
Copyright Statement - ResearchSpace@Auckland
264
Libraries and Learning Services University of Auckland Research Repository, ResearchSpace Copyright Statement The digital copy of this thesis is protected by the Copyright Act 1994 (New Zealand). This thesis may be consulted by you, provided you comply with the provisions of the Act and the following conditions of use: • Any use you make of these documents or images must be for research or private study purposes only, and you may not make them available to any other person. • Authors control the copyright of their thesis. You will recognize the author's right to be identified as the author of this thesis, and due acknowledgement will be made to the author where appropriate. • You will obtain the author's permission before publishing any material from their thesis. General copyright and disclaimer In addition to the above conditions, authors give their consent for the digital copy of their work to be used subject to the conditions specified on the Library Thesis Consent Form and Deposit Licence.
-
Upload
khangminh22 -
Category
Documents
-
view
1 -
download
0
Transcript of Copyright Statement - ResearchSpace@Auckland

Libraries and Learning Services
University of Auckland Research Repository, ResearchSpace
Copyright Statement
The digital copy of this thesis is protected by the Copyright Act 1994 (New Zealand).
This thesis may be consulted by you, provided you comply with the provisions of the Act and the following conditions of use:
• Any use you make of these documents or images must be for research or private study purposes only, and you may not make them available to any other person.
• Authors control the copyright of their thesis. You will recognize the author's right to be identified as the author of this thesis, and due acknowledgement will be made to the author where appropriate.
• You will obtain the author's permission before publishing any material from their thesis.
General copyright and disclaimer
In addition to the above conditions, authors give their consent for the digital copy of their work to be used subject to the conditions specified on the Library Thesis Consent Form and Deposit Licence.

TheDevelopmentandAgingoftheCircadianClock
inDrosophilamelanogaster
JiaZhao
AthesissubmittedinfulfilmentoftherequirementsforthedegreeofPhD
inAnaesthesiology,theUniversityofAuckland,2019.

II
Abstract
Thecircadianclockistheendogenouspacemakerthatcontrolsdailyrhythmsinbehaviourand
physiology,anditisdrivenbytheauto-regulatedtranscription-translationfeedbackloops
ofclockgenes.Themolecularmechanismcanbefoundinalmosteverycell,whichlargely
constitutestheperipheraloscillatorsasidefromthespecializedcentralclockinthebrain.
Developmentistheseriesofchangesthatorganismsundergointheirpassagefromthe
embryonic state to maturity.Aging is the process of time-related deterioration of the
physiological functions necessary for survival.We hypothesize that the circadian clock
develops,maturesandagesinalignmentwiththechangesofthewholebodythroughout
lifespan,inasystemic,interactive,andhierarchicalmanner.
ByusingDrosophilamelanogasterastheanimalmodel,weaimedtostudywhenthemolecular
circadianclockand its lightsensitivitydevelop,howtheagedclockchanges intrinsically
underconstantconditionsaswellasundercyclicentrainment,andhowtheclockreacts
immediately preceding death. Transgenic luciferase-reporter fruit flies were used to
measurethereal-timeexpressionoftwokeyclockgenesperiodandtimelessinvivo.We
foundthatfirst,PERIODexpressioninthepresumptivecentralclockdorsalneuronsstarted
tooscillate intheembryos,whilePERIODintheperipheraltissues increasedduringthe
embryonic stage but only started to oscillate in the adult stage. Secondly, PERIOD
expressioninthecentralclockneuronsstayedrobustinagedfruitflies,however,rhythms
ofPERIODandTIMELESSintheperipheraltissuesthroughoutthebodyshowedreduction

III
in both expression level and rhythmicity. Thirdly, in the days prior to death TIMELESS
expressionincreasedandlostcircadianrhythmicity.
Thesefindingsprovethatthecentralclockisalreadyfunctionalduringembryogenesis,while
the peripheral clocks develop later, maturing only after eclosion when cyclic and
synchronizedexpressionofPERIODthroughouttheanimalcommences.Significantly,we
revealthatcyclicclockgeneexpression,presumablyinprecursorsofdorsalclockneurons
occurs during the embryonic stage, which is earlier than previously thought. Equally
important,whentheanimalsgrowold,theagedmolecularclockstillfunctionswellatthe
centrallevelbutdeclinesgraduallyattheperipherallevel.Itindicatesthattheperipheral
clocks aremore seriously damagedunder aging that they should be considered as the
potentialtargetforfurtherexplorationandpossibleintervention.Furthermore,wereport
anovelmarkerofimminentdeathintheexpressionoftheclockgeneTIMELESS.It isof
importance that thismarker in the expression of TIMELESS is not age dependent and
predictsdeathequallywellinfruitfliesofdifferentagesandunderdifferentcircumstances.
Hereweshowadynamicnetworksystemofthecircadianclockchangingthroughoutlifespan,
spatiallyfromthecentretotheperipheryandtemporallyfromthebirthtothedeath.In
summary,thecircadianclockdevelopsgraduallyduringearlystages,declineswithageand
breaksdowndaysbeforedeath, showingdistinguishing characteristicsbetweencentral
andperipheraloscillators.

IV
Acknowledgements
Specialthankstotworespectableandreliablegentlemen,DrJamesCheesemanandDrGuy
Warman,whoarekindenoughtobemysupervisorsandgenerousenoughtoleadmeto
the beautiful world of Chronobiology. Their professionalism helps pave the way, their
encouragementhelpseasethepain,andtheiropen-mindednesshelpspreservethetrue
colourofme.Ofanyachievementshavebeenmadeorwillbemade,theycontributea
substantialpart.
Thankstomyteamwhohaveorhadbeenthereforme,DongniLi,NicolaLudin,AlmaOrts-
Sebastian, Victoria King, Diana Grieve, Raewyn Poulsen, Quirino Shin and Jizhong Bai.
ThankstoDebbieBeaumontwhohasbeenextremelyniceandhelpful.ThankstoAmyZhu
whoiswillingtosharelabresourcesandexperience.ThankstotheHeadofDepartmentof
AnaesthesiologySimonMitchell,whohasneverbeendistantwhenhelpisneeded.
Thankstomybelovedones,myhusbandandourparents,whoseunconditionallovemakesme
survivenotjustthedoctoralstudy,butanychallengesImeet.Beingtherolemodelsofmy
life,theyteachmetobespirited,courageous,of integrity,andmost importantly,tobe
defeatedsometimes,toyieldnever.
SincerelovetomyhusbandJiaWang(Terry)whoneverlosesfaithinmeevenImyselfdrift
away fromtime to time,whostandsbymyside,growsupstrong toprotectme, takes
excellentcareofme,andlovesmeindefinitely.

V
Thankstomydear friendsLihuaYe,YingxinWang,HuiwenZhang,YunerWu, JianhuiChen,
Benyu Tang,WenfenChen, YonggeHuang, Jing Li, Lun Luo, BeibeiGu, XiaomingRong,
HongxuanWang,AnastasiiaArtuyants, andShabihahShahrudin,who letmebepartof
theirlivesandinreturncolourmylifewithallthesweetlittlemoments.
SpecialthankstoDrLingLi,DrZhongPeiandDrJunLiu,whosewiseguidanceandgenerous
supporthelpmeflyhigher.ThankstothosewhoevercontributetomakewhoIamand
whatIam,andthosewhoeverofferhelp,evenmostslightly,lightingupmylifewithhope.
Thanks for the opportunity to enjoy high-quality education provided by the University of
Auckland,makingmefeelvalued,respectedandsupported,financiallyandmuchmore.
Herbreathingconstantlyinspiresme.

VI
Acknowledgements
ThisstudywassupportedbytheUniversityofAucklandDoctoralScholarship
2015-2018

VII
Acknowledgements
献给赠与我名的父母,张巧女士和赵伟中先生
感谢您们的言传身教,让我时刻体会到坚毅,自律,宽容是如此贵重的品格
同样感恩我们的父母,李艾凤女士和王栋才先生
感谢您们的信任与包容,让我感到温暖,在漆黑中亦不曾恐惧和颤抖
还有我的小妹妹,张慧小姐
感谢你美丽健康的成长,以及无尽的善意和无私的陪伴
愿你们一如既往的被命运善待
祝好
ThisiswritteninChinese.Ihopethemessagecanbeconveyedtomyparentsandparents-in-law,
sayingthathowmuchIappreciatetheirendlessloveandsupport,andhowmuchIlovethem.

VIII
TABLEOFCONTENTS
Page
ABSTRACT………………………………………………......………………………………………………………………....II
ACKNOWLEDGEMENTS…………………………………………………………………………………………………...IV
TABLEOFCONTENTS.......................................................................................................VIII
LISTOFTABLES...................................................................................................................X
LISTOFFIGURES................................................................................................................XI
GLOSSARY.......................................................................................................................XIV
CHAPTER
1THEORGANIZATIONANDMOLECULARMECHANISMOFTHECIRCADIAN
CLOCK.................................................................................................................1
1.1Summary...................................................................................................................4
1.2TheCentralClockandthePeripheralClocks............................................................7
1.3TheCircadianClockanditsTickingMechanism.....................................................15
1.4TheCircadianClockanditsEntrainment................................................................22
1.5TheCircadianClockanditsOutputRhythms.........................................................28
1.6TheGoals................................................................................................................31
1.7References..............................................................................................................34
2DEVELOPMENTOFTHEMOLECULARCIRCADIANCLOCKANDITSLIGHT
SENSITIVITYINDROSOPHILAMELANOGASTER..................................................44
2.1Abstract..................................................................................................................47
2.2Introduction............................................................................................................48
2.3Objectives...............................................................................................................59
2.4Methods.................................................................................................................61
2.5Results....................................................................................................................71
2.6Discussion...............................................................................................................82
2.7Conclusion..............................................................................................................87
2.8References..............................................................................................................90

IX
3PERIPHERALBUTNOTCENTRALCLOCKCOMPONENTSDECLINEWITHADVANCING
AGEINDROSOPHILAMELANOGASTER..............................................................97
3.1Abstract................................................................................................................100
3.2Introduction..........................................................................................................101
3.3Objectives.............................................................................................................108
3.4Methods...............................................................................................................109
3.5Results..................................................................................................................114
3.6Discussion.............................................................................................................125
3.7Conclusion............................................................................................................129
3.8References............................................................................................................132
4ATIMELESSCHAOSBEFOREDEATH:THEABERRANTEXPRESSIONOFCLOCK
GENESDURINGTHELASTDAYSOFLIFEINDROSOPHILAMELANOGASTER......136
4.1Abstract................................................................................................................139
4.2Introduction..........................................................................................................140
4.3Objectives.............................................................................................................144
4.4Methods...............................................................................................................146
4.5Results..................................................................................................................151
4.6Discussion.............................................................................................................159
4.7Conclusion............................................................................................................162
4.8References............................................................................................................165
5THELIFEHISTORYOFTHECIRCADIANCLOCK...................................................169
5.1Summary..............................................................................................................172
5.2FromtheCentretotheEdge................................................................................176
5.3FromtheBeginningtotheEnd.............................................................................181
5.4TheDynamicNetwork..........................................................................................191
5.5TheImplicationinReality.....................................................................................196
5.6MessagetotheFuture.........................................................................................199
5.7References............................................................................................................201
APPENDIX.......................................................................................................................210

X
LISTOFTABLES
Page
Table2-1.CharacteristicsofDifferentLuciferase-reporterLines...........................................63
Table2-2.Free-runningPeriodsandRhythmicityAmongDDandLight-treatedFlies............81

XI
LISTOFFIGURES
Page
Figure1-1.AnOverviewoftheCircadianSystem......................................................................6
Figure1-2.TheCentralClockNeuronsandPeripheralTissuesinDrosophila...........................9
Figure1-3.Transcription-translationFeedbackLoopsofClockGenesinDrosophila..............19
Figure1-4.TheOrganizationoftheCircadianClockThroughoutLifespan.............................33
Figure2-1.DevelopmentofCentralClockNeuronatEmbryonic,LarvalandPupalStagesin
Drosophila.............................................................................................................53
Figure2-2.LifeCycleofDrosophilaandExistingFindingsonClockDevelopment..................60
Figure2-3.ExperimentalDesignoftheDevelopmentStudy...................................................61
Figure2-4.BioluminescenceAssayConductedinEarlyDevelopmentalDrosophila...............67
Figure2-5.TheLight-pulseRegimeninEarlyDevelopmentalDrosophila...............................69
Figure2-6.BioluminescenceTime-seriesDataofIndividual8.0-lucFruitFliesMeasuredinDD
at25°C...................................................................................................................73
Figure2-7.BioluminescenceTime-seriesDataofIndividual8.0-lucFruitFliesMeasuredinDD
at16°C...................................................................................................................74
Figure2-8.BioluminescenceTime-seriesDataofIndividualXLG-lucFruitFliesMeasuredinDD
at25°C...................................................................................................................76
Figure2-9.BioluminescenceTime-seriesDataofIndividualXLG-lucFruitFliesMeasuredinLD
Cyclesat25°C........................................................................................................77
Figure2-10.BioluminescenceTime-seriesDataofIndividualXLG-lucFruitFliesMeasuredat
16°C.......................................................................................................................78

XII
Figure2-11.TheCircularDistributionsoftheLocomotorActivityAcrophases,Average
ActogramsandPeriodogramsat25°Cand16°C...................................................80
Figure2-12.LifeCycleofDrosophilaandNewFindingsonClockDevelopment.....................89
Figure3-1.OverviewofExistingFindingsontheAgingoftheCircadianClock.....................107
Figure3-2.ExperimentalDesignoftheAgingStudy.............................................................109
Figure3-3.BioluminescenceAssayConductedinAdultDrosophila......................................112
Figure3-4.MeanBioluminescenceTime-seriesDatafromYoung(10-day),Middle-aged(30-
day)andOld(50-day)8.0-lucFruitFliesinDD....................................................115
Figure3-5.MeanBioluminescenceTime-seriesDatafromYoung(10-day),Middle-aged(30-
day)andOld(50-day)XLG-lucFruitFliesinDD..................................................116
Figure3-6.MeanBioluminescenceTime-seriesDatafromYoung(10-day),Middle-aged(30-
day)andOld(50-day)tim-lucFruitFliesinDD...................................................117
Figure3-7.MeanBioluminescenceTime-seriesDatafromYoung(10-day),Middle-aged(30-
day)andOld(50-day)8.0-lucFruitFliesinLDCycles.........................................119
Figure3-8.MeanBioluminescenceTime-seriesDatafromYoung(10-day),Middle-aged(30-
day)andOld(50-day)XLG-lucFruitFliesinLDCycles........................................120
Figure3-9.MeanBioluminescenceTime-seriesDatafromYoung(10-day),Middle-aged(30-
day)andOld(50-day)tim-lucFruitFliesinLDCycles.........................................121
Figure3-10.MeanBioluminescenceTime-seriesDatafromYoung(10-day),Middle-aged(30-
day)andOld(50-day)8.0-lucFruitFliesinLD/temperatureCycles...................122
Figure3-11.MeanBioluminescenceTime-seriesDatafromYoung(10-day),Middle-aged(30-
day)andOld(50-day)XLG-lucFruitFliesinLD/temperatureCycles..................123
Figure3-12.MeanBioluminescenceTime-seriesDatafromYoung(10-day),Middle-aged(30-
day)andOld(50-day)tim-lucFruitFliesinLD/temperatureCycles...................124

XIII
Figure3-13.OverviewofNewFindingsontheAgingoftheCircadianClock........................131
Figure4-1.OverviewofExistingFindingsonChangesoftheClockBeforeDeath................145
Figure4-2.ExperimentalDesignoftheDeathStudy.............................................................146
Figure4-3.RepresentativeBioluminescenceTime-seriesDataoftim-lucFruitFliesShowing
AberrantTIMSignalattheLastDaysofLife.......................................................152
Figure4-4.RepresentativeBioluminescenceTime-seriesDataofXLG-lucFruitFliesShowing
AberrantPERSignalattheLastDaysofLife.......................................................153
Figure4-5.MeanBioluminescenceTime-seriesDatafromtheLastTenDaysofLifeintim-luc
FruitFliesShowingElevatedSignalsBeforeDeath.............................................155
Figure4-6.MeanBioluminescenceTime-seriesDatafromtheLastTenDaysofLifeinXLG-luc
FruitFliesShowingElevatedSignalsBeforeDeath.............................................156
Figure4-7.LocomotorActivityofRepresentativeWildTypeFruitFliesShowingArrhythmic
BehaviourBeforeDeath......................................................................................158
Figure4-8.OverviewofNewFindingsonChangesoftheClockBeforeDeath.....................164
Figure5-1.TheDynamicNetworkoftheCircadianSystem..................................................192
Figure5-2.SummaryofOurFindingsontheLongitudinalandCross-sectionalChangesofthe
CircadianClockThroughoutLifespaninDrosophila...........................................194

XIV
GLOSSARY
AVP argininevasopressin
Circadiantime timescalecoveringonefullperiodofanoscillation
clk/CLK clock/CLOCK
CNS centralnervoussystem
CPS countspersecond
cry/CRY cryptochrome/CRYPTOCHROME
cyc/CYC cycle/CYCLE
DAM Drosophilaactivitymonitoring
DBT DOUBLE-TIME
DD constantdarkness
DN dorsalneurons
Eclosion theemergenceofaninsectadultfromthepupalcase
Entrainment thesynchronizationofanendogenousoscillationtotheperiod
ofaZeitgeber
FFT fastFouriertransform
Free-running arhythminitsun-entrainedstate(isolatedfromZeitgeber)
HBeyelets Hofbauer-Buchnereyelets
ipRGCs intrinsicallyphotoreceptiveretinalganglioncells
LD light-dark
l-LNv largeventrallateralneurons
LN lateralneurons

XV
LNd dorsallateralneurons
LNv ventrallateralneurons
LPN lateralposteriorneurons
MT Malpighiantubules
MUA multiunitactivity
PDF pigmentdispersingfactor
per/PER period/PERIOD
PG prothoracicgland
PVN paraventricularnucleus
RHT retinohypothalamictract
RT-PCR reversetranscriptionpolymerasechainreaction
SCN suprachiasmaticnucleus
s-LNv smallventrallateralneurons
tau free-runningperiod
tim/TIM timeless/TIMELESS
VIP vasoactiveintestinalpolypeptide
Zeitgeber theforcinggeophysicaloscillationwhichentrainsabiological
oscillation
ZT Zeitgebertime




CHAPTER 1
Introduction
1

The Organization and Molecular
Mechanism of the Circadian Clock

IntroductionoftheCircadianClock
3
Thecontentsofthischapterhavebeenpublished*(pleaseseeAppendixfordetails).
*ZhaoJ,WarmanGR,CheesemanJF.Thefunctionalchangesofthecircadiansystem
organizationinaging.AgeingResRev.2019Apr29.pii:S1568-1637(18)30304-0.DOI:
10.1016/j.arr.2019.04.006.

IntroductionoftheCircadianClock
4
1.1Summary
CircadianrhythmicityisthenaturalsignatureoflivesresidingonEarth,respectingtherule
of 24-hour light-dark cycles according to the Earth’s rotation. It occurs as periodic
oscillations at different levels of organization from genes to systems. This allows
physiological activities to anticipate and adjust to endogenous and environmental
demandsinatimelymanner.Therulerofsuchatimingsystemisthecircadianclock,
whichispervasiveinmosttypesofcellsandconservedamongvariousorganisms.The
circadianclockistheendogenouspacemakerthatgeneratesthe~24-hourdailyoutput
rhythms in behaviour, physiology, metabolism and cellular processes. Phenomena
controlled by these intrinsicmechanisms not only synchronizewith environmental
rhythms (entrainment) but also persist under constant conditions (free-running).
StudiesonDrosophilamelanogasteraswellasmammals,astheclassicrepresentatives
ofbothinvertebratesandvertebrates,arebothintroduced.
A functional circadian system requires three basic components, including 1) the
endogenous circadian oscillator, comprising a number of clock genes that regulate
theirownexpression,2)theinputpathwayinwhichenvironmentalinformationsuch
as light is transmitted to and entrains the circadian oscillator, and 3) the output
pathwaywhichisthedownstreammanifestationoftimedbehaviourandphysiology
(Figure1-1).Theauto-regulatedtranscription-translationfeedbackloopsofcoreclock
genes are the main driving force of the circadian clock. The cell-autonomous

IntroductionoftheCircadianClock
5
characteristicmakes almost every cell endowedwith a ticking clock, which largely
comprisestheperipheraltissuesexcludingthespecializedcentralstructure.It isthe
centralpacemaker,togetherwiththecoordinatedperipheralclocksthatformawell-
organizedsystemwhichisimportantfordailyfunctionandhealth.Thecircadianclock
is therefore never an isolated system but instead within the integrated web of
numerousbiologicalprocesses.
Thisthesisrecordsthejourneyofstudyingtheprofoundworldofthecircadianclockand
its organization composedof different parts. The aim is to explore someunknown
areassuchashowthesecomponentschangespecificallyatparticularagethroughout
lifespan.Theunderstandingofthebasicknowledgeofthecircadianclockdescribed
hereispavingthewayforathoroughinsightofthefollowingchapters.Thelastsection
illustratesthegoalsandstructureofthethesisforbetterguidanceofreading.

Central Clock
Peripheral Clocks
Molecular Clockwork
Output PathwayInput Pathway
Entrainment Rhythms
Figure 1-1. An Overview of the Circadian System
• The Central Clock and the Peripheral Clocks
• The Circadian Clock and its Ticking Mechanism
• The Circadian Clock and its Entrainment
• The Circadian Clock and its Output Rhythms
6
Arr
ange
men
t o
f C
hap
ter
On
e 1
2
3
4
1
2
3 4
The endogenous circadian oscillator with its clock genes, the input pathway that transmits
entrainment information, and the output rhythms comprise a functional circadian system, which
can be divided into the central pacemaker and peripheral clocks. All these components of the
circadian clock are introduced in corresponding sections of Chapter One.

IntroductionoftheCircadianClock
7
1.2TheCentralClockandthePeripheralClocks
Inmulticellularorganisms,thegenerationand/ormaintenanceofcircadianrhythmicityis
located in specialized tissues, which are referred to as the central clock. In the
Drosophilaadultbrain, thecentral circadianclock is composedofapproximate150
pacemakerneurons,whicharedividedintomultipleclustersbasedontheirlocation,
size,neuropeptideexpressionand function (Figure1-2).Theseare12dorsal lateral
neurons (LNd),eightpigmentdispersing factor (PDF)-expressing largeventral lateral
neurons (l-LNv), six lateralposteriorneurons (LPN),eightPDF-positive small ventral
lateralneurons (s-LNv), twoPDF-negative s-LNv, and four anteriordorsalneurons1
(DN1a),28posteriordorsalneurons1(DN1p),fourdorsalneurons2(DN2),and80dorsal
neurons3(DN3)[Helfrich-Förster,2005;PeschelandHelfrich-Förster,2011;Liuetal.,
2015].
Ithasbeensuggestedthatdifferentclustersofclockneuronswork indifferentrolesto
control behavioural output rhythms. On one hand, the dorsal neurons (DN) are
believedtoberesponsibleforcontrollingthecrepuscularactivitypatternsunderlight-
dark(LD)conditionsinthelaboratory[Velerietal.,2003;Helfrich-Försteretal.,2007].
Ontheotherhand,theventral lateralneurons(LNv)havebeenreportedtoplayan
essential role inmaintaining rhythmic locomotoractivityunderconstant conditions
[Velerietal.,2003;Helfrich-Försteretal.,2007].Ithasalsobeenproposedthatthe
centralnetworkisorganizedintotwocoupledoscillators,thePDF-expressingLNvthat

IntroductionoftheCircadianClock
8
controlthemorningpeakofactivityandtheremainingLNvthatcontroltheevening
peakofactivity [Grimaetal., 2004;Stoleruetal., 2004].Thisdual-oscillatormodel
predictsthatthePDF-positiveneuronsserveasmasterpacemakersthatregulatethe
PDF-negativeneurons[Stoleruetal.,2005].
Nonetheless,thelabeloftheLNvastheprincipalcircadianpacemakersthatarenecessary
andsufficientforthegenerationofdailyactivity[Rennetal.,1999]hasbeenremoved
byrecentstudies.Theyrevealthatthecircadianneuronalnetworkisnotorchestrated
byasmallgroupofmasterpacemakers,instead,itconsistsofmultipleindependent
oscillators,eachiscapableofgeneratingboutsofactivity.Coordinatedfree-running
rhythmsrequiretheparticipationandinteractionofallgroupsofclockneurons,which
displayuniqueandcomplexcouplingrelationshipslargelyviaPDFsignalling[Disselet
al., 2014; Yao and Shafer, 2014; Yao et al., 2016]. LNv, and specifically the PDF
neuropeptidethattheysynthesize,areindeedimportantincoordinatingacircadian
cellularnetworkwithinthebrain,butthemselvesaloneareinsufficienttosupportthe
entirecircadianprogram[Pengetal.,2003].

Figure 1-2. The Central Clock Neurons and Peripheral Tissues in Drosophila
Approximate 150 pacemaker neurons localised in Drosophila brain are specified as the central
clock (upper panel), including dorsal neurons (DN1, DN2, and DN3), lateral posterior neurons (LPN),
dorsal lateral neurons (LNd), large ventral lateral neurons (l-LNv), and small ventral lateral neurons
(s-LNv), shown as dots in corresponding colour. Apart from that, circadian oscillators throughout
the body (including non-central-clock neurons in the brain) are under the category of peripheral
clocks (lower panel). [Images drawn by Jia Zhao, 2018]
Central Clock Neurons
Peripheral Tissues
Brain
Head
9

IntroductionoftheCircadianClock
10
Thecentralclockinmammalsisthesuprachiasmaticnucleus(SCN),whichiscomposedof
roughly20,000neurons,situatedinthehypothalamusdirectlyabovetheopticchiasm
[CornélissenandOtsuka,2017].Itallstartedin1972,whenalesionstudyidentifieda
singletissue,theSCNasnecessaryforcontrollingcircadianphysiologyandbehaviour
[StephanandZucker,1972].Further,permanentlydisruptedcircadianrhythmsfrom
SCN-lesionedanimalsarerestoredbyimplantationoffoetalSCN[Lehmanetal.,1987].
Moreover, graft studies usingmutant hamster strains with short circadian periods
demonstrate that the circadian rhythmicity is determined by the genotype of the
transplantSCNratherthanthehost,furtherestablishingtheprimacyoftheSCNinthe
circadianorganization[Ralphetal.,1990].
Themechanismofthecircadianclock iscell-autonomouswhich indicatesthatthebasic
unitofcircadiantimekeepingisthecell.Itisnotsurprisingthatasidefromtheticking
machineryatthecentrallevel,thecircadianclockhasbeenfoundtobeubiquitously
conservedinnearlyallcellsofthebody[BrownandAzzi,2013].Itisalsohighlypossible
that the circadian oscillators within individual cells can respond differently to
entrainingsignals,controldifferentphysiologicaloutputs,andinteractwitheachother
and within the system as a whole. These individual cells, and/or the functional
tissues/organstheycomprise,aretheoriginofperipheraloscillators.
PeripheralclocksinDrosophilaarealltissuesshowingcircadianpropertyexclusiveofthe
centralpacemakerneurons(Figure1-2). Ithasbeenshownthatdifferentflytissues
display rhythmic bioluminescence signals of clock gene expression in luciferase-

IntroductionoftheCircadianClock
11
reporter lines, suggesting that autonomous circadian oscillators are present
throughoutthebody[Plautzetal.,1997].Likewise,peripheralclocksinmammalsare
allcircadiancells/tissues/organsexceptfortheSCN,includingvariousnon-SCNbrain
regions (suchas thehypothalamus, forebrain, olfactorybulb andpineal gland) and
non-neuronalstructures(suchastheliver,kidney,muscle,adiposetissueandblood
cells[O’NeillandReddy,2011])[CermakianandBoivin,2009].
One of the most distinctive characteristics of the central clock is its ability to run
independently, even without entraining signals, making it self-sustaining.Whereas
peripheral clocks are oscillators that require internal or external signals (including
signalfromthecentralclock)tosynchronizethustosustaintheircircadianrhythms
[Balsalobre, 2002].Under this circumstance, the rapid dampening of oscillations in
peripheralclocksinconstantconditionshasbeenreportedinDrosophila[Plautzetal.,
1997; Peng et al., 2003]. Similarly, clock gene expression dampens in mammalian
peripheraltissues(suchastheliver,spleen,kidney,heartandlung)invitrowithindays,
butsustains intheSCNexplants forweeksorevenyears inconstantdarkness (DD)
[Yamazakietal.,2000].
Thedifferenceonsustainabilityofclockgeneoscillationsinconstantconditionsbetween
thecentralandperipheraltissuesillustratesamorefundamentaldissimilarityontheir
networkstructure.Thereasonunderlyingthedampeningofperipheralsignalsisaloss
ofsynchronyamongindividualcellsandthusthedeclineoftheensemblerhythmat
the population level [Welsh et al., 2004], rather than a weakening of circadian

IntroductionoftheCircadianClock
12
rhythmicity at the cellular level. It is supported by the finding that fibroblasts
(individualperipheralcells)showpersistentoscillationsinculturethatevenexceedthe
robustnessofindividualSCNneurons[Welshetal.,2004].Asconsequence,theunique
tight-couplingsystemthatinterconnectsthecentralclockneuronsisaprimaryfeature
ofthecentralpacemakerroleanddistinguishesitfromperipheraltissues.Ontheother
sideofthecoin,theloose-couplingofcellswithintheperipheralnetworkanswersfor
itstendencyofdesynchronization[BrownandAzzi,2013].
Speakingof the relationshipbetween theperipheralandcentraloscillators, it isalways
more than a simple story. Due to their heterogeneous nature, the oscillatory
machineryandentrainmentmechanismofperipheralclocksvarybetweendifferent
tissuesandorgansinDrosophila[ItoandTomioka,2016],leadingtothediversityof
theirrelationshiptothecentralclock.Peripheraloscillatorsinfruitfliescanrespond
directly to thesameentrainmentcues thatset thephaseof thecentralpacemaker
neuronsinthebrainvialight-andtemperature-sensitiveintracellularpathways[Allada
andChung,2010].Inthiscase,someperipheraltissuesareindependentofthecentral
clock, such as theMalpighian tubules (MT) [Giebultowicz andHege, 1997] and the
antenna[Krishnanetal.,1999;Tanoueetal.,2004].Forexample,theperipheralclock
inMTiscell-autonomousandabletobeentrainedtoenvironmentallightwithoutany
cuesfromthecentralclock[Hegeetal.,1997].Inaddition,theoriginalphaseofMTis
maintainedinDDevenwhenit istransplantedintotheabdomenoffliespreviously
entrainedtoantiphaseLDcycles[Giebultowiczetal.,2000].

IntroductionoftheCircadianClock
13
Someperipheraloscillatorsaredrivenbythecentralclock,similartothoseinmammals,
such as prothoracic gland (PG). Mechanism that couples the central clock to PG
underlies the circadian control of adult emergence, when central clock exerts a
dominant role on the peripheral clock through PDF signalling [Myers et al., 2003;
Selchoetal.,2017].Similarly,thecentralclocksendsoutPDFtoremotelycontrolthe
clockinoenocytetoregulatereproductivebehaviour[Kruppetal.,2008;Kruppetal.,
2013].Moreover,amorecomplexrelationshiphasbeenrecentlyreportedinthefat
body,where thecyclingofsomegenesdependsonthe localclockwhereas thatof
othergenesoccurs inresponsetocentralbrainsignals[Xuetal.,2008;Erionetal.,
2016].
Inmammals,theSCNdoesnotgeneratetheperipheralrhythms,rather,itisacoordinator
andsynchronizerattheheadofadistributedorganizationofindividualclocks.Many
of the fundamental properties of circadian oscillations in peripheral clocks in vivo
[Taharaetal.,2012]andinvitro[Izumoetal.,2014]aremaintainedintheabsenceof
SCNfunction.However,inawell-organizedcircadiansystem,acentralregulationof
peripheraloscillatorsfromtheSCNisstillcrucial.Naturally,an“orchestra”modelhas
been introduced to describe a balanced relationship between the central and
peripheralclocks[Osteretal.,2002].Inthismodel,thecentralclockSCNbehavesasa
conductor,witheachperipheralclockactsasindividualmemberswhohavetheability
to play their own rhythms. Harmony is only possible under the guidance of the
conductorandthecoordinationoftheteam.Inaword,althoughperipheralclockscan

IntroductionoftheCircadianClock
14
adapt to their own external signals at particular conditions, they are dominantly
regulatedbythecentralclock[RichardsandGumz,2012].

IntroductionoftheCircadianClock
15
1.3TheCircadianClockanditsTickingMechanism
Themolecularmechanismofthecircadianclockiscomposedofasetofspecializedgenes,
named clock genes, which are required for the running of the clock. It is widely
acceptedthattheclockisdrivenbythetranscription-translationfeedbackloopsofthe
expression of these clock genes. In Drosophila, the clock genes period (per) and
timeless(tim)arethoughttobekeyandnecessaryplayersofthemolecularoscillator,
giventhefactthatper-nullandtim-nullmutantseachabolishesclockfunction[Sehgal
etal.,1996;Stanewskyetal.,2002;Velerietal.,2003].
Asidefromperandtim,otherclockgenesandaccessorygenesarealso involved inthe
mechanismoftheauto-regulatedfeedbackloopsinDrosophila,thetheoryhasbecome
moreandmoreclearattributedtopreviousstudies [Hardinetal.,1990;Zerretal.,
1990;Ederyetal.,1994;Sehgaletal.,1995;Hunter-Ensoretal.,1996;Myersetal.,
1996;Zengetal.,1996;Haoetal.,1997;Alladaetal.,1998;Darlingtonetal.,1998;
Klossetal.,1998;Priceetal.,1998;Leeetal.,1999;Klossetal.,2001;McDonaldetal.,
2001;Wangetal.,2001;Shaferetal.,2002;Ashmoreetal.,2003;Nawatheanand
Rosbash,2004;KimandEdery,2006;Yuetal.,2006]andreviews[Hardin,2005;Hardin,
2006;HardinandPanda,2013;YuandHardin,2006].Thefirstcorefeedbackloopisa
negatively-regulated one composedmainly of geneper and tim, their translational
productsPERIOD(PER)andTIMELESS(TIM)incytoplasmformaheterodimerwhich
entersthenucleustoinhibitthetranscriptionofperandtimthemselves.Thesecond

IntroductionoftheCircadianClock
16
corefeedbackloopisapositively-regulatedonecomposedmainlyofthegeneclock
(clk) and cycle (cyc), whose corresponding proteins CLOCK (CLK) and CYCLE (CYC)
heterodimerize in thenucleusand then facilitate transcriptionofotherclockgenes
suchasperandtimbyrecognizingtheirspecializedpromoterregionsknownastheE-
box.
These two feedback loops work closely together in an antiphase manner to generate
circadianrhythms,andthedetailshavebeenreviewedinotherpapers[e.g.Yuand
Hardin,2006].Withina24-hourLDcycle(Figure1-3),transcriptionofperandtim is
initiated around midday, when CLK-CYC heterodimer acts as the key activator
[Darlingtonetal.,1998;Haoetal.,1997;McDonaldetal.,2001;Wangetal.,2001].
Subsequently,perandtimmRNAlevelsaccumulateandreachpeakvaluesintheearly
evening.Accordingly,inthecytoplasm,PERandTIMproteinlevelscontinuetoriseand
reachtheirmaximumlevelsinthemiddleofthenight,delayedbyseveralhoursofthe
peaklevelofperandtimmRNA[Ederyetal.,1994;Hardinetal.,1990;Hunter-Ensor
etal.,1996;Myersetal.,1996;Sehgaletal.,1995;Zengetal.,1996;Zerretal.,1990].
Accumulation of PER and TIM only happens after dark because the indirect light-
sensitivityofTIM(refertoSection1.4).PERisphosphorylatedbyDOUBLE-TIME(DBT)
kinase and, without TIM, is targeted for degradation. Thus, both proteins are
progressivelyphosphorylated,leadingtotheirdegradationwhenlightispresent.After
sundown,TIMisabletoaccumulateinthedarkandformacomplexwithPERandDBT,
givingthepossibilityofPER’sstabilization[Klossetal.,1998;NawatheanandRosbash,
2004; Price et al., 1998]. As DBT-PER-TIM accumulates, phosphorylation of TIM is

IntroductionoftheCircadianClock
17
believed to be a crucial step that triggers the entry of DBT, PER and TIM into the
nucleus,wheretheyinhibitthefunctionoftheirtranscriptionalactivators,theCLK-CYC
complex[Ashmoreetal.,2003;Klossetal.,2001;Shaferetal.,2002].ByremovingCLK-
CYCfromE-boxesandpromotingDBT-dependenthyper-phosphorylationofCLK,per
andtimrepresstranscriptionoftheirownaswellasothergenes[KimandEdery,2006;
Nawathean and Rosbash, 2004; Yu et al., 2006]. At dawn, a light-induced
conformationalchangeinCRYPTOCHROME(CRY)promotestheformationofCRY-TIM
complexes,leadingtoTIMdegradationandCRYdestabilization[AshmoreandSehgal,
2003]. Promoted by DBT-dependent phosphorylation, PER and CLK are also under
degradationduringtheearlymorning.PERthusgraduallyfallstoitslowestlevelsby
themiddleoftheday,however,CLKlevelsremainrelativelyconstantbecausehypo-
phosphorylated CLK is generated by new CLK synthesis, or de-phosphorylation of
hyper-phosphorylatedCLK[KimandEdery,2006;Yuetal.,2006].Backtothebeginning,
the degradation of PER and TIM relieves transcriptional inhibition, hypo-
phosphorylated CLK forms a heterodimer with CYC and binds to E-boxes, thereby
initiatesanewcycleofperandtimtranscription.
ThecircadiantimekeepingmechanismisconservedinmammalsasinDrosophila[Hardin
and Panda, 2013]. A pair of transcriptional factors CLOCK and BMAL1 form
heterodimertoactivatetranscriptionofgenesencodingtheirrepressors,PERIOD1-3
(PERs) and CRYPTOCHROME 1-2 (CRYs). Complexes of PER-CRY accumulate in the
cytoplasmandmoveintothenucleus,bindingtoandinactivatingtheCLOCK-BMAL1
torepresstheirowntranscription.PERsandCRYsarethendegraded,whichpermits

IntroductionoftheCircadianClock
18
theactivators tostart thenextcycleof transcription.Similar toDrosophila,CLOCK-
BMAL1 also activate a second interlocked feedback loop that controls rhythmic
expressionofactivatorgeneswhicharetranscribedintheoppositecircadianphaseas
repressorgenes.

DAWN MID-DAY
SUNDOWNLATE NIGHT
NUCLEUS
PER
tim
TIM
per
CLK
TIM
PER
TIM
TIM
TIM
TIM
PERPER
PERPER
TIM
PER
PER
TIM
CYC
CYTOPLASM
DBT
CYCCLKP
P
P
P
CRY
Figure 1-3. Transcription-translation Feedback Loops of Clock Genes in Drosophila
Starting from mid-day (top-right) and continuing clockwise until dawn (top-left), transcription of
per and tim is initiated, their proteins PER and TIM rise and reach peak levels after sundown, then
enter into the nucleus, where they repress transcription of their own. PER and TIM are then
degraded after dawn, which relieves transcriptional inhibition, thereby staring another cycle of
transcription-translation. per/PER: period/PERIOD, tim/TIM: timeless/TIMELESS, CLK: CLOCK, CYC:
CYCLE, CRY: CRYPTOCHROME, DBT: DOUBLE-TIME, P: phosphorylation. [Modified from Yu and
Hardin, 2006]
19

IntroductionoftheCircadianClock
20
Themolecularmechanismsthatgeneratecircadianoscillationsarelikelytobesimilarbut
notidenticalinthecentralpacemakerandintheperipheraltissues[Balsalobre,2002].
Analysisofindividualclockproteindeficiencyshowsclearlythatthebroadoutlineof
clockmechanism is the same between the central and peripheral clocks, including
feedback loops of transcription, translation, and post-translational modification
[Yagitaetal.,2001].However,itisalsoevidentthatthecell-autonomousclocksticking
ineachtissuehavetheirowncomplementofcoreandassociatedclockgenesthatmay
differ in abundanceor function. These subtle variations arebelieved tobedirectly
involved in the downstream timekeeping events, leading to pronounced tissue
specificity in clock-controlled genes and therefore the output physiology that they
direct[BrownandAzzi,2013].
Bywayofexample,thephotoreceptorCRYinDrosophilacontributestooscillatorfunction
intheperipheraltissuesbutnotinthecentralclock[Krishnanetal.,2001].Further,
comparisonofrhythmicallyexpressedgenesinheadandbodyrevealstissue-specific
controlofcircadianoutputfunctions[Cerianietal.,2002].Forsomecases,different
homologsofcertainclockgenesseemtoplayimportantrolesinclockmechanismin
some tissuesbut is redundant inothers.Oneexample is themammalianPer3, the
deletionofwhichhasonlysmallimpactonthecentralclockmechanism[Shearmanet
al.,2000],yetshowsapronouncedeffectinspecifictissueslikethepituitary,liverand
aorta[Pendergastetal.,2012].AnotherexampleisthatmammalianCLOCKdepletion
abolishes circadian rhythm generation in peripheral organ slices [DeBruyne et al.,
2007],butnotinculturedSCNslices.Inaddition,theobservationofdifferentcircadian

IntroductionoftheCircadianClock
21
phases[Yamazakietal.,2000;Yooetal.,2004]aswellasdifferentfree-runningperiods
[Pendergastetal.,2012]ofclockmachineryinperipheraltissueexplants[Yamazakiet
al.,2000;Yooetal.,2004]alsosupporttheexistenceoftissue-specificdiversityofclock
mechanism.

