
Crystal Structure of the TNF-α-Inducing Protein (Tipα) from Helicobacter pylori: Insights into Its...
7
Crystal Structure of the TNF-α-Inducing Protein (Tipα) from Helicobacter pylori: Insights into Its DNA-Binding Activity Jun Young Jang 1 †, Hye-Jin Yoon 1 †, Ji Young Yoon 1 , Hyoun Sook Kim 1 , Sang Jae Lee 1 , Kyoung Hoon Kim 1 , Do Jin Kim 1 , Soonmin Jang 2 , Byeong-Gu Han 3 , Byung Il Lee 3 and Se Won Suh 1,4 ⁎ 1 Department of Chemistry, College of Natural Sciences, Seoul National University, Seoul 151-742, Korea 2 Department of Chemistry, Sejong University, Seoul 143-747, Korea 3 Cancer Cell and Molecular Biology Branch, Division of Cancer Biology, Research Institute, National Cancer Center, Goyang, Gyeonggi 410-769, Korea 4 Department of Biophysics and Chemical Biology, College of Natural Sciences, Seoul National University, Seoul 151-742, Korea Received 6 March 2009; received in revised form 1 July 2009; accepted 6 July 2009 Available online 10 July 2009 Helicobacter pylori infection is one of the highest risk factors for gastroduodenal diseases including gastric cancer. Tumor necrosis factor- alpha (TNF-α) is one of the essential cytokines for tumor promotion, and thus, an H. pylori protein that induces TNF-α is believed to play a significant role in gastric cancer development in humans. The HP0596 gene product of H. pylori strain 26695 was identified as the TNF-α-inducing protein (Tipα). Tipα is secreted from H. pylori as dimers and enters the gastric cells. It was shown to have a DNA-binding activity. Here, we have determined the crystal structure of Tipα from H. pylori. Its monomer consists of two structural domains (“mixed domain” and “helical domain”). Tipα exists as a dimer in the crystal, and the dimeric structure represents a novel scaffold for DNA binding. A positively charged surface patch formed across the two monomers of the Tipα dimer by the loop between helices α1 and α2 may be important in DNA binding. © 2009 Elsevier Ltd. All rights reserved. Edited by K. Morikawa Keywords: tumor necrosis factor-α-inducing protein; Tipα; HP0596; Helicobacter pylori; gastric cancer Introduction Helicobacter pylori is a spiral-shaped, Gram-negative bacterium that lives in the human stomach and duodenum. H. pylori infection is one of the highest risk factors for gastroduodenal diseases including gastric cancer. 1 It has long been known that H. pylori infection results in an inflammatory response in the stomach by induction of various cytokines such as tumor necrosis factor-alpha (TNF-α), interleukin-1, and interleukin-8. 2 TNF-α is one of the essential cytokines for tumor promotion, and thus, a gene product of H. pylori that induces TNF-α is believed to play a significant role in gastric cancer develop- ment in humans. 3–5 The HP0596 gene product of H. pylori strain 26695 was identified as the TNF-α- inducing protein (Tipα). 6 The HP0596 gene encodes a protein of 192 amino acid residues with a signal *Corresponding author. E-mail address: [email protected] . † J.Y.J and H.-J.Y. contributed equally to this work. Abbreviations used: TNF-α, tumor necrosis factor-alpha; Tipα, TNF-α-inducing protein; MAD, multiwavelength anomalous diffraction; SeMet, selenomethionine; PDB, Protein Data Bank; NTD, N-terminal domain; dsDNA, double-stranded DNA. doi:10.1016/j.jmb.2009.07.010 J. Mol. Biol. (2009) 392, 191–197 Available online at www.sciencedirect.com 0022-2836/$ - see front matter © 2009 Elsevier Ltd. All rights reserved.
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Crystal Structure of the TNF-α-Inducing Protein (Tipα) from Helicobacter pylori: Insights into Its...

doi:10.1016/j.jmb.2009.07.010 J. Mol. Biol. (2009) 392, 191–197
Available online at www.sciencedirect.com
Crystal Structure of the TNF-α-Inducing Protein (Tipα)from Helicobacter pylori: Insights into ItsDNA-Binding Activity
Jun Young Jang1†, Hye-Jin Yoon1†, Ji Young Yoon1, Hyoun Sook Kim1,Sang Jae Lee1, Kyoung Hoon Kim1, Do Jin Kim1, Soonmin Jang2,Byeong-Gu Han3, Byung Il Lee3 and Se Won Suh1,4⁎
1Department of Chemistry,College of Natural Sciences,Seoul National University,Seoul 151-742, Korea2Department of Chemistry,Sejong University,Seoul 143-747, Korea3Cancer Cell and MolecularBiology Branch, Division ofCancer Biology, ResearchInstitute, National CancerCenter, Goyang, Gyeonggi410-769, Korea4Department of Biophysics andChemical Biology, College ofNatural Sciences, SeoulNational University,Seoul 151-742, KoreaReceived 6 March 2009;received in revised form1 July 2009;accepted 6 July 2009Available online10 July 2009
*Corresponding author. E-mail [email protected].† J.Y.J and H.-J.Y. contributed equAbbreviations used: TNF-α, tumor
Tipα, TNF-α-inducing protein; MADanomalous diffraction; SeMet, selenoProtein Data Bank; NTD, N-terminaldouble-stranded DNA.
