
Human immunodeficiency virus-associated plasmablastic lymphoma
Upload
independentCategory
view
0download
0
[CANCER RESEARCH 53, 3343-3348. July 15, 1993]
Differential Expression of Cell Adhesion Molecules CD54/CDlla and CD58/CD2 by
Human Melanoma Cells and Functional Role in Their Interaction with CytotoxicCells1
Maresa Altomonte, Annunciata Gloghini, Giulio Bertela, Aldo Gasparollo, Antonino Carbone, Soldano Ferrone, andMichele Maio2
Advanced Immunotherapeutics Unit and Division of Experimental Oncology 2 [M. A., A. Ca., M. M.], Pathology [A. GÌ,A. C.], Surgical Oncology I [G. B.J, and MedicalOncology 1 ¡A.Ga.¡,C. R. O., Aviano, Italy 33081, and Department of Microbiology and Immunology. New York Medical College, Valhalla, New York 10595 [S. F.J
ABSTRACT
Immunohistochemical staining with monoclonal antibodies showed adifferential distribution of intercellular adhesion molecule 1 (ICAM-1/CD54) and lymphocyte function-associated antigen 3 (LFA-3/CD58) andtheir respective counterreceptors lymphocyte function-associated antigens1 (LFA-1/CDlla) and 2 (LFÀ-2/CD2) on ten melanoma cell lines and in 46
surgically removed metastatic melanoma lesions. CDlla and CD2 werenot detected on melanoma cells while CD54 and CD58 were coexpressedon the majority of the melanoma cell populations investigated. CD54showed a higher degree of intra- and intertumor heterogeneity than CD58.•¿�y-Interferonand/or tumor necrosis factor a upregulated the expression of
CD54 by melanoma cells, but neither modulated that of CD58 nor inducedthat of CDlla and CD2. Anti-CD54 and anti-CD58 monoclonal antibodies
partially inhibited the lysis of melanoma cells by allogeneic natural killercells, lymphokine-activated killer cells and, to a greater extent, by autol-ogous tumor-infiltrating lymphocytes. Soluble CD54 (cCD54) purified
from serum of patients with melanoma inhibited the lysis of melanomacells FO-1 by natural killer cells in a dose-dependent fashion. These resultssuggest that membrane-bound CD54 and CD58 and cCD54 play a role inhost-tumor interactions in patients with malignant melanoma and may
account for the relationship between CD54 expression in primary lesionsand the clinical course of disease.
INTRODUCTION
Several cell membrane proteins that mediate homotypic and het-erotypic cell-to-cell adhesive phenomena have been assigned to thelarge family of CAM3 (1). Among CAM, ICAM-1/CD54 with its
counterreceptor LFA-1/CDlla and LFA-3/CD58 with its counter-receptor LFA-2/CD2 are of note for their role in cell-cell interactionsrequired to generate an immune response (2). Furthermore, the het-
erotypic interaction of CD54/CDlla and CD58/CD2 moleculesstrengthens the lysis of target cells by MHC antigen-restricted CTL(3, 4), and the two ligand-receptor interacting pairs can act synergis-
tically in this phenomenon (5). In contrast, conflicting data have beenreported on the role of these CAM in the NK and LAK cell-mediatedlysis of various types of target cells (6-12).
The potential involvement of CAM in the interaction of melanomacells with the host's immune system and the previously described
Received 12/30/92; accepted 5/6/93.The costs of publication of this article were defrayed in part by the payment of page
charges. This article must therefore be hereby marked advertisement in accordance with18 U.S.C. Section 1734 solely to indicate this fact.
1This work was supported by the Associazione Italiana per la Ricerca sul Cancro and
by USPHS Grant CA39559 awarded by the National Cancer Institute.2 To whom requests for reprints should be addressed, at Advanced Immunotherapeutics
Unit, C. R. O., Aviano, Italy 33081.3 The abbreviations used are: CAM, cell adhesion molecules; APAAP, alkaline
phosphatase-antialkaline phosphatase; CTL, cytotoxic T-lymphocytes; DTAF, di-chlorotriazynylaminofluorescein; ELISA, enzyme-linked immunosorbent assay; PCS,fetal calf serum; ICAM-1, intercellular adhesion molecule 1; IFN, interferon; IgG,immunoglobulin G; IIP, indirect immunofluorescence; IL, interleukin; LAK, lymphokine-activated killer; LFA, lymphocyte function-associated antigen; mAb, monoclonal antibody; MACAM, melanoma-associated cell adhesion molecules; MHC, major histocom-patibility complex; NK, natural killer; PBMC, peripheral blood mononuclear cells; PBS,phosphate-buffered saline; rIFN, recombinant interferon; rIL, recombinant interleukin;rTNF, recombinant tumor necrosis factor; TIL, tumor-infiltrating lymphocytes; TNF,
tumor necrosis factor.
association of the clinical course of the disease with CD54 expressionin primary melanoma lesions (13-15) and with the levels of soluble
CD54 (cCD54) in the serum of patients with malignant melanoma(16, 17) have stimulated interest in the characterization of the expression of CAM in melanoma cells and of their modulation by cytokines.Therefore, in the present study, we have investigated the expression ofCD54 and CD58 and their counterreceptors CDlla and CD2 on apanel of melanoma cell lines and in a large series of surgically removed metastatic melanoma lesions. In addition, we have analyzedthe role of membrane-bound CD54 and CD58 in the lysis of mela
noma cells by allogeneic and autologous cytotoxic cells. Lastly, wehave evaluated the role of soluble CD54, purified from the serum ofpatients with melanoma, in the lysis of melanoma cells by NK cells.
MATERIALS AND METHODS
Cells. The human melanoma cell lines A375, Colo 38, FO-1, Me Wo and itshighly metastatic variant MeM 50-10, SK-MEL-19, SK-MEL-33, SK-MEL-93, 3S5, and 70-W were grown in RPMI Medium 1640 (Flow Laboratories,Inc., McLean, VA) supplemented with 10% heat-inactivated PCS (Flow) and 2mm L-glutamine.
PMBC were separated from heparinized blood of healthy volunteers byFicoll-Hypaque (Pharmacia Fine Chemicals AB, Uppsala, Sweden) density
gradient centrifugation (400 X g for 30 min) and used either as a source of NKcells or for LAK cell generation. LAK cells were generated by culturing PBMCin the continuous presence of IL-2 (1000 units/ml); half of the medium waschanged every 48 h with an equal volume of fresh medium containing IL-2.
