
Differences in crop bacterial community structure between hoatzins from different geographical...
10
Differences in crop bacterial community structure between hoatzins from different geographical locations Filipa Godoy-Vitorino a,b , Sara J. Leal c , Wilmer A. Dı ´az c , Judith Rosales c , Katherine C. Goldfarb d , Marı ´a A. Garcı ´a-Amado e , Fabı ´an Michelangeli e , Eoin L. Brodie d , Marı ´a G. Domı ´nguez-Bello a, * a Department of Biology, University of Puerto Rico, P.O. Box 23360, San Juan, PR 00931, USA b Microbial Ecology Program, DOE Joint Genome Institute, Bldg 400 Rm 445Q (office), 2800 Mitchell Dr, Walnut Creek, CA 94598, USA c Centro de Investigaciones Ecolo´gicas de Guayana (CIEG), Universidad Nacional Experimental de Guayana (UNEG), Avenida Las Ame´ricas Puerto Ordaz, Estado Bolı´var, Venezuela d Ecology Department, Earth Sciences Division, Lawrence Berkeley National Laboratory, Lawrence Berkeley National Laboratory, MS 70A-3317, Berkeley, CA 94720, USA e Instituto Venezolano de Investigaciones Cientı´ficas (IVIC), San Antonio de los Altos, Carretera Panamericana, Km 11, Altos de Pipe, Estado Miranda, Venezuela Received 15 February 2011; accepted 30 December 2011 Available online xxx Abstract The hoatzin is the only known folivorous bird with foregut fermentation, and is distributed in Venezuela in rivers of the central savannas to the eastern Orinoco River. Differences in diet are expected to affect the digestive microbiota and we hypothesized that hoatzins from different habitats might have a different crop microbiota. We thus characterized the microbiota of six birds from the Cojedes and Orinoco Rivers using the G2 PhyloChip and, in parallel, we compared plant availability and foraging behavior of the hoatzins from the two locations. Plant composition differed between the 2 locations, which shared 5 out of 18 plant families and 1 plant genus e Coccoloba e that was highly consumed in both locations. The PhyloChip detected w1600 phylotypes from 42 phyla. There was a core microbiota with w50% of the OTUs shared by at least 4 of the 6 individuals, but there were also differences in the crop microbiota of animals from the two regions. There existed a higher relative abundance of Alphaproteobacteria and Actinobacteria in the crops of birds from the Cojedes River and of Clostridia and Deltaproteobacteria in the crops of birds from the Orinoco River. The results showed both a core crop microbiota and also the bacterial taxa responsible for geographical differences among individuals from the two locations with different vegetation, suggesting an effect of both diet and geography in shaping the crop bacterial community of the hoatzin. Ó 2012 Institut Pasteur. Published by Elsevier Masson SAS. All rights reserved. Keywords: Microbiota; Crop; Bird; Diet; Populations 1. Introduction The hoatzin (Opisthocomus hoazin) is a South American folivorous bird, and it is the only known bird with a well- developed fermenting crop in which digestion of dietary plant components occurs (Grajal et al., 1989). The crop ecosystem is complex and, in addition to bacteria, it also harbors archaea, protozoa and fungi (Godoy-Vitorino et al., 2006; Wright et al., 2009). As in ruminants, long digestive retention times enable hoatzin bacteria to ferment the dietary * Corresponding author. Tel.: þ1 787 7640000x4884; fax: 1 787 7643875. E-mail addresses: fi[email protected] (F. Godoy-Vitorino), [email protected] (S.J. Leal), [email protected] (W.A. Dı ´az), [email protected] (J. Rosales), [email protected] (K.C. Goldfarb), [email protected] (M.A. Garcı ´a-Amado), fmichelangeli@gmail. com (F. Michelangeli), [email protected] (E.L. Brodie), maria.dominguez1@ upr.edu, [email protected] (M.G. Domı ´nguez-Bello). Please cite this article in press as: Godoy-Vitorino, F., et al., Differences in crop bacterial community structure between hoatzins from different geographical locations, Research in Microbiology (2012), doi:10.1016/j.resmic.2012.01.001 Research in Microbiology xx (2012) 1e10 www.elsevier.com/locate/resmic + MODEL 0923-2508/$ - see front matter Ó 2012 Institut Pasteur. Published by Elsevier Masson SAS. All rights reserved. doi:10.1016/j.resmic.2012.01.001
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Differences in crop bacterial community structure between hoatzins from different geographical...

+ MODEL
Research in Microbiology xx (2012) 1e10www.elsevier.com/locate/resmic
Differences in crop bacterial community structure between hoatzins fromdifferent geographical locations
Filipa Godoy-Vitorino a,b, Sara J. Leal c, Wilmer A. Dıaz c, Judith Rosales c,Katherine C. Goldfarb d, Marıa A. Garcıa-Amado e, Fabıan Michelangeli e, Eoin L. Brodie d,
Marıa G. Domınguez-Bello a,*
aDepartment of Biology, University of Puerto Rico, P.O. Box 23360, San Juan, PR 00931, USAbMicrobial Ecology Program, DOE Joint Genome Institute, Bldg 400 Rm 445Q (office), 2800 Mitchell Dr, Walnut Creek, CA 94598, USA
cCentro de Investigaciones Ecologicas de Guayana (CIEG), Universidad Nacional Experimental de Guayana (UNEG), Avenida Las Americas Puerto Ordaz,
Estado Bolıvar, VenezueladEcology Department, Earth Sciences Division, Lawrence Berkeley National Laboratory, Lawrence Berkeley National Laboratory, MS 70A-3317, Berkeley, CA
94720, USAe Instituto Venezolano de Investigaciones Cientıficas (IVIC), San Antonio de los Altos, Carretera Panamericana, Km 11, Altos de Pipe, Estado Miranda, Venezuela
Received 15 February 2011; accepted 30 December 2011
Available online xxx
Abstract
The hoatzin is the only known folivorous bird with foregut fermentation, and is distributed in Venezuela in rivers of the central savannas tothe eastern Orinoco River. Differences in diet are expected to affect the digestive microbiota and we hypothesized that hoatzins from differenthabitats might have a different crop microbiota. We thus characterized the microbiota of six birds from the Cojedes and Orinoco Rivers using theG2 PhyloChip and, in parallel, we compared plant availability and foraging behavior of the hoatzins from the two locations.
Plant composition differed between the 2 locations, which shared 5 out of 18 plant families and 1 plant genus e Coccoloba e that was highlyconsumed in both locations. The PhyloChip detected w1600 phylotypes from 42 phyla. There was a core microbiota with w50% of the OTUsshared by at least 4 of the 6 individuals, but there were also differences in the crop microbiota of animals from the two regions. There existeda higher relative abundance of Alphaproteobacteria and Actinobacteria in the crops of birds from the Cojedes River and of Clostridia andDeltaproteobacteria in the crops of birds from the Orinoco River. The results showed both a core crop microbiota and also the bacterial taxaresponsible for geographical differences among individuals from the two locations with different vegetation, suggesting an effect of both diet andgeography in shaping the crop bacterial community of the hoatzin.� 2012 Institut Pasteur. Published by Elsevier Masson SAS. All rights reserved.
Keywords: Microbiota; Crop; Bird; Diet; Populations
* Corresponding author. Tel.: þ1 787 7640000x4884; fax: 1 787 7643875.
E-mail addresses: [email protected] (F. Godoy-Vitorino),
[email protected] (S.J. Leal), [email protected] (W.A. Dıaz),
[email protected] (J. Rosales), [email protected] (K.C. Goldfarb),
[email protected] (M.A. Garcıa-Amado), fmichelangeli@gmail.
com (F. Michelangeli), [email protected] (E.L. Brodie), maria.dominguez1@
upr.edu, [email protected] (M.G. Domınguez-Bello).
Please cite this article in press as: Godoy-Vitorino, F., et al., Differences in crop
locations, Research in Microbiology (2012), doi:10.1016/j.resmic.2012.01.001
0923-2508/$ - see front matter � 2012 Institut Pasteur. Published by Elsevier Ma
doi:10.1016/j.resmic.2012.01.001
1. Introduction
The hoatzin (Opisthocomus hoazin) is a South Americanfolivorous bird, and it is the only known bird with a well-developed fermenting crop in which digestion of dietaryplant components occurs (Grajal et al., 1989). The cropecosystem is complex and, in addition to bacteria, it alsoharbors archaea, protozoa and fungi (Godoy-Vitorino et al.,2006; Wright et al., 2009). As in ruminants, long digestiveretention times enable hoatzin bacteria to ferment the dietary
bacterial community structure between hoatzins from different geographical
sson SAS. All rights reserved.

