
Edible coatings based on Sour Prickly Pear mucilage (Opuntia ...

Harmful Algae 2 (2003) 301–316
Correlation between the presence ofGonyaulax fragilis(Dinophyceae) and the mucilage phenomena of the
Emilia-Romagna coast (northern Adriatic Sea)
Marinella Pompeia, Cristina Mazziottib, Franca Guerrinic, Monica Canginia,Silvia Pigozzia, Margherita Benzib, Simona Palamidesic,
Laurita Bonic, Rossella Pistocchic,∗a Centro Ricerche Marine, Viale Vespucci 2, 47042 Cesenatico, FC, Italy
b Arpa, Struttura Oceanografica Daphne, Viale Vespucci 2, 47042 Cesenatico, FC, Italyc Centro Interdipartimentale di Ricerca per le Scienze Ambientali, Università di Bologna, Via Tombesi dall’Ova 55, 48100 Ravenna, Italy
Received 27 February 2003; received in revised form 3 April 2003; accepted 13 June 2003
Abstract
The formation of massive amounts of suspended mucilaginous organic matter which periodically affects the Adriatic Sea,has been regarded as a complex physico-chemical phenomenon resulting from the production of extracellular material byphytoplankton. Although the exact cause has remained obscure, the mechanism of its formation has usually been consideredto be a long-term process, starting after the late winter-early spring blooms, and involving the participation of various algalspecies, mainly within the diatom group. In this paper we report on the results of a phytoplankton monitoring programme innorthern Adriatic seawaters off the Emilia-Romagna coast of Italy which revealed the constant concomitant presence of thedinoflagellateGonyaulax fragilis (Schütt) Kofoid and mucilaginous formations. In the early stages of the phenomenon thedinoflagellate was clearly observable by microscopic examination in the mucilage, but as the mucilage aged this alga almostcompletely decomposed and diatom cells increased in number and became predominant. Although characterized by a slowgrowth rate in culture, in natural seawaterG. fragilis was observed to reach cell densities of up to 7.0 × 106 cells l−1. Theresults of this study lead us to propose the hypothesis that the appearance of mucilage in the water column of the Adriatic Seais the consequence of a seasonal growth of this dinoflagellate favoured by specific environmental circumstances.© 2003 Elsevier B.V. All rights reserved.
Keywords: Adriatic Sea; Dinoflagellates; Extracellular polysaccharides;Gonyaulax fragilis; Mucilage
1. Introduction
The northern Adriatic Sea is periodically affectedby the massive presence of organic matter, called
∗ Corresponding author. Tel.:+39-0544-213831;fax: +39-0544-31204.
E-mail address: [email protected] (R. Pistocchi).
mucilage, consisting of a white gelatinous materialinitially suspended in the water column, which ac-cumulates until it covers a large area of the watersurface. The first record of this phenomenon datesback to 1729, and episodes which occurred in the19th century and in the first-half of the 20th centurywere reported mainly by fishermen and naturalistswho provided descriptions and a classification of the
1568-9883/$ – see front matter © 2003 Elsevier B.V. All rights reserved.doi:10.1016/S1568-9883(03)00059-3

302 M. Pompei et al. / Harmful Algae 2 (2003) 301–316
phenomenon (Molin et al., 1992). After an intervalof about 40 years, large-scale events occurred in1988, 1989 and 1991, during which mucilage cov-ered hundreds of square kilometres of sea surface.Until the 1980–1990s, this phenomenon seemed tohave a long-term periodicity; recently, however, itsfrequency has increased, and occurred to varyingdegrees of intensity in 1997, 2000, 2001 and 2002(Regione Emilia-Romagna, 1997-2002). Both thehistorical and the more recent episodes of mucilageaccumulation were limited to the summer months,mainly between June and September (Giani et al.,1992; Molin et al., 1992). The northern Adriatic Seahas important fishery and tourist industries that areseverely affected in the years of mucilage formation.Therefore a considerable amount of concern and sci-entific efforts have been devoted to understandingthe phenomenon. Studies performed after the late1980s related the presence of the organic mucilagi-nous material to phytoplankton production withoutan exact identification of the organism responsiblefor its formation. It was assumed that several speciescontributed to mucilage formation, and that the char-acteristic features of the Adriatic basin, together withparticular weather conditions, were the most criticalfactors (Mingazzini and Thake, 1995; Degobbis et al.,1995, 1999). Mucilage was, in fact, present in yearswith prolonged periods of high atmospheric pressure,of calm and sunny weather, and with waters having astrong picnocline (Vollenweider et al., 1992; Rinaldiet al., 1995). The hypothesis was put forward thatsuch conditions contributed to the sedimentation ofgelatinous filaments produced by phytoplankton; thefilaments accumulated at the picnocline, and increasedin size due to collisions facilitated by currents andwinds (Giani et al., 1992). Thus, mucilage startedwith the formation of flocs (marine snow) duringthe spring, with a subsequent transition to the largerforms (Rinaldi et al., 1995), although it was believedthat some other unidentified triggering mechanismhad to be present in critical years (Degobbis et al.,1999).
The presence of small mucilaginous aggregates,known as marine snow, is very common in coastaland oceanic waters worldwide (Alldredge and Silver,1988), whereas the appearance of large amounts offoam, like those of the Adriatic Sea, is more limited.One of the most conspicuous and better characterized
events is the one occurring in the North Sea. The phe-nomenon has been clearly correlated to the abnormalgrowth ofPhaeocystis pouchetii (Primnesiophyta); inits colonial form, the cells are embedded in abundantpolysaccharidic material. The copious growth of thisalga has been attributed to anthropic eutrophication,while winds and currents are thought to contributeto the accumulation of the great amount of foamalong the shoreline (Lancelot, 1995). Accumulationof large amounts of marine slime also occurs periodi-cally in New Zealand waters, where these events havebeen documented since 1840. More recently theseevents have been attributed to the extracellular releaseof polysaccharidic substances by the dinoflagellateGonyaulax hyalina (Mackenzie et al., 2002).
Instead of being attributed to one causative or-ganism, mucilage in the Adriatic is considered to becaused by a variety of diatom species for the followingreasons: (1) these organisms constitute a large portionof phytoplankton biomass of the northern AdriaticSea, and exhibit extensive blooms, usually in earlyspring (Revelante et al., 1984; Pompei et al., 1995);(2) diatoms are known to produce and release largeamounts of polysaccharides, especially in the pres-ence of unbalanced N/P ratios (Myklestad and Haug,1972; Myklestad, 1977, 1995); (3) the microscopicobservation of mucilage has often revealed the pres-ence of several diatom species; of theseCylindrothecaclosterium appeared to be the most healthy and recur-rent (Rinaldi et al., 1995; Viviani et al., 1995a,b).
The presence, in fairly high numbers, of the di-noflagellateGonyaulax fragilis was reported in severalpapers relative to the mucilage events affecting theAdriatic Sea (Cabrini et al., 1992; Honsell et al., 1992;Viviani et al., 1995a,b), even though this organismwas not considered to be a producer of polysaccha-ridic exudates. Recently, however, a relevant role hasbeen attributed to the dinoflagellate group in marinesnow and mucilage production:Alldredge et al. (1998)observed the occurrence of a form of marine snowlargely dominated by thecate dinoflagellates whichwas unusually cohesive and mucus-rich, whilePassow(2002) showed that dinoflagellates could generateconsiderable amounts of TEP (transparent exopolymerparticles). This is considered to be an important step inmucilage formation (Alldredge, 1999), and was previ-ously studied only in diatoms andPhaeocystis sp. Fi-nally, Mackenzie et al. (2002)attributed mucilage

M. Pompei et al. / Harmful Algae 2 (2003) 301–316 303
accumulation observed along the coasts of NewZealand to the polysaccharides produced byG.hyalina.