Melanoma Lesions and Sera. Metastatic melanoma lesions were obtainedfrom 43 patients with no history of chemotherapy or immunotherapy who had
been admitted for surgery at the National Cancer Institute of Aviano, Italy.Tissues were processed within 30 min following surgical removal. Each specimen was divided into three parts under sterile conditions. One third was fixedin Bouin's solution, embedded in paraffin, and processed for routine histopa-
thology. One third was snap frozen in liquid nitrogen and stored at -80°C for
immunohistochemical analysis. The remaining tissue was used to isolate TIL
and melanoma cells as previously described and with minor modifications(18). For the generation of primary cultures of melanoma cells and for TILisolation, cells (5 X 105/ml) obtained by mechanical mincing and enzymatic
digestion of tumor specimens were seeded in T25 tissue culture flasks in RPMIMedium 1640 supplemented with 10% PCS, 10% AB-type human serum, 1%nonessential amino acids (Flow), and 2 min t-glutamine. Following a 24-hincubation at 37°Cin a 5% CO2-humidified atmosphere, culture supernatant
was harvested and spun at 400 x g for 10 min. Nonadherent cells wererecovered and used as effector cells in the cytotoxicity assay with autologousmelanoma cells.
Sera were collected from patients with melanoma and frozen at -20°C until
use.Monoclonal Antibodies and Conventional Antisera. The anti-CD54
mAbs RR1/1 and CL203.4, the anti-CDlla mAb TS1/22, the anti-CD58 mAbTS2/9, the anti-CD2 mAb TS2/18, and the anti-HLA Class I mAb W6/32 weredeveloped as described elsewhere (19-22). mAbs were purified from ascitic
fluid by sequential precipitation with caprylic acid and ammonium sulfate (23).The purity of mAb preparations was monitored by sodium dodecyl sulfate-
polyacrylamide gel electrophoresis (24) under reducing and nonreducing conditions.
3343
on June 26, 2015. © 1993 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
CELL ADHESION MOLECULES IN MELANOMA-HOST INTERACTION
DTAF F(ab')2 fragments of goat anti-mouse IgG antibodies, F(ab')2 frag
ment-specific antibodies, and ChromePure mouse immunoglobulin were pur
chased from Jackson ImmunoResearch Laboratories, Inc. (West Grove, PA).Reagents for APAAP were purchased from Dakopatts (Glosturp, Denmark).Human Recombinant Cytokines and Reagents. rIFN-a and i ll-V-v were
obtained from Hoffmann LaRoche, Inc. (Nutley. NJ). rIL-la was purchasedfrom Genzyme Biochemicals, Ltd. (Maidstone, England). rIL-2 and rTNF-a
were obtained from Cetus Corporation (Emeryville, CA). Human hemoglobinwas purchased from Sigma Chemical Co. (St. Louis, MO).
Serological Assays. IIP was performed as previously described (25). Samples were analyzed for cell surface fluorescence utilizing a FACScan flowcytometer (Becton Dickinson Immunocytometry System, Mountain View, CA)equipped with a Model 9I35C Hewlett-Packard computer (Hewlett-PackardCo., Palo Alto. CA). Fluorescence was collected by using a four-decade logarithmic amplifier. Viable cells (1 X IO4, volume gated) were collected in a list
mode fashion for data analysis. The latter was performed with Consort C32software (Becton-Dickinson). Results are expressed as the percent of positive
cells and mean values of fluorescence intensity on a logarithmic scale. Asample was classified as positive when more than 5% of the cells were stainedwith relevant mAb.
Immunohistochemical staining of cryostat tissue sections by the APAAPmethod was performed as previously described (26).
The double determinant immunoassay to measure cCD54 was performedutilizing an ELISA kit from Bender MedSystems (Wien, Austria) according tothe manufacturer's instructions.
Cytotoxicity Assays. All the assays were performed in triplicate in round-bottomed 96-well microtiter plates (Costar, Cambridge, MA) as previouslydescribed (27). After labeling with 5lCr, target cells (1 x IO4) were incubatedfor 4 h at 37°Cin a total volume of 200 /il of RPMI Medium 1640 containing
10% FCS, mAb (10 /xg/ml), and different numbers of effector cells to obtain100:1 effectortarget ratios in the NK- and 20:1 ratios in the LAK- and TIL-
mediated cytotoxicity assay. Plates were then centrifuged for 5 min at 30 x gand supernatant (100 (xl) was harvested from each well and counted in agamma counter. The percentage of cytotoxicity was calculated utilizing the
formula:
MCr experimental release —¿�spontaneous release
maximum release —¿�spontaneous release X 100
Spontaneous release represents the radioactivity released by 1 X IO4 targetcells labeled with "Cr during a 4-h incubation; maximum release is theradioactivity released by 1 X IO4 target cells lysed with 1% Nonidet-40
(Sigma). Only experiments with a spontaneous release lower than 15% wereused for data analysis.
Purification of cCD54. cCD54 was purified from the serum of clinicalStage IV (distant métastases)melanoma patients as described elsewhere (28).Briefly, 4 ml of patient's serum diluted 1:10 with PBS were passed through an
immunoaffmity column with anti-CD54 mAb CL203.4 coupled to cyanogenbromide-activated Sepharose 4B (Pharmacia) to a final concentration of 5mg/ml of packed resin. All chromatography steps were at 4°C.The column was
washed, and bound CD54 was eluted with 50 HIMtriethanolamine-HCl:0.15 M
NaCl (pH 9.0) and neutralized immediately. The amount of cCD54 recoveredwas determined by the double determinant immunoassay described above.
Statistical Analysis. Data were analyzed by the Student paired t test utilizing the StatWorks statistical package from Cricket Software, Inc. (Philadelphia. PA). Differences with P < 0.05 were considered statistically significant.
