
Different Roles of Flowering-Time Genes in the Activation of Floral Initiation Genes in Arabidopsis

Folia Microbiol. 55 (2), 126–132 (2010) http://www.biomed.cas.cz/mbu/folia/
Core Promoters of the Penicillin Biosynthesis Genes and Quantitative RT-PCR Analysis of These Genes in High and Low Production Strain of Penicillium chrysogenum
R. ŠMIDÁKa, M. JOPČÍKa, M. KRALOVIČOVÁa, J. GAJDOŠÍKOVÁb, J. KORMANECa, J. TIMKOa, J. TURŇAa,c
aInstitute of Molecular Biology, Slovak Academy of Sciences, 845 51 Bratislava 45, Slovak Republic
fax +421 2 5930 7416 e-mail [email protected]
bBiotika a.s., 976 13 Slovenská Ľupča, Slovak Republic cDepartment of Molecular Biology, Faculty of Natural Sciences, Comenius University, 842 15 Bratislava, Slovak Republic
Received 3 July 2009 Revised version 9 December 2009
ABSTRACT. The transcription start points of the penicillin biosynthesis genes from Penicillium chryso-genum were mapped using the primer extension method. For each of the three genes consensus sequences of the core promoter elements were identified, supporting the notion that the basal transcription of these genes is mediated separately. Interestingly, transcription start of the pcbC gene is located within the potential Inr ele-ment with no TATA box-like sequence being found at expected position. This is in contrast to pcbAB and penDE genes with proposed TATA boxes or even to Aspergillus nidulans ipnA (pcbC) gene indicating pos-sible differences in basal transcription regulation. Using the quantitative RT-PCR analysis the expression of all three biosynthesis genes was monitored in both the high and low production strain of P. chrysogenum du-ring a 3-d cultivation under production conditions. The differences were found between the strains in time regulation and transcript levels of the biosynthesis genes. Furthermore, we showed that the effect of higher gene dosage on productivity in the production strain is amplified by more efficient transcription of the bio-synthesis genes with the RNA levels ≈37- and 12-times higher, respectively, than in a low production strain.
Penicillin and many of its semisynthetic derivates are still worldwide widely used -lactam antibio-tics. This compound is naturally synthesized as a secondary metabolite by filamentous fungi of the genera Penicillium and Aspergillus (Nemec et al. 1964; Nüesch et al. 1987). Industrial production is performed with Penicillium chrysogenum strains with high penicillin titers. These strains were obtained by random mutage-nesis from the original ancestor P. chrysogenum NRRL 1951 isolated in 1943 from a molded cantaloupe (Elander 2003; Rodríguez-Sáiz et al. 2005). The large number of strains with different productivity wakes P. chrysogenum a good model organism for studying penicillin biosynthesis. Although the initial analyses were performed mostly with Aspergillus nidulans the biosynthetic process in both P. chysogenum and A. nidulans has recently been well characterized in terms of molecular genetics, enzymology and biochemistry (Martín 2000; Nielsen 1997).
All three penicillin biosynthesis genes from P. chrysogenum denoted pcbAB, pcbC and penDE were cloned and characterized (Gutiérrez et al. 1999); they are clustered within the ≈15 kbp region (Aharonowitz et al. 1992; Smith et al. 1990a). The first two genes pcbAB and pcbC (encoding L-α-aminoadipyl-L-cys-teinyl-D-valine synthetase and isopenicillin N-synthetase) are divergently transcribed from the same ≈1-kbp bidirectional promoter region (Gutiérrez et al. 1991). The third penDE gene (encoding isopenicillin N-acyl-transferase) is located downstream of pcbC, with the promoter residing in an independent region.
Penicillin production in P. chrysogenum and A. nidulans was reported to be controlled by several factors, such as glucose and ammonium repression effect or alkaline pH activation, affecting one or both the expression of the biosynthesis genes and enzyme activities (Brakhage 1998; Leiter et al. 2001). A continu-ous research revealed many DNA-binding proteins interacting with promoter regions in the penicillin cluster (Chu et al. 1995; Feng et al. 1995; Kosalková et al. 2000). Some of these trans-acting factors and the cor-responding cis-regulatory elements were characterized in more detail and are proposed to enhance or suppress the expression of the penicillin biosynthesis genes in response to certain cultivation conditions (Brakhage 1998; Kosalková et al. 2007, 2009). The complex transcription control of the penicillin biosynthesis genes in

2010 PENICILLIN BIOSYNTHESIS GENES IN HIGH AND LOW PRODUCTION STRAIN OF P. chrysogenum 127
P. chrysogenum remains to be explained. To the best of our knowledge the basal promoter elements for these genes have not been precisely localized.