0022-2836/$ - see front matter © 2009 E
Helicobacter pylori infection is one of the highest risk factors forgastroduodenal diseases including gastric cancer. Tumor necrosis factor-alpha (TNF-α) is one of the essential cytokines for tumor promotion, andthus, anH. pylori protein that induces TNF-α is believed to play a significantrole in gastric cancer development in humans. The HP0596 gene product ofH. pylori strain 26695 was identified as the TNF-α-inducing protein (Tipα).Tipα is secreted from H. pylori as dimers and enters the gastric cells. It wasshown to have a DNA-binding activity. Here, we have determined thecrystal structure of Tipα from H. pylori. Its monomer consists of twostructural domains (“mixed domain” and “helical domain”). Tipα exists as adimer in the crystal, and the dimeric structure represents a novel scaffold forDNA binding. A positively charged surface patch formed across the twomonomers of the Tipα dimer by the loop between helices α1 and α2 may beimportant in DNA binding.
© 2009 Elsevier Ltd. All rights reserved.
Keywords: tumor necrosis factor-α-inducing protein; Tipα; HP0596;Helicobacter pylori; gastric cancer
Edited by K. MorikawaIntroduction
Helicobacter pylori is a spiral-shaped, Gram-negativebacterium that lives in the human stomach and
ess:
ally to this work.necrosis factor-alpha;, multiwavelengthmethionine; PDB,domain; dsDNA,
lsevier Ltd. All rights reserve
duodenum. H. pylori infection is one of the highestrisk factors for gastroduodenal diseases includinggastric cancer.1 It has long been known that H. pyloriinfection results in an inflammatory response in thestomach by induction of various cytokines such astumor necrosis factor-alpha (TNF-α), interleukin-1,and interleukin-8.2 TNF-α is one of the essentialcytokines for tumor promotion, and thus, a geneproduct of H. pylori that induces TNF-α is believedto play a significant role in gastric cancer develop-ment in humans.3–5 The HP0596 gene product of H.pylori strain 26695 was identified as the TNF-α-inducing protein (Tipα).6 The HP0596 gene encodesa protein of 192 amino acid residues with a signal
d.
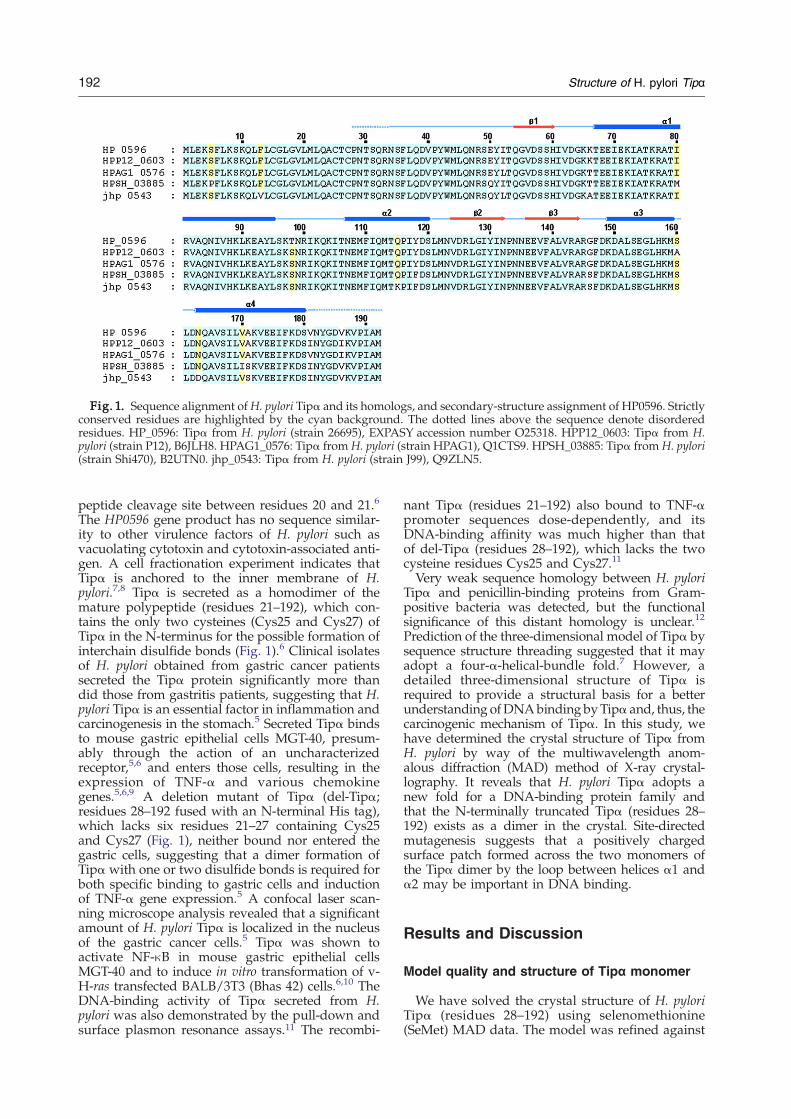
Fig. 1. Sequence alignment ofH. pylori Tipα and its homologs, and secondary-structure assignment of HP0596. Strictlyconserved residues are highlighted by the cyan background. The dotted lines above the sequence denote disorderedresidues. HP_0596: Tipα from H. pylori (strain 26695), EXPASY accession number O25318. HPP12_0603: Tipα from H.pylori (strain P12), B6JLH8. HPAG1_0576: Tipα from H. pylori (strain HPAG1), Q1CTS9. HPSH_03885: Tipα from H. pylori(strain Shi470), B2UTN0. jhp_0543: Tipα from H. pylori (strain J99), Q9ZLN5.