RESULTS
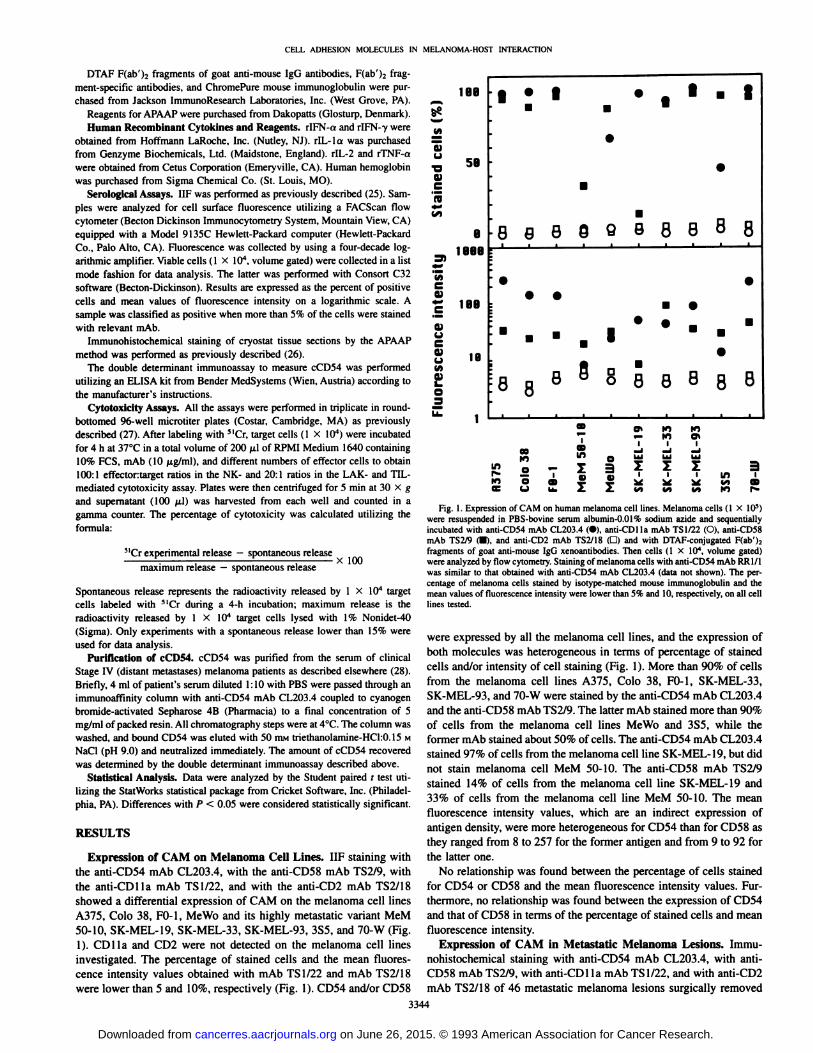
Expression of CAM on Melanoma Cell Lines. IIP staining withthe anti-CD54 mAb CL203.4, with the anti-CD58 mAb TS2/9, withthe anti-CDlla mAb TS1/22, and with the anti-CD2 mAb TS2/18
showed a differential expression of CAM on the melanoma cell linesA375, Colo 38, FO-1, MeWo and its highly metastatic variant MeM50-10, SK-MEL-19, SK-MEL-33, SK-MEL-93, 3S5, and 70-W (Fig.
1). CDlla and CD2 were not detected on the melanoma cell linesinvestigated. The percentage of stained cells and the mean fluorescence intensity values obtained with mAb TS1/22 and mAb TS2/18were lower than 5 and 10%, respectively (Fig. 1). CD54 and/or CD58
IBB2ue'ñM8>
1BBB3)'MU~
IBBu
u§
IBM£o
au.i1I
•¿�1;.•8
8 8 8Q-
900'
•¿�« .»B
8 8 8s.s
1s s 0S 1 3 11Å“
u u. Z Z•
, l .t•f888889•
•¿�*•¿�•¿�•¿�"88838er*
fofo—Kl O»
1 11_l—¿�i—¿�1LULULUIII
3i i i initt^ ^ MSu» «o M io r>
Fig. 1. Expression of CAM on human melanoma cell lines. Melanoma cells (1 X IO5)were «suspended in PBS-bovine serum albumin-0.01% sodium azide and sequentiallyincubated with anti-CD54 mAb CL203.4 (•),anti-CDlla mAb TS1/22 (O), anti-CD58mAb TS2/9 (•).and anti-CD2 mAb TS2/18 (D) and with DTAF-conjugated F(ab')2fragments of goat anti-mouse IgG xenoantibodies. Then cells (1 X IO4, volume gated)were analyzed by flow cytometry. Staining of melanoma cells with anti-CD54 mAb RR1/1was similar to that obtained with anti-CD54 mAb CL203.4 (data not shown). The percentage of melanoma cells stained by isotype-matched mouse immunoglobulin and the
mean values of fluorescence intensity were lower than 5% and 10, respectively, on all celllines tested.
were expressed by all the melanoma cell lines, and the expression ofboth molecules was heterogeneous in terms of percentage of stainedcells and/or intensity of cell staining (Fig. 1). More than 90% of cellsfrom the melanoma cell lines A375, Colo 38, FO-1, SK-MEL-33,SK-MEL-93, and 70-W were stained by the anti-CD54 mAb CL203.4and the anti-CD58 mAb TS2/9. The latter mAb stained more than 90%
of cells from the melanoma cell lines MeWo and 3S5, while theformer mAb stained about 50% of cells. The anti-CD54 mAb CL203.4stained 97% of cells from the melanoma cell line SK-MEL-19, but didnot stain melanoma cell MeM 50-10. The anti-CD58 mAb TS2/9stained 14% of cells from the melanoma cell line SK-MEL-19 and33% of cells from the melanoma cell line MeM 50-10. The mean
fluorescence intensity values, which are an indirect expression ofantigen density, were more heterogeneous for CD54 than for CD58 asthey ranged from 8 to 257 for the former antigen and from 9 to 92 forthe latter one.
No relationship was found between the percentage of cells stainedfor CD54 or CD58 and the mean fluorescence intensity values. Furthermore, no relationship was found between the expression of CD54and that of CD58 in terms of the percentage of stained cells and meanfluorescence intensity.
Expression of CAM in Metastatic Melanoma Lesions. Immunohistochemical staining with anti-CD54 mAb CL203.4, with anti-CD58 mAb TS2/9, with anti-CDl la mAb TS1/22, and with anti-CD2
mAb TS2/18 of 46 metastatic melanoma lesions surgically removed
3344
on June 26, 2015. © 1993 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
CELL ADHESION MOLECULES IN MELANOMA-HOST INTERACTION
from 43 patients showed a differential distribution of the four CAM(Table 1). Neither GDI la nor CD2 was detected in the melanomalesions tested. In contrast, CD54 and CD58 were detected in 42 of the46 tumor specimens tested (Table 1). CD54 showed a higher degree ofintra- and intertumor heterogeneity than did CD58. The percentage ofCD54-positive melanoma cells was lower than 25% in four lesions,
ranged between 2 and 50% in five and between 51 and 75% in nine,and was higher than 76% in the remaining 24 lesions. In contrast, thepercentage of CD58-positive melanoma cells ranged between 51 and
75% in two lesions and was higher than 76% in the remaining 40lesions. Moreover, the expression of CD54 for percentage of melanoma cells stained and/or intensity of staining differed in autologousmelanoma lesions removed from distinct anatomical sites from Patients Mel 8, Mel 11, and Mel 31, while the expression of CD58displayed no detectable difference (Table 1). No relationship wasfound between the percentage of melanoma cells expressing CD54
Table 1 Immunohistochemical analysis of CAM expression in metastalicmelanoma lesions
Cryostat sections of surgically removed metastatic melanoma lesions were stained withanti-CD54 mAb CL203.4, anti-CDlla mAb TSI/22, anti-CD58 mAb TS2/9. and anti-
CD2 mAb TS2/18 by APAAP.
