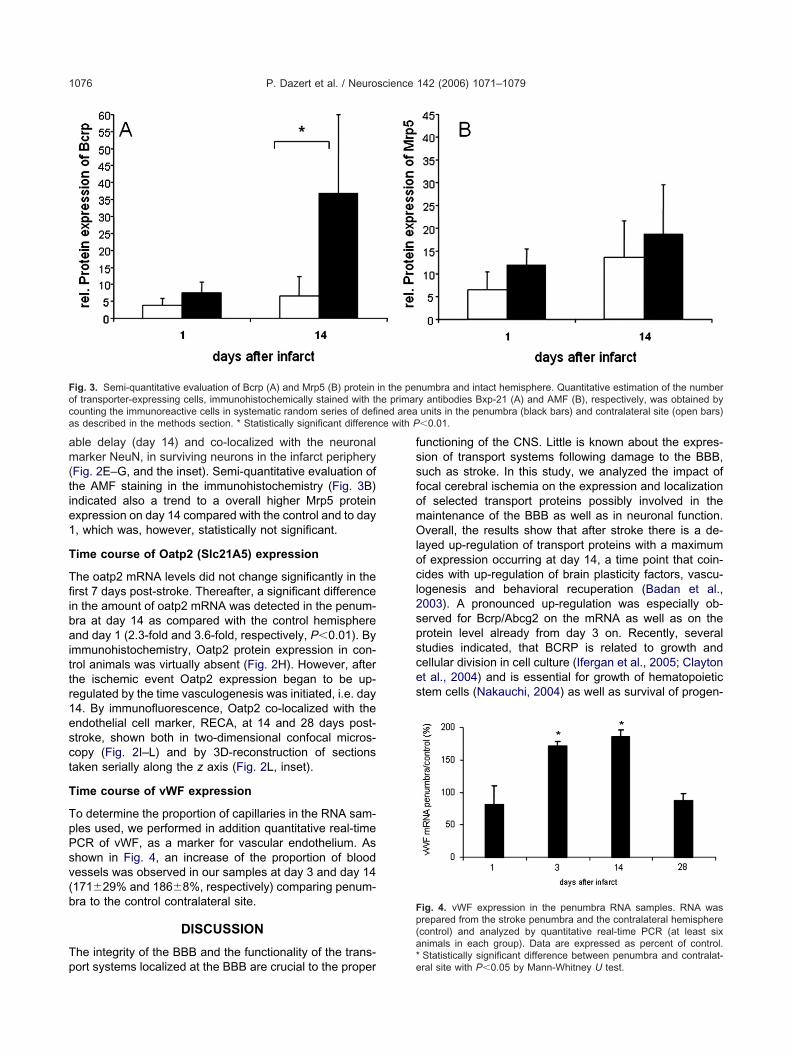
ime course of vWF expression
o determine the proportion of capillaries in the RNA sam-les used, we performed in addition quantitative real-timeCR of vWF, as a marker for vascular endothelium. Ashown in Fig. 4, an increase of the proportion of bloodessels was observed in our samples at day 3 and day 14171�29% and 186�8%, respectively) comparing penum-ra to the control contralateral site.
DISCUSSION
he integrity of the BBB and the functionality of the trans-
ig. 3. Semi-quantitative evaluation of Bcrp (A) and Mrp5 (B) proteinf transporter-expressing cells, immunohistochemically stained with thounting the immunoreactive cells in systematic random series of defis described in the methods section. * Statistically significant differen
ort systems localized at the BBB are crucial to the proper*e
unctioning of the CNS. Little is known about the expres-ion of transport systems following damage to the BBB,uch as stroke. In this study, we analyzed the impact ofocal cerebral ischemia on the expression and localizationf selected transport proteins possibly involved in theaintenance of the BBB as well as in neuronal function.verall, the results show that after stroke there is a de-
ayed up-regulation of transport proteins with a maximumf expression occurring at day 14, a time point that coin-ides with up-regulation of brain plasticity factors, vascu-
ogenesis and behavioral recuperation (Badan et al.,003). A pronounced up-regulation was especially ob-erved for Bcrp/Abcg2 on the mRNA as well as on therotein level already from day 3 on. Recently, severaltudies indicated, that BCRP is related to growth andellular division in cell culture (Ifergan et al., 2005; Claytont al., 2004) and is essential for growth of hematopoietictem cells (Nakauchi, 2004) as well as survival of progen-
umbra and intact hemisphere. Quantitative estimation of the numbery antibodies Bxp-21 (A) and AMF (B), respectively, was obtained byunits in the penumbra (black bars) and contralateral site (open bars)�0.01.
ig. 4. vWF expression in the penumbra RNA samples. RNA wasrepared from the stroke penumbra and the contralateral hemispherecontrol) and analyzed by quantitative real-time PCR (at least sixnimals in each group). Data are expressed as percent of control.
in the pene primar
Statistically significant difference between penumbra and contralat-ral site with P�0.05 by Mann-Whitney U test.