In this work we mapped the TSPs (transcriptional start points) and identified the core promoter con-sensus sequences for each of the three biosynthesis genes from P. chrysogenum. A subsequent real-time PCR (RT-PCR) analysis focused on transcript level changes of these genes during cultivation of P. chrysogenum under production conditions. In order to study the transcriptional deregulation in production strains, this analysis was performed with the high-producer strain from Biotika a.s. (Slovakia), P. chrysogenum NMU2/40 with four copies of penicillin cluster (unpublished results) and the original ancestor P. chrysogenum NRRL 1951.
MATERIALS AND METHODS
Strains, media and cultivation conditions. The production strain P. chrysogenum NMU2/40 (Biotika a.s., Slovenska Lubča, Slovakia) and the original strain P. chrysogenum NRRL 1951 (ATCC 9480; Cen-traalbureau voor Schimmelcultures (CBS), Ultrecht, The Netherland) were cultured on the plates with solid LCSB media (g/L: lactose 15, corn steep extract 2.5, peptone 5, NaCl 4, MgSO4·7H2O 0.5, KH2PO4 0.6, FeCl3·6H2O 0.005, CuSO4·5H2O 0.002, pH 4.8) with 30 g/L agar at 25 C. For all experiments a two-stage cultivation was performed with both strains: initial culture grew after inoculation of spore suspension (5 × 106 spores) in 50 mL in 500 mL glass flasks of liquid SEED media (g/L: lactose 10, glucose 30, phar-mamedia 10, corn steep liquor 30, (NH4)2SO4 2, CaCO3 5, KH2PO4 0.5, yeast extract 10; pH 5.7) in rotary shaker (200 rpm, 1 d, 26 C). Production was then carried out in 500 mL glass flasks filled with 50 mL of liquid PLP media (g/L: lactose 120, pharmamedia 25, (NH4)2SO4 10.5, CaCO3 10, K2SO4 5, Na2SO4 4, KH2PO4 0.5, sodium phenoxyacetate 12.5; pH 6.6) in rotary shaker (200 rpm, 26 C) after inoculation with 3 mL of seed culture.
Total RNA isolation. For primer extension mapping total RNA was isolated from production cul-tures of P. chrysogenum NMU2/40 after a 1-d cultivation. RNA was extracted from disrupted mycelia by hot acid–phenol method (Kormanec 2001). The integrity of RNA was assessed on agarose gel. Concentra-tion was evaluated spectrophotometrically by measuring absorbance at 260 nm with determining the purity of the samples based on A260/A280 ratio. Total RNA for RT-PCR was isolated using RNeasy Plant Mini Kit (Qiagen) and the contaminating DNA was removed by DNAse I treatment (RNase-Free DNase Set; Qia-gen). All the procedures followed the manufacturer’s instructions. All RNA samples were stored at –70 C.
Primer extension. The exact procedure was according to Skovierova et al. (2006). The 5´ ends of RNA transcripts were amplified using 5´--32P labeled oligonucleotides pcbABRev for pcbAB, pcbCRev for pcbC and penDERev for penDE gene. Primer extension products were gel-separated together with the G+A and T+C sequencing ladders derived from the corresponding end-labeled promoter regions prepared by PCR amplification with the labeled and unlabelled primers. Unlabeled primers were pcbCRev for pcbAB, pcbABRev for pcbC and penDEDir for penDE gene (for primer sequences see Table I). Each experiment was performed twice using independent sets of RNA with similar results.
