
Demographic consequences of anthropogenic influences: Florida black bears in north-central Florida
8
Demographic consequences of anthropogenic influences: Florida black bears in north-central Florida Jeffrey A. Hostetler a, * , J. Walter McCown b , Elina P. Garrison b , Aletris M. Neils a , Mark A. Barrett b , Melvin E. Sunquist a , Stephanie L. Simek c , Madan K. Oli a a Department of Wildlife Ecology and Conservation, University of Florida, P.O. Box 110430, Gainesville, FL 32611, USA b Florida Fish and Wildlife Conservation Commission, 4005 South Main Street, Gainesville, FL 32601, USA c Florida Fish and Wildlife Conservation Commission, 620 S. Meridian St., Tallahassee, FL 32399, USA article info Article history: Received 3 July 2008 Received in revised form 29 April 2009 Accepted 20 May 2009 Available online 12 June 2009 Keywords: Anthropogenic influences on wildlife Demographic effects of habitat fragmentation Matrix population model Population growth rate Population dynamics Removal modeling Ursus americanus floridanus abstract Anthropogenic habitat fragmentation poses a serious threat to conservation of large carnivores, due to their extensive movements and potential conflicts with humans. We studied the population ecology of Florida black bears (Ursus americanus floridanus) for 6 years in two study areas in north-central Florida: Ocala National Forest (ONF), a contiguous forested habitat, and an adjacent residential community of Lynne, a fragmented habitat with substantial human activities. We estimated age-specific survival and fecundity rates of bears using data from radio-collared bears, and parameterized and analyzed stage- structured matrix population models for the two study sites and also for data pooled from both sites. Annual survival rates of adult females were lower in Lynne (0.776 ± 0.074) than in ONF (0.966 ± 0.023). While cub survival rates were higher in Lynne (0.507 ± 0.135) than in ONF (0.282 ± 0.109), the rates at both sites were substantially lower than those reported for other black bear populations. Age-specific fecundities did not vary between sites. The asymptotic population growth rate for ONF was greater than one, whereas that for Lynne was less than one. Our results suggest that anthro- pogenic influences (primarily road density and vehicular traffic, through their effect on adult survival) can substantially affect the population dynamics of Florida black bears and other large carnivores with large home ranges. We recommend efforts such as constructing highway underpasses, which could reduce road-related mortalities, to ensure long-term persistence of Florida black bears facing threats from rapidly increasing human influences. Ó 2009 Elsevier Ltd. All rights reserved. 1. Introduction The loss and fragmentation of habitat due to human activities is one of the most important factors contributing to loss of biodiver- sity throughout the world (Diamond, 1989; Lawton and May, 1995; Laurance and Bierregaard, 1997; Young and Clarke, 2000; Fahrig, 2001). Consequently, many species of conservation concern currently occur in fragmented habitats (Young and Clarke, 2000). Habitat fragmentation can disrupt social structure and dispersal patterns, increase mortality, and reduce genetic diversity, repro- ductive rates, and persistence probabilities of wildlife populations (Goosem, 1997; Jules, 1998; Harrison and Bruna, 1999; Srikwan and Woodruff, 2000; Cale, 2002). Anthropogenic influences on habitat can include loss, degrada- tion, subdivision, and isolation of habitat; increased edge effects; and other direct and indirect effects of human presence; each of these can affect dynamics and persistence of populations (Lindenmayer and Fischer, 2006). Among various anthropogenic activities, roads can have especially dramatic effects on wildlife, including direct mortalities, increased air, water, and noise pollu- tion, indirect increase in mortality (e.g., through increased access for hunters), and barriers to or conduits of movement (Trombulak and Frissell, 2000; Forman et al., 2003; Coffin, 2007). Florida is home to 109 federally listed endangered and threa- tened plant and animal species (USFWS, 2007), many of which oc- cur in fragmented habitats. Florida’s human population is projected to grow by 79% between 2000 and 2030 (US Census Bu- reau, 2005); this will undoubtedly lead to further loss and frag- mentation of remaining wildlife habitat. Understanding how habitat fragmentation influences demography and dynamics of wildlife populations is of paramount importance, because such an understanding will help guide conservation efforts. The Florida black bear (Ursus americanus floridanus) was histor- ically distributed throughout Florida and southern Georgia, Ala- bama, and Mississippi (Brady and Maehr, 1985). Large scale logging and unregulated hunting greatly reduced their numbers in Florida by the mid-1900s (McDaniel, 1974). Currently, bears 0006-3207/$ - see front matter Ó 2009 Elsevier Ltd. All rights reserved. doi:10.1016/j.biocon.2009.05.029 * Corresponding author. Tel.: +1 352 846 0648; fax: +1 352 392 6984. E-mail addresses: hostetle@ufl.edu, [email protected] (J.A. Hostetler). Biological Conservation 142 (2009) 2456–2463 Contents lists available at ScienceDirect Biological Conservation journal homepage: www.elsevier.com/locate/biocon
-
Upload
conservationcatalyst -
Category
Documents
-
view
0 -
download
0
Transcript of Demographic consequences of anthropogenic influences: Florida black bears in north-central Florida

Biological Conservation 142 (2009) 2456–2463
Contents lists available at ScienceDirect
Biological Conservation
journal homepage: www.elsevier .com/locate /b iocon
Demographic consequences of anthropogenic influences: Florida black bearsin north-central Florida
Jeffrey A. Hostetler a,*, J. Walter McCown b, Elina P. Garrison b, Aletris M. Neils a, Mark A. Barrett b,Melvin E. Sunquist a, Stephanie L. Simek c, Madan K. Oli a
a Department of Wildlife Ecology and Conservation, University of Florida, P.O. Box 110430, Gainesville, FL 32611, USAb Florida Fish and Wildlife Conservation Commission, 4005 South Main Street, Gainesville, FL 32601, USAc Florida Fish and Wildlife Conservation Commission, 620 S. Meridian St., Tallahassee, FL 32399, USA
a r t i c l e i n f o a b s t r a c t
Article history:Received 3 July 2008Received in revised form 29 April 2009Accepted 20 May 2009Available online 12 June 2009
Keywords:Anthropogenic influences on wildlifeDemographic effects of habitatfragmentationMatrix population modelPopulation growth ratePopulation dynamicsRemoval modelingUrsus americanus floridanus
0006-3207/$ - see front matter � 2009 Elsevier Ltd. Adoi:10.1016/j.biocon.2009.05.029
* Corresponding author. Tel.: +1 352 846 0648; faxE-mail addresses: [email protected], jhostetler@anim
Anthropogenic habitat fragmentation poses a serious threat to conservation of large carnivores, due totheir extensive movements and potential conflicts with humans. We studied the population ecology ofFlorida black bears (Ursus americanus floridanus) for 6 years in two study areas in north-central Florida:Ocala National Forest (ONF), a contiguous forested habitat, and an adjacent residential community ofLynne, a fragmented habitat with substantial human activities. We estimated age-specific survival andfecundity rates of bears using data from radio-collared bears, and parameterized and analyzed stage-structured matrix population models for the two study sites and also for data pooled from both sites.Annual survival rates of adult females were lower in Lynne (0.776 ± 0.074) than in ONF(0.966 ± 0.023). While cub survival rates were higher in Lynne (0.507 ± 0.135) than in ONF(0.282 ± 0.109), the rates at both sites were substantially lower than those reported for other black bearpopulations. Age-specific fecundities did not vary between sites. The asymptotic population growth ratefor ONF was greater than one, whereas that for Lynne was less than one. Our results suggest that anthro-pogenic influences (primarily road density and vehicular traffic, through their effect on adult survival)can substantially affect the population dynamics of Florida black bears and other large carnivores withlarge home ranges. We recommend efforts such as constructing highway underpasses, which couldreduce road-related mortalities, to ensure long-term persistence of Florida black bears facing threatsfrom rapidly increasing human influences.