PatientMel
1Mel2
Mel 3Mel4Mel5Mel
6Mel 7Mel8aMel
8bMel9Mel
10MelIlaMel
libMel12Mel13Mel14Mel
15Mel 16Mel 17Mel 18Mel 19Mel20Mel
21Mel22Mel23Mel24Mel
25Mel 26Mel27Mel28Mel29Mel
30Mel 3laMel31bMel
32Mel 33Mel 34Mel35Mel
36Mel37Mel38Mel39Mel40Mel
41Mel 42Mel 43Site
ofmetastasis Pigment CD54 CD Ila CD58CD2Skin
-"+-H-W*Lymph
node + +-H-Lymph node -/+ -Lvmph node -/+ ++++w
Skin++++Lymphnode - ++ -
Lymph node + +++wSkin+wLymph
node +Skin -/+++Lymph
node - +++Skin - --Lymph
node - +++ -
SkinSkin++-M-Skin
-/+ ++W++++++
-+++
NTf+++-+++
-tt+
-+++
-++W--M-+Lymph
node -/+ +++ ++w
Lymph node ++++ ++++Lymph node - ++++ ++++ -
Lymph node ++++ ++++Lymph node - ++++ ++++Muscle - +-Lymph
node + +++ NTSkin +++++WMucosa
+ ++++ - ++++W-Skin-/+ ++++ ++++WNTLymph
node -/+ ++++w ++++Lymph node + +-M-+WSkin + ++++ —¿� ++++—¿�Skin
- +++++++Lymphnode - MM - ++++ -
Lymph node - ++ - ++++ -
Skin ++++++Gut—¿�/+ ++++ —¿� ++++—¿�Lymph
node - ++++ ++++Lymph node + ++++ - ++++ -Lymph node -/+ +++ - ++++ -Skin ++++W++++Lymph
node - ++++ ++++W
Skin ++++++++Skin-++++Lymph
node - ++++ MMSkin -/+ +MMLymph
node -n-++w M MWLymph node - MM ++++WLymph node - ++++ + ++++ -
" Results were classified as - when no melanin was present in malignant cells and as
+/- or + when less than or more than 50% of malignant cells contained melanin,
respectively.* Results were classified as - when less than 5% of malignant cells were stained: as
+ when 6 to 25% of malignant cells were stained; as ++ when 26 to 50% of malignant cellswere stained; as +++ when 51 to 75% of malignant cells were stained; and as ++++ when76 to 100% of malignant cells were stained. W, weak intensity of staining.
'' NT, not tested.
leee
tea
ia
CD54
cosa
= = =4
12 24 48 72
Time (hours)
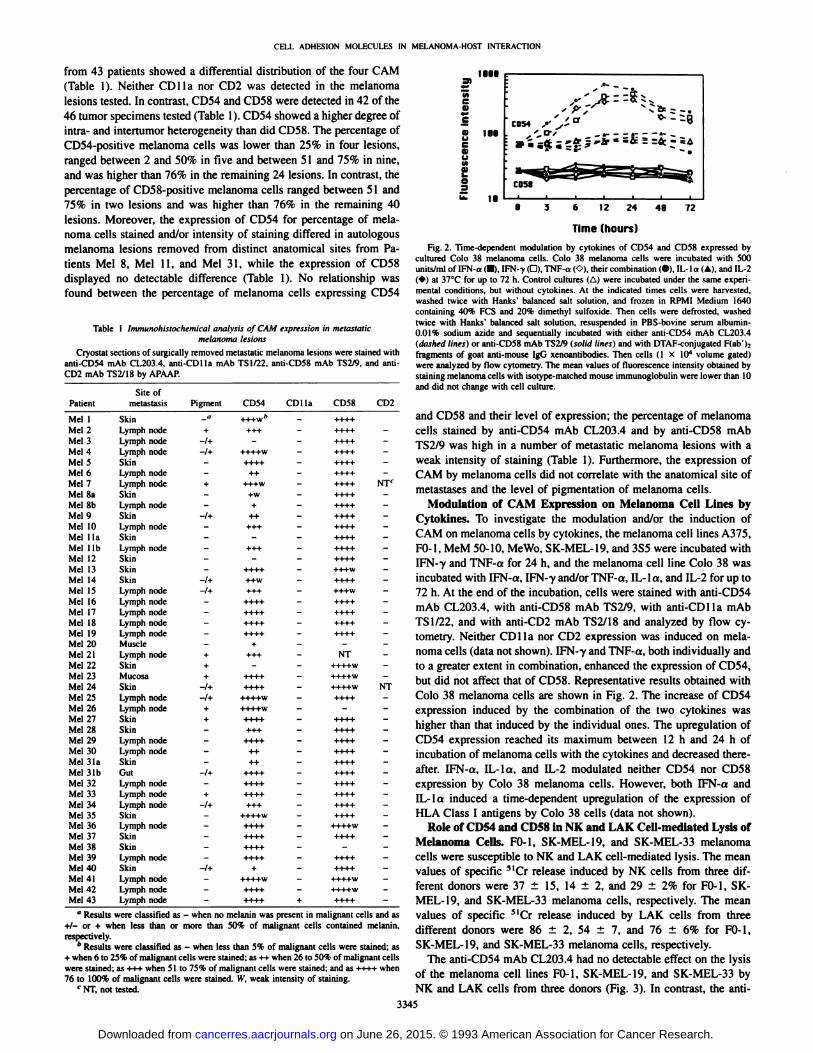
Fig. 2. Time-dependent modulation by cytokines of CD54 and CD58 expressed bycultured Colo 38 melanoma cells. Colo 38 melanoma cells were incubated with 500units/ml of IFN-a (•).IFN--y(G). TNF-a ('.•),their combination (•).IL-la (A), and 1L-2(*) at 37°Cfor up to 72 h. Control cultures (Al were incubated under the same experi
mental conditions, but without cytokines. At the indicated times cells were harvested,washed twice with Hanks' balanced salt solution, and frozen in RPMI Medium 1640
containing 40% PCS and 20% dimethyl sulfoxide. Then cells were defrosted, washedtwice with Hanks' balanced salt solution, resuspended in PBS-bovine serum albumin-
0.01% sodium azide and sequentially incubated with either anti-CD54 mAb CL203.4(dashed lines) or anti-CD58 mAb TS2/9 (solid lines) and with DTAF-conjugated F(ab'):
fragments of goat anti-mouse IgG xenoantibodies. Then cells (1 X IO4 volume gated)
were analyzed by flow cytometry. The mean values of fluorescence intensity obtained bystaining melanoma cells with isotype-matched mouse immunoglobulin were lower than 10and did not change with cell culture.
and CD58 and their level of expression; the percentage of melanomacells stained by anti-CD54 mAb CL203.4 and by anti-CD58 mAb
TS2/9 was high in a number of metastatic melanoma lesions with aweak intensity of staining (Table 1). Furthermore, the expression ofCAM by melanoma cells did not correlate with the anatomical site ofmétastasesand the level of pigmentation of melanoma cells.