In this paper, we report the results of both mon-itoring activities and laboratory research performedduring the most recent episodes of mucilage accu-mulation in the Adriatic Sea. The hypothesis is putforward that the dinoflagellateG. fragilis could makea substantial contribution to the formation of thegelatinous organic material accumulated along theEmilia-Romagna coast.
2. Material and methods
2.1. Field sampling and observations
The sampling area, located in the north-westernAdriatic coastal waters, in front of the Emilia-Romagnaregion, has been monitored, since 1978, by means ofthe vessel “M.N. Daphne II” (Arpa, Emilia-Romagna)with a monitoring network of 33 stations (Fig. 1). Thestations are situated at different distances from thecoast, with bathymetries varying from 3 to 22.5 m.Sampling was performed at weekly intervals all yearround.
The physico-chemical characteristics (temperature,salinity, pH, dissolved oxygen) of the water columnwere measured using the “Idronaut” Ocean Sevenmodel 316 probe. Concentrations of chlorophyll awere determined in situ by a TURNER 10 AU fluo-rometer.
Water and mucilage samples were collected fromthe water column with Niskin bottles or directly byscuba divers; the mucilage was sampled from the sur-face using bottles or syringes. Qualitative and quanti-tative analysis of phytoplankton was performed by theUtermöhl (1931)method on water samples stored indark glass bottles.
At a few stations, observations were performed us-ing a specially protected “Telesub Lanterna” remotelycontrolled underwater videocamera; the images weretransmitted to an on-board monitor and video-recorderin order to visualize the distribution, shape and den-sity of the aggregates at different levels of the watercolumn.
Data were represented graphically using the Mi-crosoft Excel and Surfer Software package.
2.2. Phytoplankton cultures
Gonyaulax fragilis (Schütt) Kofoid, Lingulo-dinium polyedrum Stein (Dodge), Prorocentrummicans Ehrenberg andProtoceratium reticulatum(Chlaparède and Lachmann) Buetschli were iso-lated from natural phytoplankton associations incoastal waters of north-western Adriatic Sea, Emilia-Romagna, Italy. Strains were maintained in sterilef/10 medium at 20◦C under a 16:8 h light–dark periodat about 150�mol m−2 s−1 from cool white lamps(McLachlan, 1973). For the experimental work, thecultures were transferred into sterile Erlenmeyer flaskssealed with cotton plugs and containing the mediumpreviously tested for optimal algal growth; this wasf/2 (Guillard and Ryther, 1962) for L. polyedrum andP. micans, f/2 with a five-fold lowered P and N con-tent for P. reticulatum and medium K (Keller et al.,1985) for G. fragilis. All cultures were grown underthe temperature and the light conditions describedabove, except those ofG. fragilis which were usuallykept at a light intensity of 65�mol photons m−2 s−1.Cell counts were made in settling chambers by theUtermöhl (1931)method.
2.3. Polysaccharide analysis
Extracellular carbohydrates were analysed, accord-ing to Myklestad et al. (1997), on 1 ml of supernatantobtained after centrifuging 5 ml cultures at 8260 g for10 min in a Beckman J2-HS centrifuge (rotor JS-13.1).Carbohydrate concentrations are expressed as glucoseequivalents.
For each culture condition there were two replicateflasks and each experiment was repeated two to threetimes.
2.4. Alcian Blue staining
Cultured and fieldG. fragilis samples were stainedwith Alcian Blue, a specific stain for acidic polysac-charides, added as 0.1% solution (w/v) in 0.5 N aceticacid according toCrayton (1982).
2.5. Biovolume measurement
Cell volume of cultured strains was determinedaccording to Edler (1979). Mean values obtained
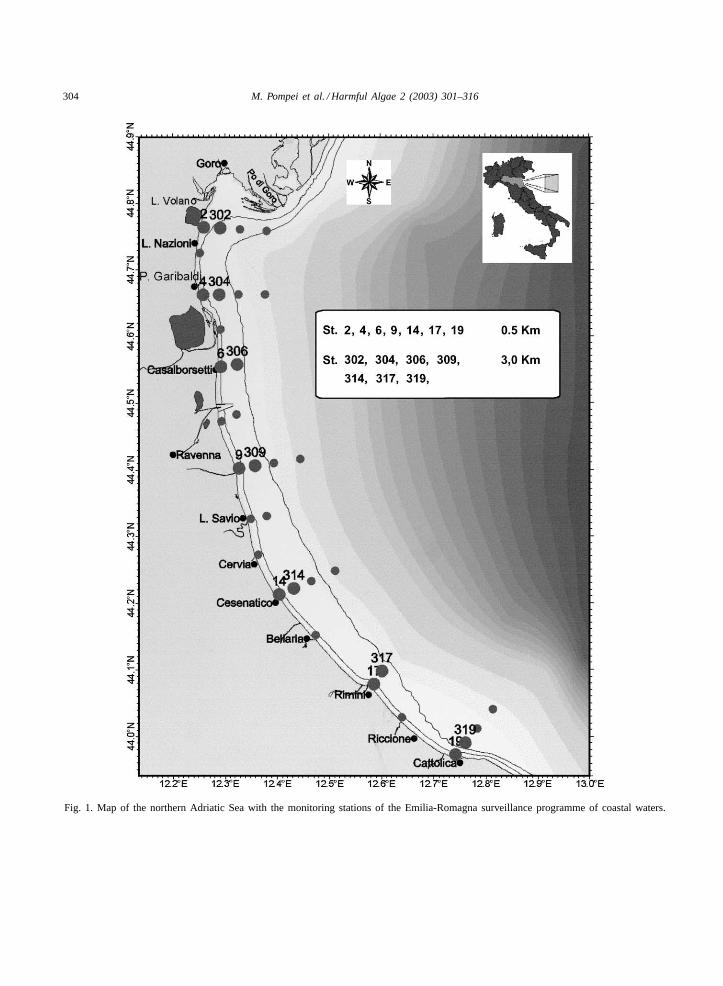
304 M. Pompei et al. / Harmful Algae 2 (2003) 301–316
Fig. 1. Map of the northern Adriatic Sea with the monitoring stations of the Emilia-Romagna surveillance programme of coastal waters.

M. Pompei et al. / Harmful Algae 2 (2003) 301–316 305
by measuring 20–40 cells were:L. polyedrum15,300�m3, P. micans 16,070�m3, P. reticulatum8,280�m3, G. fragilis 12,290�m3.
3. Results
3.1. Mucilage appearance from 2000 to 2002 andGonyaulax fragilis presence
Since 1988, based on the phytoplankton monitoringactivity, a positive correlation between the presence ofthe dinoflagellateG. fragilis and that of mucilage hasbeen observed in the Adriatic Sea waters along theEmilia-Romagna coast. InTable 1, the years duringwhich the gelatinous formations were clearly observ-able and those in whichG. fragilis was present in thewater column, are reported.
