
CSU GE-Breadth Certification Courses List (All CSU GE-Breadth ...
Upload
khangminh22Category
view
1download
0
CROSSABILITY, GE-NOME RELATIONSHIPS AND INHERITANCE STUDIES IN INTERGENERIC
HYBRIDS OF PIGEONPEA
A THESIS SUBMITTED FOR THE DEGREE OF
Doctor of Philosophy
BY
P: SATEESH KUMAR, M-sc. ( A Q
SCHOOL OF LIFE SCIENCES UNIVERSITY OF HYDERABAD
HYDERABAD - 600 134 INDIA

DECLARATION OF ORIGINALITY
T h i s t h e s i s r e p o r t s t h e o r i g i n a l work of t h e a u t h o r ,
excep t a s o the rwise s t a t e d . I t has not been submit-
t e d p r e v i o u s l y f o r a degree a t any U n i v e r s i t y .
P. Sa teesh Kumar

C E R T I F I C A T E
T h i s i s t o c e r t i f y t h a t t h e T h e s i s e n t i t l e d
"CROSSABILITY, GENOME RELATIONSHIPS AND INHERITANCE
STUDIES I N INTERGEhERIC HYBRIDS OF PIGEONPEA" i s
b a s e d o n t h e r e s u l t s o f t h e w o r k c a r r i e d o u t b y
M r . P . S a t e e s h Kumar, M . S c . ( A g . ) , f o r t h e d e g r e e
o f DOCTOR OF PHILOSOPHY u n d e r my s u p e r v i s i o n . T h i s
w o r k has n o t b e e n s u b m i t t e d t o a n y d e g r e e o r d i p l o m a
o f a n y o t h e r U n i v e r s i t y .
31 January 1985
DR. N.C. SUBRAHMANYAM (SUPERV I SOR School o f L i f e Sciences U n i v e r s i t y of Hyderabad Hyderabad-500134, I n d i a

I wish to express my gratitude to Dr. N.C. Subrahmanyam, University of Hyderabad and Dr. Donald G. Faris, ICRISAT for their encouragement and advice during the course of this investigation. I am indebted to than for the time and efforts spent on ny behalf.
I wish to express my thanks to Dr. J.P. Moss, ICRISQ. for keeping at my disposal the facilities in the groundnut cytogenetics laboratory at I W W .
I thank the Director of Research, IClUW; Program Leader (Pulses) ,. ICRIW and the Dean, School of Life Sciences, University of Xyderabad for providing the necessary research facilities.
My + M s are due to Drs. L.J. Reddy, D. Shanm, A.K. Singh, R.P.S. Pundir, K.B. Saxena and D.C. Sastri of ICFUSAT for helpful suggestions during the course of this investigation. I am also thankful to Dr. Ian Dudas for helpful discussions during the final stages of preparation of this dissertation.
Iwish to thank Drs. L.J.G. van der Maesen and P. Renwandan for the supply of Atylmia seed and for help on taxonomic details.
I would like to express my sincere =reciation to Mr. G. Shinde for all the help rendered during the course of this investigation.
My a~preciation is extended to the personnel of the Photolab, ICICISAT for their excellent processing and printing of the photographs presented in the deeaertation.
My amreciation is also extended to Messrs R. Vijaya Kurrrar, A. Nageshwar Rao M. Chenchi Reddy and to the field staff of the Pigeonpea Breeding Subprogram of IGUSAT and to the staff,of the Plant Culture Unit of University of Elyderabad for their uninhibited help during the course of field experiments.
I acknowledge with thanks the help rdered by ny frierds MeSSrs M.K. R q , D.V. Ranga Rao, A.N. Murthy, V.B. Reddy and P. Muralikrlshna.
I gratefully acknowledge the financial generosity of ICRIsA'r.
I would like to express my a~reciation to Mrs. K. Syamalanba for typing the dissertation.
Finally, I owe a personal debt to my late grand parents for their encou:agemt and involvement in my research efforts.

The present investigation w a s ini t iated with the brmd objectives of, assessment of genome relat ionship between the G&LUS and its related wild species, screening the wide crosses for haploids tbrough selective chromsome elimination, developnent of m W s to inprove crossability between species, studying the inheritance of descernible characters in the hybrids obtained and develop in VitEP regeneration techniques in pigeonpea. The present study involved crossing C. & a ~ with twelve v i e s of &ylcsia and two species of m. A t o t a l of 24 intergeneric cross conbinations including reciprocals were attenpted.
Eight (A. . . . r A. A. lanceolata, A.
-, A. =, A. and A. se&ed out of the 12 species hybridized with C. a when Cajarus w a s the female parent. From the reciprocal crosses a low rate of success was obtained in two conbinations (A. albicans x C.
and A. sericea x C. u. When fajmus was the female parent the degree of crossability varied not only with the
species in question, but also with the genotype. Che of the'- cultivars, ICP-102 failed t o cross with any of the species. The ra te of success w a s very high (1.37-19.5%) when A. lineata w a s the male parent followed by C. a x A. albicans crosses (1.09-9.6%). In the other six anbinations the success was low ranging from 0.9-3.05%. C.
cultivars which had a c o m n female parent in their pedigree did not vary mch in their abi l i ty t o cross with
species (R. cothii and R.
Post-pllinatiion homne treatments were used t o inprove the crossability in successful C&rus-&yl~~L~ crosses and also t o assess their influence in developing new hybrid con-binatiops. Effects of gibberellic acid (GA31, kinetin and their mixture on S&mw, x crossability. were tested. GIU and kinetin were used independently a t concentrations ranging from 10 ppn to 80 ppn or as a 1:l mixture keeping the net homne concentration in the same range. Following hormone treatments the rate of success w a s increased through increased pod sett ing and n&r of seeddpod. GA3 a t a concentration of 50-60 ppa was found t o be the m s t effective treatment. Higher hormone concentrations had a detrimental effect. In unsuccessful crosses homne treatments delayed bud drop by 3-4 days, permitting prolonged ovule developent. Resu l t s indicate tha t the hormone treatment helps in post-fertilization dwelopnent leading t o an increased rate of success. Results suggest that the Auetralian species of Atyhsh are more diverged fran Cajanua than the Asian & y l d a .
A l l the proqeny were subjected t o a detailed cytological andlysie. There waa no evidence of chromaom elimination in any of the m & y l Q a i B crosses.

Cytolcgical studies i n pigeonpa unequivocally established the p r s e n c e of tw nucleolar organizers per genome in S&r.u and A!&!~Q& species. Detailed meiotic s tudies in the hybriris revealed a high degree of gemme homlogy and recorrbinati~n between the a i l t iva ted and the wild species. In the hybrids eleven bivalents were ccmrcn a t metaphase-I with o c c a s i o ~ a l univalents. Univalents were m r e fresuent i n t h e hvbrids with the Australian &yl&ai (A. arandifoliaand8. latlsecala). C h i a m freauencv w a s also low i n ' these hvbrids in comrarison with the ohers: Anaphase separation was ;egular in mst 'of the hybrids again with the exception of the hybrids w i t h A. arandifolia and A. latisepala. In both these cases m u l t i ~ o l a r spindle abnormalities were ammn. Inversion loops a t pachytene and anaphase I bridges were found i n hybrids of C. s&n with A. arandifolia indicating inversion heterozygosity in t h e F, hybrids.
W r i d s between C. +an and A. were t h e most f e r t i l e while the hybrids wrth A. arandifolia and A. l m were highly s t e r i l e . Other hybrids exhibited a considerable arount of s t e r i l i t y in s p i t e of regular bivalaent formation a t metaphaseI and normal disjunction. The genotype of the f m i e parent a l so influenced the pollen f e r t i l i t y . Hybrids with the genotype, 18-7035 were consistently l e s s f e r t i l e in all c o ~ m a t 1 0 n s .
Nucleolar var iat ion in n h r , s i z e and dis tr ibut ion was recorded a f te r division-I and division-I1 i n three hybrids (C. f&dU.x A. m r C. U X A. lUEh and C. X A. $uuL&lS. I n these three hybrids three t o four brvalents were found attached t o the diplotene/diakinesis nucleolus as against one t o two bivalents i n t h e parents. Variation i n nucleolar nrrmber and dis tr ibut ion a r e interpreted to have originated from pairing and reconbination between nucleolar organizer chromosome(s) of one parental species with t h e nonnucleolar o r g h i z e r chromosme(s) of the other. Detailed analysis of these hybrids provided evidence for "crypt ic s t ruc tura l hybridityn and f o r intergenomic (allosyncetic) rearbinat ion. Nucleolar behaviour was used as a mrker t o t r a c e t h e products of recodinat ion in these hybrids. The nuc lwlar variat ion in the hybrids suggests t h a t t h e s t e r i l i t y i n t h e hybrids inspi te of regular meiosis is a consequence of duplicatirma and/or deficiencies originating from allosyndetic reambination part icular ly between t h e s t ruc tura l ly a l t e red chromsomes i n t h e hybrids.
Most of the &yJaaia characters l i k e seed strophiole, seed mottling, and pod hairiness were expressed in t h e hybrids indicating the i r daninant nature while l e a f l e t shape and twining nature shared incorrplete dominance. The F2 segregation f o r l e a f l e t &ape and pod hairiness rwealed that these t r a i t s a r e governed by one locus while twining nature, seed strophiole and seed nott l ing were governed by tuo loci . Substantial var iat ion for quantitative t r a i t s was found in the F2 generation which include8 transgressive segreganta f o r leaf l e r r g t h r leaf width, pod length, seed w e i g h t and protein content in the

hybrids, underlining the iap6rtance of Atylsda species in tk iaprovmt of fajanua. The F2 variation in these hybrids sbtantiated the cytological ,evidence for a close relationship between c. and A. -.
Techniq~es were standardized for regeneration of plants from immture ecjcryos (&ryos at least 11 &ys.oldl of C. ,@ and from wtyl&s of C. &+n, A. -, A. albuxxs and A. -. Genotypic differences were wident with respect to frequency of regeneration in both arbryo and cotyledon cultures. In cotyledcr. cultures the regeneration potential varied with the region Of ti.e cotyledonary explant with nodal halves being mre effective bi-ile in errbryo cultures an age dependent response was wident .

C O N T E N T S
LIST OF TABLES
LIST OF FIGURES
1. SCOPE OF THE INVESTIGATION
2. MATERIAL AND METHODS
3. SPECIES CROSSABILITY AND HYBRID MORPHOLOGY
3.1 . RESULTS
3.1.1 CROSSABILI'PI
3.1.2 EFFECT OF HORMONE TREATMENTS
3.1.3 HYBRID MORPHOLOGY
3.2 CiSCUSSION
4. MEIOTIC BEHAVIOUR
4.1 PARENTS
4.1.1 RESULTS
4.1.2 DISCUSSION
4.2 HYBRIDS
4.2.1 RESULTS
4.2.1.1. NUCLEOLAR DISTRIBUTION AFTER TELOPHASE-I AND I1 IN F1 HYBRIDS
4.2.1.2 CHROMOSOME BEHAVIOUR IN F2 POPULATIONS
4.2.2 DISCUSSION
4.2.2.1 F2 GENE~TION
5. INHERITANCE OF QUALITATIVE TRAITS
5.1 RESULTS
5.2 DISCUSSION
6. VARIATION FOR QUANTITATIVE TRAITS
6.1 RESULTS
6.2. DISCUSSION

Page
7. IN VITRO REGENERATION
7.1 RESULTS
7.1.1 TISSUE CULTURE
7.1.2 EMBRYO CULTURE
7.1.3 ANTHER CULTURE
7.2 ' DISCUSSION
8. GENERAL DISCUSSION AND CONCLUSION
REFERENCES
PUBLICATIONS FROM THE THESIS

LISP OF TABLES
PAGE
12 Table I. Studies on Intergeneric hybridization in pigeonpea.
Table 11. Origin and salient economic features of the species used in the study.
Table
Tab1 e
111. Seed germinability in E2 ~opllations.
IV. Constituents of basal mdia used in In . regeneration studies.
Table V. Crossability of and Rhyn&&a species with five genotypes of Caidwa dm-
Table VI. Crossability of species with C&mus a having a c o m n £ a l e background.
Table VII. Percent pod-set in intergeneric crosses of S & u s &.an and &y&iia species followed by horrrone treatments.
VIII. Expression of qualitative traits in parental species and F1 hybrids between faiarucajan and ALyhski species.
Table
Table IX. Expression of quantitative traits in pareptal species and F1 hybrids between S & b sajm and ALyLxla species.
Table X. Expression of quantitative traits in parental species acd F1 hybrids between S&nw and ALyhski species.
XI. Meta-1 configurations and pollen fertility in fajmu c&in and species.
XII. Metaphase-I configurations, Andphase1 abnodities, pcllen sterility and ovule fertility in El hybrids betwea C. s&n and different species of &yJ.g&.
Table
Table XIII. Nucleolar variation at telophase-I1 in a hybrid between C. and
Table XIV. Inheritance 1; ttern of seed strophlole in C. x ~ & & a a species nybrlds.

Table XV. Inheritance pattern of seed mottles in C. x species hybrids.
Table XVI. Inheritance pattern of pod hairiness in C. s.ajdn x A t y h i a species hybrids.
Table XVII . Inheritance pattern of twining nature i n C. mjan x At&& species hybrids.
lrPble XVIII. Inheritance pattern of lea£let shape in C. x &&ski species hybrids.
Table XIX. Variation for pod length (an) in the F2 population of hybrids between C. and &&sia species.
Table XX. Variation for 100-seed weight (g) in the F2 population of hybrids between C. and ALy&ia species.
W l e XXI. Variation for seeds per pod i n the F2 population of hybrids between C. siim and &ylgsia species.
l ab le XXII . Variation for mid-leaf width (an) in the F2 population of hybrids between C. 0 and A t y l d a species.
m l e XXIII. Variation for mid-leaf length (an) i n the F2 p p l a t i o n of hybrids between C. and Atyhaia species.
W l e XXN. Percent shoot regeneration from wtyleQn cultufes of different cul t ivars of and "qecies of A&&& on IS medium sqpleinented with 2,4-D (0.5 mg/l) and BR (2 nq/l).
Table XXV. Percent plantlet regeneration and shoot rweneration from cotyledon cultures of different f&nus culti&rs and species of &&.sia on MS Wum supplenented with 2r4-D (0.5 41) BA(2 na/l) and NA?i (1 Iq/l).
TAble XXVI. Percent plantlet r w v e r y i n d r y 0 c ~ l t u r e s of pigeonpea on MS and media s;;~plemented with 1 rg/l of 2,4-D.
PAGE
107
109
110
110
116

Fig. 1. Effect of gibberellic acid andlor kinetin on percent pod and seed set in four ~-~ crosses.
Fig. 2. Effect of gibberellic acid andlor kinetin on delaying bud abscission.
a. Caianusrojan as fanale parent.
b. Ca.ianus a as. male parent. Fig. 3. ltJigs and leaflets of parents and hybrids.
Fig. 4. Twigs and leaflets of parents and hybrids.
Fig. 5. Variation for leaf shape and ultrastructure in C. X 8. hybrids.
Fig. 6. Meiosis in C. sajm cv Pant A2 and I I E t ~ ~ in cultivars of C. &.
Fig. 7. Meiosis in Aty.&ia species.
Fig. 8. Meiosis in At&&& species.
Fig. 9. Nucleolar distribution in pollen nother cells of C Esljan and species of A t y h b .
Fig. 10. Frequency distribution of nucleoli at telo-JI in pollen mpther cells of C, cajm and & y h h species.
Fig. 11. Meiosis in hybrids between C. EBian and A. abisals.
Fig. 12. Meiosis fn hybrids between C. &an and A. serieea and C. &an and A. s&&Ua.
Fig. 13. Meiosis in hybrids between Cr &an and A. scarabaeoides and C* Enjan and A* line9ta.
PAGE
33
Fig. 14. Meiosis in hybrids betwen C. raipn a d A. arandifolia*

Fig. 15. k i o s i s in hybrids between c. & and A. lahsala.
Fig. 16. Nucleolar variation in pollen mother cells of hybrids between C. &an and A. il'blcans.
Fig. 17. Nucleolar variation in pollen mother cel ls of hybrids between C. and A. l h a b and C. 0 and A. gmnd&L~.
Fig. 18. Frequency distribution of nucleoli a t telo-11 in daughter nuclei of hybrids
' between C. sajm and .&yl&.a species.
Fig. 19. Frequency distribution of F2 segregants i n different fe r t i l i ty groups.
Fig. 20 ~romosome pairing a t pachytene in hybrids between C. 0 and species of A t y h b .
Fig. 21 Meiosis in tetraploid C. 0.
Fig. 22 Possible nucleolar variation i n pollen mther cel ls of the hybrids between C. and
species with out reconbination and follawing recabination between nucleolar organizing and non-nucleolar organizing chromomlres.
Fig. 23. YitrP regeneration of sajanu plants from cotyledons and h m t u r e d r o y s .
PAGE
75


Plants of greatest irrportance to agriculture belong to the
families Graminae and Legrmrtnosae. Leglnninosae amprises a wide
array of genera (600) and q e c i e s (1300) (Delwiche, 1978) and
occupies a unique place in the plant kingdom. Sweral legumes
particularly from the subfamily Papilionoidae are of great
m m i c importance next t o cereals (Cobley and Steele, 1976).
Grain legums or plses are the major source of dietary protein
in developing countries. The high protein content and relatively
low cost of these food legumes have earned than the t i t l e "Pcor
m's Meat" which effectively underlines their v i t a l importance
in developing nations. legmes have also played a major role in
patterning the agricultural s y s t m in the tropics. Their
nitrogen fixing abil i ty enables then to sustain high yields in
the face of mFnimrm inpts, inprove so i l f e r t i l i t y along with
their adaptability to diverse e n v i r o m t s .
Pigempea &an (L.) Millsp.1 the only cultivated
species in -, subtribe Cajaninae is an important pulse crop in
the tropics and ranks f i f t h emong the edible legumes. India has
the largest .area in the world under grain legume cultivation
where pigeonpea oocupies an area of 2.8 million ha. Pigeonpea is
m s t l y ansumed in the form of s p l i t p l s e (dhal) . Green pods
and seeds are used as vegetable in sotw parts of the tropics.
Seed protein content in pigeonpea averages about 23%. The
average yield of pllees on a national basis anpared t o cereals
baa been very low and th is gap continues to widen. The .green
revolution" has not increased the pulse yields because of the
aphasia on cereals. Apart from socio-economic co~siderations ir!

cultivating legumes on marginal lands without fertilizer and poor
plant protection measures, genetic factors have also contributed
to the slow pace of pigeonpea irrproveuent. In pigmnpea as with
m y other leg- the inbreeding behaviour resulted in severe
restriction on genetic variability and inprovement has therefore
been confined to selection and perpetuation of useful kinds of
gene action. Plant breeding history shows that diverse gene
p l s are the foundations for effective crop improvement
programnes. The primary objective in plant breeding is to widen
the genetic base of a cultivated species. If the needed
variation is limited as in the case of pigeonpa the options for
the breeders are:
1. Incorpration of alien vaiation,
2. Induction of nutations or
3. Exploitation of somclonal variation
The a~plicebility of nutation breeding in crop irrgrovment
i s limited since a.,vast majority of mutations are deleterious and
it still r a i n s a hit and trial method with l M t e d scope for
directed attenpts.
clonal variation is a recent technique which offers a
lot of potential for the future but it has not been possible to
exploit these variants on a field scale except in a few crop
(Scowcroft and Larkin, 1982) . Tbe otkr and possibly the most viable recourse of
introducing variation into a species is through transfer of
genetic material from one species to another by hybridization.

The genetic potential of wild relatives is widely damnstrated in
plant breeding and in evolutionary studies. Wild relatives have
helped to fill the voids in traditional breeding programres and
have helped to exploit the potential in several crops (Stalker,
1980).
The first reo~rded interspecific hybrid was made in 1717
between carnation and sweet William by Thomas Fairchild (see
Allard, 1960). Since then there has been extensive literature on
distant hybridization and the first a-made cereal, "Triticale"
was an outcome of intergeneric hybridization. Extensive studies
on distant hybridization have been made in crops like wheat
(Sears, 1972; Sears, 1975;) barley (see Bothmr and Hagberg,
1983), Maize (see MangelsdDrf, 1974; Harlan and de Wet, 1977) , Eblanurn (see Swaminathan and Magoon, 1961; Atskaitis and
Vinitskus, 19751, cotton (Blank & el, 1972; Meyer, 1973;
Meyer, 1974), Nicotiana (seesmith, 1968; Mann & f., 1963;
berbec, 1974) , . : f ~ t o (see Rick, 1982) , and rice (see Nayar, - 1973) .
Distant hybridization studies have also been carried out in
several important legumes such as, Phhsealus, Yigna, W,
E h m , Arachis (5nartt. 1979). Species in the genus E h a s s d u
have been a subject of wide interest. The possibility of gene
exchange between species has led to several studies on
interspecific hybridization especially between P. and
E. coccinwfi (lendel, 1866; Tschermak-Seysenegg, 1942;
Lwprecht, 19481 Rudorf, 1953; Kedar and -is, 1960; Thornas,
1964; Al-yasiri and Coyne, 1966; Rutger and Becknw, 19701

Sna~tt, 1970; m c h a n d , 1971; Marechal, 1971; Ibrahim and
m e , 19751 Elaq & a, 1980; Savwa, 1981; Shii & d r 1982;
Conti, 1983). Hybridization with other ph;rseolus species - including the wild forms has also received wide attention (erz,
1952; Elonma, 1956; m e , 1964; Smrtt, 1970; Braak and
Kogtra, 1975; Le Merchand & dl, 1976; Tan Boun Suy, 1979;
Mrang, 1979). Interspecific hybridization in Phaseolus in recent
years has been widely used in the irrprovement of Phaseolus sp.
with respect to disease resistance, insect resistance, nitrogen
fixation and several agronomic characters (F-li & dl, 1981;
Bmerot & dl, 1981; Zapata e,k al, 1982; Hunter eL al, 1982; - Alvarez, 1981; Lapinskas, 1980).
Wide hybridization has received a fair degree of attention
in the genus -. Studies on hybridization between G. max
and G. soja have been extensive (Karasawa, 1936; Ting, 1946;
William, 1948; Tang and Chen, 1959; Tang and Tai, 1962; Ahmed
& fir 1977 and 1979; Kiazuma stt dl, 1980; Malik and Singh,
1982: Gai gL a, 1982). Atmad & dl (1977 and 1979) made a
detailed cytolcqical analysis of these hybrids and found
structural differences between the two genome6 in the form of
inversions. There have &so been attempts at hybridization
between the wild species (Palmer, 19651 and in 1968 Palmr and
Hadley reparted a successful cross between G. tabacina and
E. latFfolia (E. tomentosa). Later Broue & al. (1979) and - Putiwsky and Broue (1979) obtained two intraspecific Md six
interspecific hybrids. Recently -mil and Eyrmitz (1983)
reprted a range of intra and interspecific hybrids in the

subgenus Glgche along with detailed cytological studies in the
hybrids. Attaopts at hybridization between cultivated m w
and the wild species were m t successful with the conventional
methods (Ladizinsky & & 1979; Hood and Allen, 1980). Newell
and Hymswitz (1982) were able to obtain a hybrid between E. mix
and P. tomentella with the aid of ovule culture. Apart fran
alding in understanding the species relationships, interspecific
hybridization in G l y s = h has also helped in widening the genetic
base for quality characters of P. max particularly protein
content (Ala &dl, 1979; Erikson and Beversdorf, 1981; Kiazuma
& al, 1980).
%e genus Y i d a exhibits dysploidy with x= 5, 6 or 7. Host
of the interspecific crosses m far involve species with
chrmsome nuhers x=6 (subspecies aativa, &d nigra) . Hybrids between 2n=10 subspecies (e have also been studied
(Mettin and Hanelt, 1973). Very little informtion is available
on crosses awng.' subspecies with the chromsome nrnober 2n=14.
Crosses betwea~ types with the same chromosome nuher result in
fertile or sanifertile hybrids and meiosis in the F1 hybrid$
indicated the presenceof chromoscinal structural hybridity as
evidenced by heteromrphic bivalents, trivalents, quadrivalents
etc (Mettin and Hanelt, 1973). Crosses involving forms with
different chramsome n h r s have also been attaqrted (Zohary and
Plitmann, 1979) and hybrids were obtained in numerous
ca7binations. Same of these hybrids erhibited hybrid vigour . Hybrid between a&bza (6) and llxdaka (rcf) shoved pentavalents
in the rl :cieratioii aid ir. F2, segregants st~~wetl diffrru.~t= in

chrcmeome nlmbers ranging from 11-14 (Nerson, 1970). W a t a n a b e
and Yamada (1958) reported crosses between aativa and
ancrustifolia in which the pollen fertility was less than 5% and
the F2 segregated alsrost exclusively for parental types.
Yamamto (1971, 1974 a,b) studied crosses between e, m&kaz&a, gihaa and and in the hybrids he found
poor meiotic pairing and consequently low fertility. He reported
a stable kasyotype produced by the substitution of one chromsome
of satFva in a karyotype derived from the
interspecific hybrid. Later x e (1975) studying the anylose
isoyme pattern in a Pilosa x -gm hybrid showd the
occurrence of reconbination In the hybrid progeny. Cubero (1982)
has sumnarized the interspecific hybridization studies in the
genus W. In general hybridzation studies in this genus have
shawn that the various chromsome types are not fully
reproductively isolated from one another, and hybridization
studies have also.: shown that hybridization and recurrent
reconbination between distinct karyotypes antributed to the
build up of chrolnosomdl polymorphism. Species crosses in this,
genus helped in the generation of variability for traits like
seed hardiness (Donnelly, 1980).
In the genus Yigna the closest relative of green and black
gram (Y. t.adiata and Y. mu& is Y. s&hkU. Hybridization
between these species is achiwed with relative ease (AIIRDC,
1977) . Eybridization studies betxeen Y. w, y. nuugrz,
y. LmS3ellulata and Y. mgubds have been numerous (Dana,
1966a,b,c; Al-yasiri and CoyneL1966; Biswas and Dana, 1975,

1976; Smrat 1973; <hen & & 1977; and Chowdhry,
1977; Md H a r m , 1977, 197&, 1978b; machado & pl,
1982; Chen & 1983) . Although Machado EL f. (1982) f ran
their cytological studies could not conclude whether the pairing
observed in the interspecific hybrid was autosyndetic or
allosyndetic, Chen & fir (1983) from their F2 studies concluded
that reemhination and gene exchange takes place between these
species. In !,?igna also the related species have been frequently
used for the transfer of cha.racters like disease resistance and
protein content (Singh, 1981; Gill & &, 1983; and Lukoki and
Otoule, 1981).
Several wild Arachin species have been successfully crossed
with the cultivated species A. &agaea. The cultivated species
crosses easily with A. mrrentina. The first report of this
cross came from Krapvickas and Rigoni (1952)' with subsequent
reports from Kumar & al (1957) and Raman (1959). Gregory & al
(1973) have produced hybrids between A. and wild .;I
diploids in the section &a&h. These hybrids have also been
reported by Mss (1977). Raman (1976) has reported hybrids of
a with A. y i J . l ~ ~ and A. duarmda. Intersectional
crosses sUCh BS Arachin . x -, Eredoidefi x
Ereceoidesx-,-XErectoides.
and x Erectoids are possible (Gregory et & 1973). In
addition Raman (1976) reported hybrids behveen A. and
A- -, A. hme&&di and A. d b s i and between
A- montimle and A. gl&r&a though the authenticity of m y of
these has been questioned. Inspite of ploidy differences the

wild species of Amdais have gained a lot of impbrtance with
respect to transfer of specific characters like resistance to
insects, viruses, nematodes, fungi, etc. (ICRIW, 1982).
Rybridization between species of Elaun (Mtiwm, elatius,
hknnik, f u h m and &sshhml has also received sane attention
(Zohary and Hopf, 19731 Gritton and Wiezbicka, 1975; Belova,
1979 and 1980). Success has also been repxted from crosses
involving Eislrm and U c h (Stegajlo, 1963; Sobolev eL a, 1968; Sabolev and Burgil, 1970) . The chromosome nlmber of these
hybrids was between 12 and 16 (2n) . Sobolev et al (1968)
reported mnhtmulogous chromsome pairing in the h*rids and the
chromosomes formed distinct groups.
Cultivated pigeonpea is bestowed with a wealth of related
wild species in the genus m. The genus is also
cansidered close to -. All the species of and
have the same basic chromomme nlmber as that of
(X=ll). Ther6 are about 34 species of Atylg&a, of which
18 are +Bnic to Lndia, 15 are natives of Australia and one
species is found in Africa (van der Maesen, pers. comn.) . The
taxanomic delimitation of fajmw and into distinct
genera which has been dbne on the basis of the seed stro@dole
has been a point of contention (Mac ChbL1975; van der Maesen,
1980). Many of the A Q l m h species possess ~ e ~ e r a l desirable
characters such as disease and pest r e s m c e , high protein
content, plmtoperiod insensitivity, drOUght tolerance
(Remmandan, 1980). Inspite of limited variability for certain
imp3rtant qualitative traits in C. Eirjan and the existence of a

wide wealth of species, studies on intergeneric
hybridization in pigeonpea are surprisingly few and far btween.
Naithani (1941) was the first to give details atout the
meiotic division of pigeonpea although it was M y (1933) vtso for
the first tine reported the chromsome n-r of cajanus as n-11
which was donfirmed by Akinola & al. (1972). Krishnaswamy and
Ayyangar (1935) provided information about meiotic pairing and
&aim frequencies in pigeonpea and this was followed by the
studies of Kumar & al (1945, 1966) , Bhattachax jet? (1956) and
Reddy and De (1983) all of which mentioned normal &om-
pairing and regular formation of eleven bivalents at meiosis.
The karyotype description of Naithani (1941) showed no
variation for chromsome morphology between variet'ies. This
attempt was followed by that of w i k a r and Tbakur (1956) who
gave the total conplemnt length as 75.4um.
These initial attenpts were followed by the works of
Shrivastava & al (1973) and Sinha and Kumar (1979) wlw reported
significant varietal variation for a m ratios and &orno&
lengths and by those of Sharrna and Gupta (1982) and Pundir
(1981).
There have been two publications on pachytene karyotyp
analysis (Reddy, 1981a and Dundaa ek al 1983). While Reddy
identified the pachytene chronosomes by the length and centramere
positions, Dundas & al (1983) used chronmmere patterns for
chromsore i6e~t i f ication. Only recently the C i e ~ ~ a C banding
techniaue was attempted for Riseonpea without much success

(mania and Lavania, 1982).
The satellite chrotmsome n h r s in pigeonpa have been a
subject of controversy and from the literature one can see that
thenmber varies from0 to 2. Sinha and Kcmvlr (1979) a d
Shrivastava s.t dl (1973) reported that some cultivars used in
their studies had one Satellite (SAT) chromsoxre while others
have none. The karyotypes presented by Shanm and Gupta (1982)
do not show any SAT chromsome while those of Pundir (1981)
showed t w . ArrPng the species of Atvlosia. A. lineata was used
extensively by Kunrar & dl (19581, Sikder and De (1967) and Reddy
and De (1983) in their studies on meiotic pairing. K u t m r & &.
(19581, Deodikar and Thakur (19561, Sikder and De (1967)
Shrivastava & (1973) and Pundir (1981) studied the somatic
karyotype of A. lineata and reported a close relationship between
the karyotypes of Gujaws and mineatn. Sikder and De
(1967), Reddy (1973) and Pundir (1981) studied the karyotypes of
A. lineata. A. a and A. ecarabaeoides. The karyotypes of.
these species were also presented by Reddy (1981 a,b,c) based on
pachytene chromsorne analysis. The other species for
which chrcmsome nmbers were'reported were A. (Bir
and K m i , 1973, 1977; k d i r and Singh, 1981); A. mgaa
( S a n j e and Satyananda, 1979); A. vblubllifi (Rao, 1978 and
Pundir and Singh, 1978) 1 A. f ' *% (Radir and Singh, 19781,
Atvlofiia (Rao, 1978 and Pundir and Singh 1978) and
A. caianifalia (punclir, 1981).

