
Diatom species abundance and morphologically-based dissolution proxies in coastal Southern Ocean...
8
Research papers Diatom species abundance and morphologically-based dissolution proxies in coastal Southern Ocean assemblages Jonathan P. Warnock a,n , Reed P. Scherer b a Institute for the Environment, Sustainability, and Energy, Northern Illinois University, DeKalb, IL 60115 USA b Department of Geology and Environmental Geosciences, Northern Illinois University, DeKalb, IL 60115, USA article info Article history: Received 11 September 2014 Received in revised form 25 March 2015 Accepted 10 April 2015 Available online 16 April 2015 Keywords: Diatom dissolution Taphomony Silica cycle Fragilariopsis curta abstract Taphonomic processes alter diatom assemblages in sediments, thus potentially negatively impacting paleoclimate records at various rates across space, time, and taxa. However, quantitative taphonomic data is rarely included in diatom-based paleoenvironmental reconstructions and no objective standard exists for comparing diatom dissolution in sediments recovered from marine depositional settings, in- cluding the Southern Ocean’s opal belt. Furthermore, identifying changes to diatom dissolution through time can provide insight into the efficiency of both upper water column nutrient recycling and the biological pump. This is significant in that reactive metal proxies (e.g. Al, Ti) in the sediments only ac- count for post-depositional dissolution, not the water column where the majority of dissolution occurs. In order to assess the range of variability of responses to dissolution in a typical Southern Ocean diatom community and provide a quantitative guideline for assessing taphonomic variability in diatoms recovered from core material, a sediment trap sample was subjected to controlled, serial dissolution. By evaluating dissolution-induced changes to diatom species’ relative abundance, three preservational ca- tegories of diatoms have been identified: gracile, intermediate, and robust. The relative abundances of these categories can be used to establish a preservation grade for diatom assemblages. However, changes to the relative abundances of diatom species in sediment samples may reflect taphonomic or ecological factors. In order to address this complication, relative abundance changes have been tied to dissolution-induced morphological change to the areolae of Fragilariopsis curta, a significant sea-ice indicator in Southern Ocean sediments. This correlation allows differentiation between gracile species loss to dissolution versus ecological factors or sediment winnowing. These results mirror a si- milar morphological dissolution index from a parallel study utilizing Fragilariopsis kerguelensis, sug- gesting that results are applicable to a broad spectrum of diatoms typically preserved in the sediments. & 2015 Elsevier Ltd. All rights reserved. 1. Introduction Silicon is a significant nutrient in the photic zone of most marine environments, especially the Southern Ocean, where dia- toms can comprise a majority of primary productivity, in that it is required for diatom growth and provides ballast to dead diatom cells (Nelson et al., 1995; Ragueneau et al., 2000). Biogenic silica flux to the seafloor and subsequent burial represent the largest sinks of bioavailable silica in the marine system (Tréguer et al., 1995). Diatoms’ frustules also contain interstitial carbon, and hence carry organic carbon out of the photic zone and sequester it in sediments, even after utilization of the cellular organic material by zooplankton and bacteria (e.g. Wassmann, 1998; Assmy et al., 2006). As such, diatom frustules serve as an important organic carbon sink (Sarthou et al., 2005; DeMaster, 2002). Diatom pre- servation, and hence Si and C cycling, is known to be variable across space and time (e.g. Lončarić et al., 2007; Warnock et al., 2007), as well as variable between taxa (e.g. Shemesh et al., 1989). Assessing variability in diatom dissolution is therefore significant to understanding changes to the Si and C cycles on glacial/inter- glacial timescales. The majority of diatom dissolution occurs in the upper water column (Tréguer et al., 1995), but reactive metal proxies, e.g. Al and Ti, can only account for a minor amount of post-depositional dissolution (Dymond et al., 1997). Given the impact of upper water column diatom dissolution on carbon flux as well as on paleoceanographic interpretation of fossil diatom assemblages, several strategies for estimating water column dis- solution and diatom preservation have been devised. However, each previously published approach has limited application. A common strategy in preservational studies is to identify successive changes to diatom morphology, placing diatoms into preservational ‘bins’ (e.g. Ryves et al., 2009; Warnock et al., 2007; Contents lists available at ScienceDirect journal homepage: www.elsevier.com/locate/csr Continental Shelf Research http://dx.doi.org/10.1016/j.csr.2015.04.012 0278-4343/& 2015 Elsevier Ltd. All rights reserved. n Corresponding author. E-mail address: [email protected] (J.P. Warnock). Continental Shelf Research 102 (2015) 1–8
Transcript of Diatom species abundance and morphologically-based dissolution proxies in coastal Southern Ocean...

Continental Shelf Research 102 (2015) 1–8
Contents lists available at ScienceDirect
Continental Shelf Research
http://d0278-43
n CorrE-m
journal homepage: www.elsevier.com/locate/csr
Research papers
Diatom species abundance and morphologically-based dissolutionproxies in coastal Southern Ocean assemblages
Jonathan P. Warnock a,n, Reed P. Scherer b
a Institute for the Environment, Sustainability, and Energy, Northern Illinois University, DeKalb, IL 60115 USAb Department of Geology and Environmental Geosciences, Northern Illinois University, DeKalb, IL 60115, USA
a r t i c l e i n f o
Article history:Received 11 September 2014Received in revised form25 March 2015Accepted 10 April 2015Available online 16 April 2015
Keywords:Diatom dissolutionTaphomonySilica cycleFragilariopsis curta
x.doi.org/10.1016/j.csr.2015.04.01243/& 2015 Elsevier Ltd. All rights reserved.
esponding author.ail address: [email protected] (J.P
a b s t r a c t
Taphonomic processes alter diatom assemblages in sediments, thus potentially negatively impactingpaleoclimate records at various rates across space, time, and taxa. However, quantitative taphonomicdata is rarely included in diatom-based paleoenvironmental reconstructions and no objective standardexists for comparing diatom dissolution in sediments recovered from marine depositional settings, in-cluding the Southern Ocean’s opal belt. Furthermore, identifying changes to diatom dissolution throughtime can provide insight into the efficiency of both upper water column nutrient recycling and thebiological pump. This is significant in that reactive metal proxies (e.g. Al, Ti) in the sediments only ac-count for post-depositional dissolution, not the water column where the majority of dissolution occurs.