2 F. Godoy-Vitorino et al. / Research in Microbiology xx (2012) 1e10
polysaccharides (Grajal and Parra, 1995) and provide the birdwith dietary energy to survive on an herbivorous diet. A fer-menting crop is remarkable among birds, given the body sizerestrictions on digestive fermentation. In the hoatzin, modifi-cations of the sternum accommodate the voluminous crop(Grajal, 1995). We previously characterized the crop bacterialcommunity of adult hoatzins at a single geographical locationand found that 94% of species-level OTUs were novel at thetime (Godoy-Vitorino et al., 2008).
The hoatzin is a selective feeder that browses on freshyoung shoots and leaves of up to 52 plant species, often withhigher nitrogen and lower fiber content than mature leaves(Grajal et al., 1989). Many of the neo-tropical plants containtoxic chemicals. We had previously shown that the hoatzinmicrobiota can detoxify plant defense compounds such astannins (Jones et al., 2000) and saponins (Garcıa et al., 2003).That diet can modulate the gut microbial composition has beenevidenced in the human intestine (Hayashi et al., 2002), themouse intestine (Apajalahti et al., 2002) and the cow rumen(Tajima et al., 2001). However, variation in the hoatzin cropmicrobiota due to dietary changes across temporal or spatialdimensions had not been previously addressed.
We hypothesized that the crop microbial communities aremore similar within than between hoatzin populations. To testthis hypothesis, we studied two groups of hoatzins living intwo different habitats in Venezuela, compared plant avail-ability and diet and characterized the crop bacterialcommunities.
2. Materials and methods
2.1. Foraging behavior of the hoatzin
The field study was performed in the Venezuelan states ofCojedes (8� 570 000 N, 68� 170 000 W) and Bolivar (7� 180 3600
North, 63� 250 1000 West), nearly 500 km apart. In terms ofclimate, the mean annual temperature at both sites is 27.5 �C(J. Rosales, personal communication, November 20, 2010) andthe mean precipitation is w1400 mm/year in Cojedes(Espinoza et al., 1993) and w1200 mm/year in Orinoco(J. Rosales, personal communication, November 20, 2010).Observational work on plant diversity and foraging activity inthe Orinoco River was done in the San Ignacio Stream, nearLas Galderas town, Bolıvar State in August 2008. Identifica-tion of tree species diversity was done in two 100 m lineartransects, one in each Orinoco River margin where hoatzinnested. Data on hoatzin foraging activity in the Orinoco wasobtained from 4:30 to 7:00 a.m. and from 3:30 to 6:30 p.m. forfive consecutive days. Tree diversity and dietary choices ofhoatzins from the Orinoco River were compared to previouslypublished data from the Cojedes River (Domınguez-Belloet al., 1994), considering the same transect size.
2.2. Crop sample collection
Six adult hoatzins were captured during the rainy season,three in the Orinoco River (San Ignacio stream, Las Galderas;
Please cite this article in press as: Godoy-Vitorino, F., et al., Differences in crop
locations, Research in Microbiology (2012), doi:10.1016/j.resmic.2012.01.001
coded Ga1-3) and three in the Cojedes River (coded Co 1e3), asallowed by the permits obtained from the Venezuelan Ministryof Environment (permit numbers 3187 and 1224). Six animalsrepresent a small sample size, but since each animal harborsa bacterial community, microbiome studies typically involvesmall number of hosts, including our own studies on the hoatzin(Godoy-Vitorino et al., 2010). The sampling protocol wasapproved by the UPR-IACUC (601-2007). Captures were donein the early morning by shooting the animals that were roostingin the top branches of trees. We immediately dissected the cropin situ. Crop contents with pieces of leaves and stems still intactindicated that birds had recently eaten. Whole crops with theircontents were sealed (tied with a cord) in their posterior andanterior ends and stored in 95%ethanol for less than 5weeks andtransported to the University of Puerto Rico where DNA wasextracted. All samples were treated in the same way; thus, inter-individual comparisons are valid.
With the purpose of confirming some of the rare taxadetected by the PhyloChip, we captured four additionalanimals from the Cojedes River (since we lacked enoughmaterial from the animals in this study) and immediatelydissected the crops in situ. Whole crops were frozen in liquidnitrogen and stored at �80 �C until DNA extraction.
2.2.1. DNA extractionDNA from crop contents of all six captured individuals was
extracted using the QIAamp DNA stool mini kit (Qiagen,Valencia, CA), with a modified first step where we mixed 0.5 gof sterile 0.1 mm-diameter zirconium beads (Biospec Prod-ucts, Barttlesville, Ok, USA), w200 mg of single cropcontents (previously pressed against sterilized filter paper forw20 s for alcohol absorption) and 1 ml Qiagen buffer ASL.The mixture was homogenized in a beadbeater (BiospecProducts), set at 5000 rpm for 2 min at room temperature formicrobial cell disruption. From step 3 onwards we followedthe exact recommendations of the manufacturer. The extractedDNA was frozen (w�20 �C) until use.
2.3. Amplification of 16S rRNA genes
Polymerase chain reactions were performed using ribo-somal universal bacterial primers 27F (50-AGR GTT TGATCM TGG CTC AG) and 1492R (50-GGT TAC CTT GTTACG ACT T) (Lane, 1991). The PCR mix of 50 ml mixturescontained 25 ml of PCR Master Mix (Promega, Madison, WI,USA) with 50 units/ml Taq DNA polymerase, 400 mM of eachdNTP, 3 mM MgCl2, w50 ng of DNA template and 10 pmolof each primer. Cycling was performed with an initial dena-turation at 95 �C (3 min), followed by 25 cycles of denatur-ation at 95 �C (30 s), an annealing gradient from 48 to 58 �C(25 s), an extension at 72 �C (2 min) and a final extension at72 �C (10 min). For each sample, we pooled the PCR productsfrom the 8 different annealing temperatures and verifiedcorrect product formation by electrophoresis on a 2% agarosegel. The pooled PCR products were purified using a PCRpurification kit (Qiagen, Valencia, CA, USA), and used forhybridization onto the 16S DNA microarray.
bacterial community structure between hoatzins from different geographical

3F. Godoy-Vitorino et al. / Research in Microbiology xx (2012) 1e10
2.4. The PhyloChip DNA microarray
DNA microarrays represent an excellent choice for rapidcomparison of spatial, temporal or physiological differences inbacterial communities. They offer high sensitivity to detectlow-abundance species (Brodie et al., 2006). We used the G2PhyloChip, a 16S rRNA gene microarray developed by theLawrence Berkeley National Laboratory and synthesized byAffymetrix Inc. (Santa Clara, CA, USA). The PhyloChip hasbeen validated (Brodie et al., 2006, 2007; DeSantis et al.,2007) and is highly sensitive, detecting 90% of the clonedsubfamilies and unveiling 2.5-fold more diversity thancloning. The array had 506,944 probes arranged in 712 rowsand columns representing w8700 bacterial and archeal taxa.The PhyloChip possesses oligonucleotide probes that repre-sent w8741 OTUs (targeting 16S rRNA gene sequences).Each of the spots on the array contains probe pairs (w24)consisting of perfectly matching probes (PMs) with corre-sponding mismatch controls (MMs) representing a certaintaxa. The probes were designed, where possible, to identifyonly one sequence, but some spots contain probes that covera few or many sequences (0e3% sequence divergence). Thesesequences are then contained in an OTU and a representativeorganism/sequence is chosen for that OTU.
Each chip has additional probes that serve as controls: (1)targeting amplicons of prokaryotic and eukaryotic metabolicgenes (added just before fragmentation) and (2) pre-labeledoligonucleotides added into the hybridization mix. The firstcontrol tests the fragmentation, biotinylation, hybridization,washing, staining and scanning efficiency altogether. It alsoallows for normalization of the fluorescent intensity of allarrays in the experiment. The second control accounts forvariation in hybridization, washing, staining and scanning. Theamplicon pools for each of the six samples were fragmented to50e200 bp using DNase I (0.02 U/mg DNA; Invitrogen, USA)and One-Phor-All buffer (NJ, USA). Biotin labeling wasperformed with deoxyribonucleotransferase (Promega, USA)as per the manufacturer’s instructions. Denaturing occurredlater at 99 �C for 5 min and hybridization onto the PhyloChipoccurred overnight at 48 �C at 60 rpm. The arrays weresubsequently washed, stained and scanned. Detailed method-ology is specified elsewhere (Brodie et al., 2006).