iatalapoif
oidipqo
ipceatDbtm
diecfTMemmllwn
pitliidtceab1l
Otsfie
ctpcp
lnat(KOFTsdtaaFtdkLeiTtwupttmtigp
agnasstB
AF
P. Dazert et al. / Neuroscience 142 (2006) 1071–1079 1077
tor/stem cells in the human mammary gland (Clayton etl., 2004) and murine heart (Martin et al., 2004). Localiza-ion of Bcrp in brain has been examined before (Cooray etl., 2002; Zhang et al., 2003), indicating localization in the
uminal membrane of capillaries (Cisternino et al., 2004)nd thus involvement in brain-to-blood efflux of com-ounds. In our analysis, Bcrp was localized along the wallf the lateral and 3rd ventricle but also in surviving neurons
n the periinfarcted area, an observation that deservesurther attention.
To examine, if the increase in transporter mRNA notnly reflects an increase of the proportion of blood vessels
n the samples, we determined the expression of the en-othelial marker vWF and found an approximately twofold
ncrease at day 3 and day 14. The increase in Bcrp ex-ression followed the same time course, however, wasuantitatively significantly more pronounced with a factorf about 5.
P-gp was detected mainly in newly formed capillariesn close contact with astrocytes. This is in line with arevious study by Samoto et al. (1994) which also indi-ated the activation of endothelial cell restoration and P-gpxpression by astrocytes. In the rat brain, previous studieslso described a different expression and localization ofhe P-gp isoforms Mdr1a and Mdr1b (Croop et al., 1989;evault and Gros, 1990). It should be noted that the anti-ody we used in the immunofluorescence microscopy de-ects both proteins, while we have detected only the mdr1aRNA by real-time PCR.
The localization and possible function of MRP5 areifferent from that of P-gp and BCRP. In previous studies
n the human brain, MRP5 was detected in the capillaryndothelium, but also in pyramidal neurons and in astro-ytes indicating a role in cell signaling in addition to barrierunctions (Vogelgesang et al., 2004; Nies et al., 2004).his is in line with the observations of this study detectingrp5 especially in surviving neurons in the infarct periph-ry. MRP5 may be involved in the release of nucleotideediators or other substances involved in the develop-ent of newly generated vascular or neuronal cells after
esions. The immunofluorescence images also suggest aocalization of Mrp5 in intracellular vesicular structures,hich could represent transmitter storage vesicles. Thiseeds to be clarified in further investigations.
Besides the abundance of ABC efflux pumps, the ex-ression of transport proteins of the SLC family is likely to
nfluence function of the BBB. Among these transporters,he expression of organic anion transporting polypeptidesike the human OATP-A or the rat Oatp1 and 2 is of specialnterest, because these transport proteins are probablynvolved in the uptake of opioid peptides like DPDPE andeltorphin II (Kakyo et al., 1999; Gao et al., 2000). While
he expression of Oatp1 in rat brain is controversially dis-ussed (Kusuhara et al., 2003; Choudhuri et al., 2003; Gaot al., 1999), the expression of Oatp2 (Slc22a5), which islso expressed in other tissues like liver or kidney, haseen described for the BBB and choroid plexus (Gao et al.,999). However, nothing is known so far about the regu-
ation of the expression of these proteins. In our study, r
atp2 expression was virtually not detectable in the controlissue. However, after the ischemic event, Oatp2 expres-ion began to be significantly up-regulated in the newlyormed blood vessels at a time point that is coincident withnitiation of vasculogenesis. (Schroeder et al., 2003; Badant al., 2003, 2004).
Drug delivery to the brain remains one of the mosthallenging problems of pharmacotherapy. Therefore, fur-her knowledge about the differential regulation of trans-ort proteins in brain may help to understand the pharma-okinetics of brain-targeted drugs especially under patho-hysiological conditions.