In order to assess the range of variability of responses to dissolution in a typical Southern Oceandiatom community and provide a quantitative guideline for assessing taphonomic variability in diatomsrecovered from core material, a sediment trap sample was subjected to controlled, serial dissolution. Byevaluating dissolution-induced changes to diatom species’ relative abundance, three preservational ca-tegories of diatoms have been identified: gracile, intermediate, and robust. The relative abundances ofthese categories can be used to establish a preservation grade for diatom assemblages.
However, changes to the relative abundances of diatom species in sediment samples may reflecttaphonomic or ecological factors. In order to address this complication, relative abundance changes havebeen tied to dissolution-induced morphological change to the areolae of Fragilariopsis curta, a significantsea-ice indicator in Southern Ocean sediments. This correlation allows differentiation between gracilespecies loss to dissolution versus ecological factors or sediment winnowing. These results mirror a si-milar morphological dissolution index from a parallel study utilizing Fragilariopsis kerguelensis, sug-gesting that results are applicable to a broad spectrum of diatoms typically preserved in the sediments.
& 2015 Elsevier Ltd. All rights reserved.
1. Introduction
Silicon is a significant nutrient in the photic zone of mostmarine environments, especially the Southern Ocean, where dia-toms can comprise a majority of primary productivity, in that it isrequired for diatom growth and provides ballast to dead diatomcells (Nelson et al., 1995; Ragueneau et al., 2000). Biogenic silicaflux to the seafloor and subsequent burial represent the largestsinks of bioavailable silica in the marine system (Tréguer et al.,1995). Diatoms’ frustules also contain interstitial carbon, andhence carry organic carbon out of the photic zone and sequester itin sediments, even after utilization of the cellular organic materialby zooplankton and bacteria (e.g. Wassmann, 1998; Assmy et al.,2006). As such, diatom frustules serve as an important organic
. Warnock).
carbon sink (Sarthou et al., 2005; DeMaster, 2002). Diatom pre-servation, and hence Si and C cycling, is known to be variableacross space and time (e.g. Lončarić et al., 2007; Warnock et al.,2007), as well as variable between taxa (e.g. Shemesh et al., 1989).Assessing variability in diatom dissolution is therefore significantto understanding changes to the Si and C cycles on glacial/inter-glacial timescales. The majority of diatom dissolution occurs in theupper water column (Tréguer et al., 1995), but reactive metalproxies, e.g. Al and Ti, can only account for a minor amount ofpost-depositional dissolution (Dymond et al., 1997). Given theimpact of upper water column diatom dissolution on carbon fluxas well as on paleoceanographic interpretation of fossil diatomassemblages, several strategies for estimating water column dis-solution and diatom preservation have been devised. However,each previously published approach has limited application.
A common strategy in preservational studies is to identifysuccessive changes to diatom morphology, placing diatoms intopreservational ‘bins’ (e.g. Ryves et al., 2009; Warnock et al., 2007;

J.P. Warnock, R.P. Scherer / Continental Shelf Research 102 (2015) 1–82
Shimada et al., 2003), whereas biostratigraphic studies generallyassign arbitrary and rarely defined designations (e.g., “good” or“poor”). By utilizing the ratios of binned preservational states,semi-quantitative data may be produced, which marks a sig-nificant improvement over arbitrary assignations.
The process of dissolution is complex and progressive, so uti-lizing preservational bins has the potential to miss importantthreshold events that may be relevant to the flux and preservationprocesses. These can be captured by more careful measurement ofdissolution-induced changes to valve morphology rather thansimple morphological binning. No side-by-side comparisons be-tween measured and binned data have been undertaken to assessthe validity of using preservational bins to assess diatom dissolu-tion, however studies comparing preservational bin data to dis-solved silica content and absolute abundance show a strong cor-relation (Ryves et al., 2001). Additionally, very few previous stu-dies of the variability of diatom preservation include comparisonto fresh, undissolved diatom material (e.g. Ryves et al., 2009;Warnock et al., 2007; Lončarić et al., 2007; Ryves et al., 2006;Seeberg-Elverfeldt et al., 2004; Shimada et al., 2003; Flower, 1993).Because diatom dissolution primarily takes place in the upperwater column (Tréguer et al., 1995), studies that utilize diatommaterial from core tops and sediment traps miss the critical initialstages of dissolution.
A further set of studies (e.g. Lončarić et al., 2007; Seeberg-El-verfeldt et al., 2004; Ryves et al., 2003) compares plankton tows tocore-top samples or sediment trap material. These studies identifytaphonomic changes to assemblages and provide data on themodern functioning of the biological pump by focusing on bothrelative species abundance and absolute diatom abundancechanges with depth in the water column. However, these surfacewater/deep water comparison studies cannot address whetherchanges in diatom assemblages and abundances observed in se-diment cores reflect changes in productivity, preservation, or both.Indeed, although surface water/deep water comparison studies doprovide useful tools for interpreting the fossil record and remindresearchers to consider taphonomy, they do not provide anygrounds for determining the extent to which dissolution has al-tered a fossil assemblage. A final style of experiment involves se-rial dissolution of plankton tow or core-top material (Ryves et al.,2003; Pichon et al., 1992; Tréguer et al., 1995; Shemesh et al.,1989). A transfer function is created relating progressive dissolu-tion to species loss. This is, again, a useful step, but when appliedto species abundance data generated from sediment cores, it doesnot allow for the differentiation of taphonomic and ecologicalsignals down core. Any useful record of diatom dissolution mustbe able to distinguish between ecological and taphonomic effectson diatom assemblages.