Scanning of the arrays was performed using a GeneArrayscanner (Affymetrix, CA, USA) and the intensity of all probeswas treated as previously reported (Brodie et al., 2006).Positive probe pairs met two criteria: 1) the fluorescence of theperfectly matched probe was at least 1.3 times greater thanthe intensity of the control (mismatch probe); 2) the value ofthe difference between perfectly matched probe and mismatchprobe intensities was at least 130 times greater than thesquared noise value.
The normalized intensities were then scaled by theaverage overall array intensity to account for variation inPCR amplicon quantification, and those values were log-transformed to normalize variance at different intensities.
The normalized log-transformed intensity values were usedfor further analysis (Ivanov et al., 2009). For a taxon to be
Please cite this article in press as: Godoy-Vitorino, F., et al., Differences in crop
locations, Research in Microbiology (2012), doi:10.1016/j.resmic.2012.01.001
reported as present, 90% of probe pairs in the set must havea perfect match intensity at least 1.3 times the mismatch, anddifferences between perfect match and mismatch intensity thatare 130 times the square of noise value.
2.5. Confirmatory 454 pyrotag sequencing to 4 Cojedes-captured animals
We extracted DNA from crop contents of 4 samples with2e5 technical replicates using the MoBio Power Soil DNAExtraction Protocol with modifications. We used w250 mg ofcrop bulk fiber with 3 freeze-thawing cycles (�20 �C and65 �C) followed by a Beadbeater horizontal shaking for 40 s.The protocol that followed was as suggested by the manu-facturer. DNA was eluted in PCR water and kept frozen untiluse. Amplicons of w400 nucleotides in length were producedusing broadly conserved 16S primers 926F (50-aaactYaaaK-gaattgacgg-30)and 1392R (50-acgggcggtgtgtRc-30). To multi-plex amplicons for inclusion on a single sequencing run, thereverse primer (1392R) was bar-coded on the 50 end with fiveunique bases between the 454 A-adapter sequence (50-gcctccctcgcgccatcag-30) and the conserved 16S rRNA primersequence. The forward primer in each pair was not bar-codedbut did incorporate the 454 B-adapter (50-gccttgccagcccgctca-30) at its 50 end. The detailed polymerase chain reaction isdetailed elsewhere (Engelbrektson et al., 2010). To obtaina similar number of reads from each sample, amplicons weremixed in equal concentrations. Emulsion PCR and sequencingwere performed using a GS FLX PCR amplicon kit accordingto the manufacturer’s protocols.
2.6. Data analyses: phylogenetic and multivariateanalyses
A representative sequence of each detected taxon wasexported aligned from Greengenes (DeSantis et al., 2006) andimported to ARB for neighbor-joining tree construction.
Taxa that significantly changed between the two locationswere determined using Anova, with p-values corrected formultiple observations using the Holm procedure, a little lessstringent than Bonferroni, thus decreasing the false discovery(Storey and Tibshirani, 2003). The significantly differentOTUs in the two locations were used to draw a heatmap withdendrograms using Pearson’s correlation as the similaritymetric and average linkage clustering in “R” using thepackage made4 (Eisen et al., 1998).
Rank-abundance curves were drawn from the data tovisualize species richness and overall diversity.
We also used the Vegan package in “R” to performpermutational multivariate analysis of variance (using theAdonis function), to draw box plots to visually interpret thevariance of the data as well as to calculate the Pielou index(which measures how evenly distributed the abundance isamong the species that exist in a community). Within Vegan,we used a BrayeCurtis distance metric within the function‘vegdist’ to calculate a distance matrix from the normalizedlog-transformed intensity values of the PhyloChip OTUs, to
bacterial community structure between hoatzins from different geographical

4 F. Godoy-Vitorino et al. / Research in Microbiology xx (2012) 1e10
perform statistical analysis of similarities (ANOSIM) and tocalculate and plot the dissimilarity ranks between and withinclasses.
2.7. Bacterial community analyses
To determine the common bacterial community composi-tion, core taxa (shared OTUs between the birds in both loca-tions) were defined as taxa present in at least 4 of the 6 cropsamples. Bacterial communities from the six crops werecompared using UniFrac (Lozupone and Knight, 2005). Uni-Frac community analyses are based on an ARB neighbor-joining tree of all 16S rRNA gene sequences and an environ-ment file with the PhyloChip detected taxa for each individual.
To determine whether the individuals or age groups weresignificantly different from each other, we performed theUnifrac significance test which determines whether sequencesin a tree are significantly different from each other. To clustersamples with related bacterial communities, we clusteredenvironments and verified node support using Jackknife.
Principal components analyses was performed to determineclustering of crop bacterial communities from individualswithin a single population and between populations. We per-formed weighted analyses in which the algorithm accounts forthe relative abundances of bacterial lineages, as well as takinginto account the shared branch lengths between samples.
2.8. Taxonomic classification of the pyrosequencingflowgrams
Pyrosequencing flowgrams were converted to sequencereads using the standard software provided by 454 LifeSciences. Reads were processed using the computationalpipeline (Pyrotagger) described in Kunin and Hugenholtz(2010), consisting of read quality filtering and length trim-ming (reads shorter than 220 nt were excluded from furtheranalyses), dereplication and clustering at 97% sequenceidentity. Representative sequences from each cluster were alsoclassified in Pyrotagger by blastn comparisons againstGreengenes (DeSantis et al., 2006) and SILVA (Pruesse et al.,2007) databases for bacteria/archaea and eukaryotes respec-tively. The full taxonomy string for each reference sequencewas exported from the databases to allow taxonomic identifi-cation of pyrotag sequences. Histograms for the bacterial readswere drawn in Excel.
3. Results
3.1. Foraging behavior of the hoatzin
We first assessed the diversity of plants in the two habitatsand the foraging preferences of the birds. We compared ourfield observations obtained in the first of two e 100 m tran-sects in Orinoco with our previously published data from theCojedes River (Domınguez-Bello et al., 1994).
Plant richness was similar for the two rivers, with 24species and 17 families in the Orinoco and 26 species and 18
Please cite this article in press as: Godoy-Vitorino, F., et al., Differences in crop
locations, Research in Microbiology (2012), doi:10.1016/j.resmic.2012.01.001
families in the Cojedes River. The evenness of plantcommunities was also similar between the two locations(0.946 and 0.949 for Orinoco and Cojedes respectively; Table1). In the previous study at the Cojedes River, along 4.7 km,40% of the total number of species appeared in the first 100 mtransect and included the Coccoloba sp., Lecythis ollaria andMachaerium sp. (Domınguez-Bello et al., 1994). The first100 m transect of the Orinoco River had 24 plant speciesincluding the dominant Piranhea trifoliata (w30% out of the144 plants) and Coccoloba ovata (w13% out of the 144plants). The second 100 m transect had 12 tree species, 10 ofwhich were shared with the first transect, including thedominant C. ovata, (representing herew76% of the 219 plantsin this second transect; Table S1).
In terms of the richness and evenness of their plantcommunities, the two river locations were similar. Howeverthey differed greatly in species composition, with only five ofthe 18 families (Combretaceae, Fabaceae, Flacourtiaceae,Rubiaceae and Polygonaceae) and three plant genera (Coc-coloba, Machaerium and Combretum) shared between the twosites, and with Coccoloba being the most abundant tree genusin both rivers (Table 1).Therefore, plant availability in the tworivers was different, with the exception of two common plantfamilies, the Polygonaceae and the Fabaceae.
Field observations indicated that hoatzins chose to eatabundant tree species in both locations which, in Cojedes,included species such as Coccoloba, Machaerium sp. andCombretum fructicosum, while birds from Orinoco ate abun-dantly from Coccoloba, Nectandra pichurim and Calyp-tranthes multiflora (Table 1).
