
Determinants of thermal balance in the Hawaiian giant rosette plant, Argyroxiphium sandwicense
8
International Association for Ecology Determinants of Thermal Balance in the Hawaiian Giant Rosette Plant, Argyroxiphium sandwicense Author(s): P. J. Melcher, G. Goldstein, F. C. Meinzer, B. Minyard, T. W. Giambelluca and L. L. Loope Source: Oecologia, Vol. 98, No. 3/4 (1994), pp. 412-418 Published by: Springer in cooperation with International Association for Ecology Stable URL: http://www.jstor.org/stable/4220716 . Accessed: 09/10/2013 17:43 Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at . http://www.jstor.org/page/info/about/policies/terms.jsp . JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range of content in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new forms of scholarship. For more information about JSTOR, please contact [email protected]. . Springer and International Association for Ecology are collaborating with JSTOR to digitize, preserve and extend access to Oecologia. http://www.jstor.org This content downloaded from 147.129.130.129 on Wed, 9 Oct 2013 17:43:11 PM All use subject to JSTOR Terms and Conditions
Transcript of Determinants of thermal balance in the Hawaiian giant rosette plant, Argyroxiphium sandwicense

International Association for Ecology
Determinants of Thermal Balance in the Hawaiian Giant Rosette Plant, ArgyroxiphiumsandwicenseAuthor(s): P. J. Melcher, G. Goldstein, F. C. Meinzer, B. Minyard, T. W. Giambelluca and L. L.LoopeSource: Oecologia, Vol. 98, No. 3/4 (1994), pp. 412-418Published by: Springer in cooperation with International Association for EcologyStable URL: http://www.jstor.org/stable/4220716 .
Accessed: 09/10/2013 17:43
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
Springer and International Association for Ecology are collaborating with JSTOR to digitize, preserve andextend access to Oecologia.
http://www.jstor.org
This content downloaded from 147.129.130.129 on Wed, 9 Oct 2013 17:43:11 PMAll use subject to JSTOR Terms and Conditions

Oecologia (1994) 98:412-418
ORIGINAL PAPER
? Springer Verlag 1994
PJ. Melcher ? G. Goldstein ? F.C. Meinzer ? ?. Minyard T.W. Giambelluca ? L.L. Loope
Determinants of thermal balance in the Hawaiian giant rosette plant,
Argyroxiphium sandwicense
Received: 1 October 1993/Accepted: 14 April 1994
Abstract The effects of leaf pubescence and rosette ge- ometry on thermal balance were studied in a subspecies of a Hawaiian giant rosette plant, Argyroxiphium sand- wicense. This species, a member of the silversword alli-
ance, grows above 2000 m elevation in the alpine zone of two Hawaiian volcanoes. Its highly pubescent leaves are
very reflective (absorptance in the 400-700 nm wave- band = 0.44). Temperature of the expanded leaves was
very similar to, or even lower than, air temperature dur-
ing clear days, which was somewhat surprising given that solar radiation at the high elevation sites where this
species grows can exceed 1100W nr2. However, the
temperature of the apical bud, which is located in the center of the parabolic rosette, was usually 25?C higher than air temperature at midday. Experimental manipula- tions in the field indicated that incoming solar radiation
being focussed towards the center of the rosette resulted in higher temperatures of the apical bud. Attenuation of wind speed inside the rosette, which increased the thick- ness of the boundary layer surrounding the apical bud, also contributed to higher temperatures. The heating ef- fect on the apical bud of the large parabolic rosette, which apparently enhances the rates of physiological processes in the developing leaves, may exclude the spe- cies from lower elevations by producing lethal tissue
temperatures. Model simulations of apical bud tempera- tures at different elevations and laboratory estimates of the temperature threshold for permanent heat injury pre-
dieted that the lower altitude limit should be approxi- mately 1900 m, which is reasonably close to the lower limit of distribution of A. sandwicense on Haleakala vol- cano.
Key words Argyroxiphium sandwicense
Energy balance ? Heat tolerance ? Leaf pubescence Silversword
Introduction
Giant rosette plants have evolved independently in sever- al high elevation tropical regions. This taxonomically di- verse assemblage of species with similar overall morpho- logical features represents a well-known example of con-
vergent evolution. For example, giant rosettes belonging to the genera Senecio and Lobelia grow at elevations over 5000 m in the equatorial mountains of Africa (Hed-
berg 1964), while members of the genus Espeletia are dominant in the South American Andes between 3200 and 4600 m elevation (Goldstein and Meinzer 1983, Goldstein et al. 1984, Meinzer and Goldstein 1986). Ar-
gyroxiphium sandwicense (silversword) is the only giant rosette species found in the alpine zone of the Hawaiian
archipelago (Carr 1985, Robichaux et al. 1990). The
high tropical environments where these giant rosettes
grow are characterized by greater diurnal than seasonal
temperature fluctuations, frequent nocturnal frosts, high radiant energy input, and relatively low daytime air tem-
peratures. Argyroxiphium sandwicense possesses a terminal ro-
sette of spirally arranged leaves separated by very com-
pressed internodes, and a central water storage system made up of parenchymatous pith tissue. The apical bud is massive and contains a large bank of unexpanded leaves. As the leaves expand and mature, the apical bud remains near the focus of a paraboloid formed by the
surrounding leaves. Considerable daytime heating of the
apical bud has been documented in A. sandwicense
(Robichaux et al. 1990), and in neotropical giant rosettes
PJ. Melcher ? G. Goldstein (Kl) Department of Botany, University of Hawaii, 3190 Maile Way, Honolulu, 96822, USA
EC. Meinzer Hawaiian Sugar Planters' Association, PO Box 1057, Aiea, 96701, USA
B. Minyard ? T.W. Giambelluca Department of Geography, University of Hawaii, 2424 Maile Way, Honolulu, 96822, USA
L.L. Loope Haleakala National Park, Box 369, Makawao, 96768, USA
This content downloaded from 147.129.130.129 on Wed, 9 Oct 2013 17:43:11 PMAll use subject to JSTOR Terms and Conditions

413
(Larcher 1975, Meinzer et al. 1985). It has been suggest- ed that the parabolic geometry of the rosette enhances
capture and retention of incident long and shortwave ra- diation (Meinzer et al. 1985). However, the influence of rosette geometry on energy storage in the apical region has never been experimentally tested.