One limnological study utilized fresh diatom samples to eval-uate the effects of diatom dissolution on species loss, morpholo-gical change, absolute abundance change and sediment biogenicsilica content (Ryves et al., 2001). Ryves et al. (2001; 2006) foundthat morphological change, even when binned into only 4 or asfew as 2 dissolution categories was a robust measure of diatomdissolution. Ryves et al. (2001) further found that a morphologicalapproach is a more effective means of assessing diatom dissolutionthan a transfer function based on categorizing species into ro-bustness groups. This work is valuable in assessing diatom dis-solution and generated dissolution proxies for the environmentstudied, however its conclusions are not directly applicable tomarine sediments. No morphologically-based diatom dissolutionindex yet exists for diatoms recovered from marine sediments,including Southern Ocean sediments. This is a major deficit, asdiatom-based paleoclimate and paleoecological studies based onSouthern Ocean sediment cores are common, and the SouthernOcean is a significant driver of both global climate and nutrient
cycling. Furthermore, the presence of electrolytes is known to af-fect the dissolution kinetics of amorphous silica, highlighting theneed for a separate marine proxy (Dove et al., 2008; Icenhowerand Dove, 2000).
To specifically address the lack of a diatom preservation indexfor coastal Southern Ocean diatom communities and test the va-lidity of morphologically-binned data in assessing diatom pre-servation, we present an analysis of serial dissolution of a diatomcommunity, simultaneously assessing species loss and dissolution-induced morphological change. A diatom community from theRoss Sea was dissolved in the laboratory, and sampled at regularintervals to assess the character and sequence of species lossthrough progressive dissolution. This procedure mimics that ofRyves et al. (2001) and Shemesh et al. (1989), who subjecteddiatom assemblages to serial dissolution and tracked specieschange over time. However, as noted above, ecological change canbe as significant as preservational change in establishing a diatomthanatocoenosis. In order to separate ecological and taphonomicinfluences on diatom assemblages preserved in sediment cores,dissolution-induced morphological changes to the robust sea-ice-associated diatom Fragilariopsis curta were assessed at each stageof dissolution, paralleling the morphological approach of Warnocket al. (2007) and Ryves et al. (2001). Fully quantified and binnedmorphological data are considered, to test the utility of the in-expensive and less time-consuming binning approach. The mor-phological results found here compare favorably with those ofsimilar analyses of Fragilariopsis kerguelensis, the most commonopen ocean diatom species in the Southern Ocean and SouthernOcean sediments, indicating that the approach is valid across aspectrum of diatom morphologies and ecologies (Warnock, 2013).
2. Methods
2.1. Dissolution
The diatom community used in this study was taken from the“Adélie” sediment trap described in Dunbar et al. (2003). Adéliewas deployed in the Ross Sea (76° 42.4’S, 169° 1.34’E), at 772 mdepth. The sample utilized for this study was collected on 24December 1996 and preserved in formalin. The extent to whichstorage of diatom valves in formalin will alter dissolution is un-known, however previous taphonomic studies have utilized pre-served diatom material (gluteraldehyde – Ryves et al., 2001). Thepreservational state of the sample was assessed prior to any al-teration in the lab via light microscopy. Despite the water depth,the majority of the cellular membranes were still intact, indicatingthe diatoms present in the sample were not significantly dissolvedbefore reaching the trap (Fig. 1). Additionally, gracile diatom spe-cies not typically preserved in the sediments (i.e. Chaetoceros de-bilis and Psuedo-nitzschia turgiduloides) were present in the sam-ple. It is known that some diatom species that grow in the Ross Seaare rapidly recycled in the upper water column and never reachthe sea floor (Leventer and Dunbar, 1996). The sediment trapsample studied is believed to accurately reflect the portion of thebiocoenosis of the Ross Sea in December capable of inclusion inthe sediments.
Formalin was removed from the sample via repeated washingthrough a 0.8 μm Whatman filter. The washed diatom communitywas then placed in a clean plastic flask with 30% H2O2 to strip theorganics and expose the silica walls. In nature, this step is fa-cilitated by bacteria, which strip the organic matter and exposesilica to dissolution (e.g. Bidle and Azam, 1999). H2O2 is known toattack diatom silica (Morales et al., 2013), however this step isnecessary to complete the dissolution process within a reasonabletime frame. Furthermore, the preservational state of the diatoms

Fig. 1. Intact diatom cells from sediment trap ‘Adelie’ deployed in the Ross Sea. Thecytoplasm and organelles of this Chaetoceros are still intact, indicating that thesample was not highly altered before experimental treatment.
Fig. 2. Morphological guide. This figure shows an individual valve of Fragilariopsiscurta at two magnifications. Morphology significant to this study is labeled asfollows: A – Areola, B – Costa, C – Transapical rib, D – Margin, E – Girdle band, F –
Valve face.
J.P. Warnock, R.P. Scherer / Continental Shelf Research 102 (2015) 1–8 3
was assessed prior to the dissolution experiment to verify thatperoxide did not damage the frustules. Following the peroxidetreatment, the stripped diatom community was rewashed viamultiple rinsing, and placed in a clean, plastic flask containingstandard f/2 artificial sea water in order to emulate natural salt-mediated dissolution. The diatom suspension was thoroughlymixed and immediately subsampled into a 10 ml plastic centrifugetube. Subsequently, the plastic vessel containing the suspension ofcleaned diatom frustules was placed in an 80 °C water bath. Thediatom suspension was subsampled every hour for thirteen hoursinto individual, clean 10 ml plastic centrifuge tubes and allowed tocool. After cooling, each sample was filtered. Both the altereddiatom community and the dissolution medium were retained.The diatoms were subsequently cleaned of salts via repeated rin-sing through 0.8 μm Whatman filters prior to imaging.