� 2009 Elsevier Ltd. All rights reserved.
1. Introduction
The loss and fragmentation of habitat due to human activities isone of the most important factors contributing to loss of biodiver-sity throughout the world (Diamond, 1989; Lawton and May,1995; Laurance and Bierregaard, 1997; Young and Clarke, 2000;Fahrig, 2001). Consequently, many species of conservation concerncurrently occur in fragmented habitats (Young and Clarke, 2000).Habitat fragmentation can disrupt social structure and dispersalpatterns, increase mortality, and reduce genetic diversity, repro-ductive rates, and persistence probabilities of wildlife populations(Goosem, 1997; Jules, 1998; Harrison and Bruna, 1999; Srikwanand Woodruff, 2000; Cale, 2002).
Anthropogenic influences on habitat can include loss, degrada-tion, subdivision, and isolation of habitat; increased edge effects;and other direct and indirect effects of human presence; each ofthese can affect dynamics and persistence of populations
ll rights reserved.
: +1 352 392 6984.ail.net (J.A. Hostetler).
(Lindenmayer and Fischer, 2006). Among various anthropogenicactivities, roads can have especially dramatic effects on wildlife,including direct mortalities, increased air, water, and noise pollu-tion, indirect increase in mortality (e.g., through increased accessfor hunters), and barriers to or conduits of movement (Trombulakand Frissell, 2000; Forman et al., 2003; Coffin, 2007).
Florida is home to 109 federally listed endangered and threa-tened plant and animal species (USFWS, 2007), many of which oc-cur in fragmented habitats. Florida’s human population isprojected to grow by 79% between 2000 and 2030 (US Census Bu-reau, 2005); this will undoubtedly lead to further loss and frag-mentation of remaining wildlife habitat. Understanding howhabitat fragmentation influences demography and dynamics ofwildlife populations is of paramount importance, because suchan understanding will help guide conservation efforts.
The Florida black bear (Ursus americanus floridanus) was histor-ically distributed throughout Florida and southern Georgia, Ala-bama, and Mississippi (Brady and Maehr, 1985). Large scalelogging and unregulated hunting greatly reduced their numbersin Florida by the mid-1900s (McDaniel, 1974). Currently, bears

J.A. Hostetler et al. / Biological Conservation 142 (2009) 2456–2463 2457
occupy 17% of their historic range (Wooding et al., 1994) and arelisted as threatened in Florida, occurring in several fragmentedand geographically isolated populations with limited gene flow be-tween them (Dixon et al., 2006, 2007).
The Ocala National Forest (ONF) harbors one of the largest pop-ulations of black bears in Florida (Simek et al., 2005; Dixon et al.,2007). This population, however, also experiences strong anthro-pogenic influences, such as vehicular mortality and illegal harvest.The human population in the four counties surrounding ONF (Mar-ion, Lake, Putnam, and Volusia counties) is expected to grow by75% between 2000 and 2030 (Smith and Rayer, 2005). This increasein human population is likely to result in an increase in traffic andtraffic-related bear fatalities, which is already substantial(McCown et al., 2009).
Accordingly, an understanding of demographic and populationdynamic consequences of anthropogenic effects, and population-level influences of anticipated increases in traffic-related bearfatalities, would help guide management of Florida black bear pop-ulations in ONF and other increasingly human-dominated land-scapes. Our overall goal was to understand demographicconsequences of anthropogenic influences using Florida blackbears as a model system. Specifically, we parameterized and ana-lyzed matrix population models for the Florida black bears inhab-iting a contiguous forested habitat and an adjacent residentialcommunity. We estimated and compared population growth rates,examined the proportional sensitivity (i.e., elasticity) of growthrates to vital demographic parameters, and simulated the effectsof increased mortality (perhaps due to an increase in traffic-relatedmortalities) on Florida black bear population dynamics. Wehypothesized that human activity would have no effect on fecun-dity rates, but that the bears inhabiting the more fragmented hab-itat would have higher mortality rates, and thus lower populationgrowth rates, than those in the less fragmented habitat. Thesehypotheses stemmed from previous studies showing substantialdemographic effects of habitat fragmentation and other anthropo-genic influences, including reduced survival and populationgrowth for a variety of vertebrate species (e.g., Kerley et al.,2002; Kramer-Schadt et al., 2004; Letcher et al., 2007; Ruiz-Gutiérrez et al., 2008).
Fig. 1. Map of Ocala National Forest (dark gray) in north-central Florida. The two studyclasses within the study areas includes private lands (white), water bodies (light gray),
2. Methods
2.1. Study area
This study was conducted in ONF and the adjacent unincorpo-rated community of Lynne, both located in north-central Florida(Fig. 1). The primary study area consisted of approximately468 km2 of nearly contiguous bear habitat at the center of ONF, bi-sected by State Road 40 (SR-40), a two-lane paved road with anapproximate traffic rate of 5100 vehicle trips per day (Fig. 1; Flor-ida Department of Transportation, 2003). The primary study areawas approximately 60% sand pine (Pinus clausa)/scrub oak (Quercusspp.), 22% slash pine (P. elliotti) flatwoods, 10% wetland hardwoods,and 5% prairies (Aydelott et al., 1975; Moyer et al., 2008). The for-est is managed for timber, and a variety of recreational activities(camping, hiking, canoeing, swimming, off-road vehicles use,horseback riding, hunting, and fishing). Ages of stands ranged fromrecent clear cuts (0–2 years) to P25 years. Bear harvest has notbeen permitted at ONF since 1971.
The secondary study area consisted of approximately 211 km2
in the small residential community of Lynne, which straddles SR-40 3.75 km west of the primary study area. Bear habitat in Lynneconsists of small parcels of slash pine flatwoods under ownershipof ONF, corporations, and individuals. Bear habitat in Lynne is dis-continuous and dispersed in a network of moderate volume roadsand residential, industrial, and commercial development.
The study sites were close, and occasional movements of bearsbetween the two sites did occur; thus, we make no claim that theseareas represent genetically distinct populations. However, basedon our long-term radio-tracking data, we are confident that all fe-males maintained their home ranges within the study area inwhich they were captured.
2.2. Quantification of habitat fragmentation and anthropogenicinfluences
Using a Geographic Information System (ESRI ArcMap 9.2), weobtained land cover data for the two study areas from a statewide(30 m resolution) digital layer for Florida (Kautz et al., 2007). Based
areas are ONF (eastward polygon) and Lynne (westward polygon). Other land coverand roads (black lines).

2458 J.A. Hostetler et al. / Biological Conservation 142 (2009) 2456–2463
on findings of Moyer et al. (2008) and our long-term radio-telem-etry data, we reclassified 24 land cover types into two categoriesusing ArcGIS Spatial Analyst extension (ESRI): suitable habitat(consisting of xeric oak scrub, sand pine scrub, sandhill, dry prairie,mixed pine-hardwood forest, hardwood hammocks and forest,pinelands, freshwater marsh and wet prairie, shrub swamp, bayswamp, cypress swamp, mixed wetland forest, hardwood swamp,shrub and brushland, bare soil/clearcut, and low impact urban),and unsuitable habitat (consisting of high impact urban, improvedpasture, unimproved pasture, citrus, row/field crops, other agricul-ture, and grassland). Gilbert and Stys (2004) provide detaileddescriptions of land cover types.
