
Darwin’s concepts in a test tube: Parallels between organismal and in vitro evolution
8
The International Journal of Biochemistry & Cell Biology 41 (2009) 266–273 Contents lists available at ScienceDirect The International Journal of Biochemistry & Cell Biology journal homepage: www.elsevier.com/locate/biocel Review Darwin’s concepts in a test tube: Parallels between organismal and in vitro evolution Carolina Díaz Arenas, Niles Lehman ∗ Department of Chemistry, Portland State University, Portland, OR, USA article info Article history: Available online 3 September 2008 Keywords: Evolution Selection In vitro evolution Ribozymes RNA abstract The evolutionary process as imagined by Darwin 150 years ago is evident not only in nature but also in the manner in which naked nucleic acids and proteins experience the “survival of the fittest” in the test tube during in vitro evolution. This review highlights some of the most apparent evolutionary patterns, such as directional selection, purifying selection, disruptive selection, and iterative evolution (recurrence), and draws parallels between what happens in the wild with whole organisms and what happens in the lab with molecules. Advances in molecular selection techniques, particularly with catalytic RNAs and DNAs, have accelerated in the last 20 years to the point where soon any sort of complex differential hereditary event that one can ascribe to natural populations will be observable in molecular populations, and exploitation of these events can even lead to practical applications in some cases. © 2008 Elsevier Ltd. All rights reserved. Contents 1. Introduction ......................................................................................................................................... 266 2. In vitro evolution .................................................................................................................................... 268 3. Directional selection ................................................................................................................................ 269 4. Purifying selection .................................................................................................................................. 270 5. Disruptive selection ................................................................................................................................. 271 6. Iterative evolution ................................................................................................................................... 271 7. The future: unexplored evolutionary parallels ..................................................................................................... 272 Acknowledgements ................................................................................................................................. 272 References ........................................................................................................................................... 272 1. Introduction Darwin (1859) outlined a mechanism by which evolutionary change is realized in nature. The process requires four circum- stances, which are rarely violated in the biosphere: 1. There is variation among individuals in a population. 2. This variation is heritable in that offspring resemble their par- ents. 3. More individuals are born to a population than the environment can support. ∗ Corresponding author at: Department of Chemistry, Portland State University, PO Box 751, Portland, OR 97207, USA. Tel.: +1 503 725 8769. E-mail address: [email protected] (N. Lehman). 4. The survival and reproduction of individuals is dependent on which traits they inherit. We now know that the ultimate basis for heritable variation is a consequence of the nucleotide sequence, structure, and func- tion of nucleic acids. Thus in the last half century, biochemists have embarked on the endeavor of applying the principles of natural selection to naked molecules, in an attempt to evolve new molec- ular activities. What happens in the wild can just as easily happen in the test tube, but is now manifest as in vitro evolution, in vitro selection, or SELEX, depending on the means and motives of the experiment. Many of the milestones in this field have been nicely summarized by Wilson and Szostak (1999) and Joyce (2004, 2007). In the current review, we note that – despite the fact that some or all of Darwin’s postulates can in fact be disobeyed in the lab – inevitably certain evolutionary patterns still develop, akin to those 1357-2725/$ – see front matter © 2008 Elsevier Ltd. All rights reserved. doi:10.1016/j.biocel.2008.08.034
-
Upload
independent -
Category
Documents
-
view
5 -
download
0
Transcript of Darwin’s concepts in a test tube: Parallels between organismal and in vitro evolution

R
Da
CD
a
AA
KESIRR
C
1
cs
2
3
P
1d
The International Journal of Biochemistry & Cell Biology 41 (2009) 266–273
Contents lists available at ScienceDirect
The International Journal of Biochemistry& Cell Biology
journa l homepage: www.e lsev ier .com/ locate /b ioce l
eview
arwin’s concepts in a test tube: Parallels between organismalnd in vitro evolution
arolina Díaz Arenas, Niles Lehman ∗
epartment of Chemistry, Portland State University, Portland, OR, USA
r t i c l e i n f o
rticle history:vailable online 3 September 2008
a b s t r a c t
The evolutionary process as imagined by Darwin 150 years ago is evident not only in nature but also in themanner in which naked nucleic acids and proteins experience the “survival of the fittest” in the test tube
eywords:volutionelectionn vitro evolutionibozymesNA
during in vitro evolution. This review highlights some of the most apparent evolutionary patterns, suchas directional selection, purifying selection, disruptive selection, and iterative evolution (recurrence), anddraws parallels between what happens in the wild with whole organisms and what happens in the lab withmolecules. Advances in molecular selection techniques, particularly with catalytic RNAs and DNAs, haveaccelerated in the last 20 years to the point where soon any sort of complex differential hereditary eventthat one can ascribe to natural populations will be observable in molecular populations, and exploitation
of these events can even lead to practical applications in some cases.© 2008 Elsevier Ltd. All rights reserved.
ontents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2662. In vitro evolution. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2683. Directional selection . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2694. Purifying selection . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2705. Disruptive selection . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2716. Iterative evolution . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2717. The future: unexplored evolutionary parallels . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 272
Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 272References. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 272
. Introduction
Darwin (1859) outlined a mechanism by which evolutionaryhange is realized in nature. The process requires four circum-tances, which are rarely violated in the biosphere:
4. The survival and reproduction of individuals is dependent onwhich traits they inherit.
We now know that the ultimate basis for heritable variationis a consequence of the nucleotide sequence, structure, and func-tion of nucleic acids. Thus in the last half century, biochemists have
1. There is variation among individuals in a population.. This variation is heritable in that offspring resemble their par-
ents.. More individuals are born to a population than the environment
can support.
∗ Corresponding author at: Department of Chemistry, Portland State University,O Box 751, Portland, OR 97207, USA. Tel.: +1 503 725 8769.