2.2. Microscopy
The partially dissolved diatom community from each hourlysubsample was resuspended in deionized water. Light microscopeslides from each hourly subsample were made by pipetting ap-proximately 0.5 ml of the cleaned, resuspended diatoms ontocoverslips. The coverslips were allowed to dry, and then perma-nently mounted to glass slides with Norland Optical Adhesive.Relative abundances of each diatom species at each step along thedissolution process was assessed using a Leica DMLB microscopeat 1000� under oil immersion. At least 300 diatoms were countedfrom each slide. A second 0.5 ml subsample of each cleaned, re-suspended diatom sample was also pipetted onto 11 mm roundglass coverslips and allowed to dry. These coverslips were affixedto 9 mm round aluminum slugs with semiconductive carbon paintfor scanning electron microscope (SEM) analysis. A Polaron SEMCoating System sputter coater loaded with a 50% gold:50% palla-dium alloy was used to coat each sample before viewing.
The center of the valve face of 20 valves of F. curta was imagedat 10,000� magnification in the first nine samples with aJEOL5610LV SEM operating in high vacuum mode. Later samples,in which only valve margins and girdle bands of F. curta remained,were not imaged. The perimeter of all of the visible areolae withinthe 10,000� magnification image, or the perimeter of the gapresulting from the conjoining of neighboring areolae, was mea-sured as a proxy for dissolution. Fig. 2 presents a guide diagram-ming the relevant morphology. These measurements were madeusing the ‘Freehand Selections’ tool in ImageJ (a free software
package produced by the National Institutes of Health). This toolallows the user to draw a line of any shape, and, after calibrationwith the scale bar in the image, provides a length of that line.These perimeter data were averaged for each valve.
Mean perimeter data were compared to the amount of dis-solution time (Fig. 3A) and the amount of silica in the dissolutionmedium, measured via photospectrometry with a Milton RoySpectronic 401 photospectrometer (Fig. 3F). Representative valvesshowing progressive dissolution are provided on Fig. 3B–E. Laterdissolution stages, where only valve margins and girdle bands of F.curta remained were not considered in the silica analysis as dia-toms dissolved to this point would not be included in a typicalpaleoceanographic study. Paring diatom relative abundance datato morphological data allows for the direct comparison of dis-solution-induced change to the sedimentary diatom assemblage tobe linked to a physical signal of dissolution in F. curta, a speciesused as a proxy for sea-ice and sea surface temperature (Armandet al., 2005; Crosta, 2009).
2.3. Ground-truthing of dissolution
Finally, a set of Holocene sediment samples was prepared for SEMin order to validate the progression of dissolution-induced morpho-logical changes seen in the laboratory experiments and to test thedifference between measured areolae and binned data. Sedimentwas taken from core NBP 0101-JPC24 (68°41.637’S, 76°42.712’E), re-covered from Prydz Bay (Leventer et al., 2006). A mass of 0.05 g ofsediment was sampled every 20 cm to a depth of 1580 cmbsf fromNBP 0101-JPC24. Each sediment sample was treated with H2O2 forone hour to dissolve organic matter and HCl for one hour to dissolvecarbonates. The resultant sediment slurry was plated onto 11 mmround glass coverslips and prepared for SEM as the above sedimenttrap samples. 20 flat lying, unobstructed valves of F. curta were im-aged in each sample. The perimeter of each areola, or fused areolaegap, was measured as above, and each valve was assigned to one ofthree preservational bins: (1) Expanded and irregularly shaped areolawith no more than two sets of conjoined, neighboring areolae – highquality; (2) multiple sets of neighboring conjoined areolae up to oneentirely conjoined costa – intermediate quality; and (3) Costate silicabetween trans-apical ribs dissolved in multiple costae – low quality.

Fig. 3. Measured change to F. curta morphology over time in laboratory experiments. Dissolution, as expressed in terms of the mean perimeter of the gap in the valve face, isplotted as a function of time (A) and as a function of the amount of silica lost to solution from the diatom community (F). The bars above and below each data point representthe mean plus and minus one standard deviation. Compared to an unaltered valve (B), initial dissolution widens the areolae (C). Areolae eventually conjoin, starting at themargin (D), which causes a rapid increase in the rate of dissolution. Continued conjoining (E) causes all of the areolae in each costa to combine.
J.P. Warnock, R.P. Scherer / Continental Shelf Research 102 (2015) 1–84
3. Results
Controlled dissolution, as recorded in hourly aliquot examina-tion, was complete by hour 13, with no remaining identifiablediatoms. Some particulate diatomaceous silica remained in sus-pension, but was unidentifiable even to the genus level. Dissolu-tion-induced morphological changes to F. curta from the laboratoryclosely matched those naturally preserved in Holocene-aged sedi-ments from core NBP0101-JPC24 (Fig. 4). The striking similaritybetween laboratory-induced morphological change and the natu-rally produced morphological change preserved in NBP0101-JPC24validates the laboratory technique, despite preservation in formalin,the accelerated removal of organic material and 80 °C dissolution
medium. Furthermore, research on the dissolution kinetics ofamorphous silica has shown that although temperature does affectthe kinetics of dissolution, it does not affect the process of dis-solution (Icenhower and Dove, 2000). The solution did not reachsaturation and reprecipitation of silica was not seen, as has hap-pened in other studies (Mikkelsen, 1977). Additionally, the surfaceof the diatom silica was smooth prior to artificial dissolution, sug-gesting that the formalin did not meaningfully alter the opal.
The pre-dissolution diatom assemblage was dominated bysea-ice related species (Leventer, 1992; Armand et al., 2005) Fra-giliariopsis cylindrus (47.6%) and F. curta (39.4%) (Table 1). Pro-gressive dissolution revealed a change in the relative abundancesof diatom species (Fig. 5). A species was considered to be at 0%

Fig. 4. Validation of laboratory technique. The dissolution-induced morphological changes to Fragilariopsis curta from laboratory experiments (top) closely match those seenin naturally preserved material from core NBP0101-JPC24 (bottom). The areolae of unaltered valves are small and round (A and E). Progressive dissolution widens areolae,which become more irregularly shaped and being to conjoin, starting at the margin (B and F). Further dissolution progressively conjoins more neighboring areolae (C and G),until the costate silica has been dissolved and only trans-apical ribs remain of the valve face (D and H).
Table 1Species and abundances through dissolution. This table presents the relative abundances (%) of species as dissolution progressed.