Considering the most recent period, mucilage wasalways present during the summertime of the last 3years. The organic material observed during these pe-riods had the same appearance of that described in the
Table 1Years and periods during which the mucilage was observed in theAdriatic Sea and presence or absence ofG. fragilis in the watersof the Emilia-Romagna coast in the same periods
Years ofmonitoring
Presence of mucilage Presence ofG. fragilis
1988 From August to the endof September
Present
1989 From the end of June tothe beginning of August
Present
1990 Absent Absent1991 From the end of June to
the end of AugustPresent
1992 Absent Absent1993 Absent Absent1994 Absent Absent1995 Absent Absent1996 Absent Absent1997 August and September Present1998 Absent Absent1999 Absent Absent2000 From the beginning of
June to the beginning ofJuly
Present
2001 From the end of June tothe middle of July
Present
2002 From the end of June tothe beginning ofSeptember
Present
years 1988, 1989 and 1991, with formations such asflocs and macroflocs, stringers (2–3 m long), clouds(4–5 m diameter) and gelatinous layers; it was whiteand sticky, while at later stages it became brown andsparse. The floating material came to the surface inthe warmest hours of the day while it stayed in sub-surface layers during cooler periods, such as the earlymorning or the late afternoon, presumably due to gasbubbles coming from the intense metabolic activityinside the mucus.
In 2000 the first mucilage appeared offshore atthe beginning of June. The phenomenon successivelyincreased and was localized at the thermocline; atthe beginning of July the foam disappeared as itwas dispersed by sea storms. In 2001 the mucilagestarted again offshore, towards the end of June, andthe aggregated material was observed above the ther-mocline. The foam covered different zones alongthe Emilia-Romagna coast and was present until themiddle of July when the weather conditions favouredits dispersion again. In 2002 the event started in thesecond-half of June and lasted until the first days ofSeptember; the sea area covered by the gel was par-ticularly wide so that the phenomenon was observedfrom the north Adriatic to its southern part (TremitiIslands). During that summer several sea storms wereregistered but the gelatinous formations persisted.
The weekly monitoring activity performed at 14stations along the Emilia-Romagna coast permitted aqualitative and quantitative analysis of phytoplanktonin the mentioned 3 years. InTables 2–4the periodsin which G. fragilis was observed and its cell numberper litre are reported; these observations refer to sam-pling sites located at 0.5 and 3 km from the seashore(Fig. 1) and to surface water samples (0.5 m). It isworth noting that the presence ofG. fragilis, even atnot extremely high numbers, was usually followed bythe first mucilaginous aggregation appearance nearly20 days afterwards and the increase in algal cell num-bers correlated well with the period during which themucilage was present in this part of the Adriatic coast(evidenced in italics in tables). A similar trend wasobserved also in 1997 although no data are reportedas the monitoring activity related toG. fragilis in thatyear was not so detailed.
As Tables 2–4show, the increase inG. fragilis cellnumbers usually started in June or July; the perma-nence of this species was highly dependent on weather
306 M. Pompei et al. / Harmful Algae 2 (2003) 301–316
Table 2G. fragilis cell number per litre in 14 sampling stations of the Emilia-Romagna coast weekly monitored in the year 2000
Cell number per litre
Station May June July August September October
29 9 13 19 27 3 10 17 3 8 14 30 7 13 18 6
2 120 1360 100 1280 40302 40 43191 120
4 120 1240 1440 640 80304 17148 112115
6 80 480 720 160 40306 40 5816
9 80 160 13086 40 40 80 80309 640 4014 400 440 80
314 240 1240 120 240 8017 240 280 480
317 40 44019 1000 2908 40 40
319 3120 21879 120
Only the days when the alga was present in seawater are reported and the periods during which mucilaginous formations were observedare evidenced in italics. Sampling sites are shown in the map ofFig. 1, sampling depth was 0.5 m.
conditions in that its dispersion was often caused bysea storms. However, in the years 2001 and 2002 par-ticularly high temperatures were registered in autumnand this favouredG. fragilis growth and, unusually,some aggregate formations were observed also in thatperiod.
Table 3G. fragilis cell number per litre in 14 sampling stations of the Emilia-Romagna coast weekly monitored in the year 2001
Cell number per litre
Station May June July August September October November December
2 22 14 20 3 10 17 1 9 20 27 29 2 10 15 23 30 5 30 3
2302 200 40
4 120 120 120 80 40 40304 2908 40 40 120 80
6 80 1320 3040 240 80306
9 360 880 520 80 520 320 120309 120 1120 40 80 8014 120 240 40 280
314 200 760 80 480 240 10017 40 640 640 40 40 1520 720 40
317 8019 40 80 80 80 400 440 40
319 400 160 240
Only the days when the alga was present in seawater are reported and the periods during which mucilaginous formations were observedare evidenced in italics.
3.2. G. fragilis growth in the environment
In the years during which the mucilage was notobserved,G. fragilis was never observed in seawatersampled for the monitoring activity. In the summerscharacterized by the presence of gelatinous masses, an
M. Pompei et al. / Harmful Algae 2 (2003) 301–316 307
Table 4G. fragilis cell number per litre in 14 sampling stations of the Emilia-Romagna coast weekly monitored in the year 2002
Cell number per litre
Station May June July August September October November
6 24 3 16 22 30 5 13 27 4 18 3 7 29 5
2 40 900302 8724 200
4 2908 11632 10902 100 280 600304 1454 14536 200 40
6 80 40 10178 13086 14540 900 100 600 100 80 120306
9 7270 2908 40712 14536 1000 300 1700 240309 2908 7400 1500 10014 40 23264 7268 6400 400 240
314 880 7400 400 200 10017 5816 47982 10902 4600 600 80
31719 40 68592 56706 10902 1817 1100 1900 80
319 320 440 1200 2000 300
Only the days when the alga was present in seawater are reported and the periods during which mucilaginous formations were observedare evidenced in italics.
increase inG. fragilis number was usually correlatedwith an increase in seawater temperature and the val-ues which favoured this increase were those equal orabove 20◦C. G. fragilis cells were first observed off-shore at a distance of 10–20 km from the coast and thealgae were most abundant at a depth corresponding to
Fig. 2. A mucilage sample with several brokenG. fragilis cells: scale bar 10�m.
the thermocline. When the algae reached a more ac-tive growth they could be observed also in surface andcoastal waters.
The highest numbers of liveG. fragilis cells werenot usually present within the mucilage, which wasmost often populated by several diatom species. One
308 M. Pompei et al. / Harmful Algae 2 (2003) 301–316
Fig. 3. G. fragilis cells: scale bar 10�m.
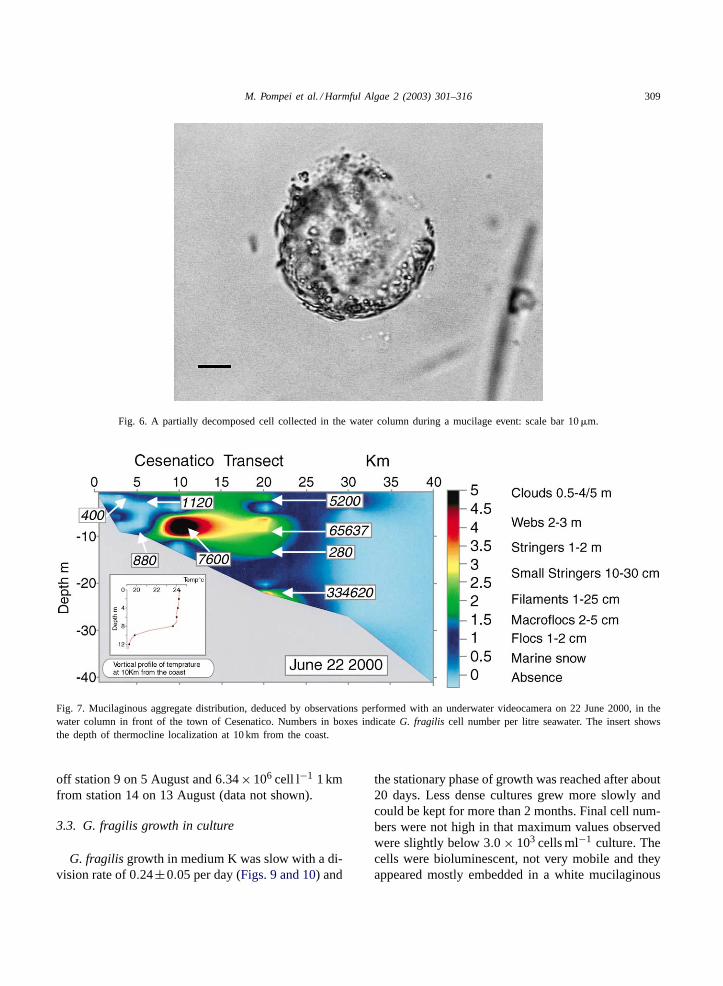
of the more recurrent wasCylindrotheca closterium,although also high numbers of different species(i.e. Asterionellopsis glacialis, Cerataulina pelagica,Pseudonitzschia spp.) could be found. In several mu-cilage episodes since 1988 mucilage contained a highnumber of deadG. fragilis organisms (Fig. 2), on thecontrary, at various depths along the water columneither healthyG. fragilis cells (Figs. 3 and 4), cellshaving a small cloud of material escaping from theirapical part (Fig. 5) or partially decomposed organisms(Fig. 6) were present.