E-mail address: [email protected] (N. Lehman).
esuisesIoi
357-2725/$ – see front matter © 2008 Elsevier Ltd. All rights reserved.oi:10.1016/j.biocel.2008.08.034
mbarked on the endeavor of applying the principles of naturalelection to naked molecules, in an attempt to evolve new molec-lar activities. What happens in the wild can just as easily happen
n the test tube, but is now manifest as in vitro evolution, in vitroelection, or SELEX, depending on the means and motives of the
xperiment. Many of the milestones in this field have been nicelyummarized by Wilson and Szostak (1999) and Joyce (2004, 2007).n the current review, we note that – despite the fact that somer all of Darwin’s postulates can in fact be disobeyed in the lab –nevitably certain evolutionary patterns still develop, akin to those
C. Díaz Arenas, N. Lehman / The International Journal of Biochemistry & Cell Biology 41 (2009) 266–273 267
Fig. 1. Parallel chronologies of molecular (top) and organismal (bottom) lineages. The diagram contrasts the reconstruction of the evolutionary pathway between a molecularpopulation (ligase class I ribozymes) and an example organism (Testudines). The reconstruction of the evolutionary path of the ligase ribozymes is smooth because enormousamounts of data can be collected and kept. Conversely, in the dramatic case of the Testudines, the reconstruction of the evolutionary pathway has not yet achieved consensus.There is no certainty whether the members of the group lack the temporal fenestrae since ancient times (Anapsids and Parareptilia) or if they lost them secondarily (Diapsido paredP Thus,i onoidee
tpow
nrtTbp
(
rigin). Data are that of Gaffney (1990), Rieppel (1993), and Romer (1956) who comroganochelys and other turtles is not like that of the Anapsids from the Paleozoic.s polemic. There is no consensus if the closest relative is Pareiasauria or Procolophvolutionary pathway when there is a lack of registry.
hat have taken place during the historical evolution of organismalopulations. Thus both beasts and molecules are bound by manyf the same underlying constraints, and it is useful to scientistsorking at both levels to investigate what these may be.
A major problem facing evolutionary biologists is the disconti-uity in the registry of species, because most species that have beeneplaced by other species have been lost from records throughoutime exacerbating the impossibility to replay the evolutionary tape.his problem is evident at multicellular and unicellular levels, andecomes more critical as we travel back in time. Two examples arerovided:
1) At the multicellular level, the lack of registry has imposedserious difficulties in classifying the orders Testudines andSaurischia of Mesozoic reptiles. Inclusion of fossil records in
cladistic analysis has the advantage of providing more sam-pling data points and makes the inferred homologies morerobust. The downside of this is that the topology of the inferredcladograms can change dramatically when compared withcladograms done without fossil data (Smith, 1994). Another(
the bone configuration in the skull of these reptiles, and showed that the skull ofthey may have a Diapsid origin. Also, the most recent known relative of Testudinesa. The case of Testudines is an example of how difficult it can be to reconstruct an
problem is that the cladograms based on molecular studies donot have the same output as cladograms based on morpholog-ical studies. A broadly accepted view was to consider turtlesanapsid reptiles (those without temporal fenestrae in the skull),with the Captorhinidae as its closer extinct relative. More recentdata, however, support a closer affinity of the Testudines to thediapsid reptiles (with fenestrae in the skull). Molecular stud-ies relate turtles to archosaurs (crocodiles and birds), whereasmorphological studies relate them to the lepidosaurs (tuatara,lizards, and snakes). This uncertainty with turtles has implica-tions in reconstruction of the evolutionary path that they havefollowed. They can be considered as living fossils of the prim-itive (late Carboniferous/early Permian) reptiles – all of whichare otherwise extinct – or they can be considered modern rep-tiles, with secondarily loss of their temporal fenestrae (Zardoya
and Meyer, 2001). Fig. 1 depicts some of these issues juxtaposedto progress made in one realm of in vitro selection.2) At the unicellular level, there is uncertainty regarding thearchaeal and eubacterial roots in the tree of life. Archaea andEubacteria are the most primitive organisms on the Earth

268 C. Díaz Arenas, N. Lehman / The International Journal of Biochemistry & Cell Biology 41 (2009) 266–273
lution
ocmb1pt
2
(sa1hf
itvtatoacpagta
mnwd(blrtovIrt
ao
Fig. 2. The in vitro evo
and could be classified in the same high hierarchical groupof “prokaryotes.” In spite of their antiquity they have majordifferences in structure and genetics, leading Woese and Fox(1977) to propose their division into Archaea and Bacteria. Onestriking difference is the constitution of the cell membrane: inArchaea, the phospholipid backbone is built on a sn-glycerol-1-phosphate (G1P) instead of sn-glycerol-3-phosphate (G3P).The archaeal hydrophobic lipid chain is based on isoprenoidsinstead of fatty acids, and the isoprenoids are linked to the G1Pby an ether linkage instead of ester linkage. The attachment ofthe isoprenoid chain to the head group is done by the (S)-3-O-geranylgeranylglyceryl phosphate synthase (GGGPS) enzyme.This enzyme has probably evolved by duplication and fusion ofa half-barrel ancestor (Payandeh and Pai, 2007), and may havedriven the speciation of the Archaea in ancient times. The evo-lutionary history of GGGPS mirrors the emergence of Archaeafrom the last universal common ancestor, but without a timemachine, this will eternally remain speculation.
By studying evolutionary processes in the laboratory we can freeurselves from some of these historical constraints. We sacrificeomplexity, but gain resolution when we trace the responses ofolecules in solution to evolutionary pressures. From its auspicious
eginnings with the work of Spiegleman and Eigen in the 1960s and970s, evolution in a test tube has itself evolved into an incredibleowerful tool to examine the intricacies of heritable change overime (Joyce, 2007).
. In vitro evolution
The elements of artificial evolution, as applied to nucleic acidsor other information-bearing polymers), are threefold: mutation,
election, and amplification (Fig. 2). The PCR and related tools suchs automated nucleic-acid and gene synthesis, 3SR (Guatelli et al.,990) and RT-PCR that became widely available in the late 1980sas allowed the isolation of target DNA, RNA, and protein sequencesrom starting pools as large as 1015 different variants (1 nanomole)
phtfi
process (Joyce, 1989).
n a few days’ time. Molecules that with desired catalytic abili-ies can be fished out of these large pools by simple culling (initro selection), as can molecules with desired binding affinitiesowards exogenous molecular substrates (SELEX). But if there isny process that generates new mutations during the experiment,hen evolution can happen “on the fly” and the experiment is reallyne of in vitro evolution. In truth this is practically unavoidable,s any amplification process including, and in many cases espe-ially, the PCR will generate occasional mistakes in the copyingrocess, thereby generating new genetic diversity. Less appreci-ted but no less important is that chimeric molecules are oftenenerated either intentionally or unintentionally during the selec-ion process by some sort of recombination mechanism (Lehmannd Unrau, 2005).