Attempts at --- hybridization are presented in
Table I. The first report on intergeneric hybridization in
pigeonpea dates back to 1956 when Deodikar and Thakur crossed
C. cajm with -. The hybrid was fairly fertile.
K- EL al (1958) extended the earlier work on to hybrid
cytology and found regular bivalent fonmtion in the hybrid. A
hybrid betwem C. and A. Bcarabaeoides was obtained by Roy
and (1967) who expressed doubts about the generic status of
A t y b s h . Rec?dy (1973) analysed pachytene chromosome pairing in
Ca ianusa , Btvlos ia l ineata ,A. -andA.ser icea
and their hybrids. These pachytene studies in general revealed a
high degree of chromosome homDlogy between C. and the three
species of -. Studies on the krheritance of a few
qualitative traits was also done in this study. Miya~yagarn and
Spence (1978) reported hybrids between Qjaua and A.
while further attagits (Pundir,l98l; Re&3y et al, 1980 and in
the present study) to cross Sajdnus with & failed.
Further attempts at fajmus - hybridization by Pundir
(1981) involved karyotype conprisons between the cultivated 'and
the wild species and meiotic pairing in the F1 hybrids. These
studies revealed a great degree of karyotypic similarities
between species. All the studies on ~ - & y & A a
hybridization revealed a close relationship between the species
of the two genera and regular pairing in their hybrids which
nevertheless exhibited a fair degree of sterility.

T a b l e I. Studies on i n t e r g e n e r i c h y b r i d i z a t i o n i n plgeonpea.
Year &ykxJa species used Authors
1956 A. Deodikar and Thakur
1958 A. Kumar at al
1967 A. scarabaecldea Roy and De
1973* A. w. A. and Reddy A. sar5aa
A r l yanayagam and Spence
1981 A- a l b t c a n s r A. serlcea, A. Pundi r A. scarabecldes, A. trlnervea and A. calanlfolla
1984 A. ceLldAa (sub-sp. retlculata). A. Dundas 0-a and A. auilhLh
*This work was publlshed I n 1981.

bcept at ICRfSA!T there bas been no report on the use of
AtylQah species in pigeonpa breeding progrms. At ICRISX!
their use is mstly confined to breeding high protein lines
(Reddy et al, 1979) and to a limited extent for breeding for
insect resistance, dwarfs and isolation of cytoplasmic m l e
steriles (L.J. Reddy pers. cam.). Only a few &yh~& species
have been used in this direction so far. While there has been a
rapid accurmlation of literature on distant hybridization in
other crops, the limited interest in pigeonpea in this regard
stew from the small chromosome size and the paucity of attanpts
in distant hybridizations in pigeonpea. Genome relationships
between C a j a u ~ and Atvlafiia species is still obscure.
A. &mi&Ua, which is mrphologically very similar to
except for the seed strophile, was identified.as early as 1920
(Van der Maesen, 1980) but an attempt to cross these two species
was not reported until 1981 (Pundir) . A clear .understanding of the cytcgenetic relationships
between the cultivated and wild species is a prerequisite for
the effective use of wild species in the crop's inprovement.
Distant hybridization is nostly aimd at introducing new genetic
variability or to achieve a new g a m i c constitution in such a
way that the characters of the parental species m e recmbined
effectively. These possibilities are directly related to the
degree of genetic relatedness between the parents. It has been
found that, the closer the gemme relationship between tbe
cultivated and the wild species the greater the m t of genetic
recmbmation and consequentry variability.

Apart from being a swrce of novel genes and variability,
wide hybridizations have also prove3 to be a potential means of
haploid production. For instance Kasha and Rao (1970) reported a
high frequency of haploids from crosses betweM 8. yulgazs and
B. kulhsun. These mlgxs haploids originate from fer t i l iza t ion
followed by selective elimination of bultaim chrorrosom during
early stages of &ryo devel-t (Subrahrmnym and Kasha,1973) . Selective chrctrosome e l m t i o n leading t o haploid formation is
widespread auong ktxiiem interspecific crosses (Subrahnwyam,
1977, 1979, 1980) and also from intergeneric crosses (Barclay,
1975; Shigenobu and Sakaaoto, 1981; Botkmer & &, 1984).
Barclay (1975) reported wheat haploids (polyhaploids) from
crosses between X. aestivMl and El. hulk^^^^. Selective
chromosome e l m b a t i o n is also known in hybrids between
a. h k a m and B. . . . ( G w and G u p t a , 1973).
Haploid production via chromeome elimination has become a
successful ccanrercial venture in Barley breeding (Kasha and
Reinbergst 1980) ':
. Pigeonpa being a long duration and pbtosensit ive plant can
nonmlly be grown only once in a year. This places a severe
constraint in pigeonpea breeding p r o g r m s . Developnent of
haploids in pigeonpea would have an inmeme value in shortening
the time required for breeding a cultivar. UnfortuMtelyt the
anther culture technique has mt shown promise in any of the
l a w species. From the experience in barley it a d
worth while t o screen Gaj.am5 - A t y h s h crosses for selective
chrorroale e i h L . ~ ~ i u r r as a a u r c e of haplcids.

Pundir (1981) reported that g ~ ~ e of the species
used in his study failed to cross in any direction. Natural
barriers to interspecific hybridization may be of p r e or
post-fertilization nature. The advent of the d r y 0 culture
technique has helped in overcaning such problens especially the
post-fertilization basriers. Rescue of interspecific progeny
through ' d r y o culture was done for the first time in a Liam
x L. austtiaenn cross by Liaba_- (1929). Raghavan (1977)
lists 75 species (updated by Stewart,l981) in which the &ryo
culture techniques have been standardized. New hybrid
ambinations through the use of d r y o culture include sweral
legumes (lionma, 1955; Bajaj and Bow, 1971; Braak and Kooistra,
1975; Alvarez et al, 1981; Mok & al, 1978; Ann and Hartman,
1978; Newell and Hymwitz, 1982; Sastriand Eloss, 1982; Gosal
and Bajaj, 1983; Chen et al, 1983).
In several instances it has been fcund that the &ryo
culture technique helps in rescuing the hybrid progeny only in
dination with h o m n e treatments (Islam, 1964; Kruse, 1974;
Bajaj 'k al, 1980; Alonso and K k b e r , 19801 Sastri st al, 1981 r
Sastri and MOSS, 1982; Kuwada and Mabuchi, 1976) . Honmne
treatments prolong the enbryo deve1-t on the fenale parent so
that the d r y 0 at excision is more enenable to culture.
Further, hormone t r e a w t s are also known to *rove the rate of
success in compatible crosses. In tbe light of the literature it
seaaed worth while to standardize eubryo culture technique for
pigeonpea and to study the effect of bornrme treatments in
intergener~c crosses invoiving pigeonpea.

Cbtaining species hybrids and assessment of g m
relationshfps is a first step in the exploitation of wild species
in the Lrprwement of any cultivated species. The next logical
step is the utilization of such hybrids in the breeding programne
before which it would be essential to study the inheritance
pattern and also assess the quantum of variability generated.
Studies on inheritance provide information on the ~ossible n&r
of genes governing a character and their interaction. Evaluation
of the variation in the F2 generation help in understanding the
extent of reconbination and variability. Genetic studies provide
a clear direction to the handling of segregating generations.
There have been several studies on the genetics of qualitative
and quantitative traits in pigeonpea (Deshpande and Jeswani,
1956; D'cruz and Deokar, 1970; l-kmoz and Abram, .1971; Pandey,
1972; Sharma & aL 1972; Joshi, 1973; Chaudhary and
lbmbrer1977; Dahiya and Brar, 1977; Dahiya & aL 1977;
Kapr, 1977; Malhotra and Sodhi, 1977; Re&3y eL Bl, 19791
Sidhu and d u 1980; Saxena and Shame, 1981 to n a ~ a few).
Only tm studies have so far be^ amducted on the genetics of
x &yUxia crosses (RedCty F& tt, 1980 and Pundir, 1981).
Tissue culture technique are emerging as a major
sqplanentary aid to traditional plant breeding p~ocedures in
crop inprwement prograrmres (Vasil SL al., 1982) due to their
increasing importance in clonal propagation (Hura- shige, 1974) , productim of haploids (CMh-Chhg Chu, 1982), production of
disease free plants (Gengenbach ek dl, 1977) and induction of
genetic variants (Scowcroft and Lah~nr 1982).

A basic knowledge of differentiation ie essential for the
effective use of this technique. Tissue culture techniques have
been developed for several grain legumes (Kartha & B;LI 1981) but
such studies on pigeonpea are limited (Rao and Swwr 1975;
Mehta and Ram, 1981). - In.the light of the above considerations and constraints the
present investigation was undertaken to understand the
relationship between Cajaus and & & h s h and Rhvnchosia in order
to allow easier transfer of useful t ra i t s from the wild t o the
cultivated species with the following broad objectives:
1. Assessment of crossability relationships between
Cajaus, and certain Atvlofiia and species.
2. Devel-t of methods to inprove species crossability.
3, Assessment of genome relationships between c. and
its wild relatives.
4. Screening for haploids produced by selective c h t m m
eliminathn in Ujmus - Atvlofiia crosses.
5. Study of the inheritance and variation for qualitative
and quantitative t rai ts in the intergeneric hybrids and
6. Develop& of in vitra regeneration techniques in
pig-npea*


S- m: The Genetic Resources Unit, I(IRISAT was the swrce
of seed for all the &Q&a and &yncb&a species while the
seed of the cultivars of C. w a s obtained from the pigeonpea
breeding subprogram a t IW-. The salient morphological
features and eammic inprtance of the species used in this
study are presented in Tables VIII, M and X I and Table I1
respectively.
PLANP CULTURE: The f ield investigations in the present study
were carried out a t the IClUW research farm which is situated
180N, 78 E a t an altitude of 536 m. Prior to planting the land
was ploughed, harrowed and ridged. The f ie ld was divided into 4m
plots and the seed was sown rnanually a t a spacing of 3 0 6 0 an
between plants depending upon the plant habit and 75 rn between
rows. Support w a s provided wherever necessary for the clinbing
types. In ckses where plants were gram i n pots the potting
mixture consisted of three parts of m i l and one part of farm
yard manure.
Seed of the wild species and also the F18s possess a hard
seed coat, hence their seed w a s sacrified with a scalpel before
eowing to fac i l i ta te easy germination. Irrigation was provided
whenever necessary. In f ield plantings hand weeding was done
tvice to keep the crop clean. msulpban (0.2%) was sprayed a t
regular intervals to check insect damge.
E Y B F U D I ~ O N : In C. G&ZJ flower opening begins i n the morning
a t about 7 A.R. with Li.6 anthesis continuing unti l l a t e in the

Table 11. Or ig in and s a l i e n t economic features o f t h e AtvloE(a species used i n t h e study.
Species Acc. No. Or ig in Important Charac te r l s t i cs --------------------------------------*--*---*-----------------------------
A. alblcans NKR-177 Asia S t e r i l i t y mosaic r e s i s t a n t and high seed p ro te in content
A. calanlfolla PR-4876 Asia High seed p ro te in content
A. g C a U f d a PR-4221 Austra l ia High seed p ro te ln content
A. lanceolata CQ-1619 Aus t ra l ia Frost and drought t o l e r a n t
A. J d L b g a h CQ-1618 Aus t ra l ia F ros t and drought t o l e r a n t
A. JiL-mh JM-3366 Asia S t e r l l l t y mosaic r e s i s t a n t and h igh seed p ro te ln content
A. rnnuls JM-4331 Asia
A. @&SUQA PR-4557 Asia B l l g h t res is tant , annual. high seed p ro te in content
8- LuQQ= KM-4180 Asia An t lb ios is t o W&&t& armlaera and h igh seed p ro te in content
A. scarabaeoCdes JM-2367 Asia An t fb ios ls t o srmlaera and h igh seed p ro te ln content
A. sericea JM-1961 Asia B l i g h t and s t e r i l i t y mosaic res is tan t and high seed p ro te ln content
A. Yolublllr JM-4208 Asia S t e r l l i t y mosaic res is tant ' and high seed p ro te in content

afternoon. The flowers raMin open for &xut 20-24 h and the
anthers dehisce before flower opening. The species of
have a similar floral biology except that they have delayed
flower opening in some cases. Anther dehiscence in all the
Atyhs ia species occurs before flower opening.
FOE hybridization, buds of the awropriate size were opened
with the help of a forceps and anthers -re rerroved without
injuring the stigma. The forcep was dipped in spirit after each
emasculation. Initially two methods of pollination were
attanpted.
1. ESMsculation and imnediate pollination and
2. Ebsculation and pollination on the next mrdng.
The first method proved to be muginally better and it was
followed in all the later studies.
EXMXE -: Cultivar Pant A2 was us& as the Gajaws
parent for ho-ti: treatment studie6. Gibberellic acid (GA3) and
kinetin were used independantly at concentrations ranging from 19
to 80 p or as a 1:l mixture keeping the net honmhe
concentration in the szaw range (10 to 80 Am) . A ~o:22
hypodermic needle was used to place the hormone solution so as to
fill the bud cavity surrounding the pistil. The tredtments were
given twice, 24 and 48 h after pollination. Care was taken to
avoid physical injury to the buds &ring hormone application.
Cne hundred pollinations were made for each treatment. Bud drop,
p d set, pod length, and seeds per pod were recorded for each
cross and treatnent .

CX'KWSZCAL lEVE9lXGRTI(TZS: Por cytological studies buds of the
eropriate size were fixed between 9 to 12 h in Carnoy's fluid
(6 parts ethyl alcohol: 3 parts chloroform: 1 part glacial
acetic acid). After 48 hours in the fixative the buds were
transferred to 70% alcohol and stored in a cool place. For
chromsoml studies the anthers were squashed in 1% acetocarmine.
For nucleolar -ring the anthers were first squashed in 1%
acetocannine followed by destaining with 459 acetic acid and
restaining with 2% acetocarmine. Detailed observations were made
on metaphase configurations, anaphase disjunctions and nucleolar
size and distribution in the parents and hybrids. Chi-
frequencies were recorded in both parents and hybrids.
Observations were also made on pachytene chromosome pairing.
KELEN FERJ!ZITY: Pollen stainability in 1% acetocannine was
taken as an index of pollen fertility. All shrivelled, unstained
and poorly stained grains were counted as steriles, Pollen
fertility was recbrded for all the parents and hybrids.
-: Desirable -11s were photcgraphed using .a
piwtomicro8~0pe with oil intersion lens end a green filter.
Kodak high contrast copy film (ASA 64) was used for micro
EuK?lXm WICKSaXY: Laaf bits were made to adbere over a thin
film of dolite silver paint on specimen stubs, dried and were
sputter coated with a thin film of gold. Photograph were taken
on a JOEG35 scanning electron microscope.

FEWlXIN ESFDMTON: Seed protein percentage was determined
with the aid of Technicon Auto Analyzer. Fifty grsms of seed
q l e was needed for the estirmtion of protein content.
GENETIC STUDIES: The F2 poplation was raised from the croases
C. G&II X A. db,h~IS, C. & x 8. caianifoliat C. X
A. sericea, C. rajan x 8 . - and C.- x
A. UneaLa. The germinability of F2 seed is presented In
Table 111. In all the above five hybrids the Ca.ianus parent used
was Pant A2. The F2 populations were scored for seed strophiole
(presence, absence) , seed mttles (presence* absence), pod
hairiness (short hairs, long hairs), leaflet shape (lanceolate,
intermediate, type) and twining nature (ere,
intermediate, twining). The plants were classifie into distinct
groups for the above characters and the goodness of fit of the
&served segregation was calculated by the standa~d chi-square
test using the following formula
Where T = Sumnation
Observed frequency and
& Expected frequency
The variation in the F2 popllation was oorrptted for the
characters, mid-leaf length, mibleaf width8 @-length8 seeds
per pod and ICr-seed weight. Pie leaf measurmcnts were I X ~ C :r.
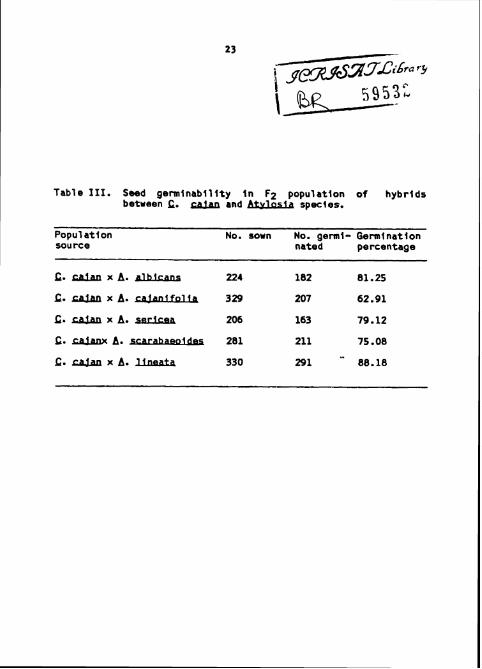
Table 111. Seed germinabi l i ty I n F2 populat ion of hybr ids between C. and Atvfasla species.
Populat ion No. sown No. germl- Germlnatlon source nated percentage

the terminal leaflet and the megsurements were on the average of
measurements based on five -lea. Observations on pod and seed
characters were from the meture pods.
IN VITFQ -ON SIODIES: Ms medium (Murashige and Skoog,
1962), B5 medium (Ganbrg & & 1968) and White's d i m (White,
1939) &re used as the basal media. The capsition of these
media is' presented in Table IV. Apart fran these media, potato
starch extract d i m (Anonymus, 1976) was used in anther
culture experbts. Stock solutions were prepared for the
different hormones used in the study and used to swlenent the
basal media depending upon the requirement. Sucrose at 39
concentration was used as the arbon source. Solidification of
the medium was achieved by Difco grade agar (8%) .. The gS3 of the media was adjusted to 5.6-5.8 by means of 0 . N IKZl or O.lN m. The media were sterilized by autodaving.
In uitrp .&eneration response was evaluated from the
cotyledonary explants of the fdjaua cultivars I B 4726,
I B 7035, Pant A2 and GS 4 and from A. caianifalia, A. aUk=am
and A. m. The regeneration potential of distal Md nodal
halves of cotyledons was evaluated. The Gajama cultivars,
Pant A2, ICP 7035, Prabhat and C 11 were used to stfvldardize the
mlture conditions for regeneration from h t u r e errbryos. The
in yifrP response of the anthers of C. &an (Pant A2),
A. rlbicans. A. atandFfdlia and A. Mlubilis yea tested.
GZXXE C(;W)ITIQB: S& (for c,.,;&ri and enbryo u r i i u r u )

Tablm I V . Constltumnts of dlffmront basal w d l a used In W ragenration rtudlms.
Constituents Murrshlgm Whltm Gamborg (mg/ll and Skoog (19191 SZ dl
(1962) MS (1968)85

and flower buds (for anther culture) were surface sterilized for
15 minutes in 10% chlorox ((BL(3WaX, USA), washed thrice i n
s ter i le water and retained for about half an bwr in sterile
water and dried before inoculation. For ootyledon cultures, the
seed coat was rem~ved and the seed was spl i t vertically to get
t m distal and two nodal halves. A l l the operations were
conductd under asceptic ~ 0 n d i t i 0 ~ in a laminar flow hood. The
a l t u r e s were maintained a t a tenperature of 25 G ' C under cool
fluorescent llght (6.8 watts m-*) .


Eight species of (8. -, A. caianifolia,
A. lineatat A. sericea, A. scarabaeoides, A. orandifolia,
A. lanceolata and A. latisepala) hybridized successfully with
C.- while the other four AQAda species (A. mollis, A.
M1UbiliR. A. w- and A. ruaosa) and the two &ynch6h
species (B. rothii and R. minirrra) used in this study failed to
cross with the cultivated pigeonpea. The - crossability was influenced by the gemtype of the Cajanu parent
( 9 ) and the cross anrbination and the hybridization attempts in
all resulted in 24 successful corrbhations (Table W .
Fumng the 8 successful Cajmm - A t y h a h crosses, the
crossability was highest when A. lineata was the male parent with
19.5%, 12.30%, 2.37% and 15.20% of the pollination resulting in
hybrids in crosses with Pant A2, Bdigani, ICP 7035 and C 11
respectively. This was followed by crosses involving A. rlbicans
ae the male parent with the successful pollinations averaging
9.16% with Pent A2, 7.23% with Baigani, 1.09% with I B 7035 and
5.12% in crosses with C 11.
The degree of success was low in crosses involving
a. A. scarabaeoides and A. caianifolia as d e parents.
A. scricea as a pollen parent yielded hybrids in 2.36%, 3.03%,
0.75% and 1.7% of pollinations with Pant A2, Baigani, ICP 7035
and C 11 respectively ar,~ when urr polirri sour-cr w a s


A. scaraboeides the crossabi l i ty with the respective C&ws
parents w a s 4.61%, 2.35%, 0.47% and 1.46%, while in crosses
hVolv.ving A. caianFfolia 2.70, 3.05%, 0.58% and 2.09% of t he
pollination with Pant A2, Baigani, ICP 7035 and C 11 respectively
resulted in pod set. Crosses involving the genotype I B 102 with
t he above mentioned species of fai led. In crosses
involving the three Australian the crossabi l i ty was low
with a pod s e t of 1.72% and 3.07% in Pant A2 x A. and
C 11 x A. arandifolia crosses respectively, 0.9% and 1.96% in the
crosses Pant A2 x A. and C 11 x A. and
1.69% and 0.93% in PantA2 x 8.- and C 1 1 x
A. lanceolata conbinatiow.
Reciprocal success of a very low frequency was obtained in
two cross oonbinations viz. A. albicans x S. &.an ( .22%) and
A. aericea x G. & (.IS%) (Table V) . In the crosses A. mollis
x C. rajr7n and A. volubilis x S. pod dwelopnent in the
crossed buds was mrmal but the seeds from such pods were
extremely shrivel& and inviable.
Seed set per pod also varied with the f e e genotype, with
ICP 7035 m i s t e n t l y averaging a higher n m b r of seeds (about 3
per pod) (Table V) . In the study in which Ii-aUkana and A-serfcea were used a s
the male parents in crosses with 1- 32, ICPL 47, ICPL 59 and
1- 95 all of which have a commn female background in t h e i r
parentage variation in the crossabi l i ty was narrow ranging from
2.5% to 5.5C m crosses k l t n A. slblcans and 1.5 t o 4% i n those

Table V I . Crossabi l i ty o f specles with G&nus having a comnon female background*.
Cu7 t i v a r Parentage A. albLanS A. adGm -------------- -------------- a b c a b c .................................................................
ICPL 32 T 21 x B r a z i l 1465 200 5.5 31.6 200 3.5 32.9
ICPL 47 T 21 x Ja 2772 200 5.0 36.2 200 1.5 34.7
ICPL 59 T 21 x EC 100467 200 4.5 34.9 200 4.0 29.8
ICPL 95 T 21 x NP(WR)lS 200 2.5 33.2 200 2.5 31.4
--
*a: ~urn* o f p o l l lnat lons attempted b: Percentage o f successful p o l l l n a t l o n s c: P o l l e n s t e r i l i t y I n t h e F1 hybrids

3 1
involving A. (Table VI) .
C. cv Pant A2 was the fanale parent in the hormone
studies. The successful conbinations exhibited a unif o m
response to hormone treatments. GA3 was found to be suprior
irrespective of the cross &ination. For e x q l e when
A. arandifolia was the pllen parent, *set increased from 2%
in the control to 14% in treatments with M p of GA3. The
optimum concentration of GA3 was found to be 40-50 m. Concentrations above 50 @ were detrimental with ocmplete
failure of pcd-set at 80 m or above. Kinetin trf?atments showed
no oonsistent response at the concentrations tested. Treatments
with GA3 + Kinetin mixture also did not inprove pod-set but were
detrimental at higher concentrations. Similar trends were .d
aparent in crosses of C. s&an with A. albicaw,
A. s+d&lh, or A. sericea as m l e parents (Table VII) . In the four successful crosses studied for hormone response,
GA3 alone or in dination with kinetin increased pod length and
seeds per pod (Fig. 1). In these crosses the average pod length
at physiological maturity increased from 4.5 to 5.5 an in the
control to 7 cm. Furthemre, the seeds per pod increasd from
the range of 1.6-2.2 in controls to 3.5-4 when Gh3 or GA3 +
kinetin treatments were given. W w e r , honmne treatments did
n: t i~fluarce reed size.