During the monitoring activities several observa-tions, performed by means of an underwater video-
Fig. 4. G. fragilis cells suspended in the water column embeddedin mucilaginous material: scale bar 100�m.
Fig. 5. An apically brokenG. fragilis cell found in a field sampleand stained with Alcian Blue: scale bar 10�m.
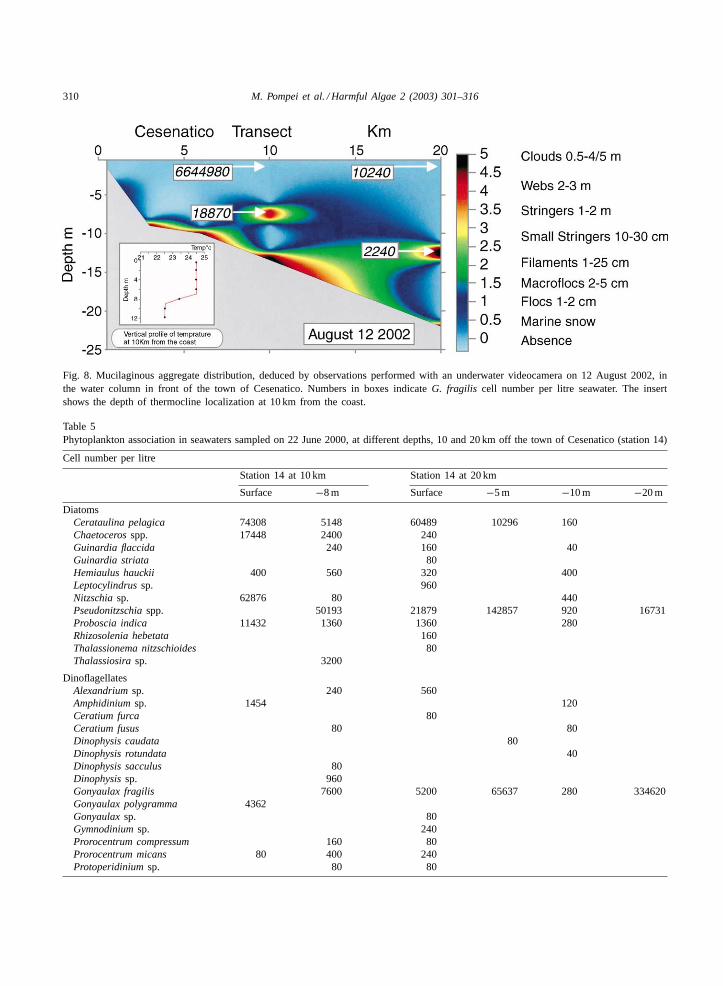
camera, permitted the drawing of graphic representa-tions of the aggregate distribution. InFigs. 7 and 8two representative graphs, relative to one event of theyear 2000 (22 June) and one of 2002 (12 August), areshown. The data were recorded in a transect in frontof Cesenatico, monitored from the coast to 20 km dis-tance and the distribution of the aggregates togetherwith the amount ofG. fragilis in the water column (ascell number per litre) is reported. Most of the biggestmucilaginous formations could be found either at thethermocline or in the bottom, while the highestG.fragilis numbers were not always localized at the samelevel. InTables 5 and 6the whole phytoplankton com-position of the water sampled at 10 and 20 km in thesame days ofFigs. 7 and 8, is reported. Dinoflagel-late numbers were low except forG. fragilis whichwas the dominant species; various diatoms were alsoabundant but the predominant species was different indifferent samplings: the large mucilaginous aggregateslocalized at the thermocline at 10 km (Figs. 7 and 8)presented an enrichment ofPseudonitzschia spp. in2000 and ofCerataulina pelagica andCylindrothecaclosterium in 2002 (Tables 5 and 6).
As shown inFigs. 7 and 8, G. fragilis could bepresent with high numbers also in surface waters; thevalue of about 6 millions cell l−1 registered on 12 Au-gust 2002, and corresponding to a chlorophyll concen-tration of 160�g l−1, was not an exception in that inthe same year were counted: 3.04×106 cell l−1 20 kmoff station 14 on 31 July, 6.70 × 106 cell l−1 15 km
M. Pompei et al. / Harmful Algae 2 (2003) 301–316 309
Fig. 6. A partially decomposed cell collected in the water column during a mucilage event: scale bar 10�m.
Fig. 7. Mucilaginous aggregate distribution, deduced by observations performed with an underwater videocamera on 22 June 2000, in thewater column in front of the town of Cesenatico. Numbers in boxes indicateG. fragilis cell number per litre seawater. The insert showsthe depth of thermocline localization at 10 km from the coast.
off station 9 on 5 August and 6.34× 106 cell l−1 1 kmfrom station 14 on 13 August (data not shown).