There are in part controversial studies on the regu-ation of P-gp expression at the nuclear receptor level. Aumber of xenobiotics as rifampin, phenobarbital, st-tins, and St. John’s wort can induce the MDR1 gene
hrough activation of the nuclear pregnane X receptorPXR) (recently reviewed in Klaassen and Slitt, 2005;ullak-Ublick and Becker, 2003). PXR can also regulateatp2 expression (Kullak-Ublick and Becker, 2003).urthermore, the effect of pro-inflammatory cytokines asNF�, IL1�, IL6 and INF�, has been investigated ineveral in vitro and in vivo models. The data differependent on the model, however, there is a general
rend toward decreased P-gp expression and function-lity. Several transcription factors and nuclear receptorsre probably involved in this regulation (reviewed inernandez et al., 2004). In connection with differentia-
ion processes, we could recently demonstrate that epi-ermal growth factor (EGF) induces BCRP via the MAPinase cascade (Meyer zu Schwabedissen et al., 2006).ittle is known so far about the regulation of MRP5xpression. We found an up-regulation of MRP5 upon
schemia before in human heart (Dazert et al., 2003).his up-regulation could be related to the enhanced
issue levels of the MRP5 substrate cGMP accompaniedith ischemia. NO and cGMP have been shown to reg-late expression of several genes through cGMP-de-endent protein kinase (PKG)-mediated activation of
ranscription factors as AFT-1 and NF-kappaB. Alterna-ively, MRP5 could be up-regulated together with otherembrane transporters through a more general protec-
ive response. Data from human MRP5 construct exper-ments suggest that MRP5 could be induced by thelucocorticoid receptor activators dexamethasone andrednisolone (Neo et al., 2006).
The temporal delay in the up-regulation of export pumpss P-gp and BCRP in the brain capillaries after stroke sug-ests that drug distribution to the brain in the affected area isot limited by these transporters in the first days resulting inn increased permeability of the BBB. Two weeks post-troke, however, compounds may be expelled more exten-ively in the newly formed blood vessels overexpressinghese export pumps. In addition, some transporters likeCRP and MRP5 may play a role in neurogenesis.
cknowledgments—This work was supported by the Germanederal Ministry for Education and Research (NBL3 program,
eference 01 ZZ 0103). The anti-Oatp2 serum was kindly provided
bovH
B
B
C
C
C
C
C
D
D
D
D
E
F
G
G
H
I
J
K
K
K
K
K
M
M
M
N
N
N
N
O
P
P
R
R
P. Dazert et al. / Neuroscience 142 (2006) 1071–10791078
y Dr. B. Stieger, Division of Clinical Pharmacology and Toxicol-gy, Zürich, Switzerland. The anti-MRP5 serum AMF was pro-ided by Prof. D. Keppler, Deutsches Krebsforschungszentrum,eidelberg, Germany.
REFERENCES
adan I, Dinca I, Buchhold B, Suofu Y, Walker L, Gratz M, Platt D,Kessler CH, Popa-Wagner A (2004) Accelerated accumulation ofN- and C-terminal beta APP fragments and delayed recovery ofmicrotubule-associated protein 1B expression following stroke inaged rats. Eur J Neurosci 19:2270–2280.
adan I, Platt D, Kessler C, Popa-Wagner A (2003) Temporal dynam-ics of degenerative and regenerative events associated with cere-bral ischemia in aged rats. Gerontology 49:356–365.
houdhuri S, Cherrington NJ, Li N, Klaassen CD (2003) Constitu-tive expression of various xenobiotic and endobiotic transportermRNAs in the choroid plexus of rats. Drug Metab Dispos31:1337–1345.
isternino S, Mercier C, Bourasset F, Roux F, Scherrmann JM (2004)Expression, up-regulation, and transport activity of the multidrug-resistance protein Abcg2 at the mouse blood-brain barrier. CancerRes 64:3296–3301.
layton H, Titley I, Vivanco M (2004) Growth and differentiation ofprogenitor/stem cells derived from the human mammary gland.Exp Cell Res 297:444–460.
ooray HC, Blackmore CG, Maskell L, Barrand MA (2002) Localisa-tion of breast cancer resistance protein in microvessel endotheliumof human brain. Neuroreport 13:2059–2063.
roop JM, Raymond M, Haber D, Devault A, Arceci RJ, Gros P,Housman DE (1989) The three mouse multidrug resistance (mdr)genes are expressed in a tissue-specific manner in normal mousetissues. Mol Cell Biol 9:1346–1350.
agenais C, Zong J, Ducharme J, Pollack G (2001) Effect of mdr1aP-glycoprotein gene disruption, gender, and substrate concen-tration on brain uptake of selected compounds. Pharm Res18:957–963.