192 Structure of H. pylori Tipα
peptide cleavage site between residues 20 and 21.6
The HP0596 gene product has no sequence similar-ity to other virulence factors of H. pylori such asvacuolating cytotoxin and cytotoxin-associated anti-gen. A cell fractionation experiment indicates thatTipα is anchored to the inner membrane of H.pylori.7,8 Tipα is secreted as a homodimer of themature polypeptide (residues 21–192), which con-tains the only two cysteines (Cys25 and Cys27) ofTipα in the N-terminus for the possible formation ofinterchain disulfide bonds (Fig. 1).6 Clinical isolatesof H. pylori obtained from gastric cancer patientssecreted the Tipα protein significantly more thandid those from gastritis patients, suggesting that H.pylori Tipα is an essential factor in inflammation andcarcinogenesis in the stomach.5 Secreted Tipα bindsto mouse gastric epithelial cells MGT-40, presum-ably through the action of an uncharacterizedreceptor,5,6 and enters those cells, resulting in theexpression of TNF-α and various chemokinegenes.5,6,9 A deletion mutant of Tipα (del-Tipα;residues 28–192 fused with an N-terminal His tag),which lacks six residues 21–27 containing Cys25and Cys27 (Fig. 1), neither bound nor entered thegastric cells, suggesting that a dimer formation ofTipα with one or two disulfide bonds is required forboth specific binding to gastric cells and inductionof TNF-α gene expression.5 A confocal laser scan-ning microscope analysis revealed that a significantamount of H. pylori Tipα is localized in the nucleusof the gastric cancer cells.5 Tipα was shown toactivate NF-κB in mouse gastric epithelial cellsMGT-40 and to induce in vitro transformation of v-H-ras transfected BALB/3T3 (Bhas 42) cells.6,10 TheDNA-binding activity of Tipα secreted from H.pylori was also demonstrated by the pull-down andsurface plasmon resonance assays.11 The recombi-
nant Tipα (residues 21–192) also bound to TNF-αpromoter sequences dose-dependently, and itsDNA-binding affinity was much higher than thatof del-Tipα (residues 28–192), which lacks the twocysteine residues Cys25 and Cys27.11
Very weak sequence homology between H. pyloriTipα and penicillin-binding proteins from Gram-positive bacteria was detected, but the functionalsignificance of this distant homology is unclear.12
Prediction of the three-dimensional model of Tipα bysequence structure threading suggested that it mayadopt a four-α-helical-bundle fold.7 However, adetailed three-dimensional structure of Tipα isrequired to provide a structural basis for a betterunderstanding ofDNAbinding byTipα and, thus, thecarcinogenic mechanism of Tipα. In this study, wehave determined the crystal structure of Tipα fromH. pylori by way of the multiwavelength anom-alous diffraction (MAD) method of X-ray crystal-lography. It reveals that H. pylori Tipα adopts anew fold for a DNA-binding protein family andthat the N-terminally truncated Tipα (residues 28–192) exists as a dimer in the crystal. Site-directedmutagenesis suggests that a positively chargedsurface patch formed across the two monomers ofthe Tipα dimer by the loop between helices α1 andα2 may be important in DNA binding.
Results and Discussion
Model quality and structure of Tipα monomer
We have solved the crystal structure of H. pyloriTipα (residues 28–192) using selenomethionine(SeMet) MAD data. The model was refined against

Table 1. Data collection and refinement statistics
Data collectionData set SeMet I (inflection) SeMet I (peak) SeMet I (remote) SeMet II (remote)X-ray source PF BL-5A PF BL-5A PF BL-5A PLS BL-4AX-ray wavelength (Å) 0.97912 0.97885 0.96000 0.96000Temperature (K) 100 100 100 100Space group P212121 P212121 P212121 P212121a, b, c (Å) 52.55, 66.43, 90.55 52.55, 66.47, 90.54 52.58, 66.49, 90.58 53.52, 67.11, 91.26Resolution range (Å) 50.0–2.70 (2.80−2.70) 50.0–2.70 (2.80−2.70) 50.0–2.70 (2.80−2.70) 30.0–2.40 (2.49−2.40)Total/unique reflections 116,922/9199 231,055/9137 117,732/9096 214,284/13,650Rmerge
a (%) 8.3 (37.4) 9.8 (41.4) 9.0 (36.4) 9.3 (42.2)Data completeness (%) 99.8 (90.0) 98.7 (89.4) 98.3 (90.3) 100.0 (100.0)Average I/σ(I) 56.7 (3.7) 76.7 (4.7) 57.1 (4.0) 42.9 (7.2)Figure of meritb for MAD phasing: 0.73/0.88 for 20.0–2.70 Å data (before/after density modification)
RefinementResolution range (Å) 30.0–2.40No. of reflections (working/free set) 12,433/714No. of protein non-hydrogen atoms 2391No. of water molecules 70Rwork/Rfree
c (%) 23.3/24.1r.m.s. deviations from ideal geometry
Bond length (Å) 0.0067Bond angle (°) 1.15
Average B-factor (Å2)Protein 41.1Water molecules 43.6
Ramachandran plot for non-glycine and non-proline residuesMost favorable regions (%) (chains A/B) 92.8/90.6Allowed regions (%) (chains A/B) 6.5/9.4Generously allowed regions (%) (chains A/B) 0.7/0.0
Values in parentheses refer to the highest resolution shells [2.80–2.70 Å for the SeMet I MADdata and 2.49–2.40 Å for the SeMet II (remote)data].