IntroductionoftheCircadianClock
22
1.4TheCircadianClockanditsEntrainment
The circadian clock is endogenousbut it has alsoevolved sophisticatedmechanisms to
reacttoenvironmentstimuli.Externalfactorssuchas light,temperature,andsocial
cues,aresocalledZeitgeberswhichcanentrainthecircadianclocktoitsenvironmental
rhythms[PeschelandHelfrich-Förster,2011].
LightisanimportantandpowerfulZeitgeberthatitcansynchronizecircadianclockswith
the24-hourrotationoftheEarth.Lightsensitivityisimplicatedasacharacteristicofa
fully functional clock that can receive stimuli from the environment, transmit this
informationtotheoscillator,thencontroldownstreamoutput[Plautzetal.,1997].A
shortlightpulseinthelatenightmimicsprematuredawn,causingaphaseadvance,
whileashortlightpulseintheearlynightcausesaphasedelay,foritmimicsdelayed
dusk[Shangetal.,2008].
AdultDrosophilahaveatleastthreephotoreceptiveorgansforlightreceivingandsignal
transductiontothecentralclockneurons,includingthecompoundeyes,theocelliand
theHofbauer-Buchner(HB)eyelets,withRhodopsinsexpressed[Chang,2006;Peschel
and Helfrich-Förster, 2011]. The compound eyes are the largest photoreceptive
structureandarethoughttobethemostimportanttolightentrainment[Yoshiietal.,
2015]. Both humoral signals such as histamine and/or direct nervous connection
between the visual organs and clock neurons are believed to be involved in the

IntroductionoftheCircadianClock
23
transmissionofphoticinformation.Infact,ourunderstandingonthepathwaysthat
lightreachesthecentralclockneuronsandtheirconsequentmolecularreactionhas
beenextending.TIMproteinhasbeenfoundtobeimportantinthispathwayandthen
CRY protein is introduced to make the story more complete. Light-dependent
degradationofTIMistheveryresponsetodailyorseasonallightchanges,leadingto
phaseshiftsinthemolecularandbehaviouralrhythms.Italsoexplainswhyaflyacts
arrhythmically in constant bright-light conditions resulting from the lack of TIM.
However,TIMitselfisnotdirectlyresponsivetolight,itisCRYwhichactsasacircadian
photoreceptor[Ivanchenkoetal.,2001].CRYisablue-lightphoto-pigmentexpressed
inasubsetsofclockneuronsandthecompoundeyes,whichcandirectlyinteractwith
TIMtoresetthemolecularclock.Light-activatedCRYbindstoTIMandTIMdegradation
isthustriggered[PeschelandHelfrich-Förster,2011].Moreover,TIMoscillationscan
alsobesynchronizedviaCRY-independentRhodopsinphoto-transduction,aswellas
morenoveltransductionpathwaysthathavebeenrecentlyuncovered[Oguetaetal.,
2018]. Aside from TIM and CRY, PDF has been found to be essential for light
entrainment in the central clock. The compound eyes andHB eyelets convey light
signalstothePDF-expressingneurons(LNv)andleadtotheresettingofthemolecular
clock. The light-reset LNv in turn synchronize other clock neurons through the
intercellularmessengerPDF[Yoshiietal.,2015].
Similartothecentralclock,directlight-entrainingpathwayalsoexistsinperipheraltissues
ofDrosophilawhichhavesemi-transparentbodies.Ithasbeenreportedthatthecell-
autonomous expression and action of the photoreceptor CRY togetherwith TIM is

IntroductionoftheCircadianClock
24
implicatedinthephoticinputpathwaynotonlyinthecentralbutalsoperipheralclocks
[Ivanchenkoetal.,2001;Krishnanetal.,2001;DubruilleandEmery,2008;Agrawalet
al., 2017]. Each tissue and even each cell are believed to possess photoreceptive
propertieswhichcontributetophoticsignaltransductionthatallowstheperipheral
oscillatorstobedirectlyresetby light.Thefindingofthenearly identicalwaveform
andphaseof therhythmsfromthreebodyparts (head, thorax,orabdomen)when
comparedwithwhole-animalrecords[Plautzetal.,1997],andtheresultofthesame
pertranscriptionpeakingtimeinallflytissues[HardinandPanda,2013],bothsupport
the theory that peripheral clocks have their own photoreceptors as well as
mechanismsthatcandirectlyreceiveandreacttolightstimuli.
Inmammals,lightisthemainsynchronizeroftheSCN.Photiccuesareperceivedbythe
retinaandsignalisthentransmittedtotheSCNviatheretinohypothalamictract(RHT).
TheSCNreceivesdirectphoticinputfromaphotoreceptorcelltypeinretinaknownas
intrinsically photoreceptive retinal ganglion cells (ipRGCs), and ipRGCs-expressing
photo-pigmentmelanopsin isconsideredtheprincipalcircadianphotoreceptors [Do
andYau,2010].
Different to peripheral clocks in Drosophila, mammalian peripheral clocks cannot be
directlyentrainedbylight.OneevidenceisthatmammalianPeriods(Pers)peakaround
4to8hours later inperipheraltissuesthanintheSCNunderLDcycles[Hardinand
Panda,2013].SuchtimelagreflectstheprocessthatlightinitiallyresetstheSCNwhich
then acts to synchronize peripheral oscillators [Balsalobre, 2002]. In other words,

IntroductionoftheCircadianClock
25
peripheral tissues inmammals are largely entrained by the SCN, the loss ofwhich
resultsinperipheralclocksthatbecomedesynchronized[Yooetal.,2004;Izumoetal.,
2014].Forinstance,rhythmsofcircadiangeneexpressionarelostinperipheraltissues
ofSCN-lesionedrats[Sakamotoetal.,1998].TheSCNcontrolscircadianrhythmicityof
peripheral clock via two major pathways, neural and humoral signals [Balsalobre,
2002]. Both sympathetic andparasympathetic nervous systems are involved in the
neuralpathway.Sympatheticefferentshavebeendocumentedforthebrownadipose
tissue,thyroid,kidney,bladderandspleen,andparasympatheticefferentsreportedin
theliver,pancreasandsubmandibulargland.Sometissuesareeveninnervatedboth
sympatheticallyandparasympatheticallybytheSCN,suchas theheartandadrenal
gland[BrownandAzzi,2013].Thesecondmajorpathwayisthroughhormones,which
hasbeenwiselyprovedintheSCNlesionstudywherearrhythmicbehaviourcanbe
rescuedbyimplantationoffoetalSCNtissueencapsulatedinporousplastic[Silveret
al., 1996]. Circadian gene expression is also observed in circulating mononuclear
leukocytes, supporting that humoral signals must be involved in peripheral
entrainment [Oishi et al., 1998]. Further, the role of glucocorticoids in the
synchronizationoftheperipheralclockshasbeenrevealed[Balsalobreetal.,2000;Son
etal.,2011].
AsidefromacceptingsignalsfromtheSCN,localclockssometimestakecuesdirectlyfrom
their external and internal environment, among all themost important peripheral
Zeitgeberisfood[BrownandAzzi,2013].Patternsoffoodintakecandirectlyentrain
clocks inperipheralorganssuchastheliver. Italso impliesthepossibilityofcertain

IntroductionoftheCircadianClock
26
peripheraltissuestoescapefromthecentralcontrolinparticularconditionssuchas
restrictedfeeding.Thisallowslocalclockstoadaptthemselvestocuesincompatibleto
thecuesSCNreceives(mainlylight)[Balsalobre,2002].Elegantexperimentshavebeen
designedtoprovethisfeatureofperipheralclocksbychangingthetimingoffeeding
andtherebyforcinganocturnalrodenttoeatduringdaytime.Iffoodisavailablead
libitum,miceandratseatmainly(80%)duringthedarkphaseofthedaywhenthey
areactive.Inversionofthetimingoffoodavailabilitylimitedtothelightphasewillturn
thetimingofclockgeneexpressioninperipheraltissuesupto180degreesinphase
comparedtoanimalsfedduringthedarkphaseoradlibitum.Atthesametime,itshifts
theperipheralclockstotheantiphaseoftheSCN,whichremainsentrainedtotheLD
cycles[Damiolaetal.,2000;Stokkanetal.,2001].Noticeably,thespeedaswellasthe
degreeofphaseshiftinducedbyfeedingvaryamongdifferentorgans[BrownandAzzi,
2013].
Apartfromlight,itiswellknownthatanotherZeitgeber,temperaturecanentraincircadian
clocks. However, temperature only acts as a weak Zeitgeber and the clock is
characterized as temperature-compensated [Refinetti, 2010]. It has been proposed
thatwithinthecentralclockofDrosophila,s-LNvaretemperature-insensitivewhileDN
are temperature-sensitive, such a two-oscillator model helps to explain how the
circadian clock maintains both temperature entrainment and temperature
compensation[BarberandSehgal,2018].Inmammals,theSCNisstrictlytemperature-
resistanthoweverperipheralclocks,includingthefibroblasts, liver,kidneyandlung,
areexquisitelysensitivetobodytemperaturechanges[Brownetal.,2002;Abraham

IntroductionoftheCircadianClock
27
etal.,2010;Buhretal.,2010].Bothincellsandinlivingmammals,subtlefluctuations
in body temperature (1°C to 4°C) are sufficient to entrain peripheral circadian
oscillators [Brown et al., 2002; Abraham et al., 2010; Buhr et al., 2010]. Inversing
circadianbodytemperaturefluctuationsinmicebyenvironmentaltemperaturecycles
will inverse circadian gene expression in peripheral cells independent of the SCN
[Brownetal.,2002].
Insummary, thecentralandperipheralphoticentrainment inDrosophila sharessimilar
pathwaysinvolvinglight-sensitiveCRYanditscloseinteractionwithTIM.Entrainment
inmammaliancentralclock ismainlyvia light,andthatoftheperipheralclockshas
proventobeawebofbothdirectandindirectsignalsthatcanevenvaryfromtissue
totissue[BrownandAzzi,2013].Feedingrhythmsarethedominantphaseresetting
cuesformostperipheralorgans.Evenso,foodintakeandbodytemperaturestillarise
asaconsequenceofcircadianbehaviourdrivenbytheSCN.Althoughthemechanism
by which peripheral oscillators can be entrained by food and temperature is still
unclear,thepathwaysarebelievedtobedifferentintheSCNandvariousperipheral
clocksinatissue-dependentmanner.

IntroductionoftheCircadianClock
28
1.5TheCircadianClockanditsOutputRhythms
Awidearrayofphysiologicalandbehaviouralprocessesareundercircadiancontrol.These
includeenergymetabolism,bodytemperature,endocrinesecretion,immunefunction,
renalactivity,gastrointestinaltractmotility,cardiovascularparameterssuchasblood
pressure and heart rates, and among all, sleep-wake cycles and locomotor activity
[Gachonet al., 2004; Froy,2011]. Thanks toDNAmicroarray studies, theextentof
circadianregulationhasbeenrevealed,inwhichupto5%oftranscriptsinDrosophila
headsandup to10% inmouse tissuesareexpressed ina circadianmanner. These
downstream cycling genes can be mapped to a broad range of intracellular and
intercellular functions including peptide synthesis, vesicle trafficking, nutrient
metabolism,detoxification,cytoskeleton,andsynapticstructure[Valloneetal.,2007].
Ofinterest,tissuespecificityisimplicatedbecausemanyofthegenescyclinginone
tissuebeingexpressedatconstantlevelsinothertissues.
Oneofthemostimportantbehaviouraloutputsinthestudyofthegeneticsandmolecular
biologyofcircadianclockinDrosophilaisthelocomotoractivityrhythms,whichcould
be considered as the equivalent of sleep-wake cycles. Importantly, one cannot
conclude a rhythm is circadian unless it persists under constant environmental
conditions,duetothemaskingeffectofLDcycles.InDrosophila,rhythmsoflocomotor
activitypersistinDD,whichisthereforebelievedtobeendogenouslyclock-regulated.
Asaresult,themeasurementoflocomotiverhythmshasbeenwidelyusedasthebasis

IntroductionoftheCircadianClock
29
forassessingcircadianfunction[Shaw,2003].Otherclock-controlledrhythmicevents
includingcourtship,egglaying,eclosion,olfactoryandgustatorysensitivity, learning
andmemory,havealsobeenstudied[PeschelandHelfrich-Förster,2011].
It has generally observed that in standard conditions of 12h:12h LD cycles at 25°C,
Drosophiladisplaytwopeaksoftheirlocomotoractivity,onearound“dawn”(usually
anartificiallyset light-ontime)andonearound“dusk” (light-off time),showingthe
classic“crepuscular”characteristics[RosatoandKyriacou,2006;RosatoandKyriacou,
2008].Duringdaylighthoursbetweendawnanddusk,theactivitylevelislow[Peschel
and Helfrich-Förster, 2011]. At night, flies tend to enter a sleep-like rest state
[Hendricksetal.,2000].Preciselyspeaking,theoscillationsofenhancedactivityofflies
areobservedunderlaboratoryinsteadofnaturalenvironment,whichprovidesmuch
richer cycling environmental stimuli. As consequence, some assumptions may be
renewedaccordingtoafindingrevealingtheDrosophilacircadianbehaviourthrough
naturalobservations.Theseincludethechangeofthefly’screpuscularactivity,athird
major locomotor component in addition to “dawn” and “dusk”, the fly’s nocturnal
behaviour under moonlight, and the dominance of light stimuli over temperature
[Vaninetal.,2012].
Circadianrhythmicityhasbeenobserved inmostaspectsofmammalianphysiologyand
behaviour, some are controlled by peripheral clocks and others directly by central
signals [Balsalobre, 2002]. Being at the top of the hierarchical organization of the
circadian timing system, theSCNare important for regulatingkey targets including

IntroductionoftheCircadianClock
30
rhythmic hormone secretion (especially night-time synthesis of melatonin by the
pineal gland), body temperature and locomotor activity [Albrecht, 2012]. As to
peripheralclocks,whichplayan integralanduniquerole ineachoftheirrespective
tissues,arebelievedtopossesstissue-specificcyclinggeneprofilestofitintheirown
physiologicalfunctions.Thusitisnotdifficulttoimaginethatcircadianphysiologyin
localtissuesislargelycontrolledbyperipheraloscillatorsthemselves[BrownandAzzi,
2013].Forinstance,peripheralclocksincardiactissuescontrolexpressionofmultiple
kinasesandionchannels,andclockmutationchangescardiacphysicalactivity[Koet
al.,2011].Inaddition,circadianclockablationinpancreaticisletsleadstodefectsin
couplingofbetacellstimulustoinsulinsecretion.Moreover,clockfunctionisessential
forcircadianproductionofglucocorticoidsinadrenalglandandcircadianexpression
ofglucocorticoidreceptorsintargettissues[BrownandAzzi,2013].

IntroductionoftheCircadianClock
31
1.6TheGoals
Theaimoftheworkdescribedinthisthesisistoexplorethenatureofthecircadianclock
and its changes throughout lifespan, from the centre to the periphery, from the
entrainmentinputtothemolecularclockandtotheoutputrhythms,andfromthepre-
maturedevelopmenttotheage-relateddeterioration(Figure1-4),askingquestionsas
follows:
• Whendoesthemolecularclockstarttotickandberesponsivetolight?
• Dothecentralandperipheralclocksdevelopatthesamepace?
• HowdoestheZeitgeberlightaffectthedevelopmentofthecircadianclock?
• Whichpartofthecircadianclockisaffectedbyaging,thecentreortheperiphery?
• Canstrongentrainmentrescuetheage-relateddeclineofthecircadianclock?
• Whathappenstotheclockatbothmolecularandbehaviourallevelstowardsdeath?
Inordertofindouttheanswers,weuseDrosophilamelanogasterastheanimalmodel,the
luciferasereporterstorecordthereal-timechangesofclockgeneexpression,andthe
locomotor activity monitoring system to measure the behavioural rhythms.
Experimentshavebeenconductedinvivoforalongdurationoftimetocollectasmuch
detailed information as possible, reflecting the cross-sectional as well as the

IntroductionoftheCircadianClock
32
longitudinal changes of the circadian clock. The adult circadian clock is well
characterized at both themolecular and behavioural level,which can serve as the
standardforcomparisonbetweenchangesfromspecificlifestagesthatweinterestin.
Correspondingly,therestofthesisisorganizedintothreeseparatechapterseachfocuses
onDEVELOPMENTofthecircadianclock(ChapterTwo),AGINGofthecircadianclock
(ChapterThree),andchangesofthecircadianclockprecedingDEATH(ChapterFour).
Lastbutnotleast,itcomesthegeneralDISCUSSION(ChapterFive).
Webelievethesetopicsareofsubstantial importance. Insightofthecircadianclock, its
development and its aging, contributes to a better understanding of the biological
rhythms that most organisms live on. Especially when it comes to a number of
pathological states causedby circadiandysfunctionat certain timingof specific life
stages. It has further implication on revealing the underlyingmechanism of clock-
relateddiseases,whichmaypointatnewdirectionstowardspotentialpreventionand
treatmentstrategies.
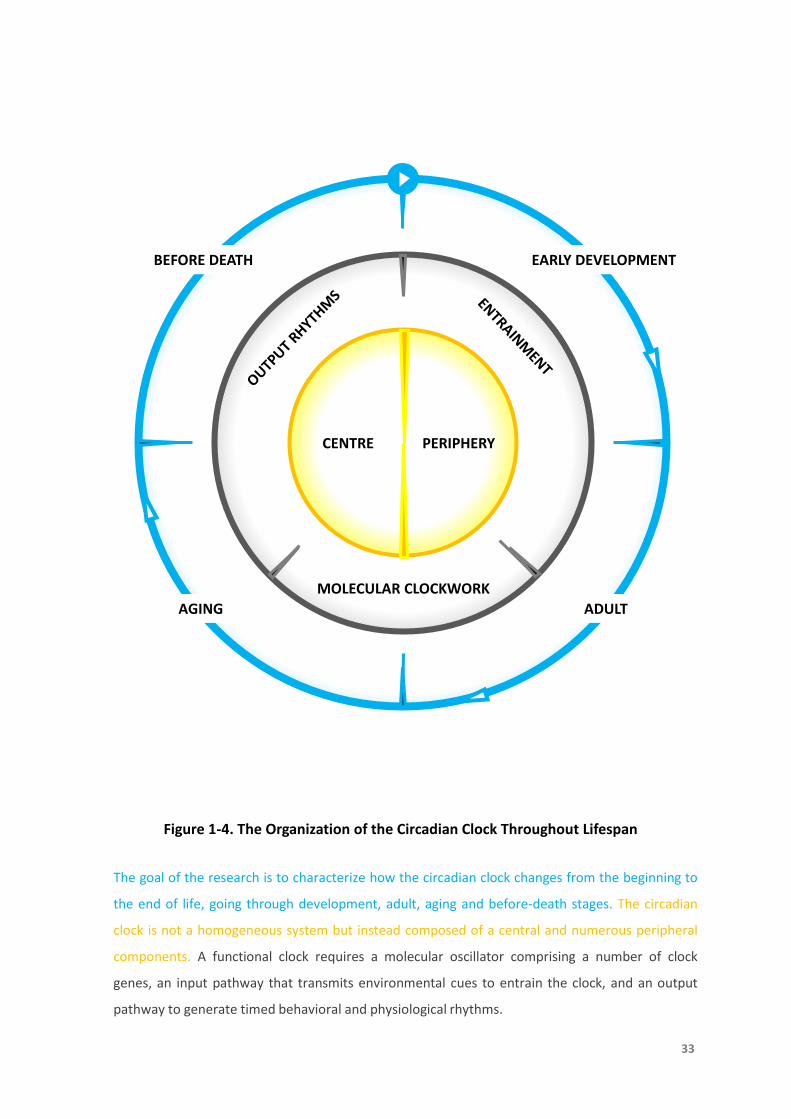
Figure 1-4. The Organization of the Circadian Clock Throughout Lifespan
CENTRE PERIPHERY
MOLECULAR CLOCKWORK
EARLY DEVELOPMENT
ADULTAGING
BEFORE DEATH
33
The goal of the research is to characterize how the circadian clock changes from the beginning to
the end of life, going through development, adult, aging and before-death stages. The circadian
clock is not a homogeneous system but instead composed of a central and numerous peripheral
components. A functional clock requires a molecular oscillator comprising a number of clock
genes, an input pathway that transmits environmental cues to entrain the clock, and an output
pathway to generate timed behavioral and physiological rhythms.

IntroductionoftheCircadianClock
34
1.7References
AbrahamU,Granada AE,Westermark PO,Heine M,Kramer A,Herzel H. Coupling governs
entrainment range of circadian clocks. Mol Syst Biol.2010Nov 30;6:438. DOI:
10.1038/msb.2010.92.
AgrawalP,HoulJH,GunawardhanaKL,LiuT,ZhouJ,ZoranMJ,HardinPE.Drosophilacryentrains
clocksinbodytissuestolightandmaintainspassivemembranepropertiesinanon-clockbody
tissue independent of light. Curr Biol.2017 Aug 21;27(16):2431-2441.e3. DOI:
10.1016/j.cub.2017.06.064.
Albrecht U. Timingtoperfection: thebiologyofcentralandperipheralcircadianclocks. Neuron.
2012Apr26;74(2):246-60.DOI:10.1016/j.neuron.2012.04.006.
Allada R,ChungBY. Circadian organization of behavior and physiology inDrosophila. AnnuRev
Physiol.2010;72:605-24.DOI:10.1146/annurev-physiol-021909-135815.
Allada R,White NE,So WV,Hall JC,Rosbash M. A mutant Drosophila homolog of mammalian
Clockdisruptscircadianrhythmsand transcription of period and timeless. Cell.1998 May
29;93(5):791-804.
AshmoreLJ,SathyanarayananS,SilvestreDW,EmersonMM,SchotlandP,SehgalA.Novelinsights
intotheregulationofthetimelessprotein.JNeurosci.2003Aug27;23(21):7810-9.
Ashmore LJ,Sehgal A. Afly'seyeviewofcircadianentrainment. J Biol Rhythms.2003
Jun;18(3):206-16.DOI:10.1177/0748730403018003003.
BalsalobreA.Clockgenesinmammalianperipheraltissues.CellTissueRes.2002Jul;309(1):193-9.
DOI:10.1007/s00441-002-0585-0.
BalsalobreA,BrownSA,MarcacciL,TroncheF,KellendonkC,ReichardtHM,SchützG,SchiblerU.
Resettingofcircadiantimeinperipheraltissuesbyglucocorticoidsignaling.Science.2000Sep
29;289(5488):2344-7.
Barber AF,Sehgal A. Coldtemperaturesfireupcircadianneurons. Cell Metab.2018 May
1;27(5):951-953.DOI:10.1016/j.cmet.2018.04.016.

IntroductionoftheCircadianClock
35
Brown SA,Azzi A. Peripheralcircadianoscillatorsinmammals. Handb Exp Pharmacol.
2013;(217):45-66.DOI:10.1007/978-3-642-25950-0_3.
BrownSA,Zumbrunn G,Fleury-Olela F,Preitner N,Schibler U. Rhythms of mammalian body
temperaturecansustainperipheralcircadianclocks.CurrBiol.2002Sep17;12(18):1574-83.
BuhrED,YooSH,TakahashiJS.Temperatureasauniversalresettingcueformammaliancircadian
oscillators.Science.2010Oct15;330(6002):379-85.DOI:10.1126/science.1195262.
Ceriani MF,Hogenesch JB,YanovskyM,Panda S,StraumeM,Kay SA. Genome-wide expression
analysisinDrosophilarevealsgenescontrollingcircadianbehavior. J Neurosci.2002 Nov
1;22(21):9305-19.
CermakianN,BoivinDB.Theregulationofcentralandperipheralcircadianclocksinhumans.Obes
Rev.2009Nov;10Suppl2:25-36.DOI:10.1111/j.1467-789X.2009.00660.x.
Chang DC. NeuralcircuitsunderlyingcircadianbehaviorinDrosophila melanogaster. Behav
Processes.2006Feb28;71(2-3):211-25.DOI:10.1016/j.beproc.2005.12.008.
CornélissenG,OtsukaK.Chronobiologyofaging:AMini-review.Gerontology.2017;63(2):118-128.
DOI:10.1159/000450945.
Damiola F,Le Minh N,Preitner N,Kornmann B,Fleury-Olela F,Schibler U. Restricted feeding
uncouples circadianoscillatorsinperipheraltissuesfrom thecentral pacemaker in the
suprachiasmaticnucleus.GenesDev.2000Dec1;14(23):2950-61.
DarlingtonTK,Wager-SmithK,CerianiMF,StaknisD,GekakisN,SteevesTD,WeitzCJ,TakahashiJS,
KaySA.Closingthecircadianloop:CLOCK-inducedtranscriptionofitsowninhibitorsperand
tim.Science.1998Jun5;280(5369):1599-603.
DeBruyneJP,Weaver DR,Reppert SM. Peripheral circadian oscillators requireCLOCK. Curr Biol.
2007Jul17;17(14):R538-9.DOI:10.1016/j.cub.2007.05.067.
Dissel S,Hansen CN,Özkaya Ö,Hemsley M,Kyriacou CP,Rosato E. The logic of circadian
organization in Drosophila. Curr Biol.2014 Oct 6;24(19):2257-66. DOI:
10.1016/j.cub.2014.08.023.

IntroductionoftheCircadianClock
36
Do MT,Yau KW. Intrinsicallyphotosensitiveretinal ganglion cells. Physiol Rev.
2010Oct;90(4):1547-81.DOI:10.1152/physrev.00013.2010.
Dubruille R,Emery P. A plastic clock: how circadian rhythms respond to environmental cues in
Drosophila.MolNeurobiol.2008Oct;38(2):129-45.DOI:10.1007/s12035-008-8035-y.
EderyI,ZwiebelLJ,DembinskaME,RosbashM.TemporalphosphorylationoftheDrosophilaperiod
protein.ProcNatlAcadSciUSA.1994Mar15;91(6):2260-4.
Erion R,King AN,Wu G,Hogenesch JB,Sehgal A. Neural clocks and Neuropeptide F/Y regulate
circadiangeneexpressioninaperipheralmetabolictissue.Elife.2016Apr14;5.pii:e13552.
DOI:10.7554/eLife.13552.
FroyO. Circadian rhythms, aging, and life span in mammals. Physiology (Bethesda).
2011Aug;26(4):225-35.DOI:10.1152/physiol.00012.2011.
GachonF,NagoshiE,BrownSA,RippergerJ,SchiblerU.Themammaliancircadiantimingsystem:
from gene expression to physiology. Chromosoma. 2004 Sep;113(3):103-12. DOI:
10.1007/s00412-004-0296-2.
GiebultowiczJM,HegeDM. Circadian clock in Malpighian tubules. Nature.1997Apr
17;386(6626):664.DOI:10.1038/386664a0.
GiebultowiczJM,Stanewsky R,Hall JC,Hege DM. Transplanted Drosophila excretory tubules
maintaincircadianclockcyclingoutofphasewiththehost.CurrBiol.2000Jan27;10(2):107-
10.
GrimaB,ChélotE,XiaR,RouyerF.Morningandeveningpeaksofactivityrelyondifferentclock
neurons of the Drosophila brain. Nature.2004 Oct 14;431(7010):869-73. DOI:
10.1038/nature02935.
Hao H,Allen DL,Hardin PE. A circadian enhancer mediates PER-dependent mRNA cycling in
Drosophilamelanogaster.MolCellBiol.1997Jul;17(7):3687-93.
HardinPE.ThecircadiantimekeepingsystemofDrosophila.CurrBiol.2005Sep6;15(17):R714-22.
DOI:10.1016/j.cub.2005.08.019.
HardinPE.Essentialandexpendablefeaturesof thecircadiantimekeepingmechanism.CurrOpin
Neurobiol.2006Dec;16(6):686-92.DOI:10.1016/j.conb.2006.09.001.

IntroductionoftheCircadianClock
37
HardinPE,HallJC,RosbashM.FeedbackoftheDrosophilaperiodgeneproductoncircadiancycling
ofitsmessengerRNAlevels.Nature.1990Feb8;343(6258):536-40.DOI:10.1038/343536a0.
HardinPE,Panda S. Circadiantimekeepingandoutputmechanismsinanimals. Curr Opin
Neurobiol.2013Oct;23(5):724-31.DOI:10.1016/j.conb.2013.02.018.
HegeDM,Stanewsky R,Hall JC,Giebultowicz JM. Rhythmic expression of a PER-reporter in the
Malpighian tubules of decapitated Drosophila: evidence for a brain-independent circadian
clock.JBiolRhythms.1997Aug;12(4):300-8.DOI:10.1177/074873049701200402.
Helfrich-Förster C. Neurobiology of the fruit fly's circadian clock. Genes Brain Behav. 2005
Mar;4(2):65-76.DOI:10.1111/j.1601-183X.2004.00092.x.
Helfrich-Förster C,Shafer OT,Wülbeck C,Grieshaber E,Rieger D,Taghert P. Development and
morphologyoftheclock-gene-expressinglateralneuronsofDrosophilamelanogaster.JComp
Neurol.2007Jan1;500(1):47-70.DOI:10.1002/cne.21146.
HendricksJC,FinnSM,PanckeriKA,ChavkinJ,WilliamsJA,SehgalA,PackAI.RestinDrosophilais
asleep-likestate.Neuron.2000Jan;25(1):129-38.
Hunter-Ensor M,Ousley A,Sehgal A. Regulation of the Drosophila protein timeless suggests a
mechanismforresettingthecircadianclockbylight.Cell.1996Mar8;84(5):677-85.
Kim EY,Edery I. Balance between DBT/CKlepsilon kinase and protein phosphatase activities
regulate phosphorylationandstabilityofDrosophilaCLOCKprotein. Proc Natl Acad Sci U S
A.2006Apr18;103(16):6178-83.DOI:10.1073/pnas.0511215103.
KlossB,Price JL,SaezL,Blau J,RothenfluhA,WesleyCS,YoungMW.TheDrosophilaclock
gene double-time encodesaproteincloselyrelatedtohuman caseinkinaseIepsilon. Cell.
1998Jul10;94(1):97-107.
KlossB,RothenfluhA,YoungMW,SaezL.Phosphorylationofperiod is influencedbycycling
physical associationsofdouble-time,period, andtimelessin theDrosophilaclock. Neuron.
2001Jun;30(3):699-706.
KoML,Shi L,Tsai JY,Young ME,Neuendorff N,Earnest DJ,KoGY. Cardiac-specific mutation of
Clockaltersthequantitativemeasurementsofphysicalactivitieswithoutchangingbehavioral
circadianrhythms.JBiolRhythms.2011Oct;26(5):412-22.DOI:10.1177/0748730411414170.

IntroductionoftheCircadianClock
38
KrishnanB,Dryer SE,Hardin PE. Circadian rhythms in olfactory responses of Drosophila
melanogaster.Nature.1999Jul22;400(6742):375-8.DOI:10.1038/22566.
KrishnanB,LevineJD,LynchMK,DowseHB,FunesP,HallJC,HardinPE,DryerSE.Anewrolefor
cryptochromeinaDrosophilacircadianoscillator.Nature.2001May17;411(6835):313-7.DOI:
10.1038/35077094.
ItoC,TomiokaK.HeterogeneityoftheperipheralcircadiansystemsinDrosophilamelanogaster:a
review.FrontPhysiol.2016Jan29;7:8.DOI:10.3389/fphys.2016.00008.
IvanchenkoM,StanewskyR,GiebultowiczJM.Circadianphotoreception inDrosophila: functions
ofcryptochromeinperipheralandcentralclock.JBiolRhythms.2001Jun;16(3):205-15.DOI:
10.1177/074873040101600303.
IzumoM,PejchalM,SchookAC,LangeRP,WalisserJA,SatoTR,WangX,BradfieldCA,TakahashiJS.
Differential effects of light and feeding on circadian organization of peripheralclocksin
aforebrainBmal1mutant.Elife.2014Dec19;3.DOI:10.7554/eLife.04617.
KruppJJ,BilleterJC,WongA,ChoiC,NitabachMN,LevineJD.Pigment-dispersingfactormodulates
pheromoneproduction in clock cells that influencemating inDrosophila.Neuron.2013 Jul
10;79(1):54-68.DOI:10.1016/j.neuron.2013.05.019.
Krupp JJ,Kent C,Billeter JC,Azanchi R,So AK,Schonfeld JA,Smith BP,Lucas C,Levine JD. Social
experience modifies pheromone expression and mating behavior in male Drosophila
melanogaster.CurrBiol.2008Sep23;18(18):1373-83.DOI:10.1016/j.cub.2008.07.089.
Lee C,Bae K,Edery I. PERandTIMinhibittheDNAbindingactivityof a Drosophila CLOCK-
CYC/dBMAL1heterodimer withoutdisruptingformationof theheterodimer: a basis for
circadiantranscription.MolCellBiol.1999Aug;19(8):5316-25.
Lehman MN,Silver R,Gladstone WR,Kahn RM,Gibson M,Bittman EL. Circadian rhythmicity
restoredbyneuraltransplant. Immunocytochemicalcharacterizationof thegraftand its
integrationwiththehostbrain.JNeurosci.1987Jun;7(6):1626-38.
Liu T,MaheshG,Houl JH,Hardin PE. Circadianactivatorsareexpresseddaysbefore they initiate
Clock functioninlatepacemaker neuronsfromDrosophila. J Neurosci. 2015 Jun
3;35(22):8662-71.DOI:10.1523/JNEUROSCI.0250-15.2015.

IntroductionoftheCircadianClock
39
McDonaldMJ,RosbashM,EmeryP.Wild-typecircadianrhythmicityisdependentoncloselyspaced
E boxes in theDrosophilatimelesspromoter. Mol Cell Biol.2001 Feb;21(4):1207-17. DOI:
10.1128/MCB.21.4.1207-1217.2001.
MyersEM,Yu J,Sehgal A. Circadian control of eclosion: interaction between a central and
peripheralclockinDrosophilamelanogaster.CurrBiol.2003Mar18;13(6):526-33.
Myers MP,Wager-Smith K,Rothenfluh-Hilfiker A,Young MW. Light-induced degradation of
TIMELESS andentrainmentof theDrosophilacircadian clock. Science.1996 Mar
22;271(5256):1736-40.
OguetaM,HardieRC,StanewskyR.Non-canonicalphototransductionmediatedsynchronizationof
theDrosophilamelanogastercircadianclockandretinal lightresponses.CurrBiol.2018Jun
4;28(11):1725-1735.e3.DOI:10.1016/j.cub.2018.04.016.
OishiK,SakamotoK,OkadaT,NagaseT,IshidaN.Humoralsignalsmediatethecircadianexpression
of rat period homologue (rPer2) mRNA inperipheraltissues. Neurosci Lett. 1998Nov
6;256(2):117-9.
O’NeillJS,ReddyAB.Circadianclocksinhumanredbloodcells.Nature.2011Jan27;469(7331):498-
503.DOI:10.1038/nature09702.
OsterH,MarondeE,AlbrechtU.Thecircadianclockasamolecularcalendar.ChronobiolInt.2002
May;19(3):507-16.
NawatheanP,RosbashM.ThedoubletimeandCKIIkinasescollaboratetopotentiateDrosophila
PERtranscriptionalrepressoractivity.MolCell.2004Jan30;13(2):213-23.
Pendergast JS,Niswender KD,Yamazaki S. Tissue-specific function of Period3 in circadian
rhythmicity.PLoSOne.2012;7(1):e30254.DOI:10.1371/journal.pone.0030254.
Peng Y,Stoleru D,Levine JD,Hall JC,Rosbash M. Drosophilafree-running rhythms require
intercellular communication. PLoS Biol.2003 Oct;1(1):E13. Epub 2003 Sep 15. DOI:
10.1371/journal.pbio.0000013.
Peschel N,Helfrich-Förster C. Settingtheclock - bynature:circadianrhythmin the fruitfly
Drosophilamelanogaster. FEBS Lett.2011 May 20;585(10):1435-42. DOI:
10.1016/j.febslet.2011.02.028.

IntroductionoftheCircadianClock
40
Plautz JD,Kaneko M,Hall JC,Kay SA. Independentphotoreceptive circadian clocks throughout
Drosophila.Science.1997Nov28;278(5343):1632-5.
PriceJL,BlauJ,RothenfluhA,AbodeelyM,KlossB,YoungMW.double-timeisanovelDrosophila
clockgenethatregulatesPERIODproteinaccumulation.Cell.1998Jul10;94(1):83-95.
Ralph MR,Foster RG,Davis FC,Menaker M. Transplantedsuprachiasmatic nucleus determines
circadianperiod.Science.1990Feb23;247(4945):975-8.
RefinettiR.Entrainmentofcircadianrhythmbyambienttemperaturecyclesinmice.JBiolRhythms.
2010Aug;25(4):247-56.DOI:10.1177/0748730410372074.
RennSC,ParkJH,RosbashM,HallJC,TaghertPH.Apdfneuropeptidegenemutationandablation
ofPDFneuronseachcausesevereabnormalitiesofbehavioralcircadianrhythmsinDrosophila.
Cell.1999Dec23;99(7):791-802.
Richards J,Gumz ML. Advancesinunderstandingtheperipheralcircadian clocks. FASEB J.
2012Sep;26(9):3602-13.DOI:10.1096/fj.12-203554.
Rosato E,Kyriacou CP. Analysisoflocomotor activityrhythmsinDrosophila. Nat Protoc.
2006;1(2):559-68.DOI:10.1038/nprot.2006.79.
Rosato E,KyriacouCP. Sleep, arousal, and rhythms in flies. ProcNatl Acad SciU SA.2008Dec
16;105(50):19567-8.DOI:10.1073/pnas.0811124106.
SakamotoK,NagaseT,FukuiH,HorikawaK,OkadaT,TanakaH,SatoK,MiyakeY,OharaO,KakoK,
IshidaN.Multitissuecircadianexpressionofratperiodhomolog(rPer2)mRNAisgovernedby
themammaliancircadianclock, the suprachiasmaticnucleusin thebrain. JBiolChem.1998
Oct16;273(42):27039-42.
SehgalA,OusleyA,Hunter-EnsorM.Controlofcircadianrhythmsbyatwo-componentclock.Mol
CellNeurosci.1996Mar;7(3):165-72.DOI:10.1006/mcne.1996.0013.
Sehgal A,Rothenfluh-Hilfiker A,Hunter-Ensor M,Chen Y,Myers MP,Young MW. Rhythmic
expression of timeless: a basisforpromotingcircadiancyclesinperiodgene autoregulation.
Science.1995Nov3;270(5237):808-10.

IntroductionoftheCircadianClock
41
SelchoM,MillánC,Palacios-MuñozA,RufF,UbilloL,ChenJ,BergmannG,ItoC,SilvaV,Wegener
C,EwerJ.CentralandperipheralclocksarecoupledbyaneuropeptidepathwayinDrosophila.
NatCommun.2017May30;8:15563.DOI:10.1038/ncomms15563.
ShaferOT,RosbashM,TrumanJW.Sequentialnuclearaccumulationoftheclockproteinsperiod
and timeless in thepacemakerneuronsofDrosophilamelanogaster. J Neurosci.2002 Jul
15;22(14):5946-54.DOI:20026628.
ShangY,Griffith LC,RosbashM. Light-arousal and circadianphotoreception circuits intersect at
thelargePDFcellsof the Drosophila brain. Proc Natl Acad Sci U S A.2008 Dec
16;105(50):19587-94.DOI:10.1073/pnas.0809577105.
Shaw P. Awakeningto thebehavioralanalysisofsleepinDrosophila. J Biol Rhythms.2003
Feb;18(1):4-11.DOI:10.1177/0748730402239672.
ShearmanLP,JinX,LeeC,ReppertSM,WeaverDR.TargeteddisruptionofthemPer3gene:subtle
effectsoncircadianclockfunction.MolCellBiol.2000Sep;20(17):6269-75.
Silver R,LeSauter J,Tresco PA,Lehman MN. Adiffusiblecouplingsignalfrom the transplanted
suprachiasmaticnucleuscontrollingcircadian locomotorrhythms. Nature.1996 Aug
29;382(6594):810-3.DOI:10.1038/382810a0.
Son GH,Chung S,Kim K. Theadrenalperipheralclock:glucocorticoidand thecircadian timing
system.FrontNeuroendocrinol.2011Oct;32(4):451-65.DOI:10.1016/j.yfrne.2011.07.003.
Stanewsky R,Lynch KS,Brandes C,Hall JC.Mapping of elements involved in regulating normal
temporalperiodandtimelessRNA expression patterns in Drosophila melanogaster.J Biol
Rhythms.2002Aug;17(4):293-306.DOI:10.1177/074873002129002609.
Stephan FK,Zucker I. Circadian rhythmsindrinking behaviorandlocomotor activity of rats are
eliminatedbyhypothalamiclesions.ProcNatlAcadSciUSA.1972Jun;69(6):1583-6.
StokkanKA,YamazakiS,TeiH,SakakiY,MenakerM.Entrainmentofthecircadianclockintheliver
byfeeding.Science.2001Jan19;291(5503):490-3.DOI:10.1126/science.291.5503.490.
Stoleru D,Peng Y,Agosto J,Rosbash M. Coupledoscillatorscontrolmorningandevening
locomotor behaviourofDrosophila. Nature.2004 Oct 14;431(7010):862-8. DOI:
10.1038/nature02926.

IntroductionoftheCircadianClock
42
StoleruD,PengY,NawatheanP,RosbashM.A resettingsignalbetweenDrosophilapacemakers
synchronizes morning andeveningactivity. Nature.2005 Nov 10;438(7065):238-42. DOI:
10.1038/nature04192.
Tahara Y,Kuroda H,Saito K,Nakajima Y,Kubo Y,Ohnishi N,Seo Y,Otsuka M,Fuse Y,Ohura Y,
Komatsu T,Moriya Y,Okada S,Furutani N,Hirao A,Horikawa K,Kudo T,Shibata S. In vivo
monitoringofperipheralcircadianclocksinthemouse.CurrBiol.2012Jun5;22(11):1029-34.
DOI:10.1016/j.cub.2012.04.009.
TanoueS,Krishnan P,Krishnan B,Dryer SE,Hardin PE. Circadian clocks in antennal neurons are
necessaryandsufficientforolfactionrhythmsinDrosophila.CurrBiol.2004Apr20;14(8):638-
49.DOI:10.1016/j.cub.2004.04.009.
ValloneD,LahiriK,DickmeisT,FoulkesNS.Starttheclock!Circadianrhythmsanddevelopment.
DevDyn.2007Jan;236(1):142-55.DOI:10.1002/dvdy.20998.
VaninS,BhutaniS,MontelliS,MenegazziP,GreenEW,PegoraroM,SandrelliF,CostaR,Kyriacou
CP.UnexpectedfeaturesofDrosophilacircadianbehaviouralrhythmsundernaturalconditions.
Nature.2012Apr4;484(7394):371-5.DOI:10.1038/nature10991.
Veleri S,Brandes C,Helfrich-Förster C,Hall JC,Stanewsky R. Aself-sustaining,light-entrainable
circadianoscillatorintheDrosophilabrain.CurrBiol.2003Oct14;13(20):1758-67.
Wang GK,Ousley A,Darlington TK,Chen D,Chen Y,Fu W,Hickman LJ,Kay SA,Sehgal A.
Regulationofthecyclingoftimeless(tim)RNA.JNeurobiol.2001Jun5;47(3):161-75.
WelshDK,YooSH,LiuAC,TakahashiJS,KaySA.Bioluminescenceimagingofindividualfibroblasts
revealspersistent,independentlyphasedcircadianrhythmsofclockgeneexpression.CurrBiol.
2004Dec29;14(24):2289-95.DOI:10.1016/j.cub.2004.11.057.
XuK,ZhengX,SehgalA.Regulationoffeedingandmetabolismbyneuronalandperipheralclocks
inDrosophila.CellMetab.2008Oct;8(4):289-300.DOI:10.1016/j.cmet.2008.09.006.
YagitaK,TamaniniF,vanDerHorstGT,OkamuraH.Molecularmechanismsofthebiologicalclock
in cultured fibroblasts. Science.2001Apr 13;292(5515):278-81. DOI:
10.1126/science.1059542.