As the process shown in Fig. 2 operates, the population ofolecules undergoes genotypic changes and corresponding phe-
otypic shifts. This relationship is easily seen in the case of RNA,here the genotype (the primary sequence of nucleotides) has airect, and effectively instantaneous, bearing on the phenotypethe catalytic and/or binding activity). As generational time goesy, the population can be viewed as exploring a Wrightian fitnessandscape, as first described by Maynard Smith (1970). A simplisticepresentation of this would be a three-dimensional graph, withhe x and y coordinates determining in some fashion the genotypef a particular individual, with the z coordinate being the fitnessalue (W) of the phenotype that would result from this genotype.n reality of course, a genotype of n monomer units in length wouldequire a multidimensional graph with n dimensions for the geno-ype.
In any event, one could – in theory – track the progression ofpopulation by tracing the path of a point (e.g., the dominant
r wildtype individual) or group of points (e.g., the population or
opulational mean) over time. This would be watching evolutionappen in real time. Of the hundreds of in vitro evolution studieshat have been performed to date, only a few have actually beenollowed in enough genotypic or phenotypic detail through time tonfer evolutionary subtleties. Usually the goal is to obtain a novel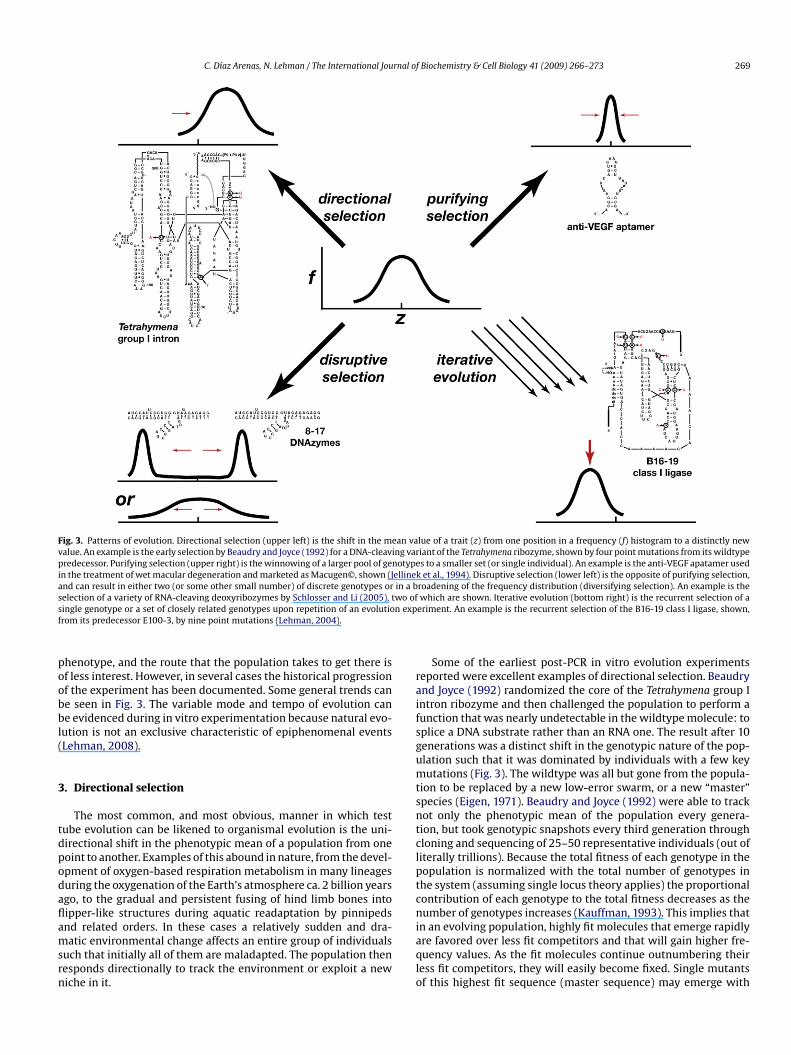
C. Díaz Arenas, N. Lehman / The International Journal of Biochemistry & Cell Biology 41 (2009) 266–273 269
Fig. 3. Patterns of evolution. Directional selection (upper left) is the shift in the mean value of a trait (z) from one position in a frequency (f) histogram to a distinctly newvalue. An example is the early selection by Beaudry and Joyce (1992) for a DNA-cleaving variant of the Tetrahymena ribozyme, shown by four point mutations from its wildtypepredecessor. Purifying selection (upper right) is the winnowing of a larger pool of genotypes to a smaller set (or single individual). An example is the anti-VEGF apatamer usedin the treatment of wet macular degeneration and marketed as Macugen©, shown (Jellinek et al., 1994). Disruptive selection (lower left) is the opposite of purifying selection,and can result in either two (or some other small number) of discrete genotypes or in a broadening of the frequency distribution (diversifying selection). An example is thes wo ofs n expf
poobbl(
3
tdpodaflamsrn
raifsgumtsntclptcn
election of a variety of RNA-cleaving deoxyribozymes by Schlosser and Li (2005), tingle genotype or a set of closely related genotypes upon repetition of an evolutiorom its predecessor E100-3, by nine point mutations (Lehman, 2004).
henotype, and the route that the population takes to get there isf less interest. However, in several cases the historical progressionf the experiment has been documented. Some general trends cane seen in Fig. 3. The variable mode and tempo of evolution cane evidenced during in vitro experimentation because natural evo-
ution is not an exclusive characteristic of epiphenomenal eventsLehman, 2008).
