
Rapid Differentiation of Closely Related Citrus Genotypes by Fluorescence Spectroscopy
Upload
independentCategory
view
2download
0
Aust. J. Bot., 1993,41,23-34
Comparative Geographical Variation in Seedling Morphology of Three Closely Related Red Mahoganies, Eucalyptus urophylla, E. pellita and E. scias
K. Pinyopusarerk, B.V . Gunn, E. R. Williams and L. D. Pryor
Division of Forestry, CSIRO, PO Box 4008, Queen Victoria Terrace, ACT 2600, Australia.
Abstract Twenty two populations of Eucalyptus urophylla, 13 of E. pellita and two of E. scias were selected throughout the species' natural distributions in Indonesia, Papua New Guinea and Australia and the morphology of seedlings compared under glasshouse conditions. Leaf length, width, 1engWwidth ratio, base angle, stem shape and, to a lesser extent, intranode length provided good discrimiiation between species and provenances.
Canonical variate analysis revealed four groups, two of which contain E. urophylla, one E. pellita and one E. scias. Wetar Island provenances formed one of the two E. urophylla groups, distinguished from the other group consisting of Alor, Adonara, Flores, Pantar and Timor provenances on the basis of narrower leaves, greater lengwwidth ratios, more acute leaf base angles and square stems. Seedling leaves of E. pellita were generally longer and broader than the other species, with a tendency for a separation between the northern occurrences (New Guinea and northern Cape York Peninsula, Queensland) and southern occurrences (Helenvale to Rockhampton, Queensland); those from the north had smaller leaves and more distinctly square stems. E. scias is clearly distinguished by its narrower leaves and longer intranode length.
Introduction The genus Eucalyptus has been divided (Pryor and Johnson 1971) into several (informal)
subgenera. Eucalyptus urophylla, E. pellita and E. scias are included in the subseries Resiniferinae within the subgenus Symphyomyrtus. Other species in the subseries are E. notabilis, E. resinifera and an undescribed species.
E. urophylla S.T. Blake is one of only two eucalypts not indigenous to Australia (the other being E. deglupta). Throughout most of its range E. urophylla is a forest tree, reaching 25-45 m in height and up to 1 m diameter, with a straight bole for half to two-thirds of the tree height. Exceptional specimens attain 55 m in height and up to 2 m diameter (Turnbull and Brooker 1978; Jacobs 1981). The species is economically important for wood production on moderate to good quality sites, especially in the seasonally dry lowland tropics. It has been planted extensively in Brazil, and is receiving increasing interest in South Africa, China and South-east Asia.
The known natural occurrence of E. urophylla is restricted to seven islands of the lesser Sunda Achipelago, viz. Flores, Adonara, Lomblen (Lembata), Pantar, Alor, Wetar and Tirnor (Fig. 1). It grows extensively on Alor, Wetar and on Timor above 1000 m, but is less common on the other islands. Latitudinal range is 7'30'-10"s and longitudinal range 122L127'E. It has the greatest altitudinal range of any eucalypt and occurs from 90 m on Wetar to nearly 3000 m on Mount Tatamailau, Timor. Its main occurrence is between 300 and 1100 m. E. urophylla occurs on slopes and valleys with varying aspects, soil types and parent material (basalts, schists and slates) but it is largely absent from calcareous soils.

K. Pinyopusarerk et al.
Fig. 1. Natural distribution of E. urophylla in Indonesia. Numbers indicate location of provenances used in the study.
E. urophylla appears to be phenotypically polymorphic, segregating into different forms on the basis of variation in altitude and aspect. The populations are diverse in a number of features, especially in the degree to which rough bark persists on the stem and branches. The appearance of the bark seems to vary with the altitude and dryness of the site. Martin and Cossalter (1976) reported that stands growing at low altitude and under drier conditions had a strong tendency towards a smooth bark to within 2 m of the base. Conversely, the occurrence of rough bark, though variable, is common at higher elevations and under moister conditions. It has been suggested that E. alba, which commonly occurs in association with E. urophylla at lower elevations, may have an influence on the smooth bark characteristic in interspecific hybrids. Although occasional interspecific hybrids have been detected in the field, it seems unlikely that there is extensive genetic breakdown between the species and this is unlikely to be the source of the bark variation. There is also considerable variation in other features such as fruit shape and size, bud characters and adult leaf characteristics such as lengthbreadth ratio and expression of a 'drip tip'. Nevertheless, there appears to be no consistent pattern that readily suggests the recognition of additional taxa, although further detailed study may lead to this conclusion.
E. pellita F. Muell. is regarded as a species related closely to E. urophylla (Turnbull and Brooker 1978), and has shown great potential for plantation forestry. The tree varies in size from a stunted multistemmed form to a large tree up to 40 m in height and over 1 m in diameter. The species has two main natural occurrences. In New Guinea it is found from as far north as the Muting area of Irian Jaya and Morehead District of Western Province, Papua New Guinea. In Queensland it extends from Iron Range near the top of Cape York Peninsula to just north of Townsville (latitude 12'45'-19"s). Boland et al. (1984) also reported isolated occurrences in southern Queensland including Byfield north of Rockhampton. Recent taxonomic review of some of the botanical material in E. pellita has led to the view (L.A.S. Johnson 1991, personal communication) that the material from New Guinea and the northern part of Cape York Peninsula should be recognised as a separate taxon within the subseries Resiniferinae. Fig. 2 shows the approximate division between northern and southern E. pellita.