Leaves of A. sandwicense, like those of several other
giant rosette species, are densely pubescent. The thick
pubescent layer on leaves of several Espeletia species modifies energy balance mainly by increasing boundary layer thickness and resistance to convective heat transfer
(Meinzer and Goldstein 1985, 1986, Goldstein et al.
1989). As a result, the temperature of Espeletia leaves tends to be substantially higher than air temperatures during the day. The trichomes of A. sandwicense are, however, shorter and apparently more reflective than those of Espeletia species, and it is possible that their role in temperature regulation is different.
Here we investigate the effects of leaf pubescence and
parabolic rosette geometry on thermal balance of A. sandwicense. Argyroxiphium sandwicense grows above 2000 m elevation on Haleakala (Maui) and Mauna Kea
(Hawaii) vol?anos (Powell 1992). Within Haleakala Na- tional Park, A. sandwicense subsp. macrocephalum grows between 2000 and 3000 m in elevation, and is lo-
cally dominant on cinder cones within the erosional cra- ter of the dormant volcano. Microclimatic variables and leaf temperatures were intensively monitored for 2 months and the results were compared with predictions of an energy balance model developed to assess the role that radiation, wind speed, and transpiration play in regu- lating temperatures in the apical region of the rosette. In addition, experimental manipulations were performed in the field to evaluate the effect of rosette geometry and solar radiation on apical bud temperatures, and laborato-
ry estimates were made of the temperature threshold for
permanent heat injury of leaf tissues.
Materials and methods
Plant material and site characteristics
Field studies on Argyroxiphium sandwicense DC (Asteraceae) (Wagner et al. 1990) were carried out in Haleakala National Park, on the island of Maui (latitude 20? 44' N, longitude 156? 15' W, elevation 2835 m). Five mature plants about 0.4 m high, with the radial symmetry typical of unbranched plants, were selected for intensive study. The vegetation of the site is open scrub with four dominant species: A. sandwicense, Dubautia menziesii, Vaccinium reticulatum, and Styphelia tameiameiae. The midday maximum solar radiation, air temperature, and mean relative humidity for the month of June 1992 were 938 W nr2, 16.6? C, and 41.9% respec- tively. This climatological information was obtained from a station located downslope from the study area at 2600 m elevation. The station is located above the mean inversion height and is similar to the study site in slope and aspect (Giambelluca and Nullet 1991).
Field studies
Several aspects of microclimate and plant response were intensive- ly monitored during June, July, and August 1992. Temperatures
were measured on the most recently expanded leaves and on inter- mediate-size expanding leaves within the rosette. For each leaf size class, the temperatures of leaves located nearest each cardinal point (N, E, S, and W) were measured. A 0.08 mm diameter (40 gauge) copper-constantan thermocouple was inserted beneath the pubescent layer to contact the upper leaf surface. Two thermocou- ples were also inserted in the apical bud in the center of the ro- sette. Global radiation was measured with Eppley (black and white) pyranometers (Model 8-48, The Eppley Laboratory, Inc., Newport, RI), net radiation with Fritschen-type net radiometers (Model Q-5, Radiation & Energy Balance Systems, Inc., Seattle, WA), air temperature and humidity with Vaisala MD-30-YB sen- sors (Vaisala, Inc., Woburn, MA), and wind speed with 3-cup ane- mometers (Met One, Inc., Grants Pass, OR), and with orthogonal- ly-arranged propeller-type anemometers (Model 27005UVX, R.M. Young Company, Traverse City, MI). Soil and air temperature were measured with copper-constantan thermocouples shielded from direct solar radiation. Data were sampled at 1 min intervals and average values were recorded every 10 min with two datalog- gers (Model CR-10, Campbell Scientific, Logan, UT). Total resis- tance to water vapor was measured with a steady state porometer (Model 1600, LI-COR, Lincoln, Nebraska) on both the adaxial and abaxial surfaces of the amphistomatous leaves. Sky tempera- ture was measured several times during the day and night with an infrared thermometer (Model 4000ALCS, Everest Interscience, Inc., Fullerton, CA) aimed towards each cardinal point at an eleva- tion angle of 30? (sufficient to eliminate ground influence).
Three experimental manipulations were performed in the field. One experiment was intended to determine the effect of rosette ge- ometry on apical bud temperature. Recently expanded leaves were either opened or closed 10? with respect to their normal position, and the plant and microenvironmental variables described above were monitored for 24 h. This experiment was repeated three times on each of two different plants. A second experiment con- sisted of shading the entire rosette with a board that eliminated di- rect solar radiation for approximately 30-min intervals. At the end of each interval the board was removed for another 30 min to al- low the plant to gradually reach thermal balance under unobstruct- ed conditions. The board was small enough to shade only the plant and it was positioned 1 m above the upper part of the rosette to avoid a substantial decrease in wind speed at the plant level. This experiment, aimed at determining the effect of abrupt changes in incoming radiation on the thermal balance of the plant, was car- ried out on clear days between 0900 and 1300 hours. The third ex- periment consisted of monitoring incoming short wave radiation inside the rosette near the apical bud and at an unobstructed site outside the rosette. The objective was to evaluate the possibility of enhanced capture of incoming solar radiation by the parabolic ro- sette. This experiment was repeated five times on different plants using two recently calibrated silicon photodiode pyranometers (model LI-190SZ ,LI-COR, Lincoln, Nebraska).