Table I. Primers used
Gene Primer extension (5´→3´) RT-PCR (5´→3´)
pcbAB Crev: GGG GAC ATT GGC CTT GGG GG AR1: ACG TTC CAT GGC CTC GTT ATC ABrev: 32P-CGG TGG CTT CAG TTG AGT CAT G AR2: CGT GCT ATT GTG GAC CTG TTG
pcbC ABrev: CGG TGG CTT CAG TTG AGT CAT G NR1: TCG AAC AAG ACC AGG GAG TTC Crev: 32P-GGG GAC ATT GGC CTT GGG GG NR2: TAG TAT CCG GCA CGG ATC TG
penDE DEdir: GGA GGA GCC GCC ACC ATG CC TR1: ATC GGG AAG GTT GGA TTT AAC AG DErev: 32P-GGG AGT GCC TTG ACA GAG GAT G TR2: GGG CAA TAT GCG AAG GAA CTC
cDNA amplification and RT-PCR. For RT-PCR analysis cDNA was amplified from extracted RNA
by ImProm-IITM Reverse Transcription System (Promega) with random primers according to the manufac-turer’s protocol. The RT-PCR was performed with ABI 7900HT Fast RT-PCR system (Applied Biosystems) and Power SYBR® Green PCR Mix (Applied Biosystems). Primers used in analysis were designed using Primer Express software (Applied Biosystems). Primer pair AR1/2 was used for pcbAB gene, NR1/2 for pcbC and TR1/2 for penDE gene (see Table I). Each reaction contained 6 µL of appropriately diluted (20–120×) cDNA sample with 2 µmol/L of gene specific primer pair at total volume of 20 µL and was run in

128 R. ŠMIDÁK et al. Vol. 55
triplicate. The PCR conditions were: initial cycle of denaturation (10 min at 95 C) followed by 40 cycles (15 s, 95 C and 1 min, 60 C). Samples were tested for nonspecific amplifications by dissociation curve determination with an additional cycle for 15 s at 95 C, 15 s at 60 C and 15 s at 95 C. Data analysis was done with the bundled SDS software, ver. 2.3 (Applied Biosystems) using standard curve method (ABI User Bulletin no. 2) involving serial dilutions of P. chrysogenum genomic DNA. Relative transcript level values for penicillin biosynthesis genes were obtained after normalization of values calculated for the target genes against those of actA gene (-actin encoding gene) used as endogenous control. Amplification efficiencies for both the target and reference gene were verified in each cDNA sample analyzed. All the calculations were performed as in ABI User Bulletin no. 2; results are expressed as mean SD.
RESULTS AND DISCUSSION
Penicillin biosynthesis genes are tightly clustered with all DNA regulatory regions required for their substantial expression. We intended to identify the separate core promoters for all three genes based on com-plete TSPs mapping. For this purpose, young cultures grown for 1 d were found suitable for isolation of total RNA exhibiting high stability and in sufficient amount; production cultivation conditions were used for en-suring higher RNA levels of the biosynthesis genes. Studies describing the precise localization of the tran-scription start for either pcbAB or penDE gene are scarce; similarly, only two could be found reporting the primer extension analysis of pcbC gene with determining the TSP at the position –11 (Barredo et al. 1989; Brakhage et al. 2004).
Our primer extension experiments localized the 5´ end of the pcbAB transcript at the position –106 and the TSP of the penDE at –73 upstream of the ATG codons of these genes. Transcription of the pcbC gene was found to initiate at position –15 upstream of the start codon. The previously published hybridiza-tion study (Díez et al. 1990) indicated that the TSP of pcbAB is localized within the XhoI–XhoI fragment spanning and the ATG codon between the nucleotide positions –84 and +83. Here we show the more distant position of the TSP upstream from the start codon for pcbAB and slightly for pcbC. These differences could be explained by higher degradation of RNA and/or abortive termination of amplification in primer extension analysis in previous studies, in which we obtained longer amplicons. Together, although we did not detect any minor signals for the genes, the variable position of TSPs across the strains in different studies cannot be ruled out. Both the pcbAB and penDE genes contain TATA box-like motifs at expected positions at around 30 bp upstream of the TSP’s identified here. Although pcbC promoter was proposed to contain TATA box –198 bp upstream from the ATG codon (Smith et al. 1990b) this is too far from the TSP of this gene and no other TATA box-like sequences were found in close proximity. Instead, transcription of the pcbC initiates at A nucleotide within the pyrimidine-rich region representing the consensus sequence (PyPyAN(T/A)PyPy) of the less well-characterized eukaryotic basal promoter element – initiator (Inr) ele-ment (Fig. 1). It is also important to note that in A. nidulans the potential TATA boxes were identified for all three genes (Fernández-Cañón and Peñalva 1995; Pérez-Esteban et al. 1993). These findings, though insuf-ficient to form complex conclusions, indicate the potential differences in basal expression control between these genes in P. chrysogenum and possibly between P. chrysogenum and A. nidulans.