In New Guinea E. pellita is known to occur from latitude 7'30' to 8'35's and from longitude 140'56' to 141°45'E, at altitudes of 30-70 m. It is common as scattered populations in this area, occupying a specific niche between open areas of impeded drainage and rainforest. It is therefore confined mostly to narrow strips less than 20 m wide. On Cape York Peninsula it is an uncommon component of transitional forests and woodlands mainly on the western side of the ranges, at latitudes from 12'30's to c.14'S (J. Morse 1991, personal communication) and an altitude range from near sea level to about 600 m.

Seedling Morphology of Three Red Mahoganies
I I
1 20° 130" 140' 150' 0"
0'
I
10
10-
NORTHERN
TERRITORY 2 0
20 QUEENSLAND
WESTERN A U S T R A L I A
AUSTRALIA
AUSTRALIA 30
30-
Pig. 2. Natural distribution of E. pellita in New Guinea and Queensland and E. scias in New South Wales. Broken line indicates approximate division between northern and southern populations of E. pellita. Numbers indicate location of provenances used in the study.
E. scias L. Johnson & K. Hill, with three subspecies, scias, callimastha and apoda was previously designated E. pellita (Johnson and Hill 1990). E. scias is a tree to 20 m high, or mallee in exposed situations. According to Johnson and Hill (1990), the species occurs in New South Wales, having a sporadic and disjunct distribution usually in coastal ranges or hilly country from Tenteri5eld in the north, and from Cessnock to Batemans Bay.
Variation in seedling characters, including leaf size and shape, has been found between populations for Eucalyptus species such as E. leucoxylon (Boland 1978), E. ovata and E. brookeriana Udiges et al. 1981) and E. nitens (Cook and Ladiges 1991). Earlier studies of seedling variation in E. urophylla revealed provenance differences in many leaf characters. Moura (1977) reported that leaf size of E. urophylla seedlings decreased with an increase in altitude of the seed sources. Charomaini (1990), using provenance samples from a limited range, found provenance differences in leaf characters of 10 month old plants of E. urophylla from Timor (three provenances), Flores and Adonara, particularly leaf width, which were

K. Pinyopusarerk et al.
si-cant, while variations in leaf length, length/width ratio and petiole were suggested but not statistically significant. Given the taxonomic relationship and economic importance of E. urophylla and E. pellita, it is desirable to examine the intraspecific variation of these species over their natural geographic range. A study of such variation can also help in the selection of provenances to be included in future provenance trials. The aim of this study was to determine the pattern of variation, based on seedling morphology, in E. urophylla, E. pellita and E. scias and assess the relationships of the three taxa as revealed by seedling features. Of special interest was the comparison between New Guinea E. pellita and E. urophylla because of their geographical proximity to each other.
Materials and Methods Twenty two provenances of E. urophylla covering the range of the species' natural distribution were
used in the study. Of these, seed of individual trees was kept separate (family-identified) in 16 provenances and bulked in the others. Thirteen provenances of E. pellita were examined. These were split into two groups: seven provenances from New Guinea and northern Cape York Peninsula and six from the southern part of its range. Two provenances of E. scias were included. Seed of each provenance of the latter two species was bulked except for three family identified provenances of E. pellita from New Guinea. Two other eucalypts, E. alba and E. grandis, were grown in the same glasshouse for general observation but not included in the analysis. Details of origins of provenances are given in Table 1.
The experimental design was a generalised lattice design for 200 entries, four replicates divided into incomplete blocks of size four. Thus there were 50 incomplete blocks per replicate. The computer- generated lattice design was modified so that there were added properties for the arrangement as a row and column design (Williams and John 1989).
The 800 seedlings (four replicates of each of 200 treatments) used in the study consisted of the following:
(1) four seedlings (replicates) from each of five families in each of the 19 family-identified provenances (four replicates each of 95 entries). These were used to examine variation between families within provenances;
(2) twenty seedlings from each of the 21 bulked provenances (four replicates of each of 105 entries). Seeds were sown in November 1990 in punnets containing vermiculite which were placed on a
warm bed (25OC) under a regulated mist spray. Three weeks after sowing, seedlings were pricked out into 15 cm pots (one seedling per pot) containing 2 parts loam, 1 part river sand, 1 part vermiculite, 1 part perlite and 1 part peat moss. Newly transplanted seedlings were placed in a shade house for 3 weeks before being raised in a temperature controlled glasshouse in Canberra with a 30125OC daylnight regime in natural daylight from January to March 1991.