IntroductionoftheCircadianClock
43
YamazakiS,NumanoR,AbeM,HidaA,TakahashiR,UedaM,BlockGD,SakakiY,MenakerM,Tei
H.Resettingcentralandperipheralcircadianoscillatorsinthetransgenicrats.Science.2000
Apr28;288(5466):682-5.
Yao Z,Bennett AJ,Clem JL,Shafer OT. The Drosophila clock neuron network features diverse
couplingmodesandrequiresnetwork-widecoherenceforrobustcircadianrhythms.CellRep.
2016Dec13;17(11):2873-2881.DOI:10.1016/j.celrep.2016.11.053.
Yao Z,Shafer OT. TheDrosophilacircadian clockis a variably coupled network of multiple
peptidergicunits.Science.2014Mar28;343(6178):1516-20.DOI:10.1126/science.1251285.
YooSH,YamazakiS,LowreyPL,ShimomuraK,KoCH,BuhrED,SiepkaSM,HongHK,OhWJ,YooOJ,
Menaker M,Takahashi JS. PERIOD2::LUCIFERASEreal-time reporting of circadian dynamics
revealspersistentcircadianoscillatorsinmouseperipheraltissues.ProcNatlAcadSciUSA.
2004Apr13;101(15):5339-46.DOI:10.1073/pnas.0308709101.
Yoshii T,Hermann-Luibl C,Helfrich-Förster C. Circadianlight-input pathways in Drosophila.
CommunIntegrBiol.2015Dec4;9(1):e1102805.DOI:10.1080/19420889.2015.1102805.
YuW,HardinPE.CircadianoscillatorsofDrosophilaandmammals. JCellSci.2006Dec1;119(Pt
23):4793-5.DOI:10.1242/jcs.03174.
YuW,ZhengH,HoulJH,DauwalderB,HardinPE.PER-dependentrhythmsinCLKphosphorylation
andE-boxbindingregulatecircadiantranscription.GenesDev.2006Mar15;20(6):723-33.DOI:
10.1101/gad.1404406.
Zeng H,Qian Z,Myers MP,Rosbash M. Alight-entrainmentmechanismfor the Drosophila
circadianclock.Nature.1996Mar14;380(6570):129-35.DOI:10.1038/380129a0.
ZerrDM,HallJC,RosbashM,SiwickiKK.Circadianfluctuationsofperiodproteinimmunoreactivity
intheCNSandthevisualsystemofDrosophila.JNeurosci.1990Aug;10(8):2749-62.

王维
终南别业

CHAPTER 2
Development
44

Development of the Molecular Circadian
Clock and its Light Sensitivity in
Drosophila melanogaster

TheDevelopmentoftheCircadianClock
46
Thedatafromthischapterhavebeenpublished**(pleaseseeAppendixfordetails).
**ZhaoJ,WarmanGR,StanewskyR,CheesemanJF. DevelopmentoftheMolecularCircadian
ClockandItsLightSensitivityinDrosophilaMelanogaster. JBiolRhythms.2019Mar
18:748730419836818.DOI:10.1177/0748730419836818.

TheDevelopmentoftheCircadianClock
47
2.1Abstract
Althoughmuch isknownabout themolecularmechanismof thecircadianclockamong
diverseorganisms,thequestionsofwhen,where,andhowdailyrhythmsemergein
earlydevelopmentarestilllargelyunanswered.Weaimedtostudywhenthecircadian
clockand its light sensitivity start todevelop inDrosophilamelanogaster.Here the
initialexpressionpatternofthekeyclockgeneperiodwasrecordedinDrosophilafrom
embryotoadultinvivo,usingtransgenicluciferasereporters.PERIODexpressioninthe
presumptive central clock dorsal neurons started to oscillate in the embryos in
constantdarkness.Inbehaviouralexperimentsasingle12-hourlightpulsegivenduring
the second half of embryonic stage synchronized adult locomotor activity rhythms
implying the early development of the entrainment mechanism. Together these
findingssuggestthatthecentralclockisalreadyfunctionalduringembryogenesis.In
contrasttocentralbrainexpression,PERIODintheperipheraltissuesincreasedduring
the embryonic stage and peaked during the pupae without showing circadian
oscillations.Itsrhythmicexpressiononlybeganintheadult.Thus,weconcludethat
cyclic expression of PERIOD in the central clock neurons starts in the embryos,
presumablyinthedorsalneurons1(DN1)ortheirprecursors.Itisnotuntilshortlyafter
eclosion when cyclic and synchronized expression of PERIOD in peripheral tissues
throughouttheanimalcommences.

TheDevelopmentoftheCircadianClock
48
2.2Introduction
Anumberofstudiesinthepastfewdecadeshavebeentryingtorevealthemysteryofthe
developmentofthecircadianclockinDrosophilaandmammals.Ofparticularinterest
isdeterminingthetimingofwhenthecentralmolecularclockstartstooscillateand
when the peripheral clocks are synchronized throughout the whole animal.
Developmentofthemajorcomponentsofthecircadianclock,includinginputpathway,
molecularclockwork,andoutputrhythmshasbeeninvestigated.
Beingan importantdownstreamoutputofthecircadianclock,behavioural rhythmsare
primarily studied.Rhythmsof locomotor activityhavebeenwell described in adult
Drosophila,andithasbeenproposedthatbehaviouralrhythmsmayexistinpre-adult
developmental stages. Forexample,a clock-drivenphotophobic responsehasbeen
describedinthelarvae[Mazzonietal.,2005],althoughthelight-avoidancebehaviour
itselfdoesnotfollowacircadianpattern.Inaddition,despitethefactthateclosion(the
emergenceofanadultfromthepupalcase)occursonlyonceinasinglefly’slifetime,
the multiple events within a population follow a circadian pattern [Skopik and
Pittendrigh, 1967]. Furthermore, a few studies have indicated that environmental
factorsadministratedduringpre-adultstagescanaltertheadultcircadianbehaviour
andmolecularstate,evenafterthetreatmentisterminated.Forinstance,abrieflight
pulseorlight-dark(LD)cyclescansynchronizeeclosionrhythmsaswellaslocomotor
activityrhythmsinadultwhenprovidedasearlyasthefirst-instarlarvalstage(butnot

TheDevelopmentoftheCircadianClock
49
duringembryonicstage),eventhoughthefliesareotherwisekeptinconstantdarkness
(DD)[Brett,1955;Sehgaletal.,1992].Subsequently,ithasbeendemonstratedthata
10-minute light pulseor different light regimen in larvae can affect the adult free-
runningperiodorshiftthephaseofadultrhythms[Matsumotoetal.,1997;Kanekoet
al.,2000;Sheebaetal.,2002].Asidefromlight,otherstimulisuchasethanolexposure
during larval development cause sustainable shortening of the adult free-running
period[Seggioetal.,2012;Ahmadetal.,2013].Theresultsfromtheabovestudiesof
pre-entrainedadultbehaviouralrhythmsimplythatcircadianclockispresentfromthe
firstlarvalstage,butwhetheritisalreadyfunctionalatearlierdevelopmentisunclear.
Drosophilalarvaepossesstheabilitytoshiftthecircadianclockinthepacemakerneurons
withalightpulse,whichcanthusentraineclosionrhythms,developmentalhormones
andadultbehaviour [KeeneandSprecher,2012].Presumably, the larvaeshouldbe
equippedwithanefficientinputpathwaythatcantransmitenvironmentalsignalsto
theclock.Suchinputpathwayisrequirednotonlyfortheentrainmentofthecircadian
clock,butalsoforrapidinitiationofbehaviours(suchasthephotophobicresponse).It
hasbeenreportedthatthelarvalvisualsystemiscapableofbothfunctions[Keeneet
al., 2011], and the larval pacemaker neurons also show corresponding capability.
Additionally, previous works have revealed sensory and central brain neurons
necessaryforpre-adultvisualreactions,andthespecificlightinputsforeachsubgroup
ofclockneuronshavebeenidentifiedinDrosophilalarvae[Klarsfeldetal.,2011].

TheDevelopmentoftheCircadianClock
50
Thevisualsystemispartoftheinputpathwaysforlightsignalsenteringintothecircadian
clockofthebrain.Thelarvalopticnerve(Bolwig’snerve)whichprojectstolarvalclock
neurons is thought tobe theorgan that receivesphotic input from the larval eyes
[Kaneko et al., 1997]. It has been revealed later that the contact between larval
Bolwig’snerveand clockneuronsexists as early as embryonic stage [Malpel et al.,
2002].Moreover,asidefromthelarvalvisualorgans,CRYPTOCHROMEisbelievedto
cruciallyparticipateinlarvalcircadianphotoreception,makingtheentrainmentofthe
circadian clock possible [Kaneko et al., 2000; Malpel et al., 2004]. The pigment-
dispersingfactor(PDF)neuropeptidesignallingisalsosuggestedtobenecessaryinthe
light inputpathwayat thisstage[Picotetal.,2009].Subsequentlyduringthepupal
stage, visual fibres that come from the Hofbauer-Buchner eyelet, a putative
photoreceptiveorganoftheadultcircadianclock,arefoundtobealreadycontacting
the pacemaker neurons [Malpel et al., 2002]. In addition, a pair of giant visual
projection neurons which have special properties in light sensitivity and rhythmic
activities are also found in the pupae [Yan et al., 2013]. We thus infer that the
establishmentoftheinputpathwaytothecentralclockpossiblystartsinembryonic
stage,thengoesthroughrapiddevelopmentduringlarvalandpupalstages.
Existingevidenceontheoutputandinputpathwayssupportsthepresenceofthecircadian
clock during early development, further interests are therefore placed on the
emergence of the clockmachinery. This includes the anatomy basis of pacemaker
neuronsandimportantly,theircyclicexpressionofcoreclockgenes.In1997,circadian
pacemakerneuronshavebeenidentifiedinthe4-hour-oldfirst-instarlarvae,withtheir

TheDevelopmentoftheCircadianClock
51
roles incontrollingrhythmicprocesses in larvae,pupae,andadult [Helfrich-Förster,
1997].Combiningwithfindingsoflaterstudies,itshowsthatintherelativelysimple
larval brain, there are nine clock neurons per hemisphere, including five lateral
neurons (LN), fourofwhichexpress thePDFneuropeptide,and twopairsofdorsal
neurons(DN1andDN2)[Klarsfeldetal.,2011].Amongtheseneurons,theLNarethe
onlycellsthatmaintainastrongcyclinginclockgeneexpressionfromfirst-instarlarvae
onward, thenthroughoutmetamorphosisand intoadulthood[Kanekoetal.,1997].
Further,thePDF-expressingLNareimplicatedtobethemasterpacemakerneurons
andsubsequentlyrepresentthesmallventrallateralneurons(s-LNv)oftheadultbrain
[Valloneetal.,2007].Nonetheless,theremaininggroupsofmainpacemakerneurons
necessary forcircadianmechanism,suchas the largeventral lateralneurons (l-LNv)
and dorsal lateral neurons (LNd), only differentiate duringmid-pupae (Figure 2-1A)
[Helfrich-Förster et al., 2007]. This finding implies that a mature circadian clock
composedofthewholepackageofpacemakerneuronsmaynotexistbeforeeclosion.
Furthermore,despitesimilaritiesbetweentheadultandlarvalmolecularoscillators,as
wellastheneuronalprojections[KanekoandHall,2000],adultandlarvaemaydiffer
substantiallyinthecomplexityandorganizationoftheirpacemakerneuronalcircuits
[Nitabachetal.,2005].
Thedrivingforceofthecircadianclockisthemolecularoscillator,whichisbasedonthe
transcription-translationfeedbackloopsofseveralclockgenes,includingperiod(per)
andtimeless(tim)(refertoChapterOne,Section1.3).WorkfromKanekoin1997has
demonstratedthatcyclicexpressionofPERIOD(PER)andTIMELESS(TIM)occursinthe

TheDevelopmentoftheCircadianClock
52
Drosophila larval centralnervous system (CNS) in several groupsofneurons (s-LNv,
DN1a,andDN2),bothinLDcyclesandinfollowingDDconditions(Figure2-1B)[Kaneko
et al., 1997]. The technique of immunohistochemistry used in this study requires
animalstobesacrificedeverysixhoursanddataatthesametimepointtobepooled
fromdifferentbrainsamples[Kanekoetal.,1997].Thequestionremainswhetherthis
technique has sufficient resolution to represent the key parameters of circadian
oscillationsfaithfully.
Evenbeforelarvaldevelopment,ithasbeenrevealedthatasignificantexpressionofper
mRNAexists inmid-latedevelopingembryos(after11-hourofembryogenesis).This
earlyexpressionofperislimitedtotheCNSandlocalisedwithinthebrainandventral
ganglia[Jamesetal.,1986].ExpressionoftimisalsodetectedintheembryonicCNS.
Thenumber, location,originand lineageofper and tim expressingcellshavebeen
definedthroughoutembryogenesisinarecentstudy[Ruizetal.,2010].CLOCK(CLK)is
showntobeinitiallyexpressedinpresumptives-LNv,DN1,andDN2atembryonicstage,
andafewhoursaftertheoccurrenceofCLK,PERaccumulates inallCLK-expressing
cells except presumptiveDN2 during late embryos (Figure 2-1C) [Houl et al., 2008;
Glossopetal.,2014].

Figure 2-1. Development of Central Clock Neurons at Embryonic, Larval and Pupal Stages in Drosophila
• l-LNv and LNd differentiate at pupal brain [Helfrich-Förster et al., 2007]
• s-LNv, DN1 and DN2 exist at larval brain [Kaneko et al., 1997]
• s-LNv and DN1 develop at embryonic brain [Houl et al., 2008]
Adult Drosophila brain and all groups of central clock neurons, including dorsal neurons (DN1, DN2,
and DN3), lateral posterior neurons (LPN), dorsal lateral neurons (LNd), large ventral lateral
neurons (l-LNv), and small ventral lateral neurons (s-LNv) are shown as dots in corresponding
colour. Various clusters of neurons develop at different stages shown as the (A) pupa, (B) larva
and (C) embryo.
DN1
DN2 DN3
s-LNv
l-LNv
LNd
LPN
53
Brain
A
B
C
De
velo
pm
en
tal Ord
ers
C
B
A
A
C
Central Clock Neurons
Developmental Stages

TheDevelopmentoftheCircadianClock
54
ExpressionofcoreclockgenesandtheirproteinproductsfoundintheembryonicCNSof
Drosophilasuggeststhatbrainoscillatorneuronsmaybegintheirdevelopmentduring
embryogenesis.However,thefactthatembryonicdevelopmentonlytakes24hoursat
25°C (typical environmental rearing temperature) makes it difficult to prove the
existenceofacircadianpatternofclockgeneexpression[Jamesetal.,1986;Houlet
al.,2008;Ruizetal.,2010;Glossopetal.,2014;Liuetal.,2015].
Thenegativecorrelationbetweentemperatureanddevelopmentaltimingiswelldefined
in Drosophila [Kuntz and Eisen, 2014]. This offers the possibility to extend the
embryonic stage in order to potentially reveal circadian rhythmicity. Higher
temperaturespeedsupdevelopmentandlowertemperatureslowsitdownwithina
rangeof 16°C to 29°C [Economos and Lints, 1986]. At 16°C the embryonic stage is
prolongedapproximatelydoublythedevelopmentaltimecomparedto25°C[Ghoshet
al.,2013].Circadianclocks,beingtemperaturecompensated,donotshowachangein
periodofmolecularandbehaviouralrhythmswithafluctuationintemperature[Hall,
1997].Therefore,manipulationoftemperaturecanbeusedasatooltoexaminethe
presenceof a functionalmolecular clock in the early embryos beyond the 24-hour
temporalconstraint.
Inmammals,thedevelopmentoftheclocksystemseemstobemorecomplicatedbutin
general, still shares a similar rulewithDrosophila. Overt circadian rhythms usually
occuratpostnatalages,suchastheday-nightchangesofcorticosterone,rest-activity
and body temperature [Weinert, 2005]. In rodents, it is weeks after birth when

TheDevelopmentoftheCircadianClock
55
circadianoutputstarttoemerge.Forexample,rhythmicproductionofmelatonin is
firstdetectedattheendofthefirstweekofpostnataldevelopment[Valloneetal.,
2007].Inprimates,includinghumans,certaincircadianrhythmscanevenbedetected
prenatally,suchasrest-activity,breathing,heartrateandurineproduction[Weinert,
2005].
Development of the central endogenous pacemaker suprachiasmatic nucleus (SCN) is
presumablythebasisofclockmaturityinmammals.Itgoesthroughagradualprogress
ofstructureformation,neurogenesisandsynaptogenesis[Weinert,2005;Valloneet
al.,2007;Christetal.,2012].ThestructureofSCNisformedduringthethirdwaveof
hypothalamicneurongenerationanddevelopsasacomponentoftheperiventricular
cellgroup.Thesubdivisionofventrolateralanddorsomedialregionshappens inthe
embryos[Weinert,2005].Neurogenesisstartsinmid-lateembryonicstage(60-80%of
gestation inrodents)whenSCNneuronsstarttodevelopanddifferentiate,andthe
adult morphology of the SCN is established at early postnatal stage [Bedont and
Blackshaw,2015].Synaptogenesis intheSCNprogressesslowlyduring lateprenatal
developmentand ismainlyapostnatalprocess [Weinert,2005].Synapse formation
indicatestheeffectivecommunicationandsynchronizationwithintheSCNneuronal
network,makingrobustandcoherentcircadianrhythmicitypossible.
Despite that a fully-developed SCN structure as well as many overt rhythms presents
postnatally,thefunctionaloutputofthecentralclockseemstoexistprenatally.For
instance,circadianactivityintheSCNcanbedetectedalreadyinutero.ThefoetalSCN

TheDevelopmentoftheCircadianClock
56
showsrhythmsinmetabolism[ReppertandSchwartz,1984],neuropeptidearginine
vasopressin(AVP)mRNAexpression[ReppertandUhl,1987],andneuronalfiringrate
[ShibataandMoore,1987].Interestingly,itishypothesizedthatsuchSCNoutputmay
evenemergebeforethecompletematurationofcoreclockmechanism[Valloneetal.,
2007].
ThemolecularclockworkintheSCNstartstodevelopataround90%ofgestationinrodents
and primates. Intrinsic circadian rhythms in some clock genes are detected during
foetalages,suchasPeriod1(Per1)inmouseandratSCN,withothergenesoscillating
later,aroundbirthorshortlyafter[Christetal.,2012;Arellanes-Liceaetal.,2014].For
instance,circadianprofilesofPeriod2(Per2),Bmal1andClock,areallexpressedinthe
ratSCNprenatallywithoutdetectablecircadianrhythm[Sládeketal.,2004;Weinert,
2005]. Genes in the positive arm seem to become rhythmic later than that in the
negativearm.Clockisparticularlynotableinthisregardbecauseitscyclingonlybegins
aftertenpostnataldaysinbothmouseandrat[Sládeketal.,2004].Takentogether,
thoughdetailsvarysomewhatamongdifferentreports[BedontandBlackshaw,2015],
datashowthattheendogenousmolecularoscillatorsintheSCNstartstodevelopfrom
thelatefoetalstageandmaturesgraduallyduringthepostnatalage[Christetal.,2012].
Itiswellacceptedthattheontogenesisofperipheraloscillatorshappenslaterthanthatin
the central pacemaker. Detection of clock genes in the foetal rat liver by in situ
hybridizationhasnotbeenrevealedanyrhythmsexceptforalowamplitudeoscillation
in Cryptochrome 1 expression before birth. Themajormolecular clock in the liver

TheDevelopmentoftheCircadianClock
57
maturesslowlyduringpostnataldevelopment[Sládeketal.,2007].Therhythmicityof
Per1 andBmal1 expression startsa fewdaysafterbirth in theheart, and thedaily
rhythmsofPer1andPer2expressionappearmuchlaterinthecerebralcortex[Weinert,
2005].Itseemslikelythatthedevelopmentofclockmechanismsoccursindependently
indifferentorgansatvarioustiming[Weinert,2005],mainlypostnatally[Valloneetal.,
2007].
Oneinterestingfindingisthatalthoughinvivostudiesshownocircadianrhythmsofclock
geneexpression, ineitherwholeembryosor individual tissues, rhythmshavebeen
detectedinembryonictissuesinvitro[Dolatshadetal.,2010].Ithasbeenshownthat
embryonictissueshavethecapacitytoexpressactivecircadianregulatorycyclesofkey
clock genes by the continuous assay of bioluminescence. The implication is that
individualcellsveryearlyindevelopmentmightalreadyexpressoscillations[Dolatshad
etal.,2010].Thereasonwhycircadiancyclesarenotevidentatthetissuelevelmay
duetothedesynchronyamongdifferentcells.Synchronizationofperipheraloscillators
is presumed to take place later in development in a tissue-dependent manner
[Arellanes-Liceaetal.,2014].
It isan important fact that thegenerationof foetal rhythmsatboththemolecularand
output level is intrinsic and is not a direct consequence of maternal rhythmicity
[Weinert,2005].However,itisalsotruethatcircadianphysiologyduringfoetallifeis
dependent on non-photicmaternal cues via the placental system. Foetal SCN and
possiblyperipheralclocksareentrainedbyrhythmicallydeliveredmaternalmessages,

TheDevelopmentoftheCircadianClock
58
includingdopamine,melatonin,stresssignalsandmaternalbehaviour[Arellanes-Licea
etal.,2014]. It isstillamatterofdebatewhetherrhythmicoutputsignalsfromthe
foetalSCNarepowerfulenoughtoentrainperipheraloscillators[Christetal.,2012].
Speakingoflightentrainment,ithasnodirecteffectonthefoetalSCNmetabolicactivity
[Reppert and Schwartz, 1983] and this is consistent with the observations that
intrinsicallyphotoreceptiveretinalganglioncells(ipRGCs)andlightinputpathwayinto
theSCNonlydeveloppostnatally[Weinert,2005;Arellanes-Liceaetal.,2014].Inthe
earlypostnatalstage,theintensesocialinteractionwiththemotheraswellasfeeding
still provide the primary signals for the developing circadian system. Until after
weaning,theLDcyclestakeoverandbecomethemainZeitgeber[Weinert,2005].The
besttimingforthistransitiontobefinishediswhenthelightinputpathwayandthe
endogenousrhythmgeneratorintheSCNhavefullydeveloped[Christetal.,2012].

TheDevelopmentoftheCircadianClock
59
2.3Objectives
Beforetheworkdescribedhere,ithasnotbeenestablishedwhetheranentrainableclock
exits during the embryonic development ofDrosophila. In this study,we aimed to
determinewhenthecentralandperipheralcomponentsofthecircadianclockstartto
oscillate(Figure2-2).Firstofall,PERexpressionwasmeasuredcontinuouslyfromthe
beginning of the embryonic stage in individual fruit flies at 25°C and 16°C, using
luciferasereporters.Secondly,theresponseofadultlocomotoractivitytotimedlight
treatment during pre-adult stageswas investigated. Thirdly,while it is known that
establishmentofbehaviouralrhythmsinindividualfruitfliesdoesnotrequireexposure
to light during development,we investigatedhere if this is also true formolecular
rhythmsofclockgeneexpression.

Embryo
Larva
Pupa
12h12h
4d
4d
12h12h
12h
12h
WHEN
Embryo
Larva
Pupa
Adult
Light Response
Overt Rhythms
Central Gene Expression and Oscillation
Peripheral Gene Expression and Oscillation?
×2
×2
16℃
25℃
Figure 2-2. Life Cycle of Drosophila and Existing Findings on Clock Development
Start Point
Drosophila go through embryo (approx. 24 hours), larva (approx. 4 days), and pupa (approx. 4
days) as early developmental stages at 25℃, before eclose into adult. Development in Drosophila
is temperature-dependent and it doubles the time to develop at 16℃. Cyclic clock gene
expression and light-sensitivity of the central clock neurons have been reported in larvae, yet the
emergence timing of peripheral oscillators has not been specified.
60
De
velo
pm
en
tal O
rde
rs
h: hourd: day

TheDevelopmentoftheCircadianClock
61
2.4Methods
2.4.1ExperimentalDesign
Figure2-3.ExperimentalDesignoftheDevelopmentStudy
In order to find out the emergence timing of light-sensitivity in Drosophila circadian clock,
behaviouralrhythmsofwildtypeflieswhichhadbeentreatedbylightorkeptinDD(whitebars
indicatelightsonandblackbarsindicatelightsoff)duringearlydevelopmentweremeasuredby
DAMsystemfrompupalstageonwardsat25°Cand16°Crespectively.Tofindouttheemergence
timing of molecular oscillations, clock gene expression was measured in transgenic flies by
bioluminescenceassayfromembryonicstageonwardsunderDDorLDcyclesat25°Cand16°C.

TheDevelopmentoftheCircadianClock
62
2.4.2AnimalModel
Drosophilamelanogaster(fruitfly)isanidealmodelforstudyingthephenomenaofthe
circadianclockaswellasitstickingmechanism.Drosophilahasbeenestablishedasan
excellenttoolforgeneticstudiesoverahundredyears,andtheuseofDrosophilafor
theanalysisofcircadianrhythmsandmolecularclockworkhasbeenwellestablished
[Cirelli,2006;LeGlouetal.,2012].Firstofall,thecircadianclockislargelyconserved
inorganismsasdiverseasbacteria,insects,mammalsandhumans[Pandaetal.,2002;
Looby and Loudon, 2005; Yu andHardin, 2006]. Secondly, fruit flies exhibit readily
assayedcircadianbehaviours,suchaslocomotoractivityrhythmswhicharesimilarto
humansleep-wakecycles.Moreimportantly,mostoftheclockgenesexpressedinfruit
fliesaresimilartotheirmammaliancounterparts[PeschelandHelfrich-Förster,2011].
Thirdly, powerful genetic tools and the simplicity of neuronal organization make
Drosophilaanexcellentsystemtodissect[RobertsonandKeene,2013]. Inaddition,
fruitfliesliveforonly50-80daysinstandardlaboratoryconditions,allowingbehaviour
andgeneexpressiontobemonitoredoveranentire lifecycle [Kohetal.,2006]. In
conclusion,due tonumerousadvantages,Drosophila isexpected tobeaneffective
modelinelucidatingbiologicalmechanismsunderlyingthecircadianclock,whichmay
eventuallybegeneralizedtootherspecies.
TransgenicLuciferase-reporterLines8.0-luc,XLG-lucandtim-lucwereusedforthewhole
project.ThesefruitfliescontainaperortimpromoterdrivenfireflyluciferasecDNA,

TheDevelopmentoftheCircadianClock
63
which exhibits a similar cyclic expression pattern and tissue distribution as their
endogenouscounterparts(Table2-1)[Stanewsky,2007].Inthedevelopmentalstudy,
we used 8.0-luc and XLG-luc flies to measure PER expression in the central clock
neurons and peripheral tissues, respectively. 8.0-luc flies specifically reflect PER
expression in subsets of the central clock dorsal neuronal clusters (DN1-3) and LNd
[Velerietal.,2003;Yoshiietal.,2009].Adult8.0-lucfliesexhibitrelativelylowoverall
bioluminescence levels, however show robust rhythmicity in DD, with little or no
dampeningofrhythmicity[Velerietal.,2003].Inordertodistinguishtheeffectfrom
thecentralclock,asecondpertransgenicluciferasereporter,XLG-lucwasused,which
showsPERexpressioninallknownper-expressingcellsthroughoutthebody,including
central and peripheral clocks [Veleri et al., 2003]. However, it mainly reports PER
signalsinperipheraloscillatorsbecauseofthesmallnumberofcentralclockneurons
in the brain compared to thousands of clock cells in the rest of the body. Though
bioluminescencesignalinXLG-lucfliesdampensrapidlyinDD,itshigh-levelexpression
ofPERcanhelpidentifywhetherclockgeneisexpressing,especiallywhenPERlevelis
presumablylowinpre-adultstages.
Table2-1.CharacteristicsofDifferentLuciferase-reporterLines
Name Gene SignalLevel Remarks
8.0-luc period Verylow RobustinDD Asubsetofneuronsinthebrain
Almosttheentirepercodingregion,no5’-UTR
XLG-luc period High RapidlydampeninDD
Mostper-expressingcells
Almosttheentirepercodingregion,plusperpromotor
tim-luc timeless Veryhigh DampeninDD tim5’-UTR(RNArhythms)

TheDevelopmentoftheCircadianClock
64
FlyStocksincludingwildtypeandtransgeniclineswererearedongrowth-media(Carolina,
USA) loaded instandardplasticvials.Fruit flieswerekeptat12:12LDcycleswitha
constanttemperatureof25˚C.DuringLDcycles,Zeitgebertime(ZT)isdefinedaslight
onat8amNZST(ZT0)andlightoffat8pmNZST(ZT12).
DevelopmentalStageswererecordeddailybymonitoring individual fruit fliesreared in
separatewellsofclear96-wellmicroplates(GreinerBio-one).Thegrowthenvironment
of these flies were identical to those going through bioluminescence assay and
locomotoractivitymonitoring.

TheDevelopmentoftheCircadianClock
65
2.4.3MeasurementofClockGeneExpression
BioluminescenceAssayusesluciferaseasareal-timereporterofgeneexpressioninvivo,
making the continuous monitoring of clock gene levels in living animals possible
[Stanewskyetal.,1997].Theadvantagesofthistechniqueincludeshort-intervaland
long-termmeasurements,aswellastime-seriesdatacollectedfromasingleliveanimal.
Methods for directly measuring gene expression include Northern blots, RNase
protection assays, quantitative real-time reverse transcription polymerase chain
reaction(RT-PCR),aswellastechniquesusedforproteinandenzymelevelsdetection,
allrequirethesacrificeofanimalsatdifferentcircadiantimes.Consequently,itisnot
practicaltotracegeneexpressionatshortenoughintervalsandoverlongenoughtime
to accurately reflect the key parameters of circadian oscillation. Furthermore, it is
impossible to analyse molecular rhythms within an individual organism over time
[Stanewsky,2007;YuandHardin,2007].
Thesubstrateluciferinfortheenzymereactioncanbeeasilyadministeredtolivefliesby
addingittotheirfood,ortoisolatedflytissuesbyaddingitdirectlytotheirculture
media [Stanewsky, 2007; Yu and Hardin, 2007]. It has been reported that live
Drosophilaembryoscanabsorb luciferinwhen immersed in the luciferin-containing
solutionforonly5minutes[Akiyoshietal.,2014].

TheDevelopmentoftheCircadianClock
66
Real-time bioluminescence recording of per was conducted as described previously
[Stanewsky,2007]andillustratedinFigure2-4.Individualeggswerecollectedwithin
fourhoursofbeinglaidinDDandtransferredintoasinglewellofwhiteopaque96-
wellmicroplates(GreinerBio-one)under100-luxwhitelight.Minimallightwasused
becauseeggscannotbedistinguishedunderDDwiththeaidofinfrarednightvision
equipment or under red light. This short-term operation falls in the first half of
embryonic stage, when neither the molecular clock or its light-response has
establishedaccordingtoour (refer toResultsandDiscussion)andprevious findings
[Houl et al., 2008]. Each well contained growth-media fortified with D-Luciferin,
Potassiumsalt(15mM),andthosewithoutanimalswereusedascontrols.Plateswere
loadedintoaMultimodePlateReader(EnSpire,PerkinElmer)keptinsideanincubator
(Percival Scientific Inc, USA) and subjected to DD or light cycles (LD 12:12).
Bioluminescencelevelsemanatingfromeachwellwererecorded,fromtheembryonic
stageonward.Experimentswereconductedfor20daysat25°Cand10daysat16°C.
Theshorterdurationatthelowertemperatureencompassedtheembryonicandearly
larvalstageswhichweretheprimaryfocusofthestudy.

96-well Plate (side-view)
Drosophila Large Vials
96-well Plate with Eggs
Film
Eggs
Luciferin-mixed Media
67
Figure 2-4. Bioluminescence Assay Conducted in Early Developmental Drosophila
The recording of clock gene expression started from embryonic stage. Individual eggs were
collected in large vials within four hours of being laid by fertilized females and transferred into
every other well of white opaque 96-well microplates. Each well contained growth-media fortified
with luciferin. Plates were sealed with plastic films and loaded into a Multimode Plate Reader.
Drosophila melanogaster

TheDevelopmentoftheCircadianClock
68
2.4.4MonitoringofLocomotorActivity
Drosophila Activity Monitoring (DAM) System was used for continuously recording
locomotoractivityoffruitflies.Thisdeviceisbasedonusingtheinterruptionofabeam
of infrared light to record the locomotoractivityof individual fliescontained inside
smalltubes[Zordanetal.,2007;Chiuetal.,2010].
EggsofwildtypefruitflieswerecollectedinstandardvialsafterbeinglaidallunderDD
withtheaidofinfrarednightvisionequipment.FliesfromonegroupwerekeptinDD
duringtheentireexperiment,andothergroupsweregivena12-hour1000luxlight
pulseatdifferentdevelopmentaltimeswithinembryonicandearlylarvalstages.When
aparticulargroupofflieswasunderlighttreatment,theothergroupswerekept in
darknesssimultaneously(Figure2-5).Experimentswereconductedat25°Cand16°C
respectively.LocomotoractivityofflieswasmonitoredautomaticallyusingTrikinetics
DrosophilaActivityMonitors(Waltham,MA,USA),aspreviouslydescribed[Zordanet
al.,2007].Onthedaysbeforeeclosion,withtheaidofinfrarednightvisionequipment
in DD, individual pupae were transferred into glass tubes (8 cm long and 5 mm
diameter)withgrowthmediaatoneend,sealedwithplasticplugs,andpluggedwith
cottonattheotherend.Locomotoractivity,temperatureandrelativehumiditywere
recordedeveryfiveminutesforatleastsevendaysinDD.

69
Figure 2-5. The Light-pulse Regimen in Early Developmental Drosophila
Larva
Embryo
Embryo
25℃ 16℃
12 h
12 h
12 h
12 h
12 h
12 h
12 h
12 h
Embryo
Embryo
Embryo
Embryo
Larva
0
GROUP A B C D E F G H I J
Darkness Light Pulse
Groups of wild type flies kept in DD at either 25℃ or 16℃ were given light treatment during early
development. Flies from group A and group E were kept in DD during the entire experiment, and
other groups were given a 12-hour light pulse at different developmental times within embryonic
and early larval stages as illustrated. When a particular group of flies was under light treatment,
the other groups were kept in darkness simultaneously.

TheDevelopmentoftheCircadianClock
70
2.4.5DataAnalysis
Datawereshowninmeanandstandarderror(Mean±SEM).ClockLab(Actimetrics,USA)
wasusedtoanalyseforfree-runningperiods(tau),phasesandrhythmicitystrengthof
wildtypeflies’ locomotoractivityinDD.Lomb-ScargleperiodograminClockLaband
MetaCycle [Wuetal.,2016]wereusedfortheanalysisofperiodicityandrhythmicity
ofbioluminescencesignals,andP<0.05wereconsideredrhythmic.Bioluminescence
datawerenormalisedbyamovingaverageofsixhours,asdescribedbyLevineetal
[Levineetal.,2002].
AsthephasesofPERexpressioninindividualflies(both8.0-lucandXLG-luc)raisedinDD
donotnecessarilysynchronize,rhythmsofPERexpressionwerealignedpost-hocat
thedataanalysisstage.Thiswasachievedbymanuallyaligningtheacrophaseofflies
acrosstherecordingtime.Toensurethatnolarvalsignalwasincorrectlyassumedto
beanembryonicsignal,wechoseacut-offofembryonicexpressiontobe24-hour(as
thisisthestandarddevelopmentaltimeofembryosat25°C).

TheDevelopmentoftheCircadianClock
71
2.5Results
2.5.1ClockGenePeriodExpressionintheCentralClockDuringEarlyDevelopment
TomeasurePERexpressioninsubsetsofthecentralclockneuronsweused8.0-lucflies,
whichexpressaPER-luciferasefusionprotein inthedorsalneuronalclusters(DN1-3)
and the LNd [Veleri et al., 2003; Yoshii et al., 2009]. During the 20-day continuous
recording at 25°C, starting with 8.0-luc embryos, we found that 71.3% of 8.0-luc
individuals(n=108)showedthestartofrhythmicPERexpressionfromtheembryonic
stageonwardsinDD(p<0.05).Oscillationsintheremainingindividualsdidnotreach
significance (Figure 2-6). Similarly, during the10-day continuous recording at 16°C,
87.8%of8.0-luc flies (n=41) showedthestartof rhythmicPERexpression fromthe
embryonicstageonwardsinDD(p<0.05).Oscillationsintheremainingindividualsdid
not reach significance (Figure 2-7). At this lower temperature, where embryonic
developmentaltimeisdoubled,clearpeaksofPERexpressionoccurredafter24hours
and 48 hours, demonstrating that the molecular clock starts operating during
embryogenesis. Because rhythmic PER expression in flies reared in DD was not
synchronizedwitheachotherateithertemperature,wemanuallysynchronizedthe
signalsbyaligningtheiracrophasesandgroupedthemasanaveragesignal,tovisualise
rhythmicity.At25°C,groupedPERexpressionduringembryonicstage(24hours)and
earlylarvalstage(stages1and2)wereemphasizedandshown(Figure2-6A),witha
tauof23.1-hour.At16°C,groupedPERexpressionduringembryonicstage(48hours)

TheDevelopmentoftheCircadianClock
72
and1stlarvalstage(48hours)wereemphasizedandshown(Figure2-7A),withatau
of25.1-hour.