The biosynthesis genes within one cluster can possess different activity in the time (Brakhage 1998). Previous reporter gene studies on acvA(pcbAB) and ipnA(pcbC) genes from A. nidulans and P. chrysogenum (Brakhage et al. 1992; Feng et al. 1994) indicated the sequential activation of the biosynthesis genes. The quantitative RT-PCR analysis of two P. chrysogenum strains (Fig. 2A) showed that, whereas the transcript level of pcbAB gene increased the most rapidly during the first 20 h, the level of penDE transcript was not increasing until the 20 h time point. These findings could be expected, considering that the pcbAB and pcbC genes are involved in the initial stage and penDE in the final step of the biosynthesis pathway. In PLP me-dium the nonrepressive C-source (lactose) was used and RNA level of the genes was increasing within the grow the phase; no separation between trophophase and idiophase was these observed in many antibiotic-producing bacteria (Litzka et al. 1995). This indicates that the separation of phases in P. chrysogenum obser-ved with glucose was due to the presence of repressive C-source as proposed by Revilla et al. (1984) and Brakhage (1998).
The growth and expression analysis revealed interesting characteristics of the high-production P. chrysogenum NMU2/40 compared with low-production P. chrysogenum NRRL 1951 (Fig. 2A,B). Pro-duction cultures of both strains grew rapidly during the first 20 h. The differences between the strains occurred after 12 h from the beginning of the cultivation. The production strain exhibited slower growth that continued from 20 h to 40 h with a significant rate decrease, and reached the stationary phase after 40 h. Almost no mycelial dry-mass increase occurred in the NRRL 1951 culture after 20 h of cultivation and after

2010 PENICILLIN BIOSYNTHESIS GENES IN HIGH AND LOW PRODUCTION STRAIN OF P. chrysogenum 129
Fig
. 1.
P
rim
er e
xten
sion
map
ping
of
tran
scri
ptio
n in
itia
tion
sit
es f
or p
enic
illi
n bi
osyn
thes
is g
enes
in
Pen
icil
lium
ch
ryso
genu
m.
A:
Org
aniz
atio
n of
th
e pe
nici
llin
bi
osyn
thes
is
gene
cl
uste
r in
P
enic
illi
um
chry
soge
num
wit
h th
e 5´
RN
A e
nds
(P1,
P2
and
P3)
ampl
ifie
d in
pri
mer
ext
ensi
on e
xper
imen
ts (
5´ e
nd o
f am
plic
on l
abel
ed i
s m
arke
d w
ith
an a
ster
isk)
. T
he
TS
Ps
iden
tifi
ed (
pcbA
Bp,
pcb
Cp
and
penD
Ep)
and
di
rect
ions
of
the
tra
nscr
ipti
on a
re i
ndic
ated
by
bent
ar
row
s. B
: P
rim
er e
xten
sion
ana
lysi
s. P
rim
er e
xten
-si
on (
3rd
lane
s, P
1–P
3) p
rodu
cts
wer
e am
plif
ied
afte
r hy
brid
izat
ion
of 5
´-la
bele
d pr
obes
hyb
ridi
zed
wit
h 60
µ
g R
NA
and
run
on
DN
A s
eque
ncin
g ge
l al
ongs
ide
G+
A (
lane
A)
and
T+
C (
lane
T)
sequ
enci
ng l
adde
r de
rive
d fr
om t
he e
nd-l
abel
ed f
ragm
ent.
Hor
izon
tal
arro
ws
indi
cate
the
pos
itio
ns o
f pr
imer
ex
tens
ion
prod
ucts
and
thi
ck b
ent
vert
ical
arr
ows
indi
cate
the
nu
cleo
tide
s co
rres
pond
ing
to T
SP
of t
he i
ndic
ated
pr
omot
ers
(1.5
nu
cleo
tide
s su
btra
cted
fr
om
the
leng
th o
f fr
agm
ent)
. C
: N
ucle
otid
e se
quen
ce o
f th
e
P. c
hrys
ogen
um p
cbA
B,
pcbC
, an
d pe
nDE
pro
mot
er
regi
ons
wit
hin
the
clus
ter.