Time (h) 0 1 2 3 4 5 6 7 8 9 10 11
Actinocyclus actinochilus 0.03 0.03 0.03 0.31 0.71 0.29 0.01 0.44 0.02 0.01 6.67 11.11Asteromphalus spp. 0.03 0.30 0.34 0.31 0.03 0.29 0.01 0.02 0.02 0.01 6.67 11.11Chaetoceros bulbosum 0.03 0.03 0.34 0.03 0.35 0.03 0.01 0.02 0.45 0.00 0.00 0.00Chaetoceros debilis 4.49 6.84 10.39 4.65 5.31 2.90 10.97 0.00 0.00 0.00 0.00 0.00Chaetoceros dichaeta 0.03 0.89 0.03 0.62 0.03 0.87 2.19 0.00 0.00 0.00 0.00 0.00Corethron spp. 0.03 0.89 0.03 0.03 0.03 0.03 0.01 0.00 0.00 0.00 0.00 0.00Eucampia antarctica 0.03 0.03 0.03 0.03 0.03 0.03 4.39 0.44 0.02 0.67 6.67 0.00Fragilariopsis curta 39.39 51.45 45.08 62.67 59.80 65.04 28.53 67.97 70.82 66.93 6.67 11.11Fragilariopsis cylindrus 47.62 23.50 33.51 20.94 18.05 9.99 0.01 1.33 0.00 0.00 0.00 0.00Fragilariopsis kerguelensis 0.03 0.15 0.17 0.62 0.03 0.87 0.01 0.67 0.02 0.01 6.67 11.11Fragilariopsis obliquecostat 1.35 2.38 0.50 0.78 1.95 1.01 2.74 2.89 3.36 5.05 6.67 0.00Fragilariopsis psuedonana 0.30 0.03 0.34 0.03 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00Fragilariopsis ritzscheri 1.05 4.46 3.35 3.72 3.89 9.27 39.50 14.22 12.10 14.80 6.67 11.11Fragilariopsis separanda 1.20 2.38 1.17 0.93 4.25 2.75 3.29 3.55 4.93 4.37 0.00 0.00Fragilariopsis sublinearis 0.45 1.19 0.84 1.09 1.77 2.46 2.19 3.78 3.14 2.02 0.00 0.00Paralia sulcata 1.20 0.03 0.03 0.03 0.03 0.03 0.01 0.02 0.02 0.01 6.67 11.11Proboscia spp. 0.03 0.03 0.34 0.03 0.03 0.29 2.19 0.02 0.02 0.01 6.67 0.00Psuedonitzchia spp. 1.50 1.04 0.84 0.93 1.06 0.43 0.55 0.02 0.02 0.00 0.00 0.00Stellarima microtrias 0.03 0.03 0.03 0.03 0.03 0.29 0.01 0.44 0.00 0.00 0.00 0.00Thalassiosira antarctica 0.90 1.19 0.67 1.24 1.06 1.16 0.01 1.33 2.24 3.36 6.67 0.00Thalassiosira gracilis 0.03 1.49 0.67 0.62 0.71 1.45 3.29 2.67 2.24 0.67 6.67 0.00Thalassiosira lentiginosa 0.03 0.30 0.67 0.03 0.71 0.29 0.01 0.02 0.45 1.35 6.67 0.00Thalassiosira oliverana 0.03 0.03 0.03 0.03 0.03 0.03 0.01 0.02 0.00 0.00 0.00 0.00Thalassiosira tumida 0.03 0.59 0.03 0.03 0.03 0.03 0.01 0.02 0.02 0.67 6.67 11.11Thalassiothrix spp 0.03 0.03 0.17 0.16 0.03 0.03 0.01 0.02 0.02 0.01 6.67 11.11
J.P. Warnock, R.P. Scherer / Continental Shelf Research 102 (2015) 1–8 5
relative abundance when only valve margins and girdle bandsremained, i.e., when the valve face silica was lost. This point waschosen because isolated girdle bands and margins are generallynot readily identifiable and therefore not included in diatomcensus counts.
Serial dissolution of a typical sea-ice related diatom communityrevealed three dominant trends in relative abundance. Thesetrends were used to define diatom robustness groups based on amodification of definitions from a similar experiment conductedby Shemesh et al. (1989). First, “robust” diatom species (e.g. F.curta and F. obliquecostata) are defined as those whose relativeabundances increased throughout dissolution until finally rapidlydropping to zero when all other diatom species had dissolved.“Intermediate” diatom species (e.g. Fragilariopsis sublinearis andThalassiosira gracilis) are those whose relative abundances increaseslightly then gradually decreases during dissolution. T. gracilis was
included in the intermediate category because the increase inabundance towards the end of the dissolution experiment wascaused by a single high-abundance sample at odds with the de-clining trend in abundance since hour 6. Third, “gracile” diatomspecies (e.g. F. cylindrus and C. debilis) are defined as generallydecreasing in relative abundance throughout dissolution.
A set of diatoms whose relative abundances were below 1% forthe majority of the experiment are not classified, as their relativescarcity caused erratic changes to their relative abundances duringdissolution e.g. (F. kerguelensis and Actinocyclus actinochilus). Manyof these unclassified species are ecologically significant and fre-quently used as paleoceanographic proxies, especially F. kergue-lensis, highlighting the need to replicate these experiments withdiatom assemblages from other oceanographic settings (Warnock,2013). The assemblage used in this study initially containedroughly 50% gracile, 45% robust, and 5% intermediate and

0 3 6 9 12
Hour
0
2
4
6
8
10
12
0 15 30 45
0.0 0.5 1.0 1.5
0 15 30
Hour
0
2
4
6
8
10
120 1 2 3 4
0 1 2 3 4 5 6 7 0 15 30 45 60
Hour
0
2
4
6
8
10
120 1 2 3 4 5 6 7
0 1 2 3 4 5
01 2 34 5 6 7
Fig. 5. Diatom species groups and preservational bins. Diatoms are categorized as gracile when their relative abundances generally decline, intermediate when their relativeabundances increase then decline slowly, or robust when their relative abundances increase then quickly drop at the end of dissolution. The proportions of these three typesof diatoms can be used to assess preservation generally as high quality, intermediate quality, and low quality.