a Rmerge =∑h ∑i | I(h)i – bI(h)N|/∑h ∑i I(h)i, where I(h)i is the intensity of the i-th measurement of reflection h and bI(h)N is the meanvalue of I(h) for all i measurements.
b Figure of merit = b|∑ P(α) exp(iα)/∑P(α)|N, where α is the phase angle and P(α) is the phase probability distribution.c R = ∑||Fobs|–|Fcalc||/∑|Fobs|, where Rfree is calculated for randomly chosen 10% of reflections, which were not used for
structure refinement, and Rwork is calculated for the remaining reflections.
193Structure of H. pylori Tipα
the SeMet II (remote) data (Table 1); it yielded Rworkand Rfree values of 23.3% and 24.1% for 30.0−2.40 Ådata, respectively. The model contains 296 residuesin two monomers of Tipα (residues 34–181 for bothchains A and B) and 70 water molecules in thecrystallographic asymmetric unit (Table 1). Themissing residues of Tipα and the N-terminal fusiontag are probably disordered in the crystal and areinvisible in the electron density map. An N-terminalloop region (residues 34–36 and 46–51 for chain A;residues 34–35 and 48–51 for chain B), a loopbetween β1 and α1 (residues 62–63 for chain B), aloop between β2 and α2 (132–136 for chain B), and aC-terminal region (179–181 for chain B) showrelatively high B-factors (N60 Å2).A monomer of Tipα (residues 28–192) consists of
four α-helices, three β-strands, and a number ofloops. Each monomer is somewhat elongated andcurved. It may be loosely divided into two structuraldomains (“mixed domain” and “helical domain”),with the longest helix α1 being shared between themand providing a platform for building the twostructural domains (Fig. 2a). The first domain(mixed domain) consists of a central three-strandedantiparallel β-sheet (β1↑–β3↓–β2↑) that covers theN-terminal half of the longest helix α1. A long N-terminal loop covers the other face of the central β-sheet. The two β-strands β2 and β3 are inserted
between helicesα2 andα3. The other domain (helicaldomain) is a four-helix bundle formed by the helicesα1 (the C-terminal half), α2, α3, and α4. Thehydrophobic core of the bundle is formed by Val82,Leu90, Ile117, Leu152, Leu156, Val169, and Val173.The four-helix bundle is a common structural motifthat occurs in many structural contexts and inproteins that are functionally diverse.14
Structure of the Tipα dimer
The crystallographic asymmetric unit containstwo monomers (chains A and B) of H. pylori Tipα.They show essentially the same conformation.Between chains A and B, the root-mean-square (r.m.s.) deviation is 0.56 Å for 148 Cα atom pairs and amaximum Cα deviation of 3.2 Å occurs at Ser49. Thetwo monomers of Tipα in the asymmetric unit arerelated by a 2-fold non-crystallographic symmetry,and they form a tight dimer (Fig. 2b). The dimerformation buries an accessible surface area of1560 Å2. There are no other possible dimers in thecrystal. Our dynamic light-scattering measurementwas consistent with the existence of recombinantTipα as a dimer in the solution.The Tipα dimer in the asymmetric unit resembles a