Num
beri
ng i
s w
ith
resp
ect
to 3
´ en
d of
the
pcb
AB
gen
e w
ith
the
inte
rrup
ted
line
s w
hich
rep
rese
nt t
he s
eque
nce
not
show
n he
re.
The
ini
tiat
ion
codo
ns a
re i
n b
old
and
the
ded
uced
pr
otei
n pr
oduc
ts c
orre
spon
ding
to
gene
s ar
e gi
ven
in
sing
le-l
ette
r am
ino
acid
cod
e in
the
sec
ond
posi
tion
of
eac
h co
don.
Bol
d a
nd u
nder
line
d re
sidu
es r
epre
-se
nt t
he T
SPs
indi
cate
d by
ben
t ar
row
s. T
he T
ATA
bo
x-lik
e se
quen
ces
upst
ream
of
the
pcbA
B a
nd p
cbC
ge
nes
and
prop
osed
Inr
ele
men
t of
pcb
C a
re u
nder
-li
ned.
130 R. ŠMIDÁK et al. Vol. 55
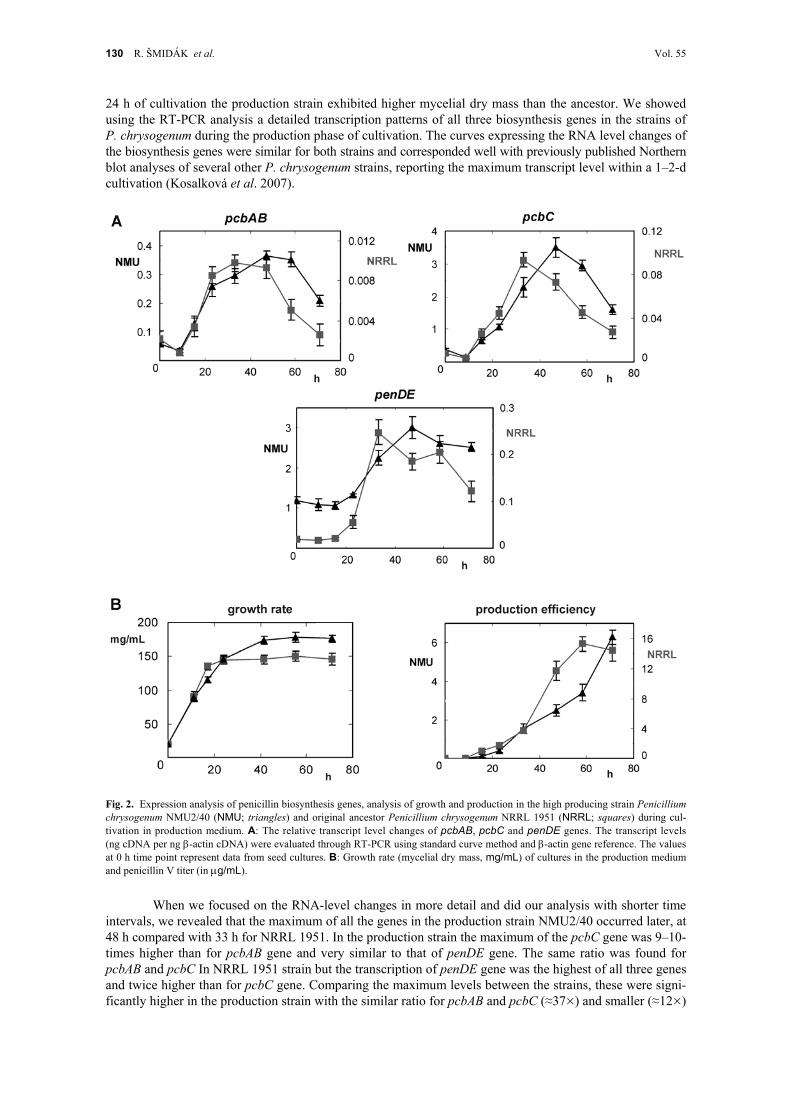
24 h of cultivation the production strain exhibited higher mycelial dry mass than the ancestor. We showed using the RT-PCR analysis a detailed transcription patterns of all three biosynthesis genes in the strains of P. chrysogenum during the production phase of cultivation. The curves expressing the RNA level changes of the biosynthesis genes were similar for both strains and corresponded well with previously published Northern blot analyses of several other P. chrysogenum strains, reporting the maximum transcript level within a 1–2-d cultivation (Kosalková et al. 2007).