Pots were watered from overhead sprinklers three times a day (at 0800, 1300 and 1800 hours), 5 min per session. Slow-release fertiliser in pellet form (Osmocote, N14:P6.1:K1.6) was applied at the amount of 4.5 g per pot. Additional application of YZ strength Hoagland (No. 2) nutrient solution was provided. Insects, mainly psyllids, were controlled by a regular spray of pyrethrum.
Assessments were made on each seedling following 2 months in the glasshouse; the seedlings were about 3 months of age from germination. The lower leaf at node 9 was used to measure all leaf variates and a number of stem characters was also assessed. Characters measured are given in Table 2. The location and nomenclature of these measurements are shown in Fig. 3.
Statistical Analyses
Analyses of variance were carried out for each of the variates to assess (1) differences between families within provenances and (2) differences between provenances. The statistical package GENSTAT 5 (Payne et al. 1987) was used for the analyses of data. The data for stem glaucousness were omitted because of the general lack of this character except for the Cessnock provenance of E. scias.
Estimated provenance means were subjected to a canonical variate analysis. By using this multivariate technique, it is possible to summarise the differences between islands (in the case of E. urophylla) as well as differences between species. In addition, regression analyses were conducted to investigate relationships between variates and the geographic locations of provenances (latitude, longitude and altitude).
Seedling Morphology of Three Red Mahoganies
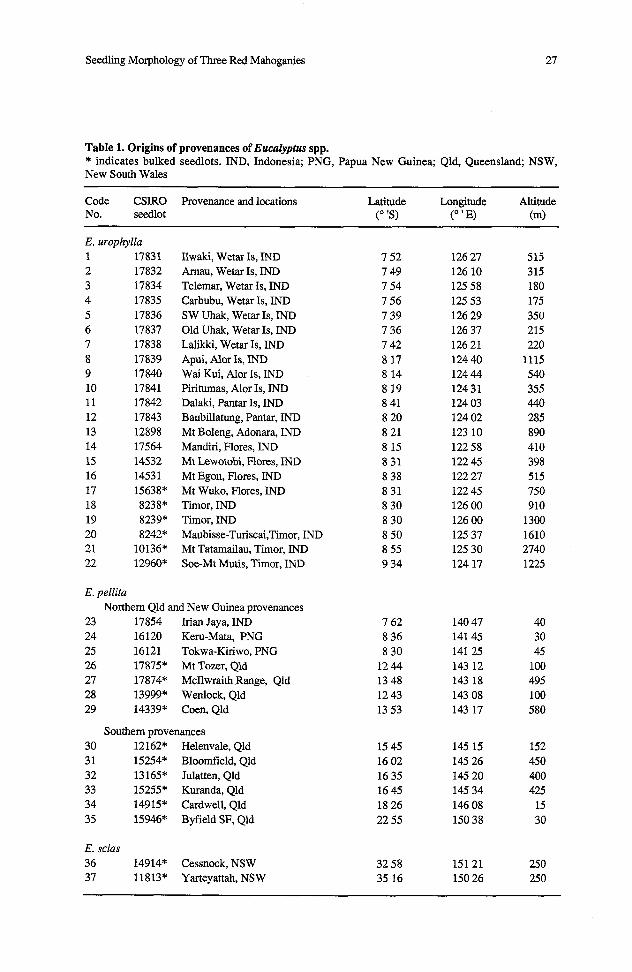
Table 1. Origins of provenances of EucaZypfus spp. * indicates bulked seedlots. IND, Indonesia; PNG, Papua New Guinea; Qld, Queensland; NSW, New South Wales
Code CSIRO No. seedlot
Provenance and locations Latitude (" ' 9
Longitude ("'E)
Altitude (m)
E. urophylla 1 17831 2 17832 3 17834 4 17835 5 17836 6 17837 7 17838 8 17839 9 17840 10 17841 11 17842 12 17843 13 12898 14 17564 15 14532 16 14531 17 15638* 18 8238* 19 8239* 20 8242" 21 10136* 22 12%0*