3.2. Crop bacterial communities in the hoatzin
The comparative study of the crop microbiota resulted ina total of 1639 bacterial OTUs from an average of 38 þ 3bacterial phyla found in the crop of the birds regardless of thelocation. Crop bacteria were dominated by only 4 phyla,namely Proteobacteria, Firmicutes, Bacteroidetes and Acti-nobacteria (Fig. 1A). Other detected phyla included Spiro-chetes, Synergistes or Acidobacteria and some putativelythermophilic bacteria, such as Aquificae, Coprothermobac-teria, Thermodesulfobacteria and Caldithrix (Fig. 1A).
Confirmatory PCRs using 454 pyrotags made on samplesfrom 4 birds yielded nearly 86,000 reads that were classifiedinto 21 bacterial phyla present in the crops of all 4 animals(13 samples, including technical replicates).Among the detectedphyla, Firmicutes, Bacteroidetes, Proteobacteria and Actino-bacteria were dominant, while the rare phyla includedSpirochetes, Chlorobi, Acidobacteria, TM7, Chloroflexi, Ver-rucomicrobia, Synergistes, Planctomycetes, Chlamydiae, OP9/JS1, Lentisphaerae and Gemmatimonadetes (Suppl. Fig. 1).
There were similar levels of taxonomic diversity in bothlocations according to the average number of groups detectedat each taxonomic level (from phyla to OTU) (Table 2). Thethree individuals in Cojedes had 41 bacterial phyla in the cropand those in Orinoco had 42, with Dictyoglomi present in onlyone animal (Ga3; Fig. 1A). The bars that represent pooled
bacterial community structure between hoatzins from different geographical
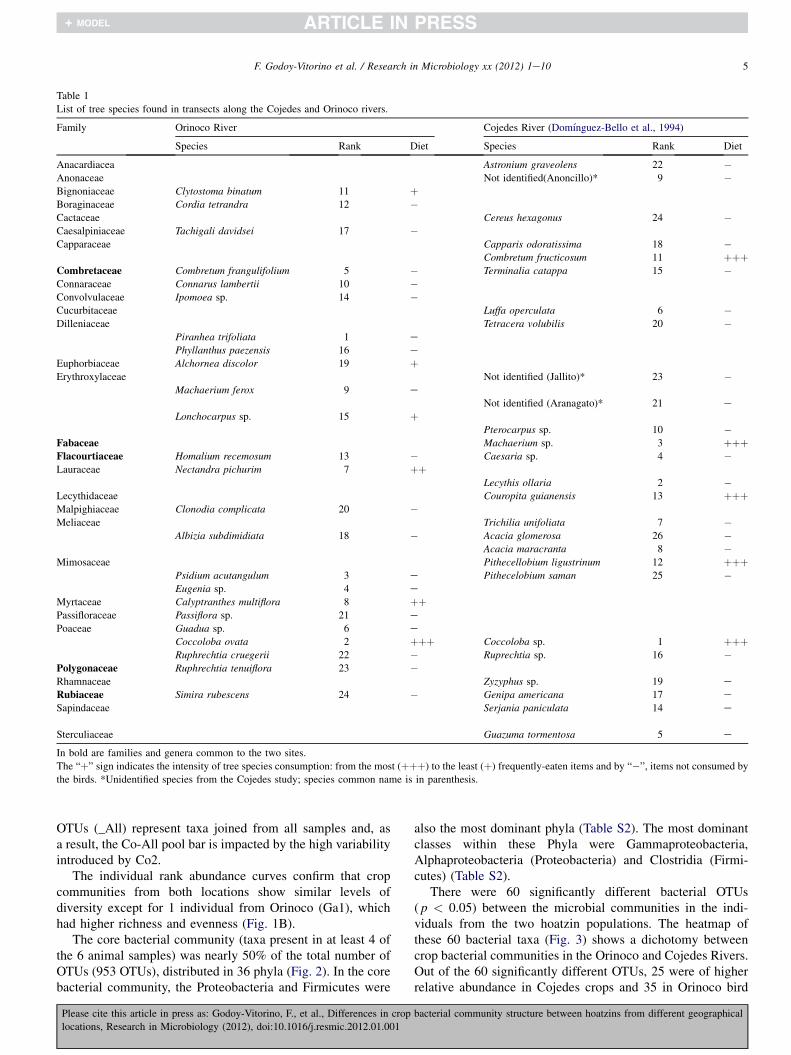
Table 1
List of tree species found in transects along the Cojedes and Orinoco rivers.
Family Orinoco River Cojedes River (Domınguez-Bello et al., 1994)
Species Rank Diet Species Rank Diet
Anacardiacea Astronium graveolens 22 �Anonaceae Not identified(Anoncillo)* 9 �Bignoniaceae Clytostoma binatum 11 þBoraginaceae Cordia tetrandra 12 �Cactaceae Cereus hexagonus 24 �Caesalpiniaceae Tachigali davidsei 17 �Capparaceae Capparis odoratissima 18 �
Combretum fructicosum 11 þþþCombretaceae Combretum frangulifolium 5 � Terminalia catappa 15 �Connaraceae Connarus lambertii 10 �Convolvulaceae Ipomoea sp. 14 �Cucurbitaceae Luffa operculata 6 �Dilleniaceae Tetracera volubilis 20 �
Piranhea trifoliata 1 ePhyllanthus paezensis 16 e
Euphorbiaceae Alchornea discolor 19 þErythroxylaceae Not identified (Jallito)* 23 �
Machaerium ferox 9 eNot identified (Aranagato)* 21 e
Lonchocarpus sp. 15 þPterocarpus sp. 10 �
Fabaceae Machaerium sp. 3 þþþFlacourtiaceae Homalium recemosum 13 � Caesaria sp. 4 �Lauraceae Nectandra pichurim 7 þþ
Lecythis ollaria 2 �Lecythidaceae Couropita guianensis 13 þþþMalpighiaceae Clonodia complicata 20 �Meliaceae Trichilia unifoliata 7 �
Albizia subdimidiata 18 � Acacia glomerosa 26 �Acacia maracranta 8 �
Mimosaceae Pithecellobium ligustrinum 12 þþþPsidium acutangulum 3 e Pithecelobium saman 25 �Eugenia sp. 4 e
Myrtaceae Calyptranthes multiflora 8 þþPassifloraceae Passiflora sp. 21 e
Poaceae Guadua sp. 6 eCoccoloba ovata 2 þþþ Coccoloba sp. 1 þþþRuphrechtia cruegerii 22 � Ruprechtia sp. 16 �
Polygonaceae Ruphrechtia tenuiflora 23 �Rhamnaceae Zyzyphus sp. 19 eRubiaceae Simira rubescens 24 � Genipa americana 17 e
Sapindaceae Serjania paniculata 14 e
Sterculiaceae Guazuma tormentosa 5 e
In bold are families and genera common to the two sites.
The “þ” sign indicates the intensity of tree species consumption: from the most (þþþ) to the least (þ) frequently-eaten items and by “�”, items not consumed by
the birds. *Unidentified species from the Cojedes study; species common name is in parenthesis.
5F. Godoy-Vitorino et al. / Research in Microbiology xx (2012) 1e10
OTUs (_All) represent taxa joined from all samples and, asa result, the Co-All pool bar is impacted by the high variabilityintroduced by Co2.
The individual rank abundance curves confirm that cropcommunities from both locations show similar levels ofdiversity except for 1 individual from Orinoco (Ga1), whichhad higher richness and evenness (Fig. 1B).
The core bacterial community (taxa present in at least 4 ofthe 6 animal samples) was nearly 50% of the total number ofOTUs (953 OTUs), distributed in 36 phyla (Fig. 2). In the corebacterial community, the Proteobacteria and Firmicutes were
Please cite this article in press as: Godoy-Vitorino, F., et al., Differences in crop
locations, Research in Microbiology (2012), doi:10.1016/j.resmic.2012.01.001
also the most dominant phyla (Table S2). The most dominantclasses within these Phyla were Gammaproteobacteria,Alphaproteobacteria (Proteobacteria) and Clostridia (Firmi-cutes) (Table S2).