Fig. 2. Expression analysis of penicillin biosynthesis genes, analysis of growth and production in the high producing strain Penicillium chrysogenum NMU2/40 (NMU; triangles) and original ancestor Penicillium chrysogenum NRRL 1951 (NRRL; squares) during cul-tivation in production medium. A: The relative transcript level changes of pcbAB, pcbC and penDE genes. The transcript levels (ng cDNA per ng -actin cDNA) were evaluated through RT-PCR using standard curve method and -actin gene reference. The values at 0 h time point represent data from seed cultures. B: Growth rate (mycelial dry mass, mg/mL) of cultures in the production medium and penicillin V titer (in g/mL).
When we focused on the RNA-level changes in more detail and did our analysis with shorter time
intervals, we revealed that the maximum of all the genes in the production strain NMU2/40 occurred later, at 48 h compared with 33 h for NRRL 1951. In the production strain the maximum of the pcbC gene was 9–10-times higher than for pcbAB gene and very similar to that of penDE gene. The same ratio was found for pcbAB and pcbC In NRRL 1951 strain but the transcription of penDE gene was the highest of all three genes and twice higher than for pcbC gene. Comparing the maximum levels between the strains, these were signi-ficantly higher in the production strain with the similar ratio for pcbAB and pcbC (≈37×) and smaller (≈12×)

2010 PENICILLIN BIOSYNTHESIS GENES IN HIGH AND LOW PRODUCTION STRAIN OF P. chrysogenum 131
for penDE gene (Table II). Higher ratios for pcbAB and pcbC in comparison with penDE indicate that the activity of the first two genes contributes in production strains remarkably to the production increase. This is consistent with the findings that introduction of additional copies of pcbAB and pcbC genes can lead to a significant production increase in transformants of P. chrysogenum (Theilgaard et al. 2001). Together with very low expression of pcbAB gene found for both strains analyzed this corresponds well with the proposal that pcbAB is rate-limiting for biosynthesis in P. chrysogenum and A. nidulans (Brakhage et al. 2004).
The production strain P. chrysogenum NMU2/40 possesses four copies of the entire penicillin clus-ter. Much higher and unequal increase in RNA levels of all three genes showed that the biosynthesis genes in this strain are transcribed more efficiently than in the single-copy NRRL 1951. Almost identical increase in pcbAB and pcbC trancripts and different from penDE implies at least partial sharing of cis-regulatory ele-ments for the first two genes.
In P. chrysogenum NMU2/40, an unknown enhancing mechanism can be suggested to act within the bidirectional promoter region, with similar effect on pcbAB and pcbC genes. Furthermore, the production strain exhibited a slower and longer growth with the expression of the biosynthesis genes peaking later but at much higher maximum transcript levels. This supports the suggestions that more balanced primary and secon-dary metabolic processes occur in higher-production strains (Spížek and Tichý 1995; Gunnarsson et al. 2004), resulting in several hundred-times higher production in NMU2/40 strain.
This work was supported by the project of Progressive Technology Development for Executive Economics, no. 2004 SP 26 028 0A 03 under the National Research Programme of the Ministry of Education of the Slovak Republic.
REFERENCES
AHARONOWITZ Y., COHEN G., MARTÍN J.F.: Penicillin and cephalosporin biosynthetic genes: structure, organization, regulation, and evolution. Ann.Rev.Microbiol. 46, 461–495 (1992).
BARREDO J.L., CANTORAL J.M., ALVAREZ E., DÍEZ B., MARTÍN J.F.: Cloning, sequence analysis and transcriptional study of the iso-penicillin N synthase of Penicillium chrysogenum AS-P-78. Mol.Gen.Genet. 216, 91–98 (1989).
BRAKHAGE A.A., BROWNE P., TURNER G.: Regulation of Aspergillus nidulans penicillin biosynthesis and penicillin biosynthesis genes acvA and ipnA by glucose. J.Bacteriol. 174, 3789–3799 (1992).