3.3. G. fragilis growth in culture
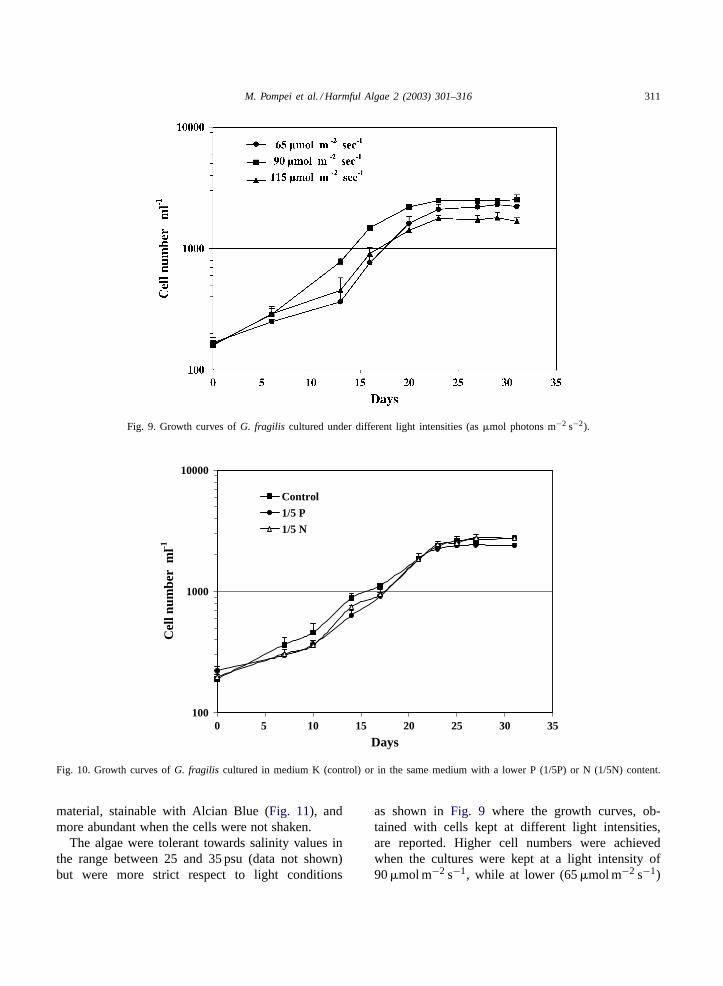
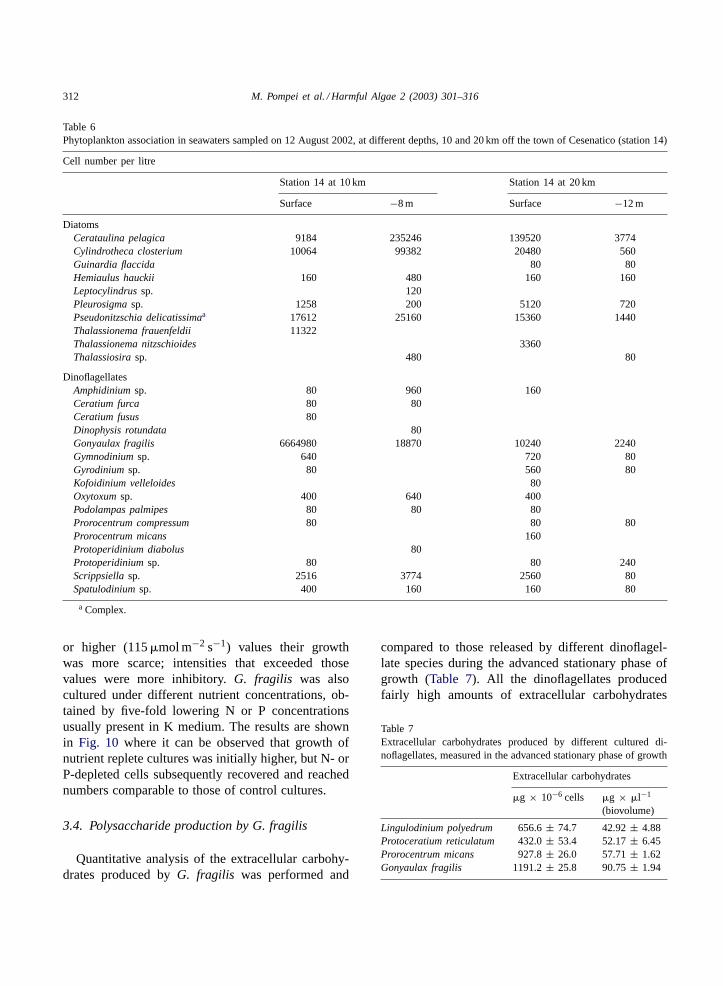
G. fragilis growth in medium K was slow with a di-vision rate of 0.24±0.05 per day (Figs. 9 and 10) and
the stationary phase of growth was reached after about20 days. Less dense cultures grew more slowly andcould be kept for more than 2 months. Final cell num-bers were not high in that maximum values observedwere slightly below 3.0 × 103 cells ml−1 culture. Thecells were bioluminescent, not very mobile and theyappeared mostly embedded in a white mucilaginous
310 M. Pompei et al. / Harmful Algae 2 (2003) 301–316
Fig. 8. Mucilaginous aggregate distribution, deduced by observations performed with an underwater videocamera on 12 August 2002, inthe water column in front of the town of Cesenatico. Numbers in boxes indicateG. fragilis cell number per litre seawater. The insertshows the depth of thermocline localization at 10 km from the coast.
Table 5Phytoplankton association in seawaters sampled on 22 June 2000, at different depths, 10 and 20 km off the town of Cesenatico (station 14)
Cell number per litre
Station 14 at 10 km Station 14 at 20 km
Surface −8 m Surface −5 m −10 m −20 m
DiatomsCerataulina pelagica 74308 5148 60489 10296 160Chaetoceros spp. 17448 2400 240Guinardia flaccida 240 160 40Guinardia striata 80Hemiaulus hauckii 400 560 320 400Leptocylindrus sp. 960Nitzschia sp. 62876 80 440Pseudonitzschia spp. 50193 21879 142857 920 16731Proboscia indica 11432 1360 1360 280Rhizosolenia hebetata 160Thalassionema nitzschioides 80Thalassiosira sp. 3200
DinoflagellatesAlexandrium sp. 240 560Amphidinium sp. 1454 120Ceratium furca 80Ceratium fusus 80 80Dinophysis caudata 80Dinophysis rotundata 40Dinophysis sacculus 80Dinophysis sp. 960Gonyaulax fragilis 7600 5200 65637 280 334620Gonyaulax polygramma 4362Gonyaulax sp. 80Gymnodinium sp. 240Prorocentrum compressum 160 80Prorocentrum micans 80 400 240Protoperidinium sp. 80 80
M. Pompei et al. / Harmful Algae 2 (2003) 301–316 311
Fig. 9. Growth curves ofG. fragilis cultured under different light intensities (as�mol photons m−2 s−2).
100
1000
10000
0 5 10 15 20 25 30 35
Days
Cel
l num
ber
ml-1
Control
1/5 P
1/5 N
Fig. 10. Growth curves ofG. fragilis cultured in medium K (control) or in the same medium with a lower P (1/5P) or N (1/5N) content.
material, stainable with Alcian Blue (Fig. 11), andmore abundant when the cells were not shaken.
The algae were tolerant towards salinity values inthe range between 25 and 35 psu (data not shown)but were more strict respect to light conditions
as shown inFig. 9 where the growth curves, ob-tained with cells kept at different light intensities,are reported. Higher cell numbers were achievedwhen the cultures were kept at a light intensity of90�mol m−2 s−1, while at lower (65�mol m−2 s−1)
312 M. Pompei et al. / Harmful Algae 2 (2003) 301–316
Table 6Phytoplankton association in seawaters sampled on 12 August 2002, at different depths, 10 and 20 km off the town of Cesenatico (station 14)
Cell number per litre
Station 14 at 10 km Station 14 at 20 km
Surface −8 m Surface −12 m
DiatomsCerataulina pelagica 9184 235246 139520 3774Cylindrotheca closterium 10064 99382 20480 560Guinardia flaccida 80 80Hemiaulus hauckii 160 480 160 160Leptocylindrus sp. 120Pleurosigma sp. 1258 200 5120 720Pseudonitzschia delicatissimaa 17612 25160 15360 1440Thalassionema frauenfeldii 11322Thalassionema nitzschioides 3360Thalassiosira sp. 480 80
DinoflagellatesAmphidinium sp. 80 960 160Ceratium furca 80 80Ceratium fusus 80Dinophysis rotundata 80Gonyaulax fragilis 6664980 18870 10240 2240Gymnodinium sp. 640 720 80Gyrodinium sp. 80 560 80Kofoidinium velleloides 80Oxytoxum sp. 400 640 400Podolampas palmipes 80 80 80Prorocentrum compressum 80 80 80Prorocentrum micans 160Protoperidinium diabolus 80Protoperidinium sp. 80 80 240Scrippsiella sp. 2516 3774 2560 80Spatulodinium sp. 400 160 160 80
a Complex.
or higher (115�mol m−2 s−1) values their growthwas more scarce; intensities that exceeded thosevalues were more inhibitory.G. fragilis was alsocultured under different nutrient concentrations, ob-tained by five-fold lowering N or P concentrationsusually present in K medium. The results are shownin Fig. 10 where it can be observed that growth ofnutrient replete cultures was initially higher, but N- orP-depleted cells subsequently recovered and reachednumbers comparable to those of control cultures.