F i g . 1 . Effect o f gibberel l ic acid and/or kinetin on percent pod and seed se t in four C q j a m s - A t y l o s i a crosses .

norane concentration
F i p u r c 1.

AmDng the wucceesful crosses bud drop comnenced within two
days after pollination (Fig. 2) . GA3 treatments prolonged wary
Wel~pnent and delayed bud drop by varying periods depending
upon the cross carbination and the homne concentration (Fig. 2).
For wmqle, when A. and A. Mlubilis were the pollen
parents, bud drop cannenced two days after pollination in the
control. GA3 prolonged this period up to five days at 60
concentration in A. volubllls . . and at 50 ppn concentration in
A. -. When A. nallis was the male parent bud drop was
delayed by 3-4 days. Increase in ovule size was evident in cross
cabinations where bud drop was delayed following hormone
applications.
In all the reciprocal crosses there was variation in
response to hormones (Fig. Zb) . A. sericea and A. arandlfolla . . failed to respond to any of the three treatments. In
A. GA3 delayed bud drop while GA3 or GA3 + kinetin treatments were ineffective, a trend which was similar to the one
observed in unsuccessful f&ws crosses. Kinetin at a low
concentration (10 Hm) prolonged ovaq developmnt by 3 days. in
A. platvcarDa and 4 days in A. -.

Fig. 2. Effect of gibberellic acid and/or kinetin on delaying bud abscission.
a . Cajanus cajan as female parent b. Cajanus cajan as male parent

Days GA?+Kl n Kin

In gross morphology the F1 hybrids in general tended to be
intermediate. Atvlosia characters like seed strophiole, seed
mottling, pod hairiness and persistence of petals were expressed
in the F1. A oarparison of parents and their hybrids for
qualitative and quantitative traits are presented in Tables VIII,
IX and X, and a hybrid wise description is given below.
The hybrid was a twiner and had a leaflet shape intermediate
between the parents (Fig.3B) in ita initial stages of growth.
'Ihe intermediate leaf shape in the hybrid continued for about 100
days. LXlring this period the leaf tip was obtuse.. The leaves in
some of the branches (Fig.Sa-leaf 2) which pea red after 100
days resabled the Cajanw parent, having an acute tip
(Fig.5C:leavess 4-6). However, as the plant grew further the f
remaining branches aleo developed leaves similar to that in the
Qijanus parent. Although the change in leaf shape was observed
in all the three leaflets it was most p r o m & in the terminal
ones. The texture of the leaves produced after 100 days atso
reserrbled the C ~ ~ B U I E parent. Scanning electron microscopic
studies of the upper surface of the terminal leaflets revealed
slniluities in the leaf surfaces of the Sajanu parent (Fig. 5d)
a d the --like leave6 produced in the hybrid (Fig 59) . m t h ihe leaf surfaces exhibited long trichomEe with uni£orm
spread. I*?zv,es of the A. albicans -rent had a very dense
population of tr ichoims (Fig. Se) , whereas the initial


Table IX. Expression o f quant i ta t iave t r a i t s I n parental . p r i e s and F1 hybrlds batweon C. a c v . Pant A2 and A w Q A A sp.ci.s.
Character Specles/Hybrld ------------------,-------------------
Midleaf Midleaf Pe t i o l e Days t o Durat ion length width length f lower of f l o - (cm) (cm) (cm) r e r f ng .................................................................
C. a 7.13 2.85 4.1 65 Sep-Jan cv Pant A-2 - 8. alblcans 4.41 3.82 4.3 155 Dee-Mar
C. ralna x 5.3 3.3 6 .O 140 Nov-Mar A. - 8. SBJxba 2.51 0.63 1.52 126 Nov-Mar
115 Nov-Mar
76 Sep-Mar
85 Oct-Feb
118 Oct-Ma r
123 Oct-Mar
160 Dec-Mar
l30 Nov-Ap r
Maintained as perennl a1
C. U x 7.7 4.15 5.8 155 Dec-Apr A. gLum-uA
S. Gahn x 6.34 3.96 4.6 172 Dec-Ma r A*
A. h x ~ ~ k t ~ 9.4 0.82 1.5 222 Fob-Apr
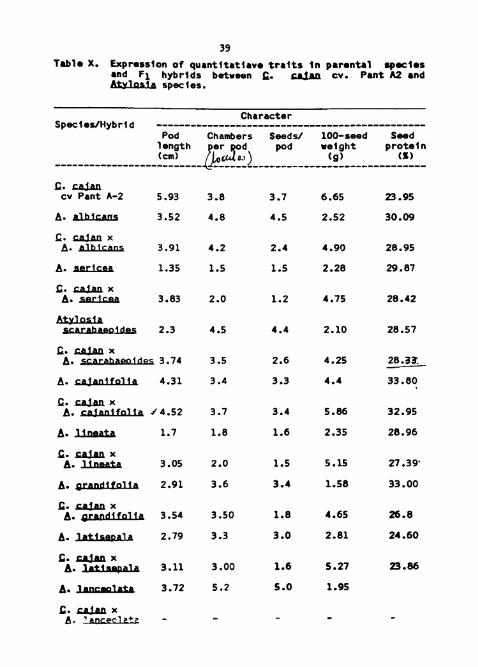
Table X. Expression o f quantltat iave t r a l t s In parental . p a l e s and F1 hybrlds between C. & cv. Pant A2 and Atvlosla species.
Character Species/Hybrld ...............................................
Pod Chambers Seeds/ 100-seed Seed pod weight p ro te ln
(cm) ( X )
c. a d n cv Pant A-2 5.93 3.8 3 .7 6.65 23.95
I;. r;aiaa x A. P 3.74 3.5 2.6 4.25 28.33.
C__

intenoediate type leaves had a eparse poprlation of short
trichomes (Fig-Sf) when caqxued to the leaves of the r;iianus
parent and the caianus like leaves in the hybrid. Floral
initiation took place only on tk branches which had developed
leaves similar in shape and texture to the parent. The
pods in the hybrid were flattened and glabrous with an average
length of 3.91 an. The chaxbers per pod averaged 4.2 and the
seeds were strophiolated and mottled. The seed weight (4.92 g)
was in between the parents with protein content (28.95%) close to
the &yb&ia parent (30.09%). Petals in the hybrid were
persistent and the hybrid flowered from N o v ~ r ~ c h with the
first flush a~pearing 140 days after sowing.
The F1 hybrid was erect with a leaflet s h w towards the 4
Akybsh parent (Fig-4D). The leaf length (7.97 cm) and the leaf
width (3.34 cm) in the hybrid were higher than the better parent
(Table MI. The first flush b. the hybrid a m e d about 130
days after sowing with the peek flowering period falling between
Mvenber and April. Petals in the hybrid were persist&. Rd
mrphology was cloee to the Uyhaia parent. The pods were
reddish green in colour with dense hairs and had 2-3 chsmbcrs
with an average of 1.6 seeds per pod. The pod length averaged
3.05 an. Tho seeds were mttled and strophiolated. A 100-scsB
weight of 5.15 g, and a protein content of 27.39% was recorded
(Table X)

F i g . 3 . Twigs and l e a f l e t s o f parents and h y b r i d s .
A) a . Cajanus cajan
b . Hybrid between Cajanus c d a n and AtyZosia scprabaeoidt
c . Aty Zosia scarabasoides
8 ) a . Cdanus cqjan
b . Hybrid between Cajanus cajan and AtyZosia aZbicans
c. AtyZosia aZb icas
C) a . Ca,janus cajan
b . Hybrid between Cajanus cajan and AtyZosia sericea
c . Aty Zosia sericea
D) a . Cajanus cajan
b . Hybrid between Cajanus cajan and AtyZosia cajanifo:
c . Aty Zosia cajanifoZia


F i g . 4. Twigs and l e a f l e t s of parents and F1 hybrids.
A ) a . Cajanus cajan
b. Hybrid between Cqianus cajan and AtyZosia grandifotia
c . AtyZosia grandifozia
B ) a . Caj;~us cqjan
b . Hybrid between Cqianus cajan and AtyZosia Zatisepata
c . Aty Zooia Zatisepata
C) a . Cajanus cujan
b . Hybrid between Cajunus cajan and AtyZosia ZanceoZata
c . A t y Zosia tanceotata
D ) a . Cajanus cqjan
p. Hybrid between Cajanus ca;'an and Atytosia Zineata
c . AtyZosia Zineata


Fig. 5 . Varation f o r leaf shape and u l t r a s t r u c t u r e i n hybrids between CaJanus c&an and AtyZosia atbicans,
a . Leaves from d i f f e r en t branches (1 t o 6 from t h e bottom) of a t yp ica l Cajanus cqjan-Atytosia aZbicans hybrid. (Note d i s t i n c t leaf shape from t h e branch two).
b . Leaves from bottom t o top o f a branch, which d i d not show d i sce rn ib l e va r i a t i on upto 5 months.
c . Leaves from bottom t o top of a branch which showed d i f f e r e n t l y shaped leaves a f t e r 100 days.
d. Leaf surface scan of Cqjanus cqjan.
e . Leaf surface scan of AtyZosia azbicans.
f . Intermediate type leaf surface scan.
g. Cajcmus type l ea f surface scan.


lh hybrid was e rec t in Wit and close t o the Cajauus
parent in leaf shape (Fig.3C) . The mid-leaf length (5.34 cm) , mid-leaf width (1.85 on) and the petiole length (3.55 cad were
all i n t e d i a t e t o the pazents (Table IX). Floral i n i t i a t i on
t o o k place 126 days a f te r sowing and the peak flowering was
recorded between Mv&xr and March. The pztals were persis tent
and the pods were hairy. Pod length in t h i s hybrid averaged 3.83
an with an average of 1.8 chatrbers and 1.2 seeds per pod. The
strophiolated and mottled seeds of the hybrid had a 100 seed
weight of 4.75 g and a protein content of 28.42% (Table X I .
The hybrid was intermediate i n its habit with t he base being
bushy and the u w r portions showing atwining tendency. The
l e a f l e t shape was intermediate (Fig. 3A) as were also the mid-leaf
l ~ g t h (4.9 an), mid-leaf width (2.0 d and the pet iole length
(3.8 on) (Table I%) . While the average pod length (3.74 ad in
t h e hybrid was in between the two parents the n a b e r of &tubers
p r pcd (3.5) and seeds per pod (2.6) were less than those of
e i t he r parents (Table X) . The seeds were mottled and
strophiolated and the seed weight and protein content were 4.25 g
and 28.33% respectively. The hybrid flowxed 85 days after
wming (October to February) as against 75 days i n E&h&a and
65 days in the !2&nu parent.

This hybrid unlike others was irrjistinguishable f r m the
parents. It has an erect habit and a lanceolate leaf shape
(Fig. 3D) similar t o the parents. The mid-leaf length (6.5 an) , mid-leaf width (2.27 an) and the petiole length (4.4 cm) were
also close t o the parental values (Table IX) . The pod length in
the hybrid averaged 4.5 cm with an average of 3.7 chaJrbers and
3.4 seeds per pod. The seed was strophiolated and mottled with a
100-seed weight of 5.86 g and a protein content of 32.95%. The
hybrid bloomed i n 125 days after sawing and the flowering which
intiated in October c~ntinued unti l March.
The hybrid resenbled the a parent in gross morphology.
The leaves were ovate to rounded i n shape (Fig.&) and the
mid-leaf length (7.7 cm) was greater than that in either of the
parents while the leaf width (4.15 cm) was intermediate between
the parents (Tablw IX) . Pods being hairy and yellwish r e s d l e d
the AtykxLa parent and the petals were persistent. The pods had
an average of 3.5 chanters and 1.8 seeds with an average l q t h
of 3.54 cm. The seeds were mottled and strophiolated and had a
100 seed weight of 4.65 g as against 1.58 g i n A. clrandifolia and
26.8% protein (Table X I . The hybrid which flowered between
Decerrber and April had its f i r s t flush 155 days after sowing.

In gross tmrpblogy the hybrid was intenrediate to the
parents. The leaves were intermediate in shape but had a m r e
pointed t i p (Fig. 4B) than those in A. and were much
l e s s densely haired than the leaves of A. m. The hybrid
leaves were 6.34 an long, 3.96 an wide with a 4.6 an long
petiole. The pods in the hybrid, which were less densely haired
than those in A. averaged 3.11 an in length w i t h an
average of 3 chanbers and 1.6 seeds per p d . The 100 seed weight
(5.27 g) was intermediate t o the parental values and the seed
protein content was 23.86% (Table X) . The hybrid flowered
between Decerrber and March with the f i s t flush a p r i n g 175 days
af ter sowing.
The hybrid, which in general was intermediate i n its gross
mrphology, had leaves shorter and broader than those of .f
A. lanceolata (Fig. 4C). The hybrid did not flower wen af ter 200
days of growth.
Eight species of Atvloaia, A- albicans~ A. aericea, A.
scarabaeoideet A. caianifalia, A- A. arandifolia, 8.
and A. latisebala hybridized w i t h G. &an with
varying degrees of success. The Australian species of Atvloaia.
8. -, A. lancealz+r a d A. have been crossed
w i t h cultivated pigeonea for tbe f i r s t time. Studies on a few

other epecica of Australian were conducted in a
parallel study at the University of Weensland (Ian mndas,
pers. cam.). Earlier atteapts at Lntergeneric hybridization of
pigeonpea were mstly concerned with crosses involving 8. lineate
(Deodikar and Thakur, 1956; K w & dl, 1958; K m and
llrrrbre, 1958; Reddy and De, 1983). There have been a couple of
successEu1 attanpts at crossing A. aericea and A. scarabaeoides
(Sikdar and De, 1967: Reddy, 1981; Pundir, 1981) and Alndir
(1981) used 8. 8. trinervea and A. caianifolia in
intergeneric studies.
Mnng eight successful fajanm - &ybsh crosses in the
present study the degree of species crossability was highest in
crosses involving A. lineata as the pollen parent followed by
those imrolving A. albicans. The crossability was.relatively low
with the other species. A. caianifolia. which is ~rorphologically
Mistinguishable from Cajxiua except for the strophiole on the
seed, did not hybridize with the orltivated species as freely as
8. lineata or A. elblcans j . . The rate of success in intervarietal crosses of pigeonpea varies from 6% to 70% depending upon the
genotypes involved. Thus the degree of species crossability in
itself is not an index of species relationship.
A wide variation in the crossability of a given &yk?sia
species with different cultivars of pigeonpa was wident.
Similar gamtypic variations for crossability are already knawn
in intervarietal crosses of pigeonpee (Singh & nl, 1980; Saxena
& b;L, 1984). The wnplete failure of the cultivar ICP 102 to
cross with any of the Atylosia species is not ~urprisir~g in the

light of its poor performance in intervarietal crosses of
pig-npea (K.B. Saxena, pers. cam.) . The w r o w range of
variation in the crossability of species with four
different gemtypes (IBL 32, ICPL 47, ICE% 59 and ICPL 95) of effect
f. is a reflection of a possible cytoplasmiclsince each of
these genotype originate from the azure f a d e parent.
Differences in the crossability of different varieties of
heat with rye (Rao, 1968; Zeven and Van Heeuert, 1970; Lange --
and Wojciechowska, 19761 W s s and Jalani, 1980) and with Hordeurn
bulbasurn (Snape SL pl, 1979) are known to be under the control of
crossability genes (Moss and J a s l , 1980; Falk and Kasha,
1980). With the developrent of genetic stocks of pig=-, it
should be possible in Cajmus - &yk&a crosses to elucidate the
inheritance pattern and the nature of crossability 'barriers.
The failure of as a male parent to cross with most
of the species might reflect a pre-fertilization barrier
as known in some bf the interapecific crosses of AraAh (Sastri
and Mae, 1982). The differences in the reciprocal crosses might
originate from the differences in the ccmaenceaent and progress
of division of endosperm and dryonic cells which has been shown
in (Robakoarihanta et & 19791 Shii & aL 1982) or
as a result of the failure of the pollen tube to grow through the
style (Muller, 1960) . In incmpatible S & ~ U E - A Q ~ M ~ CrOBsCB of the present
study our preliminary observations on pollen germination and
pollen t.:).: c,:o-,,th with tk,i: aid of a light iiicroscop usincj

acet-e s t a i n Indicated t h a t the pollen germinates and
en te rs the s tyle . Unfortunately this technique did mt help in
probing further t h e develo-t of pollen tubes. Limited
investigations on flourescence microscopy using Aniline blue did
not provide any further information because of t h e limited
clearing of t h e s ty la r t issue. I n the crosses A. voltlbilir x
C. rajan and A. rrollis x C. sajau pod dwelopnent was normal but
t h e seed w a s extremely shrivelled and w a s inviable. In these
crosses the bud drop occurs 6 days a f te r pollination as against
t h e usual 2nd or 3rd day a f t e r pollination observed in t h e other
crosses. These observations indicate t h a t t h e bar r ie r s t o hybrid
production in these two cross corcbinations might be operating a t
o r a f t e r fe r t i l i za t ion . But the fa i lu re t o recover any viable
seed w i t h honmne treatments o r errbryo cul ture in these crosses
could be taken as an indication for p r e f e r t i l i z a t i o n barr iers .
Developrent of mature pods with no seed d e v e l o m t has also been
reported in crosses involving Ebm arvense as female and
. . E. -and E. as pollen parents (Foujdar and
Tandon, 1976) . The crossed pods were shorter in length with fewer seed per
pod than selfed pods. The r a t e of pod developnent was also
slower in the crossed pods. I 8 7035 consistently averaged a
higher n d r of seeds per pod i n crosses with a l l t h e Atyks ia
species. Seeds per pod in ICP 7035 is greater than that in any
other cul t ivar used in this study inplying the predisposition of
a greater n w t r r of ovules per wary for t h e a l i e n pollen t o
f e r t i l i z e != ; i ?.; tc a k,igher seed se t i n t h e mature pods.

The results of h o m e treatmpnts indicate that in the
intergeneric crosses of and the rate of success
can be increased with hornvne treatments which also increase the
percentage of pod-set, pod length and nlmber of seeds per pod in
the cross cabinations. !R-e success rate in the untreated
wntrols was law. Moreover, the onset of flower drop amng the
unsuccessful crosses was delayed following hormone treatments.
The increased pod set associated with most of the W or
W+kinetin treatments is likely due to enhanced post-
fertilization developklt of both the ovary and ovule(s) since
treatments ccsrmenced one day after pollination when fertilization
would have been completed. The increase in pod length and ncmber
of seeds per pod following hormone treatments further
substantiate such an interpretation. Higher concentrations of
GA3 and GA3 + kinetin rdced the rate of success, but promted pod length and the nuher of seeds per pod which suggests the
role of hormones Fn post-fertilization developnent . Kruse (1967) &d Kasha et BL (1978) attributed increased
success in obtaining progeny from intergeneric or interspecific
crosses to the check by emgemus GA3 supplied to the florets on
post-fertilization break-. The increase in pd length in our
C a j a n m - m crosses may reflect increased cell n m e c and/or
cell elongation in the ovary wall after hormone treatments.
Kasha & al. (1978) observed increased mitotic activity following
GA. treatments in interspecific crosses of &)dam. Sastri and
EIoss (1982) found increases in peg nm&r as we7 1 as peg length - following GA3 treatments in Arachjn intersyeclfic crosses.

Al-Yasiri and Coyne (1964) alm rcmrded a positive influence of
honrones on pod length and diameter in a Phaseolus species cross.
(Xlr findings in pigeonpea intergeneric crosses are consistent
with these results.
The fai lure to increase further the nunher of successful
cross &inations erphasises the homna l role in the
post-fertilization processes only. Since the honmnes did not
improve the size of the mature seeds, the increase in ovule s ize
in the unsuccessful crosses is likely due to delay in bud drop
thereby permitting the ovule to develop for a longer period. In
the unsucce%sful carbinations GR3 treabnents helped to delay the
comnencement of flower bud abscission. In crosses between
P~SEQUE and E acutifolius. a ambination of G.3 and
NAA was wed to o v e r m pod abscission (Al-Yasiri and Coyne,
1964). Rnsweller and Stuart (1948) reported the Mluence of
grawth regulators on retarding the senescence of the arbryo sac,
and on the length and diameter of capsules in Lililrm LmgUhum. d
Although kinetin treatments (10 t o 80 FFn) did not show any
inprovement in *set or the nmber of seeds per pod in our
study, treatment a t the lowest concertration (10 Rn) w a s
effective in delaying bud drop in the A. albicans x C. ~ & n
and A. x C. crosses.
The data show that , amng the various crosses studied,
reeponse t o horrrone treatments is determined by the f d e parent
irrespective of the pllen parent tllsed (Table VII, Figs. 1 and
2) . The response of a, caianifalfa as a female parent was
6 - 1 ~ to that G -. These two species have more

morphological similari t ies than do the other species uaed in this
stucly and are considered one genus by mnre twonomtste (van &r
Maesen, 1980). In the unsuccessful cutbinations in spi te of
increased ovule size followhg honmne treatments, our at terpts
to culture treated ovules asceptically have failed. Furthermore,
cultures of pr-ture (< 11 day old) @fed embryos on defined
media have not developed into plants. (Details of d r y 0 culture
are presented in Chapter 7).
The present study shows that hormones can increase the
percentage of success in the crosses where the degree of success
is otherwise low. Cm the basis of the present results, it is
suggested that it would be worthwhile to t ry other hormones and
the i r cabinations in an attempt to increase the n h r of
successful crosses through further refinemnts' i n the a r y o
culture technique.
The hybrid nature of the F1 plants w a s very rmch wident
from &serving d e i r plant mrphologr. &yJ,c&a characters l i ke
seed strophiole, seed mottles, pod hairiness and twining nature
are dominant and their expression in the hybrids permitted an
ummbigous identification of hybrids and selfs. The leaf shape
in the hybrids, which' in mat of the cases was either
intenaediate or tending t o r e s d l e the male parent, helped i n
easy identification of the hybrids. MaXmb (1975) raised doubts
about the genuinity of intergeneric hybrids in l a s a e wing
to epcmictic developnent of seeds in same 1egrmLinous species but
in the present material wit? the expression of the ndle parent
t r a i t s the problem of identifying true hybrids is ciramnrented.

In the C. cajm x 8. abkma hybrid an interesting feature
was the variation for leaflet shape and texture (Fig.5) . The
deklopwnt of leaves similar to the parent after about
100 days led us to lodc for the possibility of selective
chromsome elimination similar to that of Nicotiana (Gupta and
Gupta, 1973) or Hordeum (Subralnnanym and Kasha, 1973). But our
cytological studies presented in Chapter 4 revealed eleven
bivalents and regular didunction. This could be taken as
evidence t o rule out the possibility of chromsome elimination or
in support of chrmsome doubling following chromsane
e lh imt ion . A detailed meiotic analysis (Sateesh K ~ r m a r et al,
1984) revealed the hybrid nature of the material. Thus it was
concluded that the variation in leaf mrphology accompanied by
flowering is a consequence of differential gene expression in
different branches. Since floral ini t iat ion occured only in the
branches on which !2jauu l ike leaves started developing, it is
likely that these two processes are temporal events in gene
expression . -tic variation in C. c&n was reported to be chimeral
(Rao and Reddy, 1975) which qpeared from the seedling stage.
Thus it is different from, the type encountered in the present
--&yl~& hybrid.


Qltoa~some nunber in and all the species of
atvloeia and Rhvnchoeia used in this study was found to be n=ll
or 2n=22,. Meiosis in pigecxlp (Fig.6AzG) was highly organized
with r-ar bivalent formation and normal disjunctions at both
divisions irrespective of the cultivars used. Species of
Akyb&ia and also formed regular bivalents (Figs. 7 and
8 ) with normil disjunction except for occasional asynchrony at
division-I1 in A. lancealata and A. serice9. Different cultivars
showed minor variations in their chi- frequmcy (Table XI).
In all the nine cultivare of fajmus the chiasma frequency was
around 20 per cell ranging from 19.75 i n C 11' to 20.75 in
Baigani. The chaismata per cell in A. sericea (20.85) , A. (20.50), A. (20.501, A. lanceolata
(20.301, A. acarabaeoides (20.52) and A. lirrPata (20.42) were
close to those in but different from A. (17.75)
and A. arandifolia (17.72).
In and &ykasia ~pecies studied, pollen mther
cells (PMCs) containing nucleolus aslsociated with two bivalents
were m m n (Fig&6B, 7G, 75 , 8D, 8.7) at diakinesis while two
nucleoli per Pm: (Figs.7A and 7J) were less frequent at
diakinesis and rare at pachytene (Pig. 2OA) . In C. &an the
n m r of nucleoli per PMC varied from 2 to 4 at telopha6e-I
(T-I) (Fig. 9A) and 4 to 8 at T-I1 (Fig. 9B-P) irrespective of the
cultivar. Similar variations a r e evident in A. albicans,

Tabl r XI. Ilrt.phas6-I conflguratlons* and po l l on f o r t l l l t y I n G. W c u l t l v r r r and Atvla.(l s p r l r s .
B l va l rn t s Av. Cult lvar/sprclos (Range) Pol 1 en ---------- r t d c e l l f o r t l l l t y
Ring Rod (I) ........................................... fi. tplM cv. ICP 7035 9.4 1.6 20.40S.09 96
(9-10) 10-2)
C. rPiPD cv. Bafganl
G. m cv. C 11
C. U cv. ICPl 32
G. raLnn cv. ICPL 47
C. rPL.n cv. XCPL 59
fi. rplna cv . ICPL 95
C. rntnn cv. ICP 102
A. Alhkma
A. lrneata
A. - A. - A- c&mlid-
*30 c e l l s were scorad for rach s p s l e d c u l t l v a r .

Fig. 6. , Meiosis in C. c(2vian cv. Pant A2 and metaphase I in cultivars of C. cajan. (Bar indicates 10 u).
A-G: Meiosis in C. cajan cv. Pant A2.
A) Pachytene showing regular pairing.
6) Diakinesis showing two bivalents attached to the nucleolus.
C) Metaphase I with 11 bivalents.
D) Equatorial view showing chromatic associations at late metaphase I.
EEF) Anaphase disjuncrions.
G) Late anaphase I1 showing regular disjunction.
H-0: Metaphase I in cultivars of pigeonpea showing 11 bivalents.
H) ~aigsni. I) C 11. J) ICP 1 0 2 . K) ICP 7035 .
L) ICPL 32. M) ICPL 47. N) ICPL 59. 0) ICPL 95.

- &." , . , Y*
! .fr --.... '.; 4 .,#
4, .. * 6 , ,.$; : ' ... .$, " I '9 'a ':.:, . , < , - y
, , .> ". * .'* tV . "5 . . . ' 0 " ' U."?," P -. A . B -
L b . - C - - a m , <> -JW.
- # ' .=r -e 5
-'-e ,a a- CZ
-2% m. 1'6 "A, .- D ,, !*m$g F
\ k C
hb , & dV
)r e t - H
Z . 1-

Fig. 7. Meiosis (Diakinesis, metaphase I and anaphase I) in AtyZosia species. (Bar indicates 10 u)
A-C: AtyZosia zanceotata
D-F AtyZosia grandifot ia
G - I AtyZosia Zatisepata J-L Aty los ia aZbicans
Note two nucleoli in A and J.