BRAKHAGE A.A.: Molecular regulation of -lactam biosynthesis in filamentous fungi. Microbiol.Mol.Biol.Rev. 62, 547–585 (1998). BRAKHAGE A.A., SPRÖTE P., AL-ABDALLAH Q., GEHRKE A., PLATTNER H., TÜNCHER A.: Regulation of penicillin biosynthesis in fila-
mentous fungi. Adv.Biochem.Eng.Biotechnol. 88, 45–90 (2004). CHU Y.W., RENNO D., SAUNDERS G.: Detection of a protein which binds specifically to the upstream region of the pcbAB gene
in Penicillium chrysogenum. Curr.Genet. 27, 184–189 (1995). DÍEZ B., GUTIÉREZ S., BARREDO J.L., VAN SOLINGEN P., LUCIA H.M., VAN DER VOORT L.H.M., MARTÍN J.F.: The cluster of penicillin
biosynthesis gene. Identification and characterization of the pcbAB gene encoding the -(aminoadipylcysteinyl)-valine syn-thetase and linkage to the pcbC and pen DE genes. J.Biol.Chem. 256, 16358–16365 (1990).
ELANDER R.P.: Industrial production of -lactam antibiotics. Appl.Microbiol.Biotechnol. 61, 385–392 (2003). FENG B., FRIEDLIN E., MARZLUF G.A.: A reporter gene analysis of penicillin biosynthesis gene expression in Penicillium chrysogenum
and its regulation by nitrogen and glucose catabolite repression. Appl.Environ.Microbiol. 60, 4432–4439 (1994). FENG B., FRIEDLIN E., MARZLUF G.A.: Nuclear DNA-binding proteins which recognize the intergenic control region of penicillin bio-
synthetic genes. Curr.Genet. 27, 351–358 (1995). FERNÁNDEZ-CAÑÓN J.M., PEÑALVA M.A.: Overexpression of two penicillin structural genes in Aspergillus nidulans. Mol.Gen.Genet.
246, 110–118 (1995). GUNNARSSON N., ELIASSON A., NIELSEN J.: Control of fluxes towards antibiotics and the role of primary metabolism in production
of antibiotics. Adv.Biochem.Eng.Biotechnol. 88, 137–178 (2004).
Table II. Maximum transcript levels of penicillin biosynthesis genes in the production and ancestral strain
Gene NMU2/40a NRRL 1951a Ratiob
pcbAB 0.365 ± 0.016 0.010 ± 0.001 36.5 ± 3.99 pcbC 3.50 ± 0.297 0.091 ± 0.007 38.5 ± 4.21 penDE 3.01 ± 0.267 0.248 ± 0.027 12.1 ± 1.70
aValues given as mean ±SD represent the relative transcript level determined by standard curve method normalized to the endoge- nous -actin gene reference. bThe relative transcript level in the production strain with respect to the ancestral strain.

132 R. ŠMIDÁK et al. Vol. 55
GUTIÉRREZ S., DÍEZ B., ALVAREZ E., BARREDO J.L., MARTÍN J.F.: Expression of the penDE gene of Penicillium chrysogenum encoding isopenicillin N acyltransferase in Cephalosporium acremonium: production of benzylpenicillin by the transformants. Mol. Gen.Genet. 225, 56–64 (1991).
GUTIÉRREZ S., FIERRO F., CASQUEIRO J., MARTÍN J.F.: Gene organization and plasticity of the β-lactam genes in different filamentous fungi. Antonie van Leeuwenhoek 75, 81–94 (1999).
KORMANEC J.: Analyzing the developmental expression of sigma factors with S1-nuclease mapping, pp. 481–494 in C.H. Chein (Ed.): Nuclease Methods and Protocols: Methods in Molecular Biology. Humana Press, Totowa (NJ, USA) 2001.
KOSALKOVÁ K., MARCOS A.T., FIERRO F., HERNANDO-RICO V., GUTIÉRREZ S., MARTÍN J.F.: A novel heptameric sequence (TTA GTAA) is the binding site for a protein required for high level expression of pcbAB, the first gene of the penicillin biosyn-thesis in Penicillium chrysogenum. J.Biol.Chem. 275, 2423–2430 (2000).