Modulation of CAM Expression on Melanoma Cell Lines byCytokines. To investigate the modulation and/or the induction ofCAM on melanoma cells by cytokines, the melanoma cell lines A375,FO-1, MeM 50-10, Me Wo. SK-MEL-19, and 3S5 were incubated withIFN-7 and TNF-a for 24 h, and the melanoma cell line Colo 38 wasincubated with IFN-a, IFN-y and/or TNF-a, IL-la, and IL-2 for up to72 h. At the end of the incubation, cells were stained with anti-CD54mAb CL203.4, with anti-CD58 mAb TS2/9, with anti-CDlla mAbTSI/22, and with anti-CD2 mAb TS2/18 and analyzed by flow cy
tometry. Neither CDlla nor CD2 expression was induced on melanoma cells (data not shown). IFN--y and TNF-a, both individually and
to a greater extent in combination, enhanced the expression of CD54,but did not affect that of CD58. Representative results obtained withColo 38 melanoma cells are shown in Fig. 2. The increase of CD54expression induced by the combination of the two cytokines washigher than that induced by the individual ones. The upregulation ofCD54 expression reached its maximum between 12 h and 24 h ofincubation of melanoma cells with the cytokines and decreased thereafter. IFN-a, IL-la, and IL-2 modulated neither CD54 nor CD58expression by Colo 38 melanoma cells. However, both IFN-a andIL-lo induced a time-dependent upregulation of the expression of
HLA Class I antigens by Colo 38 cells (data not shown).Role of CD54 and CD58 in NK and LAK Cell-mediated Lysis of
Melanoma Cells. FO-1, SK-MEL-19, and SK-MEL-33 melanomacells were susceptible to NK and LAK cell-mediated lysis. The meanvalues of specific 5lCr release induced by NK cells from three dif
ferent donors were 37 ±15, 14 ±2, and 29 ±2% for FO-1, SK-MEL-19, and SK-MEL-33 melanoma cells, respectively. The meanvalues of specific 31Cr release induced by LAK cells from three
different donors were 86 ±2, 54 ±7, and 76 ±6% for FO-1,SK-MEL-19, and SK-MEL-33 melanoma cells, respectively.
The anti-CD54 mAb CL203.4 had no detectable effect on the lysisof the melanoma cell lines FO-1, SK-MEL-19, and SK-MEL-33 byNK and LAK cells from three donors (Fig. 3). In contrast, the anti-
3345
on June 26, 2015. © 1993 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
CELL ADHESION MOLECULES IN MELANOMA-HOST INTERACTION
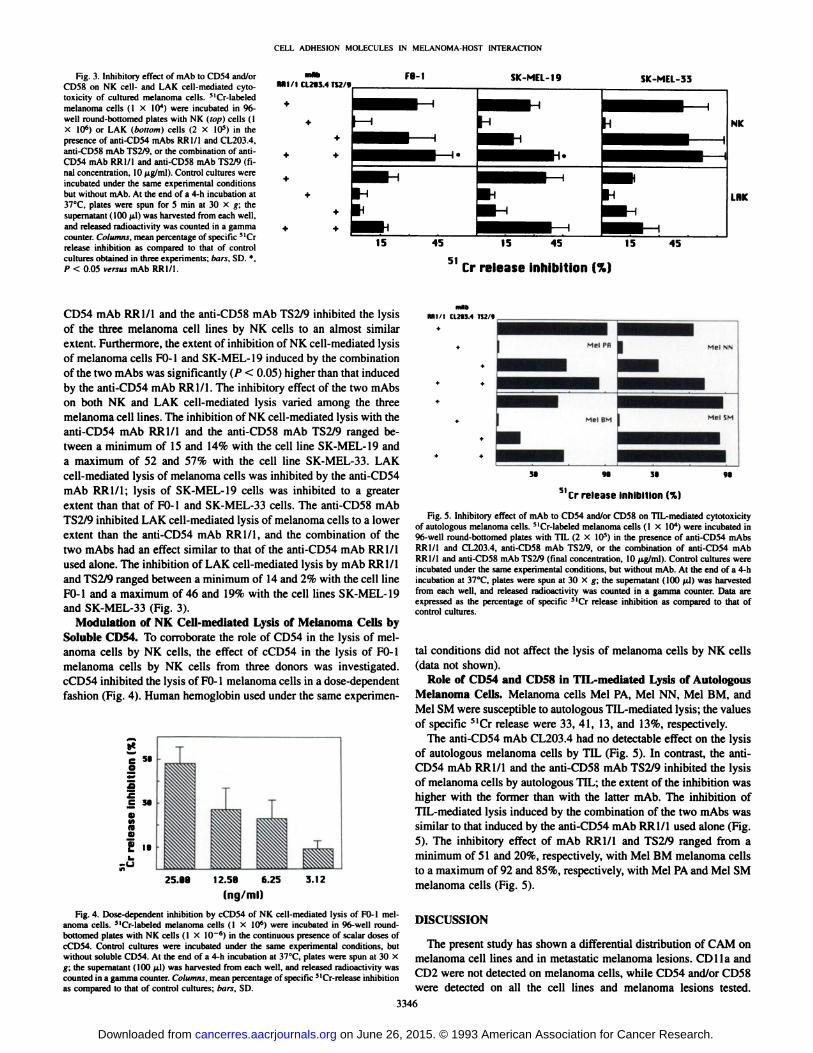
Fig. 3. Inhibitory effect of mAb to CD54 and/orCD58 on NK cell- and LAK cell-mediated cyto-toxicity of cultured melanoma cells. MCr-labeledmelanoma cells (I X IO4) were incubated in 96-well round-bottomed plates with NK (top) cells (IX IO6) or LAK (bottom) cells (2 X IO5) in thepresence of anti-CD54 mAbs RR1/1 and CL203.4,anti-CD58 mAb TS2/9. or the combination of anti-CD54 mAb RR1/I and anti-CD58 mAb TS2/9 (final concentration, 10 ^tg/ml). Control cultures wereincubated under the same experimental conditionsbut without mAb. At the end of a 4-h incubation at37°C,plates were spun for 5 min at 30 X g; the
supernatant ( 100 ^il) was harvested from each well,and released radioactivity was counted in a gammacounter. Columns, mean percentage of specific MCr
release inhibition as compared to that of controlcultures obtained in three experiments; bars. SD. *,
P < 0.05 versus mAb RR1/1
mRbMl/I CL2I3.4TS2/9
FB-1 SK-MEL-I9 SK-MEl-33
LRK
Cr release inhibition (%)
CD54 mAb RR1/1 and the anti-CD58 mAb TS2/9 inhibited the lysis
of the three melanoma cell lines by NK cells to an almost similarextent. Furthermore, the extent of inhibition of NK cell-mediated lysisof melanoma cells FO-1 and SK-MEL-19 induced by the combination
of the two mAbs was significantly (P < 0.05) higher than that inducedby the anti-CD54 mAb RR1/1. The inhibitory effect of the two mAbson both NK and LAK cell-mediated lysis varied among the threemelanoma cell lines. The inhibition of NK cell-mediated lysis with theanti-CD54 mAb RR1/1 and the anti-CD58 mAb TS2/9 ranged between a minimum of 15 and 14% with the cell line SK-MEL-19 anda maximum of 52 and 57% with the cell line SK-MEL-33. LAKcell-mediated lysis of melanoma cells was inhibited by the anti-CD54mAb RR1/1; lysis of SK-MEL-19 cells was inhibited to a greaterextent than that of FO-1 and SK-MEL-33 cells. The anti-CD58 mAbTS2/9 inhibited LAK cell-mediated lysis of melanoma cells to a lowerextent than the anti-CD54 mAb RR1/1, and the combination of thetwo mAbs had an effect similar to that of the anti-CD54 mAb RR1/1used alone. The inhibition of LAK cell-mediated lysis by mAb RR1/1
and TS2/9 ranged between a minimum of 14 and 2% with the cell lineFO-1 and a maximum of 46 and 19% with the cell lines SK-MEL-19and SK-MEL-33 (Fig. 3).