Fig. 8. Meiosis (giakinesis, metaphasc I and anaphase I) in AtyZosia species. (Bar indicates 10 u)
A-C: AtyZosia sericea
D - F : AtyZosia ca jani fo t ia
G - I : A t y Zosia Zineata
J - L : At? Zosia scarabaeoides


A. lineeta and A. Ptanairalla w h i l e in A. caianifolia,
A. lanceolata and A. scarsbaeoides, the nvutirnna nlmber of
nucleoli per FfC vks seven (Fig. S O L ) . The maxinun n m r of
nucleoli in each daughter nucleus at T-I or T-I1 never exceeded
two (Fig. 9) one of which was always bigger than the other. The
variations in frequencies of PMCs with different nlmbera of
nucleoli at -11 in different species k e presented in ~ig. 10.
The nost, freqc,--.t types were FWS with four nucleoli while the
least frequent czes were those with eight nucleoli.
The present study on the chromsome nwr&r. in C.
A. linaata. A. caianifoliat A. serice9, A. scaraboeides,
8. MlubilFs and A. confirms the earlier reports ( K m
ale 1958; Sikder and De, 1967; Shrivastava eL aL, 1973; Reddy,
1973; Bir and K L ? ~ 1973; 19773 Pundir and Singh, 1978; Rao,
1978; Sanjappa and Satyananda, 1979; Pundir, 1981) as 2n=22.
The chranosome nwrbers of A. atanalfolla . . , A. latisepalat
A. lanceolata, A. mllia, B. rathii and B . minima which have not
been r-rted eazlier has been found to be 2n=22.
The mwimum n h r of nucleoli in each daughter nucleus at
T-I or -11, never exceeded two (Fig.9) irrespective of the
cultivar or species studied suggesting thereby the presence of
two nucleolar or~anizers (N.0.) per genome in pigeonpea.
i r . L l t e ~ n ~ ~ i , .;.= b r L - ~ ~ n ~ ~ e of a naximum of two nuciwri at

Fig. 9. Nucleolar distribution in pollen mother cells of C. ca.jan and species of AtyZosia. (Bar indicates 10 m).
A-F: Nucleolar distribution in 'lollen mother cells of Cajanus ccqian.
A) Telophase-I nucleolar distribution, 2-2. Ti lophase-II nucleolar distribution. 8) 2-2-2-2.
C) 2-2-2-1. D) 2-2-1-1. E) 2-1-1-1. F) 1-1-1-1.
G-L: Nucleolar distribution in pollen mother cells of A t y Zosia species.
G) Telophase-I nucleolar distribution in A. grandifozia, 2-2. H) Telophase-I1 nucleolar distribution in A. grandifotia,
2-2-1-1 (Arrow indicates two fusing nucleoli). I) Telophase-I1 nucleolar distribution in A. ca jan i fo t i a ,
2-2-1-1 (Note the difference in the size of fused and primary nucleoli).
J) Telophase-I1 nucleolar distribution in A. t i nea ta , 2-2-2-1. K1 Telouhase-I1 nucleolar distribution in A. ZatiseoaZa. 2-2-1- ~j ~elophase-11 nucleolar distribution in A. a ~ b i c & s , i-2-2-1.


Fig. 10. Frequency distribution of nucleoli at telophase-I1 in pollen mother cells of C. cw'an and A t y Z o s i a species.


pachytene and the presence of two nucleolar bivalent8 at
diakinesis is also indicative of two pairs of nucleolar
organizers in the Qj&us and species studied.
The variation in the frequencies of PMCs at T-I1 with
different nunbers of nucleoli in different species are presented
in Figure 10. Invariably the most fr@ent type were PEW3s with
four nucleoli while the least frequent ones were those with eight
necleoli. However, the frequency of FtKb with at least one
daughter nucleus possessing the rcaxSmrnn nunber of nucleoli was
>Ma in all the species which showed up to 8 nucleoli per T-I1
PMC (Fig. 10) while in the other species that had a r m x h of 7
nucleoli, the frequencies of PMCs containhg at least one
daughter nucleus with two nucleoli was -30%. Such variation in
the frequencies of PMCs with different nunbers of nucleoli (4 to
8) is a reflection of the nucleolar fusion which is very ccmmn
(Sybenga, 1972; Bennett a&, 1973; Darvey and Driscoll, 1972;
Flavell and O'Dell, 1979). The prepondrance of PMCs with 4 /
nucleoli (1 nucleolus per daughter nucleus) at T-I1 is indicative
of the frequent occurance of the fusion of two nucleoli in each
daughfxr nucleus and the accunulation of cells with 4 nucleoli.
Miotic studies, in a spontaneous tetraploid of C. s&in
(isolated by Dr. K.B. Saxend revedld eight prinvlty nucleoli
at P I (Fiq 21 in chapter 4.2.2) further substantiating the
presence of two nucleolar organizers per genome.

!fhe s i z e difference in the two nucleoli in each of the 'ICII
nuclei (Fig-9B) indicates that t h e two nucleolar organizers (in
each of t h e gemmed d i f f e r i n t h e i r ac t iv i t i es . Cultivated
barley is a well known example f o r such differences
(Anastassova-Kristwa &AL, 1977; S u b r ~ y a m and h a d , 1978a;
Linde-Laursen, 1984; Jessop and Subrahmanyam, 19E4). Acoording
t o Beitz (1931). t h e nmber of telophase nucleoli is constant, as
well as t h e nwrber of nucleolar organizers and Sw. chromsomes
f o r a given karyotype. These observations aoquire a new meaning
i n the l i g h t of the evidence t h a t genes d i n g for riboaoual RNA
a r e located in t h e nucleolar organizer region (NOR) and t h e
m a x m nimber of nucleoli per nucleus correspond t o the n-r
of rPNA synthesizing l o c i (Anostasswa-Kristwa, 1977; Nicoloff
ge & 1977; Subratnnanyam and Azad 1978ar b; Flavel l and
OIDellr 19791 Linde-Laursen, 1984; Jessop and Subrahrnanym.
1984). For t h e karyotype of pigeonpea, there have been
mntrovers ia l reports on the n-r of SAT c h r o m m . For
instancer Sinha and K M l a r (1979) reported lack of SAT chronosomes
in certain cultivars of C. and Shrivastava e.t al (1973)
remrded one SW. drromsome per genome in Qiarus while Pundir
(1981) found two chrorrpsomes per genome in Cajaws. ard
U o - species. The present data (Figs. 9 and 10) on the
nucleolar behaviour in PMCs conclusively establ ish two nucleolar
organizers per gemme i n Caianufi and the A W l ~ s b species
studied. Ths differences i n the s i z e of t h e two nucleoli in
daughter nucleimay r e f l e c t differences i r the degree of a c t i v i t y
andlor differences in t h e mult ipl ic i ty c f - R N A cis trons in t h e
two nucleolar organizers i n each of the species.

Controversies in the nucleolar organizer nuu&r bave oft-
been resolved using the mitotic wcleolar nMber as an index of
N.O. activity (Anastasmva-Kristeva rt &.I 1978; Jessop and
Subrahmmyam, 1984: LMeLaursen, 1984). In the present study
nucleolar behaviour during meiosis has been used in dar~nstrathg
the nlmber of nucleolar organizers in a wide range of species.

All the ! 3 j a u s - ~ hybrids that flowered were analyzed
cytologically. Meiosis was studied in detail with enghasis on
metaphas-I pairing, and amphase disjunctions. The results show
the following:
Erms of this hybrid exhibited regular fonration of eleven
bivalents which were predominantly rkrgs (Fig. llE) . There were m significant differences in the chaisma frequency between
cultivar combinations (Table XII), the rwrimnn being 20.75 per
cell in Pant A? x 8. albicans and the minimrm 20.00 per cell in
hybrids with ICP 7035. A low frequency of preoocious separation
was evident in all thg cultivar colrbinations. Diplotene cells
displayed nucleoli with three or four bivalents attached
(Figs. llA-B) , while at diakinesis 20 of the 64 cells contained
nucleoli with three or four bivalmts (Fig. 11C-Dl. Ana&aae-I
and I1 separations in all b e cultivar cabinations were regular
(Fig. lU4) . Pollen sterility in these hyb~ids rang& from 27.3%
in Baigani x A. rlbicans to 46.1% in ICP 7035 x A. rlbicans while
ovule fertility ranged from 21.6% in Pant A2 x A. aU&Scl5 to
32.2% in Baigani x A. albicahs hybrids.

Fig. 11. Meiosis in the hybrid between Cajanus cujan and A t y Z o s i a aZbicans (Bar indicates 10 u): (A) Diplotene nucleolus with four bivalents attached. (8) Diplotene nucleolus with three bivalents attached. (C) Diakinesis nucleolus with four bivalents attached. (D) Diakinesis nucleolus with three bivalents attached. (E) Metaphase-I with eleven bivalents. (F).Regular disjunction at division-I. (G) Regular disjunction at division-11. (H) Quartet showing nucleolar variation, 3-2-1-1.


0 - -- WE; RE; W E ; '" 28 && d & d & 0 0 0 0 0 O O O Q


~iv&&t fonmtion was a general feature in these hybrids
with the e x m i o n of the hybrids between ICP 7035 and A. sericea
which displayed a low frequency (0.4 per W of univalents.
Chiasma frequency varied from 19.38 to 20.37 (Table XI11 . Cme or two bivalents were attached to the nucleolus (Fig. lZA) . Anaphase disjunctions were regular at both the divisions (Fig.12CID))
except h hybrids with ICP 7035. Laggards were observed in 8.6%
of the PMCa. Pollen sterility of 29.7%, 33.6%, 48.4% and 36.1%
and a owle fertility of 54.3%, 61.5%, 46.6% and 52.5% were
recorded in hybrids of A. sericea with Pant M I Ehigani, ICP
7035, and C 11 respectively (Table XII).
Hybrids of Cajaax with A. scarabaeoidea 'showed a low
frequency of univalents (Fig. 13C) (0.264.34lcell) with a nraximum
of two per cell. The chi- frequmcy varied between cultivar
carbinations rangw from 19.48 in Pant A2 x A. scarabaeoides to
20.35 in hybrids with C 11 (Table XI11 . Diplotene/Diakinesis
nucleoli were found associated with up to t m bivalents. A low
frequency (3.8 to 8.6%) of **I cells displayed laggards
(Fig.13D) and irregular disjunction irrespective of the cultivar
used. Pollen sterility ranged from 22.7% in hybrids of
A. scarabaeaides with Baigani to 41.5% in hybrids with 1 8 7035.
Ovule fertility varied from 29.3% in hybrids with ICP 7035 to
52.8% in those with C 11 (Table XII) .

Fig. 12. A-D: Meiosis in hybrids between Cajanus cajan and AtyZosia ser icea .
A) Diakinesis showing 11 bivalents. B) Metaphase-I showing 11 bivalents. C) haphase-I showing regular disjunction. D) Anaphase-I1 showing regular disjunction.
E-H: Meiosis in hybrids between Cajanus cajan and Aty2osi.a cajanifoZia.
E) Diakinesis showing 2 bivalents attached to the nucleolus.
F) Metaphase-I showing 11 bivalents. '
G) Regular disjunction at division-I. H) Regular disjunction at division-11.


Fig. 13: A-F: Meiosis in hybrids between Cajanus cqjm and Aty Zosia ~carabaeoides (bar indicates 10 p) .
A) Diakinesis. B) Metaphase-I showing 11 bivalents. CJ Metaphase-I showing 2 univalents (Arrow) D) Division-I showing laggards. E) Unequal chromosome distribution at division-I. F) Regular disjunction at division-11.
G-J: Meiosis in hybrids between Cajanus cajan and Aty Zosia Zineata.
G) Diakinesis showing 4 bivalents attakhed to the nucleolus.
H) Metaphase-I showing 11 bivalents. I) Regular disjunction at division-I. J) Regular disjunction at division-11.


Hybrids with 8. exhibited eleven bivalents in
the i r FMCs (Fig. la) w i t h a relatively high chiasma frequency
ranging from 20.49 tc, 21.20 depending on the cultivar involved in
the cross (Table XII). Mostly two bivalents were seen associated
with diplotene nucleolus. Anaphase separations a t both the
divisions were regular (Fig. l X , H ) . Pollen s t e r i l i t y in these
hybrids was relatively low (14.6% to 29.1%) with high ovule
f e r t i l i t y (63.7% t o 85.2%) (Table XII) .
These hybrids shaved regular bivalent formation (Fig 13H)
except in combination with ICP 7035 where a l o w frequency
(0.09/cell) of univalents were found. Chiamata per cell varied
from 19.70 t o 20.34 depending on the Caimu genotype
(Table XI11 . A nmximum of four bivalents attached t o the
nucleolus (Fig. l3G) were found a t diplotenddiakinesis. Amphase
segregation was r w a r a t both the divisions (Fig.131,J).
Pollen s t e r i l i t y varied from 27.5% in hybrids with C 11 t o 46.2%
in those with I B 7035 while ovule f e r t i l i t y ranged from 31.3% in
hybrids with ICP 7035 against the rwtimum of 72.7% recorded in
hybrids with C 11 (Table X I I ) .
A range of w i o t i c abmnmli t ies were encountered in
C. &an x B. hybrids (Fig. 14) . These hybr idk
disp1i:wJ a relatively hiqh frequency of univalents (up t o
(Fig.14D) and rod bivalents (3/cell). The chaiana frequency was

Fig. 14. Meiosis in hybrids between Cqianus c a j m and A t y Z o s i a grandifozia.
A) Diakinesis showing 4 bivalents attached to the nucleolus.
B) Diakinesis showing univalents (Arrow). C) Metaphase-I showing 11 bivalents. D) Metaphase-I showing 6 univalents. E-H) Laggards at division-I. I] Bridge at division-I. J) Division-I1 showing chromatic separation but
no disjunction in one of the telophase-I1 daughter nucleolus.
K] Telophase-11. L) Five daughter nuclei at division-11.


low (1Wcell) (Table XI11 . The diplotene nucleolus wae often
found with t ~ ~ r three or four bivalents attached (Fig 14A).
Bridges (Fig. 141) were found in 8.2% of the cells a t
anaphase1. Bridges were rarely accorrpenied by fragments. Other
Anaphase irregulari t ies included laggards (Fig.14EIF) mz l t ip l a r
disjunctions (Fig.14L) and occassional restitution a t division-I1
(Fig. l 4 J ) . Lagging chromosomes were commn in t h i s &ination
(Fig. 14E-H) . Division-I laggards ranged from 2 t o 6. A t
division-I1 chromosom segregation t o f ive poles was frequent.
Restitution i n one of the T-I daughter nuclei was occassionally
found (Fig. 14.3.
These hybrids whibited a high degree of pollen s t e r i l i t y
and poor ovule f e r t i l i t y . While the pollen s t e r i l i t y was 54.8%
and 48.3% in hybrids with Pant A2 and C 11 the corresponding
figures for ovule f e r t i l i t y were 23.4% and 19.9% (Table X I I ) .
Univalents were m r e frequent (Fig.15B) than in hybrids with
Asian but the maxirrPlrr1 nrmber of univalents recorded was
4 with an average of 1.12 in hybrids with Pant A2 and 0.75 in
those with C 11. R d bivalents averaged 2.89 and 3.89 per c e l l
in cabination with Pant A2 and C 11 respectively. The chiasma
frequencies were 17.97 and 17.33 involving cross6 with Pant A2
and C 11 as farrale parents.

Fig. 15. Meiosis in hybrids between Cly'anus cqjan and A t y Z o s i a Z a t i s w p a k (bar indicates 10 y) . A) Diakinesis . 0 ) Metaphase-I showing 11 bivalents. C ) Metaphase-I showing 4 univalents. D) Laggards at division-I. E) Three chromosome groups after division-I. F) Five daughter nuclei at division-11.


AMphaseI laggards were frequent (Fig.15D). A comrrcn
feature observed in these EMCs was congregation of the separated
chrwosomes at the center of the cell and subsequent segregation
Occasionally into m r e than two groups (Fig. 153. Five groups of
ChrOImsomes were often seen at division-I1 (Fig.15F).
In general precocious separation of bivalents was a general
feature in all hybrids with all A t y U s b species where ICP 7035
was the female parent and heteromrphic bivalents were c o m n in
all the hybrid conbinations. Secondary chromsome associations
were found in hybrids with the Australian At-. Differences
in staining intensities in partners of bivalents was detected in
several hybrid cahinations.
A wide variation in nucleolar nmber, size and distribution
in EmCs at division-I and division-I1 (Fiy.16.17) was recorded.
Such a variation w& detected in the hybrids between C. and
A. albicans (Fig. 161, C. and A. (Fig. l7A-D) and
C. s&dn and A. aranclifolia (Fig. 17E-H) . In these hybrids the nucleolar n&r varied from 4-8 at
-1. At T-I1 nucleoli in daughter nuclei varied from 0-4
(Fig. 18) wfiile the total nwber of nucleoli per PMC varied from
4-8. Consequently in 14% of the Pm38 of C. .&an x A. lllbicans,
6% in the FWa of C. caj,an x A. lineata and 4% in the FtCs of
C. a x A. w r d u hybrids the nucleolar distribution at

Fig. 16. Nucleolar variation in pollen mother cells of the hybrid between Cajanus cajan and A t y Z o s i a aZbicans (Bar indicates 10 PI.
A) Telophase-I nucleolar distributions, 4-4. B-I: Telophase-I1 nucleolar distribution B) 1-1-1-1. C) 2-1-1-1. D) 2-2-1-1. E) 4-2-1-0.
(Arrow indicates nucleus devoid of nucleoli). F) 2-2-2-1. G) 3-2-2-1. H) 4-2-1-1. I) 2-2-2-2.


Fig. 17. Nucleolar variation in pollen mother cells of hybrid; between Cajanus cajan and AtyZosia Zineata and Cajanus cqian and AtyZosia grandifoZia.
A-D: Cajanus cajan x AtyZosia Zineata.
A) Telophase-I nucleolar distribution: 4-4. Telophase-I1 nucleolar distribution.
B) 3-1-1-1.
C) 3-3-1-1.
D) 2-2-2-2.
E-H: Cajanus cw'm x AtyZosia grcmdifotia.
E) Telophase-I nucleolar distribution: 3-1. Telophase-I1 nucleolar distribution.
Fb3-2-1-1.
G) 2-2-2-1.
H) 4-2-1-0 (Arrow indicates nucleolus devoid of nucleolus'


Fig. 18. Frequency distribution of nucleoli in telophase-I1 daughter nuclei of hybrids between C a j a u s cajan and Aty Zosia species.

0 1 2 3 4 0 1 2 3 4 0 1 2 3 4
NO. OF NUCLEOLI

T-If w a s amfined t o three wt of the f w r daughter nuclei. The
frequencies of daughter nuclei devoid of nucleoli were 3.5% in
C. m x A . albicans, 1.5% inC. a x A. UneaUi and 1% in
C. x A. aranalfolla . . . A majority (65% in C. &an x
A* nlbicans, 59% in C. a x A. lineata a d 54% in C. &an x
A. arandifolia) of daughter nuclei contained one nucleolus while
the remainder had 2, 3 or 4 nucleoli. The frequencies of
daughter nuclei with two nucleoli were 22% in the hybrid C. a x A. albicans, 29% in tha t of C. &gm x A. linsata and 36% in
C. x A. arandifolia, while 6% in C. x A. Rlbicans, 8%
in C. c&an x A. lineata and 9% in C. Egjan x A. arandlfolra . .
included 3 nucleoli. Least frequent (2% i n C. a x
A. albicans, 3% in C. a x A. lineata and 3% in C. x
8. were the daughter nuclei w i t h 4 nucleoli.
The cells with variation i n nucleolar nrmber also showed
variation i n nucleolar size. The nucleoli could be classif ied as
large or -1. In the R3Cs with 4 nucleoli a t FII, (one in J'
each iiaughter nucleus), the nucleoli were uniformly larger
(Fig.16B) than those in the daughter nuclei w i t h 2 or more
nucleoli. There was no discernible variation in the s i ze of the
larger nucleoli, while variation was evident i n the s ize of the
wlaller nucleoli. A wide variation was recorded for nucleolar
distr ibution a t F I X (Fig. 16 and 17) as against the regular
2-2-2-2 distr ibution recorded in the parents.

8 1
4.2.1.2. BIgAVTm IR P2
F e r t i l i t y of the P2 segregants in the f i v e pplations,
X As-, C.- X A*-, C . U X
A. scarabaeoides, C. &~II x A. lineata and C. &dt~ x
8. is presented in Fig.19. The f e r t i l i t y in t h e
segregants ranged from 18 to 81% except i n those of C. &dn x
A. caianifol.ia where the segregants were relat ively m r e f e r t i l e
with 106 plants showing >71% f e r t i l i t y , 95 plants with >56%. In
t h e other four p p l l a t i o n s (Fig.191, the majority of plants
showed 41-70% f e r t i l i t y w i t h a few plants i n t h e lower (10-25%)
and t h e u p r (>71%) ranges. Meiotic analysis, such as metaphase
pairing studies, were conducted on randomly selected plants from
t h e various groups of segregants.
I n randomly analyzed plants in t h e higher f e r t i l i t y group
(>56% f e r t i l i t y ) , chromsome pairing was m s t l y mrrrurl with the
formation of eleven bivalent6 which predominantly were of t h e
ring type. Unpaired chromosomes were noticed occassionally i n
very few PMCs. Unpaired c h r o m s o m were m r e frequent in the d'
lower f e r t i l i t y g roup . Sane rrmltivalent configurations were
found in some of the segregants of t h e C. x A. sericea
p o p l a t i o n while secondacy chrom~sow associations were c o m n in
the segregants of C. sajan r A. scarabaeoiaes and a l so to a less
extent in other ppulat ions. Chrommm pairing was regular in
al l t h e analyzed segregants of the f. &au x A.
popla t ion . In general pairing was nosml in the segregants of
the popllations of C. && x A. and C. x
A. lineata except i n the i r lower f e r t i l i t y group. H ~ t e r ~ r O r p h i ~
bivhlents were camon in several cases.

Fig. 19. Frequency distribution of F2 segregants in different fertility groups.

.ty Zosia aZbicans
sericea
30
20 A t ; i Zosia cqianifoZia
10
0 -25 tC-(DILSl -lo w m
FERTILITY ( V r )

Meiotic studies in the intergeneric hybrids revealed regular
chromsome pairing in hybrids of Cajams with Asian m. In these hybrids eleven bivalents were c o m n a t uetaphase1. A s
in tbe parents ring bivalents predominated in these hybrids.
Chi=& frequencies in hybrids with Asian &yba&as was high and
m w - a b l e t o the values in the parental species. Fedak (1982)
has shown significant variation for chiasrrra frequency between
cult ivar cubinations in Barley-wheat hybrids but such
differences were not evident in the present f&aws--
hybrids.
knother interesting feature in these studies has been the
behaviour of the Cajxus cultivar I 8 7035. .Invariably in
hybriiis with all the ALykaia species where it w a s used as the
£ m e parent the pollen s t e r i l i t y w a s consistantly higher.
Chiasra frequency w a s also found to be lower in hybrids involving
this cult ivar and the occassional univalents and anaphase
i r r e g d a r i t i e s en'countered were also in hybrids w i t h this
genotlpe. In fajaws intervarietal crosses also the behavaiour
of I B 7035 is reported to be distinct (K.B. Saxena, Pers.
corn.) . Shrivastava aLr (1973) and Sinha and K- (1979)
found significant differences in k a r y o t p of G&nu6 cultivars
and -3uivastara & a, (1973) who conpaced the karyotype of
A. lir.eata with 15 cultivars of Sajmu6 found that the karyotype
of &. lhe&a was closer t o karyotypes of some !Xjanm cultivars
than t o othels. It is likely that some differences ir the
chramoscme conplement of I 8 7035 are responsible for the

distinct behaviour of this cultivar. The differences between
cultivar ambinations for pollen sterility and wule fertility
are V t e d and attributed to the effect of the genotype. A
parentage dependent pollen sterility was also reported in
interspecific crosses of (Xiazuma and Onor 1980).
In general chromsome pairing and disjunction is regular in
hybrids between and species of Asian m. Meoitic studies suggest that the Asian share a cnmron g m e
with C. &. Absence of univalents in hybrids is an indication
of chrornsome to chromosome -logy between the parental
genomes. Pairing at diakinesidmtaphkse gives only an
indication of the gross-chrorrosorce -logy but does not provide
information on the extent of homology. Pachytene FUCa of the
hybrids (Fig. 20B-F) also did not Bbow univalents indicating a
hornlogy of a high degree between C. &an and species of
m. H?wwer, intercalry unpaired regions (Fig.20B,C8E)
were evident in the pachytene chromsornes. Similar observations
were made by (1981, a,b,c).
It has been shown in som cases that chromsome horrology is
not a prerequisite for pachytene pairing. For -1e in
mna-haploids of -ley the seven chrornsoms participate in
pairing at pachytene thereby iPplying the occurrence of
Won-Hcuvlogous pairing" that does not persist until wtaphase-I
( W i v a i a h and Rasha, 1971). Such non-homlogons associations
at pachytene have also been reported inmonoploids of rice (Chu,
19671, tomto (Ecochord e% al, 1969) and maize (Ford, 1970;
weber and Uexandrr l5W.l Lr mlvhar,!c~ids, e substantial

Fig. 2 0 . A) Two nuc leo l i a t pachytene i n Cqjaus cajan.
BBC) Pachytene i n hybrids between Cajaus cajan and Atylosia grandifotia showing inversion loops ( s o l i d arrow) and unpaired regions (dotted arrow).
D) Pachytene i n hybrids between Cajanus caJm and AtyZosia ZatisepaZa showing cross shaped con- f iguration (Arrow).
E) Pachytene in hybrids between Cajanus o a j m and Atylosia l ineata.
F ) Pachytene i n hybrids between CaJ'anus cojan and AtyZosia c a j m i f o l i a .


reduction in chramb.aaal pairing from prophase-I to metaphase-1
is seen and such a raction in pairing has been attributed to
non-homologous pairing (Subrmyam, 1978).
In species hybrids univalents and other higher chromosome
associations are usually frequent and these japairments in
pairing are taken as an index of genome affilities or the lack of
it. Meiotic studies in distant hybrids have helped in
establishing evolutionary and species relationships in several
crop species which include m y leguminous species (Foujdar and
Tandon, 1976: Machado &dl, 1982; Newell and Hymwitz, 1983).
In the hybrids of Cajaui and the Asian Atvlosias.
chronvsome pairing at metaphase has been perfect implying a very
high degree of genome homlogy. A. which is
nvrpbologically very close to Qjau,s shares ' a close genome
relationship as well with the cultivated species. A. -, A. lineata and A. sericea though morphologically distinct from
pigeonpea also haves close genome relationship with fajau&. It
was only in the hybrids of C. G&U x A. scarabaeoides that
consistently univalents were found irrespective of cultivaz
ombination. This distinctly suggests that one drromsome of
A. has diverged to a relatively greater extent from
its mterpart in the Cajams wrrplerrent. The differences in
staining intensities between partners in a bivalent and also the
qpearance of heteromrphic bivalents in the hybrid PElCs is an
indication of intergenomic hornlogy .