twisted doughnut (Fig. 2c), with dimensions of52 Å×46 Å×25 Å. Contacts between the monomers
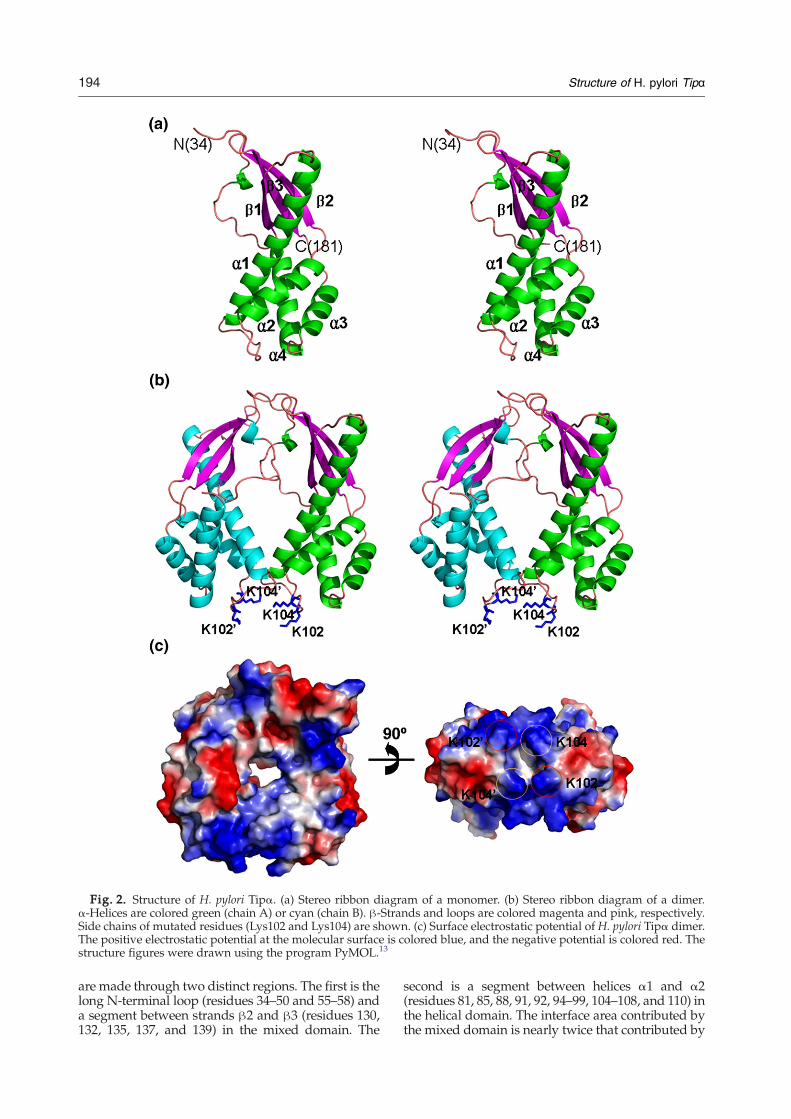
Fig. 2. Structure of H. pylori Tipα. (a) Stereo ribbon diagram of a monomer. (b) Stereo ribbon diagram of a dimer.α-Helices are colored green (chain A) or cyan (chain B). β-Strands and loops are colored magenta and pink, respectively.Side chains of mutated residues (Lys102 and Lys104) are shown. (c) Surface electrostatic potential of H. pylori Tipα dimer.The positive electrostatic potential at the molecular surface is colored blue, and the negative potential is colored red. Thestructure figures were drawn using the program PyMOL.13
194 Structure of H. pylori Tipα
are made through two distinct regions. The first is thelong N-terminal loop (residues 34–50 and 55–58) anda segment between strands β2 and β3 (residues 130,132, 135, 137, and 139) in the mixed domain. The
second is a segment between helices α1 and α2(residues 81, 85, 88, 91, 92, 94–99, 104–108, and 110) inthe helical domain. The interface area contributed bythe mixed domain is nearly twice that contributed by

Fig. 3. Electrophoretic mobility shift assay for dsDNAbinding by the mature Tipα (21–192). Detailed reactionconditions are described in Materials and Methods. Lane 1,200 bp dsDNA ladder marker; lane 2, linear dsDNA (a 545-bp PCR product; 420 ng); lane 3, wild-type Tipα (21–192)(5.6 μg); lane 4, wild-type Tipα (11.1 μg); lane 5, K102A/K104A mutant (5.6 μg); lane 6, K102A/K104A mutant(11.1 μg). Binding of the wild-type Tipα to dsDNA resultedin a dose-dependent decrease of the dsDNA band intensity,whereas the double mutant did not show such a decrease.
195Structure of H. pylori Tipα
the helical domain. This mode of dimerization resultsin a small hole of ∼8 Å in diameter (Fig. 2c). If the N-terminal residues absent in our Tipα structure areassumed to be extended and to point toward eachother in the bulk solvent, either Cys25 or Cys27 orboth could form one or two interchain disulfidebonds. It is likely that the resulting dimer structure ofTipα (residues 21–192) would not significantlydeviate from that of del-Tipα (residues 28–192).Gel electrophoresis showed that the recombinant
del-Tipα protein was monomeric without any influ-ence of dithiothreitol (DTT), while the recombinantTipα was a dimer in the absence of DTT and amonomer with DTT.6 This discrepancy between ourobservation of a stable dimer of del-Tipα in the crystaland the reportedmonomer of del-Tipα could be due tomuchdifferent protein concentrations used indifferentexperiments. Dimers of Tipα as well as del-Tipα thatexist at highprotein concentrationsmaydissociate intomonomers at very low protein concentrations, if theyare not stabilized by a disulfide bond. Thus, theinterchain disulfide bond(s) formed by Cys25 andCys27 may be necessary to prevent dissociation of thesecreted Tipα dimer into monomers.