Ilwaki, Wetar Is, IND Amau, Wetar Is, IND Telemar, Wetar Is, IND Carbubu, Wetar Is, IND SW Uhak, Wetar Is, IND Old Uhak, Wetar Is, IND Lalikki, Wetar Is, IND Apui, Alor Is, IND Wai Kui, Alor Is, IND Piritumas, Alor Is, IND Dalaki, Pantar Is, IND Baubillatung, Pantar, IND Mt Boleng, Adonara, IND Mandiri, Flores, IND Mt Lewotobi, Flores, IND Mt Egon, Flores, IND Mt Wuko, Flores, IND T i o r , IND Timor, IND Maubisse-Turiscai,Tirnor, IND Mt Tatamailau, Timor, IND Soe-Mt Mutis, T i o r , IND
E. pellita Northern Qld and New Guinea provenances
23 17854 Irian Jaya, IND 24 16120 Kern-Mata, PNG 25 16121 Tokwa-Kiriwo, PNG 26 17875" Mt Tozer, Qld 27 17874* McIlwraith Range, Qld 28 13999* Wenlock, Qld 29 14339* Coen, Qld
Southern provenances 30 12 162* Helenvale, Qld 3 1 15254* Bloomfield, Qld 32 13 165* Julatten, Qld 33 15255* Kuranda, Qld 34 14915* Cardwell, Qld 35 15946* Byfield SF, Qld
E. scias 36 14914* Cessnock, NSW 37 11813* Yarteyattah, NSW
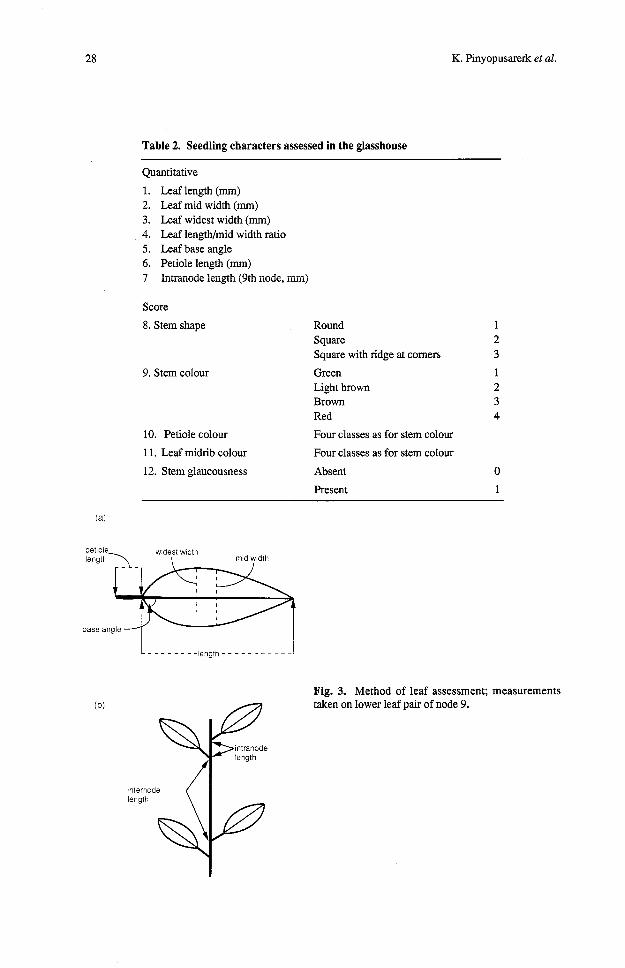
Table 2. Seedling characters assessed in the glasshouse
K. Pinyopusarerk et al.
Quantitative
1. Leaf length (mm) 2. Leaf mid width (mm) 3. Leaf widest width (mm) 4. Leaf lengwmid width ratio 5. Leaf base angle 6. Petiole length (mm) 7 htranode iengrh (9th node, ~ l m j
Score
8. Stem shape Round 1 Square 2 Square with ridge at comers 3
9. Stem colour Green 1 Light brown 2 Brown 3 Red 4
10. Petiole colour Four classes as for stem colour
1 1. Leaf midrib colour Four classes as for stem colour
12. Stem glaucousness Absent 0
Present 1
widest w~dth I mid width
base angle
Fig. 3. Method of leaf assessment; measurements taken on lower leaf pair of node 9.
Seedling Morphology of Three Red Mahoganies
Results Preliminary analyses of variance showed that for this trial the detailed blocking structure
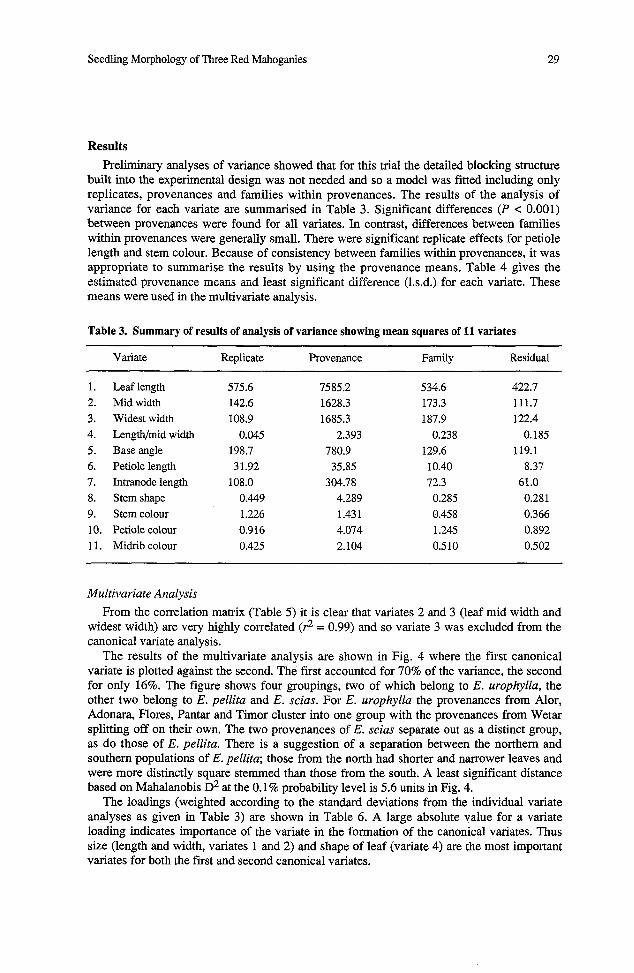
built into the experimental design was not needed and so a model was fitted including only replicates, provenances and families within provenances. The results of the analysis of variance for each variate are summarised in Table 3. Significant differences (P < 0.001) between provenances were found for all variates. In contrast, differences between families within provenances were generally small. There were significant replicate effects for petiole length and stem colour. Because of consistency between families within provenances, it was appropriate to summarise the results by using the provenance means. Table 4 gives the e s t i a t e b provename means and least significant difference (1.s.d.) for each variate. These means were used in the multivariate analysis.