Energy balance model
In order to evaluate the effect of the pubescence on the thermal balance of the leaves an energy balance model, as applied by Ehle- ringer (1989), was used. The effect of leaf orientation and solar geometry on incoming short wave radiation was modeled using al- gorithms from Jones (1992). The energy balance equation was modified to incorporate the influence of the parabolic rosette ge- ometry on apical bud temperature at midday. In the modifications, it was assumed that short- and long-wave radiation are absorbed by only one horizontal leaf surface (versus both surfaces for the fully expanded leaves) with a characteristic dimension of 0.024 m (width of apical bud), an absorptance of 0.50 (typical of small ex- panding leaves), and that the incoming short wave radiation, as measured with a silicon photodiode pyranometer imbedded in the center of the rosette, was enhanced by the parabolic geometry of the rosette.
Leaf absorptance to solar radiation was measured with a spec- troradiometer and an integrating sphere (LI-COR LI 1800, Lin-
This content downloaded from 147.129.130.129 on Wed, 9 Oct 2013 17:43:11 PMAll use subject to JSTOR Terms and Conditions
414
coin, Nebraska). Values of total shortwave (400-3000 nm) ab- sorptances needed for the energy balance simulations were ob- tained from the 400-700 nm absorptances using the empirical re- lationship obtained by Ehleringer (1981a). The thickness of the pubescence on both the adaxial and abaxial leaf surfaces was mea- sured microscopically in transverse sections. Average width was determined for three mature fully expanded leaves from each of seven plants. The contribution of the pubescence layer to total boundary layer resistance to water vapor and sensible heat transfer was calculated as
r=L/D
where r is the resistance due to the pubescence layer, L is the thick- ness of the pubescence layer, and D is the diffusivity of sensible heat or water vapor in air (Nobel 1991). Longwave radiation from the sky was calculated using average sky temperatures measured with the infrared thermometer and the Stefan-Boltzman equation.
Laboratory studies
Heat tolerance of rosette leaves was evaluated with a method that measures the amount of electrolyte leakage through heat-damaged membranes of leaf tissue (Flint et al. 1967). Sections 1 cm long were cut from the middle part of the leaf. The ends were blotted to remove solutes released from cells injured during cutting, and the sections were immediately enclosed in small sealed vials to pre- vent transpirational water loss. Each vial contained three leaf sam- ples of about 0.17 g each. The vials were sealed inside plastic bags and immersed in a water bath. The bath temperature was then in- creased from 25 to 70?C in 5?C increments every 20 min. Three vials were removed at the end of each 20 min incubation period and brought slowly to ambient temperature (20 ?C). The leaf sec- tions from each vial were weighed and quickly placed in scintilla- tion vials containing 15 ml of deionized water. The sections were shaken at 20 RPM for 12 h, removed from the bathing solutions, and the conductivity of the solutions measured with a conductance meter (YSI model 32, Yellow Springs, Ohio). Solution conductivi- ty was expressed as a percentage of that measured in solution from vials containing samples kept in boiling water for 2 h. Another set of entire leaves was enclosed in sealed plastic bags, incubated at 38?C for 2 h, and kept at ambient temperatures for 8 h. Sections 1 cm long were cut from the middle part of these leaves, and thermal tolerance was determined as described above.
Results