Structural similarity searches
A search for structural homologs of H. pylori Tipαmonomer in the Protein Data Bank (PDB) using theDALI server15 revealed that there is no highly similarstructure. The highest Z-score was only 5.7; it wasobtained with the N-terminal domain (NTD) of Nab2(residues 1–97) (PDB code: 2V75).16 The sequenceidentity between them is only 7%, and the r.m.s.deviation is 3.0 Å for 71 Cα atoms in the α1–α2 andα3–α4 regions of the Tipα helical domain and theα1–α4 region of Nab2 NTD. The arrangement of fourα-helices in the helical domain of Tipα is similar tothat of the first four α-helices in Nab2 NTD.16 Nab2NTD is largely helical with five α-helices and has noβ-strand. Compared with the helical domain of Tipα,an extra fifth C-terminal helix α5 lies approximatelyantiparallel with the N-terminal helix α1 in Nab2NTD. The helical fold of Nab2 NTD is similar to thePWI fold found in several other RNA-bindingproteins.16 Other PWI fold proteins such as the U4/U6 small nuclear ribonucleoprotein Prp3 (PDB code:1X4Q) and SRm160-PWI motif (PDB code: 1MP1) arealso mostly helical.17 Nab2 NTD mediates protein–protein interactions, and there is no evidence thatNab2 NTD binds to nucleic acids.16 In contrast,Srm160-PWI motif binds to double-stranded (ds)and single-stranded (ss) RNA/DNA.17 Furthermore,Nab2 NTD, Prp3, and SRm160-PWI motif are allmonomeric. SRm160-PWI motif contains the char-acteristic Pro-Trp-Ile sequence in helix H1,17 whereasNab2 NTD and H. pylori Tipα lack such a sequence.All of these taken together indicate that the distantstructural resemblance between the helical domain ofTipα and the PWI fold proteins such as Nab2 NTD isnot likely to be functionally relevant. Therefore, ourstructure of H. pylori Tipα appears to represent thefirst structure of a novel DNA-binding protein family.
During the revision of the manuscript, the crystalstructures of a truncated form of Tipα (TipαN34) intwo different crystal forms were reported.18 It hasbeen suggested that crystal form I dimer at pH 4 islikely a crystallization artifact and that TipαN34dimer observed in crystal from II at pH 8.5 is morelikely to be relevant in solution. The latter structure ishighly similar to our dimer structure obtained at near-neutral pH.
Insights into DNA-binding activity
The Tipα dimer is toroidal, with the C-terminal part(residues 85–92) of helix α1 and the loop region(residues 46–52) immediately before strand β1 con-tributing to form a hole of ∼8 Å in diameter (Fig. 2c).Central holes in a number of toroidal proteinsinvolved in DNA metabolism have a diameter of∼20–25 Å to hold dsDNA.19 In contrast, the centralhole of the Tipα dimer is too narrow to encircledsDNA. The surface electrostatic potential of theTipα dimer suggests that a positively chargedsurface region at the bottom of the Tipα dimer inFig. 2c, formed across two monomers by the loopbetween helices α1 and α2 (residues 96–106), couldbe important in DNA binding. To assess the role ofthis region in DNA binding, we have mutated twopositively charged residues Lys102 and Lys104 intoalanine. Electrophoretic mobility shift assay indi-cated that the dsDNA-binding ability of the doublemutant (K102A/K104A) was considerably dimin-ished compared to the wild-type Tipα (Fig. 3). Thisresult supports our proposed DNA-binding site.In summary, we have determined the crystal
structure of H. pylori Tipα (residues 28–192). It existsas a dimer in the crystal, and its dimeric structurerepresents a novel scaffold for DNA binding.

196 Structure of H. pylori Tipα
Recently, a small-molecule inhibitor of nucleophos-min oligomerization was found to inhibit prolifera-tion of cancer cells.20 The structural informationreported in this study can provide a starting pointfor discovering small-molecule inhibitors that blockhomodimerization of H. pylori Tipα and, thus,modulate its biological activity.
‡http://www.rcsb.org/pdb
Materials and Methods
Protein expression and purification
The H. pylori Tipα gene (HP0596) encoding the N-terminally truncated form (residues 28–192) was PCR-amplified and cloned into the expression vector pET-28b(+) (Novagen). This construct of the recombinant proteincontains a hexa-histidine-containing 21-residue extrasequence (MGSSHHHHHH SSGLVPRGSH M) in front ofthe starting residue Pro28. It is identical with del-Tipα.6,11
The recombinant protein substituted with SeMet wasoverexpressed in Escherichia coli Rosetta2(DE3)pLysS cellsusing M9 culture medium supplemented with SeMet.Protein expression was induced by 0.5 mM isopropyl 1-thio-β-D-galactopyranoside, and the cells were incubatedfor an additional 24 h at 30 °C after growth to the mid-logphase at 37 °C. The cells were lysed by sonication in a lysisbuffer [50 mM Tris–HCl at pH 7.9, 500 mMNaCl, and 10%(v/v) glycerol] containing 50 mM imidazole. The crudelysate was centrifuged at ∼36,000g for 60 min. Thesupernatant was applied to an affinity chromatographycolumn of HiTrap Chelating HP (GE Healthcare). Theprotein was elutedwith the lysis buffer containing 500mMimidazole, and the eluted sample was diluted 10-fold withbuffer A (20mM sodium phosphate at pH 6.0). The dilutedsample was applied to a HiTrap SP HP column (5 ml) (GEHealthcare), previously equilibrated with buffer A. Uponeluting with a gradient of NaCl in the same buffer, Tipαwas eluted at 360−480 mM NaCl concentration. The nextstep was gel filtration on a HiLoad XK-16 Superdex 200prep-grade column (GE Healthcare), previously equili-brated with 20 mM citric acid at pH 4.0 and 200 mMNaCl.The eluted protein was concentrated to 11 mg ml−1 forcrystallization using an Amicon Ultra-15 centrifugal filterunit (Millipore). Dynamic light-scattering experimentswere performed on a DynaPro-801 instrument (Wyatt) at∼1 mg ml−1 protein dissolved in the same buffer used ingel filtration on the Superdex 200 column.