-phase separations were highly organized in all the
CajaDU cultivar. curbinations involving A. alblcans (Fig. 11F),
A. aericea (Fig. 12C), A. lineata (FIg. 131) and A. caianifolia
(Fig. 1x1 with the hybrids with I 6 7035 background being the
exception where a low frequency of anaphase abmnmlities rmstly
laggards were observed (Table XII). Anaphase irregularities at a
low frequenoy were encountered in hybrids with A. acKabaeoiden
in all the cultivar oorbinations. w e n t l y the sparse horology
between one of the chsonvsomes of C. and A. scarabaeoides
as evidenced by univalents at metaphase contributes to these
irregularities at anaphase-I. AMphase-I1 separations were
aU?ost regular (Fig. 11G, 12D, lW, 13F, with occassional
asynchromy in disjunction in a few cases. These findings, i.e.
regular chromosome pairing and disjunction in the hybrids between
f. and Asian Btvlosias. are in line with those of earlier
studies on hybrids of Saiaws-Atyhxia hybrids (K- et aL,
1958; Pundir, 1981) . As against the regular meiosis encountered in the hybrids
with Asian the hybrids of fajaws with the Australian
species of for which no previous reference was found
exhibited a relatively high degree of pairing and disjunctional . .
abnonmlities. Univalents hybrids with both A. arandlfolla
end 8. were far nvre frequent. As many as 6
univalent6 were recorded in hybrids with A. arandifalia and up to
4 in those with A. -. These findings suggest that at
least 3 chromsomes of A. clrandifalia and 2 of A. have
d;-;cr.ged frop their wunterrsrts ir? thc L&i,iii& guiOl!,l!e.

m e n t l y less effective crossing w e r due to lack of adequate
hmlogy has contributed to the higher frequency of univalents.
However, from the present findings it is difficult to ascertain
whether the chsorrommes of A. and A. orandifolia that
have diverged from C. &an are similar or different from each
other. Hybrids between these species of can help in
resolving this question.
A significant obsenration has been that the univalents in
the hybrids of with A. ~ & a d & s , A. orandifolia or
A. 1- were always even nunbered (2, 4 or 6) which is
indicative of a low n m b r of pairing initiation sites
[ (Zygomeres) Sybenga, 19661 . Studies on pairing in tetraploid
C. & revealed a high degree of bivalent formation (16.54 per
cell) and a fertility of around 60%. Quadrivalents averaged only
2.73 per cell with the maximrm being 4 per cell. The selfed
progeny of the tetraploid exhibited regular disjunction (Fig.21E)
and high fertility (70%). The high freequency of bivalent
forination in the tetraploid also inplies a few pairing initiation
sites in pigeonpea similar to that found in &edkag~ sativa
(Bingham and Gillies, 1971).
An interesting feature in the PMCs of C. cajza x
A. arandifalia hybrids is the fonmtion of chromtin bridges at
division-I with or without fragments. Meiotic bridges are mstly
a consequence of reconbination within a paracentric inversion
(KKlintock, 1933; 1938) or alternatively in very rare instances
d : t to pruphase breakage of chrorasoms often referred to as 'U'
t i ~ c exchanges (Lewis and John, 1966). The distinguishing

Fig. 21. Meiosis in tetraploid Cajanus cajm.(bar indicates 10 p)
A) Diakinesis showing quadrivalents (Arrow). B) Metaphase-I showing quadrivalents. C) Irregular disjunction at division-I. D) Progeny of tetraploid showing regular bivalent
formation. E) Regular disjunction in the progeny of tetraploid. F) Eight nucleoli at telophase-I. G) Pollen grains of diploid Cajanus cujan. H) Pollen grains of tetraploid Cajanus cajan.


feature of the *U1 type excharqes fran those arising out of
crossing wer within a paracentric inversion is the presence of
fragments of variable size following 'U' type exchanges.
Observations on pachytene rn of the C. caianus x A.
arandifolia hybrid (Fig. 2OB,C) also revealed loops which indicate
that bridges at anaphase are a consequence of inversions.
Further ' both the parents involved in this cross have exhibited
regular bivalent formation and disjunction. The limitations in
chrormmme size and the inconsistent a e r a n c e of the fraqments
in the present material did not permit precise fragment size
measurements. Chromatin bridges are comaon in species hybrids
(Ahmed & dl, 1977; 1979; Mdlik and Mary, 1975; Chaudhuri gt
a, 1976; Reddy and Subrahmanyam, 1985; Sprirger and Buckner,
1982) but this is the first report in the Cajauw - Atybaia
hybrids.
These results indicate that structural variation in the form
of inversions have played a role in species differentiation
between $2. &an and A. crrandifolia. Meiotic bridges are only a
qualitative index of inversions but not quantitative. Not all
paracentric inversions mnifest themelves in the form of
bridges. A minimum of one crossover in the inverted 6-t is a
prerequisite for bridge formation and the possibility of a
crossover is directly proportional to the size of the inverted
sepent .

The structural differences between the chraw- of
C. cdjdn and A. arandifolia as evidenced by the a n a m bridges
and the relative lack of hamlogy bet- C. and
A. arandifolia and A. which is -rent from the
higher frequency of univalent6 and anaphase abnormalities in
hybrids between C. and these species explain the relatively
high deg'ree of sterility encountered in these h*rids. This
suggest that the genomes of the Australian species of Atyksja
are m r e diverged from Sajmw than tbose of Asian Atyhshs.
The regular foemation of elwen bivalents in the hybrids of
Gajanu with Asian without any detectable conventional
meiotic irregularities might be interpretted to indicate caplete
gemme b l o g y between these species which can be misleading as
will be wident from the following discussion of,our results on
nucleolar variation.
The most intriguing feature in the hybrids of S&rui with
A. dbkana, A. lineata, and A. was the association
of up to four bivalents with the nucleolus at
diplotenddiakinesis and the wide variation in nucleolar nlmber,
distribution and size. Karyotypes of L saj.au and tbse of
A. -" and A. have two pairs of satellite
chromsomes in each of their gem= (Pundir, 1981). Cur studies
on nucleolar nmbers at telophase have confirmed this finding and
have revealed a similar situation for A. crrandifolia. Thus an
association of nucleolus with a rnaxinum of two bivalents at
prophase-I and a mxinnnn of four nucleoli per PMC at telophs-I
are a-.- = i ? t h c : ~ bit*-???. The asscci~ticc sf ZG;; i. -. t,b

bivalents with the nucleolus in the hybrid -11s is suggestive of
pairing between the N.0.s of Cajmw s&an and non-N.0.s of
Atvlosfa species or a ~ e ~ s a and the variation in nucleolar
nuher and distribution at telophas-I and 11 is interpreted to
have originated from allosyndetic recombination.
The possible nucleolar distributions with 4 N.O.s, with and
without reoombination, are presented in Fig.22. If one assumes
no ~~nbhtion between N.O. and a non-N.O. (Fig. 22:I), a
meximum of 4 nucleoli can be expected at telophas-I. The
presence of 4 to 8 nucleoli at telophase-I ccrrpels us to suggest
allosyrdetic reconbination. Furthemre, if the N.O.
chromsomes are not involved in reconbination there are only
three possible telophaseII distribution patterns. However, the
observed deviation in nucleolar distribution at telophase-I1 fits
well with the theoretical expectations assuming crossing over
involving 1, 2, 3 or 4 N.0.s with non-N.0.s (Fig. 22:II-V). Our
interpretation that variation in nucleolar nuuber at telophase-I
and telo@ase-11, &d the variation in nucleolar distribution at
telophase-I1 result from reconbination between a N.O. of
c. and non-N.O. of A t y h ~ b species or vicwersa is
su~ported by the fact that the nucleolar nunher never exceeded
eight, which is the maximrm that can be expected with 4 N.0.s
(i.e. 8 satellite chromatids). Nucleolar distribution to only
three of the four daughter nuclei in a given Pm: (Fig. 16Ej 17Cr
H and Table XIII) can occur only when the N.O. chrorroEune6 are
involved in reconbination with mn-N.0.s. Additional proof for
recok4inatlnn core. ' , . . v the telophase-11 ! ,- 'ri which have L'

Fig. 22. Possible nucleolar variation in pollen mother cells of the hybrids between Cujanus cnj'an and A t y Z o s i a species without recombination and following recombination between nucleolar organizing and non-nucleolar organizing chromosomes.

nucleoli (Figs. 16Et 11 and 178) . Frcm Pig. 22 it is evident that
without reanbination there nwer exists a possibility for more
than three nucleoli per nucleus at telophase-I1 when there is
nucleolar distribution to either three or four nuclei. The
occurence of three or m r e nucleoli per telophase-11 nucleus
might suggest a possible expression of otherwise latent N.0.s of
the parehts. But the presence of a total of eight nucleoli per
EmC with a regular 2-2-2-2 distribution in the parents and the
absence of PMCs with m r e than eight nucleoli in the present
hybrid rules out such a pssibility.
A nmjority of the PMCs in the hybrids exhibiting nucleolar
variation had only 4 nucleolit one in each daughter nucleus at
telophase-11. Hence it can be presumed that the no& N.O.
chromosome disjunction without reconbination is 2-2 (Fig. 22: I-C)
resulting in 2 nucleoli in each daughter nucleus thaton fusion
give rise to one nuclwlus. Nucleolar fusion is m n in
several plant species especially in meiosis (Darvey and Driscollt
1972; Sybenga, 1972; Bennett SL al, 1973; Darvey gL al., 19731
Flavell and O'Dell, 1979; J e s q and Subrahmnyamt 1984). The
remaining P.M.C.8 undergo allosyndetic reconbination with respect
to the N.O. involving a minimrm of one N.O.-chromosome and thus
deviate from the expected nucleolar n W r at telo@mseI and
distribution pattern at telophaseI1 assuming no reconbination.
The total n-r of nucleoli at telophase1 in a Fm3 is a direct
indication of the n m b r of N.O. chromss011~~ involved in
recohination. Although we obtained r variation ranging from 4-8
?q7c!ml i at telo@s=cT (Fi? 16A, I - ' it b'r= r~r't possihl~ tc~

score emugh c e l l s a t this stsge to mable u s to arrive a t the
frequency of reconbination p r a c t s . This w a s because it is
d i f f i cu l t t o get a PMC a t telaphese-I w i t h p r h s r y (unfused)
nucleoli owing to the short meiotic cycle in pigeonpea. In the
cells containing 8 nucleoli a t telophase11 the distr ibution
pattern of 4-2-24 (Table X I I I ) is only possible when a l l the
four N.O'.s a re involved in reombination with non-N.0.s.
The presence of 5, 6 and 7 nucleoli a t telophas-I1 many a
time w i t h variable s ize is due t o nucleolar fusion. In the l ight
of the different possibi l i t ies (Fig.22) the pattern of nucleolar
fusion could not be assessed. Although in the daughter nuclei
nucleolar variation of 0-4 existed a t telophase-I1 (Figs 16 and
17), rrost of the daughter nuclei a t the quartet stage showed a
single nucleolus with a few quartets (Fig.11H) ahowing 2 o r 3
d i s t i nc t nucleoli. Since the overall variation of nucleolar
nurrber can be attributed to both fusion and/or allosyndetic
reconbination, an estimate of such recorrbination a u l d not be
obtained. r evert he less, for any given nurrber of nucleoli a t
telophase-I and nrnrber and distribution of nucleoli a t
telophaseI1, the minimin n-r of N.0.s involved i n
allosyndetic recurbination could be given.
Another interesting feature in these hybrids was the
varist ion in nucleolar size. Variation in the nucleolac size,
e s ~ ~ i a l l y the presence of BMYer nucleoli than the nonnal
f o r r , 4 by a single N.O. chromatid (Fig.161), can be attributed
t c cither a crossover involving a break within the secondary
cori;triction or a crossver between two N.0.s resulting in the

Table YIIINucleolar variation at telophase-I1 in a hybrid between C. %?a and A. albicms.
I No. of nucleoli I , -
I I
4 / 5 \ 6 ; 7 1 8 I I
I I I I t
I 11 1 26 62 1 14 I 46 I
I I I
I I I I 39 8.8 [ 28.9 6.9 16.4
I I I 1
P.M.C1s NO.
0)

formution of a chromatid with tvo N.O. termini leading to
mcleolar domFnance (differential anphiplasty). If the first
alternative is assumed one wuld expect more than 8 nucleoli.
Absence of PMCs with more than 8 nucleoli rules out such a
possibility. Thus we are inclined to suggest that nucleolar
cbminance is operative in the present h*rid. There have been
several 'reprts on differential anphiplasty in interspecific
hybrids, wherein nucleolar expression of one of the ~pecies
involved in the cross is supressed (Navashin, 1934; Keep, 1962;
Subralnmnyam and Azad, 1978a.b). Partial supression of a N.O.
leading to a reduction in nucleolar size has been reported by
Nicalof f & & (1979) in their barley translocation lines.
As indicated earlier in spite of the profuse flowering and
regular meiosis the high degree of pollen sterility and low seed
set in the hybrids of Sajaus with Asian AtyUshs is surprising.
Structural heterozygosity in the cluommmes of the parents as
evidenced by the allosyndetic recorrbination in the present
hybrids could b'e the major reason for the sterility, since the
pllen fertility was mch lower than expected on the basis of
m& bivalent formation and disjunction suggesting thereby,
complete h l o g y is not a prerequisite for regular bivalent
formation. The absence of nucleoli in som of ,the daughter
nuclei clearly indicate difficiency for the nucleolar organizer
while the presence of four nucleoli in a daughter nucleus is
suggestive of duplications for the nucleolar organizer. AlthDugh
these two are the exclusive conditions which indicate deficiency
L-t.;i..ptidi~, d . ~ .d,(je ~i k d i a t ; - . . L ~ U O L B ~ EJ;LSU inclucirs

such possibilities. It has already been demonstrated (Burnhw,
1950) that translocations involving satellite c h r o m s m ~ ~ result
in duplication-deficiency aberrations leading to pollen
sterility. Stebbins (1971) disassed the causes of sterility in
distant hybrids and proposed that "cryptic structural hybridity"
could be the major contributing factor for sterility in the
hybrids 'kssuming that the parental species differ with resped to
many .mall chrorrpx,mal rearrang-ts such as interstitial
translocations. Since then several workers on hybridization have
attributed sterility in spite of normal bivalent formation and
disjunction in their hybrids to a series of gmall structural
differences in the parental chromsoues. Our results provide a
conclusive evidence for that possibility.
The cryptic structural alterations involving .the satellite
chromosomes and chronosomes that pair with the satellite
ch~omsmes were discernible in view of the advantage of the
nucleolus act- as a marker for the products of reconbination,
while reexbination'arising out of structural alterations in
other chrcmvsomes is not traceable at later stages in the absence
of any such cytological marker and hence the structural
alterations traced with the aid of the nucleolus form only a part
of the total.
Regular meiosis in the ~-~ hybrids Micate a
dose relationship between the cultivated and the wild species.
The high frequency of recabination in the hybrids further
confirms the proximity c>f the parental species and suggests that
the species have differentiated through structural alterations of

chrowsomes. Cur findings s w r t the carclusions of - other
workers that the taxonomic separation of and
should not be wintained ~~, 1975; van der Maesen, 1981) . Cur findings on the chromosome pairing and reambination in
the S a j m u s - m hybrids clearly indicate that A.
is the closest relative of G. e. The regular ch ro l l ~sa~e
pairing in the hybrids of Sajaus with A. caianifolia and a
relatively high degree of f e r t i l i t y inplies a close genome
relationship between these two species. "Cryptic structural
differences" i f any between these genome6 are less than i n the
other species. On the other hand the relatively high degree of
pollen s t e r i l i t y in the hybrids of C a j a u s with the other species
of Asian m, which is unaccountable on the basis of
n-eiotic pairing and disjunction in their hybrids, is a
consequence of gametic duplications and deficiencies resulting
from structural differences in the chromo&me~ of the species
involved and consequent reconbination which has found clear
cytological evidence in the pairing of N.O. chromosones with non
N.O. chromsomes and the data on nucleolar variation. Hence the
four A t y k s b species, A. nlbicans, 8. Ilneata, A- Bcarabaeoidefi
and A. sericea which have differentiated through structural
heteroygosity are closely related to but m r e distantly
than 8. caianifolia. &mng these four species, a l l of which
differ from on the structural basis, it is difficult to
establish the hierarchy of relationship with respect t o CaianuR
since in th i s material the limitations in chromsome size dces
2 . . -. 1.-2::: :i t: ,-L-.tify ttre s t r b c t u r a l difftit, .~,. L s o the

differences in sterility between hybrid ombinations are not
different enough to delineate their closeness of relationship.
Since univalents though at a low frequency, acconpanied by a low
frequency of anapbase abnormalities were found in all the
cultivar cabinations of - A. Bcaraboeides hybrid it is
possible that the genome of A. scaraboeides is relatively more
distantly related to than those of the other three
species. A. albicans and 8. lineata share a om-on feature i.e.
structural heterozygosity with respect to the nucleolar
organizing chrmmnes. Both the Australian & y l ~ & ~ used in
our study are cytologically distinct £ran with respect to
the Asian &y l~& species. The structural heterozygosity
between the genomes of Qjanw and the Australian
discussed earlier in this chapter contribute to the high degree
of sterility in these hybrids.
Our results clearly krdicate that pollen sterility and ovule
fertility in the hybrids is a reliable index of genm
affinities. FO; example the hybrids with A. caIanifolia which
were cytologically mst stable had the highest pollen and ovule
fertility whereas hybrids with A. clrandifolia and A. J&bp&i
exhibited the highest degree of meiotic abnornvllities and the
highest pollen sterility 'thereby inplying that the levels of
garretic sterility reflect the degree of divergence of the
species fran C. &an.

Pgatt from the W e major general trerds the hybrids also
exhibited a few cgtological features which merit discussion in
stmne detail. OccassionaZ meiotic restitution was observed at
division-= in the figbrid C. x A. w. Restitution has been rwrtcd in several species hybrids (Li 9t Blr 1964;
Witgenarr, 1968a, b; Hann and Sasakuim, 1977; Riley and Chapan,
1958; Islam and Shgherd, 1980). Meiotic restitution could be a
result of delay in spindle fonmtion or due to spindle fusion.
Restitution is also possible when the t h + required to reach
univalent accrrrrulation exceeds the period of activity of
kinetochores resulting in failure of anaphase movanents as has
been suggested by Islam and Shepherd (1980). In the present
instance the occassional restitution might be h e to either
spindle fusion or delayed spindle fonmtion. If $he restitution
nucleus h a w to form a fertile gamete as has been reported in
species hybrids of Arachis (Singh and b@ss, 1984) a faw triploids
can be expectd in the F2 progeny of C. &an x A. m. . .
(hrormsome ~mvemmt to five poles in hybrids of !2ajaum with the
Australian is also a consequence of spindle
abnonmlities. Fosmation of chromosome sub groups resulting in
m r e than four spocade has been reprted in allopolyploids, for
exanp?le Buhus. (Bwi, 1965) and GUl&a (Lh, 1973). Such a
situation has also been reprted in a trihaploid of Hordeurn
(w m d Subrahmanym, 1985). In this case the
plyads w r e a -ace of genome specific organization at
division-I. In the present hybrids the occurrcnoe of ~ m r e than
four sporads result from alte s?indle organizatior.

In all the five --- hybrids from which a F2
population was raised, a majority of the segregants showed an
inproved level of fertility over the Fl hybrids while a few
segregants did show fertility less than that in the F1.
Redovery of fertile segregants fran the progeny of partially
sterile interspecific hybrids has been acamplished in genera
like Nicotiana, Tm&xam . . , (Stekbins, 1950) and also in
species crosses of wlilotus (Webster, 1950) and
(Steins, 1958). Such results have also barn reported in the
segregants of hybrids between x E. coccinw
(Ilaq & dl, 1980) and in crosses involving lonaialllmic; and
puaa (Yamaguchi gt Bl, 1976).
Stebbins (1958) suggested that the F2 segregants which show
inproved fertility over the Fl have a~pacently recovered a
relatively higher degree of genic balance than tbe one existing
in the F1. A failowup of this interpretation would be whether
the opt- genic balance corresponds to the balance that existed
in the parents or whether it denotes a new balance. In the
present case several of the F2 segregants which shcrwed -roved
fertility were wrpholqically distinct fran the wents, with
the F2 poprlations showing a high degree of vaciability for
different characters, a point which will be discussed in mre
detail in the cbapter 6. The remvery of fertility in the
segregants without recovery of parental morphology suggests that
rj-n recover& pciiic t~ , .~ . - .e A&. a new orrs. lnlr can be oonfrrmfu

by crossing the f e t i l e derivatives to the or ig ina l parents i.e.
back crossing and i n case t h e derivatives have acquired a new
balance, the progeny of t h e back CIOSS w i l l again be expected t o
be p a r t i a l l y s t e r i l e .
Secondary association of bivalents w a s evident in some F1
hybrids and w a s prevalent in e3me F2 segregants. Secondary
associations appear due t o the presence of genetically and
s t ruc tura l ly similar chrorro50rpes (Riley, 1960) and t h e bivalents
involved i n such associations have "residual homology"
(Darlington and Moffett, 1930; Thonvrs and Rwel l , 1946).
Kenpanna and Riley (1964) with the a id of te locentr ic chromsoms
danns t ra ted t h a t secondary associations occur between
genet ical ly related bivalents and that they a r e independent of
s imi la r i t i es in t h e s i z e of bivalents. I n t h e present material
t h e secondary associations further re f lec t t h e presence of
s t ruc tura l a l t e ra t ions in the related genomes.
The aperance of rmltivalents in some of t h e segregants
could be an a r te fac t of chromsorne s t ickiness s ince never could
we observe any clear corifigurations of higher associations.
As is evident from t h e resul ts and t h e above discussion
there is no chromosome elemination in any Saimu-Atybsia
crosses. Kasha and Kao (1970) reported recovery of haploids from
the cross Hs&em Y U ~ I ~ E x H. buEQsm and Sbr-yam ancl
Rasha (1973) reporte t h a t the haploids from this cross a r e a
consequence cf gra3,rl ant! selective elimination of the b l b ~ s u m
chrcrc.sc~r,~r frorr, t v t,ric' ert -yo-. In the last ! YePr? si.?r<

the se lec t ive chromosom elimination wthod oi haploid production
-nly kKmn as the "Bulbosun methodm has caw to l i g h t there
has been no r e p r t of haploid prcduction by this method without
t h e involvenent of a I u d a m species. I n both wheat (Barclay.
1975) and (Bother & & 1984) two other species
where haploid have been rewvered & the Bulbosum method,
species ' of tIordeLln a r e involved. In t h e genus Nicotiana
s e c t o r i a l elimination was recorded in interspecif ic m r i d s
(Gupta and Gupta, 1973) . A systanatic screening for select ive
chromosome elimination both in the present study in pigeonpea and
also i n the genus &&&is (Subralnnanym, unpblished and Singh
and Moss, perrs. comn.) there has been no evidence of
elimination. Thus select ive chrormsome elimination appears t o be
of limited w l i c a t i o n in haploid production.


The segregation and inheritance of the following qualitative
t r a i t s was studied in the hybrids of C. rainn x A. - I
C. S&II X A* lineatat C. x A. -, C. X
A. sericea and C. &an x A. caianifolia. The fanale Sdjmu
genotype i p a l l the cases was Pant A2.
SEED sl'mPHIOLE:
Strophiole on the seed is rudimMtary in C. cv Pant A2
while a prominent strophiole is present on the seed of all
BtylPsia species. In all the five hybrids the seed on the F1
plants invariably had a prominent strophiole. In the F2
generation the segregation for seed strophiole found ,a good f i t
for the ra t io 13:3 (Table X I W except in C. a x
A. caianifolia where the segregation found a good f i t for 15:l
ratio.
While the seeds of &yl.~&i species are mt t led , in cv Pan t
A2 the seeds are not rrottled. Seeds produced on the f ive F1
hybrids were mottled. Fp segregation i n C. sajm x A. &Usas,
C. &an x A. C. &an x A. scarabaeoiaes and C. &an x
A. lineata found a good f i t for 9:7 rat io (Table XV) . Whereas
the segregation pattern in the FZqs of C. &an x A . m did
not f i t into any of the standard genetic ratio's.

Table X I V . Inheritance pat tern o f seed strophiole I n C. mJm x specles hybrids.
F2 segregation Proba- Cross o d F1 ----------------- X2 b l l i t y
Pre- Ab- Rat to sent sent
C. LalM x A. &&au Absent Present Present 154 28 13:3 1.35 0.5-0.25
C. c&.l x A. Absent Present Present 189 18 15:l 2.11 0.25-0.1 - Absent Present Present 140 23 13:3 2.30 0.25-0.1 A. SeLUa
I;. r;pbn x A. Absent Present Present 165 46 13:3 1.28 0.5-0.25
C. ralan x A. l h & a Absent Present Present 245 46 13:3 1.65 0.25-0.1

Table XV. Inheritance pat tern o f seed mottles i n E. x At~ lo& species hybrids.
Cross FZ segregation Proba-
P d F1 ----------------- $ b i l i t y Pre- Ab- R a t i o sent sent
E. c&n x A. Absent Present Present 101 81 9:7 0.04 0.9-0.75
C. rnipp x A. ~ I f o l l a Absent Present Present 127 80 9:7 2.19 0.25-0.1
G. l a a n x A. zmkm Absent Present Present 110 53 .9:7 8.36 < 0.005
C. S A j m x A. Absent Present Present 118 93 9:7 0.009 > 0.9
C. !abn x A. llneata Absent Present Present 170 121 9:7 0.55 0.5-0.25

Although the pads of C&nw have hairs, the hair being very
short the pods a ~ p e a r almost glabrous, while the pods of
8. scarabaeoides, A.lineata arid A. sericea have very prominent
and dense hairs. In the Flts involving all these three BtylPsin
species, pods had dense and long hairs similar to those in the
A t y b h parents and in all the three cases the F2 segregation
found a good f i t for the s-le 3:l ratio (Table XVI ) .
!bo (A. n?bicans and 8. -) of the five &&&,a
species under study are clinbers while the rest are erect types
as in C&nw, hence the inheritance of twining nature w a s
studied i n the hybrids of C. cajm x A. albicans and C. & x
A. --ides. Hybrids with A. all&ma were twiners in their
gross morphology. In the F2 generation although minor variations
were recorded for the erect or twining nature, basically the
segregants were either twiners or erect types and the segregation
found a good f i t for 13:3 ratio (Table XVII) . The hybrids of
Cajau~ with 8. sericea were intermediate with a bushy base and
twiny nature in the upper pr t ions and the F2 segregation in this
case did not f i t into any s-le ratio.


Table XVII. Inher i tance p a t t e r n o f tw ln lng nature I n C. x dtrlrr.la rpec i hybrids.
F2 generation CMSS O d F1 "-"'--"--'-"--"--"'- Proba-
Twin- Erec t I n t e r - R a t i o x Z b i l i t y f ng mediate
------------------------*----------------.------------------------------------------
C. rPlan x A. alhlcans E r e c t Twining Twining 150 32 - 13:3 0.16 0.75-0
C. SLbn x A. scarbbasoldel Erec t Twining Twining 35 74 102 - - -
( I n t e r - mediate)
Table X V I I I , Inher i tance p a t t e r n o f l e a f l e t shape i n C, faja x &h& speci hybrids.
F2 generation Cross P u F1 --"-""""""--'-"'- X1 Proba-
I n t e r - Rat io b i l i t y type mediate t y p e
C. rninn x A. rlblcans Lancec- Obovate I n t e r - 39 90 53 1:2:1 2;16 0.5-0.;
1 a te mediate
C. rPlnn x A. shu&kd% Lanceo- Obovate I n t e r - 48 104 59 1:2:1 1.18 0.75-0.
l a t e nediate

The leaflet Bhape in C. Eajan is lanceolate while that in
A. alblcma and A. scarabaeoides is &ovate. The leaflet shape
i n the hybrids involving A. albicans or A. scarabaeoidee was
intermediate to the parents. The F2 generation segregated for a
1:2:1 ratio for leaflet shape in both the goplations
(Table XVIII) .

SEED mPH1OLE:
Our results clearly indicate that the seed ~triopbiole in
the A t y l ~ & species is a dominant trait and is controlled by two
genes. Even in the hybrids with the two Australian species where
genetic studies were not conducted the hybrid seed had a
strophiole, unequivocally establishing the dominant nature of
this trait. Re* gk al (1980) reports three ratio's, 3:1, 9:7
and 13:3 for seed strophiole expression in crosses involving
~pecies of A. sericea and A. scarabaeaides while Pundir (1981)
indicates that two genes with a duplicate gene action (15:l)
govern this character. In the present study four out of the five
hybrids segregated for a 1333 ratio indicating a inhibitory gene
action while the hybrids of C. x A. caianifolia segregated
in to 15 strophiolated to 1 nonstrophiolated types. A
significant cbservation is that the seed geminability in the F2
poplation$ of C. & x A. caianifolia which has deviated from
the 13:3 ratio has been poor (Table 111) when compared to others.
This inplies that the genetic factors reqonsible for the absence
of strophiole are linked with certain lethal factors affecting
sed germinability.

8EPDlwmtINs:
Seed mtt l ing being invariably expressed in all the hybrids
is a dauinant t r a i t . Re* & a (1980) i n hybrids of
with A. scarabaeaiaes reported a 927 ratio when I B 6915 and
ICP 7035 were used as ferrale parents and a 15:l rat io when
I B 6997 was the female parent. In our studies 4 out of the 5
ppl la t ions found a good f i t for 9:7 ratio indicating a
co11@1ement&~ gene action. The segregation in the hybrids with
8'. sericea did not f i t into any of the standard genetic ratio 's
although it w a s nearest to the 9:7 ratio but it finds a good f i t
for 2:l ra t io suggesting a close linkage with a d d o m i n a n t
lethal of the carplimentry gene(s) responsible for the
experession of this t r a i t wherein the lunmzygous daninant
genotypes do not survive.
m HAIRINESS:
Pod hairiness of the Atyk& species is dorninant over the
relative plbescence of C. s&a. Reddy (1973) and Pundir (1981)
found that pod hairiness of E & y h h species is a mnogenic
dominant t r a i t . Cur results for this character agree with the
two earlier reports since in all the three hybrids involving
p i e s of &&& with hairy pods the pis in the Flls were
hairy and in the F2 generation they segregated in a simple 3:l
ratio.