A mature, 0.4 m-tall A. sandwicense plant typically had 350-450 expanded leaves arranged in a very regular spi- ral with a divergence angle between successive leaves of about 7? horizontally (between leaves in a whorl). The three innermost whorls of fully expanded leaves formed the effective wall of the paraboloid. The diameter of the
opening formed by the leaves in this zone was 15.2 ? 0.3 cm (mean ? 1 SE, ? = 15) and the distance between the bottom of the apical bud and the distal part of the fully expanded leaves (depth) was 13.5 cm. The diameter of the opening (y) and the depth (x) were used to calculate the focal point for the incoming direct solar radiation. If the rosette were a perfect paraboloid, the focal point (p) would be
p=y/4x
which gives 4.3 cm from the bottom of the paraboloid. Apical bud length ranged from 0.5 to 2 cm, reasonably close to the focus of the paraboloid.
8/8/92 7/20/92
o "s
o co
CD =3 s Q) Cl
O
1200
? 900
: 600
" 300
0
40
30
20
10
0
40
30
20
10
0 ,^^-?
12 18 0 6 12
Local time (h)
18 24
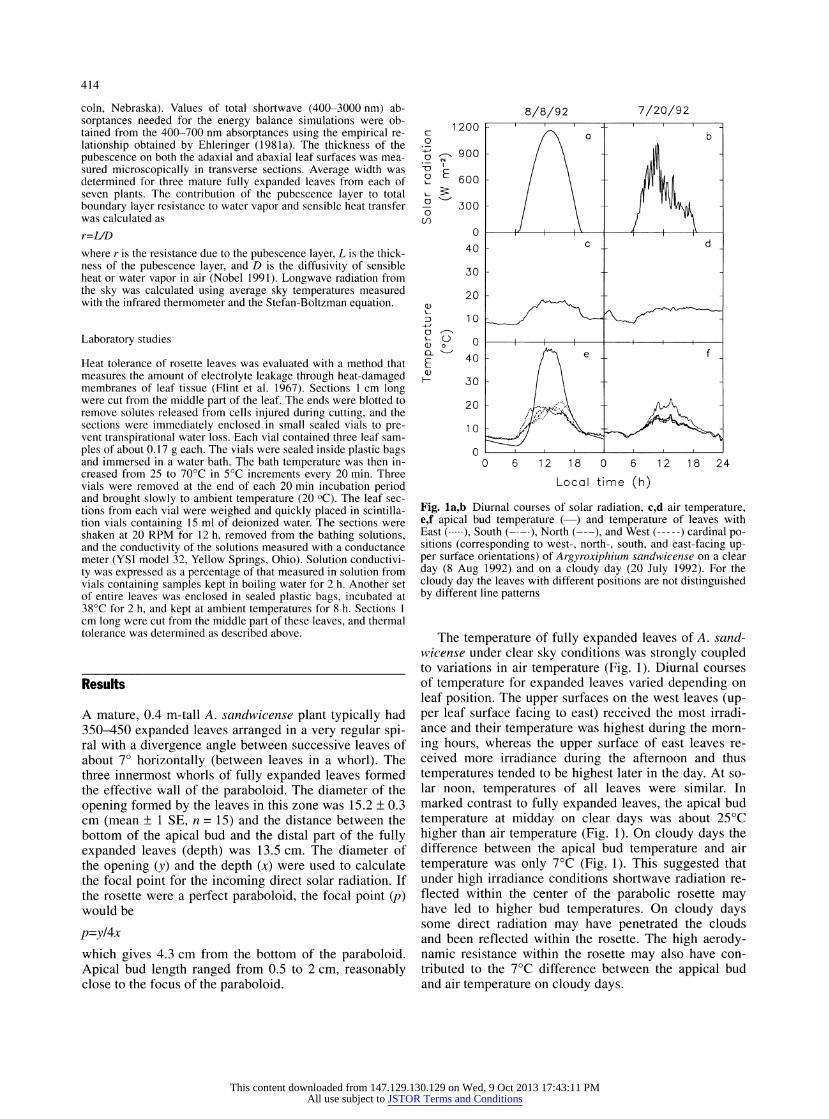
Fig. la,b Diurnal courses of solar radiation, c,d air temperature, e,f apical bud temperature (?) and temperature of leaves with East (.), South (-), North (-), and West (-) cardinal po- sitions (corresponding to west-, north-, south, and east-facing up- per surface orientations) of Argyroxiphium sandwicense on a clear day (8 Aug 1992) and on a cloudy day (20 July 1992). For the cloudy day the leaves with different positions are not distinguished by different line patterns
The temperature of fully expanded leaves of A. sand- wicense under clear sky conditions was strongly coupled to variations in air temperature (Fig. 1). Diurnal courses of temperature for expanded leaves varied depending on leaf position. The upper surfaces on the west leaves (up- per leaf surface facing to east) received the most irradi- ance and their temperature was highest during the morn-
ing hours, whereas the upper surface of east leaves re- ceived more irradiance during the afternoon and thus
temperatures tended to be highest later in the day. At so- lar noon, temperatures of all leaves were similar. In marked contrast to fully expanded leaves, the apical bud
temperature at midday on clear days was about 25 ?C
higher than air temperature (Fig. 1). On cloudy days the difference between the apical bud temperature and air
temperature was only 7?C (Fig. 1). This suggested that under high irradiance conditions shortwave radiation re- flected within the center of the parabolic rosette may have led to higher bud temperatures. On cloudy days some direct radiation may have penetrated the clouds and been reflected within the rosette. The high aerody- namic resistance within the rosette may also have con- tributed to the 7?C difference between the appical bud and air temperature on cloudy days.
This content downloaded from 147.129.130.129 on Wed, 9 Oct 2013 17:43:11 PMAll use subject to JSTOR Terms and Conditions
415
o
s '"?
? co
1200 -
800 -
400
0 6 12 1?
Local time (h)
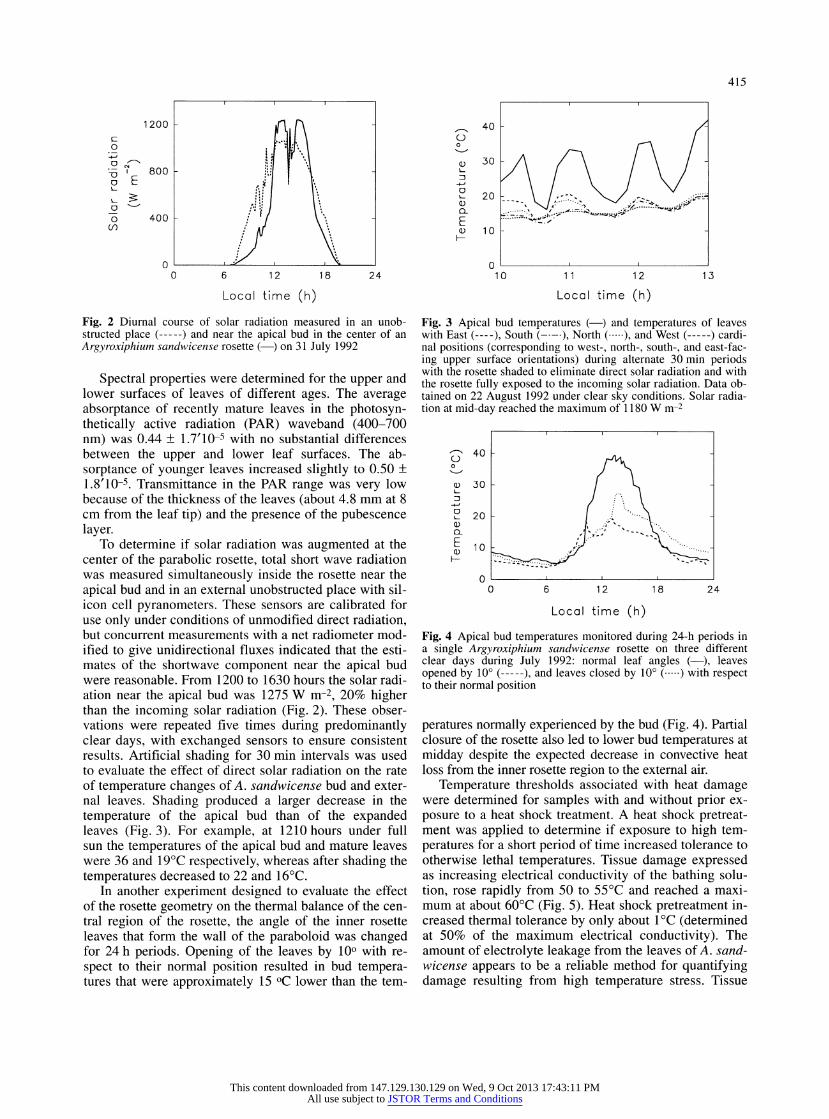
Fig. 2 Diurnal course of solar radiation measured in an unob- structed place (-) and near the apical bud in the center of an Argyroxiphium sandwicense rosette (?) on 31 July 1992
Spectral properties were determined for the upper and lower surfaces of leaves of different ages. The average absorptance of recently mature leaves in the photosyn- thetically active radiation (PAR) waveband (400-700 nm) was 0.44 ? 1.7?0-5 with no substantial differences between the upper and lower leaf surfaces. The ab-
sorptance of younger leaves increased slightly to 0.50 ? 1.8?0-5. Transmittance in the PAR range was very low because of the thickness of the leaves (about 4.8 mm at 8 cm from the leaf tip) and the presence of the pubescence layer.