There were 60 significantly different bacterial OTUs( p < 0.05) between the microbial communities in the indi-viduals from the two hoatzin populations. The heatmap ofthese 60 bacterial taxa (Fig. 3) shows a dichotomy betweencrop bacterial communities in the Orinoco and Cojedes Rivers.Out of the 60 significantly different OTUs, 25 were of higherrelative abundance in Cojedes crops and 35 in Orinoco bird
bacterial community structure between hoatzins from different geographical
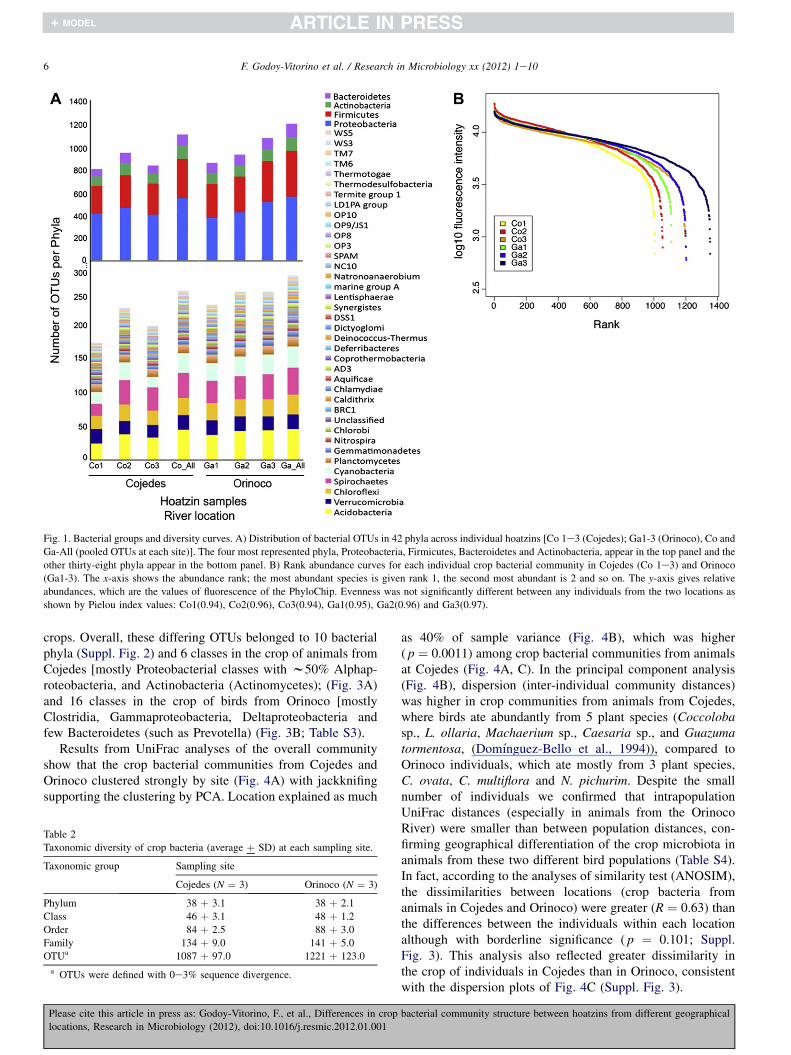
Fig. 1. Bacterial groups and diversity curves. A) Distribution of bacterial OTUs in 42 phyla across individual hoatzins [Co 1e3 (Cojedes); Ga1-3 (Orinoco), Co and
Ga-All (pooled OTUs at each site)]. The four most represented phyla, Proteobacteria, Firmicutes, Bacteroidetes and Actinobacteria, appear in the top panel and the
other thirty-eight phyla appear in the bottom panel. B) Rank abundance curves for each individual crop bacterial community in Cojedes (Co 1e3) and Orinoco
(Ga1-3). The x-axis shows the abundance rank; the most abundant species is given rank 1, the second most abundant is 2 and so on. The y-axis gives relative
abundances, which are the values of fluorescence of the PhyloChip. Evenness was not significantly different between any individuals from the two locations as
shown by Pielou index values: Co1(0.94), Co2(0.96), Co3(0.94), Ga1(0.95), Ga2(0.96) and Ga3(0.97).
6 F. Godoy-Vitorino et al. / Research in Microbiology xx (2012) 1e10
crops. Overall, these differing OTUs belonged to 10 bacterialphyla (Suppl. Fig. 2) and 6 classes in the crop of animals fromCojedes [mostly Proteobacterial classes with w50% Alphap-roteobacteria, and Actinobacteria (Actinomycetes); (Fig. 3A)and 16 classes in the crop of birds from Orinoco [mostlyClostridia, Gammaproteobacteria, Deltaproteobacteria andfew Bacteroidetes (such as Prevotella) (Fig. 3B; Table S3).
Results from UniFrac analyses of the overall communityshow that the crop bacterial communities from Cojedes andOrinoco clustered strongly by site (Fig. 4A) with jackknifingsupporting the clustering by PCA. Location explained as much
Table 2
Taxonomic diversity of crop bacteria (average þ SD) at each sampling site.
Taxonomic group Sampling site
Cojedes (N ¼ 3) Orinoco (N ¼ 3)
Phylum 38 þ 3.1 38 þ 2.1
Class 46 þ 3.1 48 þ 1.2
Order 84 þ 2.5 88 þ 3.0
Family 134 þ 9.0 141 þ 5.0
OTUa 1087 þ 97.0 1221 þ 123.0
a OTUs were defined with 0e3% sequence divergence.
Please cite this article in press as: Godoy-Vitorino, F., et al., Differences in crop
locations, Research in Microbiology (2012), doi:10.1016/j.resmic.2012.01.001
as 40% of sample variance (Fig. 4B), which was higher( p ¼ 0.0011) among crop bacterial communities from animalsat Cojedes (Fig. 4A, C). In the principal component analysis(Fig. 4B), dispersion (inter-individual community distances)was higher in crop communities from animals from Cojedes,where birds ate abundantly from 5 plant species (Coccolobasp., L. ollaria, Machaerium sp., Caesaria sp., and Guazumatormentosa, (Domınguez-Bello et al., 1994)), compared toOrinoco individuals, which ate mostly from 3 plant species,C. ovata, C. multiflora and N. pichurim. Despite the smallnumber of individuals we confirmed that intrapopulationUniFrac distances (especially in animals from the OrinocoRiver) were smaller than between population distances, con-firming geographical differentiation of the crop microbiota inanimals from these two different bird populations (Table S4).In fact, according to the analyses of similarity test (ANOSIM),the dissimilarities between locations (crop bacteria fromanimals in Cojedes and Orinoco) were greater (R ¼ 0.63) thanthe differences between the individuals within each locationalthough with borderline significance ( p ¼ 0.101; Suppl.Fig. 3). This analysis also reflected greater dissimilarity inthe crop of individuals in Cojedes than in Orinoco, consistentwith the dispersion plots of Fig. 4C (Suppl. Fig. 3).
bacterial community structure between hoatzins from different geographical
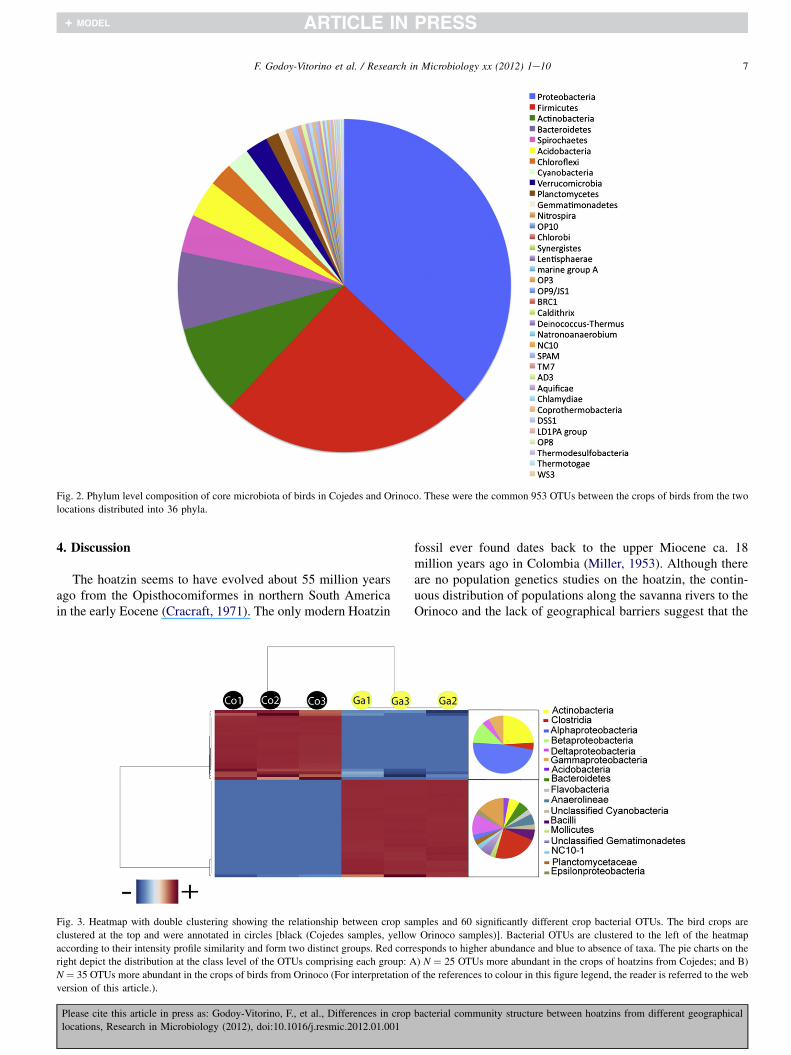
Fig. 2. Phylum level composition of core microbiota of birds in Cojedes and Orinoco. These were the common 953 OTUs between the crops of birds from the two
locations distributed into 36 phyla.