J.P. Warnock, R.P. Scherer / Continental Shelf Research 102 (2015) 1–86
unclassified diatoms. These values reflect the dominance of F. curtaand F. cylindrus in the initial assemblage.
In general, centric diatoms, other than those of the genusChaetoceros, were the last to completely dissolve, indicating theirrelative robustness over pennates. The loss of the majority ofgracile species occurs in the 6th hour, at or just before the abun-dance peak of intermediate species. As intermediate species de-cline, the relative abundance of robust species increases rapidly.Finally, the relative abundances of robust species decrease quicklywith increasing dissolution after their peak abundances areachieved.
The interplay between the gracile, intermediate, and robustdiatoms can be used to define a set of preservational grades. Theseestimates can be used to assess how dissolution has altered anassemblage. We modify the equation presented by Shemesh et al.(1989) as a guideline for tracking preservational change within adiatom assemblage:
P G G R/( ) (1)= +
where P is a value between 0 and 1, representing the preservationquality of a sample, G is the proportion of gracile species in anassemblage, and R is the proportion of robust species in an as-semblage. “High quality” preservation is represented by a largeproportion of gracile and intermediate diatoms, and correspondsto a P value Z0.7. “Intermediate quality” preservation is re-presented by a small proportion of heavily dissolved gracile dia-toms and high proportions of intermediate and robust diatoms,with 0.7ZPZ0.4. “Low quality” preservation is represented byvery few or no gracile diatoms, few intermediate diatoms, andhigh proportions of robust diatoms, with a corresponding 0.44P.These numerical values follow those established by Shemesh et al.(1989).
Variability in the relative abundances of species observedwithin a core can be interpreted to reflect changes to the livingassemblage: an ecological change, changes to the transport anddeposition of diatoms in sediment: a depositional process change,or changes to dissolution: a taphonomic change. Therefore, a shiftfrom a gracile- to a robust-dominated diatom assemblage mightbe interpreted as resulting from multiple factors. Additionally,
assigning diatom species to preservational catagories based ontheir relative abundance changes during dissolution ties the dis-solution index to the species content of the original sediment trapsample. As not all environments will have the same growth as-semblage, this adds artificial controls to the dissolution proxy. Inorder to assess these complications, a morphological dissolutionindex based on expansion and conjoining of the areolae of therobust diatom F. curta has been established.
Alterations to the valve face of F. curta followed a predictablepathway, allowing dissolution-induced morphologies to be placedinto four bins. (1) Initial dissolution caused expansion of the are-olae and thinning of the valve face (Fig. 3B and C). (2) Continueddissolution caused conjoining of neighboring areolae beginningwith those nearest the margin, resulting in a rapid increase in theaverage gap in the valve face (Fig. 3D). (3) Dissolution progresseduntil all of the areolae had conjoined, leaving only the trans-apicalribs, margins, and girdle bands (Fig. 3E). (4) At 8 h of dissolution,no F. curta valve face silica remained, leaving only margins andgirdle bands. These results emulate those of Crawford et al. (2009),who showed that differences in silica density and chemistry be-tween valve face and girdle band silica result in differing rates ofdissolution. Furthermore, serial dissolution of cultured cells of F.kerguelensis revealed the same morphological changes seen herein F. curta (Warnock, 2013).
In order to reduce the time and money spent in assessingdiatom dissolution, valves of F. curta were placed into morpholo-gical bins corresponding to the stages described above (Fig. 6).High quality valves had no more than two sets of conjoined are-olae (stages 1 and 2; Fig. 6A). Intermediate quality valves hadmultiple conjoined areolae, but no more than 1 fully conjoinedcosta (stage 3; Fig. 6B). Low quality valves had multiple completelyconjoined costae (stage 4; Fig. 6C). A valve dissolved too far to beincluded in one of these categories is shown on Fig. 6D. Pre-servation from binned data was calculated following Eq. (2):
P H I H I L( 0.5 )/( ) (2)= + + +
where P is preservation, from 1 (good) to 0 (poor), H is the numberof high quality valves, I is the number of intermediate qualityvalves, and L is the number of low quality valves for any set sample
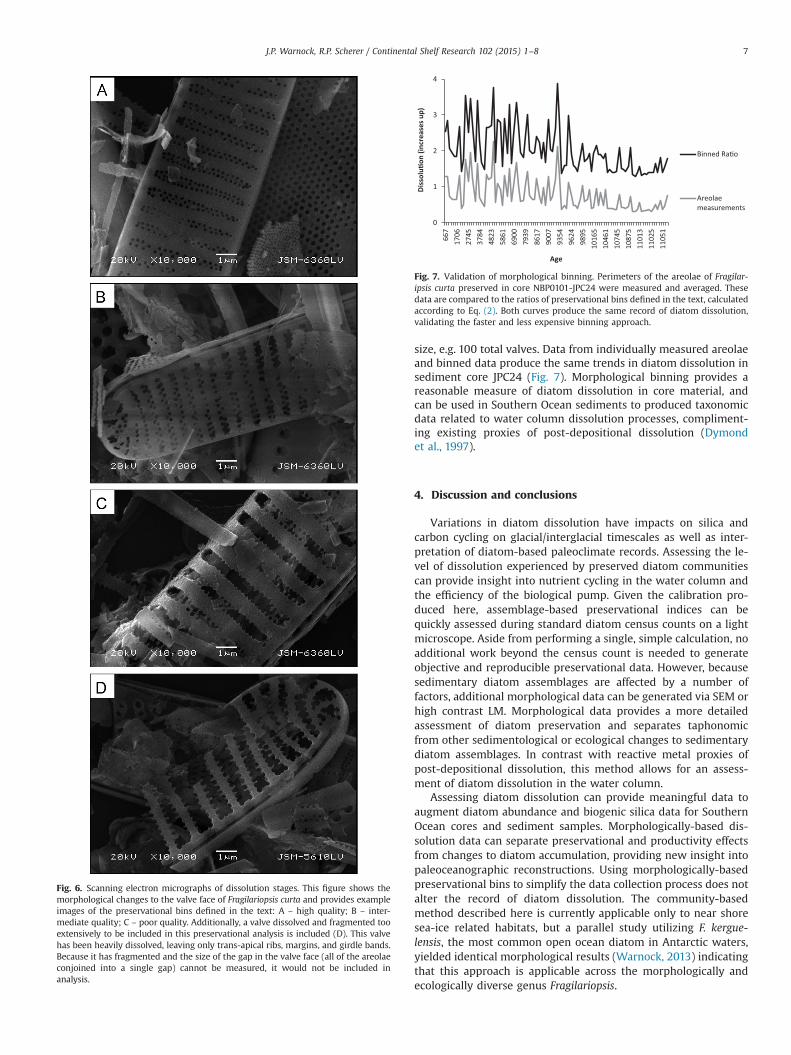
Fig. 6. Scanning electron micrographs of dissolution stages. This figure shows themorphological changes to the valve face of Fragilariopsis curta and provides exampleimages of the preservational bins defined in the text: A – high quality; B – inter-mediate quality; C – poor quality. Additionally, a valve dissolved and fragmented tooextensively to be included in this preservational analysis is included (D). This valvehas been heavily dissolved, leaving only trans-apical ribs, margins, and girdle bands.Because it has fragmented and the size of the gap in the valve face (all of the areolaeconjoined into a single gap) cannot be measured, it would not be included inanalysis.