We used four metrics generated by FRAGSTATS 3.3 (McGari-gal et al., 2002) to explore landscape fragmentation within thetwo study areas (contagion (CONTAG), percent land cover(PLAND), patch density (PD), and mean patch area (AREA_MN);see Table 1 for description of fragmentation metrics). Classesconsisted of the categories suitable habitat and unsuitable habi-tat (with open water set as background) and patches were de-fined using the eight neighbor rule (McGarigal et al., 2002).Finally, we quantified anthropogenic influences using three met-rics: (1) the density (km/km2) of major paved roads calculated aslength of roads (km) within each study area divided by total size(km2) of the study area (Florida Department of Transportation,2008); (2) traffic volume (no. of trips/day) along SR-40 withineach study area (Florida Department of Transportation, 2003);and (3) resident human population within each study area.Although the road layer consisted of 2008 data, the same majorroads (i.e., high impact urban) were present in the 2003 landcover data.
2.3. Telemetry of adult bears
We captured adult bears by modifying standard techniques de-scribed by Johnson and Pelton (1980) and by the use of culverttraps. We trapped bears from mid-May to mid-December in1999–2002 and from June to August in 2005 and 2006. Bears wereaged using an extracted premolar tooth (Willey, 1974). All adult fe-males and some adult males captured were fitted with radio collarsequipped with mortality beacons that were activated if the collarwas motionless for more than 2 h. Capture and telemetry tech-niques are described in more detail elsewhere (Garrison et al.,2007; McCown et al., 2009).
2.4. Estimation of adult survival
We tracked radio-collared adult females 1–3 times per week,using ground and aerial telemetry, from May 1999 to May 2003and June 2005 to August 2007. We documented mortalities byground checking all mortality signals. Whenever possible, we nec-ropsied bears to identify the cause of death. We used the Cox pro-portional hazard model (with staggered entry) for estimating
Table 1Quantification of area and configuration of land cover within the two study sites, Lynne andwhereas the next three variables describe suitable habitat only. FRAGSTATSa metrics are t
Study area Landscape area (km2) Road density (km/km2) Traffic volume (trips/
ONF 467.7 0.12 5100Lynne 210.8 0.30 14,700–16,700
a CONTAG (contagion) = aggregation and interspersion of class types in the landscapelandscape area; PD (patch density) = the number of patches divided by landscape area;patches.
b Traffic volume is for SR-40 only, from 2003 (Florida Department of Transportation,c The resident population for Lynne (Moyer et al., 2007) is for the town as a whole in
survival rates of collared bears, and for testing the effects of siteand year on survival (Therneau and Grambsch, 2000).
2.5. Reproductive parameters
We visited natal dens of radio-collared females from March toApril in 2000–2003, 2006, and 2007 to count and mark cubs. Weobtained the number of cubs from den visits and observationsmade during capture or tracking of adult females. Field methodswere described in detail by Garrison et al. (2007). We estimatedage-specific fecundity rates as the average number of daughters(estimated as one-half times the litter size) produced per radio-col-lared female (including non-breeders) of a given age per year. Thus,some females were included in more than one age class if they pro-duced >1 litter during the study. Females that were captured insummer with no indication of having produced cubs that year(no lactation or evidence of cubs with her, or she produced a litterthe following winter) were assumed not to have reproduced thatyear (Garrison et al., 2007). Age of first reproduction in our studypopulation was as early as 2 years, which is earlier than those re-ported from other populations (Garrison et al., 2007). We esti-mated the variance of the age-specific fecundity rates andcompared fecundity rates between study sites using the bootstrapmethod (Efron and Tibshirani, 1994).
2.6. Cub survival
During den visits in 2002, 2003, 2006, and 2007 we fitted cubsweighing more than 1.5 kg with lightweight radio-collars equippedwith mortality switches. If a cub did not meet the minimum bodyweight criterion for radio-collaring, its den was revisited and thecub was collared at a later date. The collars included a slidingmechanism designed to expand with cub growth (Vashon et al.,2003). The collar weight was <5% of cub body-mass.
After the bears emerged from their dens, we located radio-col-lared females and cubs 4–7 times per week for the first threemonths (April–June), and at least twice per week thereafter. Thisschedule provided intensive monitoring during the period withhighest expected mortality (Elowe, 1987; LeCount, 1987; Garrisonet al., 2007). We approached cubs immediately if a collar transmit-ted a mortality signal or if a cub was located away from themother. If the exact date of a cub’s death was unknown, we usedthe midpoint of the interval between the date the cub was lastknown to be alive and the date the mortality signal was first de-tected. For more details on cub capture and radio-tracking, see Gar-rison et al. (2007).
No cubs retained their collars for a full year. We right-cen-sored cubs that lost their collars or had radio failures. Most cubsdied or were censored in the first six months, and the last cubwas censored 11 months after the first cub was radio-tagged thatyear. We used parametric survival analyses (Klein and Moesch-berger, 2003) to identify appropriate survival functions and
ONF. The first five variables starting from the left correspond to the entire landscape,hose represented by all capital letters.
day)b Resident human populationc CONTAG Suitable habitat
PLAND PD AREA_MN
0 92.0 89.3 0.15 5.83760 64.4 87.3 1.33 0.7
(ranges from 0% to 100%); PLAND (% land cover) = sum of class area divided by totalAREA_MN (mean patch area (km2)) = sum of class area divided by the number of
2003).stead of the study area.

J.A. Hostetler et al. / Biological Conservation 142 (2009) 2456–2463 2459
project survival to a full year. This approach allowed us to esti-mate annual survival rates with the available data that spannedless than a year. We used the date each cub was captured asthe beginning of the study (time 0) for that cub; this approachis commonly used in medical studies and also was appropriatefor our study (Klein and Moeschberger, 2003). Unlike non-para-metric (e.g., Kaplan–Meier) and semi-parametric (e.g., Cox pro-portional-hazard) methods, parametric survival analysespresuppose a functional form to the hazard distribution. Becausewe had no strong a priori beliefs about the changes in hazard overthe first year of life for bear cubs, we tested several commonlyused survival distributions: exponential, Weibull, generalizedgamma, lognormal, and log-logistic (Allison, 1995). We usedAkaike’s Information Criterion (AIC; Burnham and Anderson,2002) to compare survival distributions, each with and withoutstudy site as a covariate. We model-averaged the annual survivalestimates from the models that converged successfully (Burnhamand Anderson, 2002). The model averaged estimates for eachstudy site were averaged over all models (models without a studysite effect were included as having the same survival for bothsites), whereas the overall model average estimated only includesmodels with no study site effect.
2.7. Construction and analysis of population models
We developed stage-structured matrix population models(Caswell, 2001) based on a post-breeding census for females inONF, Lynne, and the combined population (i.e., data pooled fromboth sites). We used a one year projection interval and fivestages, the first four of which represented age classes 1–4 andthe last of which represented bears P4 year of age. We usedthe estimate of annual survival rate obtained from the parametricsurvival analysis for the survival of the cubs (age class 1). Thesurvival of the adult age classes (2-years old and older) were esti-mated from the analysis of radio-telemetry data as described pre-viously. When data from ONF and Lynne were pooled and treatedas one population, we had sufficient data for separate survivalestimates for 2-year old and P3-year old bears. When each sitewas considered separately, however, data were insufficient forestimation of survival for two adult age classes; thus, we used asingle estimate of survival for females aged 2 and older. We onlywere able to radio-collar two yearling females in this study.Therefore, for the survival of second age class (1–2 years) we useda range of values between the annual survival rates of cubs andadult bears. We estimated the fertility rate for each age classusing the post-breeding census formulation (Caswell, 2001) asthe product of the survival and fecundity rates of that age class.To estimate the variances of the fertilities, we used Goodman’s(1960) exact method for the variance of products of randomvariables.