azert P, Meissner K, Vogelgesang S, Heydrich B, Eckel L, Bohm M,Warzok R, Kerb R, Brinkmann U, Schaeffeler E, Schwab M,Cascorbi I, Jedlitschky G, Kroemer HK (2003) Expression andlocalization of the multidrug resistance protein 5 (MRP5/ABCC5), acellular export pump for cyclic nucleotides, in human heart. Am JPathol 163:1567–1577.
e Boer AG, Van Der Sandt IC, Gaillard PJ (2003) The role of drugtransporters at the blood-brain barrier. Annu Rev Pharmacol Toxi-col 43:629–656.
evault A, Gros P (1990) Two members of the mouse mdr gene familyconfer multidrug resistance with overlapping but distinct drug spec-ificities. Mol Cell Biol 10:1652–1663.
isenblatter T, Huwel S, Galla HJ (2003) Characterisation of the brainmultidrug resistance protein (BMDP/ABCG2/BCRP) expressed atthe blood-brain barrier. Brain Res 971:221–231.
ernandez C, Buyse M, German-Fattal M, Gimenez F (2004) Influenceof the pro-inflammatory cytokines on P-glycoprotein expressionand functionality. J Pharm Pharm Sci 7:359–371.
ao B, Hagenbuch B, Kullak-Ublick GA, Benke D, Aguzzi A, Meier PJ(2000) Organic anion-transporting polypeptides mediate transportof opioid peptides across blood-brain barrier. J Pharmacol ExpTher 294:73–79.
ao B, Stieger B, Noe B, Fritschy J, Meier P (1999) Localization of theorganic anion transporting polypeptide 2 (Oatp2) in capillary endo-thelium and choroid plexus epithelium of rat brain. J HistochemCytochem 47:1255–1264.
aimeur A, Conseil G, Deeley RG, Cole SP (2004) The MRP-relatedand BCRP/ABCG2 multidrug resistance proteins: biology, sub-strate specificity and regulation. Curr Drug Metab 5:21–53.
fergan I, Jansen G, Assaraf YG (2005) Cytoplasmic confinement of
breast cancer resistance protein (BCRP/ABCG2) as a novel mech-anism of adaptation to short-term folate deprivation. MolPharmacol 67:1349–1359.
edlitschky G, Burchell B, Keppler D (2000) The multidrug resistanceprotein 5 functions as an ATP-dependent export pump for cyclicnucleotides. J Biol Chem 275:30069–30074.
akyo M, Sakagami H, Nishio T, Nakai D, Nakagomi R, Tokui T,Naitoh T, Matsuno S, Abe T, Yawo H (1999) Immunohistochemicaldistribution and functional characterization of an organic aniontransporting polypeptide 2 (oatp2). FEBS Lett 445:343–346.
laassen CD, Slitt AL (2005) Regulation of hepatic transporters byxenobiotic receptors. Curr Drug Metab 6:309–328.
onig J, Nies AT, Cui Y, Leier I, Keppler D (1999) Conjugate exportpumps of the multidrug resistance protein (MRP) family: localiza-tion, substrate specificity, and MRP2-mediated drug resistance.Biochim Biophys Acta 1461:377–394.
ullak-Ublick GA, Becker MB (2003) Regulation of drug and bile salttransporters in liver and intestine. Drug Metab Rev 35:305–317.
usuhara H, He Z, Nagata Y, Nozaki Y, Ito T, Masuda H, Meier PJ,Abe T, Sugiyama Y (2003) Expression and functional involvementof organic anion transporting polypeptide subtype 3 (Slc21a7) in ratchoroid plexus. Pharm Res 20:720–727.
artin CM, Meeson AP, Robertson SM, Hawke TJ, Richardson JA,Bates S, Goetsch SC, Gallardo TD, Garry DJ (2004) Persistentexpression of the ATP-binding cassette transporter, Abcg2, iden-tifies cardiac SP cells in the developing and adult heart. Dev Biol265:262–275.
eyer zu Schwabedissen H, Heydrich B, Linnemann K, Fusch C,Kroemer HK, Jedlitschky G (2005) Expression, localization andfunction of MRP5 (ABCC5), a transporter for cyclic nucleotides,in human placenta and cultured human trophoblasts: effects ofgestational age and cellular differentiation. Am J Pathol 166:39 – 48.
eyer zu Schwabedissen HE, Grube M, Dreisbach A, Jedlitschky G,Meissner K, Linnemann K, Fusch C, Ritter CA, Volker U, KroemerHK (2006) Epidermal growth factor-mediated activation of the mapkinase cascade results in altered expression and function of abcg2(bcrp). Drug Metab Dispos 34:524–533.