7F. Godoy-Vitorino et al. / Research in Microbiology xx (2012) 1e10
4. Discussion
The hoatzin seems to have evolved about 55 million yearsago from the Opisthocomiformes in northern South Americain the early Eocene (Cracraft, 1971). The only modern Hoatzin
Fig. 3. Heatmap with double clustering showing the relationship between crop sa
clustered at the top and were annotated in circles [black (Cojedes samples, yellow
according to their intensity profile similarity and form two distinct groups. Red corr
right depict the distribution at the class level of the OTUs comprising each group: A
N ¼ 35 OTUs more abundant in the crops of birds from Orinoco (For interpretation
version of this article.).
Please cite this article in press as: Godoy-Vitorino, F., et al., Differences in crop
locations, Research in Microbiology (2012), doi:10.1016/j.resmic.2012.01.001
fossil ever found dates back to the upper Miocene ca. 18million years ago in Colombia (Miller, 1953). Although thereare no population genetics studies on the hoatzin, the contin-uous distribution of populations along the savanna rivers to theOrinoco and the lack of geographical barriers suggest that the
mples and 60 significantly different crop bacterial OTUs. The bird crops are
Orinoco samples)]. Bacterial OTUs are clustered to the left of the heatmap
esponds to higher abundance and blue to absence of taxa. The pie charts on the
) N ¼ 25 OTUs more abundant in the crops of hoatzins from Cojedes; and B)
of the references to colour in this figure legend, the reader is referred to the web
bacterial community structure between hoatzins from different geographical
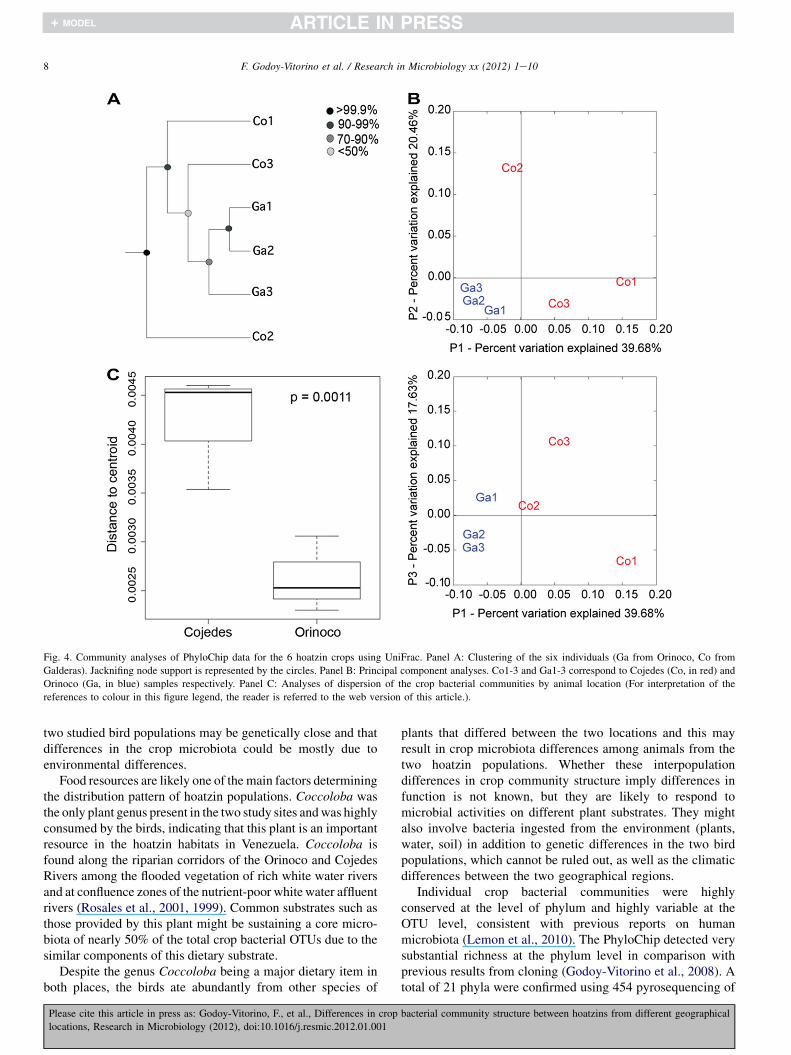
Fig. 4. Community analyses of PhyloChip data for the 6 hoatzin crops using UniFrac. Panel A: Clustering of the six individuals (Ga from Orinoco, Co from
Galderas). Jacknifing node support is represented by the circles. Panel B: Principal component analyses. Co1-3 and Ga1-3 correspond to Cojedes (Co, in red) and
Orinoco (Ga, in blue) samples respectively. Panel C: Analyses of dispersion of the crop bacterial communities by animal location (For interpretation of the
references to colour in this figure legend, the reader is referred to the web version of this article.).
8 F. Godoy-Vitorino et al. / Research in Microbiology xx (2012) 1e10
two studied bird populations may be genetically close and thatdifferences in the crop microbiota could be mostly due toenvironmental differences.
Food resources are likely one of the main factors determiningthe distribution pattern of hoatzin populations. Coccoloba wasthe only plant genus present in the two study sites andwas highlyconsumed by the birds, indicating that this plant is an importantresource in the hoatzin habitats in Venezuela. Coccoloba isfound along the riparian corridors of the Orinoco and CojedesRivers among the flooded vegetation of rich white water riversand at confluence zones of the nutrient-poor white water affluentrivers (Rosales et al., 2001, 1999). Common substrates such asthose provided by this plant might be sustaining a core micro-biota of nearly 50% of the total crop bacterial OTUs due to thesimilar components of this dietary substrate.
Despite the genus Coccoloba being a major dietary item inboth places, the birds ate abundantly from other species of
Please cite this article in press as: Godoy-Vitorino, F., et al., Differences in crop
locations, Research in Microbiology (2012), doi:10.1016/j.resmic.2012.01.001
plants that differed between the two locations and this mayresult in crop microbiota differences among animals from thetwo hoatzin populations. Whether these interpopulationdifferences in crop community structure imply differences infunction is not known, but they are likely to respond tomicrobial activities on different plant substrates. They mightalso involve bacteria ingested from the environment (plants,water, soil) in addition to genetic differences in the two birdpopulations, which cannot be ruled out, as well as the climaticdifferences between the two geographical regions.
Individual crop bacterial communities were highlyconserved at the level of phylum and highly variable at theOTU level, consistent with previous reports on humanmicrobiota (Lemon et al., 2010). The PhyloChip detected verysubstantial richness at the phylum level in comparison withprevious results from cloning (Godoy-Vitorino et al., 2008). Atotal of 21 phyla were confirmed using 454 pyrosequencing of
bacterial community structure between hoatzins from different geographical

9F. Godoy-Vitorino et al. / Research in Microbiology xx (2012) 1e10
the V8 region of the 16S RNA gene, which included a dozenrare phyla already detected by the PhyloChip. The detection ofthese poorly abundant phyla shows the sensitivity of thePhyloChip (Brodie et al., 2007). While dominant bacterialpopulations, particularly anaerobes, are expected to be part ofthe crop microbiome, we cannot determine the extent of die-tary bacteria among some of these less abundant taxa. Recentpreliminary data from metagenomics studies of the hoatzincrop support the high sensitivity of the PhyloChip, showing asmany as 36 bacterial phyla in the adult hoatzin crop (unpub-lished results). Consistent with the results of the current study,in recent studies by us, the PhyloChip also detected a highnumber of phyla in other digestive samples: 36 phyla in thecow rumen (Godoy-Vitorino et al., 2011) and 21 phyla in thececum of Capybara (Garcia-Amado et al., 2011). The Phy-loChip is therefore a sensitive and simple tool for use inhighlighting differences in the microbiota between differentgroups of hosts.