Using these population projection matrices, we calculated theasymptotic population growth rates (k), and elasticity of k to ma-trix entry and lower-level parameters (Supplementary material)using methods described by Caswell (2001). We estimated the var-iance of k using the delta method (assuming no covariation amongvital demographic rates), and constructed approximate confidenceintervals on k assuming normal distribution (Caswell, 2001).
2.8. Simulation of increased mortality
To examine the potential effects of increased mortality in thefuture (e.g., due to increased traffic fatalities, removal of nuisancebears, harvest, or other causes) on population dynamics and toevaluate a sustainable level of increased mortality, we added a re-moval term to all matrix models with k > 1 (removal in a popula-tion with k 6 1 could not be sustainable) as follows:
A¼
0 P2ð1� hÞm2 P3ð1� hÞm3 P4ð1� hÞm4 P5þð1� hÞm5þ
P1 0 0 0 00 P2ð1� hÞ 0 0 00 0 P3ð1� hÞ 0 00 0 0 P4ð1� hÞ P5þð1� hÞ
26666664
37777775
where A is the population projection matrix, Pi is the survival rate ofage class i, mi is the fecundity of age class i, and h is the removal rate(i.e., proportion of adults and yearlings removed each year). Incor-porating removal into fertility (survival times fecundity) terms isappropriate when removal precedes reproduction (Caswell, 2001,pp. 640–641), as is the case for any post-breeding census model, be-cause adults may be removed before they can reproduce.
We simulated a range of removal levels between 0 and 0.15. Be-cause yearling survival was also unknown, we varied removal andyearling survival rates simultaneously and examined the effects onthe population growth rate and lower level elasticities. Specifically,we looked at what combinations of removal and yearling survivalwould lead to k = 1.
3. Results
3.1. Fragmentation and anthropogenic influences
Although the two study areas contained similar proportions ofsuitable habitat, fragmentation metrics revealed a more frag-mented landscape with patchily distributed habitat in Lynne com-pared to ONF (Table 1). The mean patch size was much smaller inLynne, and 94% (N = 281) of patches were 60.01 km2; in ONF, only83% (N = 72) of patches were 60.01 km2. Furthermore, contagionwas lower in Lynne than ONF, indicating habitat classes in Lynnewere less aggregated and more dispersed. The density of pavedroads and traffic volume in the Lynne study area were each morethan double that in ONF study area. Finally, Lynne contained a res-ident human population of 3760, whereas there was no residenthuman population in ONF (Supplementary Table 1).
3.2. Demographic variables
We placed radio collars on 39 adult females in ONF and 18 adultfemales in Lynne, and monitored them for a total of 58,382 radio-days. We documented two mortalities of radio-collared females inONF (from intraspecific aggression) and seven among radio-col-lared females in Lynne (from vehicular accident and illegal hunt-ing) (Appendix B).
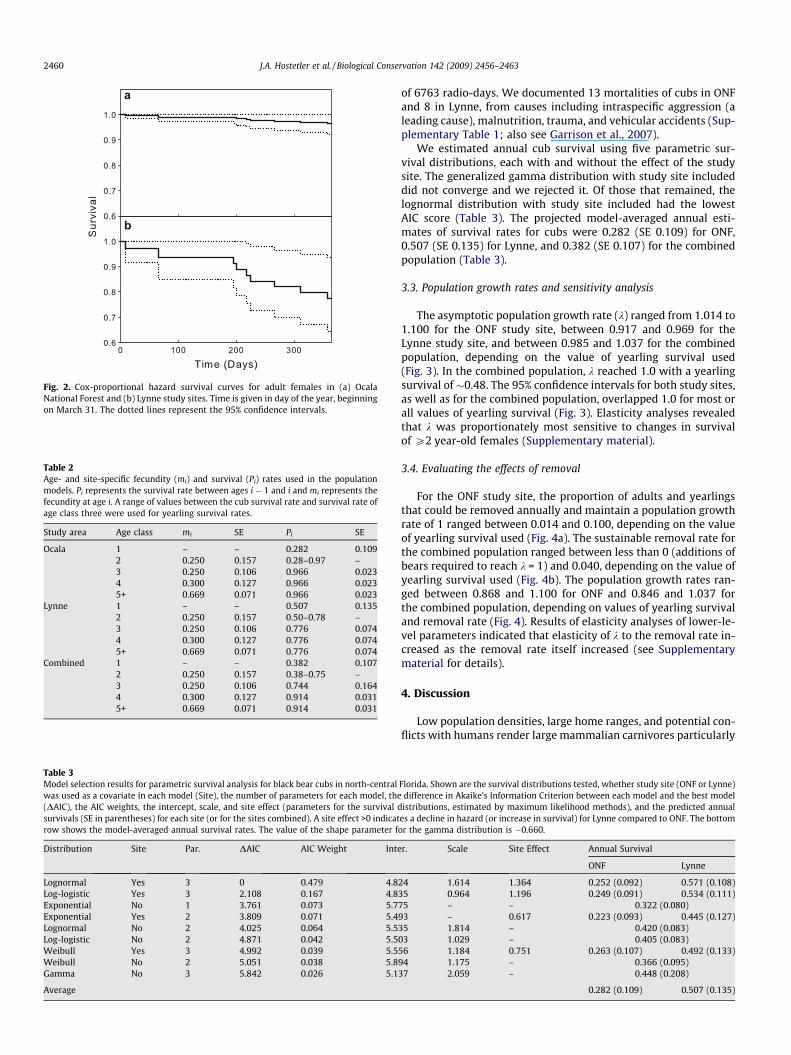
The Cox proportional-hazard analysis indicated that the sur-vival curves for Lynne and ONF females differed significantly(Fig. 2), but there was no evidence for temporal variation in sur-vival in either site. The annual survival rate for radio-collared adultfemales was 0.966 (95% CI = 0.921�1) in ONF and 0.776 (95%CI = 0.644�0.936) in Lynne (Table 2). The annual survival rate esti-mated for the combined population (i.e., data pooled from bothsites) was 0.744 (95% CI = 0.484�1.0) for 2-year old females and0.914 (95% CI = 0.855�0.977) for older females (Table 3).
Reproductive data were collected from 47 radio-tagged adultfemales of known age that were monitored from 2000 to 2003,2006, and 2007, for a total of 94 bear-reproductive seasons. Overthat period, they produced 105 cubs. The age-specific fecundityrates ranged from 0.25 for 2 and 3-year olds to 0.669 for females5-years and older (Table 2). Fecundity rates for the two sites weresimilar (overall fecundity at ONF = 0.565 and at Lynne = 0.547;p = 0.444; differences between sites by age class were also non-significant).
We placed radio collars on 26 cubs in ONF and 24 cubs in Lynnein 2002, 2003, 2006 and 2007. We monitored these cubs for a total
a
b
Fig. 2. Cox-proportional hazard survival curves for adult females in (a) OcalaNational Forest and (b) Lynne study sites. Time is given in day of the year, beginningon March 31. The dotted lines represent the 95% confidence intervals.
Table 2Age- and site-specific fecundity (mi) and survival (Pi) rates used in the populationmodels. Pi represents the survival rate between ages i � 1 and i and mi represents thefecundity at age i. A range of values between the cub survival rate and survival rate ofage class three were used for yearling survival rates.