To determine if solar radiation was augmented at the center of the parabolic rosette, total short wave radiation was measured simultaneously inside the rosette near the
apical bud and in an external unobstructed place with sil- icon cell pyranometers. These sensors are calibrated for use only under conditions of unmodified direct radiation, but concurrent measurements with a net radiometer mod- ified to give unidirectional fluxes indicated that the esti- mates of the shortwave component near the apical bud were reasonable. From 1200 to 1630 hours the solar radi- ation near the apical bud was 1275 W mr2, 20% higher than the incoming solar radiation (Fig. 2). These obser- vations were repeated five times during predominantly clear days, with exchanged sensors to ensure consistent results. Artificial shading for 30 min intervals was used to evaluate the effect of direct solar radiation on the rate of temperature changes of A. sandwicense bud and exter- nal leaves. Shading produced a larger decrease in the
temperature of the apical bud than of the expanded leaves (Fig. 3). For example, at 1210 hours under full sun the temperatures of the apical bud and mature leaves were 36 and 19?C respectively, whereas after shading the
temperatures decreased to 22 and 16?C. In another experiment designed to evaluate the effect
of the rosette geometry on the thermal balance of the cen- tral region of the rosette, the angle of the inner rosette leaves that form the wall of the paraboloid was changed for 24 h periods. Opening of the leaves by 10? with re-
spect to their normal position resulted in bud tempera- tures that were approximately 15 ?C lower than the tern-
o o
(? L_ Z5 ?+-> O v_ CD Q_ E
11 12
Local time (h)
13
Fig. 3 Apical bud temperatures (?) and temperatures of leaves with East (-), South (-), North (.), and West (-) cardi- nal positions (corresponding to west-, north-, south-, and east-fac- ing upper surface orientations) during alternate 30 min periods with the rosette shaded to eliminate direct solar radiation and with the rosette fully exposed to the incoming solar radiation. Data ob- tained on 22 August 1992 under clear sky conditions. Solar radia- tion at mid-day reached the maximum of 1180 W nr2
6 12 18
Local time (h)
24
Fig. 4 Apical bud temperatures monitored during 24-h periods in a single Argyroxiphium sandwicense rosette on three different clear days during July 1992: normal leaf angles (?), leaves opened by 10? (-), and leaves closed by 10? (.) with respect to their normal position
peratures normally experienced by the bud (Fig. 4). Partial closure of the rosette also led to lower bud temperatures at
midday despite the expected decrease in convective heat loss from the inner rosette region to the external air.
Temperature thresholds associated with heat damage were determined for samples with and without prior ex-
posure to a heat shock treatment. A heat shock pretreat- ment was applied to determine if exposure to high tem-
peratures for a short period of time increased tolerance to otherwise lethal temperatures. Tissue damage expressed as increasing electrical conductivity of the bathing solu-
tion, rose rapidly from 50 to 55?C and reached a maxi- mum at about 60?C (Fig. 5). Heat shock pretreatment in- creased thermal tolerance by only about 1?C (determined at 50% of the maximum electrical conductivity). The amount of electrolyte leakage from the leaves of A. sand- wicense appears to be a reliable method for quantifying damage resulting from high temperature stress. Tissue
This content downloaded from 147.129.130.129 on Wed, 9 Oct 2013 17:43:11 PMAll use subject to JSTOR Terms and Conditions
416
100
? ?)
"? C ? ?
s ?
? =5 ? ? ?
Temperature (?C)
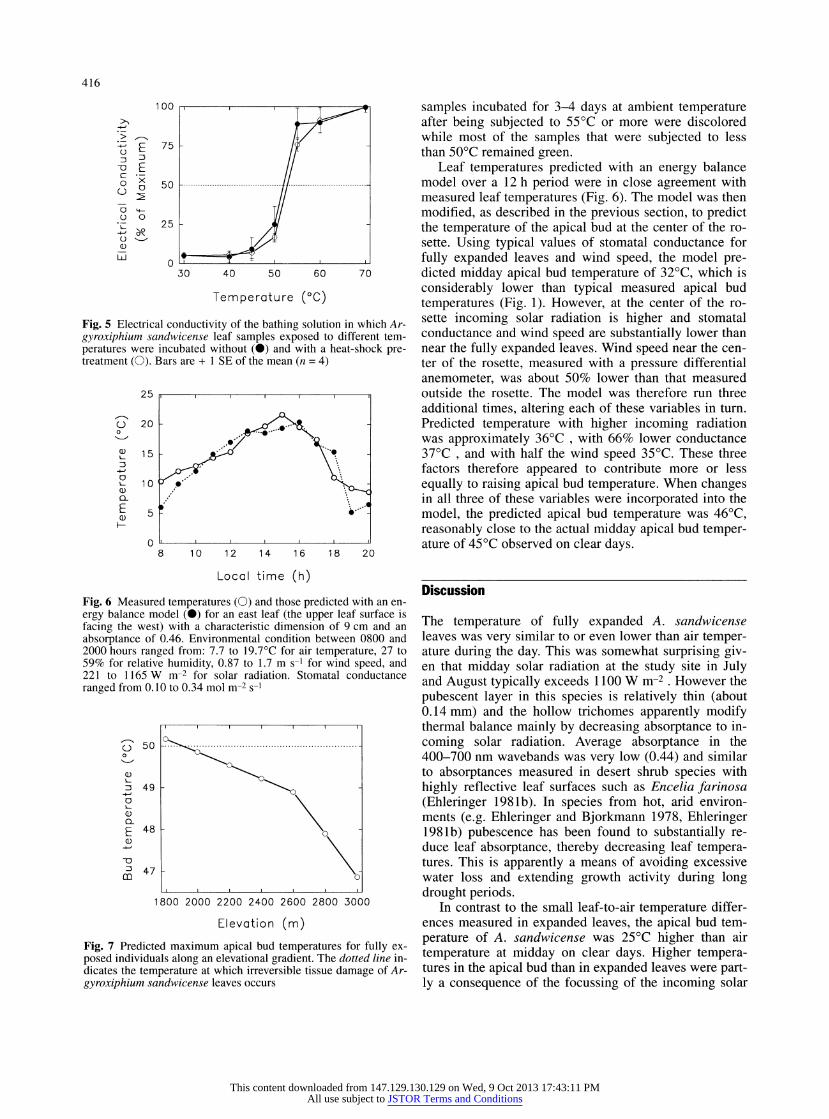
Fig. 5 Electrical conductivity of the bathing solution in which Ar- gyroxiphium sandwicense leaf samples exposed to different tem- peratures were incubated without (?) and with a heat-shock pre- treatment (O). Bars are + 1 SE of the mean (n = 4)
O o
CD L. Z3 ? V- (D CL E CD
10 12 14 16 18
Local time (h)
20
Fig. 6 Measured temperatures (O) and those predicted with an en- ergy balance model (?) for an east leaf (the upper leaf surface is facing the west) with a characteristic dimension of 9 cm and an absorptance of 0.46. Environmental condition between 0800 and 2000 hours ranged from: 7.7 to 19.7?C for air temperature, 27 to 59% for relative humidity, 0.87 to 1.7 m s-1 for wind speed, and 221 to 1165W nr2 for solar radiation. Stomatal conductance ranged from 0.10 to 0.34 mol ver2 s~l