Table 3. Summary of results of analysis of variance showing mean squares of 11 variates
Variate Replicate Provenance Family Residual
Leaf length Mid width Widest width Length/mid width Base angle Petiole length Intranode length Stem shape Stem colour Petiole colour Midrib colour
Multivariate Analysis
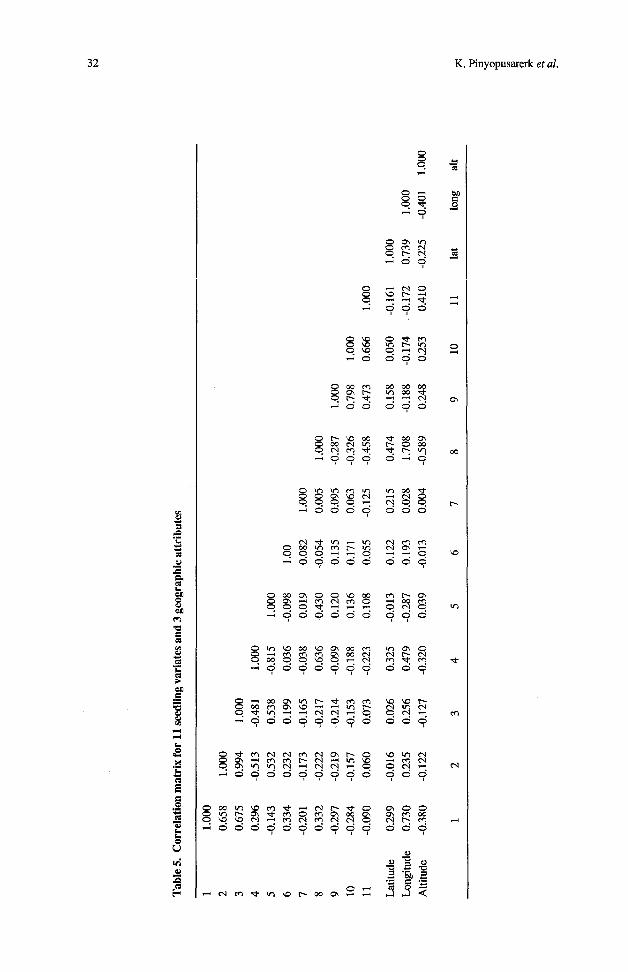
From the correlation matrix (Table 5 ) it is clear that variates 2 and 3 (leaf mid width and widest width) are very highly correlated (r2 = 0.99) and so variate 3 was excluded from the canonical variate analysis.
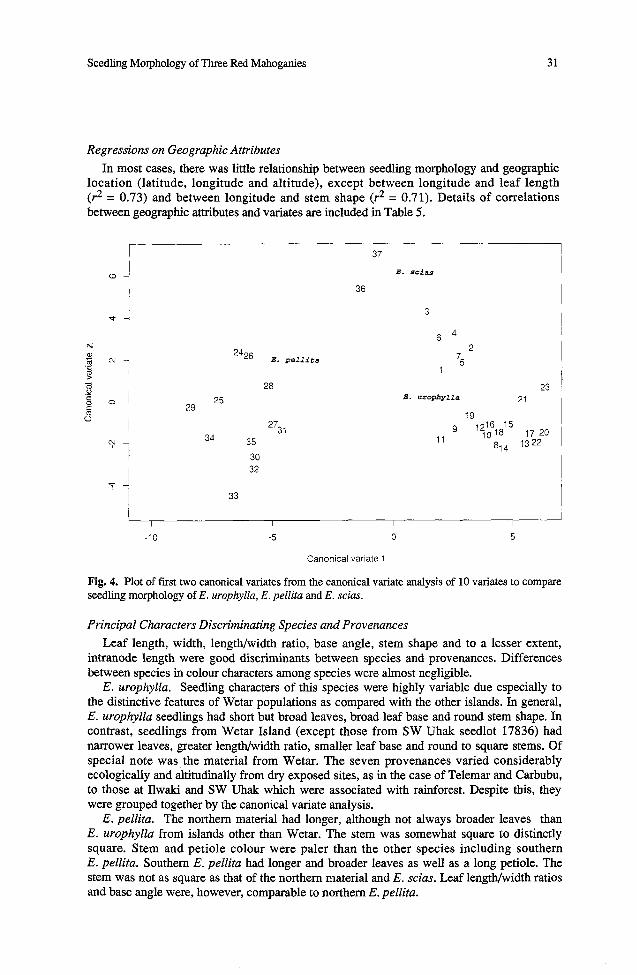
The results of the multivariate analysis are shown in Fig. 4 where the first canonical variate is plotted against the second. The first accounted for 70% of the variance, the second for only 16%. The figure shows four groupings, two of which belong to E. urophylla, the other two belong to E. pellita and E. scias. For E. urophylla the provenances from Alor, Adonara, Flores, Pantar and Timor cluster into one group with the provenances from Wetar splitting off on their own. The two provenances of E. scias separate out as a distinct group, as do those of E. pellita. There is a suggestion of a separation between the northern and southern populations of E. pellita; those from the north had shorter and narrower leaves and were more distinctly square stemmed than those from the south. A least significant distance based on Mahalanobis D~ at the 0.1% probability level is 5.6 units in Fig. 4.