The segregation pattern of twining nature in hybrids
involving A. rlbicans indicated tbat the character is gwerned by
two genes with a inhibitory gene action. In r. a x A. the F1 was intermediate to the parents in its
habit and in the F2 generation all the three types i.e. twining,
intermediate and erect types were recovered and the pattern of
segregation did not fit into any of the genetic ratio's.
Accurate classification of segregants in to distinct groups was
not possible for this character due to overlapping of characters
between the groups.
LERFLFT SHAPE:
Leaflet shape in both the hybrids studied was found to be
controlled by a single gene with a pactially &&ant gene
action. Reports on genetic studies for leaflet shape in S&au~
are variable. Deshpnde and Jeswani (1956) reported both 15:l
and 3:l ratio for lanceolate and obcordate types and the studies
of Jeswani and Deshpande (1962) and D'cruz and Deokar (1970)
indicate a 3:l ratio, while the studies of Patil and M a r
(1980) and Chaudhary and Thorrt,re (1977) indicated that m r e than
m e gene controls leaflet shape.
In intergeneric crosses involvhg C. and
A. srrarahneeides, Reddy (1973) and Pundit (1981) reported a 1:2:1
ratio which is in agreement with cur results.

Variation f o r length (Table XDU , 100-seed weight
(Table W) , seeds per pod (Tabla WI) , mibleaf lergth
(Table XXIII) and *leaf width (Table XXII) were studied i n t h e
F2 generation o f the hybrids of G -(Pant A21 w i t h 4,
a U k ~ & , A . s e r k t s a , A . -,A.lineata,andA.
The mean pod length of the F2 segregants in all the f i v e
popllations was clcse t o t h e mid parental value (Table X I X I . The
w a n f o r the p o p l a t i o n of C. a x A. rlbicans w a s 4.71 an
with a range of 2.8-6.3 an and a coefficient of var iat ion (c.v.)
of 19.5% as again& the parental values of 5.93 cm (C. -1
and 3.52 m (A. albicans). The difference between t h e parental
values was minimmar between C. c&n (5.93 cm) and A.
&mi&Ua (4.31 da) . The P2 mean in t h e p o p l a t i o n derived
from the cross between these species was 4.82 cm, t h e F2 range
being 3-86.1 an w i t h a C.V. of 13.99. The difference b e t w e e n
pwental valuesuasraximum i n C . rajnn (5.93 cm) x A. aericea
(1.35 an). The variation for p d length in fie F2 a p l l a t i o n of
this hybrid was 29.2% with the length ranging from 1.2 t o 5.4 an
w i t h an average of 3.59 m. Aln-ost ident ical resu l t s were
obtained in the ppula t ion of C. s&au x A. Jheata where the
average p d lengtbwas 3.35 an with a range of 1.3-5.1 an and a
C.V. of 29.9%. A variation of 22.47 i. .:: recorded in the F Z 1 s of
. s;aian x A. -e with t;,e pod length ra-:jing from

Table XIX. Variation fo r pod lmngth (a) i n the Fz population of hybrlds between E. mjm and spu i rs .
Mid- F2 population CFDSS 0 parental .........................
value Mean Range S.D. C.V. -----------------------------------------------*------------------------
Table XX. Variation f o r 100-seed weight (g) i n the F2 population of hybrids between C. a and &t&sh species.
Mid- FZ population Cross P parental -----------------------------
value Mean Range S.D. C.V.

Table XXI. Variatlon fo r seeds per pod i n the F2 population of hybrids between C. ajan and Atvlosrr species.
~ lb - F2 population Cross a .........................
value Mean Range S.D. C.V. ............................................................................
Table XXII. Varlatlon fo r mld-leaf wldth (cm) I n the F2 population o f hybrids between C. & and specles.
Mid- F2 population Cross. 0 parental ..........................
value Hean Range S.D. C.V.

Table XXI I I . Var lat ton f o r rnfd-leaf length (a) i n the F2 population of hybrtds between C. and spectes.
Ut d- F2 popul at ton Cross parental ..........................
value Mean Range S.D. C.V.

2.5-5.5 an with an average of 3.62 cap.
SEED WEIGHT:
A rmch greater variat ion ranging from 21.1-33.3% was
recorded for seed weight (Table XX) in the f ive p o p l a t i o m but
t he F2 mean w a s again mostly close t o t he mid parental value.
Seed weight in Pant A2 is 6.65 g. The 100 seed weight in the
~ O U K Atylosia species, A. irlbicans (2.52 g) , A. sericea
(2.28 g ) , A. scarabaeoides (2.10 g) and A. lineata (2.35 g) were
close t o each other while that in t he other species, A.
-was 4.46 4. I n the F2 generation the 100-seed weight
ranged from 3.00-7.18 g in the popllations of G. u x A.
t 4.71-6.77 in the pop l a t i on of
G. sajm x A. s & u U d h , 2.13-6.45 g in that of C . ' u x A.
sericea, 2-15-6.13 g in C. Eainn x a. scarabaeoiaes and
2.23-6.19 g in G. m j a ~ x A. lineata with 4.16 g, 5.4 g , 4.3 g,
3.7 g and 4.09 g being the respective means.
SEEDS PER POD:
None of t h e segregants in the f i v e F2 p o p l a t i o m exceeded
t h e be t te r parent value for seeds per pod (Table X X I ) .
Segregates w i t h fewer Fseeds per pod than t h e poor parent were
c o m n as is evident from the range (Table IW). The mean in the
F2 p p i l a t i o n of C. c&n x A. mjaUsUa (3.311, C. G&AR x
A. eericea (2.51) and C. G&AD x A. lineata (2.71) were close
to the mid parental value while tk~sse of C. rajan x B . a l r i w
(2.61) and C. x A. scarabaecides (3.05) were l e s s than the

mid parental value. The coefficient of variation for this
character was maxirnrm (32.6%) in the popllations of C. &an x A.
and minm (6.3%) in that of S. s&n x A. . . m, while in tbe other pgulations the variation was
21.4% in C. a x A.lineata, 22.3% in C. x
A. and 30.7% i n C. @an x A. aericea.
F2 variation for mid-leaf width was maxirnrm (31 .I%) i n the
population of C. &an x A. sericea, while the other four
poplationsr C. 4 . a ~ x A. c&dbU (5.181, C. Edjan x
A .albicans (11.4%), C. x A. scarabaeoids (17.1%) and
C. a x A. lineata (16.3%) shmed much less variation
(Table XXII) . The F2 means for mid-leaf width althdugh close t o
the mid-parental valuer fractionally exceeded that value in G
~ x A . albicanstC. b X A . scacabaeoiaesandc.
x A. sericea. The width ranged from 2.4-5.3 an in the poplation
of S. x A. w, 0.7-2.9 an inC. u X 8 .
sericear'1.4-3.3 an in C. x A. scarabaeoiaes and 1.4-3.2
an in the population of C. G&AII x A. lineata while the range
(2.4-2.8 ad was very narrow in the p p l a t i o n of C cajan x
A* caianifolia*

Studiea on F2 variation for the charactera pod lqth, seeds
per pod, 100-seed weight, mid-leaf width and mid-leaf length have
shown interesting trends. In general the Fq means with some
exceptions have been close to the mid-parental value but
transgressive segregants, i.e. those exceeding the better parent
values have been recorded for all the characters except seeds per
pod: Transgressive segregation has been most pronounced for the
leaf characters especially in the population of C. a x A.
albicans. Transgression of a marginal msgnitude has also been
recorded for pod length and 100-seed weight in the F2 '6 of s. raian X A- albicansdnd C. &an x B . c a i a n i f o l i a . A f a
-regants with leaves wider than the better parent have been
obtained from the poplation of C. x A. seric9a. As is
evident from Tables XXII and XXIII leaf length and width in some
of the F2 segregants of 6. &an x 8. is considerably
higher than that of the better parent.
Another striking feature in these studies is that the
variation m n g the five F2 populations is invariably mininun for
all the characters studied in the cross C. cajan x A. . . -, the variation being as low as 5.1% for leaf width,
7.1% for leaf lergth, 6.3% for seeds per pod, 13.9% for pod
length and 21.1% for 100-seed veight. FOE 100-seed weight and
the leaf characters the variation has been mxinunn in the
poplation of G. c&an x A. sericee while for seeds per pod it
is highe?' m the population of C. x A. abiwns. In the
h ;, ~i LA ciuan_x Pu -egreqmts wltn values

better than the better parent have been recorded for all trait6
except seeds per pod. F2 segregants poorer than the poorer
parent have been recorded in several cases.
Transgressive segregation for most of the characters
revealed the potential of the A&b&a species for pigeonpea
inprov-t. . BarBacki & al (1976) has enphasized the role of
transgressive segregation in crop improvement. Stebbins (1977)
and BarBacki & a;L (1976) have sham that transgressive
segregation front distant hybrids can give rise to extreme
characters and a range of new forms. The exploitation of the
transgressive segregantv would be a viable method for the use of
wild v i e s in the inproverrent of the cultivated species
especially when the cultivated and the wild f o m share a comron
chrcmsome n-r as in pigeonpa. Such studies i.e. those
aimed at the exploitation of transgressive segregants have been
~ ~ ~ e S 8 f u l l y attempted in other crop species (Reeves, 1950;
Raeves and BocWlolt, 1964; Efron and Everett, 1969; Harlan and
Dewet. 1976; Lawrence and Frey, 1976; Frey, 1976; Frey & al,
1983).
In the present study as indicated earlier trangressive
segrega~ts of considerable significance especially for leaf
characters have b- recorded in the C a j m u s - u F2
popllations. Such segregants for leaf characters assume greater
significance in the light of the findings of Shacm and Saxena
(1982) who have shown from intervarietal cross of pigeonpea that
lesf characters ere, k '.;hly heritable with a positive irfluence on
grain yield. The ~arginal hiproverent over the better parent in

the eeed weight in aome of the F2 populations is also a point of
significance. In a nutshell these findings indicate that the
species of -can be exploited for the inproverent of
certain quantitative t r a i t s of Chianus apart from their u t i l i t y
in incorporation of specific t r a i t s l ike disease and insect
resistance.
Further the wide range of variation recorded for the
chdracters studied with segregants better than the better parent
and poorer than the poor parent is indicative of the substantial
degree of reconhination between the gerromes of and
m. In these studies the variation recorded has been least
in the population of (laianus x A. . . which further
swstantiates the uor#mlogical and cytological evidence for the
relative genetic proximity of these two genomdspecies.
The limited investigations made on seed protein c o n t a t
reveal a dist inct trend. In all the F1 hybrids the seed protein
content is close to the values recorded in the better parent
!&y&&al w i t h the lone exception of 6. a x A. aranaifolia
where the protein content was intermediate to the parental
values. These results i.e. the proximity of the protein
percentage values in the hybrids to the values in the Aty&sia
parents indicate that the high protein content in the E Q h s h
parents is a dominant t r a i t . W h r (19%) re~or t ed part ial
domFnance for high protein content in crosses involving wild and
cultivated saybcans. It has not been possible t o analyse the
protein content in all the F2 segregants due t o limitations in
s& sar.p~r; wt analysis m sane segrqates silowed values

mrginally better than the better parat values. Haever,
without sufficient data it is not appropriate to conclude that
these are transgressive segregants since environment exerts a
strong effect on protein percentage.
Certain characteristic and interesting segregants were
&served in some of the F2 populations. In the poplation of
C. SWXI x A. pods in one of the segregants were with
and with out streaks on the sane plant. A few segregants in the
ppllations of C. sajm x A. and !2. x
&. scarabaeoides were found to have seeds of different colours.
A segregant each in the poplation of C. i&m x A. lineata and
C. C ~ J . ~ D x A. nlbicans was found to be non branching.
The occurrence of pods and seeds of different types on the
same plant is a possible case of chimera. Such chimera1
ir.dividuals especially for pod colour were also observed in the
progenies of Ca.ianus intervarietal crosses (K.B. Swena, 1984).
A non branching single stem mutant of the type observed in so=
of the F2 pplations was earlier reprted (Dahiya and Sidhu,
1979) as a spontaneous mtation in the F2 generation of a Sajaru
intervarietal hybrid. These aberrant types in the F2 popllations
l r ~ g h t also be a consequence of some freak genic constitutions as
a result of recarbination and segregation in the F, hybrids.


A t t m t s to standardize culture conditions for regeneration
of Q&aus and EQl.&a plants f r m different explants such as
a t y l e d o n s from mature seeds and leaf and epicotyl s-ts from
one week old seedlings resulted in different degrees of success.
~ o t ~ l e d o r . t i s s u e wss found t o be the best responding explant in
our i n i t i a l exploratory a t t enp ts and further detailed s tudies
were conducted on t h a t explant. The i n i t i a l a t t q t s a l s o
reveal& the superiority of H.5 M i u r n over the others hence, t h e
r e m n s e of cotyledons was studied on MS medium.
B&s.zl ( V S ) W i m su~le inen ted w i t h 2, 4 -0 ( 2 q/l) induced
copious mounts of heaithy ca l lus from all e x p l a t s i r respect ive
of the r q i o n of the cytoledon used. kaole cotyledons and nodal
halves of the qotylfdons on MS medim. su&plenented with 2 , 4-D
(0.5 q/l) and BA ( 2 q/1) developed rmlt iple W t s (3-7) in
addition t o mall m u n t s of callus (Fig.23C) in 2-46% of t h e
cul tures depending on t h e cu l t ivar (Table XXIV) . &mng the wild
re la t ives , m l t i p l e shoots developed from 21% of A. caianifolia
cul tures , while in t h e ccl tures ..of 8. and A. sericea
n.=inly s ing le shoots developed (Fig. 23B) and multiple shoots were
rare . In tkie cul tures of the d i s t a l s-ts of the cotyledons,
only shoot bud I:.itiation was observed i n l o w frequencies a f te r
profuse ca l lus i r~g . From t h e explants with nul t iple shaots,
r o o t k g w a s &t:-:: +.: cn b5 d i m supplerented with NAA ( 2 rq'l)
acd EA (0.5 &1:.

Fig. 23. In vitro regeneration of Caj'aras and A'y ios ia plants from cotyledons and immature embryos.
A) Sequence of plantlet regeneration from whole cotyledons of C d n u s ccjan.
B) Shoot regeneration from whole cotyledons of AtaZoslc sericec.
C ) Mu!tiple shoot formation in whole cotyledons of Cajcms @Gun.
D) Sequence of plantlet regeneration from immature embryos (15 day old) of Cc,icn.us ccj'cn.


T a b l e ~ x ~ \ ~ . ~ e r c e n t a g e s h o o t r e g e n e r a t i o n from c o t y l e d c n c u l t u r e s o f d i f f e r e n t c u l t i v a r s of C-s and s ? e c i e s of u c s ; e on HS rr.edium supplemented w i t h 2 ,4 -3 ( 3 . 5 mg/l) and & A t 2 n q / l J .
-- ---------------- Whole Cory1ec:n seqrnents
Spec les /cu i : i ' ; a r5 COty- ------------------ l e d on Nodal E l s t a l -----------------------------------------------------------------
G G ~ ) E D cv ICP 7335 4 6 3 1 1 3

Basal W i m n sqpl-ted w i t h 2, 4-D (0.5 u ~ / l ) , 54 (2
mg/l) and NAA (1 nq/l) also meed rml t ip le shts from whole
cotyledons and r d a l cotyledonary seguents. In this honmne
d i n a t i o n both shoots end rwts developed from 2 t o 14% of
wble-atyledon explants of C. &an and from a b u t 2% of
A. caianifolia (Table W . A l o w frequency of plant
rweneration was obtained from cul tures of IKXW cotyledctlary
s-ts i n C. &. In t h e cul tures of &. and
A. sericea only shoot regeneration was obtained.
Of t h e four C. cu l t ivars tested, three cu l t ivars
showd varying leve l s of regeneration, while the fourth cul t ivar
GS 4 , which had the smallest seed s ize , dLrrPst mmpletely fa i l ed
to respnd . The r e s p n s e of A. ca'lanifolia, which is
m r p b l o g i c a l l y q u i t e similar t o L;. e, c o n p r & well w i t h the
response of C. & cul t ivars .
A g e n e r a observation i n cotyledon cul tures has been tha t ,
wherever there was m l t i p l e shoot f o m t i o n , only one or two
shoots developed into f u l l shoots while t h e others r a i n e d
su~pressed.
Respnse of errbryos from four genotl-5 of pigeonpea
(Pant A2, ICP 7035, P r a t b t and C 11) w z s evduated on B5 nediw
t;ug.@mted with 2, 4-D (1 rrq/l). An age d-ent d r y 0
resporse was evident (Table XXVI) . Eleven t o fourteen day old
e:trvos i=-,.c:c:.<-c- r : :?;? f r c : wt.:.:? r- ICK f r x , . e=r_ , ?f p l m t l e t .

T a b l € \ u i . P e : c e i i a q c p l a n t l e t and s h o o t r e c e r . e r + t ~ o n f rom cct)!cdar , c u l t u r e s of different W ~ S c ~ l t l v a r s a r d s p e c l e s of &-8 on fi5 m e d l ~ a s ~ p p ? e - e n t e d w l t n i , 4 - C ( 0 . 5 m g / l ) , BA ( 2 mg/l) and NAJ (1 nq / l ) .
Whole C o t y l e d o n oegn ,en t s c o t y l e d o n -----------------------------
-------------- Kodal D i s t a l Cul : lva r /apec les s h o o t s p l a n t - -------------- --------------
l e t s S h o o t s P l a n t - S h o o t s P l a n t - l e t s l e t s ---------__________----------------------------------------------

regeneration wa.~ obtained. I n 15-19 day old ent~ryos p lan t le t s
were recovered d i rec t ly a c a q m i e d by d l a m t s of ca l lus a t
the base ( F i g 23D) . Direct plant let recovery and occasional
ca l lus forrration w a s observed in a b r y o s older than 19 days.
m r y o s .younger thar, 11 days fa i l ed t o r e s p n d on both the d i a
and b-1 media sup~lemented w i t h several h o m n e a r b i n a t i o n s
fa i l ed t o extract any r e p n s e from such d r y o s .
Though there was no s t r ik ing influence of genotype on
p lan t le t recovery, ICP 7035 performed marginally bet ter in all
t r eamts (Table XXVI) . ICP 7035 has the l a rges t seed s i z e of
t h e cu l t ivars used, in this study.
Cdllus w a s obtained from the mthers of C. c&n and
A. alhicuLs on M.S rredim supplimnted with 2 4 1 of 2, 4-D.
Potato s tarch extract sd iun : ~ r o m t & a l l u s developnent from 'he
anthers of A. ora.ICifolia md A. volubllls . . . Callusing was m r e
profuse from the anthers of A. aJ&ms and A. ~ Q W U than from
the oL&r slpcies . A t t q t s t o induce different ion by
s k m l t u r i n g t h e callus on k a l media supplimentd with various
horrrones and t h e i r coribiiiations did not meet with success. C k
s , a l t u r i n g the c i l l c s turn& trow. and dqu .e ra te2 .
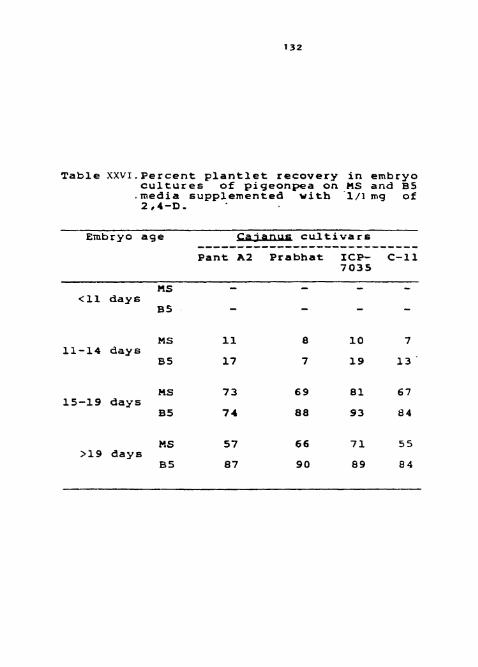
Table SXVI. Percent plantlet r e c o v e r y i n embryo c u l t u r e s of p i g e o n p e a o n - H S and BS .media supplemented with 1/1mg o f 2,4-D.
Embryo age Ca,enus c u l t i v a r s ............................ P a n t A2 Prabhat ICP- C-11
7035
ns - <11 days
B 5 -
HS 11 11-14 days
B 5 17
HS 7 3 6 9 8 1 6 7 15-19 days
B5 7 4 8 8 93 8 4
US 5 7 6 6 7 1 5 5 >19 days
B5 8 7 9 0 8 9 8 4

I n cotyledon cul tures bksal medium supplerented with 2, 4-D
induced only ca l lus while the rredium su~plemented w i t h both 2,
4-D and FA induced shoots. Robting of such explants was'achieved
on the medium when it was su~planented with W.&. When the basal
medim was scgpl-ted with bath BA and NAA in addition t o
2, 4-D a low frequency of whole plants were recovered. These
findings suggest thet while BA plays a role in s b t regeneration
while tGA helps i n rcot regeneration. CUtivar differences with
respect t o t h e percentage of regeneration (Table X X M is a
possible consequence of the genotypic e f fec t and such resu l t s
have been r e p r t e d in other species. This conclusion is
strength& by t h e f a c t t h a t the response of 8. caianifolia which
is genet ical ly close t o C. w a s similar t o the' response
obtained i n the a l t u r e s of C. -while t h e prforrrmce of
A. and A. m, w i e s Mich a r e rrore dis tant ly
related t o Caianc-+ was distinct.
Variation in respnse kas recorded w i t h respect t o the
cotyledonary s e g e n t cultured. The r e s p p s e of &dl s q e n t s
w a s c lose to t h e resLwrse ob'iained from t h e m l e cotyl&ns
while t h e r e s p r s o £LC,;: t:w d i s t z l segrmts was poor. The
irproved r.=sF<rrse cf the r.:dal s e p s n t s night be due t o the
presence of m r y c : i c cells in the nod& half.
Culture ccr.dl:ions for the regeneration of imature
pigeonpa erbrycr : rve tw scmdardized for the f i e s t tire. PLI
.:;: 5'; c-r'cnt cr.: - : c : ; . : ~ . : . \.;r cta i r .62 in t h e c:,tryc c l t - r c . -

~t vas mt possible t o regenerate plants from erbryos younger
than 11 day old. This has been a m n fea ture in other
leguminous species ( G o d and Bajaj, 1983: Newel1 and Fiymi tz ,
19821 Qlbero, 1981). Genotypic effect on percentage of
regeneration in d r y 0 cul tures was less pronouncd than Fn
cotyledon cultures.
Anther Eulture s tudies were not successful l beyond the s tage
of callus induction insp i te of a t t enp ts t o induce regeneration by
s u ~ l i m e n t i n g t h e basdl media with several b r a o n e carbinations.
Anther cul tures have been successful t o a la rge extent in
cereals , solanaceous crops e tc . but have not made an impact on
any of t h e leguminous crops. An e a r l i e r attempt on pigeonpea
anther a l t u r e s also did not progress beyond the c a l l u s induction
stage (Bajaj & a, 1980) .

8- GENERAT, DISCUSSION AND 00NCLL1SI<3N

present hestigation m Mtiated in to
understand the q=c-me relationahi~e bet= C. arrd
AQhsia species, screen -- crosses for selective
chrmaome elimination, develop methods to overcane barriers to
species crossability, study the inheritance and variation for
important traits and develop h YitEn rgeneration techniques for
pigeonpea. All these objectives have been met.
Out of the twelve species of Atyksia and two species of
used in the study eight species of hybridized
with the cultivated species. Thepreeent study has aho*m that the
species crossability is significantly Mluenced by the genotype
of the Cajmu pacent. Studies on h o m e treatments have
revealed that post-pollination hormone applications -rove
species crossability by checking the post-fertilization barrires.
Reciprocal success in these crosses is very difficult. Four of
the twelve Atyhs ia species failed to crose with Caiauu in any
direction. In the unsuccessful crosses post-pollination hormone
treatments delayed bud/@ drop but it was not sufficient to get
ovuledenbryos responding to in yitLp rgeneration. In YttEP
rescue of dry06 could be achieved only in embryos older than 11
days. Both hormone a~plications and d r y o culture techniques in
pigeonpa have been developed for the first time. The results
indicate that h o m n e amlications in conjuncture with &ryo
culture technique offer potential for obtaining new
~~ hybrids with further refinements of these
techniques.

The present study harr resolved the longstanding controvep
regarding n-r of nuc$lar organizer chromeomes in pig-.
In spi te of the limitations of short meiotic cycle and
di f f icul t ies in cbtaining good cytological preparations in
pigeonpe a detailed information on the nuker, size and
distribution of nucleoli a t T-I and T-I1 was obtained. Evidence
in the form of nucleolar n m r a t T-I and 'lr-I1 in diploid and a
spontaneous tetraploid of pigeonpea establishshed beyond doubt the
presence of two nucleolar organizer chromosomes in pigeonpea. A
similar situation exists in the species of -. This is the
f i r s t study t o use meiotic activity of the nucleolaf chrom- e
as an index of the nucleolar organizer nuker. The preqnt study
has also shown the attachment of two bivalent6 t o the
diplotenddiakinesis nucleoli and also the p r v c e of two
p r i m nucleoli a t pachytend diplotenddiakinesis t o
substantiate the presence of two nucleola I organizing
c h r - m . The chromsome n-r i n the Australian $ies of
&ylg&a is being reported for the f i r s t time.
The major accorfplishmt of t h i s investigation is the
detailed insight provided into the g e ~ m e &ationship between C.
sajan and the species,, A t y h d a . These investigations suggest
that C. &an and the spxies in the genus A t y l d a basically
have a similar genome or i n other w r d s they have evolved from a
c a m n gene pool and the differentiation has occur& as a result
of structural alterations. The closest relat ive of pigeonpea . .
among the specie6 studied is A. ca,nnlfolla. The major
oist inguishirq feat1 9 I r t w e w these t r q-~ccier is the prccc:: e

of seed strophiole. sane of the recent collections of
&an by the Genetic Resources Unit a t ICRISAT ,ahow a promihent
strophile (van der Maesen, pers. am.). In view of tbese
findings it i s likely that C. mj.an might differ fran A.
. . by only a few gene uutations. This was further
substantiated from the data on variation for quantitative t ra i t s
from the F2 populations. For a l l the characters studied the
variation .has been the lowest in the populations of C. x
A. caianifolia. The other species of that have
hybridized with C. &an are mrphologically mre distinct.
Meiotic studies of their hybrids reveal that they F e genomically
close to pigeonpee and have differentiated through structural
alteratiohs and the results have shown that the levels of gametic
s ter i l i ty reflect the degree of species divergence. Australian
species of A ~ Y ~ Q & are more diverged from than the Asian
species.
The predominantly one way success of the --&yl~&&
crosses suggest that the structural alterations and/or gene
nutations that have led to the species differentiation are
acconpanied by the differentiation of p l m n . There is no
evidence for chromosome elimination in the intergeneric hybrids
of pigeon*. The infomktion gathered in the present study is
rot sufficient to comnent upon the relatianship of C. Eajan
with the species of Fhyshsh except that they share the same
chromomme rumber.

The study on inheritance of qualitative t r a i t s has shown
that these t r a i t s are controlled by one or t m genes. Sweral
w e 1 segregants were recorded in the F2 generation and
transgressive segregants of considerable value have been
reomered in the F2 generation for characters l ike leaf length,
leaf width, seed protein content e tc underlining the importance
of species for the hprovement of the cultivated
' species.
This study for the f i r s t t h e has provided direct
cytological evidence for reambination between g a m e s by way of
studying the pairing of nucleolar organizer chromsomes and
tracing the products of recabination using the nkleolus as a
mrker. This technique can be a potential tool Fn reambination
studies in distant hybrids. These findings have also provided
the f i r s t cytological evidence for the structural hybridity
between Cajaus and B f s ! b s i a species.
'Ihe present investigation has provided anple cytological and
geneticdl evidence for reconbination between the genome6 of
Ca.ianus and A t y l ~ & species and has proved that the species of
A t y k s i a possess all the requisites (close genome relationship
with the cultivated species, a high degree of recarbination
between the genosrres and recovery of transgressive segregants from
the progeny of hybrids with the cultivated species) for effective
introgression.