Figure 2-6. Bioluminescence Time-series Data of Individual 8.0-luc Fruit Flies Measured in DD at 25℃
Bio
lum
ine
sce
nce
(CP
S)A
B
Larva Pupa Adult
Time (hour)
Bio
lum
ine
sce
nce
(CP
S)
Embryo
Development
Period (hour)
Am
plit
ud
e
tau=23.1
Individual bioluminescence counts per second (CPS) is plotted by six-hour moving average as grey
lines, and synchronized by acrophases and averaged as red lines. Control is the average signal
from wells contained only luciferin-media without any flies under the same condition, as black
lines. Corresponding developmental stages are shown. (A) Synchronized PER expression in 8.0-
luc (n=77) during embryonic and early larval stages of the first 72 hours after fertilization,
periodogram displays significant circadian rhythm (Lomb-Scargle, p<0.05) with a 23.1-hour tau,
and (B) from the embryo until adult of the first 20 days after fertilization shows cyclic expression
with a mean period of 23.9±0.13 hours, as show in the box and whisker plot.
Development
EmbryoTime (day)
73
Pe
rio
d (
ho
ur)
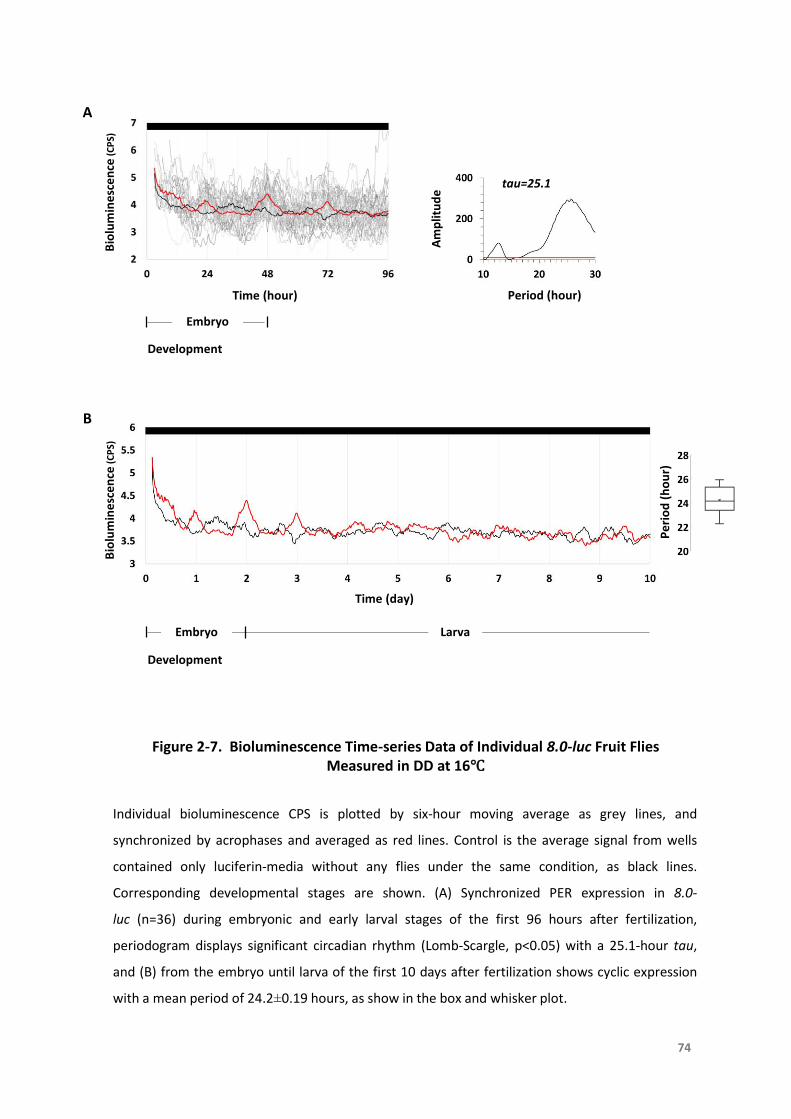
Bio
lum
ine
sce
nce
(CP
S)A
B
Larva
Time (hour)
Bio
lum
ine
sce
nce
(CP
S)
Embryo
Development
Period (hour)
Am
plit
ud
e
tau=25.1
Development
Embryo
Time (day)
Figure 2-7. Bioluminescence Time-series Data of Individual 8.0-luc Fruit Flies Measured in DD at 16℃
Individual bioluminescence CPS is plotted by six-hour moving average as grey lines, and
synchronized by acrophases and averaged as red lines. Control is the average signal from wells
contained only luciferin-media without any flies under the same condition, as black lines.
Corresponding developmental stages are shown. (A) Synchronized PER expression in 8.0-
luc (n=36) during embryonic and early larval stages of the first 96 hours after fertilization,
periodogram displays significant circadian rhythm (Lomb-Scargle, p<0.05) with a 25.1-hour tau,
and (B) from the embryo until larva of the first 10 days after fertilization shows cyclic expression
with a mean period of 24.2±0.19 hours, as show in the box and whisker plot.
74
Pe
rio
d (
ho
ur)

TheDevelopmentoftheCircadianClock
75
2.5.2ClockGenePeriodExpressioninthePeripheralClocksDuringEarlyDevelopment
After observing theseper expressionpatterns at the central clock level,wedecided to
furtherstudyhowPERoscillationsdevelopattheperipheralclockslevel.Inorderto
distinguishtheeffectfromthecentralclock,asecondperluciferase-reporterline,XLG-
lucwasused,whichisexpressedinallknownper-expressingcellsthroughouttheadult
body[Velerietal.,2003].
Againbecause rhythmicPERexpression in flies reared inDDwasnot synchronized,we
manually synchronized the signals by aligning their acrophases at 25°C.During the
continuous 20-day and 10-day XLG-luc recording at 25°C or 16°C, respectively, we
foundthatPERstartedtoriseduringtheembryonicstage.Alloftheembryoskeptin
DD(n=68)andinLD(n=67)at25°CshowedthisincreaseofPERsignal(Figure2-8&2-
9). The rise in PER peaked at the pupae and then decreased, phase differences
betweenDDandLDrecordsmostlikelywerecausedbymanualphasesortingofthe
DDrecordstoaidrecognitionofrhythmicPERexpression(seeMethodsSection2.4.5
for details). Importantly no oscillations were evident during this first rise of PER.
SubsequentlyPERbegantoincreaseagainaftereclosionandatthistimealsobeganto
cyclewithtauof22.9hoursinDDandwithperiodof23.9hoursinLD.Thestrengthof
rhythmicityofthebioluminescenceoscillationswasmorerobust inLDcomparedto
DDasexpected.At16°C,PERexpressioninXLG-luc(n=14inDDandn=82inLD)beings
torisefromtheembryonicuntillarvalstagewithoutoscillations(Figure2-10).
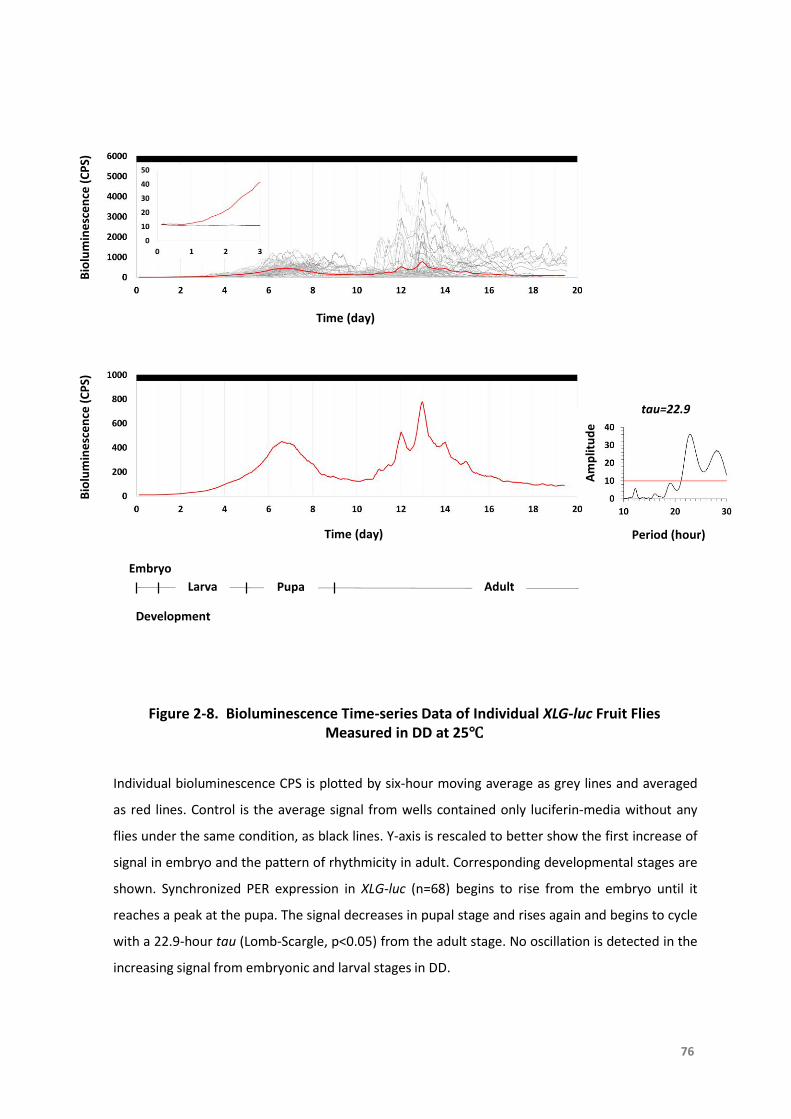
Figure 2-8. Bioluminescence Time-series Data of Individual XLG-luc Fruit Flies Measured in DD at 25℃
Bio
lum
ine
sce
nce
(C
PS)
Time (day)
tau=22.9
Am
plit
ud
e
Period (hour)
Larva Pupa Adult
Embryo
76
Individual bioluminescence CPS is plotted by six-hour moving average as grey lines and averaged
as red lines. Control is the average signal from wells contained only luciferin-media without any
flies under the same condition, as black lines. Y-axis is rescaled to better show the first increase of
signal in embryo and the pattern of rhythmicity in adult. Corresponding developmental stages are
shown. Synchronized PER expression in XLG-luc (n=68) begins to rise from the embryo until it
reaches a peak at the pupa. The signal decreases in pupal stage and rises again and begins to cycle
with a 22.9-hour tau (Lomb-Scargle, p<0.05) from the adult stage. No oscillation is detected in the
increasing signal from embryonic and larval stages in DD.
Time (day)
Bio
lum
ine
sce
nce
(C
PS)
Development
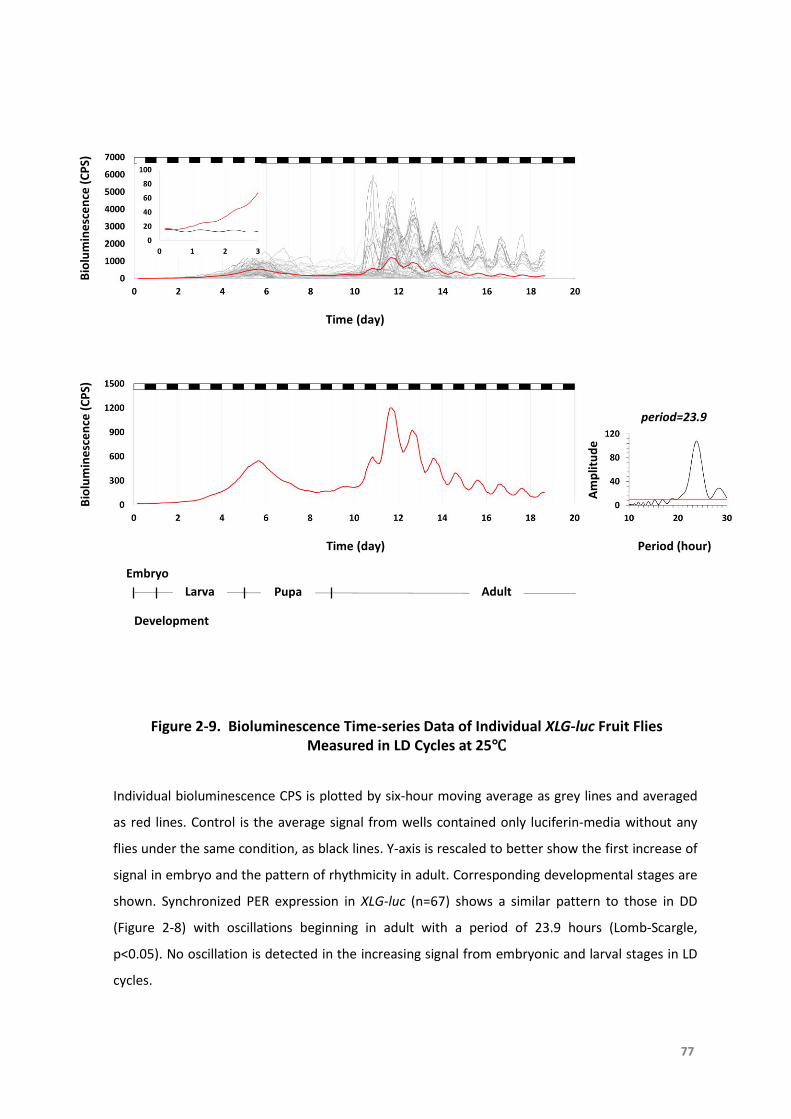
Figure 2-9. Bioluminescence Time-series Data of Individual XLG-luc Fruit Flies Measured in LD Cycles at 25℃
Bio
lum
ine
sce
nce
(C
PS)
Time (day)
Am
plit
ud
e
Period (hour)
Larva Pupa Adult
Embryo
77
period=23.9
Bio
lum
ine
sce
nce
(C
PS)
Time (day)
Individual bioluminescence CPS is plotted by six-hour moving average as grey lines and averaged
as red lines. Control is the average signal from wells contained only luciferin-media without any
flies under the same condition, as black lines. Y-axis is rescaled to better show the first increase of
signal in embryo and the pattern of rhythmicity in adult. Corresponding developmental stages are
shown. Synchronized PER expression in XLG-luc (n=67) shows a similar pattern to those in DD
(Figure 2-8) with oscillations beginning in adult with a period of 23.9 hours (Lomb-Scargle,
p<0.05). No oscillation is detected in the increasing signal from embryonic and larval stages in LD
cycles.
Development

Figure 2-10. Bioluminescence Time-series Data of Individual XLG-luc Fruit Flies Measured at 16℃
Bio
lum
ine
sce
nce
(C
PS)A
Bio
lum
ine
sce
nce
(C
PS)B
Time (day) Period (hour)
Individual bioluminescence CPS is plotted by six-hour moving average as grey lines and averaged
as red lines. Control is the average signal from wells contained only luciferin-media without any
flies under the same condition, as black lines. Y-axis is rescaled to better show the first increase of
signal in embryo. Corresponding developmental stages are shown. PER expression in (A) DD and in
(B) LD (n=14 and n=82, respectively) begins to rise from the embryo until larva without oscillations
(Lomb-Scargle, p>0.05). Periodogram in LD is calculated by average signal subtracted by control
signal to counteract the pseudo-rhythm caused by the light-induced background [Stanewsky,
2007].
Larva
Development
Embryo
Period (hour)Time (day)
Larva
Development
Embryo
78

TheDevelopmentoftheCircadianClock
79
2.5.3 Synchronized Behavioural Rhythms in Adult Flies Exposed to Light Early in
Development
Theresultsfromtheaboveexperimentsindicatedtheexistenceofafunctionalclockinthe
embryos, thusweattempted toconfirmthisby testingwhether lightgivenat such
earlydevelopmental stageshadaneffecton thephaseofadult locomotoractivity.
GroupsofwildtypeflieskeptinDDweregivenlighttreatmentatdifferenttimesof
development (Figure 2-5).We found that the locomotor activity of individual flies
raised inDDwasnot synchronized at either 25°Cor 16°C. Theprobability that the
distribution of individual phases within a population is random was calculated by
Rayleigh’s test. Probabilities for flies reared in DD being synchronized were not
significant indicating random distribution (p>0.05). Flies which received light-
treatment at the first half of embryonic stage showed desynchrony at both
temperatures,incontrast,thosethathadreceivedlight-treatmentatthesecondhalf
ofembryonicstageandearlylarvalstageshowedsynchronizeddistributionsofphases
(Figure2-11).DespitethefactthatthegroupofflieskeptinDDwasneverexposedto
light during their early development, they showed rhythmic locomotor activity as
adultssimilartoallthelight-treatedgroups(Table2-2).

Figure 2-11. The Circular Distributions of the Locomotor Activity Acrophases, Average Actograms and Periodograms at (A) 25°C on the Left Panel and (B) 16°C on the Right Panel
Larva
Embryo
Embryo
25℃
A B
16℃
12 h
12 h
12 h
12 h
12 h
12 h
12 h
12 h
DD
00:00
12:00
18:00
06:00
DD
00:00
12:00
18:00
06:00
00:00
12:00
18:00
06:00
00:00
12:00
18:00
06:00
Am
plit
ud
e
Period (hour)
tau=24.2
tau=24.1
Am
plit
ud
eA
mp
litu
de
Am
plit
ud
e
Period (hour)
Period (hour)
Period (hour)
Embryo
Embryo
Embryo
Embryo
Larva
00:00
12:00
18:00
00:00
12:00
18:00
00:00
12:00
18:00 06:00
00:00
12:00
18:00 06:00
00:00
12:00
18:00 06:00
00:00
12:00
18:00 06:00
Am
plit
ud
eA
mp
litu
de
Am
plit
ud
eA
mp
litu
de
Am
plit
ud
eA
mp
litu
de
Period (hour)
Period (hour)
Period (hour)
Period (hour)
Period (hour)
Period (hour)
tau=23.3
tau=23.3
tau=23.6
80
One group per panel was under DD throughout the experiment, 12-
hour light pulses were administrated to other groups during
different timing of early development. Black dots around the 24-
hour clock faces indicate the timing of acrophase of every single fly
in the same group. The mean phase of each population is shown by
the angle of the radius with 95% confidence intervals. Double-
plotted actograms are averaged from flies in each group. (A) The
animals kept in DD for the entire experiment (n=28) and given light
pulses at the first 12-hour development (embryo, n=34) show
random distributions (p=0.48 and p=0.35, respectively), with
arrhythmic average actograms supported by periodograms. Light
pulses given during the second 12-hour (embryo, n=38) and third
12-hour development (larva, n=35) both show significantly
concentrated distributions (p<0.001), with rhythmic average
actograms supported by periodograms (Chi-square, p<0.05). (B) The
animals kept in DD for the experiment (n=43) show random
distributions (p=0.08). Light pulses given on the first (embryo, n=48)
and second (embryo, n=45) 12-hour development both show
random distributions (p=0.82 and p=0.67, respectively), with arrhythmic
average actograms supported by periodograms. Light pulses given on the
third (embryo, n=42), fourth (embryo, n=54) and fifth (larva, n=47) 12-
hour development all show significantly concentrated distributions
(p=0.003, p=0.01 and p=0.03, respectively), with rhythmic average
actograms supported by periodograms (Chi-square, p<0.05).
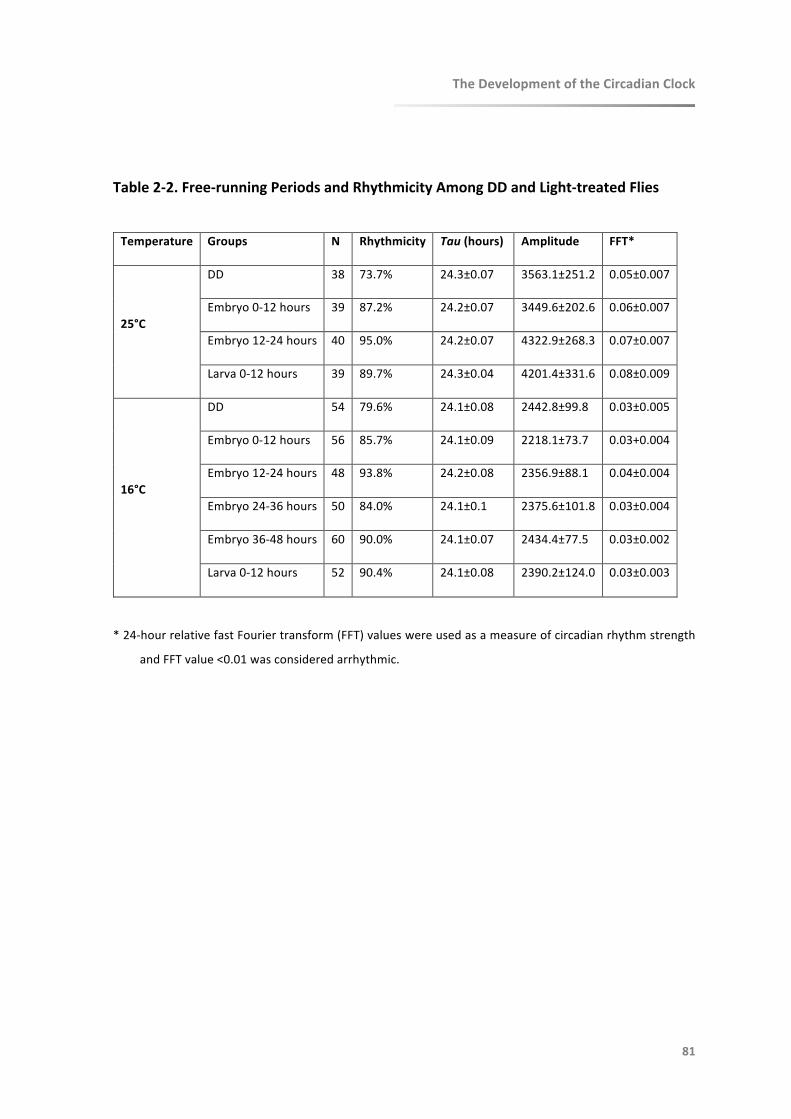
TheDevelopmentoftheCircadianClock
81
Table2-2.Free-runningPeriodsandRhythmicityAmongDDandLight-treatedFlies
Temperature Groups N Rhythmicity Tau(hours) Amplitude FFT*
25°C
DD 38 73.7% 24.3±0.07 3563.1±251.2 0.05±0.007
Embryo0-12hours 39 87.2% 24.2±0.07 3449.6±202.6 0.06±0.007
Embryo12-24hours 40 95.0% 24.2±0.07 4322.9±268.3 0.07±0.007
Larva0-12hours 39 89.7% 24.3±0.04 4201.4±331.6 0.08±0.009
16°C
DD 54 79.6% 24.1±0.08 2442.8±99.8 0.03±0.005
Embryo0-12hours 56 85.7% 24.1±0.09 2218.1±73.7 0.03+0.004
Embryo12-24hours 48 93.8% 24.2±0.08 2356.9±88.1 0.04±0.004
Embryo24-36hours 50 84.0% 24.1±0.1 2375.6±101.8 0.03±0.004
Embryo36-48hours 60 90.0% 24.1±0.07 2434.4±77.5 0.03±0.002
Larva0-12hours 52 90.4% 24.1±0.08 2390.2±124.0 0.03±0.003
*24-hourrelativefastFouriertransform(FFT)valueswereusedasameasureofcircadianrhythmstrength
andFFTvalue<0.01wasconsideredarrhythmic.

TheDevelopmentoftheCircadianClock
82
2.6Discussion
InthisstudyweanalysedthedevelopmentofthecircadianclockinDrosophilabyutilizing
luciferase reporterswhich record theessential clockcomponentPER in vivo. These
toolsallowedustomonitortheexpressionofclockgenesinreal-timeandtokeeptrack
ofthemeasurementthroughouttheearlylifeoftheflies.Herewehaveshownthat
theexpressionofPERinperipheralclocksstartsintheembryosbutdoesnotbeginto
cycleuntiltheadultstage.However,areporterthatreflectsPERexpressioninsubsets
of central clock neurons only, starts to show rhythmic circadian patterns during
embryogenesis, coinciding with the time when light begins to affect the clock. In
addition,andinagreementwithpreviousstudies,clockgeneoscillationsofPERaswell
asadultlocomotoractivityrhythmsinitiatewithouttherequirementoflight.
Slowingdowndevelopmentalspeedbyexposingembryostoconstant16°Crevealedthe
initiationofcircadianPERexpressionfromasearlyastheembryonicstageinDD.PER
in8.0-lucfliesisspecificallyexpressedwithinsubsetsoftheDNandLNd,butnotinLNv,
norinperipheralclocktissuesinadults,whichexplainsthelowbioluminescencesignal
detected.WesuggestthatsomeoftheDNand/orLNdprecursorneuronsmaypossess
a functional clock already very early during development. On the basis that per
expressionhasbeen reported inDN1ands-LNvduringembryonic stages16and17
[Houletal.,2008]whichfallwithinthesecondhalfofembryogenesisandthatLNdonly

TheDevelopmentoftheCircadianClock
83
differentiatesinthepupalstage[Helfrich-Försteretal.,2007],weinferthesignalfrom
8.0-lucfliespotentiallyreflectsembryonicPERexpressioninDN1ortheirprecursors.
ThemeaningoftheearlyemergenceofDNduringclockdevelopmentisstillunknown.In
adultDrosophila,thesignificanceofDNhasbeenshowninrecentfindings.Originally,
the DN are believed to be responsible for controlling the laboratory crepuscular
activitypatternsunderLDconditions,butunderconstantconditions,theyarenotable
tomaintainrhythmiclocomotoractivitywithouttheLNv[Velerietal.,2003;Helfrich-
Försteretal.,2007].Nonetheless,insteadoftraditionallyputtingLNvinthepositionof
principalcircadianpacemakers,recentstudieshaveplacedemphasisontheweightof
othergroupsofclockneurons,includingDN,whichareallrequiredfororchestrating
bouts of activity [Dissel et al., 2014; Yao and Shafer, 2014; Yao et al., 2016].
Furthermore,ithasbeenproposedthatDNaretemperature-sensitivewhiles-LNvare
temperature-insensitive,suchthatDNisresponsiblefortemperatureentrainmentin
Drosophila[BarberandSehgal,2018].ThusweassumethatDNmayplayanimportant
roleonclockdevelopmentanditsfunctionduringembryogenesis.
Our finding that a 12-hour light pulse given at the second half of embryogenesis can
synchronize adult behavioural rhythms is consistent with the molecular result. It
supportstheideathatatleastsomefunctionalcomponentsofthecentraloscillator
and its light sensitivityarepresent from theverybeginningofdevelopment, i.e. in
precursorsofclockneurons.Comparedtopreviousfindings,weshowthatthecentral
clockinitiatestickingattheembryonicstage.

TheDevelopmentoftheCircadianClock
84
Aside from the identified central clock neurons during early development, studies on
peripheralclockdevelopmenthasattractedlessattention.Ithasbeenrevealedthata
significantnon-cyclicexpressionofpermRNAinitiatesinmid-latedevelopingembryos,
which is limitedtothemidlineofthenervoussystem[Jamesetal.,1986;Liuetal.,
1988;Houletal.,2008;Ruizetal.,2010].ThisisconsistentwithourfindingthatPER
expressionbeginsfromtheembryosbutdoesnotoscillateineitherDDorLDatthe
wholeanimallevel.However,thelinearlyincreasingexpressionfromembryotolarva
isdifferentfrompreviousreports,whichhaveshownlowexpressioninlarvalstageand
itsre-emergenceatthemid-pupalstage[Liuetal.,1988].
Intheembryosandlarvae,PERbegantocycleinthe8.0-lucflies,whileanon-cyclicincrease
wasobserved inXLG-luc flies, suggesting that central neuronal oscillations of clock
geneexpression initiatepriortothose inperipheralclocks.BecauseXLG-luc reports
PERexpressioninbothcentralandperipheralcells,wecannotexcludethatsomeof
thesignalmaycomefromthecentralclockneurons.However,ofthetotalnumberof
approximate300per-expressingcellsintheCNSduringembryogenesis,amongwhich
onlymaximum20aredefinedascentralclockneurons[Ruizetal.,2010].Itsuggests
that the XLG-luc signal mainly indicates the periphery. Of course, the absence of
circadian rhythms in the whole animal level does not exclude the possibility that
individual tissuesorcellshavealreadyoscillatedduringearlydevelopment [Dekens
andWhitmore, 2008; Dolatshad et al., 2010], presumably out-of-phase with each
other.WefurtherinferthatPERisexpressedwithindifferentperipheralclocksaround
thebody,whichcouldbecyclicornon-cyclic,butnotsynchronizedatthewholeanimal

TheDevelopmentoftheCircadianClock
85
levelbeforeeclosion.Becauseofthenon-synchronizedPERexpressioninLDcycles,
our results also suggest that photoreceptors in peripheral clocks have not fully
developedbeforetheadultstage.Thisissupportedbyarecentfindingthatalthough
expressionofCryptochromeexistsincertainlarvalperipheraltissuebutnotothers,it
lacks the functionwhichmediates light-entrainment inboth central andperipheral
clocksinadultDrosophila[Agrawaletal.,2017].Thisisdifferentfromthevertebrate
systemwherecells inzebrafisharesensitivetoandcanbesynchronizedby lightat
early embryonic stage,while in the adult peripheral clocks of bothDrosophila and
zebrafisharedirectlylight-sensitive[Tamaietal.,2005].
ThecyclicexpressionofPERatthewholeanimallevelinearlyadultsuggeststhatamature
clocksystem,includingtheentrainablecentralclockandthesynchronizedperipheral
clocks,isfunctioningfromthepointofeclosiononward.Previousworkssuggestthat
a number of pacemaker neurons do not initiate rhythmic PER expression until the
pupal development [Kaneko et al., 1997; Kaneko and Hall, 2000; Liu et al., 2015].
Furthermore,twogroupsoflateralneuronsLNdandl-LNvdifferentiatenoearlierthan
inpupalstage[Helfrich-Försteretal.,2007].Combinedwiththefindingthatoneofthe
novelvisualprojectionneuronscanonlybeidentifiedduringpupae[Yanetal.,2013],
itimpliesthatawell-equippedcircadiansystemisestablishedaftereclosion.
Oneoftheimportantfeaturesofthecircadianclockisthatitisendogenous.Thecurrent
developmentalstudyshowsthatincertainspecies,suchasDrosophila,thecircadian
clockisabletodevelopandmaturewithouttherequirementofentrainingstimulisuch

TheDevelopmentoftheCircadianClock
86
aslight[Sehgaletal.,1992].Inourstudylightexposuredidnotinitiateorspeedupthe
developmentofthecircadianclockorchangetheclockgeneexpressionlevel,butit
synchronized the phases among individual flies. This is consistentwithmammalian
clockswhichcanstartsynchronizedoscillationswithinanindividualindependentlyof
Zeitgebers, and independently of a functional maternal circadian clock [Jud and
Albrecht,2006].Italsosharessimilaritywithclocksinzebrafishwhichcaninitiatebut
notsynchronizewithoutlight[DekensandWhitmore,2008].
Asinglelightpulsegivenduringtheembryonicstageinthecurrentstudy,orduringthe
larval stage in previous studies [Kaneko et al., 2000], is able to synchronize adult
rhythmicbehaviour.Onepossibleexplanationisa“time-memory”whichimpliesthe
specialabilityoftheclocktomemorisesuchstimuliandmanifestitlaterinlife,when
theclockbecomesmoremature[Kanekoetal.,2000].Thedirectionandmagnitudeof
phaseshiftsoflocomotoractivityhasbeenshowntobedifferentbetweenwildtype
andfast-clockpershortmutants,suggestingtheinvolvementofperintheregulationof
developmental “time-memory” [Kaneko et al., 2000]. However a reasonable
assumptionisthat,atsomepointduringearlyadultstages,synchronizationbetween
differentclockcellsisundercontrolofanasyetundescribedintrinsicmechanism.So
insummary,ratherthanatime-memory,itispossiblethatthecentralclockis‘set’by
theinitiallightbutlacksofthepowertosynchronizethemajorityofperipheralclocks,
thusthecircadiansystemthroughoutthebodyfree-runsuntiladulthood.

TheDevelopmentoftheCircadianClock
87
2.7Conclusion
An important role of the circadian clock in the regulation of development has been
reported[Kyriacouetal.,1990;Kumaretal.,2006;Valloneetal.,2007].Forinstance,
althoughknownclocknullmutantssuchasper01[KonopkaandBenzer,1971]andtim01
[Sehgal et al., 1994] show no destructive impact on development, tissue-specific
knock-downofclockgenescanblocklarvaldevelopmentthusleadingtolethality[Di
CaraandKing-Jones,2016].Inthisstudyweplaceemphasisonthedevelopmentof
thecircadianclock itself. It isassumedthat thedevelopmentof thecircadianclock
shouldbeinaccordancewiththesequentialandprecisedevelopmentalprocessofthe
entireorganism.Asaconsequence,thecircadianclockisthoughttodevelopasthe
wholebodydevelops,inagradualway[Valloneetal.,2007].Inanumberofdiverse
species,circadianclockmaturityisbeautifullylinedupwiththeneedsoftheanimalat
variouslifestagesanddevelopmentalmilestones,suchasperinatalstageinmammals
[Valloneetal.,2007],andinDrosophilaasourresultssuggest.
Throughthenewapplicationofestablishedluciferase-reportertechniques,weshowthat
cyclicclockgeneexpression,presumablyinprecursorsofdorsalclockneuronsoccurs
during the embryonic stage, which is earlier than previously thought. Light is not
requiredtostarttheclockbutitcansynchronizetheclockasearlyasintheembryos,
alsoearlier thanpreviously thought. In total, theearlycyclicexpressionofPERand

TheDevelopmentoftheCircadianClock
88
light-entrainablebehaviourinthisstudysuggestthatkeycomponentsofthecircadian
clockarepresentfromtheverybeginningoflife(Figure2-12).
Thereareofcourseotherplayersapartfromperthatmakeuptheclockandwecannot
concludefromthepresentstudywhenthesecomponentsdevelopinrelationtoper.
It is not hard to imagine that these factors interact to control the running of the
circadian system and some may play additional roles in controlling development.
Futurestudiesmayputemphasisondevelopmentaldetails in individualpacemaker
neurons,perhapsbyperforming simultaneous recordingsofdevelopmentalevents,
behaviouralrhythmsandclockgeneexpressioninthesameanimal.

Embryo
Larva
Pupa
12h12h
4d
4d
12h12h
16℃
25℃
WHEN
Embryo
Larva
Pupa
Adult
Central Light Response
Overt Rhythms
Central Gene Expression and
Oscillation
Peripheral Gene Oscillation
Peripheral Gene
Expression
Synchrony
Central Oscillations andLight Response
Synchronized Peripheral Oscillations
and Light Response
×2
×2
Start Point
Figure 2-12. Life Cycle of Drosophila and New Findings on Clock Development
Based on our findings from the current development study in Drosophila, circadian clock gene
expression in the central clock neurons and their light-sensitivity start in embryonic stage, which is
earlier than previously reported. Clock gene expression in the peripheral clocks begins in embryos
but only oscillates in synchrony after eclosion, when light-sensitivity is present.
89
De
velo
pm
en
tal O
rde
rs
Peripheral Light
Response
h: hourd: day

TheDevelopmentoftheCircadianClock
90
2.8References
AgrawalP,HoulJH,GunawardhanaKL,LiuT,ZhouJ,ZoranMJ,HardinPE.Drosophilacryentrains
clocksinbodytissuestolightandmaintainspassivemembranepropertiesinanon-clockbody
tissueindependentoflight. Curr Biol.2017 Aug 21;27(16):2431-2441.e3. DOI:
10.1016/j.cub.2017.06.064.
AhmadST,SteinmetzSB,BusseyHM,PossidenteB,SeggioJA.Larvalethanolexposurealtersfree-
runningcircadianrhythmandperLocustranscriptioninadultD.melanogasterperiodmutants.
BehavBrainRes.2013Mar15;241:50-5.DOI:10.1016/j.bbr.2012.11.035.
Akiyoshi R,Kaneuch T,Aigaki T,Suzuki H. Bioluminescenceimagingtotrackreal-time armadillo
promoteractivityinliveDrosophilaembryos.AnalBioanalChem.2014Sep;406(23):5703-13.
DOI:10.1007/s00216-014-8000-8.
Arellanes-LiceaE,CaldelasI,DeIta-PérezD,Díaz-MuñozM.Thecircadiantimingsystem:arecent
additioninthephysiologicalmechanismsunderlyingpathologicalandagingprocesses.Aging
Dis.2014Jan9;5(6):406-18.DOI:10.14336/AD.2014.0500406.
Barber AF,Sehgal A. Coldtemperaturesfireupcircadianneurons. Cell Metab.2018 May
1;27(5):951-953.DOI:10.1016/j.cmet.2018.04.016.
BedontJL,BlackshawS.Constructingthesuprachiasmaticnucleus:awatchmaker’sperspectiveon
thecentralclockworks.FrontSystNeurosci.2015May8;9:74.DOI:10.3389/fnsys.2015.00074.
BrettWJ.persistentdiurnalrhythmicityinDrosophilaemergence.AnnEntomolSocAm.1955May
1;48(3):119031.
ChiuJC,LowKH,PikeDH,YildirimE,EderyI.Assayinglocomotoractivitytostudycircadianrhythms
andsleepparametersinDrosophila.JVisExp.2010Sep28;(43).pii:2157.DOI:10.3791/2157.
Christ E,Korf HW,von Gall C. When does itstartticking?Ontogeneticdevelopmentof the
mammalian circadiansystem. Prog Brain Res.2012;199:105-18. DOI: 10.1016/B978-0-444-
59427-3.00006-X.

TheDevelopmentoftheCircadianClock
91
CirelliC.Sleepdisruption,oxidativestress,andaging:newinsightsfromfruitflies.ProcNatlAcad
SciUSA.2006Sep19;103(38):13901-2.DOI:10.1073/pnas.0606652103.
DekensMP,WhitmoreD.Autonomousonsetofthecircadianclockinthezebrafishembryo.EMBO
J.2008Oct22;27(20):2757-65.DOI:10.1038/emboj.2008.183.
Di Cara F, King-Jones K. The circadian clock is a key driver of steroid hormone production in
Drosophila.CurrBiol.Elsevier;2016Sep26;26(18):2469-77.DOI:10.1016/j.cub.2016.07.004.
Dissel S,Hansen CN,Özkaya Ö,Hemsley M,Kyriacou CP,Rosato E. The logic of circadian
organization in Drosophila. Curr Biol.2014 Oct 6;24(19):2257-66. DOI:
10.1016/j.cub.2014.08.023.
Dolatshad H,Cary AJ,Davis FC. Differentialexpressionof the circadian clock in maternal and
embryonictissuesofmice. PLoS One.2010 Mar 24;5(3):e9855. DOI:
10.1371/journal.pone.0009855.
Economos AC,Lints FA. Developmental temperature and life span in Drosophilamelanogaster.
I.Constantdevelopmental temperature: evidence for physiological adaptation in a wide
temperaturerange.Gerontology.1986;32(1):18-27.
GhoshSM,TestaND,ShingletonAW.Temperature-sizerule ismediatedby thermalplasticityof
criticalsizeinDrosophilamelanogaster.ProcBiolSci.2013Apr17;280(1760):20130174.DOI:
10.1098/rspb.2013.0174.
GlossopNRJ,GummadovaJO,GhangrekarI,HardinPE,CouttsGA.EffectsofTWIN-OF-EYELESSon
clock gene expression and central-pacemaker neuron development in Drosophila. J Biol
Rhythms.2014Jun10;29(3):151-66.DOI:10.1177/0748730414534819.
HallJC.Circadianpacemakersblowinghotandcold--but they'reclocks,not thermometers.Cell.
1997Jul11;90(1):9-12.
Helfrich-FörsterC.Developmentofpigment-dispersinghormone-immunoreactiveneurons in the
nervoussystemofDrosophilamelanogaster.JCompNeurol.1997Apr14;380(3):335-54.
Helfrich-Förster C, ShaferOT,Wulbeck C, Grieshaber E, Rieger D, Taghert P. Development and
morphologyoftheclock-gene-expressinglateralneuronsofDrosophilamelanogaster.JComp
Neurol.2007Jan1;500(1):47–70.DOI:10.1002/cne.21146.

TheDevelopmentoftheCircadianClock
92
Houl JH, Ng F, Taylor P, Hardin PE. CLOCK expression identifies developing circadian oscillator
neurons in the brains of Drosophila embryos. BMC Neurosci. 2008;9(1):119. DOI:
10.1186/1471-2202-9-119.
JamesAA,EwerJ,ReddyP,HallJC,RosbashM.Embryonicexpressionoftheperiodclockgenein
thecentralnervoussystemofDrosophilamelanogaster.EMBOJ.1986Sep;5(9):2313-20.
JudC,AlbrechtU.Circadianrhythmsinmurinepupsdevelopinabsenceofafunctionalmaternal
circadianclock.JBiolRhythms.2006Apr;21(2):149-54.DOI:10.1177/0748730406286264.
Kaneko M, Hall JC. Neuroanatomy of cells expressing clock genes in Drosophila: transgenic
manipulationoftheperiodandtimelessgenestomarktheperikaryaofcircadianpacemaker
neuronsandtheirprojections.JCompNeurol.2000Jun19;422(1):66-94.
KanekoM,HamblenMJ,HallJC.Involvementoftheperiodgeneindevelopmentaltime-memory:
effectoftheperShortmutationonphaseshiftsinducedbylightpulsesdeliveredtoDrosophila
larvae.JBiolRhythms.2000Feb;15(1):13-30.
KanekoM,Helfrich-FörsterC,HallJC.Spatialandtemporalexpressionoftheperiodandtimeless
genesinthedevelopingnervoussystemofDrosophila:newlyidentifiedpacemakercandidates
andnovelfeaturesofclockgeneproductcycling.JNeurosci.1997Sep1;17(17):6745-60.
KeeneAC,MazzoniEO,ZhenJ,YoungerMA,YamaguchiS,BlauJ,DesplanC,SprecherSG.Distinct
visualpathwaysmediateDrosophilalarvallightavoidanceandcircadianclock.entrainment.J
Neurosci.2011Apr27;31(17):6527-34.DOI:10.1523/JNEUROSCI.6165-10.2011.
Keene AC,Sprecher SG. Seeingthelight:photobehaviorinfruitflylarvae. Trends Neurosci.2012
Feb;35(2):104-10.DOI:10.1016/j.tins.2011.11.003.
KlarsfeldA,PicotM,ViasC,ChélotE,RouyerF. Identifyingspecificlightinputsforeachsubgroup
ofbrainclockneuronsinDrosophilalarvae. J Neurosci.2011 Nov 30;31(48):17406-15. DOI:
10.1523/JNEUROSCI.5159-10.2011.
Koh K, Evans JM, Hendricks JC, Sehgal A. A Drosophila model for age-associated changes in
sleep:wake cycles. Proc Natl Acad Sci U S A. 2006 Sep 12;103(37):13843-7. DOI:
10.1073/pnas.0605903103.

TheDevelopmentoftheCircadianClock
93
KonopkaRJ,BenzerS.ClockmutantsofDrosophilamelanogaster.ProcNatlAcadSciUSA.1971
Sep;68(9):2112–6.
KumarS,VazeKM,KumarD,SharmaVK.Selectionforearlyandlateadultemergencealtersthe
rateofpre-adultdevelopmentinDrosophilamelanogaster.BMCDevBiol.2006;6(1):57.DOI:
10.1186/1471-213X-6-57.
Kuntz SG, Eisen MB. Drosophila embryogenesis scales uniformly across temperature in
developmentally diverse species. PLoS Genet. 2014 Apr;10(4):e1004293. DOI:
10.1371/journal.pgen.1004293.
KyriacouCP,OldroydM,WoodJ,SharpM,HillM.Clockmutationsalterdevelopmentaltimingin
Drosophila.Heredity.1990Jun;64(Pt3):395-401.
LeGlouE,SeugnetL,ShawPJ,PreatT,GoguelV.Circadianmodulationofconsolidatedmemory
retrievalfollowingsleepdeprivationinDrosophila.Sleep.2012Oct1;35(10):1377-1384B.DOI:
10.5665/sleep.2118.
Levine JD, FunesP,DowseHB,Hall JC. Signal analysisofbehavioral andmolecular cycles.BMC
Neurosci.2002;3:1.
LiuT,MaheshG,HoulJH,HardinPE.Circadianactivatorsareexpresseddaysbeforetheyinitiate
clockfunctioninlatepacemakerneuronsfromDrosophila.JNeurosci.2015Jun3;35(22):8662-
71.DOI:10.1523/JNEUROSCI.0250-15.2015.
LiuX,LorenzL,YuQN,HallJC,RosbashM.Spatialandtemporalexpressionoftheperiodgenein
Drosophilamelanogaster.GenesDev.1988Feb;2(2):228-38.
LoobyP,LoudonAS.Geneduplicationandcomplexcircadianclocks inmammals.TrendsGenet.
2005Jan;21(1):46-53.DOI:10.1016/j.tig.2004.11.012.
Malpel S, Klarsfeld A, Rouyer F. Larval optic nerve and adult extra-retinal photoreceptors
sequentiallyassociatewithclockneuronsduringDrosophilabraindevelopment.Development.
2002Mar;129(6):1443–53.
Malpel S, Klarsfeld A, Rouyer F. Circadian synchronization and rhythmicity in larval
photoperception-defectivemutantsofDrosophila.JBiolRhythms.2004Feb;19(1):10–21.DOI:
10.1177/0748730403260621.

TheDevelopmentoftheCircadianClock
94
MatsumotoN,UwozumiK,TomiokaK.Lightcyclesgivenduringdevelopmentaffectfreerunning
periodofcircadianlocomotorrhythmofperiodmutantsinDrosophilamelanogaster.JInsect
Physiol.1997Mar;43(3):297-305.
Mazzoni EO,Desplan C,Blau J. Circadian pacemaker neurons transmit and modulate visual
informationtocontrolarapidbehavioralresponse.Neuron.2005Jan20;45(2):293-300.DOI:
10.1016/j.neuron.2004.12.038.
NitabachMN,SheebaV,VeraDA,BlauJ,HolmesTC.Membraneelectricalexcitabilityisnecessary
for thefree-runninglarvalDrosophilacircadian clock. JNeurobiol.2005 Jan;62(1):1-13.DOI:
10.1002/neu.20053.
PandaS,Hogenesch JB,Kay SA. Circadian rhythmsfromfliestohuman. Nature. 2002 May
16;417(6886):329-35.DOI:10.1038/417329a.
Peschel N, Helfrich-Förster C. Setting the clock--by nature: circadian rhythm in the fruitfly
Drosophila melanogaster. FEBS Lett. 2011 May 20;585(10):1435-42. DOI:
10.1016/j.febslet.2011.02.028.
PicotM,KlarsfeldA,ChélotE,MalpelS,RouyerF.AroleforblindDN2clockneuronsintemperature
entrainmentof theDrosophilalarvalbrain. J Neurosci.2009 Jul 1;29(26):8312-20. DOI:
10.1523/JNEUROSCI.0279-08.2009.
ReppertSM,SchwartzWJ.Maternal coordination of the fetal biological clock in utero. Science.
1983May27;220(4600):969-71.
ReppertSM,SchwartzWJ. The suprachiasmatic nuclei of the fetal rat: characterization of a
functionalcircadianclockusing14C-labeleddeoxyglucose.JNeurosci.1984Jul;4(7):1677-82.
ReppertSM,UhlGR.Vasopressinmessenger ribonucleicacid in supraopticandsuprachiasmatic
nuclei: appearanceandcircadianregulationduringdevelopment. Endocrinology.1987
Jun;120(6):2483-7.DOI:10.1210/endo-120-6-2483.
RobertsonM,KeeneAC.Molecularmechanismsofage-relatedsleeplossin thefruit fly- amini-
review.Gerontology.2013;59(4):334-9.DOI:10.1159/000348576.