1800 2000 2200 2400 2600 2800 3000
Elevation (m)
Fig. 7 Predicted maximum apical bud temperatures for fully ex- posed individuals along an elevational gradient. The dotted line in- dicates the temperature at which irreversible tissue damage of Ar- gyroxiphium sandwicense leaves occurs
samples incubated for 3-4 days at ambient temperature after being subjected to 55?C or more were discolored while most of the samples that were subjected to less than 50?C remained green.
Leaf temperatures predicted with an energy balance model over a 12 h period were in close agreement with measured leaf temperatures (Fig. 6). The model was then
modified, as described in the previous section, to predict the temperature of the apical bud at the center of the ro- sette. Using typical values of stomatal conductance for
fully expanded leaves and wind speed, the model pre- dicted midday apical bud temperature of 32?C, which is
considerably lower than typical measured apical bud
temperatures (Fig. 1). However, at the center of the ro- sette incoming solar radiation is higher and stomatal conductance and wind speed are substantially lower than near the fully expanded leaves. Wind speed near the cen- ter of the rosette, measured with a pressure differential
anemometer, was about 50% lower than that measured outside the rosette. The model was therefore run three additional times, altering each of these variables in turn. Predicted temperature with higher incoming radiation was approximately 36?C , with 66% lower conductance 37?C , and with half the wind speed 35?C. These three factors therefore appeared to contribute more or less
equally to raising apical bud temperature. When changes in all three of these variables were incorporated into the
model, the predicted apical bud temperature was 46?C,
reasonably close to the actual midday apical bud temper- ature of 45?C observed on clear days.
Discussion
The temperature of fully expanded A. sandwicense leaves was very similar to or even lower than air temper- ature during the day. This was somewhat surprising giv- en that midday solar radiation at the study site in July and August typically exceeds 1100 W nr2 . However the
pubescent layer in this species is relatively thin (about 0.14 mm) and the hollow trichomes apparently modify thermal balance mainly by decreasing absorptance to in-
coming solar radiation. Average absorptance in the 400-700 nm wavebands was very low (0.44) and similar to absorptances measured in desert shrub species with
highly reflective leaf surfaces such as Encella farinosa
(Ehleringer 1981b). In species from hot, arid environ- ments (e.g. Ehleringer and Bjorkmann 1978, Ehleringer 1981b) pubescence has been found to substantially re- duce leaf absorptance, thereby decreasing leaf tempera- tures. This is apparently a means of avoiding excessive water loss and extending growth activity during long drought periods.
In contrast to the small leaf-to-air temperature differ- ences measured in expanded leaves, the apical bud tem-
perature of A. sandwicense was 25?C higher than air
temperature at midday on clear days. Higher tempera- tures in the apical bud than in expanded leaves were part- ly a consequence of the focussing of the incoming solar
This content downloaded from 147.129.130.129 on Wed, 9 Oct 2013 17:43:11 PMAll use subject to JSTOR Terms and Conditions

417
radiation onto the apical bud. The rosette is, however, far from being a perfect parabola. Energy balance simula- tions predicted that if the rosette leaves formed a continu- ous wall (as if it consisted of a single parabolic leaf) the
temperature of the apical bud would reach 130?C on clear
days. Experimental manipulations in the field and energy balance simulations suggested that relatively low wind
speeds in the protected center of the parabolic rosette also contribute to the higher temperatures observed in the api- cal bud. The geometry of the rosette influences the atten- uation of wind, and thus the thickness of the boundary layer surrounding the apical bud. This aerodynamic effect enhances heating of the apical bud during the day. The same mechanisms also promote relatively low nocturnal bud temperatures. At high elevation sites where A. sand- wicense grows, net radiation is strongly negative at night because of rapid longwave loss into the thin, clear atmo-
sphere (Giambelluca and Nullet 1991). During a typical clear summer night, when the air temperature is 8?C, the
temperature of expanded leaves falls to 7?C, but the tem-
perature of the massive apical bud is usually below 4?C.