In a overview the present imrestigaticn has shovn that the
species of Uyhda can be of potential in pigeonpea hprwment
programnes and these species deserve m r e attention than they
have received so far.
In view of these findings future attempts rmst be directed
a t obtaining hybrids between and species of
which have not been used so far in intergeneric studies. Further
refinements in unconventional techniques l ike d r y 0 culture and
hormone treatments and developrent of new techniques l ike
r-ition and mentor pollen techniques should be given
importance t o achieve success in otherwise incompatible crosses
in an attenpt to widen the genetic base. There is also a need t o
obtain hybrids between I&yh& species to be able t o gain more
knowledge about the divergence of these species from -. There is a need t o screen the progeny from the i n t e r g e r i c
hybrids and select t h e desirable segregants, evaluate them and
incorporate thm into pigeonpea inprovenrent p r o g r m s t o
achieve introgression of the deisred character by adapting the
relevant breeding mthcdologies.
The present study has shawn tha t tetraploid pigeonpea is
cytologically stable and hence long term attenpts must also be
directed a t crosses between tetraploid C&m& and species of
tO Obtain addition andtor substitution lines of
pig=%=.

REFERENCES

Q.N., E.J. BRITPEN, and D.E. BYTB. 1977. Inversion bridges and roeiotic behaviwr in species hybrids of soybeans J. Here. 68: 360-364. d m, Q.N., and E . J . BRmTEN. 1979. Inversion heterozygosity , in the hybrid soybean x BPjB. J. Hered. 70: 358-364.
*, C . S . , and R.W. HATRMANN. 1977. Interspecific hybridization amng four gpecies of the genus w. In: Proc. First Intl. Mungbem Gympasim, Asian Vegetable Research and Dweloprient Center, Shanhua, Taiwan. p. 240-246.
'AHN, C.S., and R.W. HAKlWWW, 1978a. Interspecific hybridization between mgt?ean (yisnn ' (L.) WUCZEK) and adzuki bean YQna ~$&ULU ( W I D . ) =and -1). J. hr.Soc. Hort. Sci. 103: 3-6.
'AHN, C.S., and R.W. H?XfMMW, 1978b. Interspecific hybridization between rice bean (- ~THUNB) CHWl and OHASlI) and adzuki bean Y. XQUULLS (WILLD). OBWI and OHASU) . J. Amer., Soc. Hort. Sci. 103: 435-438.
'AKI&, J . O . , A . J . PIuTaAFD, and P.C. WHITPIAN. 1972. chromsome rider in pigeonpea ~~ (L.) Millsp.) . J. Aust. Inst. Agric. Sci. 38: 305-306.
'ALA, A. YA., and T.F. 05TAPENKO. 1979. Variability of quantitative characters in fifth generation interspecific hybrids of soybean. Vyni Zhurnal 7.65. 49.
R.W. 1960. Principles of Plant Breeding. Jhon Wiley. and,Sons, Inc. pp. 434.
/A, L.C., and c . KIMBER. ,1980. A haploid between and Trltlcum aavrnn. Cereal ms. -%???
355.-358.
8, M.K. 1981. Effect of interspecific S&S+UR hybrids on biological nitrogen fixation. Dissertation Abstracts International, B, 41: 39458.
'--KRISl'EVA, M., H. NI@X@F, G. KUNZEL and R. RIFGER. 1977. Nucleolus fomtion in structurally reconstructed barley karyotypes with six satellite chromsmres. Chronns01~ (Berl.) 623111-119.
' w - K R I S P F V A , M.. H. NICOLaFP.r aria S. -1EV. 1978. Revision of SAllcchromsone nunher in !lbtbm with respect to nucleolar organiers activity. Theor. -1. Genet. 55:229-231.
M a E K X J S . 1.7 6 . s i m p ,..- .~r;sr U-I ure irequerlcy of kollen Plant induction in wheet with Potato medium. Acta Genetika Slnice, 3 : 30-31. --

'?&-YRSIRI, S. and D.P. COYNE. 1964. Effect of growfh regulators in delaying pod abscission and &ryo ebortlon ur the inter-ific cross B ~ ~ B ~ U S x E acutlfolius. Crop Sci. 4: 433-435.
~CYASIRI, S.A., and D.P. OYYNE. 1966. Intergpecific hybridization i n the genus m. Crop. Sci. 6: 59-60.
&, R.P. and J.A. SPENCE. 1978. A possible gene source for early, day-length neutral pig-, (L.) Millsp. Eqhytica 27: 505-509.
AVRDC. 1977. (Asian Veg. R e s . Develop. Center. Progress ~eport . Taiwan.
'&AS, Y.P .S . , and M . BOPP. 1971. Gewebekulturen in der angewandten Botanik (Plant tissue cultures in applied botany). Angew. Botanik 45: 115-151.
WAJ, Y.P.S . , H. S I m , and S . S . GOsAL. 1980. HaploiC dryogenesis in anther cultures of pigeonpea (- . Theor., Appl. Genet. 58: 157-159.
W A ~ ~ Y . P . S . , M.M. VERB%, and M.S. IXIANXI. 1980. Barley x rye hybrids (Hordecdle) through eirbryo culture U. i33. 49: 362-263.
2 E&WEFOT, H., andG. FODILUXM. 1981. Breeding. french beans and scarlet sumnr beans. Present position and out look.
FW!UI, R.K. 1965. Complement fractionation +I a natural hybrid between Bvtzus &x~csus Muell. and B. lacvlatus wild. Nature 208, 608.
~ ~ I r S.G., G. rWNWEXA, T. ADAMSKI and M. SURPIIA. 1976. Transgressions in barley (H+hm s&&m JESS). V. Trangression and heterosis their umrtance for plant evolution and breeding Genet. Pol. 17 : 77-82.
B k A Y , I.R. 1975. High frequency haploid production in wheat (- aestlvrrm) by chromsome elimination. Nature, 255:410-411.
BELCWA,G.I., and K.N. NASSAL. 1979. A study of abryo fomtion following interspecific hybridization in pea Wsum L.) Referativnyi Zhurnal, 5: 65. 37.
B m , G . I . , KW. NASSAL, O.H. BODEUIN and K.P. GIKfZSt. 1980. Cytoembryologicdl study of Fez following interspecif ic crosses. Nauch.-tekhn. vses. se1ekts.-qenet . in-tar 36 :40-47.
BENNCrr, M.D., M.K. PAD, J.B. SkIITP M.W. EXiLISS. 1973. Cell develomt in the anther, the :;.~le and the young seed of wm. L.. VGL . Ci~irisr L L T ~ . ~ U I . Trarrc. R.

&c. Ser. B. 266: 39-81.
V E E , S.K. 1956. Study of autotetraploid Caianus (Linn.) Millsp. Caryologia 9: 149-159.
B I N Z W M , E.T. and C.B. GILLIES. 1971. Chromsome pair*, fertility and crossing behaviour of haploids of tetraploid alfalfa, Medlcwo sativa L. Can. J. Genet. Cytol. 13:195-202.
BIR, S.S., and SIDW. 1966. m. IOPB chromsome number repoes VI. w e l l Love (Ed.) Taxon. 15: 119.
BIR, 6.S. r and SANlXUSH KUMARI. 1973 IOW C ~ ~ O E O S ~ nyber reports XLI. &kdl Love (~d.1 Taxon. 22: 459. J
BIR, S.S., and SAtmDsH KUMARI. 1977. Evolutionary status of Legrnninosae from Pactmarhi, Cmtra.1 India. Nucleus, Calcutta 20: 94-98.
BISYSLS, M.R., and S. INNA. 1975. Black gram x ric bean cross. Cytologia 40: 787-795.
BIMAS,M.R.,andS. INNA. 1976. Phhseolus amnltlfllus . . . X
Phhseolus trilobus. Indian J. Genet. 36:125.
B L k , L.M., W.D. F 1 9W, and L.S. . 1972. Notice of release of three noncomnercial stocks of cotton, Arizona 22, 383 and 495, resistant to Soutlmestern cotton rust. hiz. Agric. Exp. Stn. and USDA Mano.
WXWtGU, R.V., and A. HXBEfG. 1983. Prereeding and wide hybridization in barley. Eucarpia workshop. Genitika. Ser. F, Suppl. 111, pp. 41-53. f
BOTHMER, R.V., N. JACOBSEN, R.B. J-EN AM) I. LINDE-IAURSEN. 1984. Haploid barley f ran the intergener ic cross x ILagULs. Euphytica 33 (In press) .
BRARK, J.P., andR.S. MATLLXX. 1961. Mungbean hybridization te-que. Agron. J. 53: 54.
d, J.P. , and E. KCX)ISP.A. 1975. A successful cross between L. andphaseolua ' Jones with the
=dry=re. Euphytica, 24 :66-
BIIOUE, P . , MMSWEL, .D.R., and J .P. GRACE. 1979. Hybridization among the Austral~an wild relatives of the soybean. J. Aust. Inst. Agric. Sci. 45.
BLlWHAM, C.R. 1950. Chx,.,i usome segregation in translocations involving chramscxne 6 - ir~ize. Genetics, 35: 446-481.
UU~UD~~EBY, A.N. and M.V. TH@&X. ' 1977. Genetic studies in

pigeonpea. Round leaf x N.P. 51. Mahatme Phule Agri. Univ. Res. J. 2: 17-20.
B.K.r S.K. -1 S.L. BASAK., and S. DAN&- 1976. Cytogenetics of a cross between two species of annual -. Cytologia, 41:lll-121.
C E D I , N.C., J.F. PARFWT, T. JA-, L.R. BAKER, and P.S. CARLSON 1977. Interspecific hybridization of food legumes by unconventional methods of plant breeding. In. Proc. First Int. Mungbean Symp. AVRDC, Tainan, Taiwan, R.O.C. gp. 247-252.
/ J
CBFN, N.C., L.R. BAKER, and S. EON@L 1983. Interspecific crossability m n g four species of Y&na food legumes. Rrphytica 32:925-993.
&~I?+CHMG CHU. 1982. Haploids in Plant Improvement. Plant inprovement and somatic cell genetics (ed.1.K. Vasil, W.R. Sowcraft and K.J. Frey) . Academic Press, New York. pp. 129-158.
-Y, R.J . , J.B. CH(3EHURYI and V.P. S?NZH: 1977. An amphidiploid between Yigm radiata, m. and nwag~ . , Crop Improv. 4: 113-114.
W Y , R.K., and J .B . -. 1977. . Intergeneric hybridization between Y i g ~ mung~ (L.) HEPPFR and Phaseolus
RXB. Indian J. Agric. Sci. 47: 117-121.
CHU, Y. 1967. Pacytene analysis and observations of chromsome associations in haploid rice. Cytologia (%kyo), 32: 87-95.
- I L.S., and W.M. ST-. 1976. An introduction to the botany of tropical crops, Longnnn, London. * B 4 D.P. 1964. Species hybridization in &asedu& J. Heredity 55: 5-6.
m, I.J. 1982. Interspecific hybridization in vicia. In Faba Bean Improvement (ed. G. Hatwin and C. Webb). ~p:91-108.
aAHIYA, B.S . , and J.S. BRAR. 1977. Diallel analysis of genetic variation in pigeonpa (Ujaus . Experimental Agriculture 13: 193-200.
-A, B.S., 3.5. ERAR, and B.S. J3KUUGt. 1977. Inheritance of protein content and its correlation with grain yield in pigeonpea (Qjaus c j a n (L.) Millsp.) . Qualitas Plantarum, Plant Foods for PC-;in Nutrition 27: 327-334.
X R A , B . S . , and T.S. ClbtFJ. 1979. Non-branching (smglc s r u z d n u a t 21 klgeonpea ( C ~ J ~ W S (L.) Millsp.) Trop. Grain legme B u l l . 15: 24-25.

'm, S. 1966a. Cross between Pfusealus aureufi RO*. and 2. uu&a L. Genetics 37: 259-274.
D?iNA, S. 1966b. Cross between Philseolus Roxb. and E. -Ten. Genet. Iber. 18: 141-156.
'm, S. 1966c. Species cross between B ~ ~ ~ Q L E aureus Roxb. P. triloblls Ait. Cytologia 31: 176-187.
DARLINCI13Nr C.D., and A.A. m E p . 1930. P r h r y and secondary chromosme balance in J. Genet. 22: 129-151.
6ARW3Y! N.L., and C.J: DRISCOLL. 1972. &cleolar behaviour i n Tritlnrm. Chromsorra, 36: 131-139.
DRRVFY, N.L., C.J. DRISMLL, and P.J. KALTSIKES. 1973. Evidence against somatic and pr-iotic association in wheat. Genetics (Suppl.), 74: 57-58.
'D'CWZ, R., and A.B. D m . 1970. Genetic s tudies in pigeonpea. R e s . J. Mahatnm Phule Agric. Univ., 1: 44-53.
'DEIWICHE, C.C. 1978. ~egumes-pkst, present and future . Bioscience 28: 565-570. J
'DDXX?V&, G.B., and C.V. 'IHAWLR. 1956. Cyto-taxonomic evidence f o r t h e a f f i n i t y betw? iadisua, S p r y g . and ce r ta in erect species of W. & A. Proc. Indxan Acad. Sci. 43 (8) : 37-45.
DESPANDE, R.B., and L.M. JPSWANI. 1952. A pros t ra te mutant in pigeonpea Ca$anus G&UI (L.1 Millsp.) . A possible s o i l conservation plant. Indian J. Genet. 12: 50-51.
J 'DEPANDE, R.B.,and L.M. JESWINI. 1956. A case of p l e i o t r o ~ in
pigeonpea. Curr. Sci. 25: 201-202.
UWNElLY, E.D. 1980. Selecting l i n e s of vetch that breed t r u e f o r hard seed m a t . Crop science. 20: 259-260. .J
'DUIXAS, I.S., E.J. BRf'lTEN, and D.E. BYlB. 1983. Pachytene chromsorne iden t i f i ca t ion by a key based on chromrneres in t h e pigeonpa. J. Hered., 74: 461-464.
n , R . , M . S . FSdGUNA, and D. DE NEITANCDURT. 1969. Detection and cytological analysis of tormto haploids. Genetica 40: 181-190.
EFRX, Y., and H.L. EVERFm'. 1969. Evaluation of exot ic gemplasm f o r inproving corn hybrids mr thern U.S. Crop Sci. 9: 44-47. .
-, S.L. , and N . W . S r A I 7 . 1946. U s e of growth regulating substances L" L L ~ L W ~ ~ ii,. .. t ~ t ; L ~ i i ~ r LA Uiiuu. P L W . MI. Soc. Hort. Sci . 51: 581-519.

L.R., and W.D. -. 1981. Early generation selection for protein in Glyd.ne mx x Glydae &m crosses. Can. J. Plant Sci. 61: 490.
FALK, D.E. and K.J. KASBA. 1980. Carparison of the crossability of rye. (secak Md &U%lRll butbaslrm onto wheat (Prlticrrm astunad.
F m , G. 1977. Haploids from Barley x Rye crosses. Can. J. Genet. Cytol. 19: 15-19. .
F m , G. 1982. Effect of cultivar &ination on meiosis in barley-wheat hybrids. Can. 'J. Genet. .Cytol. 24:575-582.
F L A W , R.B., and M. O'DELG. 1979. The genetic control of nucleolus formation in wheat. Qiromsoma, 71: 135-152.
n;wD, L. 1970. Chrorrosome association in Zea mays mnoploids. The nucleus 13: 99-105.
MUZDAR, A., and S.L. TANXIN. 1976. Cytogenetical evolution in the genus W. Cytologia, 41: 91-104.
E'RhY, K.J. 1976. Plant breeding in the seventies: Useful genes from wild plant species, F q p t J. Genet. Cytol. 5:460-482.
E3EYr K.J., T.S. COP(, D.M. Ea3lXXES and P.B.. COP(. 1983. Increasing cereal yields with genes from wild and weedy species. Genetics :New Frontiers. (Proc. XV Inter. Cong . Genet. New Delhi) IV: 51-68.
GAI, J.Y., W.R. FEfIR, and R.G. PRIMER. 1982. Genetic performance of some agronomic characters in four generations of a back crossing progr- involving ~ r a ~ and G. a. Acta Genetca Sinica, 1982. 9: 44-56.
m, O.L., R.A. MILLER and K. CUIMA. 1968. Nutrient requirements of suspension cultures of soybeans root cells. [email protected]. Cell Res. 50:151-158.
GEXGENBACH, B.G., C.E. GREEN and C.M. COtKKRN. 1977. Inheritance of selected pathotoxin resistance in mize plants regenerated from cell d tu res . Proc. Nat. Acad. Sci., L1.S.A. 74: 5113-5117.
GILL, A.S., M.M. VERWi, B.S. DHALIWAL, and T.S. SANDW. 1983. Interspecific transfer of resistance to m@em yellow msaic virus from yirrna IUXJQ to Yigm radiata. Current Science, 52: 31-33.
GOSAC, S.S., and Y.P.S. BIUAJ. 1983. Interspecific hybridization between Yigna KWESQ and Ugna zadiata through exkjryo culture. ELtpbytica, 32:129-137.
-Y, W.C., M.P. GWXXY, A. mvIW, B.W. SMITH, and

J.A. uuwcua. 1973. In 'Peanuts~ture and Uses', Am. Peanut Res. and Educ. Ass. Gtone Printing Co., Virginia. ~ p . 47-133.
;RITPDN, E.T., and B.' WIERZBI(XA. 1975. An mbryological study of piam sativrrm x feha cross. !Zuphytica 24: 277-284.
b, S.B. and P. v. 1973. Selective -tic elimination of U h c & i a m alutvlosa chrcimscanes in the F1 hybrids of N. mapdens and H. -. Genetics, 73:605.
Ed?, M.N. . G.R. and J. SMNtlT. 1980. The cytogenetics of Ba6sAus L. , mccineun L. their interspecific hybrids. derived -diploids and back cross progeny in relation to their potential exploitation in breeding. Cytologia, 45:791-789.
IWRUh, J . R . , and J.M.J. D m . 1977. Pathways of genetic trnasf er from to Zea maya. Proc. Natl. Acad. Sci. U.S.A. 74: 3494-3497.
HAWKIlS, C.M. , and A.M. EMUS. 1973. Elucidating the behaviour of pollen tubes in intra and inter specific pllinations of Phaseolus-L. mdP. LAM. Euphytica, 22: 378-385.
HEHN, E.R. 1948.' The inheritance of agronomic characters ir barley. Ph.D. dissertation. Iowa State Un'iversity, Ames, Iwoa.
~ E I T Z , E. 1931. Die ursache der gesat~~6sigen Zaht, Lage-, Form and Grosse pflanzlicher Nukleolen. Planta, 12~775-884.
S. 1955. A technique for artificial culturing of bear 6rb~~os. Proc. Am. Soc. Hort. Sci. 65: 405-408.
d, S. 1956. A bean interspecific hybrid. J. Heredity. 47: 217-220.
'BOOD. M.J., and F.L. ALLEW. 1980. Interspecific hybridizatior studies between cultivated Glycine max and a perennidl wild relative. G. falcata. Agron. Abstr. P. 58.
'HLNE%t, J . E . , M . B . DI(RSON, M.A. and J.A. CI-. 1982. Evaluation of plant introductions of E b s e d u ~ s&p. fol resistance to white mld. Plant Disease, 66: 320-322.
6, J.K. 1979. Interspecific hybridization between Pbse~U L. and A. Gray. Dissertatia
Abstqcts Internatxmal, 39(10) 46728.
IBRAIfm, A.M. . and D.P. ODYNE. 1975. Genetics of stiqna Bhape coty? don position -7d C-nder mler in reciprocal c rossel betwew. aE&eQhs ul&Lx L LL E b s e a h B mscLuus ((Lrrn,
and inplications in breeding. J . Amer. Soc. Hort. Sci

ICRIW. 1982. Ann. R&rt. International Crop Reeearch Institute for the Semi-Arid Tropics. ~ p . 215-220.
ISAM, X.8. 1964. A rare hybrid conbination through mication of h o m e and embryo culture. Nature ( L o W 201: 320.
ISAM, A.K.M.R., and K.W. SHE PBERD. 1980. Meiotic restitution in Wheat-Barley hybyids. Chromscnm (Berl.) , 79: 363-372.
JW, B~s., and J.P. -. 1980. The site of action of the crossability genes (Krl, kr2) betireen and Secale. I. pollen germination, pollen tube growth, and nuu&r of pollen tubes. -Euphytica, 29:571-579.
~ESSOP, C.M., and N.C. SUBRAHMANYAM. 1984. Nucleolar number variation in &cdem species and interspecific hybrids. Genetics, 64(2) :93-100.
JESYSLNI, L.M., and R.B. DESHPAM)E. 1962, Inheritance studies on same sterile mutants in pigeonpea. Indian J. Genet. 22 : 236-240.
JESYIANI, L.M., and R. D'CEUJZ 1965. Linkage studies in pigeonpea. mdian J. Genet. 25: 234-238.
'JOGHI, S.N. 1973. Variability and correlatien studies in p i g e o v (Cajanus cajan (L.) Millsp.1 . Fonmtion of corrposltes. Euphytica 22:373-377.
KAKTR, R. 1977. A new approach to the breeding of pigeonpea (fajama &ad (L.) Millsp.) M.Sc. thesis (unpblished) Puniab Agricultural University, Ludhiana, India.
' IGdh iA , K. 1936. Crossing experiments with Glycjne &a and E. -. Jap. 3. Botany 8: 113-118.
,KAE33A, K.K., K. PAHL, N.L. LWIG, and L.A. ~ ~ 1 . 1981. Plant regeneration f r m meristane of grain legmes: Soyban, c n w p a , peanut, chickpea and bean. Can. J. Bot., 59: 1671-1679.
&, K.J., and K.N. &. 1970. High frequency haploid production in barley. Nature (Land.)., 225: 874-856.
'KFS?A,K.J.,N.C. -AM, andA. &I. 1978. Effect of grbberellic acid treatment and nutrient sqply through detacw tillers, u p n haploid frequ~cy in barley. Theor. -1. Genet. 51: 169-175.
'RASiA, K.J., and E. REI-. 1980. Achievments wid haploids in barley research and breeding. In the Plant Genm (EB. WVIES and HORKDD) . The John Innes Charity, 215-230.

K d R , N., and W.P. m S . 1960. Bybridization betwen two species of separated by physiological and wqhlogical blocks. Proc. Amer. Soc. Bort. Sci. 76: 397-402.
KEEP, E. 1962. Satellite and nycleolar n&rs in hybrids between 8Uu n&um,& g m s a h u a and in their back- crosses.
, Can. J. Geriet. -1. 4: 206-218.
REMPAWA, C., and R. RILEY. 1964. Secondacy association b e t w e m genetically controlled equivalent bivalents. Heridity, 19: 289-299.
KIAZUMA, N. and F. W. 1980. Inheritance of some qualitative characters in F2 generation of a species cross between soybeans ( G l y c h e msx (1.) Merrill) and wild aoybeans (G. sieb. and Zucc.) J. Fac. Agric., Iwate Univ. 15: 1-9.
KIAZUMA, N., F. W, J. FUKUI, and K. SAKURAI. 1980. Inheritance of qualitative characters in the F2 of an interspecific cross between Glydne m and G. apja. J. Fac. Agric. Iwate Univ. 15: 1-9.
'kIAZUPbl, N., Y. KIUCHI, and F. C#3. 1980. Breeding high proetin soybeans frem an interspecific cross between nwx (L.) Merrill and G. &,a Sieb.and kucc. - The need for repeated back crossing. J. Fac. Agric. Iwate univ. 15: 11-28.
~APOVICP;AS. A.. and V.A. RIG(3NI. 1952. Estudios citoloaioos en el g&ero Arachis. R w . Invest. Agric. (B. irk 5: 289-293.
KRISHm%mMI, N., and G.N. AYY?!KAR. 1955. Qlrmsome nurrbers inC&mu- spreng. Curr. Sci. 3: 614-615.
~ R B E , A. 1967. Intergeneric hybrids betweM Hoxdehm mkaue I.;. spp. distichum (V. 2n = 14) and Secale ceredle L. (V. pqt*us, 2n = 14). $?;etl-hojsk Arsskr. 82-92.
KRUSE, A. 1974. A 2,4-D treatment prior to pollination eliminates the haplontic (gametic) sterility in wide intergeneric crosses with 2-r~wed~barley, mlgazs ssp. distichum as material species Herld.itas 78:319. ,
L.S.S., A. ABWWM, and V.K. SRINIVASAN. 1945. Preliminary mte on autotetraploidy in C a j a u ~ hUal.6 spreng. Proc. Indian Acad. Sci. 21(B): 301-306.
KWAFt, L.S.S., R. DnCRUZ, and J.G. CRE. 1957. A synthetic allshexaploid in Arachls. Curr. Sci. 26: 121-122.
?NMARI L.S.S., K.V. THOMBRE, and R. DnCRUZ. 1958. Cytological studies of ar, intergeneric hybrid of fajamas rajan (Linn.)

Mllsp. and A Q h d a w, W. and A. Proc. Indian Acad. Sci. 47(B) : 252-262. i
N).IAR, L.S.S., and M.V. TBOMBRE. 1958. An-btergeneric hybrid of & (L.) -1lSp. X W.& A. J. Univ. POOM, 12: U-16.
d L.S.S., B.K. SWNi RPDI and M.V. 'MCIMBRE. 1966. Interspecific and intergeneric hybridization in the breeding of crop plants. The hpct of Mendelism on agriculture, biology and medicine. S. Rarrurnujam (Ed.) Indian J. Genet. P1. Breed. 26 (A) t U4-120.
KWUl, P.S. , N.C. -AM and D.G. FARIS. 1984. Nucleolar variation in a pigeonpea intergeneric hybrid. Widence for allosyndetic recarbination. Can. J. Genet. Cytol. 26: 499-505.
&, H., and MRBWI, T. 1976. Ovule and d r y 0 culture of the seeds .obtained form the crosses. between -. H . c a n n a b i n l s and H . B a b d a r i f f a . Jpl. J. Breed. 26: 298-306.
LADIZIEKY, G . , C.A. and T. E E K W 3 Y . Z . 1979. Wide crosses in soybeans: prospects and limitations. ELlFhytica 28: 421-423.
IAXBACH, F. 1929. Ectogenesis in plants. J. Hered. 20: 200-w.
LAMPRECHI', 8 . 1948. Losung des Artsproblens. Neue tmd bisheq bekannte Ergebnisse der Kreuzung L. x Eh. ooccineus L. und Reziprok. Agric. Genet. Landskrona. 6: 87-145.
kt W., ar?d.B. K 3 J C I m . 1976. The crossing of commn wheat (m ag&xam L.) w i t h cultivated rye (Secale cereale L.) I. Crossability, pollen grain germination and pollen tube growth. Euphytica, 25: 609-620.
Z R P X ~ U S , P.U.W. 1980. Tbe potential of and of hybrids with 2. yulgans as a wee crop for the U.K. Merories, Dept. Apll. Biol. Univ. Ca&ridger 51:20-22.
F, E.N., and H. RTS. 1960. The influence of gitberellic acid on the darelopent of hybrid barley ovules inYitm. Cand. J. Genet. -1. 2: 435-441.
'IAVANIA, u.C., and S. L A W . 1982. Chromosome banding patterns in eome Lndian pulses. AM. Bot. 49: 235-239.
m, P.L., and K.J. FRFY. 1976. Inheritance of grain yield in oat species crosses Ayena sativa L. x A. mteriles L. Egypt. J. Genet. -1. 5: 400-409.