3.4. Polysaccharide production by G. fragilis
Quantitative analysis of the extracellular carbohy-drates produced byG. fragilis was performed and
compared to those released by different dinoflagel-late species during the advanced stationary phase ofgrowth (Table 7). All the dinoflagellates producedfairly high amounts of extracellular carbohydrates
Table 7Extracellular carbohydrates produced by different cultured di-noflagellates, measured in the advanced stationary phase of growth
Extracellular carbohydrates
�g × 10−6 cells �g × �l−1
(biovolume)
Lingulodinium polyedrum 656.6± 74.7 42.92± 4.88Protoceratium reticulatum 432.0± 53.4 52.17± 6.45Prorocentrum micans 927.8± 26.0 57.71± 1.62Gonyaulax fragilis 1191.2± 25.8 90.75± 1.94
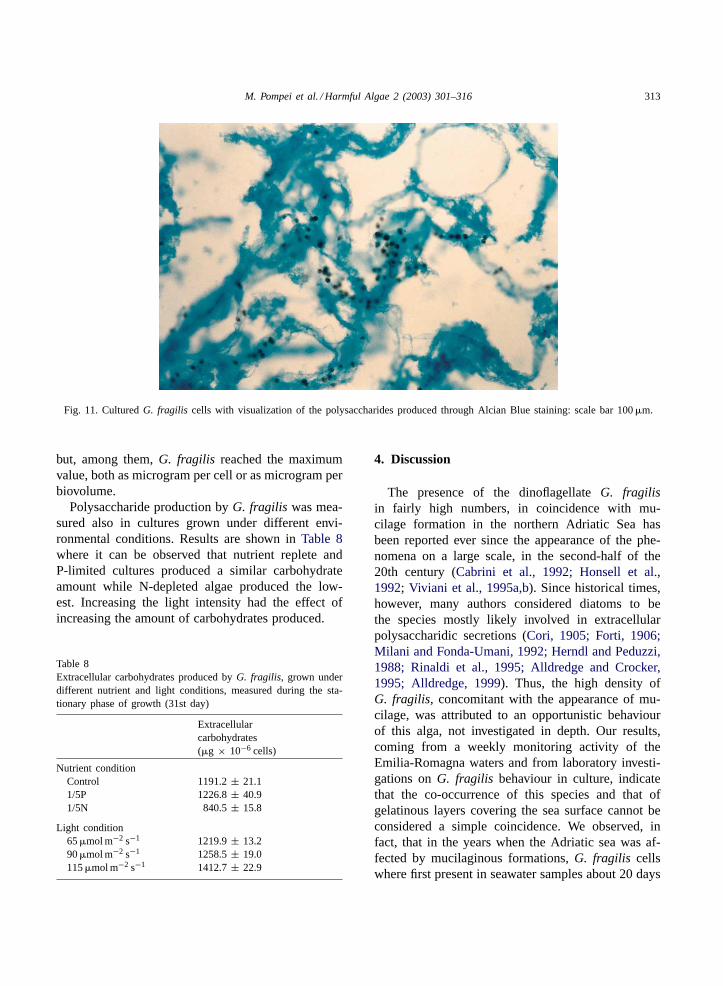
M. Pompei et al. / Harmful Algae 2 (2003) 301–316 313
Fig. 11. CulturedG. fragilis cells with visualization of the polysaccharides produced through Alcian Blue staining: scale bar 100�m.
but, among them,G. fragilis reached the maximumvalue, both as microgram per cell or as microgram perbiovolume.
Polysaccharide production byG. fragilis was mea-sured also in cultures grown under different envi-ronmental conditions. Results are shown inTable 8where it can be observed that nutrient replete andP-limited cultures produced a similar carbohydrateamount while N-depleted algae produced the low-est. Increasing the light intensity had the effect ofincreasing the amount of carbohydrates produced.
Table 8Extracellular carbohydrates produced byG. fragilis, grown underdifferent nutrient and light conditions, measured during the sta-tionary phase of growth (31st day)
Extracellularcarbohydrates(�g × 10−6 cells)
Nutrient conditionControl 1191.2± 21.11/5P 1226.8± 40.91/5N 840.5± 15.8
Light condition65�mol m−2 s−1 1219.9± 13.290�mol m−2 s−1 1258.5± 19.0115�mol m−2 s−1 1412.7± 22.9
4. Discussion
The presence of the dinoflagellateG. fragilisin fairly high numbers, in coincidence with mu-cilage formation in the northern Adriatic Sea hasbeen reported ever since the appearance of the phe-nomena on a large scale, in the second-half of the20th century (Cabrini et al., 1992; Honsell et al.,1992; Viviani et al., 1995a,b). Since historical times,however, many authors considered diatoms to bethe species mostly likely involved in extracellularpolysaccharidic secretions (Cori, 1905; Forti, 1906;Milani and Fonda-Umani, 1992; Herndl and Peduzzi,1988; Rinaldi et al., 1995; Alldredge and Crocker,1995; Alldredge, 1999). Thus, the high density ofG. fragilis, concomitant with the appearance of mu-cilage, was attributed to an opportunistic behaviourof this alga, not investigated in depth. Our results,coming from a weekly monitoring activity of theEmilia-Romagna waters and from laboratory investi-gations onG. fragilis behaviour in culture, indicatethat the co-occurrence of this species and that ofgelatinous layers covering the sea surface cannot beconsidered a simple coincidence. We observed, infact, that in the years when the Adriatic sea was af-fected by mucilaginous formations,G. fragilis cellswhere first present in seawater samples about 20 days

314 M. Pompei et al. / Harmful Algae 2 (2003) 301–316
before their appearance; algal number subsequentlyincreased, reaching as much as 6.0 × 106 cells l−1.
Microscopic observations of mucilage samplesprovided evidence thatG. fragilis was always presentwithin the polysaccharidic material, and that it wasmore abundant in the earliest stages of the phe-nomenon. When the gelatinous masses aged,G. frag-ilis decomposed and became unrecognizable, whilelarge numbers of a variety of diatom species cameto dominate the mucilage. In the Adriatic Sea mu-cilage of 1988 and 1999,Revelante and Gilmartin(1991) also found an enrichment of species, such asNitzschia longissima and Nitzschia closterium (nowCylindrotheca closterium), at higher levels than inthe surrounding seawater. They hypothesized that afew individuals of these species were embedded atearlier stages of aggregate formation, and then grewwell in the mucus environment. Similarly,Degobbiset al. (1999)reported that freshly formed aggregatescontained phytoplankton species similar to thosepresent in the surrounding water, while aged mucilagewas enriched especially inCylindrotheca closterium.Although often found in the water column, these di-atoms are, in fact, benthic species; they thrive verywell in the mucilage, which offers a false bottom anda high nutrient concentration. Thus, in aged mucilagesamples, in whichG. fragilis cells were completelydecomposed, only high numbers of diatoms couldbe observed, althoughG. fragilis was still present atdifferent levels of the water column.