Study area Age class mi SE Pi SE
Ocala 1 – – 0.282 0.1092 0.250 0.157 0.28–0.97 –3 0.250 0.106 0.966 0.0234 0.300 0.127 0.966 0.0235+ 0.669 0.071 0.966 0.023
Lynne 1 – – 0.507 0.1352 0.250 0.157 0.50–0.78 –3 0.250 0.106 0.776 0.0744 0.300 0.127 0.776 0.0745+ 0.669 0.071 0.776 0.074
Combined 1 – – 0.382 0.1072 0.250 0.157 0.38–0.75 –3 0.250 0.106 0.744 0.1644 0.300 0.127 0.914 0.0315+ 0.669 0.071 0.914 0.031
Table 3Model selection results for parametric survival analysis for black bear cubs in north-centralwas used as a covariate in each model (Site), the number of parameters for each model, the(DAIC), the AIC weights, the intercept, scale, and site effect (parameters for the survival dsurvivals (SE in parentheses) for each site (or for the sites combined). A site effect >0 indicatrow shows the model-averaged annual survival rates. The value of the shape parameter f
Distribution Site Par. DAIC AIC Weight Inte
Lognormal Yes 3 0 0.479 4.8Log-logistic Yes 3 2.108 0.167 4.8Exponential No 1 3.761 0.073 5.7Exponential Yes 2 3.809 0.071 5.4Lognormal No 2 4.025 0.064 5.5Log-logistic No 2 4.871 0.042 5.5Weibull Yes 3 4.992 0.039 5.5Weibull No 2 5.051 0.038 5.8Gamma No 3 5.842 0.026 5.1
Average
2460 J.A. Hostetler et al. / Biological Conservation 142 (2009) 2456–2463
of 6763 radio-days. We documented 13 mortalities of cubs in ONFand 8 in Lynne, from causes including intraspecific aggression (aleading cause), malnutrition, trauma, and vehicular accidents (Sup-plementary Table 1; also see Garrison et al., 2007).
We estimated annual cub survival using five parametric sur-vival distributions, each with and without the effect of the studysite. The generalized gamma distribution with study site includeddid not converge and we rejected it. Of those that remained, thelognormal distribution with study site included had the lowestAIC score (Table 3). The projected model-averaged annual esti-mates of survival rates for cubs were 0.282 (SE 0.109) for ONF,0.507 (SE 0.135) for Lynne, and 0.382 (SE 0.107) for the combinedpopulation (Table 3).
3.3. Population growth rates and sensitivity analysis
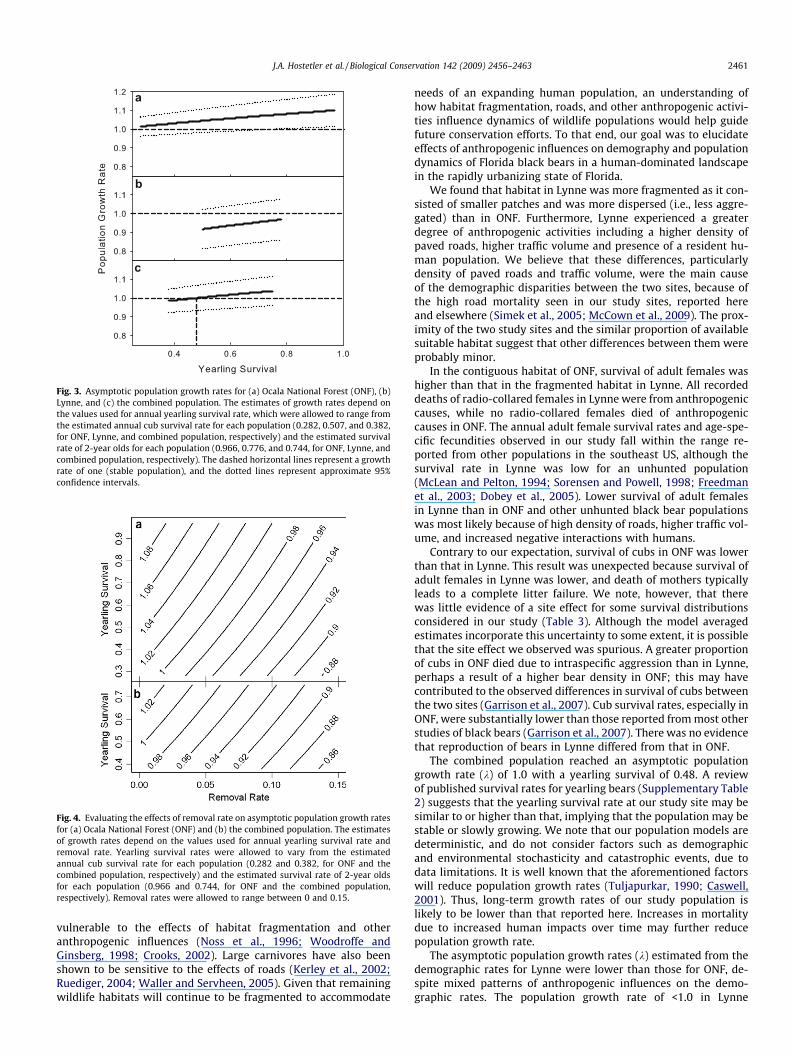
The asymptotic population growth rate (k) ranged from 1.014 to1.100 for the ONF study site, between 0.917 and 0.969 for theLynne study site, and between 0.985 and 1.037 for the combinedpopulation, depending on the value of yearling survival used(Fig. 3). In the combined population, k reached 1.0 with a yearlingsurvival of �0.48. The 95% confidence intervals for both study sites,as well as for the combined population, overlapped 1.0 for most orall values of yearling survival (Fig. 3). Elasticity analyses revealedthat k was proportionately most sensitive to changes in survivalof P2 year-old females (Supplementary material).
3.4. Evaluating the effects of removal
For the ONF study site, the proportion of adults and yearlingsthat could be removed annually and maintain a population growthrate of 1 ranged between 0.014 and 0.100, depending on the valueof yearling survival used (Fig. 4a). The sustainable removal rate forthe combined population ranged between less than 0 (additions ofbears required to reach k = 1) and 0.040, depending on the value ofyearling survival used (Fig. 4b). The population growth rates ran-ged between 0.868 and 1.100 for ONF and 0.846 and 1.037 forthe combined population, depending on values of yearling survivaland removal rate (Fig. 4). Results of elasticity analyses of lower-le-vel parameters indicated that elasticity of k to the removal rate in-creased as the removal rate itself increased (see Supplementarymaterial for details).
4. Discussion
Low population densities, large home ranges, and potential con-flicts with humans render large mammalian carnivores particularly
Florida. Shown are the survival distributions tested, whether study site (ONF or Lynne)difference in Akaike’s Information Criterion between each model and the best modelistributions, estimated by maximum likelihood methods), and the predicted annual
es a decline in hazard (or increase in survival) for Lynne compared to ONF. The bottomor the gamma distribution is �0.660.
r. Scale Site Effect Annual Survival
ONF Lynne
24 1.614 1.364 0.252 (0.092) 0.571 (0.108)35 0.964 1.196 0.249 (0.091) 0.534 (0.111)75 – – 0.322 (0.080)93 – 0.617 0.223 (0.093) 0.445 (0.127)35 1.814 – 0.420 (0.083)03 1.029 – 0.405 (0.083)56 1.184 0.751 0.263 (0.107) 0.492 (0.133)94 1.175 – 0.366 (0.095)37 2.059 – 0.448 (0.208)
0.282 (0.109) 0.507 (0.135)
a
b
c
Fig. 3. Asymptotic population growth rates for (a) Ocala National Forest (ONF), (b)Lynne, and (c) the combined population. The estimates of growth rates depend onthe values used for annual yearling survival rate, which were allowed to range fromthe estimated annual cub survival rate for each population (0.282, 0.507, and 0.382,for ONF, Lynne, and combined population, respectively) and the estimated survivalrate of 2-year olds for each population (0.966, 0.776, and 0.744, for ONF, Lynne, andcombined population, respectively). The dashed horizontal lines represent a growthrate of one (stable population), and the dotted lines represent approximate 95%confidence intervals.