LE bWtU4UDr G. 1971. Oabservations sur quelqws hybrid- dam le genre Phaseolua I. Le probleme des in-tibilities int$rspecifiques. Bull. Rech. Agron. Gembloue 6: 441-452.
LE &, 'G., R. m, arad J.C. BAMGT. 1976. Observations sur quelques hybrides dans le genre 111. P. hmaLus. Bull. Agron. Gemblauc ll: 183-200.
LEWIS, K.R., and B. JCBN. 1966. The meiotic consequences of spon+neous chrono- breakage. O-L 18: 287-304.
LI, B.W., K.K.S. YANG, dnd K.C. ED. 1964. ~ogenetical studies of Qcyza sativa and its related species. 7. Non-&chronization of mitosis and cytokinesis in relation to the fo-tion of diploid gametes in hybrids of P;LYIP &atjxa and Q. offlcinalis. Bot. Bull. Acad. Sinica 5: 142-153.
L I M , S.N. 1973. Meiotic instability in the plant genus Globba. chromsames Today 4: 321-334.
'LI~DE-LM.IRSEN, I. 1984. Nucleolar organizer polymorphism in barley, L. Heriditas, 100: 33-44.
'IUKCRI, L., and E. (7POUL. 1981. Study of the aminoacid spectrum in the seeds of U$ma raaiata (L.) wilckzek and Y. mung~ (L.) H-r. Parentdl forms and Direct line hybrids Bull. des Recherches Agronomiques de Gerbloux, 16: 299-308.
MACHADO, M., W. T A I , and L.R. BAKSR. 1982: m e n e t i c analysis of the interspecific hybrid Y i g ~ raalata x Y. uhellata J. Hered. 73: 141-146.
MALHdRA, R.S., and J.S. SODtiI. 1977. Genetic variability and discriminant function studies in pigeonpea (Cajans rajm (L.) Millap.) . Madras Agriculture Journal 64:677-678.
WiLIK, S.S. and B.B. SIN3-I. 1982. Note on the correlations in quantitative characters in an intersepecific cross of Soybean. Ind. J. Agri. Sci. 52: 608610.
WUK, C.P., and T.N. W. 1975. Qtogenetic studies in paprer Kt. Chromsome relationship between papver species of the section orthrhoeadw (P. pvrianm, f. sekuhws and 2. xinms and their mphiploids. Cytologia, 40x333-345.
MAMXE&P, P.C. 1974. Corn, Its Origin, hrolution and Inprovecent. The Belknap Press of Harvard W v . Cambridge, Plaasachusetts.
M, s.s., and T. SASAKUMA. 1977. Fertility of mphihqloids i n Triticinae. J. Hered. 68: 87-94.
'WWN; T.J., D.U. GEWEL, and J.L. APPLE. 1963. PEOC. Int. Tbb. Sci. Conar. 3rd,Sali&ury pp. 201-207.

W@&&L. R. 1971. Cbservations sur quelques hybrides dam le genre 11. Les phmmne6 meiotiques. Bull. Re&. Agron. Genbloux. 6: 461-489.
MdZINItXX, B. 1931. Cytological observations of deficiencies involving knawn genes, translocations, and an inversion in Zea mys. Univ. Mo. Agr. Exp. Sta. Bull. 163: 1-30.
McCLINIDCR, B. 1933. The association of non-hornlogous parts of chrmson-es in the mid-prophase of meiosis in Zea mays. Z. Zel.lforsch u. Mikr. mat. 19: 191-237.
MoaINlWX, B. 1938. The fusion of broken ends of sister half-chromtids follawing , chroanatid breakage at meiotic anaphases. Univ. Mo. Agr. Exp. Sta. Res. Bull. 290: 1-48. .
NcCMB, J.A. 1975. Is intergeneric hybridization in the leguninosae possible. Euphytica, 24: 497-502.
Mnrra, U., and H.Y. KBPWWl. 1980. Regeneration of plantlets from ootyledons of C. r;aian. Ind. J. Exp. Biol. , 18: 800-JOZ.
&. G. 1866. Versuche uber Pflanzenhybriden. Verh. Naturf. Brunfi. 4: 3-47.
KEJZCN, D., and P. BANEGT. 1973. Uber Speziati?nsvorgange in d e ~ , fattung y i d a L. Kulturpflanze 2l: 25-54.
MEYER, J.R. 1973. A study of reciprocal hybrids between upland cotton (- hksutm~ L.) and experimental lines with pkoplamm from ewen other species.Crop. Sci. 13: 778.
m. V.G. 1974. Emn. Bot. 28:5660.
~';D.W.S., M.C.MCP(, and A . ~ ~ ~ . 1978. Interspecific hybridization of PhasePlua ydg&.s with p. lunatus and 2. -. m r . -1. Genet. 52: 209-215.
MQSS, J.P. 1977. resistance of ArachiF; interspecific hybrids. Proc. Am. Peanut Res. EdClc. Asa. 9:34.
~ ' I ? s , YU, YU., and Z.V. VINITSVLS. 1975. Schlechtd. Dokl . Vsesor juznoi Ordena Lenina . ' Akad. Selqskokhzyaistvennykh Naik. Iueni V.I. Larina 7: 17-19.
-, G. 1980. Untersuchengen iber das pollenschalaucb Wachstm bei verschiedzen Trifolium - Art . -Kreuzungen. Der Zuchter, 30: 28-30.
MUXJZ, A.M., and R. ABPAMS. 1971. Inheritance of some quantitative characters in pigeonpea. Journal of the Agricultural Universi t!' of Puerto R i a , 551 : 22-43.

~ ~ S H I G E , T. and F. WCS. 1962. A revised medim for rapid growth and bioessays with tobacsn tissue culturee. physiol. Plant., 15:473-479.
WRASIGE, T. 1974. Plant propagation through tissue culture. Ann. Rev. Plant Physiol. 25: 135-166.
U NMlTGNI, S.P. 1941. Cytological studies on M i a n pllses. Part I. the soktic chramscsaes and prochrcamsomes of -. Proc. Nat. Acad. Sci. Mia. 11: 67-73.
WVASIIN, M. 1934. Chromsome alterations caused by hybridization and their bearing upon certain genetic problems. Cytologia, 5: 169-203.
tX&R, N.H. 1973. Origin and cytogenetics of rice. Genet. 17: 153-292.
Adv.
NERSON, C.H. 1970. A cytogenetic study in the Yicia sativa un!@lex - M.Sc. Thesis, Dept . of Genetics. The Hewbrew Un4versity - Jerusalaa.
NE%KL, C.A.8 and T. HYKWTZ. 1982. Successful wide hybridization between the soybean and a wild perennial relative, P. torrrentella Hayata. ' Crop. Sci. 22:1062-1065.
NEwEKL,C.A.,~~~T. HYKWTZ. 1983. Hybridization in the genus subgenus Glycjne Willd. ( w o s a e , papilionoideae) . Amer. J. Bot. 70: 334-348..
NI03LOFF, H . , M. w - K R I S l T X % M. KUNZEL and R. RIEGER. 1977. The behaviour of nucleolus organizers in structurally changed karyotypes of Barley. Chrowsoma (Berl.) 62:103-109.
NICQU~FFI H . , R. R I S E R , and G. KUNZEL. 1979. "Nucleolar &mimncew as observed in barley translocation lines with specifically reconstructed SW. chramsomes. Theor. -1. Genet. 55: 247-251.
PALNEB, R.G. 1965. Interspecific hybridization in the genus Glyche. MS Thesis, Univ. of Illinois, Urbana.
m. R.G., and H.K. HADLEY 1968. Interspecific hybridization in subgenus - m i Cmp. &i. 8: 55743.
h b D E Y , R.L. 1972. Inheritance of some quantitative Character€ in pigeon- (- &an (L.) M i l l s p . ) . M.Sc. thesis. J N K W Jabal~r , India.
PATIL, R.B., and A.B. D m . 1980. Inheritance studies ir pig-- G&W& (L.) Millsp.) . Journal of M.A.U. 5: 74-75.
RAOI M.V. 1968. Crossability of ccrmon wheat varieties in rye. Curr. Sci. 37:670671.

mIR, R.P.S., and R.B. SIEM. 1978. In: IOPB chroaoaome n d r reports =I. Askell m e (Ed.) Taxon. 27: 533-4.
FUNOIR, R.P.S. 1981. Relationship anwig fajanua, npsiea. *.D. mais, -as Bin& e s i g
Varanasi, -as.
LUrIEVSKY, E., andP. BWXTE. 1979. Cytogmetics of hybrids amng perennial species of subgenus GlysAne. Aust. J. Bot. 27 :713-723.
D.W.S. MX and M.C. MX. 1979. -A A ~ d early enbr-elo-t in recipr- interspecif ic crosses of -. Theor. -1. Genet. 54: 55-39.
'kSHAm, V. 1977. +lied aspects of a r y o culture. In Applied and Fundamental Aspects of Plant Cell, Tissue and Organ Culture (J. Reinert and Y.P.S. Bajaj, eds.), pp. 375-397. Springer Verlag, Berlin and New York.
RAMAN, V.S. 1959. Studies in the genus Atachifi w. Investigation on 30 chromsomed interspecific hybrids. Indian Oilseeds J.. 3: 157-161.
RAMAN, V.S. 1976. Cytogehetics and Breeding in Arachis. Today and Tonorrow's Printers and Publishers, New Delhi.
'M, B.K.S., and S. -. 1975. The effect of gamna radiation on cell proliferation and regeneration in explanated tissues of pigeonpea, Cajarus &an (L.) Millsp. Rad. Bot . , 15: 301-305.
k, .N.G.P. and R.P. REIDY. 1975. Samtic variation in Ca,enus -. Curr. Sci., 44:816-817.
RPD8 N. K. 1978. In: IOW chromosome nmber reports W I . Askell Love (Ed.) Taxon .27: 544.
REDDY,-L.J. 1973. Inter-relations hi^ of and species as revealed by hybridization and pachytene analysis. Fh.D. Thesis, Indian Institute of TecLPlology, Kharagpur, India.
'REEDY8 L.J., J.M. GREEN, U. SllGH, S.S. BISSEN8 and R. J v . 1979. Seed protein studies on Sajaus szajan,
and same hybrid derivatives. In Proceedings on Seec ~ I u p r o v e m e n t in Cereals and Grain LegLrmeS, IAFA, Vienna, 2: 105-117.
h, L.J., J.W. GREEN8 andD. SHMW&. . 1980. Genetics oj s&an (L.) Killsp. x spp. Proc. Inter
Workshop on Pigeonpeas, 2: 39-50, lWSAT, India.
' m y , L.J. 1981~. Pachytene :. ;:yse=, in S&anlls

and their hybrid. Cytologia, 46: 397-412.
- V L.J. 1981b. Pa-- rmolyecs in egEirien ~d S a b U U a x atvlosia Bericea hybrid. Cytologla. 46%
561-577.
L.J., and D.N. DE. 1983. Cytormrpbological studies in &an x -. Ind. J. G n e t . P1.
Breed., 43: 96-103.
m, M.K., and N.C. -AM. 1985. G m o m e relationship between. Wxdew gmxxx.m (6x) and lechlerl ( 6 x ) . Genetika (In press) .
'=>R.P., M.A. AZW, K.V. RAO, and N.G.P. RPD. 1979. Corrbining ability and selection index in F2 crosses of pigeonpea crosses. Indian Journal of Genetics 39: 247-254.
REEVES, R.G. 1950. The use of teosinte in the irrprov€srent of corn inbreds, Agron. J. 42: 248-251.
w, R.G.,and A.J. BOCRHDLT. 1964. Wification and inprrovmt of a maize inbred by crossing it with - Cram Sci. 4: 7-10.
'-AN, P. 1980. The wild gene p o l of & at IaSAT, present and future. Proc. Inter. Workshop on Pigeonpeas, 2: 29-36, ICRISRT, India.
RI&, C.M. 1982. The potential of exotic gemplasm for tomato inprovement. Plant lmprovanent and somatic cell genetics, Aced6nic press, New York (ed. Indta K. V a s i l , William, R. Scowcraft and Kenneth J. Frey) 1-28.
RILeY, R., and V. CHAF+lW. 1958. The production and phenotypes of wheat-rye chrmsome adition lines. fleredity 12: 301-315.
RTLEY, R. 1960. The secondary pairing of bivalents with genetically similar chronvsomes. Nature, 185: 751-752.
F¶X, >., and D. N. DE. 1965. Intergeneric hybridization of and Sci. Cult. 31: 93-95.
'IIM, B. 1933. Studies in the developumt of the fenale gametophyte in some legu?ninous crop plants of India. Indian J. AgrJc. Sci. 3: 1098-1107.
'EmUSF', W. 1953. Neue Beobachtungen an Bastarden von L. rmltiflarus a. und. S . , 1,. mngr-
Genetics: 844-845.

N., and W.C. GRDXRX. 1967. Interepecffic cross ampatibility between the cultivated peanut, +&ui L. and other menbers of the genus
hvaaaea . Oleagineur 22: 455-459.
J.N., and L.S. BM3(MAN. 1970. Natural hybridization of BaasQlU vulaarie L. x Ebaamlu -L. J. Amer. Soc. Hort. Sci. 95: 659461.
IWPSER, J .N. 1970. Interspecific hybridization bet- cultivated American species of the genus - m y t i c a 19: 480-489.
'sADAsIVAIAFI, R.S . , and K.J. KASHA. 1971. Meiosis in Haploid Barley . - An Interpretation Of Non-Bornlogous chronwscme associations. Chromsorra (Berl. , 35: 247-263.
&JAPPA, M., and N. .SA!lWYAXWX. 1979. In: IOPB chrorrwsome nw&r reports LXN. Askell Love (Ed.) Taxon. 28: 393-395.
SASrT(I* D.C., and K.R. SHIVANNA. 1976a. Att- to overcome interspecific incompatibility in seMrmrm usmg recognition pollen. Ann. Bot . (London) 41: 891-893.
'SMIF?fr D.C., M.S. WILINI, and J.P. MsS. 1981. Tissue culture and prospects of crop inpswement in &acU& and other oilseeds crops. ;Zn: Tissue Culture of Emnormcally Important Plants in Developing Countries (A.N. Rao, ed.) &p. 42-57 -
'GASPRI;D.C., and J.P. KSS. 1982. Effect of gr& regulators on incaptible crosses in the genus Arachis L. J . of Fxpt. Bot. 33: 1293-1301.
SASI'RID D.C., and J.P. M36S. 1982. Effects of. growth regulators on incompatible crosses in the genus Arachls L. J. -1. Bot. 33: 1293-1301.
SASlR.I, D.C., and J.P. IKSS. 1982. Hormonal requir-t in inampatible crosses in the genus hachis L.J. Exp. Bot. 33. 129371301.
'SA-, .N. 1981. Hybridization of the species and a. mcclneus L. 111 fertility and yield of
hybrids in F2. Genetika. Selektsiya, 14: 338-345.
M. 1973. (XI the interspecific hybridization between the adzuki bean, #rageolus , (wILLD.) W.F. WRTQ?T and the green grm, I-L. I. cro~sing betyeen a eultivar of the green gram and semi-wild relative of adzuki been, in emhnic name 'Bakaso'. Japan. J. Breed. 23: 6166.
SAXENAD K.B., and D. SHARMA. 2961. Variation for leaf ctlaracteristics and relationship klth grain yield end seed size

in pigeonpa. Ind. J. Genet. Plt. Breed. 41: 340-345.
K-B.r M.D. Md D-G- FARIS. 1984. nonflowering uniculm mtant of pigeonpea. Inter. Pigeonpea News lett. 1984. 11-12.
KUK%AET, W.R. and P.J. IARRIN. 1982. Scmaclonal variation: A new option for plant inprwement. In Plant Iroprovement and Somptic Cell Genetics. Academic Press, w.159.
SEARS- E.R. 1972b. Stadler Genet. Syxp, 4: 23-38.
&, E,R. 1975. In Handbook of -tics (R.C. King, ed.) Plenm, New York. 2: 59-91.
&AREA,D.,L. SIIGI3,S.S. BAGHEL, and H.K. m. 1972. Genetic analysis of seed size in pigeon* (- -. Can. J. Genet. Cytol. 14: 545-548.
b, P.C., and P.K. C;UPIIA. 1982. Karyotypes in some pilse crops. Nucleus, Calcutta 25: 181-185.
=I-, T., and S. -. 1981. Intergeneric hybridization between and eardelrm bulbosum. Jp. J. Genet. 56: 50-515.
SHB, C.T., A. RAWXOAFgHANIIA, M.C. MHI and D.W.S. MX. 1982. m r y o developnent in reciprocal crosses of L. and p. c- Lam. Theor. -1. met. 62259-64.
-Am, M.P., D. SHARMA, and LAXIWLN S I m . 1973. Karyotyp analysis in 15 varieties of CaSannr rpian (L.) Millsp. and AQb6i.a (W. & A.) Cytologia 38: 219-227.
S m , P.S., and T.S. SANMRI. 1980. Genetic analysis of grain yield and other characters in pigeonpea (Caianufi (L.) Millsp.). Available from the authors.
SIEW, A.K., and J.P. KSS. 1984. Utilization.of wild relatives in genetic irrprwenent of A+da L. VI. Fertility of triploids: Cytological basis and breeding inplications. Peanut science, 11: 17-21.
'SIFXX, B.V., M.P. PAM)=, and B.P. PANDYA. 1980. Pod set in pigeonpea. Ind. J. Genet. Plt. Breed. 40: 568-572.
SIhM, D.P. 1981. Breeding for resistance to diseases in Green gram and Bladc gram. Theor. -1. Genet., 59: 1-10.
~b, A.K., and D. N. DE. l9,61. Q'tologicdl studi* of two species of -. Bull. Bot. Soc. Beng. 21: 25-28.
- S m , S.S.N., and P. KVMAR. 1979. Kitetic analysis in th1rt-r. v a r i e t i e s of & (L.! ;<i:l~p. Cyt010giar

SMAlUT, J . 1970. Inter specific hybridization betweM cultivated American species of the genus E ~ ~ ~ R A L Ebg&Aca Ip; rsb-rss.
~ L ~ a r A d ~ GEGmxs J S L B- between---
L. and other &rs of the genus -hnoaaea Ezk Oleeginw 22:
4557459.
d, 8.8. 1968. Recent cytogenetic studies in the genus -. ?dv. in Genet. 14: 1-54.
SNAPE,J.W.,V. CHARWJ, J. m3SS, C.E. and T.E. MILLER. 1979. The crossability of wheat varieties with &rdew~ bulbosum. Heridity, 42:291-298.
-, N.A., and V.P. BURGIL. 1970. A spontaneous intergeneric hybrid in the vetch tribe and its characteristics (Ru.) Otdalen. gibridizatsiya rast. Mosum. USSR, Kolos. 415-421.
'SOBOLEV, N.A., E.D. ZHIIA* and G.K. ZUWCRUS. 1968. Intergeneric hybridization in the vicieae tribe (Ru.) Rastenievodstro, selektsiyai lesovodstro. No.1 Moscow, USSR, Kolos. 86-94.
GPRINZE3, D.W., and R.C. BUCRNER. 1982. A efotic examination of L~li,m rmlltiflorumm. xFestuca Schreb. F1 Hybrids. Crop Science, 22:305-309.
e m , H.T. 1980. Utilization of wild species for crop inprwement. Adv. Agron. 33: 111-147.
-INS, G.L., Jr. 1950. Variation and Evolution in Plants, 643 w. Colhia Univ. Press. New York.
SEBBIKSr G.L. 1958. The inviability, weakness, and sterility of interspecific hybrids. Adv. Genet. 9: 147-215.
SEBBIKS, G.L. 1971. Chromosomdl evolution in higher ' plants. Edward Arnold Pub. Ltd., London, 117-120.
STEBBIM, G.L. 1977. Processes of organic evolution. Third ed. Prentice-Hall, Inc. New Jersey.
what is new in nlethodology of breeding fields beens, peas & . . & m h a ~ . I D ? > \ -1 I h n r i r A a t . Mn-- 98-102-

< - SreWAFa', J.M. 1981. In ~ L L ~ Q fertiliaatian and embryo r-. gvirnn- Exp. E o t . 21: 301-305.
N.C. 1977. Eaploidy from fiqrderrm interspecific crosses. I. Polyhaploids of E. and H. A-. Theor. -1. Genet. 99: 209-217.
-At?, N.C. 1978. Ehploids and hybrids following interspecific crosses. Barley Genet. -1, 8: 97-99.
-AM, N.C. 1978. Meiosis in polyhaploid Herhizygous ineffective control Of diploid-like behaviour in a Aexaploid? Chromosorm (Berl.) 66: 185-192.
'-AM; N.C. 1979. IIELp10idy from Hordeurn interspecific crosses. 2: Dihaploids of 8- and 8. -. Theor. -1. - Genet. 55: U9-144.
'-AM, N.C. 1980. Haploidy from crosses. 3: Trihaploids of B. Theor. -1. Genet. 56: 257-263.
< -AM, N.C., and A-A. AZAD. 1978a. Trisomic analysis of ribosomal FWA cistron itultiplicity in barley (- L.) . Chroro?, 61: 255-264.
-AM, N.C., and A.A. AZAD. 1978b. Nucleoli and ribosomal EINA cistron nmbers in Ilordeum species and inter specific hybrids exhibiting supression . of secondary constriction. Chromosomal 69: 265-273.
-AM,N.C. andK.J. KAsH&. 1971. Increased barley haploid production follwing g-rellic acid treatment. Barlev Genet. Newslett. 1: 47-50.
\
-AM, N.C.# and K.J. KASHA. 1973. Selective chromosome elimination during haploid formation in barley following interspecific hybridization. C h r o w m , 42: 111-125.
-! M.S.I and M.L. m. 1961. Origin and * cytogenetics of the comnercidl potato. Adv. in Genet. 10: 217-256.
SYBEEX;AI J. 1966. The zygomere as hypothetical unit of chrmsome pairing initiation. Genetics, 37s 166.
J. 1972. General CytogeneticS. North-Holland Publ. Co. Amsterdam, 158-159.
TAN-BOUN SUY. 1979. Observation using fluorescence of the growth of the male gametophute in PtwseDlue; L. and in an interspecific hybrid. Bull. des Rechercbes Agronomiques de - 1 0 ~ ~ 14: 97-108.
'TAN;, w:T., and C.H. C-IEN. 1953.. Preliminary studies on the

hybridization of the cultivated and wild bean m r i l and E. (medtawal. J. Agr. Assoc. Qlina N.S. 281 17-23.
CAEX;, m. , and G. IIAf. 1962. Studies on the genetic behaviour of both quantitative and qualitative characters in the progeny of an interspecific cross of soybean w x G. (2nd rep.) J. Agr. Assoc. china N.S. 42: 13-25.
pyx;, W.T., and G. I1AI. 1963. Studies on the qualitative and quantitative inheritance of an interspecific cross of soybean, Glvcine lmx x G. fsmmama. Bot. Bull. Acad. Sinica 3: 39-60.
l b W , P.T., and S.A. ReVELL. 1946. Secx,""r,y association and heterodrrotmtln attraction. 1. a-. Ann. Bot., -10,: 159-164.
% b F S , H. 1964. Investigations into the inter-relationships of vulaarls L. and E - m c c i n e u s Lam. Genetica35:
59-7 4.
'kk, C.L. 1946. Genetic studies on the wild and cultivated SO-, J. Am. SOC. AgrM. 38: 381-393.
(- L.) und Feuerbone (Phaseolyh Lam.) und ihre mentuelle Praktische Verwertbarkelt. Zuchter
van2er -EN, L.J.G. 1980. Taxonomy of cajanus. proc. Inter. Nat. Worksbp on pigeonpa, 2:9-13.
'VASIL~ I .K., W.R. and K.J. F'REY. 1982. Plant hp.rovanent and -tic cell genetics. Academic Press. York.
WENAAR, E.B. 1968a. Meiotic restitution and the origin of plyploiQ. I. Influence of gmotype on Polyploid seed set ir a ! . r u W - x T. Q u g i b n hybrid. Canad. J. Genet. Cytol 10: 836-843.
E.B. 1968b. Meiotic restitution and the origin of polyploidy. 11. Prolonged duration of rnetaphases I as causal factor of restitution indhrction. Canad. J. Genet. Cytol. 10: 844-852.
WATAWBE, R., and T. WPPIW. 1958. (Studies on tht interspecific hybridization of wmmn vetch. Y k i a &&~YB, am Yahazuendo Y. anaustlfblia ~arr. (Eng. sunmuy) Bull. Net. Inst. Agric. Sciences, Japan, series G I 15: 1090145.
hTgER, C.R. 1950. Iowa State Agric. W t . Sta. Res. Bull. 374.

W5ER, D.F., Md D.E. ALEUMlER. 1972. Redundant segmmts in Zea mays detected by transactions of mnoploid origin. &txm6oma (Berl.) 39: 27-42.
G.T. 1950. Fertility relationships and meiosis of interspecific hybrids in m. Agron. J. 42: 315-322.
WMTE, P.R. 1954. The culture of animal and plant cells. New Yod: Ronald Press.
'WI~%IAMS# L.F. 1948. Inheritance in a species cross in the soybean (abst) . Genetics 33: 131-132.
yzwaczir, F. , M. HASDWlD and.1. Nishiyarm. 1976. Cytogenetics of the progeny of a highly sterile hybrid Jmg&&x& x 8. strigosa. Japan J. Genetics, 51(2):109-113.
. a
YAtWmD, K. 1971. On the secondary balanced karyotypes of Mm + S m + S and M-m + a M -m. Tec. Bull. Fac. Agr. Kagawa U. 22: 1-8.
YAWNlX)* K. 1974a. (X1 the interspecif ic hybrds between Xi& and Y. ancrustifolia, Y. gUwa and Y. -. Tec.
Bull. Fac. Agr. KagaWa U. 25: 179-190.
'YAtMWlD, K. 1974b. Ilybrid plants between two races of W having 211314 chrom~mes. Japan. J. Breed.
24: 15-22.
YAFWwlD, K. 1975. Estimation of genetic homogeneity bj isozymes from interspecific hybrids of W, Japan. J. Bred. 25:. 59-64.
ZR~&# M., and G.F. FRGYTPG. 1982. Phadeolus germplas resistant to six strains of X- in Puerto Rico. Phytopatholog~~ 72: 174.
Z m ; A.C., and C. VAN HEPIEKP. 1970. Germination of polla of weed rye (- seg&&e L.) on wheat (w aestlvlm L.) 6tigm.s and the growth of pollen tubes. Euphytica, 19:175-179.
KEWRY, D., and M. WPF. 1973. Domstication of pllses in thc Old World. Science 1982: 887-894.
ZOBZIRY, D., and U. PLIWANN. 1979. Chromsome pdlymorphism, hybridization and colonization in the Yicia sativa grOuF (Fabaceae). P1. Syst. Evol. 131: 143-156.
*CONTI, L . 1383. Results of crossing PhaseoZus vuZqaris x P72 coccineus. Genetica A ~ r a r i a , 37:155 .

PUBLICATION FROM THE THESIS

1. P. Sateesh Kumar, N.C. Subrahmanyam and D.G. faris, 1982. Studies on developing pigeon-pea haploids. International Pigeonpea llewsletter 2 : 14-15.
2. P. Sateeah Kmar, N.C. Subrahmanyam and D.G. Paris, 1984. Nucleolar variation in a pigeonpea intergeneric hybrid: Evidence for allosyndetic recombination. Can. J. Genet. Cytol. Vo1.26(5): 499-505.
3. P. Sateesh Kmar, N.C. Subrahmanyam and D.G. Faris, 1984. Inter- 'generic hybridization in pigeongea: I. Effect of hormone treatments. Field Crops Research (In Press).
4. P. Sateesh Kumar, N.C. Subrahmanyam and D.G. Faris, 1984. In vitro regeneration of pigeonpea from cotyledons. International Pigeonpea Newsletter 3:15-16.
5. P. Sateesh Kmar, N.C. Subrahmanyam and D.G. Faris, 1983. Allosyn- detic recombination in CAJANUS-ATYLOSIA hybrid. Abstracts XV International Congress of Genetics, New Delhi. pp 671.
6. P. Sateesh Kmnar, N.C. Subrahnyam and D.G. Faris, 1984. Plantlet regeneration from inmature embryo s of pigeonpea, International Pigeonpea Newsletter 4 (In Press).
7. P. Sateesh Kumar, N.C. Subrahmnayam and D.G. Faris, 1985. Morpholo- gical variation and inheritance in a pigeonpea intergeneric hybrid. Current Science (In Press).
8. N.C. Subrahmanyam. P. Sateesh Kumar and D.G. Faris. 1984. Genome relationships in pigeonpea and its implications in crop improvement. Symposium on Advances in Genetics and Crop Improvement, Meerut. Abstract. p 11.
Copyright © 2022 FDOKUMEN




