As already reported in previous work, the most abundantphyla in the crop of the hoatzin were the Proteobacteria, Fir-micutes, Actinobacteria and Bacteroidetes (Godoy-Vitorinoet al., 2010, 2008). These phyla also dominate other verte-brate guts including herbivores, omnivores and carnivores(Ley et al., 2008).
DNA microarray detected rare bacteria previouslyreported in the hoatzin crop (Godoy-Vitorino et al., 2010),including several putative thermophilic bacterial phyla suchas Aquificae (Reysenbach, 2001), Thermodesulfobacteria(Huber and Stetter, 2001), Coprothermobacteria (Rainey andStackebrandt, 2008) and Caldithrix (Wrighton et al., 2008).We cannot dismiss the possibility that some bacteria (espe-cially aerobic), detected by the PhyloChip may be associatedwith ingested soil, water or plants (Jacques et al., 1995), aswell as with animal differences resulting from different envi-ronmental factors other than diet.
The hoatzin diet is composed of plants that containcomplex toxic polyphenolic compounds (D’Mello, 1989;Domınguez-Bello et al., 1994; Grajal et al., 1989), with anti-microbial properties. The fruits and flowers of plants of thegenus Coccoloba are known to contain high concentrations ofphenols (Schall and Ressel, 1991), which might be detoxifiedby crop bacteria, since we have shown that hoatzin cropbacteria can detoxify plant tannins (Garcia-Amado et al.,2007; Jones et al., 2000). Certain Clostridia (Clostridium)and Actinobacteria (Actinomycetes), both abundant in thecrop, comprise known cellulose degraders in the rumen ofcows (Hungate, 1975).
We detected other fiber-degrading microorganisms in low-abundance, including taxa within Spirochetes, a lignindegrader in the termite hindgut (Warnecke et al., 2007) andPrevotellaceae, a common rumen hemicellulolytic group(Fondevila and Dehority, 1994). Prevotellaceae were found inhigher relative abundance in birds from the Orinoco than inthose from the Cojedes River. The current data does not enablepredicting functions beyond those relevant to known bacterialtaxa. Ongoing studies, including metagenomics, will help inunderstanding crop functions, developing hypotheses for
Please cite this article in press as: Godoy-Vitorino, F., et al., Differences in crop
locations, Research in Microbiology (2012), doi:10.1016/j.resmic.2012.01.001
interactions between community members and reconstructingcomplete or near-complete genomes of dominant organisms.
This is the first study comparing the microbiota of adulthoatzins from different geographical sites and is a first steptoward understanding the underlying principles that governcommunity assembly. Further studies using wider “-omics”(metagenomics, transcriptomics, proteomics) approaches needto identify how plant dietary resources shape communityfunctions. Our study should trigger wider-scale studies char-acterizing the microbial communities of plant leaf material,water and soil that might be sources of transient bacteria in thedigestive tract.
Acknowledgements
The authors gratefully acknowledge computational supportfrom Dr. Ulas Karaoz and the fieldwork support of JoseGonzalez-Fernandez, Juan Gonzalez-Fernandez and AntonioGonzalez-Fernandez from Hato Mataclara (Cojedes River),Capitan Jesus Perez from UNEG and Ramon Marino and hisson “Monchito” from Puerto Ordaz. This work was supportedin part by grants from NSF IOS 0716911, NSF DEB-DDIG0709840 and NSF CREST/HRD0206200. Part of this workwas performed at Lawrence Berkeley National Laboratoryunder the auspices of the University of California, contractnumber DE-AC02-05CH11231.
Appendix. Supplementary data
Supplementary data related to this article can be foundonline at doi:10.1016/j.resmic.2012.01.001.
References
Apajalahti, J.H., Kettunen, H., Kettunen, A., Holben, W.E., Nurminen, P.H.,
Rautonen, N., Mutanen, M., 2002. Culture-independent microbial
community analysis reveals that inulin in the diet primarily affects
previously unknown bacteria in the mouse cecum. Appl. Environ. Micro-
biol. 68, 4986e4995.
Brodie, E.L., Desantis, T.Z., Joyner, D.C., Baek, S.M., Larsen, J.T.,
Andersen, G.L., Hazen, T.C., Richardson, P.M., Herman, D.J.,
Tokunaga, T.K., Wan, J.M., Firestone, M.K., 2006. Application of a high-
density oligonucleotide microarray approach to study bacterial population
dynamics during uranium reduction and reoxidation. Appl. Environ.
Microbiol. 72, 6288e6298.Brodie, E.L., DeSantis, T.Z., Parker, J.P., Zubietta, I.X., Piceno, Y.M.,
Andersen, G.L., 2007. Urban aerosols harbor diverse and dynamic bacte-
rial populations. Proc. Natl. Acad. Sci. USA 104, 299e304.
Cracraft, J., 1971. A new family of hoatzin-like birds (Order Opisthocomi-
formes) from the Eocene of South America. Ibis 113, 229e233.
D’Mello, J.P., 1989. Toxic Amino Acids. In: D’Mello, J.P., Duffus, C.M.,
Duffus, J.H. (Eds.), Toxic Substances in Crop Plants. Association of
Applied Biologists, Warwick, U.K., pp. 29e54.
DeSantis, T.Z., Hugenholtz, P., Larsen, N., Rojas, M., Brodie, E.L., Keller, K.,
Huber, T., Dalevi, D., Hu, P., Andersen, G.L., 2006. Greengenes,
a chimera-checked 16S rRNA gene database and workbench compatible
with ARB. Appl. Environ. Microbiol. 72, 5069e5072.DeSantis, T.Z., Brodie, E.L., Moberg, J.P., Zubieta, I.X., Piceno, Y.M.,
Andersen, G.L., 2007. High-density universal 16S rRNA microarray
analysis reveals broader diversity than typical clone library when sampling
the environment. Microb. Ecol. 53, 371e383.
bacterial community structure between hoatzins from different geographical

10 F. Godoy-Vitorino et al. / Research in Microbiology xx (2012) 1e10
Domınguez-Bello, M.G., Michelangeli, F., Ruiz, M.C., Garcia, A.,
Rodriguez, E., 1994. Ecology of the folivorous hoatzin (Opisthocomus
hoazin) on the Venezuelan plains. Auk 111, 643e651.
Eisen, M.B., Spellman, P.T., Brown, P.O., Botstein, D., 1998. Cluster analysis
and display of genome-wide expression patterns. Proc. Natl. Acad. Sci.
USA 95, 14863e14868.
Engelbrektson, A., Kunin, V., Wrighton, K.C., Zvenigorodsky, N., Chen, F.,
Ochman, H., Hugenholtz, P., 2010. Experimental factors affecting PCR-
based estimates of microbial species richness and evenness. ISME J. 4,
642e647.
Espinoza, F.M., Argenti, P.m., Rodrıguez de, L.I., Gil, J.L., Guenni, O.,
Botaccio, R., 1993. Rendimiento en materia seca de ecotipos nativos e
introducidos del genero Centrosema, en los estados Anzoategui y Cojedes.
Zootecnia Trop. 11, 171e193.
Fondevila, M., Dehority, B.A., 1994. Degradation and utilization of forage
hemicellulose by rumen bacteria, singly in coculture or added sequentially.
J. Appl. Bacteriol. 77, 541e548.
Garcıa, M.A., Gueneau, P., Michelangeli, F., Domınguez-Bello, M.G., 2003.
Rate of detoxification of Quillaja saponins by crop bacteria from Opis-
thocomus hoazin is increased in the presence of methanogenic bacteria.
Trop. Subtrop. Agroecosyst 3, 595e598.
Garcia-Amado, M.A., Michelangeli, F., Gueneau, P., Perez, M.E., Domınguez-
Bello, M.G., 2007. Bacterial detoxification of saponins in the crop of the
avian foregut fermenter Opisthocomus hoazin. J. Anim. Feed Sci. 16,
82e85.