During the winter when nighttime air temperatures are
frequently below 0?C, the lower bud temperatures would increase the risk of freezing. Freezing exotherms, sudden, transient increases in temperature, have been observed in the apical bud of A. sandwicense during winter nights (CC. Lipp, G. Goldstein, EC. Meinzer, W. Niemczura, unpublished results). Leaf tissue of A. sandwicense toler- ates extracellular ice formation during short periods at
night because damage to the apical bud did not occur in the plants in which exothermic events were observed. Other giant rosette species have also been reported to tol- erate extracelluar freezing, and nocturnal intracellular de-
hydration (Beck et al. 1982, 1984, Azocar et al. 1988,
Squeoetal. 1991). There are several reasons why the high midday tem-
peratures observed in the apical bud of A. sandwicense could be of adaptive value during the winter season. The mean daily temperature of 7?C measured near the study site during the coldest months of the year may be subop- timal for translocation and growth (Leopold and Kriedemann 1975). Since a typical Q10 for these process- es is approximately 2, a 20 to 25?C increase in tempera- ture of the apical bud could increase the rate of leaf
growth by a factor of four. Nevertheless, enhanced day- time heating of the apical bud could be disadvantageous during the summer or at lower elevations. Temperatures greater than 50?C cause irreversible injury to leaf tissue of A. sandwicense. Maximum apical bud temperatures of
approximately 45?C were recorded on several occasions
during July in our field site at 2835 m. It is thus possible that apical bud temperature may attain levels associated with permanent tissue damage during unusually warm, still days at high elevations or under the warmer condi- tions prevailing at lower elevations. Brief exposure to
moderately elevated temperatures is known to increase tolerance to otherwise lethal high temperatures in several
plant and animal species (Lindquist 1987); however its effects on A. sandwicense were minimal (Fig. 5). Accli-
mation to increasing growth temperatures for extended
periods (days to months) is common among vascular
plants (Seemann et al. 1986, Nobel 1988). For example, the threshold for the onset of irreversible tissue damage is 63.9?C for 18 species of cacti grown at 50?C/40?C
day/night temperatures, and increases to 69.2?C per 10?C increase in growth temperatures (Nobel 1988).
Long-term acclimation treatments to moderately high temperatures were performed with small A. sandwicense
plants (G. Goldstein and D. Drake unpublished informa-
tion) but increased tolerance to high temperature stress was not observed.
In order to assess the potential role of apical bud heat-
ing in survival of A. sandwicense at lower elevations, the
energy balance model was used to predict maximum api- cal bud temperatures as a function of elevation (Fig. 7). The highest midday air temperatures observed during Ju-
ly and August were used to run the model. The tempera- ture lapse rate between 1800 and 2600 m elevation is
very small (about 0.002?C irr1) because of a nearly per- manent fog zone at those elevations during the summer months. The temperature lapse is substantially steeper above 2600 m (above the persistent subsidence inversion characteristic of the region) where air is extremely dry due to its isolation from the oceanic moisture source (Gi- ambelluca and Nullet 1991). According to the model, the lethal bud temperature of 50 ?C for fully exposed indi- viduals would be reached at about 1900 m elevation (Fig. 7). This coincides well with the lowest elevation ob- served at the beginning of the century (1890 m) for A. sandwicense prior to the substantial reduction of the
population range by human activities (Rock 1974). It is
interesting to speculate concerning possible changes in
morphology that could result in lower maximum bud
temperatures thereby extending the lower altitudinal lim- it of A. sandwicense. Changes in the architecture of the
rosette, such as an increase or decrease of the leaf an-
gles, could displace the focus of the paraboloid away from the position of the apical bud, thus lowering the
apical bud temperature. Leaf angle and opening of the rosette were measured in 40 adult plants from several
populations in Haleakala National Park and no substan- tial differences were observed (unpublished results). This
suggests that the degree of morphological variation in A. sandwicense individuals from several populations is much less than that previously reported for an Andean
giant rosette species growing along a comparable altitu- dinal and climatic gradient (Meinzer et al. 1985). Appar- ently A. sandwicense has a reduced capacity for morpho- logical adjustment. Substantial reduction of A. sand- wicense population size early in this century, due to re- moval by visitors to the volcano and browsing by cattle and goats (Loope and Crivellone 1986), may have in- duced a genetic bottleneck, and with that the loss of some of the genetic variation necessary for morphologi- cal plasticity.
Argyroxiphium grayanum (greensword), which occurs in bogs at lower elevations on Maui, is closely related to A. sandwicense. Argyroxiphium grayanum is also a giant
This content downloaded from 147.129.130.129 on Wed, 9 Oct 2013 17:43:11 PMAll use subject to JSTOR Terms and Conditions

418
rosette plant but its morphology and the reflectance of its rosette leaves are considerably different from those of A. sandwicense. For example, A. grayanum leaves are
green, suggesting that their reflectance is much lower than that of A. sandwicense. The morphological differ- ences between these two giant rosettes with A. grayanum being less parabolic in appearance may reflect adaptive modifications selected in their respective environments. Lower leaf reflectance and less focussing of the incom-
ing light would not result in extremely high apical bud
temperatures at the lower elevations where A. grayanum is found. On the other hand, the morphological charac- teristics of A. sandwicense result in higher apical bud
temperatures, which are apparently of adaptive value in
the colder habitats were it occurs.
Whereas the Mauna Kea silversword subspecies is on
the verge of extinction, the population of A. sandwicense
subsp. macrocephalum in Haleakala National Park ap-
pears to be stable or increasing in size with about 50,000 individuals (Kobayashi 1973, Loope and Crivellone
1986). Although locally numerous, the Haleakala silver-
sword is rare and potentially vulnerable to extinction be-
cause of its extremely restricted geographic range and its
narrow habitat requirements. The area within the perime- ter of its range is no more than 50 km2. Some of the mor-
phological and physiological characteristics of this spe- cies apparently contribute to its narrow range confining it
to high elevations. Tolerance of extracellular freezing al-
lows this species to survive short episodes of freezing
temperature at high elevation. On the other hand, the
heating effect on the apical bud of the large parabolic ro-
sette, which apparently enhances the rates of physiologi- cal processes in the developing leaves, may exclude the
species from lower elevations through producing lethal
tissue temperatures.