Fig. 4. Evaluating the effects of removal rate on asymptotic population growth ratesfor (a) Ocala National Forest (ONF) and (b) the combined population. The estimatesof growth rates depend on the values used for annual yearling survival rate andremoval rate. Yearling survival rates were allowed to vary from the estimatedannual cub survival rate for each population (0.282 and 0.382, for ONF and thecombined population, respectively) and the estimated survival rate of 2-year oldsfor each population (0.966 and 0.744, for ONF and the combined population,respectively). Removal rates were allowed to range between 0 and 0.15.
J.A. Hostetler et al. / Biological Conservation 142 (2009) 2456–2463 2461
vulnerable to the effects of habitat fragmentation and otheranthropogenic influences (Noss et al., 1996; Woodroffe andGinsberg, 1998; Crooks, 2002). Large carnivores have also beenshown to be sensitive to the effects of roads (Kerley et al., 2002;Ruediger, 2004; Waller and Servheen, 2005). Given that remainingwildlife habitats will continue to be fragmented to accommodate
needs of an expanding human population, an understanding ofhow habitat fragmentation, roads, and other anthropogenic activi-ties influence dynamics of wildlife populations would help guidefuture conservation efforts. To that end, our goal was to elucidateeffects of anthropogenic influences on demography and populationdynamics of Florida black bears in a human-dominated landscapein the rapidly urbanizing state of Florida.
We found that habitat in Lynne was more fragmented as it con-sisted of smaller patches and was more dispersed (i.e., less aggre-gated) than in ONF. Furthermore, Lynne experienced a greaterdegree of anthropogenic activities including a higher density ofpaved roads, higher traffic volume and presence of a resident hu-man population. We believe that these differences, particularlydensity of paved roads and traffic volume, were the main causeof the demographic disparities between the two sites, because ofthe high road mortality seen in our study sites, reported hereand elsewhere (Simek et al., 2005; McCown et al., 2009). The prox-imity of the two study sites and the similar proportion of availablesuitable habitat suggest that other differences between them wereprobably minor.
In the contiguous habitat of ONF, survival of adult females washigher than that in the fragmented habitat in Lynne. All recordeddeaths of radio-collared females in Lynne were from anthropogeniccauses, while no radio-collared females died of anthropogeniccauses in ONF. The annual adult female survival rates and age-spe-cific fecundities observed in our study fall within the range re-ported from other populations in the southeast US, although thesurvival rate in Lynne was low for an unhunted population(McLean and Pelton, 1994; Sorensen and Powell, 1998; Freedmanet al., 2003; Dobey et al., 2005). Lower survival of adult femalesin Lynne than in ONF and other unhunted black bear populationswas most likely because of high density of roads, higher traffic vol-ume, and increased negative interactions with humans.
Contrary to our expectation, survival of cubs in ONF was lowerthan that in Lynne. This result was unexpected because survival ofadult females in Lynne was lower, and death of mothers typicallyleads to a complete litter failure. We note, however, that therewas little evidence of a site effect for some survival distributionsconsidered in our study (Table 3). Although the model averagedestimates incorporate this uncertainty to some extent, it is possiblethat the site effect we observed was spurious. A greater proportionof cubs in ONF died due to intraspecific aggression than in Lynne,perhaps a result of a higher bear density in ONF; this may havecontributed to the observed differences in survival of cubs betweenthe two sites (Garrison et al., 2007). Cub survival rates, especially inONF, were substantially lower than those reported from most otherstudies of black bears (Garrison et al., 2007). There was no evidencethat reproduction of bears in Lynne differed from that in ONF.
The combined population reached an asymptotic populationgrowth rate (k) of 1.0 with a yearling survival of 0.48. A reviewof published survival rates for yearling bears (Supplementary Table2) suggests that the yearling survival rate at our study site may besimilar to or higher than that, implying that the population may bestable or slowly growing. We note that our population models aredeterministic, and do not consider factors such as demographicand environmental stochasticity and catastrophic events, due todata limitations. It is well known that the aforementioned factorswill reduce population growth rates (Tuljapurkar, 1990; Caswell,2001). Thus, long-term growth rates of our study population islikely to be lower than that reported here. Increases in mortalitydue to increased human impacts over time may further reducepopulation growth rate.
The asymptotic population growth rates (k) estimated from thedemographic rates for Lynne were lower than those for ONF, de-spite mixed patterns of anthropogenic influences on the demo-graphic rates. The population growth rate of <1.0 in Lynne

2462 J.A. Hostetler et al. / Biological Conservation 142 (2009) 2456–2463
suggests that this population may be a ‘‘sink” that would not per-sist without immigrants from the ONF. We recognize that ourstudy was based on one group of bears inhabiting highly frag-mented and human-dominated habitat and another group occupy-ing contiguous and less disturbed habitat. This situation is not idealfor rigorous statistical inference, but replicated field studies oflarge carnivores that permit rigorous statistical inferences arerarely possible. Nevertheless, two important results emerge fromour study that have broad implications. First, the population as awhole could probably not sustain itself if it faced the level of adultmortality seen in Lynne. Second, in the fragmented habitat inLynne, all deaths of adult, radio-collared females were fromanthropogenic causes (collision with vehicles and poaching). Thereis little doubt about the negative effects of anthropogenic influ-ences for many species, but the mechanisms by which this can oc-cur can vary among species (Fahrig, 2003; Letcher et al., 2007;Stuchbury, 2007). Our study provides an example of demographicmechanisms by which anthropogenic activities can influence pop-ulation dynamics.
To examine the possible effects of increased mortality, whichcan arise due to an increase in traffic volume as the human popu-lation increases or due to other anthropogenic influences, we sim-ulated a range of removal rates for the ONF study site and thecombined population. Depending on the value used for yearlingsurvival, the combined population might need addition of bearsto reach a stable population, or it might be able to sustain a re-moval rate as high as 0.040 per year. These results suggest thatthe population may sustain a low level of additional mortality(e.g., increased road-related mortality or nuisance bear removal).However, these should be interpreted cautiously because stochas-tic events such as prolonged drought, mast failure and disease canreduce survival and reproductive rates (and thus, populationgrowth rate) below replacement level.
Elasticity analyses revealed that k was proportionately moresensitive to changes in survival (particularly of reproductive ageclasses) than to reproductive parameters (Supplementary mate-rial). These results are similar to those reported by Freedmanet al. (2003) for black bear populations of the Southeastern Coast-al Plain, and also are consistent with broader elasticity patternsfor long-lived mammals with relatively low reproductive rates(Heppell et al., 2000; Oli and Dobson, 2003). Elasticity analysisof the model that included the removal term showed that theabsolute value of the lower-level elasticity of k to the removalrate was generally lower than that to changes in survival orfecundity rates, but it increased as the removal rate increased.However, these results were based on asymptotic analysis of ma-trix model and should be interpreted with caution. Asymptoticpopulation growth rates and elasticities, although informativeand generally robust (Caswell 2001), may not always reflectshort-term growth rates or elasticities (Caswell 2007; Ozgulet al., 2009). Analysis of transient dynamics requires estimatesof abundance for each age class, which are not available for ourstudy population; consequently, we could not evaluate how re-sults of transient analyses might compare to those of asymptoticanalyses reported here.