akauchi H (2004) Isolation and clonal characterization of hematopoi-etic and liver stem cells. Cornea 23:S2–S7.
eo T, Chee K, Liu E, Tan W, Tan T (2006) Transcriptional regulationof human MRP4 and MRP5 by xenobiotics. In: ATP-binding cas-sette (ABC) proteins: from multidrug resistance to genetic dis-eases, p 123. FEBS Special Meeting, 2006, Innsbruck.
ies A, Jedlitschky G, König J, Herold-Mende C, Steiner HH, SchmittHP, Keppler D (2004) Expression and immunolocalization of themultidrug resistance proteins, MRP1-MRP6 (ABCC1-ABCC6), inhuman brain. Neuroscience 129:349–360.
ies AT, Spring H, Thon WF, Keppler D, Jedlitschky G (2002) Immu-nolocalization of multidrug resistance protein 5 in the human gen-itourinary system. J Urol 167:2271–2275.
hnishi T, Tamai I, Sakanaka K, Sakata A, Yamashima T, YamashitaJ, Tsuji A (1995) In vivo and in vitro evidence for ATP-dependencyof P-glycoprotein-mediated efflux of doxorubicin at the blood-brainbarrier. Biochem Pharmacol 49:1541–1544.
opa-Wagner A, Schroder E, Schmoll H, Walker LC, Kessler C (1999)Upregulation of MAP1B and MAP2 in the rat brain after middlecerebral artery occlusion: effect of age. J Cereb Blood Flow Metab19:425–434.
opa-Wagner A, Schroder E, Walker LC, Kessler C (1998) beta-Amyloid precursor protein and ss-amyloid peptide immunoreactiv-ity in the rat brain after middle cerebral artery occlusion: effect ofage. Stroke 29:2196–2202.
eichel C, Gao B, van Montfoort J, Cattori V, Rahner C, Hagenbuch B,Stieger B, Kamisako T, Meier PJ (1999) Localization and functionof the organic anion-transporting polypeptide Oatp2 in rat liver.Gastroenterology 117:688–695.
itter CA, Jedlitschky G, Meyer zu Schwabedissen H, Grube M, Köck
K, Kroemer HK (2005) Cellular export of drugs and signaling
S
S
S
S
S
T
V
Z
P. Dazert et al. / Neuroscience 142 (2006) 1071–1079 1079
molecules by the ATP-binding cassette transporters MRP4(ABCC4) and MRP5 (ABCC5). Drug Metab Rev 1:253–278.
akata A, Tamai I, Kawazu K, Deguchi Y, Ohnishi T, Saheki A, TsujiA (1994) In vivo evidence for ATP-dependent and P-glycoprotein-mediated transport of cyclosporin A at the blood-brain barrier.Biochem Pharmacol 48:1989–1992.
amoto K, Ikezaki K, Yokoyama N, Fukui M (1994) P-glycoproteinexpression in brain capillary endothelial cells after focal ischaemiain the rat. Neurol Res 16:217–223.
chinkel AH (2001) The roles of P-glycoprotein and MRP1 in theblood-brain and blood-cerebrospinal fluid barriers. Adv Exp MedBiol 500:365–372.
chroeder E, Vogelgesang S, Popa-Wagner A, Kessler C (2003)Neurofilament expression in the rat brain after cerebral infarction:
effect of age. Neurobiol Aging 24:135–145.un H, Dai H, Shaik N, Elmquist WF (2003) Drug efflux transporters inthe CNS. Adv Drug Deliv Rev 55:83–105.
hiebaut F, Tsuruo T, Hamada H, Gottesman MM, Pastan I, Willing-ham MC (1987) Cellular localization of the multidrug-resistancegene product P-glycoprotein in normal human tissues. Proc NatlAcad Sci U S A 84:7735–7738.
ogelgesang S, Kunert-Keil C, Cascorbi I, Mosyagin I, Schroder E,Runge U, Jedlitschky G, Kroemer HK, Oertel J, Gaab MR, PahnkeJ, Walker LC, Warzok RW (2004) Expression of multidrug trans-porters in dysembryoplastic neuroepithelial tumors causing intrac-table epilepsy. Clin Neuropathol 23:223–231.
hang W, Mojsilovic-Petrovic J, Andrade MF, Zhang H, Ball M,Stanimirovic DB (2003) The expression and functional charac-terization of ABCG2 in brain endothelial cells and vessels. FASEB
J 17:2085–2087.(Accepted 14 July 2006)(Available online 25 September 2006)