Modulation of NK Cell-mediated Lysis of Melanoma Cells by
Soluble CD54. To corroborate the role of CD54 in the lysis of melanoma cells by NK cells, the effect of cCD54 in the lysis of FO-1
melanoma cells by NK cells from three donors was investigated.cCD54 inhibited the lysis of FO-1 melanoma cells in a dose-dependentfashion (Fig. 4). Human hemoglobin used under the same experimen-
Rl/l CL2H.4 TS2/9
C 38
•¿�Mm
£ 18
2s.ee 12.SB 6.25
(ng/ml)
3.12
Fig. 4. Dose-dependent inhibition by cCD54 of NK cell-mediated lysis of FO-1 melanoma cells. 51Cr-labeled melanoma cells (1 X IO6) were incubated in 96-well round-bottomed plates with NK cells ( 1 X 10~6) in the continuous presence of scalar doses of
cCD54. Control cultures were incubated under the same experimental conditions, butwithout soluble CD54. At the end of a 4-h incubation at 37°C.plates were spun at 30 X
g; the supernatant ( 100 ¿il)was harvested from each well, and released radioactivity wascounted in a gamma counter. Columns, mean percentage of specific 5lCr-release inhibition
as compared to that of control cultures; bars, SD.
Cr release inhibition (%)
Fig. 5. Inhibitory effect of mAb to CD54 and/or CD58 on TIL-mediated cytotoxicityof autologous melanoma cells. "Cr-labeled melanoma cells (1 X IO4) were incubated in96-well round-bottomed plates with TIL (2 X IO5) in the presence of anti-CD54 mAbsRR1/1 and CL203.4, anti-CD58 mAb TS2/9, or the combination of anti-CD54 mAbRR1/1 and anti-CD58 mAb TS2/9 (final concentration. 10 fig/ml). Control cultures wereincubated under the same experimental conditions, but without mAb. At the end of a 4-hincubation at 37°C,plates were spun at 30 X g; the supernatant (100 (¿1)was harvested
from each well, and released radioactivity was counted in a gamma counter. Data areexpressed as the percentage of specific "Cr release inhibition as compared to that of
control cultures.
tal conditions did not affect the lysis of melanoma cells by NK cells(data not shown).
Role of CD54 and CD58 in TIL-mediated Lysis of Autologous
Melanoma Cells. Melanoma cells Mel PA, Mel NN, Mel BM, andMel SM were susceptible to autologous TIL-mediated lysis; the valuesof specific 51Cr release were 33, 41, 13, and 13%, respectively.
The anti-CD54 mAb CL203.4 had no detectable effect on the lysisof autologous melanoma cells by TIL (Fig. 5). In contrast, the anti-CD54 mAb RR1/1 and the anti-CD58 mAb TS2/9 inhibited the lysis
of melanoma cells by autologous TIL; the extent of the inhibition washigher with the former than with the latter mAb. The inhibition ofTIL-mediated lysis induced by the combination of the two mAbs wassimilar to that induced by the anti-CD54 mAb RR1/1 used alone (Fig.
5). The inhibitory effect of mAb RR1/1 and TS2/9 ranged from aminimum of 51 and 20%, respectively, with Mel BM melanoma cellsto a maximum of 92 and 85%, respectively, with Mel PA and Mel SMmelanoma cells (Fig. 5).
DISCUSSION
The present study has shown a differential distribution of CAM onmelanoma cell lines and in metastatic melanoma lesions. CDlla andCD2 were not detected on melanoma cells, while CD54 and/or CD58were detected on all the cell lines and melanoma lesions tested.
3346
on June 26, 2015. © 1993 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

CELL ADHESION MOLECULES IN MELANOMA-HOST INTERACTION
However, CD54 and CD58 display distinct characteristics in theirexpression by melanoma cells: CD54 is more heterogeneous thanCD58 in its distribution within and among metastatic melanoma lesions than CD58. Furthermore, CD54 is susceptible to modulation bycytokines, while CD58 is not. Lastly, CD54 is shed by melanomacells, and the shedding is enhanced by cytokines (29), while CD58was not detected in the spent medium of cultured melanoma cells.4
The distributions of the CAM tested on malignant cells of the mel-
anocytic lineage imply that interactions of melanoma cells with immune cells are mediated by CD Ila and CD2 expressed on the lattercells and by CD54 and CD58 expressed on the former cells. Inaddition, the expression of CD54 and/or CD58 on melanoma cells isof potential relevance in vivo, since effector cells may use either oneof the two molecules for their contact with neoplastic cells.