. Directional selection
The most common, and most obvious, manner in which testube evolution can be likened to organismal evolution is the uni-irectional shift in the phenotypic mean of a population from oneoint to another. Examples of this abound in nature, from the devel-pment of oxygen-based respiration metabolism in many lineagesuring the oxygenation of the Earth’s atmosphere ca. 2 billion yearsgo, to the gradual and persistent fusing of hind limb bones intoipper-like structures during aquatic readaptation by pinnipeds
nd related orders. In these cases a relatively sudden and dra-atic environmental change affects an entire group of individualsuch that initially all of them are maladapted. The population thenesponds directionally to track the environment or exploit a newiche in it.
iaqlo
which are shown. Iterative evolution (bottom right) is the recurrent selection of aeriment. An example is the recurrent selection of the B16-19 class I ligase, shown,
Some of the earliest post-PCR in vitro evolution experimentseported were excellent examples of directional selection. Beaudrynd Joyce (1992) randomized the core of the Tetrahymena group Intron ribozyme and then challenged the population to perform aunction that was nearly undetectable in the wildtype molecule: toplice a DNA substrate rather than an RNA one. The result after 10enerations was a distinct shift in the genotypic nature of the pop-lation such that it was dominated by individuals with a few keyutations (Fig. 3). The wildtype was all but gone from the popula-
ion to be replaced by a new low-error swarm, or a new “master”pecies (Eigen, 1971). Beaudry and Joyce (1992) were able to trackot only the phenotypic mean of the population every genera-ion, but took genotypic snapshots every third generation throughloning and sequencing of 25–50 representative individuals (out ofiterally trillions). Because the total fitness of each genotype in theopulation is normalized with the total number of genotypes inhe system (assuming single locus theory applies) the proportionalontribution of each genotype to the total fitness decreases as theumber of genotypes increases (Kauffman, 1993). This implies that
n an evolving population, highly fit molecules that emerge rapidlyre favored over less fit competitors and that will gain higher fre-uency values. As the fit molecules continue outnumbering theiress fit competitors, they will easily become fixed. Single mutantsf this highest fit sequence (master sequence) may emerge with

2 rnal of
fismte
sgrabdrtrt“plp
mToeJoaqolpet
4
sdOtaspaiertctie
IaIpfclpr
t1s(
lidltfiD(–cbateiitfla2t
ea(tJtwgrpuio
megcwmotbTglvtsRcg
70 C. Díaz Arenas, N. Lehman / The International Jou
tness values and competing abilities alike. This specific group ofequences (the neutral network) reduces the probability of less fitutants to become fit, by increasing the error threshold. The neu-
ral network hence buffers decreases of the mean fitness due to themergence of less fit mutants.
Other early experiments did not trace the evolutionary courseo finely, but clearly demonstrated directional selection. Oneood example was the successful selection of a self-biotinylatingibozyme by Wilson and Szostak (1995). These authors first isolatedset of RNA sequences that demonstrated an enhanced ability toind biotin. From there, the most promising sequences were ran-omized and selected for self-alkylation catalytic activity. Sevenounds of in vitro selection improved the populational activity inhis regard more than three orders of magnitude, and a subsequenteselection after randomizing a particularly active isolate improvedhe activity another 100-fold. This reselection is a classic example ofdeclaring a new wildtype” (Joyce, 2004) and evokes a parallel withunctuated equilibrium (Eldredge and Gould, 1972) in which evo-
ution occurs in discrete stages from a series of ever-higher fitnesslateaus.
In the past decade, the power of in vitro evolution to generateolecules with desired properties has been even more utilitarian.
he focus has shifted far away from the process and more sharplyn pushing the boundaries of new phenotypes. The aldolase (Fuszt al., 2005) and Diels-Alderase (Tarasow et al., 1997; Seeleg andäschke, 1999) ribozymes are good case studies. Interestingly, allf these studies seem to converge on a relatively efficient solutionfter six to eight generations of selection, which is in part a conse-uence of the practical limitations of the size of the starting poolf genotypes, typically 1012 to 1015 molecules. Though seeminglyarge, these sizes rarely allow all possible sequences to be sam-led, just as very few conceivable organismal genotypes have beenxposed to the evolutionary process during the history of life onhe Earth.
. Purifying selection
Another type of selection is purifying selection, also known astabilizing selection in less severe circumstances. These occur afterirectional selection has produced an evolutionary uphill path.nce at the peak in a fitness landscape, the population has reached
he maximum possible degree of adaptation, and thus any vari-tion from that will be in the downhill direction and negativelyelected. There are numerous examples of natural populations thatersist at or near the peaks of their fitness landscapes. In humans,study done by Hughes et al. (2003) to estimate the heterozygos-
ty at 1442 single-nucleotide polymorphism (SNP) loci, used anthnically diverse sample of individuals. There was an observededuction in gene diversity at these SNP loci resulting from nega-ive selection acting on variations that caused a protein structuralhange. Similar studies show a strong purifying selection in theransmission of the mammalian genome (Steward et al., 2008), andn free-living and obligate intracellular proteobacteria (Mamirovat al., 2007).
Stabilizing selection is a common phenomenon in nature.t occurs when there is an intermediate optimum for fitness,nd thus the fitness value is lower at the phenotypic extremes.n natural populations, stabilizing selection can happen byopulation–ecosystem interactions. An example is the gall size
ormed by the larvae of Eurosta spp., on goldenrod plants. In thisase, the size of the gall produced by the plant is determined by thearvae that the female fly deposits inside the goldenrod stems. Aarasitoid wasp will attack galls small enough for its ovipositor toeach the larvae inside; at the same time birds will feed on larvaewEttd
Biochemistry & Cell Biology 41 (2009) 266–273
hat are inside galls big enough to be easily seen (Weis and Gorman,990). Human birth weight is also thought to be under stabilizingelection, as values on either extreme lead to reduced survivorshipBell, 2008).
Turning to the molecular level, most SELEX experiments or iso-ation of catalysts from completely random pools of sequences aren fact examples of purifying selection. Although the investigatoroes not know a priori which sequence(s) will satisfy a particu-
ar selection criterion, if random sequence pools are used to starthe experiment, there will be a distinct peak somewhere in thetness landscape. The first SELEX examples, RNA aptamers to T7NA polymerase (Tuerk and Gold, 1990) or polycyclic organic dyes
Ellington and Szostak, 1990), were cases in which relatively smallbut completely random – pools of RNA sequences were iteratively
ulled to a subset of active sequences. A few years later, the ground-reaking selection of a theretofore unknown ligase ribozyme fromsparse pool of RNAs with 220 completely random nucleotide posi-
ions by Bartel and Szostak (1993) stands as the quintessentialxample of purifying selection targeted towards catalytic activ-ty, as opposed to binding affinity. Subsequent experiments tomprove ligase ability by partially randomizing certain portions ofhe molecule (Ekland and Bartel, 1995; Ekland et al., 1995) wouldall under the directional selection classification. Although whenarge completely random elements were added to the ligase in thettempt to make it a self-replicating polymerase (Johnston et al.,001), this would more aptly be described again as purifying selec-ion.