Fig. 7. Validation of morphological binning. Perimeters of the areolae of Fragilar-ipsis curta preserved in core NBP0101-JPC24 were measured and averaged. Thesedata are compared to the ratios of preservational bins defined in the text, calculatedaccording to Eq. (2). Both curves produce the same record of diatom dissolution,validating the faster and less expensive binning approach.
J.P. Warnock, R.P. Scherer / Continental Shelf Research 102 (2015) 1–8 7
size, e.g. 100 total valves. Data from individually measured areolaeand binned data produce the same trends in diatom dissolution insediment core JPC24 (Fig. 7). Morphological binning provides areasonable measure of diatom dissolution in core material, andcan be used in Southern Ocean sediments to produced taxonomicdata related to water column dissolution processes, compliment-ing existing proxies of post-depositional dissolution (Dymondet al., 1997).
4. Discussion and conclusions
Variations in diatom dissolution have impacts on silica andcarbon cycling on glacial/interglacial timescales as well as inter-pretation of diatom-based paleoclimate records. Assessing the le-vel of dissolution experienced by preserved diatom communitiescan provide insight into nutrient cycling in the water column andthe efficiency of the biological pump. Given the calibration pro-duced here, assemblage-based preservational indices can bequickly assessed during standard diatom census counts on a lightmicroscope. Aside from performing a single, simple calculation, noadditional work beyond the census count is needed to generateobjective and reproducible preservational data. However, becausesedimentary diatom assemblages are affected by a number offactors, additional morphological data can be generated via SEM orhigh contrast LM. Morphological data provides a more detailedassessment of diatom preservation and separates taphonomicfrom other sedimentological or ecological changes to sedimentarydiatom assemblages. In contrast with reactive metal proxies ofpost-depositional dissolution, this method allows for an assess-ment of diatom dissolution in the water column.
Assessing diatom dissolution can provide meaningful data toaugment diatom abundance and biogenic silica data for SouthernOcean cores and sediment samples. Morphologically-based dis-solution data can separate preservational and productivity effectsfrom changes to diatom accumulation, providing new insight intopaleoceanographic reconstructions. Using morphologically-basedpreservational bins to simplify the data collection process does notalter the record of diatom dissolution. The community-basedmethod described here is currently applicable only to near shoresea-ice related habitats, but a parallel study utilizing F. kergue-lensis, the most common open ocean diatom in Antarctic waters,yielded identical morphological results (Warnock, 2013) indicatingthat this approach is applicable across the morphologically andecologically diverse genus Fragilariopsis.

J.P. Warnock, R.P. Scherer / Continental Shelf Research 102 (2015) 1–88
Acknowledgements
This research was funded by NSF Grant ANT-1043690. Thanksto Dr. Rob Dunbar for providing sample material. Thanks also toAmy Leventer and Leanne Armand for their helpful commentsalong the way. We also thank our reviewers for their valuableinsights, which strengthened the paper.
References
Armand, L., Crosta, X., Romero, O., Pichon, J.-J., 2005. The biogeography of majordiatom taxa in Southern Ocean sediments: 1. Sea-ice related species. Palaeo-geogr. Palaeoclimatol. Palaeoecol. 223, 93–126.
Assmy, P., Henjes, J., Smetacek, V., Montresor, M., 2006. Auxospore formation by thesilica-sinking, oceanic diatom Fragilariopsiskerguelensis (Bacillariophyceae). J.Phycol. 42, 1002–1006.
Bidle, K., Azam, F., 1999. Accelerated dissolution of diatom silica by marine bacterialassemblages. Nature 397, 508–512.
Crawford, S., Chiovitti, A., Pickett-Heaps, J., Wetherbee, R., 2009. Micro-morphogenesis during diatom wall formation produces siliceous nanos-tructures with different properties. J. Phycol. 45, 1353–1362.
Crosta, X., 2009. Holocene size variations in two diatom species off East Antarctica:productivity vs. environmental conditions. Deep Sea Res. 56, 1983–1993.
DeMaster, D., 2002. The accumulation and cycling of biogenic silica in the SouthernOcean: revisiting the marine silica budget. Deep-Sea Res. II 49, 3155–3167.
Dove, P.M., Han, N., Wallace, A.F., De Yoreo, J.J., 2008. Kinetics of amorphous silicadissolution and the paradox of the silica polymorphs. Proc. Natl. Acad. Sci. 105(29), 9903–9908.