The functional involvement of CD54 and CD58 in the lysis ofmelanoma cells by cytotoxic cells is demonstrated by the ability of theanti-CD54 mAb RR1/1 and of the anti-CD58 mAb TS2/9 to inhibit the
lysis of melanoma cells by allogeneic NK and LAK cells and byautologous TIL. The combination of the two mAbs had a synergisticeffect on NK cell lysis but not on LAK cell and TIL lysis of melanomacells, suggesting that the two pathways might be differentially involved in the lysis of melanoma cells by different effector cells.Furthermore, it is noteworthy that the anti-CD54 mAb RR1/1, theanti-CD58 mAb TS2/9, and their combination did not cause a com
plete inhibition of the lysis of melanoma cells by cytotoxic cells.Moreover, the inhibition of NK and LAK cell-mediated lysis of mel
anoma cells by mAb RR1/1 and mAb TS2/9 was found to be dosedependent (data not shown) and reached its maximum by using thetwo mAbs at 10 /J.g/ml in the cytotoxicity assays. The latter findingsstrongly suggest that, besides CD54 and CD58, other CAM may beinvolved in cell-mediated lysis of melanoma cells. The involvement of
CD54 and CD58 in the lysis of melanoma cells by NK cells isnoteworthy. In fact, a lymphoid cell infiltrate with the morphologicaland immunophenotypic characteristics of NK cells has been recentlyidentified in 10 of the 16 melanoma lesions investigated (30). Therelevance of CD54 in non-MHC-restricted lysis of melanoma cells isalso supported by the observation that the anti-CD54 mAb LB2 inhibited monocyte-mediated cytotoxicity of human melanoma cells
(31). The differential role of the antigenic determinants of CD54recognized by mAb CL203.4 and mAb RR1/1 in the lysis of melanoma cells by NK cells and by TIL extends similar findings we hadpreviously obtained with LAK cells (32). In all these cytotoxic systems mAb CL203.4, which recognizes the fourth extracellular domainof CD54 (33), did not affect the lysis of melanoma cells by NK andLAK cells and by TIL while mAb RR1/1, which recognizes the firstand second extracellular domain of CD54 (33), inhibited it.
Although both CD54 and CD58 are functionally involved in thelysis of melanoma cells by cytotoxic cells, the differential distributionand susceptibility to modulation by cytokines and the shedding ofCD54 but not of CD58 by melanoma cells propose that environmentalconditions might influence the expression and shedding of CD54 butnot that of CD58 by melanoma cells and suggest that CD54, comparedto CD58, is a less stable and useful melanoma cell surface receptor forcytotoxic cells. In addition, we also found that cCD54, affinity purified from the serum of melanoma patients, inhibited the lysis ofmelanoma cells by NK cells. The levels of cCD54 are increased in theserum of melanoma patients (16) and correlate with the clinical progression of the disease (17). Therefore, cCD54 could be envisaged asa potential inhibitor of the lysis of melanoma cells by cytotoxic cells.In line with this hypothesis is the observation that soluble CD54,purified from the culture supernatant of the M19 melanoma cell line,
inhibited the lysis of M26 melanoma cells by LAK cells (34). Theseobservations, although preliminary, suggest that soluble CD54 can beconsidered a molecule that melanoma cells use in vivo to escapeimmune cell surveillance by cytotoxic cells.
The data reported in this paper demonstrate that CD54 and CD58may play a critical role in melanoma-host interaction(s), although
other MACAM may be relevant in this phenomenon. However, theshedding of MACAM in a functional form should be taken intoappropriate consideration for the evaluation of their biological significance and for their potential role in influencing the clinical course ofdisease in melanoma patients.
ACKNOWLEDGMENTS
The authors thank Dr. T. A. Springer for the gift of mAb RR1/1 andacknowledge the excellent technical and secretarial assistance of FrancescaColizzi and Paola Cattaneo.
REFERENCES
10.
12.
13.
16.
17.
18.
19.
20.4 M. Altomonte and M. Maio, unpublished observation.
Albelda. S. M., and Buck. C. A. Integrins and other cell adhesion molecules. FASEBJ.. 4: 2868-2880. 1990.
Springer, T. A. Adhesion receptors of the immune system. Nature (Lond.). 346:425-134, 1990.Shaw. S., Ginther Luce, G. E., Quiñones. R., Gress, R. E., Springer. T. A., andSanders, M. E. Two antigen-independent adhesion pathways used by human cytotoxicT-cell clones. Nature (Lond.). 323: 262-264, 1986.Makgoba, M. W.. Sanders, M. E.. Ginther Luce. G. E.. Gugel. E. A.. Dustin, M. L.,Springer. T. A., and Shaw, S. Functional evidence that intercellular adhesion molecule-1 (ICAM-1) is a ligand for LFA-1-dependent adhesion in T cell-mediated cytotoxicity. Eur. J. Immunol., 18: 637-640, 1988.Makgoba, M. W., Sanders, M. E., and Shaw. S. The CD2-LFA-3 and LFA-1-ICAMpathways: relevance to T-cell recognition. Immunol. Today. 10: 417—422, 1989.Hart, M. K., Kombluth. J.. Main. E. K., Spear, B. T., Taylor, J.. and Wilson, D. B.Lymphocyte function-associated antigen 1 (LFA-1) and natural killer (NK) cell activity: LFA-1 is not necessary for all killer: target cell interactions. Cell. Immunol.,109: 306-317. 1987.Blanchard, D. K., Michelini-Norris, M. B.. Friedman, H., and Djeu, J. Y. Lysis ofmycobacteria-infected monocytes by IL-2-activated killer cells: role of LFA-1. Cell.Immunol., 119: 402^111, 1989.Nitta, T., Vagita, H., Sato, K., and Okumura, K. Involvement of CD56 (NKH-1/Leu-19 antigen) as an adhesion molecule in natural killer-target cell interaction. J.Exp. Med.. 170: 1757-1761, 1989.Une, C., Gronberg, A., Axberg, I.. Jondal. M., and Om, A. Phospholipase C treatmentof certain human target cells reduces their susceptibility to NK lysis without affectingbinding or sensitivity to lytic granules. Cell. Immunol., 133: 127-137, 1991.Lanier, L. L., Chang, C., Azuma, M., Ruitenberg. J. J.. Hemperly, J. J., and Phillips,J. H. Molecular and functional analysis of human natural killer cell-associated neuralcell adhesion molecule (N-CAM/CD56). J. Immunol., 146: 4421^426, 1991.Quillet-Mary, A., Cavarec. L.. Kermarrec, N., Marchiol-Fournigault, C, Gil, M. L.,Conjeaud, H., and Fradelizi. D. Target lysis by human LAK cells is critically dependent upon target binding properties, but LFA-1, LFA-3, and ICAM-1 are not the majoradhesion ligands on targets. Int. J. Cancer, 47: 473-179, 1991.Zarcone. D., Viale, O., Cerniti. G.. Tenca. C.. Malorni, W., Arancia. G., losi, F,Galandrini, R., Velardi, A., Moretta, A., and Grossi, C. E. Antibodies to adhesionmolecules inhibit the lytic function of MHC-unrestricted cytotoxic cells by preventingtheir activation. Cell. Immunol., 143: 389^t04, 1992.Johnson, J. P., Stade, B. G.. Holzmann, B., Schwäble,W., and Riethmuller, G. Denovo expression of intercellular-adhesion molecule 1 in melanoma correlates withincreased risk of metastasis. Proc. Nati. Acad. Sci. USA, 86: 641-644, 1989.Carrel, S., Doré,J.-F., Ruiter. D. J., Prade, M., Lejeune, F. J., Kleeberg, U. R.. Rumke,
P., and Brocker, E. B. The EORTC melanoma group exchange program: evaluation ofa muhicenter monoclonal antibody study. Int. J. Cancer, 48: 836-847, 1991.Natali, P., Nicotra, M. R.. Cavaliere, R., Bigotti, A., Romano, G., Temponi, M., andFerrone, S. Differential expression of intercellular adhesion molecule 1 in primary andmetastatic melanoma lesions. Cancer Res.. 50: 1271-1278, 1990.Haming, R., Mainolfi, E., Bystryn. J-C. Henn, M., Merluzzi, V. J., and Rothlein, R.Serum levels of circulating intercellular adhesion molecule 1 in human malignantmelanoma. Cancer Res., 51: 5003-5005. 1991.