The loadings (weighted according to the standard deviations from the individual variate analyses as given in Table 3) are shown in Table 6. A large absolute value for a variate loading indicates importance of the variate in the formation of the canonical variates. Thus size (length and width, variates 1 and 2) and shape of leaf (variate 4) are the most important variates for both the fisst and second canonical variates.
K. Piiyopusarerk et al.
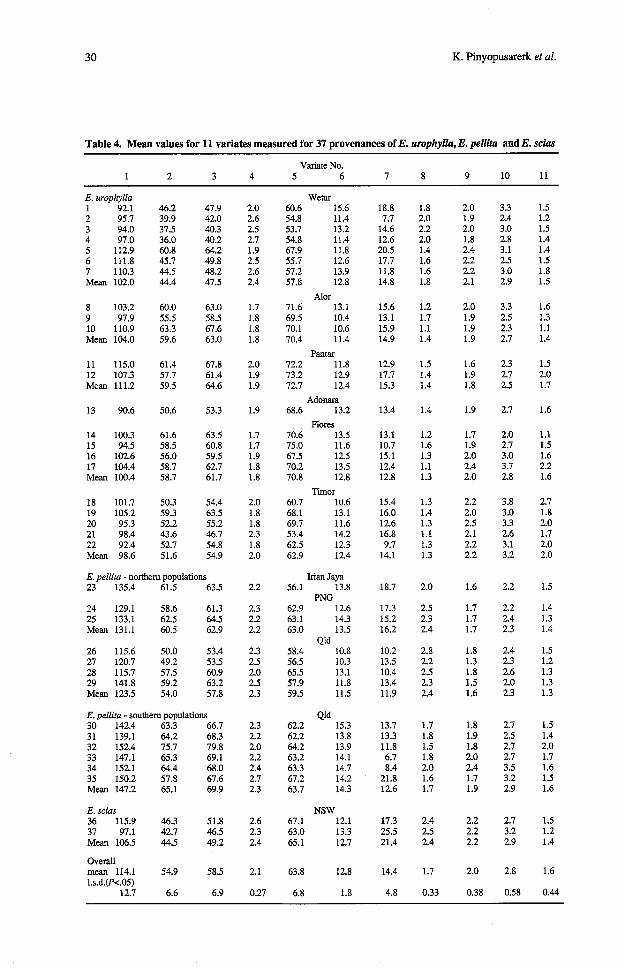
Table 4. Mean values for 11 variates measured for 37 provenances of E. urophylla, E. pellita and E. scias
Variate No. 1 2 3 4 5 6 7 8 9 10 11
E. urophylla 1 92.1 2 95.7 3 94.0 4 97.0 5 112.9 6 111.8 7 110.3 Mean 102.0
8 103.2 9 97.9 10 110.9 Mean 104.0
11 115.0 12 107.3 Mean 111.2
13 90.6
14 100.3 15 94.5 16 102.6 17 104.4 Mean 100.4
18 101.7 19 105.2 20 95.3 21 98.4 22 92.4 Mean 98.6
W e w 60.6 15.6 54.8 11.4 53.7 13.2 54.8 11.4 67.9 11.8 55.7 12.6 57.2 13.9 57.8 12.8
Alor 71.6 13.1 69.5 10.4 70.1 10.6 70.4 11.4
E. pellita - northern populations 23 135.4 61.5 63.5
Irian Jaya 56.1 13.8
PNG 62.9 12.6 63.1 14.3 63.0 13.5
Qld 58.4 10.8 56.5 10.3 65.5 13.1 57.9 11.8 59.5 11.5
24 129.1 58.6 61.3 25 133.1 62.5 64.5 Mean 131.1 60.5 62.9
26 115.6 50.0 53.4 27 120.7 49.2 53.5 28 115.7 57.5 60.9 29 141.8 59.2 63.2 Mean 123.5 54.0 57.8
E. pellita - southern populations 30 142.4 63.3 66.7 31 139.1 64.2 68.3 32 152.4 75.7 79.8 33 147.1 65.3 69.1 34 152.1 64.4 68.0 35 150.2 57.8 67.6 Mean 147.2 65.1 69.9
E. scias 36 115.9 46.3 51.8 37 97.1 42.7 46.5 Mean 106.5 44.5 49.2
NSW 67.1 12.1 63.0 13.3 65.1 12.7
Overall mean 114.1 54.9 58.5
Seedling Morphology of Three Red Mahoganies
Regressions on Geographic Attributes
In most cases, there was little relationship between seedling morphology and geographic location (latitude, longitude and altitude), except between longitude and leaf length (r2 = 0.73) and between longitude and stem shape (r2 = 0.71). Details of correlations between geographic attributes and variates are included in Table 5.