KOSALKOVÁ K., RODRÍGUEZ-SÁIZ M., BARREDO J.L., MARTÍN J.F.: Binding of the PTA1 transcriptional activator to the divergent promoter region of the first two genes of the penicillin pathway in different Penicillium species. Curr.Genet. 52, 229–237 (2007).
KOSALKOVÁ K., GARCÍA-ESTRADA C., ULLÁN R.V., GODIO R.P., FELTRER R., TEIJEIRA F., MAURIZ E., MARTÍN J.F.: The global regu-lator LaeA controls penicillin biosynthesis, pigmentation and sporulation, but not roquefortine C synthesis in Penicillium chrysogenum. Biochimie 91, 214–225 (2009).
LEITER E., EMRI T., GYÉMÁNT G., NAGY I., PÓCSI I., WINKELMANN G., PÓCSI I.: Penicillin V production by Penicillium chrysogenum in the presence of Fe3+ and in low-iron culture medium. Folia Microbiol. 46, 127–132 (2001).
LITZKA O., BERGH K.T., BRAKHAGE A.A.: Analysis of the regulation of Aspergillus nidulans penicillin biosynthesis gene aat (penDE) encoding acyl coenzyme A:6-aminopenicillanic acid acyltransferase. Mol.Gen.Genet. 249, 557–569 (1995).
MARTÍN J.F.: Molecular control of expression of penicillin biosynthesis genes in fungi: regulatory proteins interact with a bidirectional promoter region. J.Bacteriol. 182, 2355–2362 (2000).
NEMEC P., BARÁTH Z., BETINA V., KUTKOVÁ M.: Antibiotic activity of fungi isolated from soil samples from Indonesia. Folia Micro-biol. 9, 383–386 (1964).
NIELSEN J.: Physiological engineering aspects of Penicillium chrysogenum. World Scientific Publishing, Singapore 1997. NÜESCH J., HEIM J., TREICHLER H.-J.: The biosynthesis of sulfur containing -lactam antibiotics, Ann.Rev.Microbiol. 41, 51–75 (1987). PÉREZ-ESTEBAN B., OREJAS M., GÓMEZ-PARDO E., PEÑALVA M.A.: Molecular characterization of a fungal secondary metabolism pro-
moter: transcription of the Aspergillus nidulans isopenicillin N synthetase gene is modulated by upstream negative elements. Mol.Microbiol. 9, 881–895 (1993).
REVILLA G., LÓPEZ-NIETO M.J., LUENGO J.M., MARTÍN J.F.: Carbon catabolite repression of penicillin biosynthesis by Penicillium chrysogenum. J.Antibiot. 37, 781–789 (1984).
RODRÍGUEZ-SÁIZ M., DÍEZ B., BARREDO J.L.: Why did the Fleming strain fail in penicillin industry? Fungal Genet.Biol. 42, 464-470 (2005).
SKOVIEROVA H., ROWLEY G., REZUCHOVA B., HOMEROVA D., LEWIS C., ROBERTS M., KORMANEC J.: Identification of the E regulon of Salmonella enterica serovar Typhimurium. Microbiology 152, 1347–1359 (2006).
SMITH D.J., BUMHAM M.K.R., BULL J.H., HODGSON J.E., WARD J.M., BROWNE P., BROWN J., BARTON B., EARL A.J., TURNER G.: β-Lactam antibiotic biosynthetic genes have been conserved in clusters in prokaryotes and eukaryotes. EMBO J. 9, 741–747 (1990a).
SMITH D.J., EARL A.J., TURNER G.: The multifunctional peptide synthetase performing the first step of penicillin biosynthesis in Peni-cillium chrysogenum is a 421 073 dalton protein similar to Bacillus brevis peptide antibiotic synthetases. EMBO J. 9, 2743–2750 (1990b).
SPÍŽEK J., TICHÝ P.: Some aspects of overproduction of secondary metabolites. Folia Microbiol. 40, 43–50 (1995). THEILGAARD H.B., VAN DEN BERG M., MULDER C., BOVENBERG R., NIELSEN J.: Quantitative analysis of Penicillium chrysogenum
Wis54-1255 transformants overexpressing the penicillin biosynthetic genes. Biotechnol.Bioeng. 72, 379–388 (2001).
Copyright © 2022 FDOKUMEN




