Altomonte, M., Colizzi, F., Esposito, G., and Maio, M. Circulating intercellularadhesion molecule 1 as a marker of disease progression in cutaneous melanoma. N.Engl. J. Med., 327: 959, 1992.Kawakami, Y., Zakut. R., Topalian, S. L.. Stotter. H., and Rosenberg, S. A. Sharedhuman melanoma antigens. Recognition by tumor-infiltrating lymphocytes in HLA-A2.l-transfected melanomas. J. Immunol., 148: 638-643, 1992.
Rothlein, R., Dustin, M. L., Marlin, S. D., and Springer, T. A. A human intercellularadhesion molecule (ICAM-1) distinct from LFA-1. J. Immunol., 137: 1270-1274,
1986.Matsui. M., Temponi. M., and Ferrone, S. Characterization of a monoclonal antibody-defined human melanoma-associated antigen susceptible to induction by immune
3347
on June 26, 2015. © 1993 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

CELL ADHESION MOLECULES IN MELANOMA-HOST INTERACTION
interferon. J. Immunol.. 39: 2088-2094, 1987.21. Sanchez-Madrid, F., Krensky, A. M., Ware, C. F., Robbins, E., Strominger, J. L.,
Burakoff, S. J., and Springer, T. A. Three distinct antigens associated with human Tlymphocyte-mediated cytolysis: LFA-1, LFA-2, and LFA-3. Proc. Nati. Acad. Sci.USA, 79: 7489-7493, 1982.
22. Barnstable, C. J., Bodmer, W. F., Brown, G., Galfre, G., Milstein, C., Williams, A. F.,and Ziegler. A. Production of monoclonal antibodies to Group A erythrocytes. HLAand other human cell surface antigens. New tools for genetic analysis. Cell, 14: 9-20,
1978.23. Temponi, M., Kageshita. T., Perosa, F., Ono. R.. Okada, H., and Ferrone. S. Purifi
cation of murine IgG monoclonal antibodies by precipitation with caprylic acid:comparison with other methods of purification. Hybridoma, 8: 85-95, 1989.
24. Laemmli, U. K. Cleavage of structural proteins during the assembly of the head ofbacteriophage T4. Nature (Lond.). 227: 680-685. 1970.
25. Maio, M., Pinto. A., Carbone. A., Zagonel, V., Gloghini. A., Marotta, G., Grillo, D.,Colombatti. A.. Ferrara, F.. Del Vecchio. L., and Ferrone. S. Differential expression ofCD54/iniercellular adhesion molecule-1 in myeloid leukemias and in lymphoprolif-erative disorders. Blood, 76: 783-790, 1990.
26. Cordell, J. L., Falini. B., Erber, W. N., Ghosh, A. K., Abdulaziz, Z., Macdonald, S.,Pulford, K. A. F., Stein, H., and Mason, D. Y. Immunoenzymatic labeling of monoclonal antibodies using immune complexes of alkaline phosphatase and monoclonalantialkaline phosphatase (APAAP complexes). J. Histochem. Cytochem., 32: 219-
229, 1984.27. Maio, M., Allomóme, M., Tatake, R., Zeff. R. A., and Ferrone, S. Reduction in
susceptibility to natural killer cell-mediated lysis of human FO-1 melanoma cells after
induction of HLA Class I antigen expression by transfection with B^m gene. J. Clin.Invest., 88: 282-289, 1991.
28. Marlin, S. D., Staunton, D. E., Springer, T. A., Stratowa, C., Sommergruber, W., andMerluzzi, V. J. A soluble form of intercellular adhesion molecule-1 inhibits rhinovirusinfection. Nature (Lond.), 344: 70-72, 1990.
29. Temponi, M.. Romano, G., D'Urso, C. M., Wang, Z., Kekish. U.. and Ferrone, S.
Profile of intercellular adhesion molecule-1 (ICAM-1) synthesized by human melanoma cell lines. Semin. Oncol., 15: 595-607, 1988.
30. Kemohan, N. M., Sewell, H. F., and Walker, F. Natural killer cells in cutaneousmalignant melanoma. J. Pathol., 161: 35^40, 1990.
31. Jonjic, N., Alberti, S., Bemasconi, S., Peri, G., Jilek, P.. Ann him. A.. Parmiani, G.,and Mantovani. A. Heterogeneous susceptibility of human melanoma clones to mono-cyte cytotoxicity: role of ICAM-1 defined by antibody blocking and gene transfer.Eur. J. Immunol., 22: 2255-2260, 1992.
32. Maio, M., Tessitori, G., Pinto, A., Temponi, M.. Colombatti, A., and Ferrone, S.Differential role of distinct determinants of intercellular adhesion molecule-1 inimmunologie phenomena. J. Immunol., 143: 181-188, 1989.
33. Staunton, D. E., Dustin, M. L., Erickson, H. P., and Springer, T. A. The arrangementof the immunoglobulin-like domains of ICAM-1 and the binding sites for LFA-1 andrhinovirus. Cell, 61: 243-254, 1990.
34. Becker, J. C., Dummer, R., Hartmann, A. A., Burg, G., and Schmidt, R. E. Sheddingof ICAM-1 from human melanoma cell lines induced by IFN-y and tumor necrosisfactor-a. J. Immunol., 147: 4398^401, 1991.
3348
on June 26, 2015. © 1993 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

1993;53:3343-3348. Cancer Res Maresa Altomonte, Annunziata Gloghini, Giulio Bertola, et al. in Their Interaction with Cytotoxic Cellsand CD58/CD2 by Human Melanoma Cells and Functional Role Differential Expression of Cell Adhesion Molecules CD54/CD11a
Updated version
http://cancerres.aacrjournals.org/content/53/14/3343
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
on June 26, 2015. © 1993 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Copyright © 2022 FDOKUMEN




