Mutagenesis and electrophoretic mobility shift assay
The mature Tipα (residues 21–192), containing Cys25and Cys27, was mutated using the QuikChange Site-Directed Mutagenesis Kit (Stratagene), and the mutationswere confirmed by DNA sequencing. The K102A/K104Adouble mutant was expressed and purified under theconditions identical with those for the wild type. Thedouble mutant showed a similar elution profile as the wildtype upon gel filtration. SDS-PAGE analysis indicated thatthe double mutant as well as the wild-type Tipα migratedas a dimer in the absence of β-mercaptoethanol but waspresent as a monomer in the presence of β-mercaptoetha-nol.Wemixed 1.5 or 3.0 μl of Tipα (3.7 μg μl−1) and 3.0 μl ofa linear dsDNAwith 545 bp (140 ng μl−1), corresponding tothe molar ratio of 10:1 or 20:1 for Tipα dimer to dsDNA.The reaction mixtures were incubated for 20 min at roomtemperature. Binding of Tipα to dsDNAwas analyzed by
0.5% (w/v) agarose gel electrophoresis in 1× TAE buffer(45 mM Tris–acetate at pH 8.3 and 1 mM ethylenediami-netetraacetic acid). DNA bands were visualized by ethi-dium bromide staining.
Crystallization and data collection
Crystals were grown by hanging-drop vapor diffusionmethod by mixing equal volumes of the protein solutionand the reservoir solution [100 mM Tris–HCl at pH 7.0,200 mM calcium acetate, and 20% (w/v) polyethyleneglycol 3000]. They were transferred to a cryoprotectantsolution, which contained 15% (v/v) glycerol in thereservoir solution. Two sets (SeMet I and SeMet II) ofMAD data were collected at 100 K on an ADSC Quantum315 detector at the BL-5A experimental station of PhotonFactory (PF), Japan, and on an ADSC Quantum 210 CCDdetector (Area Detector Systems Corporation, Poway, CA)at the BL-4A experimental station of Pohang Light Source(PLS), Korea, respectively. The raw data were processedand scaled using the program suite HKL2000.21 Thecrystals belong to the space group P212121, with unit cellparameters of a=53.52 Å, b=67.11 Å, and c=91.26 Å forthe SeMet II (remote) data. Two monomers of Tipα arepresent in the asymmetric unit, yielding a VM of 1.93 Å3/Da and a solvent content of 36.4%.
Structure solution and refinement
Of the 14 expected selenium atoms of the twomonomers in each crystallographic asymmetric unit, 10were locatedwith the program SOLVE,22 and the seleniumsites were used to calculate and improve the phases withRESOLVE.23 The resulting electron density map wasinterpreted by the automatic model building programRESOLVE, which generated an initial model thataccounted for ∼37% of the residues in the polypeptidechain with much of the sequence assigned. Subsequentmanual model building was conducted using the programCoot.24 The model was refined with the programREFMAC,25 including the bulk solvent correction. Tenpercent of the data were randomly set aside as the testdata for the calculation of Rfree.26 The program PRO-CHECK was used to assess the stereochemistry and toassign the secondary-structure elements.27
Accession code
Atomic coordinates and structure factors of H. pyloriTipα have been deposited in the PDB‡ under accessioncode 3GIO.
Acknowledgements
We thank the beamline staffs at Pohang LightSource, Korea (BL-4A and BL-6C), and PhotonFactory, Japan (BL-5A and BL-6A), for assistanceduring X-ray diffraction experiments. This work wassupported by Korea Ministry of Education, Science,and Technology; National Research Foundation of

197Structure of H. pylori Tipα
Korea; Innovative Drug Research Center for Meta-bolic and Inflammatory Disease (grant no. R11-2007-107-00000-0); Basic Science Outstanding ScholarsProgram; and World-Class University Program(grant no. 305-20080089) to S.W.S. B.I.L. was sup-ported by a Korea National Cancer Center researchgrant (no. 0810183).
References1. IARC Working Group on the Evaluation of Carcino-
genic Risks to Humans. Infection with Helicobacterpylori. (1994). Schistosomes, liver flukes and Helicobac-ter pylori. In IARC Monographs on the Evaluation ofCarcinogenic Risk to Humans (IRAC, ed) vol. 61, pp. 177–240. IRAC, Lyon, France.
2. Peek, R. M., Jr & Blaser, M. J. (2002). Helicobacter pyloriand gastrointestinal tract adenocarcinomas. Nat. Rev.Cancer, 2, 28–37.
3. Moore, R. J., Owens, D. M., Stamp, G., Arnott, C.,Burke, F., East, N. et al. (1999). Mice deficient in tumornecrosis factor-α are resistant to skin carcinogenesis.Nat. Med. 5, 828–831.
4. Suganuma, M., Okabe, S., Marino, M. W., Sakai, A.,Sueoka, E. & Fujiki, H. (1999). Essential role of tumornecrosis factor α (TNF-α) in tumor promotion asrevealed by TNF-α-deficient mice. Cancer Res. 59,4516–4518.
5. Suganuma, M., Yamaguchi, K., Ono, Y., Matsumoto,H., Hayashi, T., Ogawa, T. et al. (2008). TNF-α-inducing protein, a carcinogenic factor secreted fromH. pylori, enters gastric cancer cells. Int. J. Cancer, 123,117–122.