Florida is experiencing a rapid population growth, and thistrend will likely continue in foreseeable future (US Census Bureau,2005). Thus, wildlife habitat will continue to be lost to commercialand residential developments, and construction or expansion ofroads will further fragment remaining habitat. In our study site,the Florida 2060 model (Zwick and Carr, 2006) predicts that the fu-ture increase in urban area by the year 2060 would result in a lossof suitable habitat within Lynne of 13.0 km2 and in ONF of 3.3 km2,as compared to the 2003 land cover layer. Thus, fragmentation willincrease as preferred habitat decreases in Lynne at a much fasterrate than may occur in ONF. In addition, the widening of SR40 from
2 to 4 lanes through the Lynne study area has been approved.These changes would likely lead to an increase in traffic volume.A series of eight large-species wildlife crossing structures isplanned for the approximately 22.8 km section of the road to mit-igate the effects of this highway on bears and other species of con-cern. We believe this is a prudent step to mitigate the potentialeffects upon bear mortality and dispersal and to help ensure demo-graphic and genetic connectivity of the population.
The future challenge for wildlife conservation in Florida andother rapidly urbanizing states or countries is one of managingwildlife populations in a fragmented, human-dominated land-scape. This requires an adequate understanding of how demo-graphic rates and population dynamics are affected by habitatfragmentation and other anthropogenic influences. In this study,we demonstrated an apparent population level effect of anthropo-genic influences, mediated through reduction in survival due pri-marily to road-related fatalities. An increase in yearling and adultmortality due to anthropogenic causes may lead this populationfrom stable or increasing to declining, especially with increasinghabitat fragmentation. We recommend efforts to maintain or in-crease survival of adult Florida black bears, such as highway under-passes, careful management of residential and commercialdevelopments, and vehicular traffic, reduced speed limits duringperiods of low visibility, and increased efforts to reduce illegal kill-ing of bears.
Acknowledgements
We gratefully acknowledge Florida Fish and Wildlife Conserva-tion Commission (FWC) for funding and logistical support. Addi-tional support was provided by the Florida Department ofTransportation, United States Forest Service, Wildlife Foundationof Florida, and Florida Agricultural Experiment Station. All proce-dures were approved by the University of Florida’s InstitutionalAnimal Care and Use Committee. We thank M. Cunningham, J. Dix-on, M. Moyer, D. Masters, B. Scheick, B. Smith, K. Isaacs, and D.Gualtieri for help in the field. FWC pilots J. Wisniesky, J. Johnston,and P. Crippen assisted with telemetry flights. We thank A. Ozgul, J.Martin, P. Kubilis, K. Pollock, and R. Fletcher for their assistancewith statistical questions and T. Eason and T. O’Meara for their sup-port during all phases of this study. K. With, R. Powell, and twoanonymous reviewers provided many insightful comments onthe manuscript, for which we are grateful.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.biocon.2009.05.029.
References
Allison, P.D., 1995. Survival Analysis Using the SAS System. SAS, Cary, NC.Aydelott, D.G., Bullock, H.C., Furman, A.L., White, H.O., Spieth, J.W., 1975. Soil Survey
of Ocala National Forest Area, Florida. US Department of Agriculture, SoilConservation Service. <soildatamart.nrcs.usda.gov/Manuscripts/FL609/0/ONF.pdf> (accessed July 2008).
Brady, J.R., Maehr, D.S., 1985. Black bear distribution in Florida. Florida FieldNaturalist 13, 1–7.
Burnham, K.P., Anderson, D.R., 2002. Model Selection and Multimodel Inference. aPractical Information-Theoretic Approach, second ed. Springer-Verlag, NewYork, NY.
Cale, P.G., 2002. The influence of social behaviour, dispersal, and landscapefragmentation on population structure in a sedentary bird. BiologicalConservation 109, 237–248.
Caswell, H., 2001. Matrix Population Models: Construction, Analysis, andInterpretation, second ed. Sinauer Associates, Sunderland, MA.
Caswell, H., 2007. Sensitivity analysis of transient population dynamics. EcologyLetters 10, 1–15.
Coffin, A.W., 2007. From roadkill to road ecology: a review of the ecological effectsof roads. Journal of Transport Geography 15, 396–406.

J.A. Hostetler et al. / Biological Conservation 142 (2009) 2456–2463 2463
Crooks, K.R., 2002. Relative sensitivities of mammalian carnivores to habitatfragmentation. Conservation Biology 16, 488–502.
Diamond, J.M., 1989. Overview of recent extinctions. In: Western, D., Pearl, M.C.(Eds.), Conservation for the Twenty-First Century. Oxford University Press, NewYork, NY, pp. 37–41.
Dixon, J.D., Oli, M.K., Wooten, M.C., Eason, T.H., McCown, J.W., Cunningham, M.W.,2007. Genetic consequences of habitat fragmentation and loss: the case of theFlorida black bear (Ursus americanus floridanus). Conservation Genetics 8, 455–464.
Dixon, J.D., Oli, M.K., Wooten, M.C., Eason, T.H., McCown, J.W., Paetkau, D., 2006.Effectiveness of a regional corridor in connecting two Florida black bearpopulations. Conservation Biology 20, 155–162.
Dobey, S., Masters, D.V., Scheick, B.K., Clark, J.D., Pelton, M.R., Sunquist, M.E., 2005.Ecology of Florida black bears in the Okefenokee-Osceola ecosystem. WildlifeMonographs 158, 1–41.
Efron, B., Tibshirani, R.J., 1994. An Introduction to the Bootstrap. Chapman and Hall,London.
Elowe, K.D., 1987. Factors affecting black bear reproductive success and cub survivalin Massachusetts. M.Sc. Thesis, University of Massachusetts, Amherst.
Fahrig, L., 2001. How much habitat is enough? Biological Conservation 100, 65–74.Fahrig, L., 2003. Effects of habitat fragmentation on biodiversity. Annual Review of
Ecology, Evolution, and Systematics 34, 487–515.Florida Department of Transportation, 2003. Florida traffic information, Tallahassee.
CD-ROM.Florida Department of Transportation, 2008. Major roads – statewide – April 2008.
FDOT. <www.dot.state.fl.us> (accessed December 2008).Forman, R.T.T., Sperling, D., Bissonette, J.A., Clevenger, A.P., Cutshall, C.D., Dale, V.H.,
Fahrig, L., France, R., Goldman, C.R., Heanue, K., Jones, J.A., Swanson, F.J.,Turrentine, T., Winter, T.C., 2003. Road Ecology: Science and Solutions. IslandPress, Washington, DC.
Freedman, A.H., Portier, K.M., Sunquist, M.E., 2003. Life history analysis for blackbears (Ursus americanus) in a changing demographic landscape. EcologicalModelling 167, 47–64.
Garrison, E.P., McCown, J.W., Oli, M.K., 2007. Reproductive ecology and cub survivalof Florida black bears. Journal of Wildlife Management 71, 720–727.
Gilbert, T., Stys, B., 2004. Descriptions of vegetation and land cover types mappedusing Landsat imagery. Florida Fish and Wildlife Conservation Commission.<http://199.250.30.194/GIS/LandCover/fwcveg_descriptions03.pdf> (accessedDecember 2008).
Goodman, L.A., 1960. On the exact variance of products. Journal of the AmericanStatistical Association 55, 708–713.
Goosem, M., 1997. Internal fragmentation: the effects of roads, highways, andpowerline clearings on movements and mortality of rainforest vertebrates. In:Laurance, W.F., Bierregaard, R.O. (Eds.), Tropical Forest Remnants. University ofChicago Press, Chicago, IL, pp. 241–255.
Harrison, S., Bruna, E., 1999. Habitat fragmentation and large-scale conservation:what do we know for sure? Ecography 22, 225–232.