TheDevelopmentoftheCircadianClock
95
RuizS,RickertC,BergerC,TechnauGM,CanteraR.Spatio-temporalpatternofcellsexpressingthe
clock genes period and timeless and the lineages of period expressing neurons in the
embryonicCNSofDrosophilamelanogaster.GeneExprPatterns.2010Sep;10(6):274-82.DOI:
10.1016/j.gep.2010.06.001.
Seggio JA, Possidente B, Ahmad ST. Larval ethanol exposure alters adult circadian free-running
locomotoractivityrhythminDrosophilamelanogaster.ChronobiolInt.2012Feb;29(1):75-81.
DOI:10.3109/07420528.2011.635236.
SehgalA,PriceJL,ManB,YoungMW.LossofcircadianbehavioralrhythmsandperRNAoscillations
intheDrosophilamutanttimeless.Science.1994Mar18;263(5153):1603–6.
SehgalA,PriceJ,YoungMW.OntogenyofabiologicalclockinDrosophilamelanogaster.ProcNatl
AcadSciUSA.1992Feb15;89(4):1423–7.
SheebaV,ChandrashekaranMK, joshiA,SharmaVK.Developmentalplasticityof the locomotor
activityrhythmofDrosophilamelanogaster.JInsectPhysiol.2002Jan;48(1):25-32.
Shibata S,Moore RY. Developmentofneuronalactivityin theratsuprachiasmatic nucleus. Brain
Res.1987Aug;431(2):311-5.
SkopikSD,PittendrighCS.Circadiansystems,II.TheoscillationintheindividualDrosophilapupa;
itsindependenceofdevelopmentalstage.ProcNatlAcadSciUSA.1967Nov;58(5):1862-9.
SládekM,JindrákováZ,BendováZ,SumováA.Postnatalontogenesisofthecircadianclockwithin
the rat liver. Am J Physiol Regul Integr Comp Physiol. 2007Mar;292(3):R1224-9. DOI:
10.1152/ajpregu.00184.2006.
SládekM,SumováA,KovácikováZ,BendováZ,LaurinováK,IllnerováH.Insightintomolecularcore
clockmechanismofembryonicandearlypostnatalratsuprachiasmaticnucleus.ProcNatlAcad
SciUSA.2004Apr20;101(16):6231-6.DOI:10.1073/pnas.0401149101.
StanewskyR.AnalysisofrhythmicgeneexpressioninadultDrosophilausingthefireflyluciferase
reportergene.MethodsMolBiol.2007;362:131-42.DOI:10.1007/978-1-59745-257-1_9.
Stanewsky R, Jamison CF, Plautz JD, Kay SA, Hall JC. Multiple circadian-regulated elements
contributetocyclingperiodgeneexpressioninDrosophila.EMBOJ.1997Aug15;16(16):5006-
18.DOI:10.1093/emboj/16.16.5006.

TheDevelopmentoftheCircadianClock
96
TamaiTK,CarrAJ,WhitmoreD.Zebrafishcircadianclocks:cellsthatseelight.BiochemSocTrans.
2005Nov;33(Pt5):962-6.DOI:10.1042/BST20050962.
ValloneD,LahiriK,DickmeisT,FoulkesNS.Starttheclock!Circadianrhythmsanddevelopment.
DevDyn.2007Jan;236(1):142–55.DOI:10.1002/dvdy.20998.
Veleri S, Brandes C,Helfrich-Förster C, Hall JC, Stanewsky R. A self-sustaining, light-entrainable
circadianoscillatorintheDrosophilabrain.CurrBiol.2003Oct14;13(20):1758–67.
Weinert D. Ontogeneticdevelopmentof themammaliancircadiansystem. Chronobiol Int.
2005;22(2):179-205.
WuG,AnafiRC,HughesME,KornackerK,HogeneschJB.MetaCycle:anintegratedRpackageto
evaluateperiodicity in large scaledata.Bioinformatics. 2016Nov1;32(21):3351-3353.DOI:
10.1093/bioinformatics/btw405.
YanY,XuY,DengS,HuangN,YangY,QiuJ,LiuJ,WangX,YangG,GuH.Apairofidentifiedgiant
visual projection neurons demonstrates rhythmic activities before eclosion. Neurosci Lett.
2013Aug29;550:156–61.DOI:10.1016/j.neulet.2013.06.035.
Yao Z,Bennett AJ,Clem JL,Shafer OT. The Drosophila clock neuron network features diverse
couplingmodesandrequiresnetwork-widecoherenceforrobustcircadianrhythms.CellRep.
2016Dec13;17(11):2873-2881.DOI:10.1016/j.celrep.2016.11.053.
Yao Z,Shafer OT. TheDrosophilacircadian clockis a variably coupled network of multiple
peptidergicunits.Science.2014Mar28;343(6178):1516-20.DOI:10.1126/science.1251285.
Yoshii T, Wulbeck C, Sehadova H, Veleri S, Bichler D, Stanewsky R, Helfrich-Förster C. The
neuropeptide pigment-dispersing factor adjusts period and phase of Drosophila's clock. J
Neurosci.2009Feb25;29(8):2597–610.DOI:10.1523/JNEUROSCI.5439-08.2009.
YuW,HardinPE.CircadianoscillatorsofDrosophilaandmammals.JCellSci.2006Dec1;119(Pt
23):4793–5.DOI:10.1242/jcs.03174.
YuW,HardinPE.Useoffireflyluciferaseactivityassaystomonitorcircadianmolecularrhythmsin
vivoandinvitro.MethodsMolBiol.2007;362:465-80.DOI:10.1007/978-1-59745-257-1_38.
Zordan MA, Benna C, Mazzotta G. Monitoring and analyzing Drosophila circadian locomotor
activity.MethodsMolBiol.2007;362:67-81.DOI:10.1007/978-1-59745-257-1_4.

to be calm and confident
When Your TALENT Outpowers Your Ego

CHAPTER 3
Aging
97

Peripheral but not Central Clock
Components Decline with Advancing Age
in Drosophila melanogaster

TheAgingoftheCircadianClock
99
Thedatafromthischapterhavebeenpublished***(pleaseseeAppendixfordetails).
***ZhaoJ,WarmanGR,CheesemanJF.Clockgeneexpressionandlocomotoractivitypredict
deathinthelastdaysoflifeinDrosophilamelanogaster.SciRep.2018Aug9;8(1):11923.
DOI:10.1038/s41598-018-30323-x.

TheAgingoftheCircadianClock
100
3.1Abstract
Thecircadianclockagesasanimalsgrowold,however,findingsabouthowagingaffects
themolecularoscillator isstillunclear.Weaimedtostudyhowtheagedmolecular
clock at both central and peripheral levels changes intrinsically under constant
conditions,andalsotoinvestigatehowitwouldbeaffectedbyregularentrainment.
Herewerecordedtheexpressionoftwokeyclockgenesinyoung(10-day-old),middle-
aged (30-day-old) and old (50-day-old) Drosophila melanogaster using transgenic
luciferase-reporter lines measuring period and timeless in vivo. We found that in
constantdarkness,PERIODexpressioninthecentralclockdorsalneuronsinagedfruit
fliesstayedasrobustasinyoungones.IncontrasttocentralexpressionofPERIOD,
PERIODandTIMELESSrhythmsintheperipheraltissuesthroughoutthebodyshowed
reduction inbothexpression level and rhythmicitywithadvancingage. Ithasbeen
reportedthatunderstrongentrainmentcuessuchastemperature-coupledlight-dark
cycles,theage-relateddeteriorationofcircadianbehaviouralrhythmscanberescued.
Nonetheless, we found no corresponding increase of clock gene expression under
light-dark cycles (12:12 at 25°C) or temperature-coupled light-dark cycles (12-hour
lightat25°Cand12-hourdarknessat21°C).Therefore,ourresultsdemonstratethat
anagedmolecularclockstillfunctionswellatthecentrallevelbutdeclinesgradually
attheperipherallevel,despitestrongenvironmentalentrainment.

TheAgingoftheCircadianClock
101
3.2Introduction
Aging is the intrinsic, inevitable, and currently irreversible process of functional
deterioration which consequently leads to the loss of viability [Fonseca Costa and
Ripperger, 2015]. The circadian clock is affected by time, showing progressive
disturbancewithadvancingage[Nakamuraetal.,2015;Banksetal.,2016].Theeffect
ofagingonthecircadianclockisbelievedtobehighlyconservedamonghumansand
otheranimals[KondratovaandKondratov,2012].
Ithasbeenwidelyobservedthatthedeclineofcircadianoutputrhythmsisaprominent
signature of aging in fruit flies and mammals, including humans. In humans, the
changesincircadianoutputsignalsassociatedwithagingincludethereductionofthe
amplitudeandearliertimingofphaseofthecircadianrhythms,bothevidentinbody
temperatureandsleep-wakecycle[WeinertandWaterhouse,2007;Brownetal.,2011;
Duffyetal.,2015;Banksetal.,2016],aswellasadisruptionofquantityandqualityof
nocturnal sleep [Arellanes-Licea et al., 2014; Hood and Amir, 2017]. Mammalian
studiesonagingdemonstratechangesinthefree-runningperiod,thephase,andthe
strengthof rhythmicitywhichhavebeen reported inbody temperature, locomotor
activitypatterns,anddrinkingbehaviour[Froy,2011;Nakamuraetal.,2015].Similarly,
prolonged free-running period, decline in sleep consolidation and the overall rest-
activityrhythmstrength,havebeenrevealedinDrosophila[Kohetal.,2006;Umezaki
etal.,2012].

TheAgingoftheCircadianClock
102
Asidefromdownstreamovertrhythms,thereisalsoevidenceforage-relatedchangesin
circadian organization at various levels, including alterations in the function of the
molecular oscillators, the neuroanatomical structures, and the transmission and
responsiveness to light [Duffy et al., 2015]. Among all, the phase resetting of the
circadian system by light is inevitably affected by aging due to impaired light
perception and transmission, as well as reduced sensitivity [Arellanes-Licea et al.,
2014]. Several studies also demonstrate impairment of photic re-entrainment at
different levels. At the behavioural level, old animals require a greater number of
transitorycyclesafteraphaseshift.At thecentral level, there isan increaseof the
cycles required for re-synchronization of the suprachiasmatic nucleus (SCN) with
increasingage[Arellanes-Liceaetal.,2014].Attheperipherallevel,organssuchasthe
esophagus,lungandthymusglandareslowertobeentrainedtoashiftedlight-dark
(LD)scheduleinoldanimals[Sellixetal.,2012].Inaddition,experimentaljetlaghas
beenreportedtoproduceabnormalphasechangesinagedperipheralclocksbutnot
inyoungones[Taharaetal.,2017].
Howthemechanismofmolecularclockanditsgenepropertieschangeduringagingare
less well known [Rakshit et al., 2012; Giebultowicz and Long, 2015]. Moreover,
whetheraging isassociatedwithdefects in thecentralclock,ordegradationof the
peripheral clocks, or loss of effective synchronization, is still a question without
confirmativeanswer.

TheAgingoftheCircadianClock
103
First, it isunclearwhetherthecentraloscillatorisaffectedbyaging, insuchcasecausal
linksbetweenclockgeneexpressionpatternsatthecentrallevelandage-associated
disturbanceincircadianrhythmscannotbeenconclusivelydemonstrated.Forinstance,
it has been reported that in the central clock neurons PERIOD (PER) is robustly
expressed insynchronyamongdifferentclockneurons inagedflies, includingsmall
ventrallateralneurons(s-LNv),dorsallateralneurons(LNd),anddorsalneurons1(DN1)
[Luoetal.,2012].However,thisischallengedbythefindingthatPERandTIMELESS
(TIM)oscillationsinalmostallcentralclockcellgroupsdeterioratewithadvancingage,
including largeventral lateralneurons (l-LNv), s-LNv, LNd,DN1anddorsalneurons2
(DN2)[Umezakietal.,2012].Inaddition,studiesperformedinagedmammalsshow
conflictingresultswitheithernormalorreducedexpressionofvariousclockgenesin
the SCN of different species [Froy, 2011; Nakamura et al., 2015], aside from a
consistent age-relateddecline inexpressionofBmal1 anda consistent lackof age-
relatedchangeintheexpressionofPeriod1(Per1)[Banksetal.,2016].
Inparticular,resultsofPeriod2/PERIOD2(Per2/PER2)expression(samegene)inagedmice
(samespecies)fromdifferentstudiesareinconsistent.Age-relatedreductionhasbeen
found inPer2mRNAmeasuredby insituhybridization invitro in free-runningaged
malemice(approximate15months)[Weinertetal.,2001],nonetheless,onlyminor
deficits of PER2 expression measured by both immunohistochemistry and
bioluminescence in vitro have been shown in free-running agedmalemice (13-16
months)[Nakamuraetal.,2011].TheamplitudeofPER2::LUCrhythmsinvitrodiffers
slightlyunderLDconditionsyetdropsmarkedlyunderconstantdarkness(DD)inaged

TheAgingoftheCircadianClock
104
malemice(13-15months)[Nakamuraetal.,2015].WhilePer2rhythmsmeasuredin
oldmaleandfemalePer2Lucmice(24-26months)invitro,thedifferenceisnotnotable
underbothDDandLD[Polidarováetal.,2017].
Secondly,thefindingsofagedmolecularclockattheperipherallevelaremoreconsistent
inDrosophila.Astudypublished in2000hasdemonstratedthataging isassociated
with reduced amplitude of circadian clock gene expression throughout the animal
[Driver,2000].Further,period(per)mRNAoscillations[Krishnanetal.,2009]aswellas
per,timeless(tim)andcryptochrome(cry)atbothmRNAandproteinlevels[Luoetal.,
2012;RakshitandGiebultowicz,2013]havebeenreportedtoreduceinheadsofold
flies. Accordingly, per promoter bioluminescence activity is found to reduce in
peripheralclocksthroughouttheflybody[Luoetal.,2012].Oneoftheinterestingfacts
liesin,despitethereductionofPERandTIM,theirnegative-feedbacktranscriptional
activatorsCLOCK-CYCLEcomplexesarestillimpairedwithincreasingage.Suchresult
suggests thatagingattenuatesthecorepropertiesof thetranscriptionalclock from
boththepositiveandthenegativelimbs[Rakshitetal.,2012].
Even thoughmost results inDrosophila studies indicatemolecular decline at the gross
peripherallevel,differenttissuesorbodypartsseemtoreacttoagingdifferently.For
example, transcriptional oscillations ofperand timhavebeenproved to reduce in
headsbutnotbodiesofagedflies[Rakshitetal.,2012].Asoneoftheperipheralclocks,
the retinal photoreceptor cells in the compound eyes of old flies show reduced
expressionofPER,yetthatisnotthecaseforanotherperipheralclock,i.e.,Malpighian

TheAgingoftheCircadianClock
105
tubules,inwhichstrongPERoscillationshasbeenobserved[GiebultowiczandLong,
2015].
Thetissue-dependentchangesofclockgeneexpressionwithadvancingageisevenmore
prominent inmammalian peripheral clocks. Based on existing data, age-associated
decline of rhythmic clock gene activity has been observed in some but not all
peripheraloscillatorsinmammals.ThecircadianexpressionprofilesofclockgenePer1,
Per2,orCryptochrome1(Cry1)mRNAmeasuredbyreversetranscriptionpolymerase
chain reaction (RT-PCR) in vitro are similar between young and aged rats (22-26
months)intheperipheraltissues(theparaventricularnucleus(PVN)andpinealgland)
[Asai et al., 2001]. This finding is supportedby subsequent studies showingalmost
identical clock genesPer2 andBmal1 expression by using similarmethodbetween
youngandagedmice(15months)intheperipheralorgans(thebrain,heartandliver)
[Oishietal.,2011],andnormalPer1andPer2mRNAinagedmice(>18months)inthe
peripheralorgans (thekidney, liverandsubmandibulargland) [Taharaetal.,2017].
Further,thesethreeperipheralclocksmeasuredinPER2::LUCagedmice(>18months)
invivoexhibitnormaloscillationamplitudesunderLDcyclesandDDconditions[Tahara
et al., 2017]. In a study using Per1 bioluminescence reporter in vitro, circadian
rhythmicityinsomeperipheraltissues(thePVN,pinealgland,arcuatenucleus,cornea,
pituitary,kidneyand liver) inagedrats (24-26months)are foundtobeunaffected,
whereassometissues(theretrochiasmaticareaandlung)aredisturbed[Yamazakiet
al., 2002]. Inaddition,amplitude,phaseandexpression levelofCLOCKandBMAL1
revealedbyimmunohistochemistryinvitroinanumberofextra-SCNregions(thePVN,

TheAgingoftheCircadianClock
106
arcuate and dorsomedial nuclei of the hypothalamus, amygdala and hippocampus)
alterinatissue-dependentmannerinagedmice(16months)[WyseandCoogan,2010].
Inahumanstudy,consistenteffectsofagingonthecircadianpatternofPERIOD1(PER1)
andPER2expressionhavebeenfoundinpost-mortembrainsamples(theprefrontal
cortex)collectedfromold(>60years)andyoung(<40years)subjects,showingaphase
advanceandanamplitudereduction[Chenetal.,2016].
Weinferthatcurrentevidenceoftheage-relatedchangesofthemolecularoscillatorsand
theirclockgenepropertiesseemnottobeconvincingenoughtoexplaintheoverall
declineofthecircadianclock,ateitherthecentralorperipherallevel.Theeffectsof
agingonbehaviouralandmolecularrhythmicityarelikelytobecomplex,itisassumed
tobetheresultsofdeclineatseverallevelsofthecircadianclockorganization(Figure
3-1).

INPUT MOLECULAR CLOCK OUTPUT
Perception
Transmission
Response
Central Clock Period
Phase
Rhythmicity
Light Overt Rhythms
AGING
Robust?
Weakened?
Figure 3-1. Overview of Existing Findings on the Aging of the Circadian Clock
Aging can affect any components within the circadian system, including the input pathway, the
molecular clockwork, and the output rhythms. Among each, impairment may occur at different
loci and to different extent. Although presumably no party can escape the influence of aging, the
weight of contribution should be diverse. The age-related decline in light entrainment as well as in
various parameters of overt rhythms have been reported, yet the changes of the clock gene
expression at the central and peripheral levels are still unclear.
107
Peripheral ClocksOverall Decline?
Tissue-dependent?

TheAgingoftheCircadianClock
108
3.3Objectives
Hereweaimedtoinvestigatehowthemolecularclock,inthebrainandintheperipheral
tissues,changesintrinsicallywithincreasingageunderconstantconditions.Wealso
examinedwhetherstrongentrainmentcues,suchasLDcyclesandLDcoupledwith
temperature cycles, rescue age-related functional decline of the clock. By using
luciferase reporters, we were able to measure real-time expression of clock gene
productsPERandTIM inyoung,middle-aged,andoldDrosophila fora longenough
durationoftime.

TheAgingoftheCircadianClock
109
3.4Methods
3.4.1ExperimentalDesign
Figure3-2.ExperimentalDesignoftheAgingStudy
InordertocomparethemolecularclockatbothcentralandperipherallevelsinyoungandagedDrosophila,
clock gene expression in flies with different age was measured by bioluminescence assay for 10 days
continuously,underthreeconditionsincludingDD,LDcycles(12:12,whitebarsindicatelightsonandblack
barsindicatelightsoff))withconstanttemperatureat25°C,andLDcouplingtemperaturecycles(lightonat
25°Candlightoffat21°C).

TheAgingoftheCircadianClock
110
3.4.2AnimalModel
Drosophila transgenic luciferase-reporter lines 8.0-luc, XLG-luc and tim-luc (refer to
ChapterTwo,Section2.4.2)wererearedongrowth-media(Carolina,USA)loadedin
standardplasticvials.Flieswerekeptat12:12LDcycleswithaconstanttemperature
of25˚C.DuringLDcycles,Zeitgebertime(ZT)isdefinedaslightonat8amNZST(ZT0)
andlightoffat8pmNZST(ZT12).Young(10-day-old),middle-aged(30-day-old)andold
(50-day-old)matedmaleflieswereused,allflieswithdifferentageswerekeptunder
identicalcircumstances.
We used both 8.0-lucand XLG-luc lines tomeasure PER expression.8.0-luc specifically
reflectsPERexpressioninsubsetsofthecentralclockdorsalneuronalclusters(DN1-3)
andLNd[Velerietal.,2003;Yoshiietal.,2009].XLG-lucreflectsPERexpressioninall
known per-expressing cells throughout the body, including central and peripheral
tissues[Velerietal.,2003].XLG-lucismainlyindicativeofperipheralclocksbecauseof
thesmallnumberofcentralclockneuronscomparedtothousandsofclockcellsinthe
rest of the body. A similar rule applies to tim-luc, which therefore reflects TIM
expressioninperipheralclocks[Beaveretal.,2002;Giebultowiczetal.,2000].

TheAgingoftheCircadianClock
111
3.4.3MeasurementofClockGeneExpression
Transgenic flies for real-time bioluminescence recording of clock gene per and tim
expressionwereusedinthisstudy,asdescribedpreviously inChapterTwo(Section
2.4.3)andillustratedinFigure3-3.Individualfliesweretransferredintoasinglewell
of white opaque 96-well microplates (Greiner Bio-one). Plates were loaded into a
Multimode Plate Reader (EnSpire, PerkinElmer) kept inside an incubator (Percival
ScientificInc,USA).ExperimentswereconductedinDD,LDcycles(12:12)atconstant
temperature(25°C),andLDcycles(12:12)coupledwithtemperaturecycles(lightonat
25°Candlightoffat21°C),respectively.

96-well Plate (side-view)
Ice
Drosophila Standard Vials
PCR Caps
96-well Plate with Flies
Film
Luciferin-mixed Media
112
Figure 3-3. Bioluminescence Assay Conducted in Adult Drosophila
Individual adult flies with known age were immobilized on ice and transferred into every other
well of white opaque 96-well microplates. Each well contained growth-media fortified with
luciferin. Flies were covered with PCR caps to limit their activity impact on bioluminescence signal
recording. Plates were sealed with plastic films and loaded into a Multimode Plate Reader.
PCR: polymerase chain reaction
Drosophila melanogaster

TheAgingoftheCircadianClock
113
3.4.4DataAnalysis
Datawereshowninmeanandstandarderror(Mean±SEM).Statisticalsignificanceofthe
amplitudeandacrophaseoftimeseriesdataamongyoung,middle-agedandoldflies
weredeterminedusingtwo-wayANOVAwithBonferroni’spost-hoctest[Rakshitetal.,
2012].

TheAgingoftheCircadianClock
114
3.5Results
3.5.1Age-relatedChangesofClockGenesPeriodExpression inCentralandPeripheral
ClocksandTimelessExpressioninPeripheralClocksUnderDD
Weanalysedtheamplitudeandacrophaseofbioluminescencesignalofeachdaytoshow
therhythmicityandlevelofclockgeneexpression,respectively. InDDtherhythmic
PERexpressionatcentrallevelin8.0-lucmiddle-agedandoldflieswascomparableto
that in theyoung flieswith robustoscillations (Figure3-4), showingnodecrease in
eitherrhythmicityorexpressionlevel.Incontrast,bothamplitudeandacrophaseof
PERatperipherallevelinXLG-luc(Figure3-5)andTIMintim-luc(Figure3-6)middle-
agedandoldfliesweresignificantlylowerthancorrespondingyoungones,showing
age-dependent decline in both rhythmicity and expression level. Despite that the
consumptionofluciferin(substrate)ledtothedecayofsignalwithineachagegroup,
bycomparingthesignalfromdifferentagegroupsatthesametimepoint,theeffect
ofagingonclockgeneexpressionwasstillevident.

Figure 3-4. Mean Bioluminescence Time-series Data From Young (10-day), Middle-aged (30-day) and Old (50-day) 8.0-luc Fruit Flies in DD
Bio
lum
ine
sce
nce
(CP
S)A
mp
litu
de
Acr
op
has
e
Time (day)
Time (day) Time (day)
115
Bioluminescence counts per second (CPS) is plotted as average of raw counts at 30-minute
intervals. Data are shown as mean ± SEM with error bars at each time point. 10-day cohort equals
black lines and columns, 30-day cohort equals mid-grey lines and columns, and 50-day cohort
equals light-grey lines and columns. 8.0-luc (reporting PER in the dorsal neurons, central clock) 30-
day cohort shows higher PER amplitude and acrophase than 10-day and 50-day cohorts (p<0.001
in all), and there is no difference between 10-day and 50-day cohorts (p=1.00 in both amplitude
and acrophase).
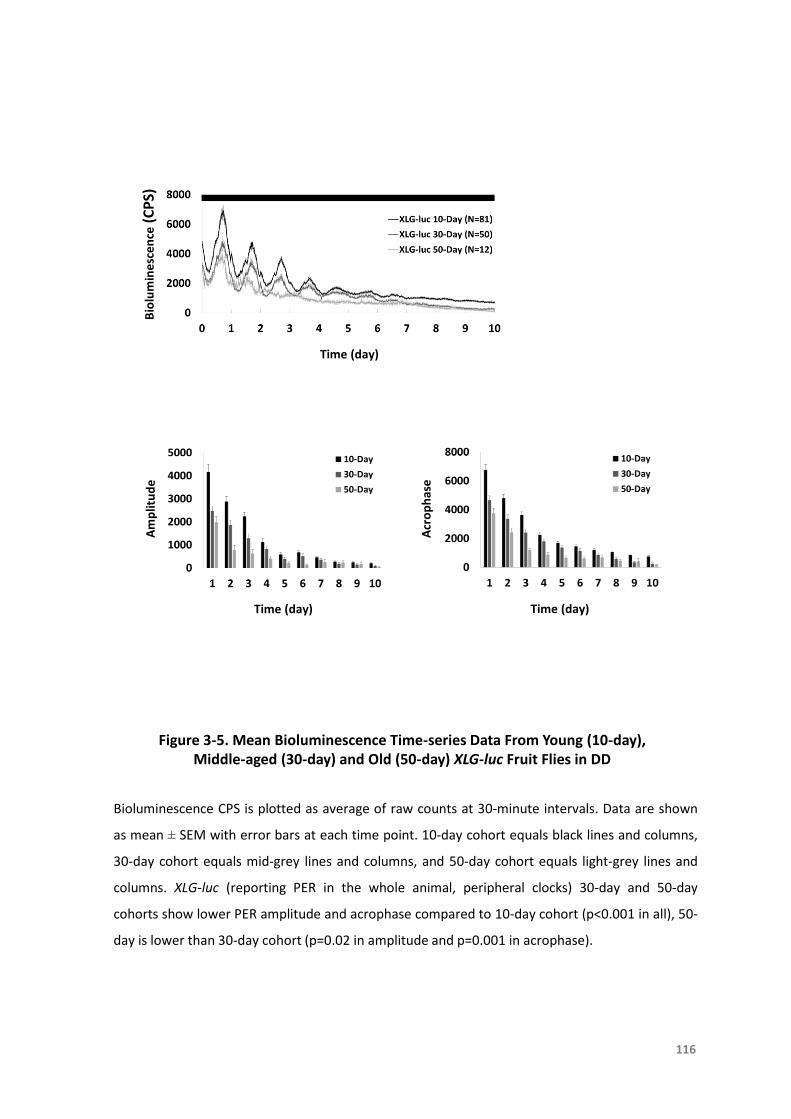
Figure 3-5. Mean Bioluminescence Time-series Data From Young (10-day), Middle-aged (30-day) and Old (50-day) XLG-luc Fruit Flies in DD
Bio
lum
ine
sce
nce
(CP
S)
Time (day)
Am
plit
ud
e
Acr
op
has
e
Time (day) Time (day)
116
Bioluminescence CPS is plotted as average of raw counts at 30-minute intervals. Data are shown
as mean ± SEM with error bars at each time point. 10-day cohort equals black lines and columns,
30-day cohort equals mid-grey lines and columns, and 50-day cohort equals light-grey lines and
columns. XLG-luc (reporting PER in the whole animal, peripheral clocks) 30-day and 50-day
cohorts show lower PER amplitude and acrophase compared to 10-day cohort (p<0.001 in all), 50-
day is lower than 30-day cohort (p=0.02 in amplitude and p=0.001 in acrophase).
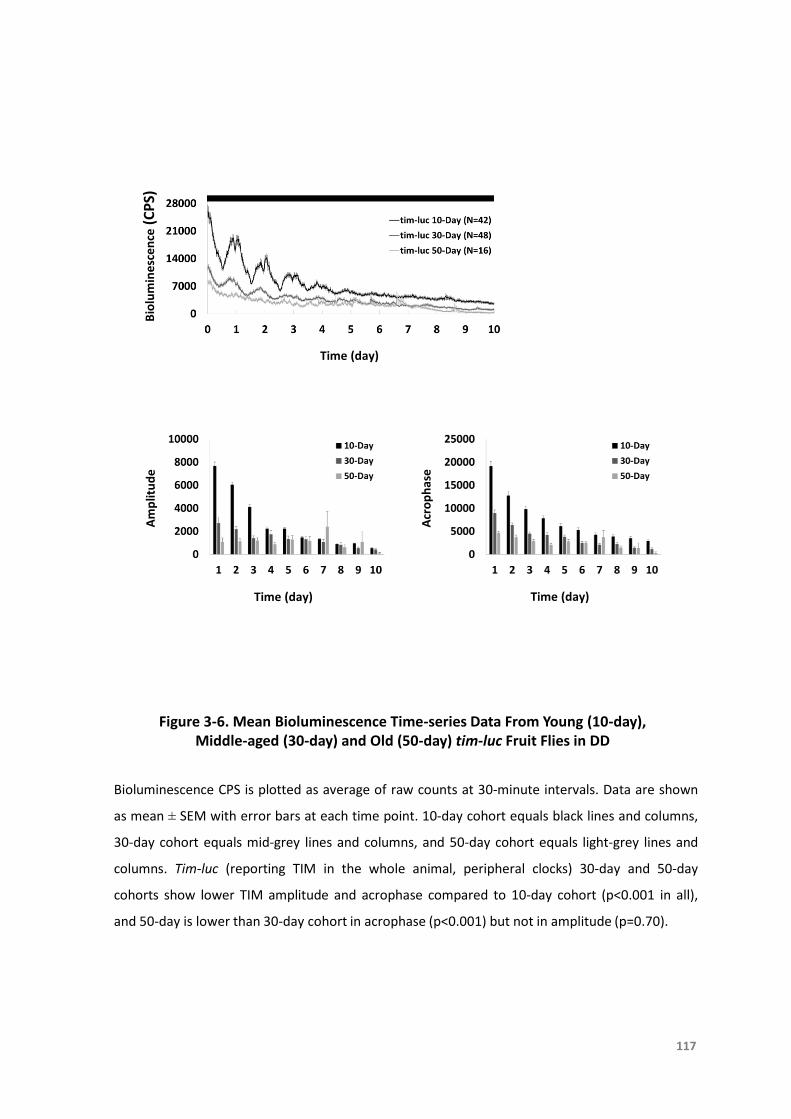
Figure 3-6. Mean Bioluminescence Time-series Data From Young (10-day), Middle-aged (30-day) and Old (50-day) tim-luc Fruit Flies in DD
Bio
lum
ine
sce
nce
(CP
S)A
mp
litu
de
Acr
op
has
e
Time (day)
Time (day) Time (day)
Bioluminescence CPS is plotted as average of raw counts at 30-minute intervals. Data are shown
as mean ± SEM with error bars at each time point. 10-day cohort equals black lines and columns,
30-day cohort equals mid-grey lines and columns, and 50-day cohort equals light-grey lines and
columns. Tim-luc (reporting TIM in the whole animal, peripheral clocks) 30-day and 50-day
cohorts show lower TIM amplitude and acrophase compared to 10-day cohort (p<0.001 in all),
and 50-day is lower than 30-day cohort in acrophase (p<0.001) but not in amplitude (p=0.70).
117

TheAgingoftheCircadianClock
118
3.5.2Age-relatedChangesofClockGenesPeriodExpression inCentralandPeripheral
ClocksandTimelessExpressioninPeripheralClocksUnderLDandLD/temperature
Cycles
Inordertotesttheinfluenceofentrainmentcuesonthisage-relatedeffect,flieswerekept
underregularLDcycleswithconstanttemperatureandLDcoupledwithtemperature
cyclesrespectively.InLDandLD/temperaturecyclestherhythmicPERexpressionat
centrallevelin8.0-lucmiddle-agedandoldflieswascomparabletothatintheyoung
flieswith robustoscillations (Figure3-7&3-10). Similar to that shown inDD,both
amplitudeandacrophaseofPERexpressionatperipherallevelinXLG-luc(Figure3-8)
andTIMexpressionatperipherallevelintim-luc(Figure3-9)middle-agedandoldflies
were significantly lower than correspondingyoungones in LD.Bothamplitudeand
acrophaseofPERexpression inXLG-luc (Figure3-11)andTIMexpression in tim-luc
(Figure3-12)old flieswere significantly lower thancorrespondingmiddle-agedand
youngonesinLD/temperaturecycles.

Figure 3-7. Mean Bioluminescence Time-series Data From Young (10-day), Middle-aged (30-day) and Old (50-day) 8.0-luc Fruit Flies in LD Cycles
Bio
lum
ine
sce
nce
(CP
S)A
mp
litu
de
Acr
op
has
e
Time (day)
Time (day) Time (day)
119
White bars represent lights on and black bars indicate lights off. Bioluminescence CPS is plotted as
average of raw counts at 60-minute intervals. Data are shown as mean ± SEM with error bars at
each time point. 10-day cohort equals black lines and columns, 30-day cohort equals mid-grey
lines and columns, and 50-day cohort equals light-grey lines and columns. The 8.0-luc 30-day and
50-day cohorts show higher PER amplitude than 10-day cohort (p<0.001 and p=0.001,
respectively). There is no difference between 30-day and 50-day cohorts in amplitude (p=1.00).
30-day cohort shows higher PER acrophase than 10-day and 50-day cohorts (p<0.001 and p=0.02,
respectively), and there is no difference between 10-day and 50-day cohorts (p=1.00).

Figure 3-8. Mean Bioluminescence Time-series Data From Young (10-day), Middle-aged (30-day) and Old (50-day) XLG-luc Fruit Flies in LD Cycles
Bio
lum
ine
sce
nce
(CP
S)
Time (day)
Am
plit
ud
e
Acr
op
has
e
Time (day) Time (day)
120
White bars represent lights on and black bars indicate lights off. Bioluminescence CPS is plotted as
average of raw counts at 60-minute intervals. Data are shown as mean ± SEM with error bars at
each time point. 10-day cohort equals black lines and columns, 30-day cohort equals mid-grey
lines and columns, and 50-day cohort equals light-grey lines and columns. The XLG-luc 30-day and
50-day cohorts show lower PER amplitude and acrophase compared to 10-day cohort (p<0.001 in
all), 50-day is lower than 30-day cohort (p<0.001 in amplitude and p=0.001 in acrophase).

Figure 3-9. Mean Bioluminescence Time-series Data From Young (10-day), Middle-aged (30-day) and Old (50-day) tim-luc Fruit Flies in LD Cycles
Bio
lum
ine
sce
nce
(CP
S)A
mp
litu
de
Acr
op
has
e
Time (day)
Time (day) Time (day)
White bars represent lights on and black bars indicate lights off. Bioluminescence CPS is plotted as
average of raw counts at 60-minute intervals. Data are shown as mean ± SEM with error bars at
each time point. 10-day cohort equals black lines and columns, 30-day cohort equals mid-grey
lines and columns, and 50-day cohort equals light-grey lines and columns. The tim-luc 30-day and
50-day cohorts show lower TIM amplitude and acrophase compared to 10-day cohort (p<0.001 in
all), 50-day is lower than 30-day cohort in amplitude (p<0.001) but not in acrophase (p=0.11).
121
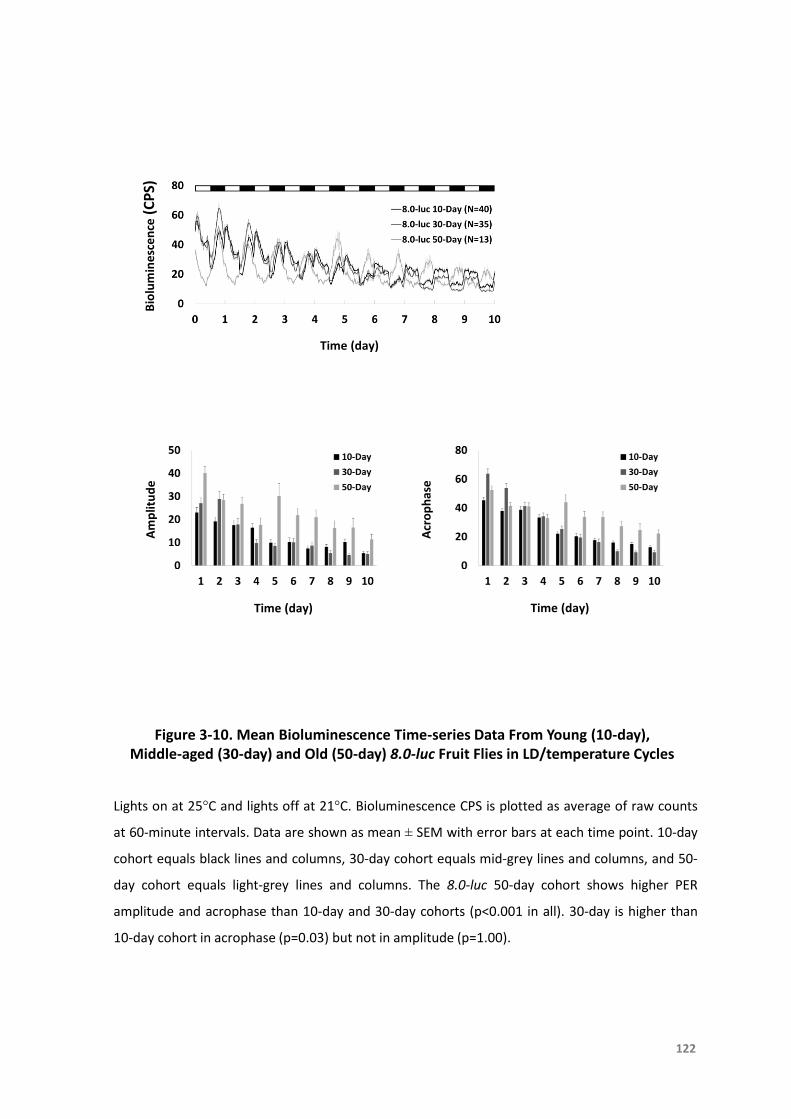
Figure 3-10. Mean Bioluminescence Time-series Data From Young (10-day), Middle-aged (30-day) and Old (50-day) 8.0-luc Fruit Flies in LD/temperature Cycles
Bio
lum
ine
sce
nce
(CP
S)A
mp
litu
de
Acr
op
has
e
Time (day)
Time (day) Time (day)
122
Lights on at 25°C and lights off at 21°C. Bioluminescence CPS is plotted as average of raw counts
at 60-minute intervals. Data are shown as mean ± SEM with error bars at each time point. 10-day
cohort equals black lines and columns, 30-day cohort equals mid-grey lines and columns, and 50-
day cohort equals light-grey lines and columns. The 8.0-luc 50-day cohort shows higher PER
amplitude and acrophase than 10-day and 30-day cohorts (p<0.001 in all). 30-day is higher than
10-day cohort in acrophase (p=0.03) but not in amplitude (p=1.00).
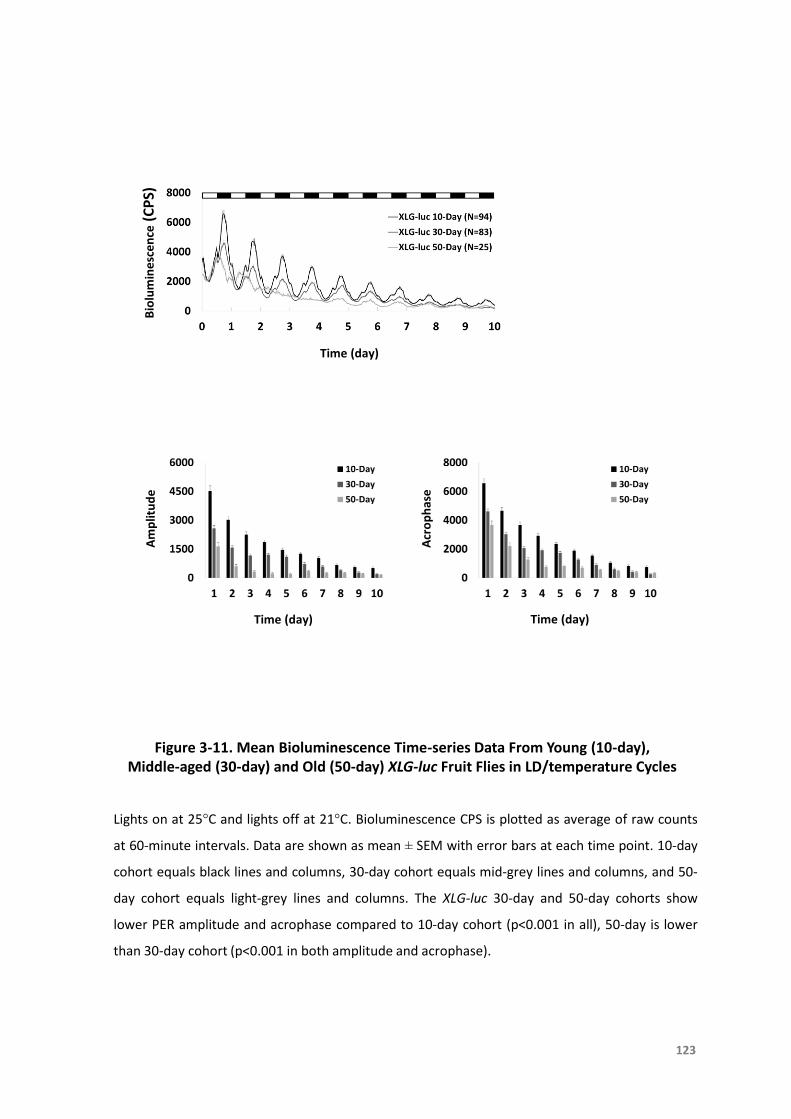
Figure 3-11. Mean Bioluminescence Time-series Data From Young (10-day), Middle-aged (30-day) and Old (50-day) XLG-luc Fruit Flies in LD/temperature Cycles
Bio
lum
ine
sce
nce
(CP
S)
Time (day)
Am
plit
ud
e
Acr
op
has
e
Time (day) Time (day)
123
Lights on at 25°C and lights off at 21°C. Bioluminescence CPS is plotted as average of raw counts
at 60-minute intervals. Data are shown as mean ± SEM with error bars at each time point. 10-day
cohort equals black lines and columns, 30-day cohort equals mid-grey lines and columns, and 50-
day cohort equals light-grey lines and columns. The XLG-luc 30-day and 50-day cohorts show
lower PER amplitude and acrophase compared to 10-day cohort (p<0.001 in all), 50-day is lower
than 30-day cohort (p<0.001 in both amplitude and acrophase).