Purifying selection can also be deliberately forced during in vitrovolution, using a population that is already known to be located inn optimum peak of the fitness landscape. Soll et al. (2007) used thethen) fastest known synthetic ligase ribozyme, in terms of kcat, andhe continuous evolution (CE) in vitro methodology (Wright andoyce, 1997) to watch a wildtype fall from grace. It was suspectedhat a particular variant of the class I ligase, termed B16-19, was soell adapted to the CE environment that there was likely no place to
o but down, as most mutations in this sequence would be delete-ious to either survival or reproduction (Joyce, 2004). Various-sizedopulations, purely composed of the 152-nt B16-19 ribozyme, weresed to test for the impacts of mutation accumulation through the
nexorable decline in fitness that results upon successive mutations,r Muller’s Ratchet.
Soll et al. (2007) observed purifying selection in the extreme:any of the lineages actually went extinct in fewer than 150 gen-
rations, and the smaller populations had a greater tendency too extinct and/or do so earlier. This is perhaps the only knownase where an in vitro evolution experiment has been startedith no genetic diversity. Here, the initial population size and theeans to introduce genetic variability played crucial roles in the
nset of Muller’s Ratchet. During replication that includes muta-ional generation by polymerase errors, larger populations have aroader genetic variability from which to sample each generation.he probability that a mutant genotype can be drawn by simpleenetic drift in one sampling event is smaller because they areess represented in proportion to the whole; if the initial geneticariability is broad, a mutant genotype will have less weight inhe genetic pool of the population. The shape of the fitness land-cape being explored also plays a role in the onset of Muller’satchet. Starting with a highly fit initial population of individuallones (e.g., B16-19) has a similar effect as starting with a wideenetic variability because (1) a mutant genotype will have less
eight overall and (2) they will have to overcome a more stringentigen’s error threshold than if there were no superior phenotypeo start with. If the wildtype is in a fitness peak, emergent geno-ypes will be largely underrepresented and can get lost by simplerift (which includes stochastisity during manual pipetting) with a

rnal of
fi1
mtsntwdotfildbtiGoC(n
tbptPRmfbOc1RoVMhmpil
5
sflvde(c
ftrnFea
aImgha
tqwtmA(aDstiothutmtottd
6
goet“ptTipcNmMbasiiltrcir
C. Díaz Arenas, N. Lehman / The International Jou
nite probability in populations below a certain size (Lynch et al.,993).
Also, the higher the mutational rate the larger the number ofutations introduced in the population per generation. During CE,
he major source for new mutations is the error rate of reverse tran-criptase, which can be on the order of one mistake every 1000ucleotides copied. A back-of-the-envelope calculation suggestshat about 1 in 100 ligases synthesized during continuous evolutionill be a new variant under standard (non-hypermutagenic) con-itions (Soll et al., 2007). For populations that have settled downn a fitness landscape, the neutral network had increased the errorhreshold such that emergent mutants can steeply go down to a lowtness valley by the effect of only a few mutations. B16-19 is already
ocated in a fitness peak; from this location it is more probable to goown the low fitness valley than to hurdle into another fitness peaky the effect of the mutations. Such a high stringency implies thathe accumulation of mutations has a negatively synergistic effectn driving genotypes down into the valley of extinction (Lynch andabriel, 1990; Gabriel et al., 1993). Analogous events have beenbserved in small populations of E. coli (Kibota and Lynch, 1996),aenorhabditis elegans (Estes et al., 2004), Drosophila melanogasterMukai, 1964), and Arabidopsis thaliana (Schultz et al., 1999), just toame a few, during mutation accumulation experiments.
It is also worth noting that purifying selection has now beguno have definite biomedical applications. The SELEX technique haseen used to successfully isolate nucleic acid sequences that haveharmacological properties in that they can bind specifically andightly to desired targets. A particularly notable example is the drugegaptanib, which is marketed under the name Macugen©. ThisNA is injected into the eye, where is can treat wet age-relatedacular degeneration by acting as a vascular endothelial growth
actor (VEGF) antagonist. It is not a cure for macular degenerationut has proven efficacy in slowing the progression of the disease.ne productive SELEX experiment started with a pool of 1014 RNAsontaining 30 completely randomized nucleotides (Jellinek et al.,994). This eventually lead to the isolation of one superior 24-ntNA motif (Fig. 3), which, when chemically modified with 2′-aminer 2′-fluoro groups to enhance in vivo stability, displayed excellentEGF binding specificity and paved the way for the formulization ofacugen (Ng et al., 2006). Despite the fact that there were several
uman interventions in this process, with accompanying decision-aking events, the ultimate “victory” by 1 sequence out of 1014
ossible is unambiguously purifying selection . . . and is comparablen likelihood to the odds that a particular sperm genotype or oysterarva will make its way to adulthood after production.
. Disruptive selection
In some sense, disruptive, or diversifying selection, is the oppo-ite of purifying selection. Here, the phenotypic distribution isattened or at least spread into two peaks away from an initialalue relatively sharply distributed around a mean. The observediversification is a consequence of the potential of the individuals toxplore the genotypic landscape. Depending on the variables at playenvironment, etc.), the selection forces acting on the populationan be of different types.
In nature, examples of disruptive selection abound, and can beound at higher taxonomic levels and within individual species. Inhe former case, the process of speciation itself is a type of dis-
uptive selection, where the (often sudden) availability of multipleiches allows an ancestral species to rapidly diversify. Darwin’sinches on the Galápagos Islands, the mammalian radiation at thend of the Mezazoic Era, and the phyletic explosion that occurredt the beginning of the Cambrian period roughly 540 million yearswmet
Biochemistry & Cell Biology 41 (2009) 266–273 271
go are well described such events at progressively larger scales.n the latter case, genotypic diversity in a single species can be
aintained by circumstances that disfavor the fixation of any sin-le type. The blood-group and immunogenetic polymorphisms inumans, most famously the sickle-cell anemia hemoglobin morphs,re classic examples.