Garcia-Amado, M.A., Godoy-Vitorino, F., Piceno, Y.M., Tom, L.M.,
Andersen, G.L., Herrera, E.A., Dominguez-Bello, M.G., 2011. Bacterial
diversity in the cecum of the World’s Largest living Rodent (Hydrochoerus
hydrochaeris). Microb. Ecol..
Godoy-Vitorino, F., Gao, Z., Pei, Z., Ley, R., Gordon, J., Hackstein, J.,
Pericchi, L., Garcia-Amado, M., Michelangeli, F., Blaser, M., Domınguez-
Bello, M., 2006. In: 11th International Symposium on Microbial Ecology:
The Hidden Powers e Microbial Communities in Action. Ecology.
I.I.S.f.M, Vienna, Austria, p. 121.
Godoy-Vitorino, F., Ley, R.E., Gao, Z., Pei, Z., Ortiz-Zuazaga, H.,
Pericchi, L.R., Garcia-Amado, M.A., Michelangeli, F., Blaser, M.J.,
Gordon, J.I., Dominguez-Bello, M.G., 2008. Bacterial community in the
crop of the hoatzin, a neotropical folivorous flying bird. Appl. Environ.
Microbiol. 74, 5905e5912.
Godoy-Vitorino, F., Goldfarb, K., Brodie, E., Garcia-Amado, M.A.,
Michelangeli, F., Dominguez-Bello, M.G., 2010. Developmental microbial
Ecology of the crop of the folivorous hoatzin. ISME J. 4, 611e620.Godoy-Vitorino, F., Goldfarb, K.C., Karaoz, U., Leal, S., Garcia-
Amado, M.A., Hugenholtz, P., Tringe, S.G., Brodie, E.L., Dominguez-
Bello, M.G., 2011. Comparative analyses of foregut and hindgut bacterial
communities in hoatzins and cows. ISME J..
Grajal, A., 1995. Structure and Function of the Digestive Tract of the Hoatzin
(Opisthocomus hoazin): a folivorous bird with foregut fermentation. Auk
112, 20e28.Grajal, A., Parra, O., 1995. Passage rates of digesta markers in the gut of the
Hoatzin, a folivorous bird with foregut fermentation. Condor 97, 675e683.
Grajal, A., Strahl, S.D., Parra, R., Dominguez, M.G., Neher, A., 1989. Foregut
fermentation in the hoatzin, a neotropical leaf-Eating bird. Science 127,
1131e1134.
Hayashi, H., Sakamoto, M., Benno, Y., 2002. Fecal microbial diversity in
a strict vegetarian as determined by molecular analysis and cultivation.
Microbiol. Immunol. 46, 819e831.
Family I. deferribacteraceae fam. nov. In: Huber, H., Stetter, K.O. (Eds.),
Bergey’s Manual of Systematic Bacteriology. (The Archaea and the
Deeply Branching and Phototrophic Bacteria). Springer, New York,
pp. 169e210.
Hungate, R.E., 1975. The rumen microbial ecosystem. Annu. Rev. Ecol. Syst.
6, 39e66.
Please cite this article in press as: Godoy-Vitorino, F., et al., Differences in crop
locations, Research in Microbiology (2012), doi:10.1016/j.resmic.2012.01.001
Ivanov II, , Atarashi, K., Manel, N., Brodie, E.L., Shima, T., Karaoz, U.,
Wei, D., Goldfarb, K.C., Santee, C.A., Lynch, S.V., Tanoue, T., Imaoka, A.,
Itoh, K., Takeda, K., Umesaki, Y., Honda, K., Littman, D.R., 2009.
Induction of intestinal Th17 cells by segmented filamentous bacteria. Cell
139, 485e498.Jacques, M., Kinkel, L.L., Morris, C.E., 1995. Population sizes, immigration,
and growth of epiphytic bacteria on leaves of different ages and positions
of field-grown endive (Cichorium endivia var. latifolia). Appl. Environ.
Microbiol. 61, 899e906.
Jones, R.J., Amado, M.A., Dominguez-Bello, M.G., 2000. Comparison of the
digestive ability of crop fluid from the folivorous Hoatzin (Opisthocomus
hoazin) and cow rumen fluid with seven tropical forages. Anim. Feed Sci.
Technol 87, 287e296.
Kunin, V., Hugenholtz, P., 2010. PyroTagger: a Fast, Accurate Pipeline for
Analysis of RRNA Amplicon Pyrosequence Data. Open J. 1, e1.
Lane, D.J., 1991. 16S/23S RRNA Sequencing. Wiley, London, pp. 115e175.Lemon, K.P., Klepac-Ceraj, V., Schiffer, H.K., Brodie, E.L., Lynch, S.V.,
Kolter, R., 2010. Comparative analyses of the bacterial microbiota of the
human nostril and oropharynx. MBio 1, e00129-e10.
Ley, R.E., Hamady, M., Lozupone, C., Turnbaugh, P.J., Ramey, R.R.,
Bircher, J.S., Schlegel, M.L., Tucker, T.A., Schrenzel, M.D., Knight, R.,
Gordon, J.I., 2008. Evolution of mammals and their gut microbes. Science
320, 1647e1651.Lozupone, C., Knight, R., 2005. UniFrac: a new phylogenetic method for
comparing microbial communities. Appl. Environ. Microbiol. 71,
8228e8235.
Miller, A., 1953. A fossil hoatzin from the Miocene of Colombia. Auk 70,
484e489.
Pruesse, E., Quast, C., Knittel, K., Fuchs, B.M., Ludwig, W., Peplies, J.,
Glockner, F.O., 2007. SILVA: a comprehensive online resource for quality
checked and aligned ribosomal RNA sequence data compatible with ARB.
Nucleic Acids Res. 35, 7188e7196.
Rainey, F.A., Stackebrandt, E., 2008. Phylogenetic analysis of the bacterial
genus thermobacteroides indicates an ancient origin of thermobacteroides
proteolyticus. Lett. Appl. Microbiol. 16, 282e286.
Reysenbach, A.-L., 2001. Aquificae phy. nov. In: Boone, D.R.,
Castenholz, R.W. (Eds.), Bergey’s Manual of Systematic Bacteriology.
Springer-Verlag, Berlin, pp. 359e367p.Rosales, J., Petts, G., Salo, J., 1999. Riparian flooded forests of the Orinoco
and Amazon basins: a comparative review. Biodivers. Conserv 8,
551e586.
Rosales, J., Petts, G., Knab-Vispo, C., 2001. Ecological gradients in riparian
forests of the lower Caura River, Venezuela. Plant Ecol. 152, 101e118.
Schall, J.J., Ressel, S., 1991. Toxic plant compounds and the diet of the
predominantly herbivorous Whiptail Lizard, Cnemidophorus arubensis.
Copeia 1991, 111e119.
Storey, J.D., Tibshirani, R., 2003. Statistical methods for identifying differ-
entially expressed genes in DNA microarrays. Methods Mol. Biol. 224,
149e157.Tajima, K., Aminov, R.I., Nagamine, T., Matsui, H., Nakamura, M., Benno, Y.,
2001. Diet-dependent shifts in the bacterial population of the rumen
revealed with real-time PCR. Appl. Environ. Microbiol. 67, 2766e2774.
Warnecke, F., Luginbuhl, P., Ivanova, N., Ghassemian, M., Richardson, T.H.,
Stege, J.T., Cayouette, M., McHardy, A.C., et al., 2007. Metagenomic and
functional analysis of hindgut microbiota of a wood-feeding higher
termite. Nature 450, 560e565.Wright, A.D., Northwood, K.S., Obispo, N.E., 2009. Rumen-like methanogens
identified from the crop of the folivorous South American bird, the hoatzin
(Opisthocomus hoazin). ISME J. 3, 1120e1126.
Wrighton, K.C., Agbo, P., Warnecke, F., Weber, K.A., Brodie, E.L.,
DeSantis, T.Z., Hugenholtz, P., Andersen, G.L., Coates, J.D., 2008. A
novel ecological role of the Firmicutes identified in thermophilic microbial
fuel cells. ISME J. 2, 1146e1156.
bacterial community structure between hoatzins from different geographical




