Figure 3-12. Mean Bioluminescence Time-series Data From Young (10-day), Middle-aged (30-day) and Old (50-day) tim-luc Fruit Flies in LD/temperature Cycles
Bio
lum
ine
sce
nce
(CP
S)A
mp
litu
de
Acr
op
has
e
Time (day)
Time (day) Time (day)
Lights on at 25°C and lights off at 21°C. Bioluminescence CPS is plotted as average of raw counts
at 60-minute intervals. Data are shown as mean ± SEM with error bars at each time point. 10-day
cohort equals black lines and columns, 30-day cohort equals mid-grey lines and columns, and 50-
day cohort equals light-grey lines and columns. The tim-luc 50-day cohort shows lower TIM
amplitude and acrophase compared to 10-day and 30-day cohorts (p<0.001 in all), 30-day group is
lower than 10-day cohort in acrophase (p<0.001) but not in amplitude (p=1.00).
124

TheAgingoftheCircadianClock
125
3.6Discussion
Inourinvestigationoftheeffectofagingontheendogenousclock,wefoundareduction
inbothamplitudeandacrophaseofPERandTIMexpressionrhythmsthroughoutthe
bodyinDrosophilawithadvancingage.Thischangeingeneexpressionlargelyreflects
the decline of circadian clock function instead of the loss of cell numbers or body
composition.Drosophilabodymassisbelievedtobegenerallyconstantacrossallages
(from15daysto65days)[Laneetal.,2014],exceptforaremarkabledecreaseinthe
firstsevendaysaftereclosionandinextremelyoldflies[Djawdanetal.,1996].Ithas
been reported thatmiddle-agedmale flies showa lengthened free-runningperiod,
reducedactivitylevelanddiminishedrhythmstrengthincomparisonwithyoungflies
[Umezakietal.,2012].Weconcludethatthereductioninexpressionandrhythmicity
of clock genesperand tim is responsible for the decay of the circadian pattern of
activityinagedanimals.Thistheoryisfurthersupportedbygenemanipulationstudies
[GiebultowiczandLong,2015],inwhichover-expressionofclockgeneinspecificclock
neuronscanpartiallyrescuebehaviouralrhythmsandshortenedfree-runningperiod
inoldflies[Umezakietal.,2012].
Therearetwopossiblereasonsforthisresultinoverallreductionofclockgeneexpression
withincreasingage,aweakenedsignalfromthecentralclockorthedisruptionofthe
peripheraloscillators.WefoundthatrobustmolecularoscillationsofPERpersistinthe
dorsalneurons(DN1-3andLNd)ofthecentralclocks.Thisisconsistentwithaprevious

TheAgingoftheCircadianClock
126
findingofrobustPERcyclingins-LNv,LNdandDN1[Luoetal.,2012],butdifferentto
Umezaki’s2012studyinwhichtheamplitudeofPERandTIMexpressiondeteriorated
with advancing age in central clock cell groups (l-LNv, s-LNv, LNd, DN1 and DN2)
[Umezakietal.,2012].Ourfindingsupportstheideathatafunctionalcentraloscillator
is still intact in aged Drosophila. Interestingly, solid evidence showing molecular
changesfromtheSCNisalsolackingdespitethecentralclockstructuresaredifferent
betweeninsectsandmammals[Froy,2011;Nakamuraetal.,2015;Banksetal.,2016].
Weinferthattheeffectofagingisontheperipherallevelratherthanthecentrallevel.We
showedreductioninPERandTIMexpressionwithoutthetotallossofrhythmicityin
peripheralclocksthroughouttheflybodies,whichisinaccordwithpreviousfindings
intheflyheads[Krishnanetal.,2009;Luoetal.,2012;Rakshitetal.,2012].Although
restoration of CRYPTOCHROME (CRY) expression in all clock cells can significantly
enhance molecular and behavioural disturbance in old flies as well as delay
physiologicalaging,overexpressionofCRYincentralclockneuronsalonehasnosuch
effect[GiebultowiczandLong,2015].Itagainsuggeststhatperipheralclocksplayan
activeroleintheprogressofage-relatedclockdecline.Moreover,therearefindings
showingthatsomeperipheraloscillatorsalsodampenwithincreasingageinmammals
[MattisandSehgal,2016],withreducedexpressionorweakenedrhythmicityofseveral
clockgenesand/ortheirproteinproductsincertainextra-SCNregions[Froy,2011].
Together it points out an important fact that peripheral clocks are more likely to be
targetedbyagingandpossiblycontributetoage-associateddecline.Theunderlying

TheAgingoftheCircadianClock
127
mechanism is still unclear but the observed dampening signal of PER and TIM
expressioncancorrespondtoalossofcircadianrhythmicityinindividualcellsortoa
desynchronizationamongcellswithintheperipheralnetwork.Anincreasedtendency
towardsinternalandexternaldesynchronizationhasbeenreportedtobeacommon
feature of aging cellular networks [Farajnia et al., 2012]. Desynchrony among
peripheralclocksmaybeexplainedbymulti-oscillatortheoryandtheimpairmentof
thecouplingmechanisminoldage[WeinertandWeinert,1998;Yamazakietal.,2002].
Since loss of synchronization could be themain cause for the peripheral decline, one
strategy to repair the damaged network is by strengthening of entrainment.
Nonetheless, this way is also challenged by the potential impairment of the input
pathway, because entrainment and re-entrainment abilities are also compromised
withadvancingage[LiuandChang,2017].Evidenceinmammalssuggeststhataged
animalshavealesssensitiveclocktoentrainmentsignals[Froy,2011],suchastissue-
specificdeclineofreactionstolight[Taharaetal.,2017].
Correspondingly,wetestedwhetherstrongentrainmentcuescanimprovethedeclineof
molecularclocksinagedDrosophila.WeusedLDcyclesandtemperature-coupledLD
cycles to try reversing the deterioration of the clock in peripheral oscillators. The
disturbanceoftheinputpathwaymaybeapossibleexplanationwhyintheseolder
fliestheclockcannotbeeffectivelyentrainedbylightortemperature.Apreviousstudy
hasreportedthatLDcyclesaloneisnotsufficienttorescuedisturbanceoflocomotor
activityinagedflies,buttheuseofLD(12:12)couplingtemperaturecycles(25°C/21°C)

TheAgingoftheCircadianClock
128
turnsouttobeeffective[Luoetal.,2012].Ourfindingsuggestsaclock-independent
temperature effect may be involved. Lower temperatures (such as 21°C) prolong
longevityinDrosophilaandslowdowntheage-associatedsleepdecline,likelythrough
non-circadianmechanisms[Kohetal.,2006;Shawetal.,2008].

TheAgingoftheCircadianClock
129
3.7Conclusion
Aged animals commonly exhibit significant changes in circadianorganizationwhich are
evident at various levels: the overt rhythmicity of different variables becomes
imprecise or fragmented, and there are alterations in the function of the central
pacemakerandperipheraloscillators,particularlytheirnetworkproperties.Herewe
studytheagedclockandreportthatonthebasisofawell-functioningcentralclock,
there is a descending trend of clock gene products PER and TIM expression with
advancing age in peripheral clocks under constant condition and even strong
entrainment(Figure3-13).Ofcourse,becausetheagingprocessdoesnotimpactall
oscillatorytissuesinauniformmanner,studiesinthefuturewillfocusonnotonlythe
system-level alterations but also changeswithin individual oscillatory tissues, even
individualcells.
Since it lies in the fact that theperipheral clocks are seriously affectedby aging,more
emphasis should be placed on these local oscillators. Through the vast amount of
circadianbiology that they control, peripheral clocksdoubtlesslyplayan important
roleindailyphysiology.Specificdisruptionofclocksinperipheraltissuescancreatea
widerangeofpathologies[Marchevaetal.,2013],includingglucoseintoleranceand
diabetes,atherosclerosis,anddefectsinrenalandcardiacfunction[BrownandAzzi,
2013].Inaddition,dampeningofperipheralrhythmsispresumedtocontributetothe
increasedriskofmetabolicdiseasesintheelderly[HoodandAmir,2017].

TheAgingoftheCircadianClock
130
Ontheothersideofthecoin,robustandentrainedcircadianrhythmscouldpotentially
lead to better health and increased longevity [Froy, 2011]. We have proved that
Zeitgeberssuchaslightortemperatureoreventhecombinationofthesetwoarenot
sufficienttoresettheagedperipheralclocksinDrosophila,wherelightisanimportant
Zeitgeber for the entrainment of peripheral clocks. In mammals, food is a more
efficientagenttoentraintheperipheralclocksthustimedfooddeliverycouldserveas
effectivemeanstoresynchronizecircadiantiminginoldindividuals.Ithasalsobeen
reportedthatagedperipheralclocksbecomemoresensitivetofoodstimuli,makingit
anevenmoreidealentrainmentforagedsubjects[Taharaetal.,2017].Anumberof
studieshaveshownthatregularroutinesofmealtimeandexerciseimprovecircadian
consolidation in elderly individuals, addition with light regimen, these timing cues
mighthelpsubstituteforthelossofinternaldesynchrony[Brownetal.,2011].Thus,
non-pharmacologicalinterventionssuchaswell-organizedlifestyle,dietmodification,
and regular exercise should be underlined for their effect on strengthening and
maintainingsynchronyofthecircadiansystem[CornélissenandOtsuka,2017].

INPUT MOLECULAR CLOCK OUTPUT
Perception
Transmission
Response
Central Clock Period
Phase
Rhythmicity
Overt Rhythms
AGING
131
Peripheral Clocks
Light/temperature Rescue
Network Breakdown
Robust
Overall Decline
Figure 3-13. Overview of New Findings on the Aging of the Circadian Clock
Based on our findings from the current aging study in Drosophila, the central molecular clockwork
is still robustly ticking in aged flies while an overall decline has been shown in the peripheral
tissues throughout the body. A broken network is likely to be the underlying mechanism. The
strategy of strong entrainment fails to rescue the peripheral deterioration, thus the impairment of
the input pathway is implicated. Finally, the changes of the molecular clock may answer for the
behavioural disruption with increasing age.
Tissue-dependent

TheAgingoftheCircadianClock
132
3.8References
Arellanes-LiceaE,CaldelasI,DeIta-PérezD,Díaz-MuñozM.Thecircadiantimingsystem:arecent
additioninthephysiologicalmechanismsunderlyingpathologicalandagingprocesses.Aging
Dis.2014Jan9;5(6):406-18.DOI:10.14336/AD.2014.0500406.
AsaiM,YoshinobuY,KanekoS,MoriA,NikaidoT,MoriyaT,AkiyamaM,ShibataS.Circadianprofile
ofPergenemRNAexpression in the suprachiasmaticnucleus,paraventricularnucleus, and
pinealbodyofagedrats.JNeurosciRes.2001Dec15;66(6):1133-9.DOI:10.1002/jnr.10010.
BanksG,NolanPM,PeirsonSN.Reciprocalinteractionsbetweencircadianclocksandaging.Mamm
Genome.2016Aug;27(7-8):332-40.DOI:10.1007/s00335-016-9639-6.
BeaverLM,GvakhariaBO,VollintineTS,HegeDM,StanewskyR,GiebultowiczJM.Lossofcircadian
clockfunctiondecreasesreproductivefitnessinmalesofDrosophilamelanogaster.ProcNatl
AcadSciUSA.2002Feb19;99(4):2134-9.DOI:10.1073/pnas.032426699.
Brown SA,Azzi A. Peripheralcircadianoscillatorsinmammals. Handb Exp Pharmacol.
2013;(217):45-66.DOI:10.1007/978-3-642-25950-0_3.
Brown SA, Pagani L, Cajochen C, Eckert A. Systemic and cellular reflections on ageing and the
circadianoscillator:amini-review.Gerontology.2011;57(5):427-34.DOI:10.1159/000320673.
ChenCY,LoganRW,MaT,LewisDA,TsengGC,SibilleE,McClungCA.Effectsofagingoncircadian
patternsofgeneexpressioninthehumanprefrontalcortex.ProcNatlAcadSciUSA.2016Jan
5;113(1):206-11.DOI:10.1073/pnas.1508249112.
CornélissenG,OtsukaK.Chronobiologyofaging:AMini-review.Gerontology.2017;63(2):118-128.
DOI:10.1159/000450945.
DjawdanM,SugiyamaTT,SchlaegerLK.Metabolicaspectsofthetrade-offbetweenfecundityand
longevity in Drosophila melanogaster. Physiol Zool. 1996 Sep-Oct; 69(5):1176-95. DOI:
10.1086/physzool.69.5.30164252.
DriverC.ThecircadianclockinoldDrosophilamelanogaster.Biogerontology.2000;1(2):157-62.

TheAgingoftheCircadianClock
133
DuffyJF,ZittingKM,ChinoyED.Agingandcircadianrhythms.SleepMedClin.2015Dec;10(4):423-
34.DOI:10.1016/j.jsmc.2015.08.002.
FarajniaS,MichelS,DeboerT,vanderLeestHT,HoubenT,RohlingJH,RamkisoensingA,Yasenkov
R,MeijerJH.Evidenceforneuronaldesynchronyintheagedsuprachiasmaticnucleusclock.J
Neurosci.2012Apr25;32(17):5891-9.DOI:10.1523/JNEUROSCI.0469-12.2012.
FonsecaCostaSS,RippergerJA.Impactofthecircadianclockontheagingprocess.FrontNeurol.
2015Mar6;6:43.DOI:10.3389/fneur.2015.00043.
FroyO. Circadian rhythms, aging, and life span in mammals. Physiology (Bethesda). 2011
Aug;26(4):225-35.DOI:10.1152/physiol.00012.2011.
GiebultowiczJM,LongDM.Ageingandcircadianrhythms.CurrOpinInsectSci.2015Feb1;7:82-86.
DOI:10.1016/j.cois.2015.03.001.
Giebultowicz JM,Stanewsky R,Hall JC,Hege DM. Transplanted Drosophila excretory tubules
maintaincircadianclockcyclingoutofphasewiththehost.CurrBiol.2000Jan;10(2):107-10.
HoodS,Amir S. Theagingclock:circadian rhythmsandlaterlife. J Clin Invest.2017Feb
1;127(2):437-446.DOI:10.1172/JCI90328.
Koh K, Evans JM, Hendricks JC, Sehgal A. A Drosophila model for age-associated changes in
sleep:wake cycles. Proc Natl Acad Sci U S A. 2006 Sep 12;103(37):13843-7. DOI:
10.1073/pnas.0605903103.
Kondratova AA, Kondratov RV. The circadian clock and pathology of the ageing brain. Nat Rev
Neurosci.2012Mar7;13(5):325-35.DOI:10.1038/nrn3208.
KrishnanN,kretzschmarD,RakshitK,ChowE,GiebultowiczJM.Thecircadianclockgeneperiod
extends healthspan in aging Drosophila melanogaster. Aging (Albany NY). 2009 Nov
19;1(11):937-48.DOI:10.18632/aging.100103.
LaneSJ,FrankinoWA,ElekonichMM,RobertsSP.Theeffectsofageandlifetimeflightbehaviour
onflightcapacityinDrosophilamelanogaster.JExpBiol.2014May1;217(Pt9):1437-43.DOI:
10.1242/jeb.095646.
LiuF,Chang HC. Physiologicallinksofcircadian clockandbiological clockofaging. Protein Cell.
2017Jul;8(7):477-488.DOI:10.1007/s13238-016-0366-2.

TheAgingoftheCircadianClock
134
LuoW,ChenW-F,YueZ,ChenD, SowcikM,SehgalA, ZhengX.Old flieshavea robust central
oscillatorbutweakerbehavioralrhythmsthatcanbeimprovedbygeneticandenvironmental
manipulations.AgingCell.2012Jun;11(3):428-38.DOI:10.1111/j.1474-9726.2012.00800.x.
MarchevaB,RamseyKM,PeekCB,Affinati A,Maury E,Bass J. Circadian clocks andmetabolism.
HandbExpPharmacol.2013;(217):127-55.DOI:10.1007/978-3-642-25950-0_6.
MattisJ,SehgalA.Circadianrhythms,sleep,anddisordersofaging.TrendsEndocrinolMetab.2016
Apr;27(4):192-203.DOI:10.1016/j.tem.2016.02.003.
Nakamura TJ, NakamuraW, Tokuda IT, Ishikawa T, Kudo T, Colwell CS, Block GD. Age-related
changesinthecircadiansystemunmaskedbyconstantconditions(1,2,3).eNeuro.2015Sep
22;2(4).pii:ENEURO.0064-15.2015.DOI:10.1523/ENEURO.0064-15.2015.
NakamuraTJ,NakamuraW,YamazakiS,KudoT,CutlerT,ColwellCS,BlockGD.Age-relateddecline
in circadianoutput. JNeurosci.2011 Jul13;31(28):10201-5.DOI:10.1523/JNEUROSCI.0451-
11.2011.
OishiK,KoyanagiS,OhkuraN.CircadianmRNAexpressionofcoagulationandfibrinolyticfactorsis
organ-dependentlydisrupted inagedmice. Exp Gerontol.2011 Dec;46(12):994-9. DOI:
10.1016/j.exger.2011.09.003.
PolidarováL,SládekM,NovosadováZ,SumováA.Agingdoesnotcompromiseinvitrooscillation
ofthesuprachiasmaticnucleibutmakesitmorevulnerabletoconstantlight.ChronobiolInt.
2017;34(1):105-117.DOI:10.1080/07420528.2016.1242491.
Rakshit K,Giebultowicz JM.Cryptochrome restoresdampened circadian rhythmsandpromotes
healthspaninagingDrosophila.AgingCell.2013Oct;12(5):752-62.DOI:10.1111/acel.12100.
Rakshit K, Krishnan N, Guzik EM, Pyza E, Giebultowicz JM. Effects of aging on the molecular
circadian oscillations in Drosophila. Chronobiol Int. 2012 Feb;29(1):5-14. DOI:
10.3109/07420528.2011.635237.
SellixMT,Evans JA,Leise TL,Castanon-Cervantes O,Hill DD,DeLisser P,Block GD,Menaker M,
Davidson AJ. Aging differentially affects the re-entrainment response of central and
peripheralcircadian oscillators. J Neurosci.2012Nov 14;32(46):16193-202. DOI:
10.1523/JNEUROSCI.3559-12.2012.

TheAgingoftheCircadianClock
135
ShawP,OcorrK,BodmerR,OldhamS.Drosophilaaging2006/2007.ExpGerontol.2008Jan;43(1):5-
10.DOI:10.1016/j.exger.2007.10.008.
TaharaY,TakatsuY,ShiraishiT,KikuchiY,YamazakiM,MotohashiH,MutoA,SasakiH,Haraguchi
A,Kuriki D,Nakamura TJ,Shibata S. Age-relater circadian disorganization caused by
sympathetic dysfunctioninperipheralclock regulation. NPJ Aging Mech Dis.2017 Jan
5;3:16030.DOI:10.1038/npjamd.2016.30.
Umezaki Y, Yoshii T, Kawaguchi T, Helfrich-Förster C, Tomioka K. Pigment-dispersing factor is
involvedinage-dependentrhythmchangesinDrosophilamelanogaster.JBiolRhythms.2012
Dec;27(6):423-32.DOI:10.1177/0748730412462206.
Veleri S,Brandes C,Helfrich-Förster C,Hall JC,Stanewsky R. Aself-sustaining,light-entrainable
circadianoscillatorintheDrosophilabrain.CurrBiol.2003Oct14;13(20):1758-67.
WeinertD,WaterhouseJ.Thecircadianrhythmofcoretemperature:effectsofphysicalactivity
andaging.PhysiolBehav.2007Feb28;90(2-3):246-56.DOI:10.1016/j.physbeh.2006.09.003.
WeinertH,WeinertD.Circadianactivityrhythmsoflaboratorymiceduringthelastweeksoftheir
life.BiolRhythmRes.1998;29(2):159-78.
WeinertH,WeinertD,Schurov I,MaywoodES,HastingsMH. Impairedexpressionof themPer2
circadian clock gene in the suprachiasmatic nuclei of aging mice. Chronobiol Int.2001
May;18(3):559-65.
WyseCA,CooganAN.ImpactofagingondiurnalexpressionpatternsofCLOCKandBMAL1inthe
mousebrain.BrainRes.2010Jun14;1337:21-31.DOI:10.1016/j.brainres.2010.03.113.
Yamazaki S, StraumeM,TeiH, Sakaki Y,MenakerM,BlockGD.Effectsof agingon central and
peripheral mammalian clocks. Proc Natl Acad Sci U S A. 2002 Aug 6;99(16):10801-6. DOI:
10.1073/pnas.152318499.
Yoshii T, Wulbeck C, Sehadova H, Veleri S, Bichler D, Stanewsky R, Helfrich-Förster C. The
neuropeptide pigment-dispersing factor adjusts period and phase of Drosophila's clock. J
Neurosci.2009Feb25;29(8):2597-610.DOI:10.1523/JNEUROSCI.5439-08.2009.

赵嘉
如若尘埃

CHAPTER 4
Death
136

A Timeless Chaos Before Death
The Aberrant Expression of Clock
Genes During the Last Days of Life in
Drosophila melanogaster

TheBreakdownoftheCircadianClock
138
Thedatafromthischapterhavebeenpublished***(pleaseseeAppendixfordetails).
***ZhaoJ,WarmanGR,CheesemanJF.Clockgeneexpressionandlocomotoractivitypredict
deathinthelastdaysoflifeinDrosophilamelanogaster.SciRep.2018Aug9;8(1):11923.
DOI:10.1038/s41598-018-30323-x.

TheBreakdownoftheCircadianClock
139
4.1Abstract
Thecircadianclockagesandeventuallydeterioratesbut findingsonhowthemolecular
clockaltersbeforedeatharerare.Weaimedtostudyhowthecircadianclockchanges
priortodeathbycontinuousrecordingtheexpressionoftwokeyclockgenesperiod
andtimelessinvivoinDrosophilamelanogasteruntilthelastdaysoflife.Wereported
anovelmarkerofimminentdeathintheexpressionoftheclockgeneTIMELESS.Inthe
daysimmediatelyprecedingdeath,TIMELESSexpressionincreasedtoatleast150%of
previous acrophase values (88.0%of n=217) and circadian rhythmicity in TIMELESS
expressiondisappeared. Importantlythismarker intheexpressionofTIMELESSwas
notagedependentandpredicteddeathequallywellinfruitfliesofdifferentagesand
under lightandtemperaturecycles.Weconsider this transientaberrantclockgene
expression to be the centralmechanism of the disturbance in circadian behaviour
beforedeath(82.7%ofn=342). Inconclusion,thecircadianclockbreaksdowndays
before death, resulting in arrhythmic activity as well as arrhythmic increase of
expressionofclockgenes.

TheBreakdownoftheCircadianClock
140
4.2Introduction
Deathisthecessationofbiologicalfunctionsthatmaintainhomeostasisandsustainaliving
organism[PozhitkovandNoble,2017].Itwasproposedacenturyagothatonlymulti-
cellularanimalshavetheconceptof“naturaldeath”becauseunicellularorganismsare
knowntobeimmortal[Pearl,1922].Thedeathofananimalmeansthatitssystems,
organs,tissues,andcellsstopfunctioningasawhole,whichimpliesthattheindividual
partsmaystillbealiveatthetimewhenthebodydies[PozhitkovandNoble,2017].As
consequence,studyingdeathatanentireanimallevelseemstobeareasonablestep
tostartwith.Weinterestedinthequestionssuchaswhathappenstothecircadian
clockwhentheanimalisapproachingdeath?Inturn,howthesechangesoftheclock
contributetotheprocessofdeath?
Althoughthedetailsarecurrentlynotfullyunderstood,increasingevidencesuggeststhat
thecircadianclockisoneofthedeterminantsoflongevity[HoodandAmir,2017].First,
abetterfunctioningclockitselfspeaksofalongerlifespan,supportedbythefactthat
fliesdisplayingrhythmiclocomotoractivitylivelongerthanarrhythmicones[Kumaret
al.,2005].Secondly,thealignmentbetweenintrinsicfree-runningperiod(tau)andthe
environmental cycles is awell-acceptedpredictorof lifespan. In aword,organisms
keptinanenvironmentthatbestmatchestheirinternalcircadianclockoutcompete
the others in longevity from the study of cyanobacteria [Ouyang et al., 1998] and
Drosophila[PittendrighandMinis,1972;KlarsfeldandRouyer,1998].Thesamerule

TheBreakdownoftheCircadianClock
141
appliestomammals.Wildtypemicewithafree-runningperiodofapproximately24
hoursshowreducedlongevitywhenhousedunderartificialshortdays(T-cycles)[Park
et al., 2012]. Tau mutant hamsters with a free-running period of 22 hours show
reduced longevitywhen housed in 24-hour light-dark (LD) cycles [Hurd and Ralph,
1998].Naturally,whenthesemutantsarekepteitherintheconstantdarkness(DD)or
under22-hourT-cycles, longevityandhealthareunaffected[HurdandRalph,1998;
Martino et al., 2008]. Finally, mammalian body temperature which is dominantly
regulated by the circadian clock displays a complex relationship with longevity. In
general,reductionofbodytemperatureusuallyindicatesanincreaseoflifespan[Keil
etal.,2015].
Theconnectionbetweenthecircadianclockanddeathhasbeenshowninafewstudies,
withclock-controlledbehaviourchangingbeforedeath.Forinstance,agedfliesintheir
lastdaysoflifehavebeenfoundtoexhibitarrhythmicityinbehaviour[Luoetal.,2012].
Inlaboratorymammals,impairmentofseveralparametersofcircadianrhythms,such
as body temperature and locomotor activity, is considered amarker for imminent
death[Bassoetal.,2016].Severalmammalianstudieshaveshownamarkeddecrease
in the amount and amplitude of activity as well as a loss of ability to maintain
entrainmentunderLDcyclesastheanimalsapproachdeath.Anumberofanimalslose
their circadian rhythm completely [Davis and Viswanathan, 1998; Morin, 1988;
WeinertandWeinert,1998].

TheBreakdownoftheCircadianClock
142
Withgeneralacceptancethatdeclineofthecircadianclockisareliablemanifestationof
aging,itisnothardtoimaginethatcollapseoftheclockcouldbeapotentialindicator
ofdeath.Nonetheless,therehasbeenverylittleinvestigationintothemechanismof
thisphenomenonpriortodeath.
Presumably,differentlevelsoftheclockorganizationmaybeinvolvedinthemechanism
ofdeath. There is strongevidence showing that the suprachiasmaticnucleus (SCN)
playsacrucialroleinagingandlongevityinmammals.Oneoftheexamplesofthisis
that an extended lifespan has been observed in aged animals given foetal
suprachiasmatic implantswhichrestorehigheramplituderhythms[HurdandRalph,
1998]. Aside from the central clock and the overt rhythms it controls, another
promisingcandidate is themolecularclockand itspropertygenesattheperipheral
level. Interesting enough, system-wide knockouts of clock genes create so-called
“acceleratedaging”phenotypesinbothDrosophilaandmice[HoodandAmir,2017].
Forinstance,nullmutationoftheperiod(per)gene[Krishnanetal.,2009]ortimeless
(tim)gene[Vaccaroetal.,2017]orcyclegene[Hendricksetal.,2003]inDrosophilais
associatedwithsignificantlyshorterlifespan,aswellasimpairedlocomotorrhythms.
Likewise,deficiencyoftheBMAL1orCLOCKproteinsignificantlyaffectslongevityand
results in the development of age-specific pathologies, such as sarcopenia,
osteoporosisandcataracts[Kondratovetal.,2006].DoublemutantsofPeriod1and
Period2 cause an arrhythmic phenotype togetherwith premature aging conditions
[Lee,2005].Anintriguingideaisalsorevealed,thatchangesinclockgeneexpression

TheBreakdownoftheCircadianClock
143
maybeimplicatednotonlyinage-associatedchangesincircadianrhythmicity,butalso
morebroadlyintheprocessesofagingoverall[HoodandAmir,2017].
A recent study has reported an increase of specific groups of genes after death in
vertebrates,despitemostofthetranscripts(99%)decreaseinabundancewithpost-
mortemtime[PozhitkovandNoble,2017].Thesegenesareassociatedwithsurvival
and stress compensation, such as heat shock, hypoxia-related, oxidative stress,
immune and apoptotic proteins. Others are associated with epigenetic factors,
developmental control, and cancer [Pozhitkov and Noble, 2017]. Accordingly, we
assumethatdeathisanextremestressandtheultimatechallengeforsurvival,sothat
certaingeneticregulatoryprogramsshouldbeactivatedtomeetthebodyneeds,and
sensibleenough,areverylikelytostartincreasingbeforedeath.Ifthisistrue,among
allgenes,aretheclockgeneschosentobeturnedonatthemostcriticaltiming?

TheBreakdownoftheCircadianClock
144
4.3Objectives
Withthecurrentlimitedknowledgeofthephenomenonandpossiblemechanismonthe
process of death (Figure 4-1),we aimed to investigate how themolecular clock in
Drosophila changes intrinsically before death under constant conditions, aswell as
under entrainment cues such as LD and temperature cycles. By using transgenic
luciferase-reporterlines,wewereabletocontinuouslymeasurereal-timeexpression
ofclockgeneproductsPERIOD(PER)andTIMELESS(TIM)inDrosophilauntiltheydied.
Meanwhile, rhythms of locomotor activity of wild type flies were recorded to
investigatethesimultaneouschangesatthebehaviourallevel.

DeathBefore After
Overt RhythmsLocomotor Activity
Body Temperature
Loss of Output Rhythmicity Rise of Gene Expression
?
Genes
Survival
Stress Compensation
Development Control
Cancer
CellOrganBody Cell Organ
Figure 4-1. Overview of Existing Findings on Changes of the Clock Before Death
The disintegration of an organism at the body level leads to the irreversible destiny of death,
when some of the cells and organs might still vital and functional at this stage. A specific group of
genes have been detected to increase shortly after death, yet the meaning is largely unknown.
The loss of circadian rhythms such as behavioural arrhythmicity can be used as markers of
imminent death, though whether the expression of clock genes shows corresponding change
needs to be further explored.
145

TheBreakdownoftheCircadianClock
146
4.4Methods
4.4.1ExperimentalDesign
Figure4-2.ExperimentalDesignoftheDeathStudy
InordertofindoutthemolecularandbehaviouralchangesofthecircadianclockinDrosophilabeforedeath,
behavioural rhythms of wild type flies with different age going through LD entrainment and DD were
measuredbyDAMsystem,clockgeneexpressionoftransgenicflieswasmeasuredbybioluminescenceassay
underDD,LDcycleswithconstanttemperatureandLDcouplingtemperaturecycles,respectively.

TheBreakdownoftheCircadianClock
147
4.4.2AnimalModel
Drosophilatransgenicluciferase-reporterlinesXLG-lucandtim-luc(refertoChapterTwo,
Section2.4.2)andwildtypeflieswererearedongrowth-media(Carolina,USA)loaded
instandardplasticvials.Flieswerekeptat12:12LDcyclesandattemperature25°C,
and transferred to freshvialsonceaweek.During LDcycles,Zeitgeber time (ZT) is
definedaslightonat8amNZST(ZT0)andlightoffat8pmNZST(ZT12).WeusedXLG-
luc and tim-luc flies to measure PER and TIM expression in peripheral tissues,
respectively.
Flieswerecollectedwithin2daysofeclosionandsurvivalofmatedmalefliesinvialswere
monitoreduntiltheirdeath,showingaveragelongevityof50-70daysat25°C.Itwas
consistent with most studies reporting 50-80 days as the maximum lifespan of
Drosophilainordinarylaboratorycircumstance[Grotewieletal.,2005].Young(10-day-
old),middle-aged(30-day-old)andold (50-day-old)matedmale flieswereused.All
fliesofdifferentageswerekeptunderidenticalcircumstances.

TheBreakdownoftheCircadianClock
148
4.4.3MeasurementofClockGeneExpression
Transgenic flies for real-time bioluminescence recording of clock gene per and tim
expressionwereusedinthisstudy,asdescribedpreviouslyinChapterThree(Section
3.4.3). Individual flies were transferred into a single well of white opaque 96-well
microplates (Greiner Bio-one). Plates were loaded into a Multimode Plate Reader
(EnSpire, PerkinElmer) kept inside an incubator (Percival Scientific Inc, USA).
ExperimentswereconductedinDD,LDcycles(12:12)atconstanttemperature(25°C),
andLDcycles(12:12)coupledwithtemperaturecycles(lightonat25°Candlightoffat
21°C),respectively.Datawerecontinuouslyrecordeduntilmostfliesdied.

TheBreakdownoftheCircadianClock
149
4.4.4MonitoringofLocomotorActivity
Locomotor activity of wild type flies was monitored automatically using Trikinetics
DrosophilaActivityMonitors(Waltham,MA,USA),aspreviouslydescribedinChapter
Two(Section2.4.4). Individual flieswithknownage(10-day-old,30-day-oldand50-
day-old)weretransferredintoglasstubes(8cmlongand5mmdiameter)withgrow
mediaatoneend,sealedwithplasticplugs,andpluggedwithcottonattheotherend.
Locomotor activity, temperature and relative humidity were recorded every five
minutesforanentrainingtimeof13daysinLDcyclesandtheninDDuntilmostflies
died.

TheBreakdownoftheCircadianClock
150
4.4.5DataAnalysis
Datawereshowninmeanandstandarderror(Mean±SEM),andp<0.05wasconsidered
significantly different. ClockLab (Actimetrics, USA) was used for the analysis of
rhythmicityinlocomotoractivityofwildtypeflies,andrhythmstrengthwasmeasured
by24-hourrelativefastFouriertransform(FFT)values[Kohetal.,2006].

TheBreakdownoftheCircadianClock
151
4.5Results
4.5.1MolecularMarkerofImminentDeathinBothPERIODandTIMELESSinPeripheral
Clocks
WefoundanincreaseofTIMsignalintim-lucflies(Figure4-3)andPERsignalinXLG-luc
(Figure4-4)attheperipherallevelwhichlastedforseveraldayspriortodeath,before
eventuallyvanishingwhentheanimalsdied.Theincreaseofsignalwasdefinedasat
least 50%higher than the acrophase of the preceding cycle. Prior to death, 87.8%
(n=82)tim-lucfliesrearedinDDshowedanincreaseinsignaloveratimeof86.2±2.2
hours, 93.5% (n=62) in LD cycles showed 98.4±3.4 hours, and 83.6% (n=73) in
LD/temperaturecyclesshowed90.0±3.0hours.Theageofdeathofthesefliesranged
from20to60days.Priortodeath,34.2%(n=79)XLG-lucfliesrearedinDDshowedan
increase in signalovera timeof84.3±5.2hours,54.3% (n=46) in LDcycles showed
81.4±3.8hours,and56.3%(n=48)inLD/temperaturecyclesshowed81.3±4.6hours.
Theageofdeathofthesefliesrangedfrom20to50days.

Age
(d
ay)
AB
Bioluminescence (CPS)
Age
(d
ay)
Bioluminescence (CPS)
Age
(d
ay)
Age
(d
ay)
Bioluminescence (CPS)
Bioluminescence (CPS)
CD
Age
(d
ay)
EF
Bioluminescence (CPS)
Age
(d
ay)
Bioluminescence (CPS)Fi
gure
4-3
. Rep
rese
nta
tive
Bio
lum
ines
cen
ce T
ime
-se
ries
Dat
a o
f tim-luc
Fru
it F
lies
Sho
win
g A
ber
ran
t TI
M S
ign
al a
t th
e La
st D
ays
of
Life
Bioluminescence
counts
per
second(CPS)
isplotted
asraw
counts
(greylines)andas
asix-hourmovingaverage(black
lines).(A)A10-day-old
and(B)a30-day-old
flyin
DD
displayarhythmicbutdescendingTIM
signaluntilseveraldaysbefore
theirdeathsat
22daysand42days,increasesofTIM
lasted
for87hours
and78hours
areobserved
,
respectively.(C)A10-day-oldand(D)a30-day-oldflyinLD
cycles
showincreasesofTIM
lasted
for91hoursand75hoursbefore
theirdeathsat
27daysand44days.(E)A10-
day-oldand(F)a30-day-oldflyinLD
/tem
perature
cycles
showincreasesofTIM
lasted
for91hoursand77hoursbefore
theirdeathsat
38daysand48days,respectively.
15
2
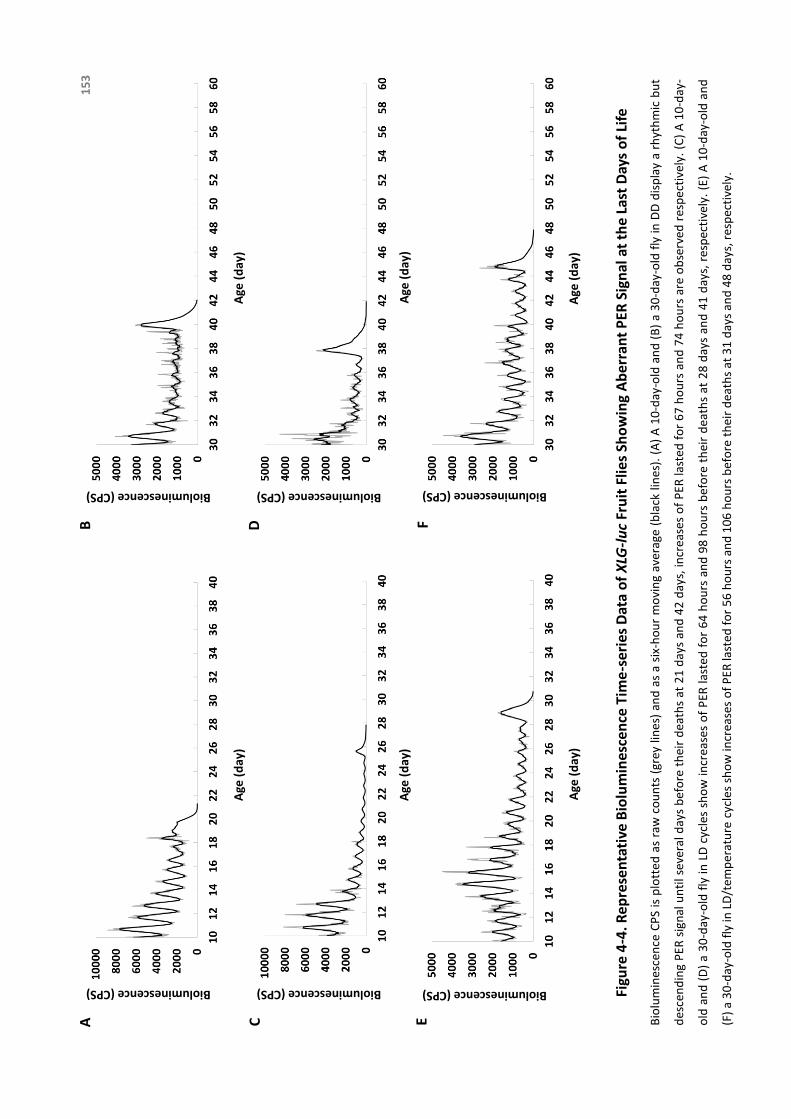
Age
(d
ay)
AB
Bioluminescence (CPS)
Age
(d
ay)
Bioluminescence (CPS)
Age
(d
ay)
Age
(d
ay)
Bioluminescence (CPS)
Bioluminescence (CPS)
CD
Age
(d
ay)
EF
Bioluminescence (CPS)
Age
(d
ay)
Bioluminescence (CPS)Fi
gure
4-4
. Rep
rese
nta
tive
Bio
lum
ines
cen
ce T
ime
-se
ries
Dat
a o
f XLG
-luc
Fru
it F
lies
Sho
win
g A
ber
ran
t P
ER S
ign
al a
t th
e La
st D
ays
of
Life
Bioluminescence
CPSisplottedas
rawcounts(greylines)andas
asix-hourmovingaverage(black
lines).(A)A10-day-oldand(B)a30-day-oldflyinDDdisplayarhythmicbut
descendingPER
signaluntilseveraldaysbefore
theirdeathsat
21daysand42days,increasesofPER
lasted
for67hoursand74hoursareobserved
respectively.(C)A10-day-
old
and(D)a30-day-oldflyinLD
cycles
showincreasesofPER
lasted
for64hoursand98hoursbefore
theirdeathsat
28daysand41days,respectively.(E)A10-day-old
and
(F)a30-day-oldflyinLD
/tem
perature
cycles
showincreasesofPER
lasted
for56hoursand106hoursbefore
theirdeathsat
31daysand48days,respectively.
15
3

TheBreakdownoftheCircadianClock
154
Inordertoconfirmthattheincreaseofsignaloccurredbeforedeath,wetestedhowlong
the bioluminescence signal would last after death under the same condition in a
numberofflies.Decapitatedtim-lucfliesshoweda11.4±1.6hourafter-deathsignal
(n=40) and XLG-luc flies showed 7.8±1.0 hours (n=41). In summary, immediately
precedingdeathmosttim-luc flies(88.0%,n=217)demonstratedan increase inTIM
signalthatlastedfor91.2±1.7hoursonaverageindependentofageorenvironment.
AtitsheightthiselevatedTIMsignalwasonaverage3.8timesashighastheprevious
acrophase (Figure 4-5A), and not concentrated at any particular ZT calculated by
RayleighTest(Figure4-5B).IncontrastthisincreaseinTIMwasnotreflectedinPERas
notasmanyfliesinXLG-luc(45.7%,n=173)showedanincreaseof82.3±2.6hourson
averagebeforedeath.AtitsheightthiselevatedPERsignalwasonaverage2.4times
ashighasthepreviousacrophase(Figure4-6A),andconcentratedwithmeanphase
aroundZT18toZT20,similartopreviouscycles(Figure4-6B).