Heppell, S.S., Caswell, H., Crowder, L.B., 2000. Life histories and elasticity patterns:perturbation analysis for species with minimal demographic data. Ecology 81,654–665.
Johnson, K.G., Pelton, M.R., 1980. Prebaiting and snaring techniques for black bears.Wildlife Society Bulletin 8, 46–54.
Jules, E.S., 1998. Habitat fragmentation and demographic change for a commonplant: trillium in old-growth forest. Ecology 79, 1645–1656.
Kautz, R., Stys, B., Kawula, R., 2007. Florida vegetation 2003 and land use changebetween 1985–1989 and 2003. Florida Scientist 70, 12–23.
Kerley, L.L., Goodrich, J.M., Miquelle, D.G., Smirnov, E.N., Quigley, H.B., Hornocker,M.G., 2002. Effects of roads and human disturbance on Amur tigers.Conservation Biology 16, 97–108.
Klein, J.P., Moeschberger, M.L., 2003. Survival Analysis, second ed. Springer, NewYork.
Kramer-Schadt, S., Revilla, E., Wiegand, T., Breitenmoser, U., 2004. Fragmentedlandscapes, road mortality and patch connectivity: modelling influences on thedispersal of Eurasian lynx. Journal of Applied Ecology 41, 711–723.
Laurance, W.F., Bierregaard, R.O., 1997. Tropical Forest Remnants. University ofChicago Press, Chicago, IL.
Lawton, J.H., May, R.M., 1995. Extinction Rates. Oxford University Press, Oxford.LeCount, A., 1987. Causes of black bear cub mortality. In: International Conference
on Bear Research and Management, pp. 75–82.Letcher, B.H., Nislow, K.H., Coombs, J.A., O’Donnell, M.J., Dubreuil, T.L., 2007.
Population response to habitat fragmentation in a stream-dwelling brook troutpopulation. PLoS One 2, e1139.
Lindenmayer, D.B., Fischer, J., 2006. Habitat Fragmentation and Landscape Change:An Ecological and Conservation Synthesis. Island Press, Washington, DC.
McCown, J.W., Kubilis, P., Eason, T.H., Scheick, B.K., 2009. Effects of traffic volume onAmerican black bears in central Florida, USA. Ursus 20, 39–46.
McDaniel, J., 1974. Reports on black bear management and research by state oruniversity. In: Second Eastern Workshop on Black Bear Management andResearch. Gatlinburg, TN.
McGarigal, K., Cushman, S.A., Neel, M.C., Ene, E., 2002. FRAGSTATS: Spatial PatternAnalysis Program for Categorical Maps. Computer software program producedby the authors at the University of Massachusetts, Amherst. <www.umass.edu/landeco/research/fragstats/fragstats.html> (accessed December 2008).
McLean, P.K., Pelton, M.R., 1994. Estimates of population density and growth ofblack bears in the Smoky Mountains. In: International Conference on BearResearch and Management, pp. 253–261.
Moyer, M.A., McCown, J.W., Oli, M.K., 2007. Factors influencing home-range size offemale Florida black bears. Journal of Mammalogy 88, 468–476.
Moyer, M.A., McCown, J.W., Oli, M.K., 2008. Scale-dependent habitat selection byfemale Florida black bears in Ocala National Forest, Florida. SoutheasternNaturalist 7, 111–124.
Noss, R.F., Quigley, H.B., Hornocker, M.G., Merrill, T., Paquet, P.C., 1996. Conservationbiology and carnivore conservation in the Rocky Mountains. ConservationBiology 10, 949–963.
Oli, M.K., Dobson, F.S., 2003. The relative importance of life-history variables topopulation growth rate in mammals: Cole’s prediction revisited. AmericanNaturalist 161, 422–440.
Ozgul, A., Oli, M.K., Armitage, K.B., Blumstein, D.T., Van Vuren, D.H., 2009. Influenceof local demography on asymptotic and transient dynamics of a yellow-belliedmarmot metapopulation. American Naturalist 173, 517–530.
Ruediger, B., 2004. Carnivore conservation and highways: understanding therelationships, problems, and solutions. In: Fascione, N., Delach, A., Smith, M.E.(Eds.), People and Predators: from Conflict to Coexistence. Island Press,Washington, DC, pp. 132–149.
Ruiz-Gutiérrez, V., Gavin, T.A., Dhondt, A.A., 2008. Habitat fragmentation lowerssurvival of a tropical forest bird. Ecological Applications 18, 838–846.
Simek, S.L., Scheick, B.K., Jonker, S.A., Endries, M.J., Eason, T.H., 2005. Statewideassessment of road impacts on bears in six study areas in Florida from May2001–September 2003. Florida Department of Transportation and Florida Fishand Wildlife Conservation Commission, Tallahassee. <myfwc.com/bear/Reports/RoadImpactsOnBearsStudy-Final-Report-2005.pdf> (accessed July2008).
Smith, S.K., Rayer, S., 2005. Population projections by age, sex, race, and hispanicorigin for Florida and its counties 2004–2030. Florida Population Studies 38.
Sorensen, V.A., Powell, R.A., 1998. Estimating survival rates of black bears. CanadianJournal of Zoology 76, 1335–1343.
Srikwan, S., Woodruff, D.S., 2000. Genetic erosion in isolated small-mammalpopulations following rainforest fragmentation. In: Young, A.G., Clarke, G.M.(Eds.), Genetics, Demography, and Viability of Fragmented Populations.Cambridge University Press, New York, NY, pp. 149–172.
Stuchbury, B.J.M., 2007. The effects of habitat fragmentation on animals: gaps in ourknowledge and new approaches. Canadian Journal of Zoology 85, 1015–1016.
Therneau, T.M., Grambsch, P.M., 2000. Modeling Survival Data: Extending the CoxModel. Springer, New York, NY.
Trombulak, S.C., Frissell, C.A., 2000. Review of ecological effects of roads onterrestrial and aquatic communities. Conservation Biology 14, 18–30.
Tuljapurkar, S., 1990. Population Dynamics in Variable Environments. Springer,New York, NY.
US Census Bureau, 2005. State Interim Population Projections by Age and Sex:2004–2030. <www.census.gov/population/www/projections/projectionsagesex.html> (accessed July 2008).
USFWS, 2007. USFWS threatened and endangered species system (TESS) [Internet].<ecos.fws.gov/tess_public> (accessed July 2008).
Vashon, J.H., Vaughan, M.R., Vashon, A.D., Martin, D.D., Echols, K.N., 2003. Anexpandable radiocollar for black bear cubs. Wildlife Society Bulletin 31, 380–386.
Waller, J.S., Servheen, C., 2005. Effects of transportation infrastructure on grizzlybears in northwestern Montana. Journal of Wildlife Management 69, 985–1000.
Willey, C.H., 1974. Aging black bears from first premolar tooth sections. Journal ofWildlife Management 38, 97–100.
Wooding, J.B., Cox, J.A., Pelton, M.R., 1994. Distribution of black bears in thesoutheastern coastal plain. In: Annual Conference of Southeastern Associationof Fish and Wildlife Agencies, pp. 270–275.
Woodroffe, R., Ginsberg, J.R., 1998. Edge effects and the extinction of populationsinside protected areas. Science 280, 2126–2128.
Young, A.G., Clarke, G.M., 2000. Genetics, Demography and Viability of FragmentedPopulations. Cambridge University Press, Cambridge.
Zwick, P.D., Carr, M.H., 2006. Florida 2060: a population distribution scenario for thestate of Florida. University of Florida GeoPlan Center. <http://www.1000friendsofflorida.org/PUBS/2060/Florida-2060-Report-Final.pdf> (accessed December2008).




