At the molecular level, diversifying selection is far less commonhan directional or purifying selection. This is partially a conse-uence of the fact that most in vitro selection studies are performedith a discrete phenotypic goal in mind, and processes that main-
ain diversity are either ignored or outright avoided. Nevertheless,olecular diversification probably occurs more often than realized.case in point is the elegant study described by Schlosser and Li
2005). Here, DNA enzymes (deoxyribozymes) with RNA-cleavingbility (Fig. 3) were selected from pools consisting of 250 pmol ofNAs containing 80 random nucleotides. Of interest is that two
elections were performed in parallel, one with selection pressurehat was increased in steps over 24 rounds of selection, and onen which the selection pressure was maintained at a constant levelver 30 generations. While the first pool converged onto a rela-ively small number of genotypes, the second pool maintained aigh degree of diversity, to the point where the generation #30 pop-lation displayed a remarkably flat frequency distribution both athe genotypic and phenotypic levels. Mechanisms to promote and
aintain diversity in the latter pool included a permissive selec-ion regime (many suboptimal genotypes could survive) and at leastne clear case of intergenotype recombination. The comparison ofhese two lineages serves to highlight the ways in which attenua-ion of environmental conditions – even in the test tube – can haveistinctive evolutionary effects.
. Iterative evolution
Iterative evolution, or recurrence, is the tendency for a particularenotype or phenotype to repeatedly appear under a specified setf selection conditions. In the wild, the environment rarely allowsxact repetition of evolution, leading Gould (1989) to ask whetherhe same species and traits would evolve on the Earth should theevolutionary tape be played twice”. Mutation itself is a stochasticrocess, and in nature one cannot even count on the same geno-ypic repertoire to be available even if the environment is recurrent.he face that mutations will adopt in a population depends on var-ous conditions. Advantageous mutations offer an advantage to theopulation during changing environmental conditions, thus, it isonvenient for the population to have a variable genetic reservoir.evertheless, a few cases of iterative evolution have been docu-ented at the organismal level. It was particularly common inesozoic ammonites, swimming cephalopods with intricate cham-
ered shells (Bayer and McGhee, 1984). A particular case is anncestral shelf-ocean ammonite that reappeared often times as theea level arose and receded. These ammonites were very similarn their morphologies and evolved from the open-ocean form thatteratively adapted to the new hydrodynamic conditions in the shal-ow waters when the sea level decreased at different times duringhe Mesozoic. In bacterial population monitored in the laboratory,ecurrence of phenotypes can also be seen. Travisano et al. (1995)hallenged parallel populations of E. coli with carbon-source lim-tations and found that after 1000 generations, the fitnesses of 36eplicate populations were not statistically different.
In vitro selection offers us the ability to see more sharply whatould happen during iterative evolution. Although the environ-ent and the phenotypes are not nearly as complex as the two
xamples described above, we do have the opportunity to play theape as many times as desired, with or without changing single vari-

2 rnal of
atsapa
fatwDtribsStiupmghsttoi1prptt
fd1sCisnI7pu16Asmc2
lepatmla
cgitrmssl
7
mbeTbmeppet2ssimtpsbebVtoc
neaimbt
A
dAt
R
B
B
72 C. Díaz Arenas, N. Lehman / The International Jou
bles. In vitro evolution gives the advantage of being able to observehe path of evolution in a time frame several orders of magnitudehorter than what happens in nature; thus, the registries of anynd all parallel lineages are observable. Therefore, it offers a com-rehensive tool to better understand all the intricate mechanismsnd factors shaping the evolution of organisms.
Laboratory cultures of catalytic RNA molecules can be sustainedor several generations under stable conditions and the emergencend fixation of mutant molecules can be evidenced. For example,he Tetrahymena ribozyme has a tendency to evolve towards some-hat predictable genotypes in certain environments (Hanczyc andorit, 2000). But the clearest case of iterative evolution have come
hrough the study of class I ligase ribozymes. The discovery of suchibozymes by Bartel and Szostak (1993) has lead to several studiesn different laboratories and a phylogeny of RNA molecules haveeen formed through years (Joyce, 2004). The initial experimentstarted from a very large pool of variable molecules (Bartel andzostak, 1993) and used separate vessels for the catalytic reaction,he amplification process, and the selection step. Subsequent exper-ments that used the original class I ligase b201 as a branch pointsed improved techniques, especially the CE method in which bothrocesses are coupled and thus evolution occurs more akin to itsode in natural populations (Fig. 1). In general, these RNA phylo-
enies show clear paths of diversification and evolution, as mayave occurred at the origins of life and evolution of organismalpecies. The rapid pace of the CE methodology makes it practicalo test evolutionary mechanism by latitudinal analysis of a popula-ion. Compared to stepwise evolution, continuous in vitro evolutionffers a more realistic model of biological evolution because theres a reduced intervention of the experimenter (Wright and Joyce,997; Johns and Joyce, 2005). There is a direct link between thehenotype and the breeding characteristics of the individuals (e.g.,ibozymes) in the molecular population: the fittest ribozyme canroduce more progeny in a given period of time. Of relevance here ishe highly fit genotype B16-19, has descended from the b201 ances-or not once, but literally dozens of times in different laboratories.
We have studied the recurrence of B16-19 mutations (Fig. 3) andound that the ligase iteratively jumps from peak to peak and hur-les the deep valley of its fitness landscape (Schmitt and Lehman,999; Lehman, 2004; Soll et al., 2007). This ribozyme thus repre-ents a deep local fitness optimum, as mentioned above. ThroughE, this genotype was evolved via nine point mutations from its
mmediate predecessor E100-3 originally in at least two distincttudies performed on opposite sides of the North American conti-ent (Ordoukhanian and Joyce, 1999; Schmitt and Lehman, 1999).
n the latter case, samples from the test tube were drawn at bursts, 13 and 17 and sequenced for genotypic characterization of theopulation. By burst 7 the precursor E100-3 dominated the pop-lation and showed no mutation in 28% of the clones. After burst3 eight mutations have already become fixed and the ninth was0% fixed. The population has clearly shifted its fittest phenotype.fter burst 16 the nine mutations were fixed and the dominantequence is that of B16-19 (clone 19 from burst 16). This experi-ent was later performed in parallel under strict contamination
ontrols and the same result occurred at least 13 times (Lehman,004), demonstrating the persistent recurrence of B16-19.