6. Suganuma, M., Kurusu, M., Suzuki, K., Nishizono, A.,Murakami, K., Fujioka, T. & Fujiki, H. (2005). Newtumor necrosis factor-α-inducing protein releasedfrom Helicobacter pylori for gastric cancer progression.J. Cancer Res. Clin. Oncol. 131, 305–313.
7. Godlewska, R., Pawlowski, M., Dzwonek, A., Mikula,M., Ostrowski, J., Drela, N. & Jagusztyn-Krynicka,E. K. (2008). Tip-α (hp0596 gene product) is a highlyimmunogenic Helicobacter pylori protein involved incolonization of mouse gastric mucosa. Curr. Microbiol.56, 279–286.
8. Yoshida, M., Wakatsuki, Y., Kobayashi, Y., Itoh, T.,Murakami, K., Mizoguchi, A. et al. (1999). Cloning andcharacterization of a novel membrane-associatedantigenic protein of Helicobacter pylori. Infect. Immun.67, 286–293.
9. Kuzuhara, T., Suganuma, M., Kurusu, M. & Fujiki, H.(2007). Helicobacter pylori-secreting protein Tipα is apotent inducer of chemokine gene expressions instomach cancer cells. J. Cancer Res. Clin. Oncol. 133,287–296.
10. Suganuma, M., Kuzuhara, T., Yamaguchi, K. & Fujiki,H. (2006). Carcinogenic role of tumor necrosis factor-αinducing protein of Helicobacter pylori in humanstomach. J. Biochem. Mol. Biol. 39, 1–8.
11. Kuzuhara, T., Suganuma, M., Oka, K. & Fujiki, H.(2007). DNA-binding activity of TNF-α inducing
protein from Helicobacter pylori. Biochem. Biophys. Res.Commun. 362, 805–810.
12. Kuzuhara, T., Suganuma, M., Tsuge, H. & Fujiki, H.(2005). Presence of a motif conserved betweenHelicobacter pylori TNF-α inducing protein (Tipα)and penicillin-binding proteins. Biol. Pharm. Bull. 28,2133–2137.
13. DeLano, W. L. (2002). The PyMOL Molecular Gra-phics System DeLano Scientific, Palo Alto, CA.
14. Kamtekar, S. & Hecht, M. H. (1995). Protein motifs. 7.The four-helix bundle: what determines a fold? FASEBJ. 9, 1013–1022.
15. Holm, L. & Sander, C. (1995). Dali: a network tool forprotein structure comparison. Trends Biochem. Sci. 20,478–480.
16. Grant, R. P., Marshall, N. J., Yang, J. C., Fasken, M. B.,Kelly, S. M., Harreman, M. T. et al. (2008). Structure ofthe N-terminal Mlp1-binding domain of the Sacchar-omyces cerevisiaemRNA-binding protein, Nab2. J. Mol.Biol. 376, 1048–1059.
17. Szymczyna, B. R., Bowman, J., McCracken, S., Pineda-Lucena, A., Lu, Y., Cox, B. et al. (2003). Structure andfunction of the PWI motif: a novel nucleic acid-binding domain that facilitates pre-mRNA processing.Genes Dev. 17, 461–475.
18. Tosi, T., Cioci, G., Jouravleva, K., Dian, C. &Terradot, L. (2009). Structures of the tumor necrosisfactor α inducing protein Tipα: a novel virulencefactor from Helicobacter pylori. FEBS Lett. 583,1581–1585.
19. Hingorani, M. M. & O'Donnell, M. (2000). A tale oftoroids in DNAmetabolism.Nat. Rev., Mol. Cell Biol. 1,22–30.
20. Qi, W., Shakalya, K., Stejskal, A., Goldman, A., Beeck,S., Cooke, L. & Mahadevan, D. (2008). NSC348884, anucleophosmin inhibitor disrupts oligomer formationand induces apoptosis in human cancer cells.Oncogene,27, 4210–4220.
21. Otwinowski, Z. & Minor, W. (1997). Processing of X-ray diffraction data collected in oscillation mode.Methods Enzymol. 276, 307–326.
22. Terwilliger, T. C. & Berendzen, J. (1999). AutomatedMAD and MIR structure solution. Acta Crystallogr.,Sect. D: Biol. Crystallogr. 55, 849–861.
23. Terwilliger, T. C. (2003). Automated main-chainmodel building by template matching and iterativefragment extension. Acta Crystallogr., Sect. D: Biol.Crystallogr. 59, 38–44.
24. Emsley, P. & Cowtan, K. (2004). Coot: model-buildingtools for molecular graphics. Acta Crystallogr., Sect. D:Biol. Crystallogr. 60, 2126–2132.
25. Murshudov, G. N., Vagin, A. A. & Dodson, E. J. (1997).Refinement of macromolecular structures by themaximum-likelihood method. Acta Crystallogr., Sect.D: Biol. Crystallogr. 53, 240–255.
26. Brünger, A. T. (1992). The free R-value: a novelstatistical quantity for assessing the accuracy of crystalstructures. Nature, 355, 472–474.
27. Laskowski, R. A., MacArthur, M. W., Moss, D. S. &Thornton, J. M. (1993). PROCHECK: a program tocheck the stereochemical quality of protein structures.J. Appl. Crystallogr. 26, 283–291.




