Figure 4-5. Mean Bioluminescence Time-series Data from the Last Ten Days of Life in tim-luc Fruit Flies Showing Elevated Signals Before Death
AB
iolu
min
esc
en
ce (
CP
S)
10 9 8 7 6 5 4 3 2 1 0
Days before death
Bio
lum
ine
sce
nce
(C
PS)
B
155
(A) X-axis indicates the days before death. Black solid line is the average signal from the last five-
days of life, and grey solid line is from the last six to ten days of life with an exponential trendline
shown as grey dash line. Tim-luc flies (n=119) show circadian rhythms of TIM expression from day
10 to day 6 before death with a descending trend. The aberrant signal from the last five days
(average height value shown in black column) breaks the circadian pattern and shows a
remarkable increase (p<0.001 by non-parametric Wilcoxon signed-rank test, compared to the
average acrophase from the preceding cycle shown in grey column). (B) The circular distributions
of the elevated signals before death in a 24-hour clock face. Black solid dots around clock face
indicate individual flies in LD cycles and hollow dots in LD/temperature cycles. ZT (where ZT0 is
lights on and ZT12 is lights off) is labelled. Flies show random distribution in LD (n=58, p=0.37) and
in LD/temperature (n=61, p=0.44).

Figure 4-6. Mean Bioluminescence Time-series Data from the Last Ten Days of Life in XLG-luc Fruit Flies Showing Elevated Signals Before Death
AB
iolu
min
esc
en
ce (
CP
S)
10 9 8 7 6 5 4 3 2 1 0
Days before death
Bio
lum
ine
sce
nce
(C
PS)
B
156
(A) X-axis indicates the days before death. Black solid line is the average signal from the last five-
days of life, and grey solid line is from the last six to ten days of life with an exponential trendline
shown as grey dash line. XLG-luc flies (n=52) show circadian rhythms of PER expression from day
10 to day 6 before death with a descending trend. The aberrant signal from the last five days
(average height value shown in black column) follows the circadian pattern and shows a
remarkable increase (p<0.001 by non-parametric Wilcoxon signed-rank test, compared to the
average acrophase from the preceding cycle shown in grey column). (B) The circular distributions
of the elevated signals before death in a 24-hour clock face. Black solid dots around clock face
indicate individual flies in LD cycles and hollow dots in LD/temperature cycles. ZT (where ZT0 is
lights on and ZT12 is lights off) is labelled. Flies show significant concentrated distribution with
mean phase pointing at ZT18 in LD (n=25, p<0.001) and at ZT20 in LD/temperature (n=27,
p=0.007).

TheBreakdownoftheCircadianClock
157
4.5.2DownstreamBehaviouralChangesofImminentDeath
Locomotoractivitychangedinthelastfewdaysbeforedeath,inaccordwiththeaberrant
PERandTIMexpressionoccurringatasimilartime.82.7%ofwellentrainedflies(n=342)
at different ages lost the circadian pattern of behavioural rhythms and became
arrhythmic over a period of 3.7±0.1 days before death (Figure 4-7). Arrhythmic
locomotoractivitywasobservedin82.9%inflieswhichstartedtobeentrainedand
recordedfrom10-day-old(n=146),85.1%infliesfrom30-day-old(n=148),and75.0%
in flies from50-day-old (n=48),showingnosignificantdifferencebyChi-squaretest
(p=0.27).Theageofdeathofthesefliesrangedfrom20to70days.24-hourrelative
FFTvalueswereusedasameasureofcircadianrhythmstrengthandFFTvalue<0.01
wasconsideredarrhythmic.FliesshowedanaverageFFTvalueof0.050±0.002under
LDentrainmentand0.039±0.002under following free-running inDD, indicatingthe
existence of circadian rhythms of locomotor activity. FFT value dropped to
0.007±0.0003 (p<0.001) in the final days preceding death, showing behavioural
arrhythmicity.

Figure 4-7. Locomotor Activity of Representative Wild Type Fruit Flies Showing Arrhythmic Behaviour Before Death
†
†
†
Time (hour)0 8 20 24
50
66
30
56
10
31
60
40
50
20
2
1
4
1
3
Age
(d
ay)
Be
fore
De
ath (d
ay)
A
B
C
1
Age
(d
ay)
Age
(d
ay)
Be
fore
De
ath (d
ay)B
efo
re D
eath
(day)
Time (hour)
Time (hour)
0 8 20 24
0 8 20 24
158
Actogram showing 13-day
of LD 12:12 followed by
free running rhythms in
DD (the white area marks
the light phase, the grey
shaded area marks the
dark phase). Left y-axis
indicates the age of each
fly and right y-axis
indicates the days before
death. Dagger is used as
label of death. Well-
entrained flies of
different ages (A) died at
66 days old, showing a 2-
day arrhythmic activity
with a FFT value of 0.0001
before death (0.049 in LD
and 0.011 in DD), (B) died
at 56 days old, showing a
4-day arrhythmic activity
with a FFT value of 0.008
before death (0.021 in LD
and 0.017 in DD), and (C)
died at 31 days old,
showing a 3-day
arrhythmic activity with a
FFT value of 0.0006
before death (0.046 in LD
and 0.099 in DD).

TheBreakdownoftheCircadianClock
159
4.6Discussion
Here, we reported for the first time that a transient increase of both PER and TIM
expressionat thewholeorganism leveloccurreddaysbeforedeath.This signalhas
been found to be most obvious in TIM, and we assume that it can be used as a
molecularmarkerofimminentdeath.Thelowsignalbackgroundof8.0-lucmakesit
difficulttotellwhethertheaberrantincreaseofPERsignalalsoexists inthecentral
clock.However ithasbeen reported that the centralmolecular clock inDrosophila
remainsefficientuntiltheverylastdayoflife[Luoetal.,2012].Inmammals,ithasalso
been shown that the key function in the central clock is preserved [Weinert and
Weinert,1998;WeinertandWaterhouse,2007].Therefore,theinferenceisthatthe
effect is on peripheral rather than the central level, which implies the further
breakdownofthecouplingsystemandlossofsynchronyamongagedperipheralclocks
beforedeath.
The elevated signals occurred in both clock gene reporters tim-luc and XLG-luc with
differenteffectsonincidence,magnitudeandphase(Figure4-5&4-6).Weinferthat
this signal reflects the actual increase of clock gene expression, rather than the
changes in luciferase reaction which should affect both reporters equally.
Furthermore, we hypothesize that the increase in TIM expression is the leading
change,whichinturnaltersPERexpressionasadownstreameffect.Priortodeath,the
increasingexpressionofPER isstill inphasewithnormal24-hourLDcyclesyetTIM

TheBreakdownoftheCircadianClock
160
expressionlosescircadianpatternanddistributesrandomly.Itimpliesthatthechange
of clock mechanism before death is starting with the continuous increasing
transcription of tim due to the loss of regulation by the transcription-translation
feedback loop. It is very likely that without the stabilization from TIM, PER in the
cytoplasm cannot enter nucleus to inhibit its own transcription (negative feedback
loop)thusresultinginanincreaseinexpression.Thismayalsoexplainwhysomebut
notalloftheflies(45.7%)showincreaseofPERexpression,ifTIMisaroundforPERat
therighttimingthenPERexpressionisnormal,otherwisePERexpressionincreases.
Correspondingly, we hypothesize the aberrant increase of TIM immediately preceding
deathiscentraltotheunderlyingmechanismofthefinalcollapseofthecircadianclock
system.Thelossincircadianbehaviouralrhythmicity(Figure4-7)canbeexplainedby
simultaneous increaseofclockgeneexpression.Suchcircadianclockchanges (both
geneexpressionandoutputbehaviour)inthelastdaysoflifearenotage-dependent
becausebothyoungandagedfliessharesimilarpattern.ThechangesofTIMbefore
deathisrelatedtoitsroleasaclockgeneessentialincircadianrhythmicity,whichis
supportedbythefindingthattimnullmutantsshowfasterlocomotorrhythmdecline
withadvancingageandshortenedlifespan[Kohetal.,2006;Vaccaroetal.,2017].
Althoughtheunderlyingmechanism isnot fully revealed, timmustbe important in the
biologyofaginganddeathbeyonditsrolesincircadianrhythmicity.Timfunctionsin
cellcycleprogression,DNAreplication,DNAdamageresponse,andtelomerelength
and integritymaintenance[Mazzoccolietal.,2016],making itamultifacetedfactor

TheBreakdownoftheCircadianClock
161
implicated in aging and death. Tim has drawnmore andmore attention in recent
studiesforitsroleincancer[Chietal.,2017;Liuetal.,2017;Zhangetal.,2017],also
indicates its versatility. It also raises the possibility that immune response towards
infectionisoneoftheunderlyingreasons,giventhattheroleoftimasaregulatorof
immuneresponsehasbeenrecentlyrevealedinPlanarian[Gutiérrez-Gutiérrezetal.,
2017].Anoveltheoryisalsorisingthatthroughreprogrammingoftranscriptomein
cells, aging turnsoff genes involved inhomeostasis and turnson those involved in
tissue-specificstresses,suchasinflammationandDNAdamage[Solanasetal.,2017].
Furthermore, a recent study has reported that about 1% of the gene transcripts
significantlyincreaseinabundancepost-morteminzebrafishandmice,amongwhich
are genes involved in survival and stress compensation as well as developmental
controlandcancer[PozhitkovandNoble,2017].Althoughthetechniqueusedlimits
thefeasibilitytoalsomeasuretheexpressionofthesegenesbeforedeath,itraisesthe
possibilityofacompensatorymechanismatthelaststageoflife.
Asaconsequence,theoutbreakoftimexpressioninthedaysprecedingdeathmightreflect
astressorimmuneresponse,oracompensatorymechanismofthebodywhenfacing
ultimatesurvivalchallenge.

TheBreakdownoftheCircadianClock
162
4.7Conclusion
Research on the topic of death is of significance to ethical, legal andmedical science
[PozhitkovandNoble,2017],whichmainlyfocusesonpost-mortemsubjects/samples.
Therearenumerousbarrierslimitingthepossibilitytomovethetimeframeforward,
sothatdatafromthemomentsbeforedeathcanbecollected.Byusinganimalmodels
such asDrosophila and techniques such as luciferase reporters give us a precious
opportunitytorevealsomeirreplaceableinformationonthechangespriortodeath.
Herewereportaremarkabledisruptionofthecircadianclockduringthelastdaysoflifeat
boththemolecularandbehavioural levels. Importantly,wereveal forthefirsttime
thatanovelmarkerofimminentdeathinDrosophila.Ourresultsindicatenotonlythe
underlyingcauseofthecircadianclockdisruptionbeforedeath,butalsotheexciting
potentialoftheclockgenestobekeyinthemechanismofdeath,andevenabasisfor
intervention.Thesefindingsallgreatlyexpandourunderstandingonthelinkbetween
thecircadianclockanddeathprocess(Figure4-8).
AlthoughwefindtheaberrantexpressionofTIMbeforedeathatthewholeanimallevel,
itdoesnotnecessarilymeanthatthesamepatternoccursineverytissue/organorcell.
It has been reported that there are significant differences in qualitative and
quantitativeofgeneexpressionbetween tissues/organsafterdeath [Pozhitkovand
Noble,2017;Ferreiraetal.,2018].Forinstance,mRNAinthemousebrainincreasesin

TheBreakdownoftheCircadianClock
163
thefirsthourandthengraduallydecreaseswhileinthelivergraduallyreduceswith
post-mortemtime.Itisreasonabletoassumethatothermulti-cellularorganismswill
displayasimilarpattern.Therefore,geneexpressionamongindividualtissues/organs
shouldbeincludedinfuturestudies.
Itisnotsensibletorelyononespecificmoleculetobethegeneralmarkerofagingordeath,
due to the substantial differencesof aging rate and lifespandependentongenetic
background among individuals [Koh et al., 2006; Giebultowicz and Long, 2015]. As
consequence,furtherexplorationonvariousgenesamongdifferentspeciesisneeded.
Based on TIM being a steppingstone, it is not hard to foresee that more genes,
includingnon-clockgenes,willbedisclosedinfuturestudiesshowingtheirimportance
onregulatingthefinalprocessoflife.Theestablishmentofanarrayofgeneexpression
attheterminalstagewillcertainlyopenthegatetostudythebiologyofdeathand
attractwideinterestacrosslifescience.

DeathBefore After
Overt RhythmsLocomotor Activity
Body Temperature
Loss of Output Rhythmicity Rise of Gene Expression
Genes
Survival
Stress Compensation
Development Control
Cancer
CellOrganBody Cell Organ
Organization Failure
Increase of Clock Gene Expression
TIMELESS
Clock Rhythmicity
Cell Cycle
DNA Repair
Telomere Integrity
Figure 4-8. Overview of New Findings on Changes of the Clock Before Death
Based on our findings from the current towards-death study in Drosophila, aberrant clock gene
expression in TIM throughout the animal may be the cause of the arrhythmic behaviour occurring
days before death. It indicates the involvement of clock mechanism in the death process.
Importantly, we reveal that the breakdown of the clock organization at the body level happens
prior to death, instead of after death.
164

TheBreakdownoftheCircadianClock
165
4.8References
BassoA,DelBelloG,PiacenzaF,GiacconiR,CostarelliL,MalavoltaM.Circadianrhythmsofbody
temperatureandlocomotoractivityinagingBALB/cmice:earlyandlatelifespanpredictors.
Biogerontology.2016Aug;17(4):703-14.DOI:10.1007/s10522-016-9635-y.
ChiL,ZouY,QinL,MaW,HaoY,TangY,LuoR,WuZ.TIMELESScontributestotheprogressionof
breast cancer through activation of MYC. Breast Cancer Res.2017 May 2;19(1):53. DOI:
10.1186/s13058-017-0838-1.
Davis FC,Viswanathan N. StabilityofcircadiantimingwithageinSyrian hamsters. Am J Physiol.
1998Oct;275(4Pt2):R960-8.
FerreiraPG,Muñoz-AguirreM,ReverterF,SáGodinhoCP,SousaA,AmadozA,SodaeiR,Hidalgo
MR,PervouchineD,Carbonell-CaballeroJ,NurtdinovR,BreschiA,AmadorR,OliveiraP,Çubuk
C,CuradoJ,AguetF,OliveiraC,DopazoJ,SammethM,ArdlieKG,GuigóR.Theeffectsofdeath
andpost-mortem cold ischemia on human tissue transcriptomes. Nat Commun. 2018 Feb
13;9(1):490.DOI:10.1038/s41467-017-02772-x.
FroyO. Circadian rhythms, aging, and life span in mammals. Physiology (Bethesda).
2011Aug;26(4):225-35.DOI:10.1152/physiol.00012.2011.
GiebultowiczJM,LongDM.Ageingandcircadianrhythms.CurrOpinInsectSci.2015Feb1;7:82-86.
DOI:10.1016/j.cois.2015.03.001.
GrotewielMS,Martin I,Bhandari P,Cook-Wiens E. Functional senescence in Drosophila
melanogaster.AgeingResRev.2005Aug;4(3):372-97.DOI:10.1016/j.arr.2005.04.001.
Gutiérrez-GutiérrezÓ,FelixDA,González-EstévezC.Planarian finds time(less) to fight infection.
Virulence.2017Mar2:1-6.DOI:10.1080/21505594.2017.1300735.
Hendricks JC,Lu S,Kume K,Yin JC,Yang Z,Sehgal A. Genderdimorphismin the role of cycle
(BAML1) inrest,restregulation, and longevity in Drosophilamelanogaster. J Biol Rhythms.
2003Feb;18(1):12-25.DOI:10.1177/0748730402239673.

TheBreakdownoftheCircadianClock
166
HoodS,Amir S. Theagingclock:circadian rhythmsandlaterlife. J Clin Invest.2017Feb
1;127(2):437-446.DOI:10.1172/JCI90328.
HurdMW,RalphMR.Thesignificanceofcircadianorganizationforlongevityinthegoldenhamster.
JBiolRhythms.1998Oct;13(5):430-6.DOI:10.1177/074873098129000255.
KeilG,CummingsE,deMagalhães JP.Beingcool:howbody temperature influencesageingand
longevity.Biogerontology.2015Aug;16(4):383-97.DOI:10.1007/s10522-015-9571-2.
Klarsfeld A,Rouyer F. EffectsofcircadianmutationsandLDperiodicityon the life span of
Drosophila melanogaster. J Biol Rhythms.1998 Dec;13(6):471-8. DOI:
10.1177/074873098129000309.
Koh K, Evans JM, Hendricks JC, Sehgal A. A Drosophila model for age-associated changes in
sleep:wake cycles. Proc Natl Acad Sci U S A. 2006 Sep 12;103(37):13843-7. DOI:
10.1073/pnas.0605903103.
KondratovRV,KondratovaAA,GorbachevaVY,VykhovanetsOV,AntochMP.Earlyagingandage-
relatedpathologies inmicedeficient inBMAL1, thecorecomponentof thecircadianclock.
GenesDev.2006Jul15;20(14):1868-73.DOI:10.1101/gad.1432206.
KrishnanN,kretzschmarD,RakshitK,ChowE,GiebultowiczJM.Thecircadianclockgeneperiod
extendshealthspaninagingDrosophilamelanogaster.Aging.2009Nov19;1(11):937-48.DOI:
10.18632/aging.100103.
Kumar S,Mohan A,Sharma VK. CircadiandysfunctionreduceslifespaninDrosophila
melanogaster.ChronobiolInt.2005;22(4):641-53.DOI:10.1080/07420520500179423.
LeeCC.Thecircadianclockandtumorsuppressionbymammalianperiodgenes.MethodsEnzymol.
2005;393:852-61.DOI:10.1016/S0076-6879(05)93045-0.
LiuSL,LinHX,LinCY,SunXQ,YeLP,QiuF,WenW,HuaX,WuXQ,LiJ,SongLB,GuoL.TIMELESS
confers cisplatin resistance in nasopharyngeal carcinoma by activating the Wnt/β-catenin
signalingpathwayandpromotingtheepithelialmesenchymaltransition.CancerLett.2017Jun
3;402:117-130.DOI:10.1016/j.canlet.2017.05.022.

TheBreakdownoftheCircadianClock
167
LuoW,ChenW-F,YueZ,ChenD, SowcikM,SehgalA, ZhengX.Old flieshavea robust central
oscillatorbutweakerbehavioralrhythmsthatcanbeimprovedbygeneticandenvironmental
manipulations.AgingCell.2012Jun;11(3):428-38.DOI:10.1111/j.1474-9726.2012.00800.x.
MartinoTA,Oudit GY,Herzenberg AM,Tata N,Koletar MM,Kabir GM,Belsham DD, Backx PH,
RalphMR,SoleMJ.Circadianrhythmdisorganizationproducesprofoundcardiovascularand
renaldiseaseinhamsters.AmJPhysiolRegulIntegrCompPhysiol.2008May;294(5):R1675-
83.DOI:10.1152/ajpregu.00829.2007.
MazzoccoliG,LaukkanenMO,VinciguerraM,ColangeloT,ColantuoniV.Atimelesslinkbetween
circadian patterns and disease. Trends Mol Med. 2016 Jan;22(1):68-81. DOI:
10.1016/j.molmed.2015.11.007.
MorinLP.Age-relatedchanges inhamstercircadianperiod,entrainmentand rhythm-splitting. J
BiolRhythms.1988Sep;3(3):237-48.
OuyangY,AnderssonCR,KondoT,Golden SS,JohnsonCH.Resonating circadian clocks enhance
fitnessincyanobacteria.ProcNatlAcadSciUSA.1998Jul21;95(15):8660-4.
ParkN,Cheon S,SonGH,Cho S,Kim K. Chronic circadian disturbance by a shortened light-dark
cycle increases mortality. Neurobiol Aging.2012Jun;33(6):1122.e11-22. DOI:
10.1016/j.neurobiolaging.2011.11.005.
PearlP.Thebiologyofdeath.Philadelphia:JBLippincottCompany.1922.
PittendrighCS,Minis DH. Circadian systems: longevity as a function of circadian resonance in
Drosophilamelanogaster.ProcNatlAcadSciUSA.1972Jun;69(6):1537-9.
Pozhitkov AE,Noble PA. Gene expressionin thetwilightofdeath: The increase of thousands of
transcriptshasimplicationstotransplantation,cancer,andforensicresearch.Bioessays.2017
Sep;39(9).DOI:10.1002/bies.201700066.
SolanasG,PeixotoFO,PerdigueroE,JardíM,Ruiz-BonillaV,DattaD,SymeonidiA,CastellanosA,
Welz PS,Caballero JM,Sassone-Corsi P,Muñoz-Cánoves P,Benitah SA. Aged stem cells
reprogram their daily rhythmic functions to adapt to stress. Cell. Elsevier Inc; 2017 Aug
10;170(4):678-678.e20.DOI:10.1016/j.cell.2017.07.035.

TheBreakdownoftheCircadianClock
168
Vaccaro A,Issa AR,Seugnet L,Birman S,Klarsfeld A. Drosophila clock is required in brain
pacemakerneuronstopreventprematurelocomotoraging independentlyof its circadian
function.PLoSGenet.2017Jan10;13(1):e1006507.DOI:10.1371/journal.pgen.1006507.
WeinertD,WaterhouseJ.Thecircadianrhythmofcoretemperature:effectsofphysicalactivity
andaging.PhysiolBehav.2007Feb28;90(2-3):246-56.DOI:10.1016/j.physbeh.2006.09.003.
WeinertH,WeinertD.Circadianactivityrhythmsoflaboratorymiceduringthelastweeksoftheir
life.BiolRhythmRes.1998;29(2):159-78.
ZhangW,HeW,ShiY,ZhaoJ,LiuS,ZhangF,YangJ,XieC,ZhangY.AberrantTIMELESSexpression
is associated with poor clinical survival and lymph nodemetastasis in early-stage cervical
carcinoma.IntJOncol.2017Jan;50(1):173-184.DOI:10.3892/ijo.2016.3784.

No snowflake in an avalanche ever
feels responsible.
Stanislaw Jerzy Lec, from More Unkempt Thoughts, 1969

CHAPTER 5
Discussion
169

The Life History of the Circadian Clock

TheLifeHistoryoftheCircadianClock
171
Thecontentsofthischapterhavebeenpublished*(pleaseseeAppendixfordetails).
*ZhaoJ,WarmanGR,CheesemanJF.Thefunctionalchangesofthecircadiansystem
organizationinaging.AgeingResRev.2019Apr29.pii:S1568-1637(18)30304-0.DOI:
10.1016/j.arr.2019.04.006.

TheLifeHistoryoftheCircadianClock
172
5.1Summary
NooneonEarthcanescapethe impactof time.Aftermillenniaofevolution,biological
clockswithineveryorganismhavedevelopedtopreciselymeasurethepassingoftime.
Among these, the circadianclock ismost importantandwell-studied, yet thisdaily
timingsystemembeddedineachbodyisnotalwayshard-and-fastasitshouldbe.The
circadianclock is shapedby time, showing remarkablechanges from itsbirth to its
death.
Weassumethattheclockisnotperfectatthebeginning.Throughgradualdevelopment,
iteventuallymaturesintoafully-functionalclockkeepingbiologicalrhythmsinternally
coupledandexternallycompatible.Thenitcomestotheinevitablefadingunderaging.
Ultimately,theclockstopstickingatthecriticaltimingofdeath.
Along with the continuous alterations as the animal develops, matures and ages, the
unequal changes amongevery component (spatially) of the circadian clock at each
stage (temporally) add particulars to this already colourful picture. Based on our
findings,werevealsomebreakthroughsonthetopicofdevelopmentandagingofthe
circadianclockinDrosophilamelanogaster,answeringquestionswehavebeenasking
(refertoChapterOne,Section1.6).Thesecanbesummarizedasfollows:

TheLifeHistoryoftheCircadianClock
173
Q: When does the molecular clock start to tick and be
responsivetolight?
A: The expression of clock gene in certain central clock
neuronsbeginstooscillateatearlydevelopmentalstagewith
light-sensitivity, when that in the peripheral tissues
throughoutthebodybeginstoincreasebutnotcyclewithout
light-sensitivity.
Q:Dothecentralandperipheralclocksdevelopatthesame
pace?
A: The molecular machinery in the central clock starts to
function at embryonic stage, earlier than that in the
peripheralclocks,whichstartstotickatearlyadultstage.
Q:Howdoes theZeitgeber lightaffect thedevelopmentof
thecircadianclock?
A:Clockgeneoscillationsaswellasadultlocomotoractivity
rhythms start without the requirement of light. Light
exposuredoesnot initiateoracceleratethecircadianclock
development, instead it synchronizes the phases among
differentindividuals.
ChapterTwo
DEVELOPMENT
ChapterTwo
DEVELOPMENT
ChapterTwo
DEVELOPMENT

TheLifeHistoryoftheCircadianClock
174
Q:Whichpartofthecircadianclockisaffectedbyaging,the
centreortheperiphery?
A:Agingaffectstheendogenousclockattheperipherallevel,
showing reduction in both expression and rhythmicity of
clockgenesthroughoutthebody,whileleavesthemolecular
clockatthecentrallevelintact.
Q:Canstrongentrainmentrescuetheage-relateddeclineof
thecircadianclock?
A:Regularandstrongentrainmentincludinglight-darkcycles
and light-dark coupled with temperature cycles are not
efficient enough to recover the age-related decline of the
peripheralmolecularclockwork.
Q: What happens to the clock at both molecular and
behaviourallevelstowardsdeath?
A:Clockgeneexpression in theperipheralclocksshowsan
aberrant pattern in both expression and rhythmicity days
before death, which is mirrored by a disturbance in
behaviouralrhythms.
ChapterThree
AGING
ChapterThree
AGING
ChapterFour
DEATH

TheLifeHistoryoftheCircadianClock
175
Inthis finalchapterwediscussthedynamicnetworkofthecircadianclockon itscross-
sectionalalterationsateachstageanditslongitudinalchangesthroughoutlifespan.In
the firstpart,we summarize thenetwork systemof the circadian clock, and in the
second part, we focus on the development- and age-related changes of different
componentswithinthisinteractiveorganization.

TheLifeHistoryoftheCircadianClock
176
5.2FromtheCentretotheEdge
TheRULEof‘CircadianResonance’wasproposedbythefounderofChronobiologyColin
Pittendrigh [Pittendrigh et al., 1958], emphasizing the importance of the harmony
betweentheinternalclockandexternalenvironmentanditsimplicationinthefitness
oforganisms.Itremindsustheexquisitely-formedarchitectureofthecircadianclock,
withinwhichallpartiesarecoordinatedwelltogethertoachievethebestperformance,
andinteractpreciselywiththeexternalworldasasystem.Thecircadianclockcontains
suchapowerfulnetworkthateverycomponentisconnected,followingtheregulation
oftheRULE.
Thecouplingmechanismtomaintainthenetworkwithinthecentralclockhasbeenwell
studied. In Drosophila, a sophisticated network is formed by the intercellular
communication among different clusters of clock neurons in the central nervous
system.Centralclockneuronshavebeenreportedtobeweakoscillatorsifwithoutan
intactcircadiancircuit,leavingthegenerationofrobustcircadianrhythmsimpossible
[Sabadoetal.,2017].Amongall,pigmentdispersingfactor(PDF)playsakeyroleinthe
communication within the central clock neuronal network to maintain rhythmic
activity[TopandYoung,2017].This issupportedbythefindingsthatclockneurons
fromflieslackingPDFareuncoupledandshowingimpairedrhythmicity[Pengetal.,
2003;Linetal.,2004;Yoshiietal.2009].Furthermore,PDFhasmultipleeffectson
differentgroupsoftheclockneurons.Althoughonlyreleasedbysmallventrallateral

TheLifeHistoryoftheCircadianClock
177
neurons(s-LNv)andlargeventrallateralneurons(l-LNv),PDFalsofeedsbacktonon-
PDF-expressing neurons. In a word, PDF is crucial for adjusting cycling amplitude,
period, and phase of different players in the circadian system [Yoshii et al., 2009;
Collinsetal.,2014].
Inmammals,thesuprachiasmaticnucleus(SCN)isknownasahierarchicalmulti-oscillator
systeminwhichindividualneuronsareinterconnectedintotheneuronalnetworkto
sendacoherenttimingsignaltotherestofthebody.Thecircadianspontaneousfiring
rhythmsaresynchronizedthroughouttheSCN[Enokietal.,2017],thoughdispersed
individualneuronsdisplayheterogeneityinperiods,phasesandstrengthofoscillation
[Welshetal.,1995;Liuetal.,1997;Honmaetal.,1998;Honmaetal.,2004;Webbet
al., 2009]. SCN neurons possess specific mechanisms to maintain synchrony as a
functional unit, and the neuropeptidergic coupling is believed to be the most
important one [Liu and Chang, 2017]. Vasoactive intestinal polypeptide (VIP) is
necessary both tomaintain circadian rhythmicity within individual neurons and to
synchronizemoleculartimekeepingamongthem[Atonetal.,2005;Maywoodetal.,
2006]. For instance, inhibitionofVIP-mediatedcoupling in theSCN leads to lossof
synchronyinPERIOD2(PER2)expressionofindividualneurons[Abrahametal.,2010].
Peripheral clocks, on the contrary, are more complicatedly formed as a system when
comparedtothecentralclockduetotheirdiversestructuresaswellasfunctions,yet
thenetworkorganization is far inferior. In fact, ithasbeenreportedthat individual
fibroblastsarerobustmolecularoscillators[Nagoshietal.,2004;Welshetal.,2004].

TheLifeHistoryoftheCircadianClock
178
Thisarguesthatatthecellularlevel,theperipheralcellsarecomparableoscillatorsto
thecentralcells,ifnotbetter[Mohawketal.,2012].Yetunderconstantconditionsin
vitro,Period1rhythmsofperipheraltissueexplantsdampenpartiallyorcompletely,
while in the SCN persist for amuch longer time [Yamazaki et al., 2000]. Likewise,
peripheral tissues in Drosophila can be clearly distinguished from the molecular
oscillatorsinthecentre,fortheirrapiddampeninginconstantconditions[Pengetal.,
2003]. Apparently at the tissue level, peripheral clocks behave like weak, small-
amplitudeoscillatorsandthecentralpacemakerbehaveslikearigid,high-amplitude
circadianoscillator[Abrahametal.,2010].Thelossofsynchronyamongindividualcells
andthusdampingoftheensemblerhythmatthepopulationlevelinperipheryisthe
mostimportantreasonaccountingforthedifference[Welshetal.,2004].Itismainly
duetothefactthatperipheralclockslackofthefunctionalcouplingmechanismthatis
unique to the central clock [Brown and Azzi, 2013]. Despite that individual
tissues/organsmaymaintainsomedegreeofcircadiansynchrony,itiswellaccepted
thatthiscouplingismuchweakerthanthatintheSCN[Yooetal.,2004].
Therefore, spontaneous synchronization among central clock neurons is a primary
requirementofitsmasterpacemakerroleanddistinguishesitfromperipheraltissues.
What’smore,thecircadiannetworkintheDrosophilabraincanserveasanessential
bufferingsystemthatprotectsagainstchallengessuchastranscriptionalimpairment
ofclockgenes,whiledisturbanceofmolecularclockworkaswellasoutputrhythmsare
shownintheperiphery[Weissetal.,2014].Similarly,theintercellularnetworkenables

TheLifeHistoryoftheCircadianClock
179
the SCN to fight against genetic perturbations which otherwise disturb circadian
rhythmsinperipheraltissuesandcells[Liuetal.,2007].
The connectivenetworkbetween the central pacemaker and local oscillators hasbeen
characterizedinDrosophilaasinmammals,butthepatternisthoughttobedistinct.
InDrosophila,coordinatedtimingbetweenindividualoscillatorsisthoughttooccurvia
lightandtemperaturesensitiveintracellularpathways,whichresponddirectlytothe
sameentrainmentcuesthatsetthephaseofthecentralpacemakerneurons inthe
brain [Allada and Chung, 2010]. The cell-autonomous expression and action of the
blue-lightphotoreceptorCRYPTOCHROME(CRY)isimplicatedinthelight-entrainment
inbothcentralandperipheralclocks[Krishnanetal.,2001;DubruilleandEmery,2008;
Agrawaletal.,2017].Inthisway,peripheralclocksmaintainsynchronywithexternal
environmental cues independent of input from the central clock with several
exceptions[Plautzetal.,1997;Kruppetal.,2013](refertoChapterOne,Section1.2).
Inmammals, ononehand, the SCN shows thedominant role and regulates peripheral
clocksthroughdirectandindirectpathways.Directpathways includeneuralcontrol
throughtheautonomicnervoussystem,aswellashumoralcontroltravellingthrough
circulationsuchasglucocorticoids[Mohawketal.,2012](refertoChapterOne,Section
1.4).Indirectpathwaysincludefeedingrhythmsandbodytemperaturerhythms,which
are regulated directly by the SCN and/or by activity cycles controlled by the SCN
[Dibneretal.,2010].Ontheotherhand,theinfluencefromoneparticularperipheral
clocktotheSCNortoothertissuesissupposedtobesmall.Ithasbeenreportedthat

TheLifeHistoryoftheCircadianClock
180
specificgeneticdisruptionofthecircadianclockmechanisminthehepatocytesofmice
leavesanintactSCNandothercelltypesthroughoutthebody[Kornmannetal.,2007].
Despitethatperipheraltissuesarerelativelyindependentwitheachotherasthisstudy
suggests,theweakeningofrhythmicityand/orshiftofphaseswouldpresumablecause
desynchronyandconsequentimpactatthesystemiclevel.
Insummary,ledbyatightly-coupledcentralpacemaker,theperipheralcircadiansystemis
composed of loosely-coupled clocks which need to be coordinated by internal or
externalcues.Correspondingly,theendogenouscircadianoscillatorsinindividualcells,
tissues and organs within the same organismmust interplay wisely to maintain a
consolidatednetwork.Innormalcircumstance,circadianoscillatorsinindividualcells
aresynchronizedwithinatissue,likewiseindividualtissuesarekeptinastablephase-
relationship with each other in order to make up a coherent circadian system.
Accordingly, loss of synchrony could happen at various levels of the circadian
organization,orcombinationofseveral,suchasinthestateofagingorcertaindiseases.

TheLifeHistoryoftheCircadianClock
181
5.3FromtheBeginningtotheEnd
A considerable amount of research has helped shed light on the circadian clock being
crucialformaintainingorganizedandhealthylifehistory.Actually,thecircadianclock
hasbeenimplicatedinaffecting lifetraitssuchaspre-adultdevelopmentaswellas
adultlifespan.Itisgenerallybelievedthatafasterclock(shorterfree-runningperiod)
speeds up pre-adult development and shortens lifespan, vice versa, a slower clock
(longerfree-runningperiod)slowsdowndevelopmentandlengthenslifespan.
Thedevelopmentalplanofananimalbodyisbuiltthroughthecoordinatedtimingofcell
divisions, cell differentiation, and cell movements [Vallone et al., 2007], and the
circadian clock is one of the important determinants that orchestrate these basic
processes.Ithasbeenreportedthatthepaceofpre-adultdevelopmentinDrosophila
is controlledby clockgenes [Kumaret al., 2006]. For instance,period (per)mutant
studiessuggestapositivecorrelationbetweenfree-runningperiodanddevelopment
timeinDrosophila.TheperShortmutants(19-hour)developfasterfromeggstoadult
than the wild type, while perLong mutants (28-hour) complete development more
slowlythanthewildtype[Kyriacouetal.,1990].Furthermore,thecircadianclockhas
beensuggestedinregulatingthetimingofeclosioninDrosophila[KonopkaandBenzer,
1971].Thiskeydevelopmentaleventisrestrictedtooccurarounddawn,evenifflies
arereadytoecloseearlier.ActingbymeansofPDF,interactionsbetweenaperipheral
clock intheprothoracicglandandthecentralclockof the lateralneurons,seemto

TheLifeHistoryoftheCircadianClock
182
answer for themechanismof the circadian timingof eclosion [Myers et al., 2003].
Likewise, circadian timing is also a featureofmammalian embryonic development.
Loss of function in clock genes disrupts developmental processes, such as growth,
proliferation and differentiation [Dolatshad et al., 2010]. For instance, knockout of
certain clock genes in mice shows embryo losses and/or delayed embryonic
development[Dolatshadetal.,2010].
Inthisstudyweshowthatthedevelopmentofthecircadianclock itselfgoesthrougha
hierarchicalprocess,withcertaincentralclockneuronsdevelopfirst,andperipheral
tissues at the body levelmature later (refer to Chapter Two DEVELOPMENT). The
meaning is presumably simple, it is that the central clock plays a relatively more
importantroleinregulatingtheearlydevelopmentoftheclocksystemitself,somuch
sothattocoordinatethedevelopmentoftheentirebody.Themechanismunderlying
is stillunknown,but thedevelopmentof tight-or loose- couplingmayexplainwhy
expression of clock gene shows circadian rhythmicitywithin central clock neurons,
whileonly increasesbutnotoscillates in synchronyamongperipheral clocksat the
samedevelopmentalstages.Wethusinferthatnetworkassemblyatthesystemiclevel
is on-going but incomplete during early development. In addition, the non-cyclic
expression of clock gene in peripheral tissues under constant darkness implies the
scarceinputfromthecentralpacemaker.Withoutinternalcues,externalstimulisuch
aslight-dark(LD)cyclesbeingincapabletoentrainclockgeneoscillationsindicatesthat
theperipheralpropertyoflight-sensitivityhasnotyetbeenestablishedbeforeeclosion.

TheLifeHistoryoftheCircadianClock
183
If pre-adult development is described as travelling towards the destination where the
beautifulscene isthewell-functioningadultstage,thataging is the journeydrifting
awayfromperfection.Agingischaracterizedasagradualdeclineinnumeroussystems,
influencedbybothgeneticandenvironmentalfactors,withchangeshappeningatboth
thephysiologicalandmolecularlevels[Banksetal.,2016].Thereareninehallmarksof
aging, which are regulated by the clock to different extent, including increased
genomic instability, diminishing of the telomeres, epigenetic alterations, loss of
proteostasis, impairment of stem cells,mitochondrial dysfunction, dysregulation of
metabolicactivity,cellularsenescenceandintercellularmiscommunication[Fonseca
CostaandRipperger,2015].
It is still not clearwhetherdeclineof clock function is the causeor the result of aging
process,orboth.Existingevidencesuggestsabidirectionalrelationshipbetweenthe
circadianclockandaging.Whilethereisevidenceonage-relateddampeningofclock
oscillationsandclock-controlledrhythms,disruptionofthecircadianclockinturnleads
to accelerated aging and increased susceptibility to age-associated pathologies
[RakshitandGiebultowicz,2013;GiebultowiczandLong,2015;HoodandAmir,2017].
When aging and the aged clock interact adversely with each other, it leads to an
irreversibleoutcomeofdeterioration.Forinstance,ithasbeenreportedthatevena
mild circadian challenge suchas chronic jet-lag, canmarkedly increasemortality in
aged but not young animals [Davidson et al., 2006]. This illustrates a “double-hit”
theorythatafurtherdisturbanceoftheagedclock(firsthit)byasecondattacksuch
aschronicjet-lagcancausecascadedconsequences.