At a finer level of detail, structural recurrence is of great evo-utionary importance because structures that are recurrent canventually serve as a scaffold for the construction of more com-lex structures capable of higher informative content (Kennedy et
l., 2008). An example of a short RNA structure that appears itera-ively can be evidenced in vitro by the selection of the UAUU affinityotif for isoleucine aptamer (Lozupone et al., 2003). It has been iso-ated from pool sequences 22, 26, 50, 70 and 90 nucleotides long,nd is interesting because this quartet contains both an isoleucine
B
B
D
Biochemistry & Cell Biology 41 (2009) 266–273
odon (AUU) and an isoleucine anticodon (UAU) in the standardenetic code, lending some modest support to the stereochem-cal theories of the code’s origins. Of course, the shortness ofhese sequences casts doubt on the statistical significance of theirecurrence (Ellington et al., 2000). Another case is that of the ham-erhead motif, which, as a class of genotypes all displaying the
ame phenotype, has been shown to repeatedly result from in vitroelection studies targeted towards RNAs that self-cleave at a bio-ogically relevant rate (Salehi-Ashtiani and Szostak, 2001).
. The future: unexplored evolutionary parallels
The above examples illustrate the parallels between organis-al and raw molecular evolution. The two processes are bound
y the same set of forces, and generate patterns that are decid-dly analogous, simply manifest at differing levels of complexity.here are however, certain trends that natural populations haveeen seen to display for which the molecular counterpart is stillissing. Frequency-dependent selection, sexual selection, and het-
rochrony are evolutionary phenomena for which we know no goodarallel in the test tube. This begs the question of whether we sim-ly have not yet investigated molecular evolutionary systems innough detail, or whether these events are truly epiphenomenahat cannot be realized at the level of single molecules (Lehman,008). Heterochrony for example, requires selection operating topeed up or slow down the developmental process in an individualuch that different life stages may experience different advantagesn various environments. What is the developmental process of a
olecule? For a catalytic RNA, it can be the acquisition of a foldedhree-dimensional conformation following synthesis as a linearolymer. But most RNAs fold on the second or millisecond timecale, and thus unless selection can operate that fast, we may note able to detect any sort of heterochronic effect. The future how-ver holds great promise of new molecular methodologies that mayring to light some of these phenomena. In the case of heterochrony,oytek and Joyce (2007) recently applied selection for ligases in
he millisecond time frame, and proliferation of this technique andthers will undoubtedly lead to new molecular evolutionary dis-overies.
Finally, we note that there are evolutionary patterns that wouldot be characterized as Darwinian that are portended by molecularvolution. Can molecules evolve in groups, acquire and transmitcquired characteristics, or exhibit altruistic behavior? Again, its likely that as our understandings of the physical chemistry of
acromolecules and of the mathematics of organismal evolutionoth advance, dynamics such as these should be uncovered withime.
cknowledgements
We would like to thank members of the Lehman lab for usefuliscussions in the preparation of this manuscript and the Nationaleronautics and Space Administration for funding that supported
his work.
eferences
artel DP, Szostak JW. Isolation of new ribozymes from a large pool of randomsequences. Science 1993;261:1411–8.
ayer U, McGhee Jr GR. Iterative evolution of middle Jurassic ammonite faunas.Lethaia 1984;17:1–16.
eaudry AA, Joyce GF. Directed evolution of an RNA enzyme. Science1992;257:635–41.
ell G. Selection: the mechanism of evolution. 2nd edition Oxford: Oxford UniversityPress; 2008.
arwin CR. The origin of species by means of natural selection, or the preservationof favoured races in the struggle for life. London: John Murray; 1859.

rnal of
E
E
E
E
E
EE
F
G
G
G
G
H
H
J
J
J
JJ
JKK
K
L
L
L
L
L
L
M
M
M
N
O
P
R
RS
S
S
S
S
S
S
S
T
T
T
V
W
W
W
C. Díaz Arenas, N. Lehman / The International Jou
igen M. Selforganization of matter and the evolution of biological macromolecules.Naturwissenschaften 1971;58:465–523.
kland EH, Bartel DP. The secondary structure and sequence optimization of an RNAligase ribozyme. Nucl Acids Res 1995;23:3231–8.
kland EH, Szostak JW, Bartel DP. Structurally complex and highly active RNA ligasesderived from random RNA sequences. Science 1995;269:364–70.
ldredge N, Gould SJ. Punctuated equilibria: an alternative to phyletic gradualism.In: Schopf TJM, editor. Models in paleobiology. San Francisco: Freeman, Cooper& Co.; 1972. p. 82–115.
llington AD, Szostak JW. In vitro selection of RNA molecules that bind specificligands. Nature 1990;346:818–22.
llington AD, Khrapov M, Shaw CA. The scene of a frozen accident. RNA 2000:485–98.stes S, Phillips P, Denver D, Thomas K, Lynch M. Mutation accumulation in popula-
tions of varying size: the distribution of mutational effects for fitness correlatesin Caenorhabditis elegans. Genetics 2004;166:1269–79.
usz S, Eisenführ A, Srivatsan SG, Heckel A, Famulok M. A ribozyme for the aldolreaction. Chem Biol 2005;12:941–50.
abriel W, Lynch M, Bürger R. Muller’s ratchet and mutational meltdowns. Evolution1993;47:1744–57.
affney ES. The comparative osteology of the Triassic turtle Proganochelys. Bull AmMus Nat Hist 1990;194:259–63.
ould SJ. Wonderful life: the Burgess shale and the nature of history. New York: WWNorton & Co; 1989.
uatelli JC, Whitfield KM, Kwoh DY, Barringer KJ, Richman DD, Gingeras TR. Isother-mal, in vitro amplification of nucleic acids by a multienzyme reaction modeledafter retroviral replication. Proc Natl Acad Sci USA 1990;87:1874–8.
anczyc MM, Dorit RL. Replicability and recurrence in the experimental evolutionof a group I ribozyme. Mol Biol Evol 2000;17:1050–60.