Acknowledgements We thank W. Buscher for assistance in the field, R. Robichaux for helpful comments and suggestions, and M. Cui for assistance with leaf spectral properties measurements.
References
Azocar A, Rada F, Goldstein G (1988) Freezing tolerance in Draba chianophila, a miniature caulescent rosette species. Oecologia 75:156-160
Beck E, Senser W, Scheibe R, Steiger HM, Pongratz ? (1982) Frost avoidance and freezing tolerance in Afroalpine 'giant ro- sette' plants. Plant Cell Environ 5:215-222
Beck E, Schulze E, Senser W, Scheibe R (1984) Equilibrium freezing of leaf water and extracellular ice formation in afroal- pine 'giant rosette' plants. Planta 162:276-282
Carr GD (1985) Monograph of the Hawaiian Madiinae (Aster- aceae): Argyroxiphium, Dubautia, and Wilkesia. Allertonia 4:1-123
Ehleringer J (1981a) Leaf absorptances of Mojave and Sonoran Desert plants. Oecologia 49:366-370
Ehleringer J (1981b) Leaf absorptances of Mohave and Sonoran desert plants. Oecologia 49:366-370
Ehleringer JR (1989) Temperature and energy budgets. In: Pearcy RW, Ehleringer J, Mooney A, Rundel PW (eds) Plant Physio- logical Ecology: Field Methods and Instrumentation. Chap- man and Hall, New York, pp 117-135
Ehleringer J, Bjorkmann O (1978) Pubescence and leaf spectral characteristics in a desert shrub, Encella farinosa. Oecologia 36:151-162
Flint HL, Boyce BR, Beattue DU (1967) Index of injury. A useful expression of freezing injury to plant tissues as determined by the electrolyte method. Can J Plant Sci 47:229-230
Giambelluca TW, Nullet D (1991) Influence of the trade-wind in- version on the climate of a leeward mountain slope in Hawaii. Climate Research 1: 207-216
Goldstein G, Meinzer FC (1983) Influence of insulating dead leaves and low temperatures on water balance in an andean gi- ant rosette plant. Plant Cell Environ. 6:649-656
Goldstein G, Meinzer FC, Monasterio M (1984) The role of ca- pacitance in the water balance of andean giant rosette species. Plant Cell Environ 7:179-186
Goldstein G, Rada F, Canales MO, Zabala O (1989) Leaf gas ex- change of two giant caulescent rosette species. Acta Oecol 10: 359-370
Hedberg O (1964) Features of afroalpine plant ecology. Acta Phytogeogr. Suecia 49:1-144
Jones HG (1992) Plants and Microclimate: A quantitative ap- proach to environmental plant physiology, 2nd edn. Cam- bridge University Press, Cambridge
Kobayashi HK (1973) Ecology of the silversword Argyroxiphium sandwicense DC. (Compositae), Haleakala Crater, Hawaii. Ph.D. Diss. Univ. of Hawaii, Honolulu
Larcher W (1975) Pflanzenokologische Beobactungen in der Para- mostufe der venezolanischen anden. Anz Oesterr Akad Wiss Math Naturwiss KL 11:194-213
Leopold, AC, Kriedemann PE (1975) Plant growth and develop- ment. McGraw-Hill, New York
Lindquist S (1987) The heat shock response. Annu Rev Biochem 55: 1151-1191
Loope LL, Crivellone CF (1986) Status of the silversword in Ha- leakala National Park: past and present. Coop Nati Park Stud- ies Unit, University of Hawaii/Manoa, Dept. Botany, Tech
Rept 58 Meinzer FC, Goldstein G (1985) Leaf pubescence and some of its
consequences in an Andean giant rosette plant. Ecology 66:512-520
Meinzer FC, Goldstein G (1986) Adaptation for water and thermal balance in Andean giant rosette plants. In: Givnish TJ (ed) On the economy of plant form and function. Cambridge Universi-
ty Press, Cambridge, pp 381-411 Meinzer FC, Goldstein G, Rundel PW (1985) Morphological
changes along an altitude gradient and their consequences for an andean giant rosette plant. Oecologia 65:278-283
Nobel PS (1988) Environmental biology of agaves and cacti. Cam- bridge University Press, New York
Nobel PS (1991) Physicochemical and environmental plant physi- ology. Academic Press, San Diego
Powell E (1992) Life history, Reproductive biology, and conserva- tion of the Mauna Kea silversword, Argyroxiphium sand- wicense DC (Asteraceae), and endangered plant of Hawaii. Ph.D. Diss. Univ. of Hawaii, Honolulu
Rock JF (1974) The Indigenous Trees of the Hawaiian Islands. National Tropical Botanical Garden, Kaui, Hawaii
Robichaux RH, Carr GD, Liebman M, Pearcy RW(1990) Adaptive radiation of the Hawaiian silverwsord alliance (Compositae- Madiinae): ecological, morphological, and physiological di-
versity. Ann Missouri Bot Gard 77:64-72 Seemann JR, Downton WJS, Berry JA (1986) Temperature and
leaf osmotic potential as factors in the acclimation of photo- synthesis to high temperature in desert plants, Plant Physiol 80:926-930
Squeo FA, Rada F, Azocar A, Goldstein G (1991) Freezing toler- ance and avoidance in high tropical Andean plants: Is it equal- ly represented in species with different plant height? Oecologia 86:378-382
Wagner WL, Herbst DR, Sohmer SH (1990) Manual of the flower- ing plants of Hawaii. (Bishop Museum Special Publication 83). Univ. of Hawaii Press, Bishop Museum Press, Honolulu
This content downloaded from 147.129.130.129 on Wed, 9 Oct 2013 17:43:11 PMAll use subject to JSTOR Terms and Conditions




