These observations suggest that this species is oneof the causative organisms of the Adriatic Sea mu-cilage, an idea supported by recent findings pointingto an increasing role of dinoflagellates in producingmucus-rich marine snow (Alldredge et al., 1998), highamounts of TEP (Passow, 2002) and as being the causeof massive mucilaginous formation in New Zealand(Mackenzie et al., 2002). The latter phenomenon, inparticular, has very similar characteristics to those ofthe Adriatic Sea and it was attributed to the extra-cellular polysaccharide production of a dinoflagellate,Gonyaulax hyalina, a species very similar toG. frag-ilis and with which it can easily be confounded. Thespecies present in the Adriatic Sea, and collected dur-ing the 1989 event, was formerly calledG. hyalina(Cabrini et al., 1992); it was later identified asG. frag-ilis by SEM observations, although a clarification ofthe taxonomy of the two species, due to contradictions
between the description given by various authors, wasdeemed necessary (Honsell et al., 1992). Electronicmicroscope examinations of recently collected strainsled this author to confirm previous observations (Hon-sell, personal communication).
The results from cell culture analyses showed thatG. fragilis produced large amounts of extracellularcarbohydrates, which gave a white colouring and a mu-coid consistence to the cultures. Results also showedthat all the different dinoflagellate species tested re-leased a fair amount of carbohydrates, although lessthan that produced byG. fragilis. L. polyedrum has fre-quently been the cause of red tides in the Adriatic Sea,especially from 1976 to 1985 (Boni, 1983; Pompeiet al., 1995), but the water never assumed the gelati-nous aspect associated with the presence ofG. fragilis.In addition, all the dinoflagellate cultures exhibiteda typical brown colour, and the cells could be easilyseparated, except those ofG. fragilis. We comparedthe amount of carbohydrates produced byG. frag-ilis with those released by the diatomCylindrothecafusiformis (Guerrini et al., 1998; Pistocchi et al.,1997) and calculated that a few thousandG. fragiliscells release the same amount of carbohydrates as thatproduced by tens of millions ofC. closterium cells.Such a cell number is not very often reached by this orother diatom species, especially in the summertime.
It is our opinion that the mucilage phenomenon ofthe northern Adriatic Sea is mainly due to the pres-ence and growth ofG. fragilis. The frequent presencein the sea of only a few thousand cells per litre sug-gests that this amount could be sufficient to sustain thephenomenon. Similarly, during the New Zealand mu-cilage formation, only 5.7×103 cell l−1 seawater werecounted. It should be noted, however, that on manyoccasions cell counts performed in the Adriatic Seawaters could be underestimated due to the presence ofmany broken and unrecognizable cells as well as tothe discontinuous sampling which could have missedthe depth at whichG. fragilis is present.
Finally, cell numbers reached by culturedG. frag-ilis were not as high as those found in seawater dur-ing mucilage episodes. In fact, cell count in culturerarely exceeded 4.0×106 cell l−1 in contrast to nearly7.0 × 106 cell l−1 (cells in the sea had larger dimen-sions). This could be due to physical concentrationof the cells or to the fact that in its natural environ-mentG. fragilis growth was sustained by unidentified

M. Pompei et al. / Harmful Algae 2 (2003) 301–316 315
factors, such as an organic nutrient or micronutrient.The foam produced in culture was not as thick as theone observed on the sea surface, we showed, however,that the secretory activity ofG. fragilis could be stimu-lated by some environmental conditions. For example,carbohydrate production increased with increasinglight intensity. We cannot exclude that other factors,as yet unidentified, play a role in mucus aggregation,although it seems possible to discard nutrient depriva-tion as one such factor. It has always been postulatedthat the mucilage phenomenon of the Adriatic Sea wasprimarily due to the excretory activity of phytoplank-ton but that a complex array of environmental factorswere also important. The role of bacteria in the thick-ening and aggregation of the gel matrix was suggestedby Herndl et al. (1999)andAzam et al. (1999), andthe particular hydrological conditions of the AdriaticSea, lack of grazing pressure (Degobbis et al., 1999)and aggregation processes were also considered im-portant (McCave, 1984; Alldredge and Gotschalk,1990).
The observations and experiments reported in thispaper suggest that, although the previous hypothesescan be still considered valid, they do not only ap-ply to a long-term process, such as that of marinesnow formation as a consequence of early springphytoplankton blooms, but also to a short-term phe-nomenon represented by the summer growth of a di-noflagellate. By monitoring the phytoplankton in theAdriatic Sea coastal waters, an increase inG. frag-ilis cell numbers, even in coastal stations which arenot the preferred sites for its localization, could pro-vide an early warning of the possibility of mucilageformation.
Acknowledgements
This research work was funded by RegioneEmilia-Romagna and by Ministero dell’Istruzionedell’Università e della Ricerca (MIUR), Italy.
References
Alldredge, A.L., 1999. The potential role of particulate diatomexudates in forming nuisance mucilaginous scums. Ann. Ist.Super. Sanità. 35 (3), 397–400.
Alldredge, A.L., Silver, M.A., 1988. Characteristics, dynamics andsignificance of marine snow. Prog. Oceanogr. 20, 41–82.
Alldredge, A.L., Gotschalk, C., 1990. The relative contribution ofmarine snow of different origins to biological process in coastalwaters. Cont. Shelf Res. 10, 41–58.
Alldredge, A.L., Crocker, K.M., 1995. Why do sinking mucilageaggregates accumulate in the water column? Sci. Total Environ.165, 15–22.
Alldredge, A.L., Passow, U., Haddock, S.H.D., 1998. Thecharacteristics and transparent exopolymer (TEP) content ofmarine snow from thecate dinoflagellates. J. Plank. Res. 20 (3),393–406.
Azam, F., Fonda Umani, S., Funari, E., 1999. Significance ofbacteria in the mucilage phenomenon in the northern AdriaticSea. Ann. Ist. Super. Sanità. 35 (3), 411–419.
Boni, L., 1983. Red tides of the coast of the Emilia-Romagna(north-western Adriatic Sea) from 1975 to 1982. Inf. Bot. Ital.15, 18–24.
Cabrini, M., Fonda-Umani, S., Honsell, G., 1992. Mucilaginousaggregates in the Gulf of Trieste (northern Adriatic Sea):analysis of the phytoplanktonic communities in the periodJune–August 1989. In: Vollenweider, R.A., Marchetti, R.,Viviani, R. (Eds.), Marine Coastal Eutrophication. Elsevier,Amsterdam, pp. 557–568.
Cori, C.J., 1905. Sul fenomeno del “mare sporco” nel Golfo diTrieste durante l’estate del 1905. Oester. Fischerei Ztg. 1, 76–80.
Crayton, M.A., 1982. A comparative cytochemical study ofvolvocacean matrix polysaccharides. J. Phycol. 18, 336–344.
Degobbis, D., Fonda-Umani, S., Franco, P., Malej, A., Precali,R., Smodlaka, N., 1995. Changes in the northern Adriaticecosystem and the hypertrophic appearance of gelatinousaggregates. Sci. Total Environ. 165, 43–58.
Degobbis, D., Malej, A., Fonda-Umani, S., 1999. The mucilagephenomenon in the northern Adriatic Sea. A critical review ofthe present scientific hypoteses. Ann. Ist. Super. Sanità 35 (3),373–381.
Edler, L., 1979. Recommendations for Marine Biological Studiesin the Baltic Sea. Phytoplankton and Chlorophyll. BMBPublishers, pp. 1–38.
Forti, A., 1906. Alcune osservazioni sul “mare sporco” ed inparticolare sul fenomeno avvenuto nel 1905. Nuovo G. Botan.Ital. N. S. 13 (4), 81–132.
Giani, M., Cicero, A.M., Savelli, F., Bruno, M., Donati, G., Farina,A., Veschetti, E., Volterra, L., 1992. Marine snow in the AdriaticSea: a multifactorial study. In: Vollenweider, R.A., Marchetti,R., Viviani, R. (Eds.), Marine Coastal Eutrophication. Elsevier,Amsterdam, pp. 511–524.