ughes AL, Packer B, Welch R, Bergen AW, Chanock SJ, Yeager M. Widespread purify-ing selection at polymorphic sites in human protein-coding loci. Proc Natl AcadSci USA 2003;100:15754–7.
ellinek D, Green LS, Bell C, Janjic N. Inhibition of receptor binding by highaffinity RNA ligands to vascular endothelial growth factor. Biochemistry1994;33:10450–6.
ohns GC, Joyce GF. The promise and peril of continuous in vitro evolution. J Mol Evol2005;61:253–63.
ohnston WK, Unrau PJ, Lawrence MS, Glasner ME, Bartel DP. RNA-catalyzed RNApolymerization: accurate and general RNA-templated primer extension. Science2001;292:1319–25.
oyce GF. Amplification, mutation and selection of catalytic RNA. Gene 1989;82:83–7.oyce GF. Directed evolution of nucleic acid enzymes. Annu Rev Biochem
2004;73:791–836.oyce GF. Forty years of in vitro evolution. Angew Chem Int Ed 2007;46:2–19.auffman SA. The origins of order. New York: Oxford University Press; 1993.ennedy R, Lladser ME, Yarus M, Knight R. Information, probability, and the abun-
dance of the simplest RNA active sites. Front Biosci 2008;13:6060–71.ibota TT, Lynch M. Estimate of the genomic mutation rate deleterious to overall
fitness in E. coli. Nature 1996;381:694–6.ehman N. Assessing the likelihood of recurrence during RNA evolution in vitro. Artif
Life 2004;10:1–22.ehman N. The molecular underpinnings of genetic phenomena. Heredity
2008;100:6–12.
ehman N, Unrau PJ. Recombination during in vitro evolution. J Mol Evol2005;61:245–54.ozupone C, Changayil S, Majerfeld I, Yarus M. Selection of the simplest RNA that
binds isoleucine. RNA 2003;9:1315–22.ynch M, Gabriel W. Mutation load and the survival of small populations. Evolution
1990;44:1725–37.
W
W
Z
Biochemistry & Cell Biology 41 (2009) 266–273 273
ynch M, Burger R, Butcher D, Gabriel W. The mutational meltdown in asexualpopulations. J Hered 1993;84:339–44.
amirova L, Popadin K, Gelfand MS. Purifying selection in mitochondria, free-livingand obligate intracellular proteobacteria. BMC Evol Biol 2007;7:17.
aynard Smith J. Natural selection and the concept of a protein space. Nature1970;225:563–4.
ukai T. The genetic structure of populations of Drosophila melanogaster.I. Spontaneous mutation rate of polygenes controlling viability. Genetics1964;72:335–55.
g EWM, Shima DT, Calias P, Cunningham Jr ET, Guyer DR, Adamis AP. Pegaptanib,a targeted anti-VEGF aptamer for ocular vascular disease. Nat Rev Drug Disc2006;5:123–32.
rdoukhanian P, Joyce GF. A molecular description of the evolution of resistance.Chem Biol 1999;6:881–90.
ayandeh J, Pai E. Enzyme-driven speciation: crystallizing Archaea via lipid capture.J Mol Evol 2007;64:364–74.
ieppel O. Studies on skeleton formation in reptiles. IV. The homology of the reptilian(amniote) astragalus revisited. J Vert Paleontol 1993;13:31–47.
omer AS. Osteology of the reptiles. Chicago: Chicago University Press; 1956.alehi-Ashtiani K, Szostak JW. In vitro evolution suggests multiple origins for the
hammerhead ribozyme. Nature 2001;414:82–4.chlosser K, Li Y. Diverse evolutionary trajectories characterize a community of RNA-
cleaving deoxyribozymes: a case study into the population dynamics of in vitroselection. J Mol Evol 2005:192–206.
chmitt T, Lehman N. Non-unity heritability demonstrated by continuous evolutionin vitro. Chem Biol 1999;6:857–69.
chultz ST, Lynch M, Willis JH. Spontaneous deleterious mutation in Arabidopsisthaliana. Proc Natl Acad Sci USA 1999;96:11393–8.
eeleg B, Jäschke A. A small catalytic RNA motif with Diels-Alderase activity. ChemBiol 1999;6:167–76.
oll S, Díaz Arenas C, Lehman N. Accumulation of deleterious mutations in smallabiotic populations of RNA. Genetics 2007;175:267–75.
mith A. Systematics and the fossil record: documenting evolutionary patterns.Boston: Blackwell; 1994.
teward JB, Freyer C, Elson JL, Wredenberg A, Cansu Z, Trifunovic A, Larsson N-G.Strong purifying selection in transmission of mammalian mitochondrial DNA.PLoS Biol 2008;6:e10.
arasow TM, Tarasow SL, Eaton BE. RNA-catalysed carbon–carbon bond formation.Nature 1997;389:54–7.
ravisano M, Mongold JA, Bennett AF, Lenski RE. Experimental tests of the roles ofadaptation, chance, and history in evolution. Science 1995;267:87–90.
uerk C, Gold L. Systematic evolution of ligands by exponential enrichment: RNAligands to bacteriophage T4 DNA polymerase. Science 1990;249:505–10.
oytek SB, Joyce GF. Emergence of a fast reacting ribozyme that is capable ofundergoing continuous evolution. Proc Natl Acad Sci USA 2007;104:15288–93.
eis AE, Gorman WL. Measuring selection on reaction norms: an exploration of theEurosta-Solidago system. Evolution 1990;44:820–31.
ilson C, Szostak JW. In vitro evolution of a self-alkylating ribozyme. Nature1995;374:777–82.
ilson C, Szostak JW. In vitro selection of functional nucleic acids. Annu Rev Biochem1999;68:611–47.
oese CR, Fox GE. Phylogenetic structure of the prokaryotic domain: the primarykingdoms. Proc Natl Acad Sci USA 1977;74:5088–90.
right MC, Joyce GF. Continuous in vitro evolution of catalytic function. Science1997;276:614–7.
ardoya R, Meyer A. The evolutionary position of turtles revised. Naturwis-senschaften 2001;88:193–200.




