Dunbar, R., Arrigo, K., Lutz, M., DiTullio, G., Leventer, A., Lizotte, M., Van Woert, M.,Robinson, D., 2003. Non-Redfield production and export of marine organicmatter: a recurrent part of the annual cycle in the Ross Sea, Antarctica. Antarct.Res. Ser. 78, 179–195.
Dymond, J., Collier, R., McManus, J., Honjo, S., Manganini, S., 1997. Can the alumi-num and titanium contents of ocean sediments be used to determine the pa-leoproductivity of the oceans? Paleoceanography 12, 586–593.
Flower, R., 1993. Diatom preservation: experiments and observations on dissolutionand breakage in modern and fossil material. Hydrobiologia 269/270, 473–484.
Icenhower, J.P., Dove, P.M., 2000. The dissolution kinetics of amorphous slica intosodium chloride solutions: effects of temperature and ionic strength. Geochim.Cosmochim. Acta 64 (24), 4193–4203.
Leventer, A., Domack, E., Dunbar, R., Pike, J., Stickley, C., Maddison, E., Brachfeld, S.,Manley, P., McClennen, C., 2006. Marine sediment record from the East Ant-arctic margin reveals dynamics of ice sheet recession. GSA Today 16 (12), 4–10.
Leventer, A., Dunbar, R., 1996. Factors influencing the distribution of diatoms andother algae in the Ross Sea. J. Geophys. Res. 101 (C8), 18489–18500.
Leventer, A., 1992. Modern distributions of diatoms in sediments form the George VCoast, Antarctica. Mar. Micropaleontol. 19, 315–332.
Lončarić, N., van Iperen, J., Kroon, D., Brummer, G.-J., 2007. Seasonal export andsediment preservation of diatomaceous, foraminiferal and organic matter mass
fluxes in a trophic gradient across the SE Atlantic. Prog. Oceanogr. 73, 27–59.Mikkelsen, N., 1977. Silica dissolution and overgrowth on fossil diatoms. Micro-
paleontology 23, 223–226.Morales, L.V., Sigman, D.M., Horn, M.G., Robinson, R.S., 2013. Cleaning methods for
the isotopic determination of diatom-bound nitrogen in non-fossil diatomfrustules. Limnol. Oceanogr. Methods 11, 101–112.
Nelson, D., Tréguer, P., Brzezinski, M., Leynaert, A., Quéguiner, B., 1995. Productionand dissolution of biogenic silica in the ocean: revised global estimates, com-parison with regional data and relationship to biogenic sedimentation. Glob.Biogeochem. Cycles 9 (3), 359–372.
Pichon, J.-J., Bareille, G., Labracherie, M., Labeyrie, L., Baudrimont, A., Turon, J.-L.,1992. Quantification of the biogenic silica dissolution in the Southern Oceansediments. Quat. Res. 37, 361–378.
Ragueneau, O., Tréguer, P., Leynaert, A., Anderson, R., Brzezinski, M., DeMaster, D.,Dugdale, R., Dymond, J., Fischer, G., François, R., Heinze, C., Maier-Reimer, E.,Martin-Jézéquel, V., Nelson, D., Quéguiner, B., 2000. A review of the Si cycle inthe modern ocean: recent progress and missing gaps in the application ofbiogenic opal as a paleoproductivity proxy. Glob. Planet. Change 26, 317–365.
Ryves, D., Battarbee, R., Fritz, S., 2009. The dilemma of disappearing diatoms: in-corporating diatom dissolution data into paleoenvironmental modeling andreconstruction. Quat. Sci. Rev. 28, 120–136.
Ryves, D., Battarbee, R., Juggins, S., Fritz, S., Anderson, J., 2006. Physical and che-mical predictors of diatom dissolution in freshwater and saline lake sedimentsin North America and West Greenland. Limnol. Oceanogr. 51 (3), 1355–1368.
Ryves, D., Jewson, D., Sturm, M., Battarbee, R., Flower, R., Anson, M., Granin, N.,2003. Quantitative and qualitative relationships between planktonic diatomcommunities and diatom assemblages in sedimenting material and surfacesediments in Lake Baikal, Siberia. Limnol. Oceanogr. 48 (4), 1643–1661.
Ryves, D., Juggins, S., Frizt, S., Battarbee, R., 2001. Experimental diatom dissolutionand the quantification of microfossil preservation in sediments. Paleogeogr.Paleoclimatol. Paleoecol. 172, 99–113.
Sarthou, G., Timmermans, K., Blain, S., Tréguer, P., 2005. Growth physiology and fateof diatoms in the ocean: a review. J. Sea Res. 53, 25–42.
Seeberg-Elverfeldt, I., Lange, C., Pätzold, J., 2004. Preservation of siliceous micro-plankton in surface sediments of the northern Red Sea. Mar. Micropaleontol. 51,193–211.
Shemesh, A., Burckle, L., Froelich, P., 1989. Dissolution and preservation of Antarcticdiatoms and the effect on sediment thanatocoenoses. Quat. Res. 31, 288–308.
Shimada, C., Hasegawa, S., Tanimura, Y., Burckle, L., 2003. A new index to quantifydiatom dissolution levels based on a ratio of Neodenticula seminae frustulecomponents. Micropaleontol. 49 (3), 267–276.
Tréguer, P., Nelson, D., Van Bennekom, A., DeMaster, D., Leynaert, A., Quéguiner, B.,1995. The silica balance in the world ocean: a reestimate. Science 268, 375–379.
Warnock, J., 2013. Diatom dissolution in the Southern Ocean: Experimental DiatomTaphonomy with Application to Pliocene to Holocene Antarctic Sediments.Northern Illinois University, p. 206, Doctoral Dissertation.
Warnock, J., Scherer, R., Loubere, P., 2007. A quantitative assessment of diatomdissolution and late quaternary primary productivity in the Eastern EquatorialPacific. Deep Sea Res. 54, 772–783.
Wassmann, P., 1998. Retention versus export food chains: processes controllingsinking loss from marine pelagic systems. Hydrobiologia 363, 29–57.




