Guerrini, F., Mazzotti, A., Boni, L., Pistocchi, R., 1998.Bacterial-algal interactions in polysaccharide production. Aquat.Microb. Ecol. 15, 247–253.
Guillard, R.L., Ryther, J.H., 1962. Studies on marine planktonicdiatoms. I.Cyclotella nana Hustedt andDetonula confervacea(Cleve) Gran. Can. J. Microbiol. 8, 229–239.
Herndl, G.J., Peduzzi, P., 1988. The ecology of amorphousaggregations (marine snow) in the northern Adriatic Sea. I.General considerations. Mar. Ecol. 9, 79–90.

316 M. Pompei et al. / Harmful Algae 2 (2003) 301–316
Herndl, G.J., Arrietta, J.M., Stoderegger, K., 1999. Interactionbetween specific hydrological and microbial activity leadingto extensive mucilage formation in the northern Adriatic Sea.Ann. Ist. Super. Sanità. 35 (3), 405–409.
Honsell, G., Cabrini, M., Darin, M., 1992. Gonyaulax fragilis(Schutt) Kofoid: a dinoflagellate from gelatinous aggregates ofnorthern Adriatic Sea. Giornale Botanico 126 (6), 749–751.
Keller, M.D., Selvin, R.C., Claus, W., Guillard, R.L., 1985. Factorssignificant to marine dinoflagellate culture. In: Anderson,D.M., White, A.W., Baden, D.G. (Eds.), Toxic Dinoflagellates.Elsevier, New York, pp. 73–77.
Lancelot, C., 1995. The mucilage phenomenon in the continentalcoastal waters of the North Sea. Sci. Total Environ. 165, 83–102.
Mackenzie, L., Sims, I., Beuzenberg, V., Gillespie, P., 2002. Massaccumulation of mucilage caused by dinoflagellate polysacc-haride exudates in Tasman Bay, New Zealand. Harmful Algae1, 69–83.
McCave, I.N., 1984. Size spectra and aggregation of suspendedparticles in the deep ocean. Deep-Sea Res. 31, 329–352.
McLachlan, J., 1973. Growth media-marine. In: Stein, J.R. (Ed.),Handbook of Phycological Methods. Culture Methods andGrowth Measurements. Cambridge University Press, New York,pp. 25–51.
Milani, L., Fonda-Umani, S., 1992. Mucilaginous agglomerationin the Gulf of Trieste (northern Adriatic Sea): analysis of themicrozooplankton population in the period June–August 1989.In: Vollenweider, R.A., Marchetti, R., Viviani, R. (Eds.), MarineCoastal Eutrophication. Elsevier, Amsterdam, pp. 569–580.
Mingazzini, M., Thake, B., 1995. Summary and conclusions ofthe workshop on marine mucilages in the Adriatic Sea andelsewhere. Sci. Total Environ. 165, 9–14.
Molin, D., Guidoboni, E., Lodovisi, A., 1992. Mucilage andthe phenomena of algae in the history of the Adriatic:periodicization and the anthropic context (17th–20th centuries).In: Vollenweider, R.A., Marchetti, R., Viviani, R. (Eds.), MarineCoastal Eutrophication. Elsevier, Amsterdam, pp. 511–524.
Myklestad, S.M., 1977. Production of carbohydrates by the marineplanktonic diatoms. II. Influence of the N:P ratio in the growthmedium on the assimilation ratio, growth rate, and production ofcellular and extracellular carbohydrates byChaetoceros affinisvar. Willei (Gran) Hustedt andSkeletonema costatum (Grev.)Cleve. J. Exp. Mar. Biol. Ecol. 29, 161–179.
Myklestad, S.M., 1995. Release of extracellular products byphytoplankton with special emphasis on polysaccharides. Sci.Total Environ. 165, 155–164.
Myklestad, S.M., Haug, A., 1972. Production of carbohydratesby the marine diatomChaetoceros affinis var. Willei (Gran)Hustedt. I. Effect of the concentration of nutrients in the culturemedium. J. Exp. Mar. Biol. Ecol. 9, 125–136.
Myklestad, S.M., Skånøy, E., Hestmann, S., 1997. A sensitiveand rapid method for analysis of dissolved mono- andpolysaccharides in seawater. Mar. Chem. 56, 279–286.
Passow, U., 2002. Production of transparent exopolymer particles(TEP) by phyto- and bacterioplankton. Mar. Ecol. Prog. Ser.236, 1–12.
Pistocchi, R., Guerrini, F., Balboni, V., Boni, L., 1997. Coppertoxicity and carbohydrate production in the microalgaeCylindrotheca fusiformis and Gymnodinium sp. Eur. J. Phycol.32, 125–132.
Pompei, M., Ghetti, A., Milandri, A., Mazziotti, C., 1995.Fioriture microalgali ed evoluzione dei principali popolamentifitoplanctonici nelle acque costiere emiliano-romagnole dal1982 al 1994. In: Proceedings of the Conference on Evoluzionedello stato trofico in Adriatico analisi di interventi attuati efuture linee di intervento, Marina di Ravenna sett, Italy, 1995,pp. 51–60.
Regione Emilia-Romagna, 1997–2002. Eutrofizzazione delle acquecostiere dell’Emilia-Romagna, Rapporti annuali. A cura di:Arpa Struttura Oceanografica Daphne. Pubbl. AssessoratoAgricoltura, Ambiente e Sviluppo Sostenibile, Bologna.
Revelante, N., Gilmartin, G., 1991. The phytoplankton compositionand population enrichment in gelatinous “macroaggregates” inthe northern Adriatic during the summer of 1989. J. Exp. Mar.Bot. Ecol. 146, 217–233.
Revelante, N., Williams, W.T., Gilmartin, G., 1984. A numericalassessment of the temporal and spatial distribution ofphytoplankton assemblages in the northern Adriatic Sea. J.Exp. Mar. Biol. Ecol. 77, 137–150.
Rinaldi, A., Vollenweider, R.A., Montanari, G., Ferrari, C.R.,Ghetti, A., 1995. Mucilages in Italian Seas: the Adriatic andTyrrhenian Seas, 1988–1991. Sci. Total Environ. 165, 165–183.
Utermöhl, H., 1931. Neue Wege in der quantitativen Erfassung desPlanktons. Int. Ver. Theor. Angew. Limnol. Verh. 5, 567–597.
Viviani, R., Boni, L., Cattani, O., Milandri, A., Pirini, M., Poletti,R., Pompei, M., 1995a. Fatty acids, chlorophylls and totalsilicon in mucilaginous aggregates collected in a coastal area ofnorthern Adriatic Sea facing Emilia-Romagna in August 1988.Sci. Total Environ. 165, 193–201.
Viviani, R., Boni, L., Cattani, O., Milandri, A., Poletti, R., Pompei,M., Sansoni, G., 1995b. ASP, DSP, NSP and PSP monitoring in“mucilaginous aggregates” and in mussels in a coastal area ofnorthern Adriatic Sea facing Emilia-Romagna in August 1988,1989 and 1991. Sci. Total Environ. 165, 201–211.
Vollenweider, R.A., Rinaldi, A., Montanari, G., 1992. Eutro-phication, structure and dynamics of a marine coastal system:results of a 10-year monitoring along the Emilia-Romagna coast(northwest Adriatic Sea). In: Vollenweider, R.A., Marchetti,R., Viviani, R. (Eds.), Marine Coastal Eutrophication. Elsevier,Amsterdam, pp. 63–106.
Copyright © 2022 FDOKUMEN




