37
E . scias
36
3
Canonical variate 1
Fig. 4. Plot of first two canonical variates from the canonical variate analysis of 10 variates to compare seedling morphology of E. urophylla, E. pellita and E. scias.
Principal Characters Discriminating Species and Provenances
Leaf length, width, lengthlwidth ratio, base angle, stem shape and to a lesser extent, intranode length were good discriminants between species and provenances. Differences between species in colour characters among species were almost negligible.
E. urophylla. Seedling characters of this species were highly variable due especially to the distinctive features of Wetar populations as compared with the other islands. In general, E. urophylla seedlings had short but broad leaves, broad leaf base and round stem shape. In contrast, seedlings from Wetar Island (except those from SW Uhak seedlot 17836) had narrower leaves, greater lengtwwidth ratio, smaller leaf base and round to square stems. Of special note was the material from Wetar. The seven provenances varied considerably ecologically and altitudinally from dry exposed sites, as in the case of Telemar and Carbubu, to those at Ilwaki and SW Uhak which were associated with rainforest. Despite this, they were grouped together by the canonical variate analysis.
E. pellita. The northern material had longer, although not always broader leaves than E. urophylla from islands other than Wetar. The stem was somewhat square to distinctly square. Stem and petiole colour were paler than the other species including southern E. pellita. Southern E. pellita had longer and broader leaves as well as a long petiole. The stem was not as square as that of the northern material and E. scias. Leaf lengthlwidth ratios and base angle were, however, comparable to northern E. pellita.
Tab
le 5
. C
orre
latio
n m
atri
x fo
r 11
seed
ling
vari
ates
and
3 g
eogr
aphi
c att
ribu
tes
Lat
itude
0.
299
-0.0
16
0.02
6 0.
325
-0.0
13
0.12
2 0.
215
0.47
4 0.
158
0.05
0 -0
.161
1.
000
Lon
gitu
de
0.73
0 0.
235
0.25
6 0.
479
-0.2
87
0.19
3 0.
028
1.70
8 -0
.188
-0
.174
.-
0.17
2 0.
739
1.00
0
Alti
tude
-0
.380
-0
.122
-0
.127
-0
.320
0.
039
-0.0
13
0.00
4 -0
.589
0.
248
0.25
3 0.
410
-0.2
25
-0.4
01
1.00
0
1 2
3
4 5
6 7
8 9
10
11
lat
long
al
t
Seedling Morphology of Three Red Mahoganies
E. scias. Leaves were generally shorter and narrower than the other species. Another notable feature was the distinctly square stem. E. scias and E. urophylla from Wetar are similar in leaf size and shape, and are closest to each other according to the canonical variate analysis (Fig. 4).
There appeared to be no clear pattern of variation in intranode length between leaves at the 9th node although differences between species and provenances existed. Among provenances of E. urophylla the shortest intranode length (7.7 mm) and the longest (20.5 mm) were from Wetar. Intranode length of southern E. pellita ranged from 6.7 to 21.8 mm. Queensland populations of northern E. pellita had shorter intranodes (average 11.9 mm) thaii those from Maii Jaya (18.7 rim) and P a p a New Guinea ji6.2 mmj. Both E. scias provenances had long intranodes (17.3 and 25.5 mm).
Table 6. Canonical variate loadings weighted according to the standard deviations of original variates Variate 3 (leaf widest width) is excluded
Discussion
Variate No. Loadings cv 1 CV2
In this study, differentiation was evident between some species in the subseries Resiniferinae (E. urophylla, northern and southern E. pellita, and E. scias) as well as between provenances within E. urophylla. Leaf length, width, lengthfwidth ratio, base angle, stem shape and to a lesser extent intranode length provided good discriminating characters for comparing species and provenances.
The Wetar provenances are distinct from the rest of the E. urophylla provenances, having narrower leaves and greater lengthlwidth ratios. Other features which differentiated the provenances from Wetar from the five other islands were the small leaf angle and square stem. The Wetar populations represented by seedlots 17832,' 17834, 17835 and 17837 of E. urophylla are therefore anomalous in relation to the array of populations of the species as a whole. The harsh sites, with shallow rocky soils on steep slopes, on the island and perhaps climatic stresses (Martin and Cossalter 1976) due to its location at the easternmost extension of the range of the species suggest that the narrow leaves are an adaptation to this area.
On the basis of seedling morphology, E. pellita appears to be less variable than E. urophylla. However, there was a trend, as shown in the canonical variate analysis, towards a separation between the northern and southern populations; those from the north had shorter and narrower leaves and more distinctly square stems than those from the south.
Variation in seedling morphology in eucalypts has been shown in many cases to be correlated with geographical origins of the seeds (Burley et al. 1971; Turnbull 1979).

K. Pinyopusarerk et al.
Martin and Cossalter (1976), in an analysis of the classical determinants of weather patterns in these latitudes and with these geographical dispositions, mentioned a gradient of increasing aridity from west to east in the Lesser Sunda islands brought about by the influence of the high mountains of Timor. The correlation of seedling morphology at least to some extent with longitude found in this study would suggest a selective environmental pressure through increasing aridity. However, eastern Wetar is at odds with this trend and seems to have moister conditions than those of the western part of that island Until meteorological data are available for Wetar, any attempt to correlate the Wetar form of E. urophylla with some unique climatic condition of the island must remain conjectural. Further investigation of these populations using isozyme techniques may reveai their intemiationships witn more precision.
Acknowledgments This study was financially supported by AIDAB under Seeds of Australian Trees Project
and the Australian Centre for International Agricultural Research. David Lea, Maurie Mill and Jorg Dippel provided technical assistance during the glasshouse experiment. We thank Doug Boland, John Doran, Chris Harwood, Jock Morse and John Turnbull for their valuable comments on the manuscript and Alan House for preparing Figs 1,2 and 3.
References Boland, D.J. (1978). Geographic variation in Eucalyptus leucoxylon F. Muell. Australian Forest
Research 8,25-46. Boland, DJ., Brooker, M.I.H., Chippendale, G.M., Hall, N., Hyland, B.P.M., Johnston, R.D., Kleinig, D.A.,
and Turner, J.D. (1984). 'Forest Trees of Australia.' (Nelson / CSIRO: Melbourne.) Burley, J., Wood, P.J., and Hans, A.S. (1971). Variation in leaf characteristics among provenances of
Eucalyptus camaldulensis Dehn. grown in Zambia. Australian Journal of Botany 19,239-49. Chappill, J., Ladiges, P.Y., and Boland, DJ. (1986). Eucalyptus aromaphloia Pryor & W i s - a redefmition
of geographical and morphological boundaries. Australian J o w l of Botany, 34,395-412. Charomaini, M. (1990). Geographic variation of juvenile leaf morphological characteristics of
Eucalyptus urophylla from Eastern Nusa Tenggara islands grown in south Sumatra. Indonesian Forest Research Bulletin 530, 1-10.
Cook, I.O., and Ladiges, P.Y. (1991). Morphological variation within Eucalyptus nitens s. lat. and recognition of a new species, E. denticulata. Australian Systematic Botany 4,375-90.
Jacobs, M.R. (1981). Eucalyptus for planting. FA0 Forestry Series No. 11. (FAO: Rome.) Johnson, L.A.S., and Hill, K.D. (1990). New taxa and combinations in Eucalyptus and Angophora
(Myrtaceae). Telopea 4,37-108. Ladiges, P.Y., Gray, A.M., and Brooker, M.I.H. (1981). Pattern of geographic variation, based on
seedling morphology, in Eucalyptus ovata Labill. and E. brookeriana A.M. Gray and comparisons with some other Eucalyptus species. Australian Journal of Botany 29,593-603.
Martin, B., and Cossalter, C. (1976). Les Eucalyptus des Iles de la Sonde. Bois et Forets des Tropiques 165,3-20.
Moura, V.P.G. (1977). Altitudinal variation in Eucalyptus urophylla S.T. Blake. M.Sc. Thesis, University of Melbourne.
Payne, R.W., Lane, P.W., Ainsley, A.E., Bicknell, K.E., Digby, P.G.N., Harding, S.A., Leech, P.K., Simpson, H.R., Todd, A.D., Verrier, P.J., White, R.P., Gower, J.C., and Tunnicliffe Wilson, G. (1987). 'Genstat 5 Reference Manual.' (Oxford University Press: New York.)
Pryor, L.D., and Johnson, L.A.S. (1971). 'A Classification of the Eucalypts.' (Australian National University Press: Canberra.)
Turnbull, J.W. (1979). Geographic variation in E. cloeziana. Ph.D. Thesis, Australian National University.
Turnbull, J.W., and Brooker, I. (1978). Timor mountain gum - Eucalyptus urophylla S.T. Blake. Forest Tree Series No. 214. (CSIRO Division of Forest Research: Canberra.)
Williams, E.R., and John, J.A. (1989). Construction of row and column design with continuous replicates. Applied Statistics 38, 149-54.
Manuscript received 10 June 1992, accepted 19 October 1992
Copyright © 2022 FDOKUMEN




