TheLifeHistoryoftheCircadianClock
184
Unsurprisingly,theeffectofagingonthecircadianclockmustbemorethanjustasimple
story,especiallywithbroaderanddeeperexploration.Infact,agingmayactonevery
componentoftheclockorganization,includingthecentreandtheperiphery,atthe
levelsoftissue,intercellularconnection,andthecell.Althoughpresumablynoparty
canescapetheinfluenceofaging,theweightofcontributioncanbediverse.
From the existing results of bothmammalian andDrosophila studies including ours, a
generalimpressionisthatthemolecularmachineryinthecentralclockislessorlater
damaged,ifthereisany,bytheagingprocess.Inthecurrentstudy(refertoChapter
ThreeAGING),wefindthattheexpressionofclockgeneinthecentralclockneurons
ofagedDrosophilastaysasrobustasinyoungones.Ithasalsobeenproposedthat
molecularclockintheSCNasawholeisrelativelyresistanttoaging[MattisandSehgal,
2016].As consequence, causal linksbetweenclockgeneexpressionpatterns in the
central clock and age-associated disturbance in circadian rhythms, cannot been
conclusivelydemonstrated.
Nonetheless, itdoesnotmean that thecentral clock is “off thehook”when facing the
challengeof aging. The first clue that the central clock is involved in agingprocess
comes from the transplantation studies. Implantation of foetal SCN tissue in old
animals rescue circadian output function in hamsters [Van Reeth et al., 1994;
Viswanathan and Davis, 1995] and rats [Cai et al., 1997]. Moreover, animals that
restorehigherclockfunctionendupwithincreasedlongevity[HurdandRalph,1998].
Thereareafewpossibilitiesasidefromtheintactmolecularclockwork,includingthe

TheLifeHistoryoftheCircadianClock
185
loss of structural integrity, theweakening of the output signal, the interruption of
connectionsbetweentheneurons,andthediminishingofsinglecellrhythmswithin
theSCN[Farajniaetal.,2014].
Whether age-related clock decline is paralleled by age-associated alterations in the
structureoftheSCNisunclear.TheresultsofthechangesoftheSCNinbothvolume
andtotalcellnumberswithincreasingageinhumansandrodentsaremixed[Hood
andAmir,2017].Bycontrast,theevidenceofSCNneuronaloutputdeteriorationwith
advancingageatthetissuelevelissolidandconsistent.Theelectricalactivitypatterns
of SCN neuronsmeasured bymultiunit activity (MUA) recordings indicate that the
amplitudeofSCNactivitydecreasesunderaginginbrainslicepreparations[Satinoffet
al.,1993;Watanabeetal.,1995;Nygårdetal.,2004;Biello,2009]andculturedSCN
neurons [Aujard et al., 2001] in vitro, as well as in vivo [Nakamura et al., 2011].
Importantly,thereductionofSCNoutputmeasuredbyneuronalactivityrhythmsat
thetissuelevelisdirectlyaccountforovertoutputdeclineattheanimallevel.
The coupling system in the SCN which undergoes significant age-related degradation
[MattisandSehgal,2016]mayanswerforsuchdeteriorationoftheSCNdownstream
neuronal output [Nakamura et al., 2016]. Themain neurotransmitters such asVIP,
whichmakesindispensablecontributiontothesynchronizationwithintheSCNcellular
network [Vasalou et al., 2009], is affected by aging. It has been reported that the
expressionlevelandtheamplitudeofVIPmRNA[Kawakamietal.,1997;Kallóetal.,
2004]and theexpression levelof its receptormRNA in theSCN [Kallóetal.,2004]

TheLifeHistoryoftheCircadianClock
186
decreaseinthecourseofmammalianaging.Suchattenuationisassociatedwithboth
declineinthelevelofmRNApercellandinthenumberofVIP-producingcellsinthe
SCN [Krajnak et al., 1998]. This is well supported by human research, showing a
decreasednumberofVIPneuronsintheSCNofmiddle-agedmales[Zhouetal.,1995;
ZhouandSwaab,1999].ThedisruptionofVIPexpressionhasfunctionalimplicationin
circadian clock aging. It has been reported that the extent of changes of circadian
amplitude of behaviour with advancing age is correlated with the number of VIP-
expressingSCNneuronsinhumans[Wangetal.,2015].Correspondingly,thepresence
ofVIP-expressingtransplantedcellssuccessfullyrestorescircadianoutputbehaviour
andphysiologyinagedmice[LiandSatinoff,1998].
Existing findings suggest a disrupted cellular coupling of the SCN, yet the circadian
rhythmicitywithinindividualcellsseemstoplaylessimportantrole.Accordingly,the
molecularclockworkisbelievedtobeintactatthecellularlevel[Nakamuraetal.,2016].
Arecentstudyusinganultra-sensitivecamerasystemiscapabletorecordsinglecell
imaging in PER2::LUC reporter mice, revealing clear circadian rhythms and similar
amplitudeineachcellsinbothyoungandagedSCN.Furthermore,eachrhythmrapidly
becomesdesynchronizedanddissociated,graduallydriftsoutofphasewitheachother
inassociatedagedSCNcells[Nakamuraetal.,2015],againindicatingafaultednetwork.
ResultsoftheagingstudiesinDrosophilaaresomehowsimilartothoseinmammals,and
thereisaconvincingpatternofdeclineinsynchronizationamongcentralclockneurons.
ThereductionofthePDFlevelsinthecentralclocknetworkisconsistent[Luoetal.,

TheLifeHistoryoftheCircadianClock
187
2012;Umezakietal.,2012].Moreover,overexpressionofPDFinPDF-positiveneurons
increasesclockgeneexpressioninspecificcentralclockneurons,andpartiallyrescues
rhythmicityandperiodchangesofoutputbehaviourinoldflies[Umezakietal.,2012].
These findings indicatethe importantroleofPDF in theagingprocess,aswellas it
implication in age-associated attenuation of intercellular communication in the
circadianneuronalnetwork.Thetheoryis,withadvancingage,itbecomesincreasingly
difficultfortherhythmsofindividualneuronstoremainsynchronizedwitheachother
andthustherhythmicoutputofthenetworkbecomesfragmented[Banksetal.,2016].
Theexistingevidencestronglypointsatadamagednetworkofthecircadiansystemwith
advancing age. Here we predict a hypothesis that the network may serve as a
protectivemechanismratherthanapurevictimundertheattackofaging.Itcannotbe
deniedthatthenetworkistargetedbytheagingprocess,showingfunctionaldecline,
nonethelessitisstillacrucialplayerintheanti-agingfight.Age-induceddampeningof
circadianrhythmsofneuronalactivityisgreateronthecellularlevelcomparedtothe
wholeSCNinvitro[Farajniaetal.,2012]orinvivo[Nakamuraetal.,2011],suggesting
acompensatoryroleoftheSCNnetworkwhichiscapabletocoverforcellulardefects
underagingtoacertainextent[Farajniaetal.,2014].Anotherstrongpieceofevidence
lies inthefindingofnormalmolecularoscillation intheagedSCN invivobut faster
decline in vitro [Tahara et al., 2017], indicating the maintenance of the systemic
network is a compensating mechanism towards aging. In conclusion, the delicate
networksystemholdstheentireclockorganizationtightlytogetherwhenchallenged
byaging,evenundergoinginevitabledeteriorationitself.

TheLifeHistoryoftheCircadianClock
188
Asidefromtheimpairmentofthecentralclock,anotherpossiblemechanismunderlying
circadiandisruptionwithincreasingageisthedegradationoftheperipheralclocks.It
isgenerallybelievedthatperipheralclocksgothroughmoreseriousdamagethanthe
centralpacemaker,asweshowinthecurrentDrosophilastudy(refertoChapterThree
AGING).Despitethereasonshavingnotbeenfullyrevealed,oneofthepossibilityis
thatperipheraltissueslackofthetight-couplingnetworksystemthatisuniquetothe
centralclock,whichmayworkasaprotectivemechanismaswementionedabove.It
isnothardtoimaginethatthebreakdownofthenetworkshouldnotonlyexistwithin
thecentral,but,inevitablyandmoredramaticallyoccurintheperipheralclocks,since
they are far more complicatedly structured yet far less efficiently coordinated. As
consequence,suchloosenetworkstructureputperipheralclocksinamorevulnerable
status,makingthemeasilytargetedbypathologicalprocessessuchasaging.
Ithasbeenreportedthatrhythmsofneuronaloutputactivityareclearlydegradedinone
of the peripheral tissues (the sub-paraventricular zone) in the mammalian brain
[Nakamuraetal.,2011].Nonetheless,arecentstudyhasshownthatneuronalfiringis
notaffectedbyagingateitherthesinglecell levelorastheoverallpattern inmice
motor cortex [McKillop et al., 2018]. So far there are a relatively small number of
studiesfocusingonthemolecularchangesinagedperipheralclocks.Inaddition,itis
evenmoredifficulttoseeauniformpatternduetothediversitythatagingimpactson
all oscillatory tissues [Yamazaki et al., 2002]. Based on existing data, changes of
rhythmic clock gene activity have been observed in some but not all peripheral

TheLifeHistoryoftheCircadianClock
189
oscillators in a strong tissue-dependent manner in rodents and humans (refer to
ChapterThree,Section3.2).
Itisknownthatorgansageatdifferentrateswithtissue-specificsignatures[Girardotetal.,
2006], adding the complexity of age-related propertieswithin individual oscillatory
tissues. Further, the changes in peripheral oscillator behaviourmaynotonly result
fromintrinsicalterationswithinthetissuesthemselves,butalsobeaconsequenceof
thereductionoftheirresponsivenesstointernalorexternaltemporalcues.Ourresult
showstheinefficiencyofstronglightandtemperatureentrainmentsonrescuingthe
age-relateddeteriorationoftheperipheraloscillators(refertoChapterThreeAGING).
It indicatesthat the impairmentof theperipheral inputpathwaymaybeapossible
mechanism involved in the aging process. It is supported by the finding that the
photoreceptorCRYreducesatbothmRNAandprotein levels inperipheralclocksof
aged Drosophila. Further, restoration of an effective input signal through
overexpressingCRYintheperipheralclockscanreversedeclineofcircadianrhythms
andbenefithealth[RakshitandGiebultowicz,2013].
Finally,wefindthattheclockstopstickingproperlynotatthetimeofdeath,rather, its
functionalcollapsehappensaheadofdeathasaredflagsign(refertoChapterFour
DEATH).Themolecularclockworkattheperipherythroughoutthebodyincreasesthe
clockgeneexpressioninthelastfewdaysbeforedeath,oppositetoitsdecliningtrend
withadvancingage.Itcouldreflectastressresponseoracompensatorymechanism.
Additionwiththetotallossofbehaviouralrhythms,theytogetherillustrateadeeply

TheLifeHistoryoftheCircadianClock
190
dysfunctionalclockatthefinalstageoflife.Insteadofbeingasacascadeeventunder
theprocessof death, the intense reactionof the circadian clockoccurs before the
systemic breakdown of the entire body. Beyond the role as a marker predicting
imminentdeath,theclockisverylikelytocontributetotheeventscausalfordeath.Of
courseitstillleavesanintriguingquestionthat,asoneoftheonce-in-a-lifetimeevents
suchaseclosioninearlyDrosophiladevelopment,whetherthetimingofdeathisalso
underclock-regulated?
Insummary,thedeteriorationofvariouslevelofcircadianclocksystematvariouslayers,
occurring at different rates and at different times, all contributes to the aging and
deathprocess.Withoutdoubt,theentireclockorganizationisaffected,showingage-
related alternations at the cellular, intercellular, tissue levels at both central and
peripheral layers, to different extent. The combination of these impairments is
reflected by the layout at the body level, resulting in a disrupted behavioural or
physiologicaloutputrhythmsseeninelderlypeopleoragedanimals.

TheLifeHistoryoftheCircadianClock
191
5.4TheDynamicNetwork
Thetwodimensionsofthecircadianclock,spatiallyfromthecentretotheperiphery(each
comprisingtheinput,themolecularoscillatorandtheoutput),aswellastemporally
fromthebirthtothedeath,havebeencharacterizedinthisstudy.Weemphasizethe
dynamicchangesofthecircadiannetworkaswellaseachinterconnectedcomponent,
asshowninthedynamicnetworkmodel(Figure5-1).

Figure 5-1. The Dynamic Network of the Circadian System
MOLECULAR CLOCKWORK
EARLY DEVELOPMENT
ADULTAGING
BEFORE DEATH
NETWORK
192
The circadian network is built on the dynamic interaction between the central and peripheral
components, each is composed of the input entrainment, the molecular clockwork and clock
genes, and output rhythms. Importantly, the network of circadian organization goes through
dynamic changes from birth to death, along with the progress of development, maturity, aging,
and the final stage before the end of life.
Network Formation
Network Perfection
Network Degradation
Network Breakdown

TheLifeHistoryoftheCircadianClock
193
At the longitudinal view throughout lifespan in Drosophila (Figure 5-2A), the central
molecularclockdevelopsearlyatembryonicstageandcontinuestostayrobustwhen
theanimalages.Bycontrast,theperipheralmolecularclockmatureslaterattheadult
stageandshowsdeclinewithadvancingage,endingupwithanoutbreakinthelast
daysoflife.Entrainmentofthecentralclockestablishesduringembryogenesis,which
is earlier than that of the peripheral clock which occurs in the adult. Rhythms of
locomotoractivityemergefromadultonwards,deterioratewithincreasingage[Koh
etal.,2006]andloserhythmicitycompletelyprecedingdeath.
Atthecross-sectionalviewofspecificageinDrosophila(Figure5-2B),earlydevelopment
stageshowsarhythmiccentralclockandemergingbutnotyetsynchronizedperipheral
clocks. Adult stage shows amature clock system composed of the well-functional
centralandperipheralcomponents.Theagingstageshowsarobustcentralclockand
declinedperipheralclocks,andfinallyatthestagebeforedeath,severedisruptionis
foundattheperipherallevelbutnotatthecentrallevel[Luoetal.,2012].
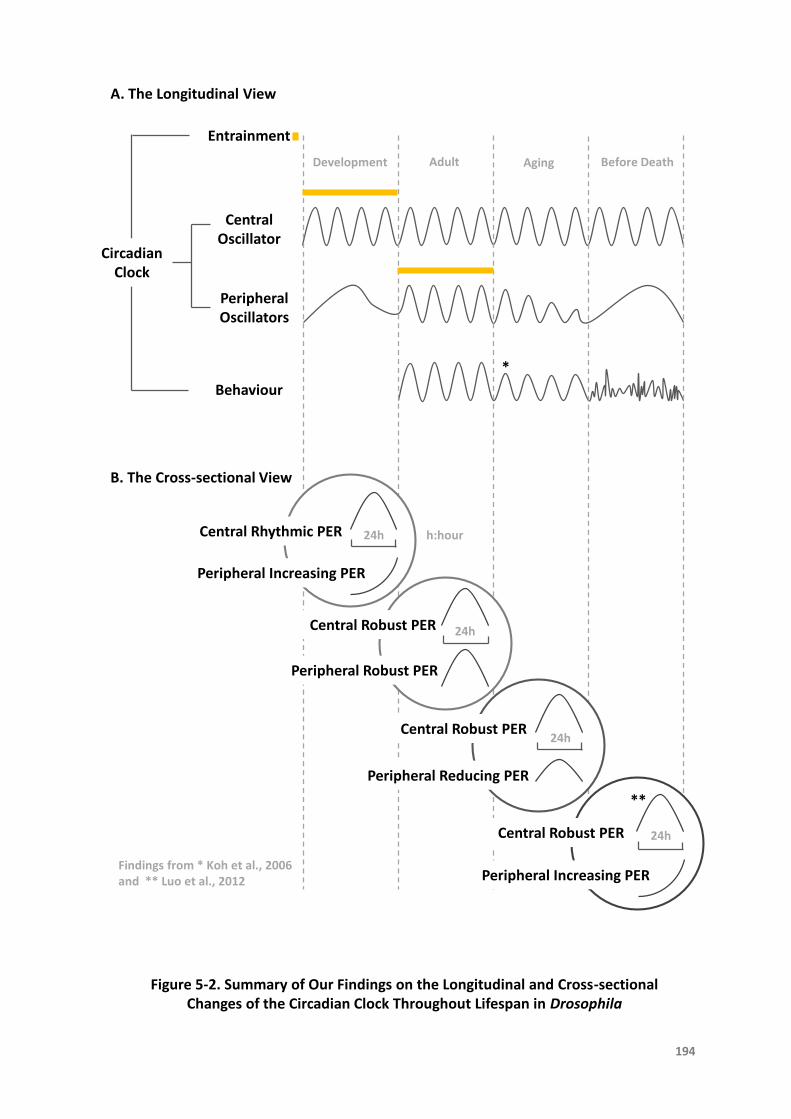
Circadian Clock
A. The Longitudinal View
B. The Cross-sectional View
Entrainment
Central Oscillator
Peripheral Oscillators
Behaviour
Development Adult Aging Before Death
*
Central Rhythmic PER
Peripheral Increasing PER
Central Robust PER 24h
Peripheral Robust PER
Central Robust PER24h
Peripheral Reducing PER
24h
Central Robust PER 24h
**
Peripheral Increasing PER
Figure 5-2. Summary of Our Findings on the Longitudinal and Cross-sectional Changes of the Circadian Clock Throughout Lifespan in Drosophila
194
Findings from * Koh et al., 2006 and ** Luo et al., 2012
h:hour

TheLifeHistoryoftheCircadianClock
195
Within thenetwork systemof the circadian clock, peripheral oscillators areoneof the
crucialelements.Whenthespotlighthasalwaysbeenprojectedonthecentralclock,
itmakesperipheralclockslessthoroughlystudiedeventhoughtheyconstitutealarge
proportion of the circadian organization. Our findings have shed light on these
peripheral clocks,notonlypointingout their laterdevelopmentat thewholebody
level,butalsomoreimportantly,revealingtheirroleasthemajorvictimatbothcritical
times of aging and death. As an important player within the circadian network,
peripheralclocksshouldbevaluedmoreintheirnormalfunctionandtheirmalfunction
duringcertainpathologicalconditions.

TheLifeHistoryoftheCircadianClock
196
5.5TheImplicationinReality
Circadianrhythmsstructurelife.
The coordination of the circadian network has a pronounced impact on metabolic,
physiological and neurological functions, overall health, and disease susceptibility,
throughout life history among various organisms including humans [Gibson et al.,
2009].ItisnotdifficulttoimaginehowhealthwouldsufferwhentheRULEof‘Circadian
Resonance’ischallenged.Thereissubstantialevidenceshowingthatanimpairedclock
can lead to negative consequences, including sleep disorders (with subsequent
cognitiveandotherbehaviourproblems)[VanSomerenandRiemersma-VanDerLek,
2007; Wu and Swaab, 2007; Rakshit and Giebultowicz, 2013], metabolic diseases,
cancer[SaharandSassone-Corsi,2009;Mauryetal.,2010],onsetoflatelifediseases,
increaseofmorbidityandmortalityrisk[Kondratovetal.,2006;Antochetal.,2008;Yu
andWeaver,2011],andreductioninlifespan[LiuandChang,2017].
Circadiandisruptioniscommonlycausedbythemisalignmentoftheexternalinfluenceof
Zeitgebers and internal circadian clock. Symptoms of fatigue, disorientation, and
insomniaareseeninjet-laggedtravellers,aswellasalteredhormoneprofiles,elevated
risksofcardiovasculardiseaseandcancer,andhighmorbidityseen inshiftworkers
[Froy, 2011; Banks et al., 2016]. Aside from the desynchronization between
endogenoustimingsystemandastronomicaltime,conflictscouldalsoexistbetween

TheLifeHistoryoftheCircadianClock
197
centralandperipheralclocksduetotheirdifferentialratesofadjustmentindifferent
circumstances[Davidsonetal.,2009].Inthecaseofaging,misalignmentbetweenthe
centralpacemakerandoscillatorsintheperipheraltissuesisassumedtobeprominent.
Harmonious interaction between the centre and periphery is needed to maintain
criticalhomeostasisandassurehealthyphysiology[BrownandAzzi,2013].Itislikely
thatadditionalpathologiesmayresultfromuncouplingbetweentheprincipalandthe
secondarycircadianoscillators,suchascardiovasculardiseases,metabolicdisorders,
neuropsychiatric disorders and cognitive deficits, sleep disorders, gastrointestinal
diseases, fertility problems, immune dysfunction, and cancer tendency [Arellanes-
Liceaetal.,2014].
Ontheothersideofthecoin,aregularandstrongenvironmentalZeitgeberisespecially
important to the reintroduction of healthy rhythms which may help fight the
deleterious effects of circadianmisalignment. The clinical applications of light and
melatoninascircadiansynchronizershaveshownpromisingresultonhealthbenefits
in aged people with neurodegenerative diseases such as Alzheimer’s disease
[Riemersma-vanderLeketal.,2008;KondratovaandKondratov,2012;Duffyetal.,
2015;Banksetal.,2016].
Considering that the peripheral clocks are a definite target of aging from our study in
Drosophila, more resource should be placed on this relatively unploughed area of
Chronobiology.Especiallywhenspeakingaboutintervention,strategymightneedto
adjust accordingly, based on the different characteristic between the central and

TheLifeHistoryoftheCircadianClock
198
peripheralclocks.Forinstance,foodisamoreefficientagenttoentraintheperipheral
clocks inmammals compared to light. Thus timed food delivery could serve as an
effectivemeanstoresynchronizecircadiantiminginoldindividuals.Furthermore,once
the peripheral clocks are damaged, it inevitably comeswith the desynchronization
between themselves and the central pacemaker. Correspondingly, this couldmake
peripheralclocksbeapotentialcandidateforrescuingcircadianclockimpairment,and
moregenerallyforhealthimprovementindifferentpathologicalsituations.

TheLifeHistoryoftheCircadianClock
199
5.6MessagetotheFuture
Scienceisaboutexplorationoftheunknownsandtruth-seeking.Thisfour-yearjourneyfor
mehasbeenintimidatingyetimpressive,withthebeliefthatwhatevercomestoan
endisjustthebeginningofthenextexpedition.
When we look back a century ago, an inspiring theory was proposed that unicellular
organisms are actually immortal, and the concept of mortality is only applied to
multicellularorganisms[Pearl,1922]. It illustratesthesacrificeoftheevolution into
complexity.Itremindsmeofthisstudythatagingandthusdeathmaylargelydueto
the failureof thesystemrather than individual cellsoreven individual tissues.The
network fails because communication is necessary yet inadequate, leading to a
disintegratedsystem.
Accordingly,wehavestudiedthecircadianchangesatthebodyleveltostartwith,despite
thatthecentralandperipheralplayerscanbedistinguished,thedetailswithineach
tissueandcellarelefttobedisclosedinfuturestudies.Hopefullyintheupcomingera,
every gear and every hinge of the circadian clock can go through high-resolution
dissectionandtheirtime-serieschangescanbecontinuouslyrecordedfrombirthto
deathinfreely-movinganimalsinvivo.
Whenlookintothefuture,understandingthelifetraitsofthecircadianclockhorizontally

TheLifeHistoryoftheCircadianClock
200
andverticallyshouldhaveimplicationonhealthbenefits.Studiesondevelopment-or
age-relateddiseasesmaybeinspiredbytheperspectivesfromChronobiologists,and
thegateforinterdisciplinarycollaborationwillbeopened.

TheLifeHistoryoftheCircadianClock
201
5.7References
AbrahamU,Granada AE,Westermark PO,Heine M,Kramer A,Herzel H. Coupling governs
entrainment range of circadian clocks. Mol Syst Biol.2010Nov 30;6:438. DOI:
10.1038/msb.2010.92.
AgrawalP,HoulJH,GunawardhanaKL,LiuT,ZhouJ,ZoranMJ,HardinPE.DrosophilaCRYentrains
clocksinbodytissuestolightandmaintainspassivemembranepropertiesinanon-clockbody
tissue independent of light. Curr Biol.2017 Aug 21;27(16):2431-2441.e3. DOI:
10.1016/j.cub.2017.06.064.
Allada R,ChungBY. Circadian organization of behavior and physiology inDrosophila. AnnuRev
Physiol.2010;72:605-24.DOI:10.1146/annurev-physiol-021909-135815.
Antoch MP,Gorbacheva VY,Vykhovanets O,Toshkov IA,Kondratov RV,Kondratova AA,Lee C,
NikitinAY.DisruptionofthecircadianclockduetotheClockmutationhaseffectsonagingand
carcinogenesis.CellCycle.2008May1;7(9):1197-204.DOI:10.4161/cc.7.9.5886.
Arellanes-LiceaE,CaldelasI,DeIta-PérezD,Díaz-MuñozM.Thecircadiantimingsystem:arecent
additioninthephysiologicalmechanismsunderlyingpathologicalandagingprocesses.Aging
Dis.2014Jan9;5(6):406-18.DOI:10.14336/AD.2014.0500406.
AtonSJ,ColwellCS,HarmarAJ,WaschekJ,HerzogED.Vasoactiveintestinalpolypeptidemediates
circadianrhythmicityandsynchronyinmammalian clockneurons. Nat Neurosci. 2005
Apr;8(4):476-83.DOI:10.1038/nn1419.
Aujard F,Herzog ED,Block GD. Circadian rhythms in firing rate of individual suprachiasmatic
nucleusneuronsfromadultandmiddle-agedmice.Neuroscience.2001;106(2):255-61.
BanksG,NolanPM,PeirsonSN.Reciprocalinteractionsbetweencircadianclocksandaging.Mamm
Genome.2016Aug;27(7-8):332-40.DOI:10.1007/s00335-016-9639-6.
Biello SM. Circadian clock resetting in the mouse changes with age. Age (Dordr).2009
Dec;31(4):293-303.DOI:10.1007/s11357-009-9102-7.

TheLifeHistoryoftheCircadianClock
202
Brown SA,Azzi A. Peripheralcircadianoscillatorsinmammals. Handb Exp Pharmacol.
2013;(217):45-66.DOI:10.1007/978-3-642-25950-0_3.
CaiA,ScarbroughK,HinkleDA,WisePM.Fetalgraftscontainingsuprachiasmaticnucleirestorethe
diurnal rhythmofCRHandPOMC mRNAinagingrats. Am J Physiol. 1997 Nov;273(5 Pt
2):R1764-70.
CollinsB,KaplanHS,CaveyM,LelitoKR,BahleAH,ZhuZ,MacaraAM,RomanG,ShaferOT,BlauJ.
Differentiallytimedextracellularsignalssynchronizepacemakerneuronclocks.PLoSBiol.2014
Sep30;12(9):e1001959.DOI:10.1371/journal.pbio.1001959.
DavidsonAJ,Castanon-CervantesO,LeiseTL,MolyneuxPC,HarringtonME.Visualizingjetlaginthe
mousesuprachiasmaticnucleusandperipheralcircadiantimingsystem.EurJNeurosci.2009
Jan;29(1):171-80.DOI:10.1111/j.1460-9568.2008.06534.x.
DavidsonAJ,Sellix MT,Daniel J,Yamazaki S,Menaker M,Block GD. Chronicjet-lagincreases
mortalityinagedmice.CurrBiol.2006Nov7;16(21):R914-6.DOI:10.1016/j.cub.2006.09.058.
Dolatshad H,Cary AJ,Davis FC. Differentialexpressionof the circadian clock in maternal and
embryonictissuesofmice. PLoS One.2010 Mar 24;5(3):e9855. DOI:
10.1371/journal.pone.0009855.
Dibner C,Schibler U,Albrecht U. The mammalian circadian timing system: organization and
coordination of central and peripheral clocks. Annu Rev Physiol.2010;72:517-49. DOI:
10.1146/annurev-physiol-021909-135821.
Dubruille R,Emery P. A plastic clock: how circadian rhythms respond to environmental cues in
Drosophila.MolNeurobiol.2008Oct;38(2):129-45.DOI:10.1007/s12035-008-8035-y.
DuffyJF,ZittingKM,ChinoyED.AgingandCircadianRhythms.SleepMedClin.2015Dec;10(4):423-
34.DOI:10.1016/j.jsmc.2015.08.002.
EnokiR,Oda Y,Mieda M,Ono D,Honma S,Honma KI. Synchronous circadian voltage rhythms
withasynchronouscalciumrhythmsinthesuprachiasmaticnucleus.ProcNatlAcadSciUSA.
2017Mar21;114(12):E2476-E2485.DOI:10.1073/pnas.1616815114.
FarajniaS,Deboer T,Rohling JH,Meijer JH,Michel S. Aging of the suprachiasmatic clock.
Neuroscientist.2014Feb;20(1):44-55.DOI:10.1177/1073858413498936.

TheLifeHistoryoftheCircadianClock
203
FarajniaS,MichelS,DeboerT,vanderLeestHT,HoubenT,RohlingJH,RamkisoensingA,Yasenkov
R,MeijerJH.Evidenceforneuronaldesynchronyintheagedsuprachiasmaticnucleusclock.J
Neurosci.2012Apr25;32(17):5891-9.DOI:10.1523/JNEUROSCI.0469-12.2012.
FonsecaCostaSS,RippergerJA.Impactofthecircadianclockontheagingprocess.FrontNeurol.
2015Mar6;6:43.DOI:10.3389/fneur.2015.00043.
FroyO. Circadian rhythms, aging, and life span in mammals. Physiology (Bethesda).
2011Aug;26(4):225-35.DOI:10.1152/physiol.00012.2011.
GibsonEM,WilliamsWP,KriegsfeldLJ.Aging inthecircadiansystem:considerationsforhealth,
disease prevention and longevity. Exp Gerontol. 2009 Jan-Feb;44(1-2):51-6. DOI:
10.1016/j.exger.2008.05.007.
GiebultowiczJM,LongDM.Ageingandcircadianrhythms.CurrOpinInsectSci.2015Feb1;7:82-86.
DOI:10.1016/j.cois.2015.03.001.
Girardot F,Lasbleiz C,Monnier V,TricoireH. Specific age-related signatures inDrosophila body
partstranscriptome.BMCGenomics.2006Apr4;7:69.DOI:10.1186/1471-2164-7-69.
HonmaS,NakamuraW,ShirakawaT,HonmaK.Diversityinthecircadianperiodsofsingleneurons
oftheratsuprachiasmaticnucleusdependsonnuclearstructureandintrinsicperiod.Neurosci
Lett.2004Apr1;358(3):173-6.DOI:10.1016/j.neulet.2004.01.022.
Honma S,Shirakawa T,Katsuno Y,Namihira M,Honma K. Circadian periods of single
suprachiasmaticneuronsinrats.NeurosciLett.1998Jul10;250(3):157-60.
HoodS,Amir S. Theagingclock:circadian rhythmsandlaterlife. J Clin Invest.2017Feb
1;127(2):437-446.DOI:10.1172/JCI90328.
HurdMW,RalphMR.Thesignificanceofcircadianorganizationforlongevityinthegoldenhamster.
JBiolRhythms.1998Oct;13(5):430-6.DOI:10.1177/074873098129000255.
Kalló I,Kalamatianos T,Piggins HD,Coen CW. Ageingand the diurnal expression of mRNAs for
vasoactiveintestinalpeptideand for theVPAC2andPAC1receptorsin the suprachiasmatic
nucleusofmalerats. J Neuroendocrinol.2004 Sep;16(9):758-66. DOI: 10.1111/j.1365-
2826.2004.01232.x.

TheLifeHistoryoftheCircadianClock
204
KawakamiF,OkamuraH,TamadaY,MaebayashiY,FukuiK,IbataY.Lossofday-nightdifferences
in VIP mRNA levels in the suprachiasmatic nucleus of aged rats. Neurosci Lett.1997 Jan
31;222(2):99-102.
Koh K, Evans JM, Hendricks JC, Sehgal A. A Drosophila model for age-associated changes in
sleep:wake cycles. Proc Natl Acad Sci U S A. 2006 Sep 12;103(37):13843-7. DOI:
10.1073/pnas.0605903103.
Kondratova AA, Kondratov RV. The circadian clock and pathology of the ageing brain. Nat Rev
Neurosci.2012Mar7;13(5):325-35.DOI:10.1038/nrn3208.
KondratovRV,KondratovaAA,GorbachevaVY,VykhovanetsOV,AntochMP.Earlyagingandage-
relatedpathologies inmicedeficient inBMAL1, thecorecomponentof thecircadianclock.
GenesDev.2006Jul15;20(14):1868-73.DOI:10.1101/gad.1432206.
KonopkaRJ,BenzerS.ClockmutantsofDrosophilamelanogaster.ProcNatlAcadSciUSA.1971
Sep;68(9):2112–6.
KornmannB,SchaadO,ReinkeH,SainiC,SchiblerU.Regulationof circadiangeneexpression in
liver by systemic signals and hepatocyte oscillators. Cold Spring Harb Symp Quant Biol.
2007;72:319-30.DOI:10.1101/sqb.2007.72.041.
KrajnakK,KashonML,RosewellKL,WisePM.Agingalterstherhythmicexpressionofvasoactive
intestinalpolypeptidemRNAbutnotargininevasopressinmRNAinthesuprachiasmaticnuclei
offemalerats.JNeurosci.1998Jun15;18(12):4767-74.
KrishnanB,LevineJD,LynchMK,DowseHB,FunesP,HallJC,HardinPE,DryerSE.Anewrolefor
cryptochromeinaDrosophilacircadianoscillator.Nature.2001May17;411(6835):313-7.DOI:
10.1038/35077094.
Krupp JJ,Billeter JC,Wong A,Choi C,Nitabach MN,Levine JD. Pigment-dispersing factor
modulatespheromoneproductioninclockcellsthatinfluencematinginDrosophila. Neuron.
2013Jul10;79(1):54-68.DOI:10.1016/j.neuron.2013.05.019.
KumarS,VazeKM,KumarD,SharmaVK.Selectionforearlyandlateadultemergencealtersthe
rateofpre-adultdevelopmentinDrosophilamelanogaster.BMCDevBiol.2006;6(1):57.DOI:
10.1186/1471-213X-6-57.

TheLifeHistoryoftheCircadianClock
205
KyriacouCP,OldroydM,WoodJ,SharpM,HillM.Clockmutationsalterdevelopmentaltimingin
Drosophila.Heredity.1990Jun;64(Pt3):395-401.
LiH,SatinoffE. Fetal tissue containing the suprachiasmatic nucleus restores multiple circadian
rhythmsinoldrats.AmJPhysiol.1998Dec;275(6Pt2):R1735-44.
LinY,StormoGD,TaghertPH.Theneuropeptidepigment-dispersingfactorcoordinatespacemaker
interactionsin theDrosophila circadiansystem. J Neurosci.2004 Sep 8;24(36):7951-7. DOI:
10.1523/JNEUROSCI.2370-04.2004.
LiuAC,WelshDK,KoCH,TranHG,ZhangEE,PriestAA,BuhrED,SingerO,MeekerK,Verma IM,
Doyle FJ 3rd,Takahashi JS,Kay SA. Intercellular coupling confers robustness against
mutationsin theSCNcircadian clocknetwork. Cell.2007 May 4;129(3):605-16. DOI:
10.1016/j.cell.2007.02.047.
Liu C,Weaver DR,Strogatz SH,Reppert SM. Cellularconstructionof acircadian clock: period
determinationinthesuprachiasmaticnuclei.Cell.1997Dec12;91(6):855-60.
LiuF,Chang HC. Physiologicallinksofcircadian clockandbiological clockofaging. Protein Cell.
2017Jul;8(7):477-488.DOI:10.1007/s13238-016-0366-2.
LuoW,ChenW-F,YueZ,ChenD, SowcikM,SehgalA, ZhengX.Old flieshavea robust central
oscillatorbutweakerbehavioralrhythmsthatcanbeimprovedbygeneticandenvironmental
manipulations.AgingCell.2012Jun;11(3):428-38.DOI:10.1111/j.1474-9726.2012.00800.x.
MattisJ,SehgalA.Circadianrhythms,sleep,anddisordersofaging.TrendsEndocrinolMetab.2016
Apr;27(4):192-203.DOI:10.1016/j.tem.2016.02.003.
Maury E,Ramsey KM,Bass J. Circadian rhythmsandmetabolic syndrome: from experimental
geneticstohumandisease. Circ Res.2010 Feb 19;106(3):447-62. DOI:
10.1161/CIRCRESAHA.109.208355.
MaywoodES,ReddyAB,WongGK,O'Neill JS,O'Brien JA,McMahonDG,HarmarAJ,OkamuraH,
HastingsMH.Synchronizationandmaintenanceoftimekeepinginsuprachiasmaticcircadian
clock cells by neuropeptidergic signaling. Curr Biol.2006Mar 21;16(6):599-605. DOI:
10.1016/j.cub.2006.02.023.

TheLifeHistoryoftheCircadianClock
206
McKillopLE,FisherSP,CuiN,PeirsonSN,FosterRG,WaffordKA,VyazovskiyVV.Effectsofagingon
corticalneuraldynamicsandlocalsleephomeostasisinmice. J Neurosci.2018 Apr
18;38(16):3911-3928.DOI:10.1523/JNEUROSCI.2513-17.2018.
MohawkJA,GreenCB,TakahashiJS.Centralandperipheralcircadianclocksinmammals.AnnuRev
Neurosci.2012;35:445-62.DOI:10.1146/annurev-neuro-060909-153128.
MyersEM,Yu J,Sehgal A. Circadian control of eclosion: interaction between a central and
peripheralclockinDrosophilamelanogaster.CurrBiol.2003Mar18;13(6):526-33.
NagoshiE,SainiC,BauerC,LarocheT,NaefF,SchiblerU.Circadiangeneexpressioninindividual
fibroblasts: cell-autonomousandself-sustainedoscillatorspass time todaughter cells.Cell.
2004Nov24;119(5):693-705.DOI:10.1016/j.cell.2004.11.015.
Nakamura TJ, NakamuraW, Tokuda IT, Ishikawa T, Kudo T, Colwell CS, Block GD. Age-related
changesinthecircadiansystemunmaskedbyconstantconditions(1,2,3).eNeuro.2015Sep
22;2(4).DOI:10.1523/ENEURO.0064-15.2015.
NakamuraTJ,NakamuraW,YamazakiS,KudoT,CutlerT,ColwellCS,BlockGD.Age-relateddecline
in circadianoutput. JNeurosci.2011 Jul13;31(28):10201-5.DOI:10.1523/JNEUROSCI.0451-
11.2011.
Nakamura TJ, Takasu NN, Nakamura W. The suprachiasmatic nucleus: age-related decline in
biologicalrhythms.JPhysiolSci.2016Sep;66(5):367-74.DOI:10.1007/s12576-016-0439-2.
Nygård M,Hill RH,Wikström MA,Kristensson K. Age-related changes in electrophysiological
propertiesof themousesuprachiasmatic nucleusin vitro. Brain Res Bull.2005 Mar
15;65(2):149-54.DOI:10.1016/j.brainresbull.2004.12.006.
PearlP.Thebiologyofdeath.Philadelphia:JBLippincottCompany.1922.
Peng Y,Stoleru D,Levine JD,Hall JC,Rosbash M. Drosophilafree-running rhythms require
intercellular communication. PLoS Biol.2003 Oct;1(1):E13. Epub 2003 Sep 15. DOI:
10.1371/journal.pbio.0000013.
PittendrighC,BruceV,KausP.Onthesignificanceoftransientsindailyrhythms.ProcNatlAcadSci
USA.1958Sep15;44(9):965-73.

TheLifeHistoryoftheCircadianClock
207
Plautz JD,Kaneko M,Hall JC,Kay SA. Independentphotoreceptive circadian clocks throughout
Drosophila.Science.1997Nov28;278(5343):1632-5.
Rakshit K,Giebultowicz JM.Cryptochrome restoresdampened circadian rhythmsandpromotes
healthspaninagingDrosophila.AgingCell.2013Oct;12(5):752-62.DOI:10.1111/acel.12100.
DOI:10.1111/acel.12100.
Riemersma-vanderLekRF,SwaabDF,TwiskJ,HolEM,HoogendijkWJ,VanSomerenEJ.Effectof
lightandmelatoninoncognitiveandnoncognitivefunctioninelderlyresidentsofgroupcare
facilities: a randomized controlled trial. JAMA.2008 Jun 11;299(22):2642-55. DOI:
10.1001/jama.299.22.2642.
Sabado V,Vienne L,Nagoshi E. Evaluating the autonomy of theDrosophilacircadian clock in
dissociated neuronal culture. Front Cell Neurosci.2017 Oct 12;11:317. DOI:
10.3389/fncel.2017.00317.
SaharS,Sassone-CorsiP.Metabolismandcancer:thecircadianclockconnection.NatRevCancer.
2009Dec;9(12):886-96.DOI:10.1038/nrc2747.
SatinoffE,LiH,TchengTK,LiuC,McArthurAJ,MedanicM,GilletteMU.Do the suprachiasmatic
nucleioscillateinoldratsastheydoinyoungones?AmJPhysiol.1993Nov;265(5Pt2):R1216-
22.DOI:10.1152/ajpregu.1993.265.5.R1216.
TaharaY,TakatsuY,ShiraishiT,KikuchiY,YamazakiM,MotohashiH,MutoA,SasakiH,Haraguchi
A,Kuriki D,Nakamura TJ,Shibata S. Age-relater circadian disorganization caused by
sympatheticdysfunctioninperipheralclock regulation. NPJ Aging Mech Dis.2017 Jan
5;3:16030.DOI:10.1038/npjamd.2016.30.
Top D,Young MW. Coordinationbetweendifferentiallyregulatedcircadian clocks generates
rhythmicbehavior. Cold Spring Harb Perspect Biol.2017 Sep 11. DOI:
10.1101/cshperspect.a033589.
Umezaki Y, Yoshii T, Kawaguchi T, Helfrich-Förster C, Tomioka K. Pigment-dispersing factor is
involvedinage-dependentrhythmchangesinDrosophilamelanogaster.JBiolRhythms.2012
Dec;27(6):423-32.DOI:10.1177/0748730412462206.
ValloneD,LahiriK,DickmeisT,FoulkesNS.Starttheclock!Circadianrhythmsanddevelopment.
DevDyn.2007Jan;236(1):142-55.DOI:10.1002/dvdy.20998.

TheLifeHistoryoftheCircadianClock
208
VanReethO,ZhangY,ZeePC,TurekFW.Graftingfetalsuprachiasmaticnucleiinthehypothalamus
of oldhamstersrestores responsivenessof thecircadian clockto a phase shifting stimulus.
BrainRes.1994Apr18;643(1-2):338-42.
VanSomerenEJ,Riemersma-VanDerLekRF.Livetotherhythm,slavetotherhythm.SleepMed
Rev.2007Dec;11(6):465-84.DOI:10.1016/j.smrv.2007.07.003.
VasalouC,HerzogED,HensonMA.Small-worldnetworkmodelsof intercellularcouplingpredict
enhancedsynchronizationin thesuprachiasmatic nucleus. J Biol Rhythms.2009
Jun;24(3):243-54.DOI:10.1177/0748730409333220.
Viswanathan N,Davis FC. Suprachiasmatic nucleus grafts restore circadian function in aged
hamsters.BrainRes.1995Jul17;686(1):10-6.
WangJL,LimAS,ChiangWY,HsiehWH,LoMT,SchneiderJA,BuchmanAS,BennettDA,HuK,Saper
CB.Suprachiasmaticneuronnumbersandrest-activitycircadianrhythmsinolderhumans.Ann
Neurol.2015Aug;78(2):317-22.DOI:10.1002/ana.24432.
Watanabe A,Shibata S,Watanabe S. Circadian rhythm of spontaneous neuronal activity in the
suprachiasmaticnucleusofoldhamsterinvitro.BrainRes.1995Oct16;695(2):237-9.
Webb AB,Angelo N,Huettner JE,Herzog ED. Intrinsic,nondeterministiccircadian rhythm
generation inidentifiedmammalianneurons. Proc Natl Acad Sci U S A.2009 Sep
22;106(38):16493-8.DOI:10.1073/pnas.0902768106.
WeissR,BartokO,MezanS,MalkaY,KadenerS.Synergisticinteractionsbetween themolecular
and neuronalcircadiannetworksdriverobust behavioralcircadian rhythmsinDrosophila
melanogaster.PLoSGenet.2014Apr3;10(4):e1004252.DOI:10.1371/journal.pgen.1004252.
Welsh DK,Logothetis DE,Meister M,Reppert SM. Individual neurons dissociated from rat
suprachiasmatic nucleusexpressindependentlyphased circadianfiringrhythms. Neuron.
1995Apr;14(4):697-706.
WelshDK,YooSH,LiuAC,TakahashiJS,KaySA.Bioluminescenceimagingofindividualfibroblasts
revealspersistent,independentlyphasedcircadianrhythmsofclockgeneexpression.CurrBiol.
2004Dec29;14(24):2289-95.DOI:10.1016/j.cub.2004.11.057.

TheLifeHistoryoftheCircadianClock
209
WuYH,SwaabDF.Disturbanceandstrategiesforreactivationof the circadian rhythm system in
aging andAlzheimer'sdisease. Sleep Med.2007 Sep;8(6):623-36. Epub 2007 Mar 26. DOI:
10.1016/j.sleep.2006.11.010.
YamazakiS,NumanoR,AbeM,HidaA,TakahashiR,UedaM,BlockGD,SakakiY,MenakerM,Tei
H. Resettingcentralandperipheralcircadianoscillatorsintransgenic rats. Science. 2000Apr
28;288(5466):682-5.
Yamazaki S, StraumeM,TeiH, Sakaki Y,MenakerM,BlockGD.Effectsof agingon central and
peripheral mammalian clocks. Proc Natl Acad Sci U S A. 2002 Aug 6;99(16):10801-6. DOI:
10.1073/pnas.152318499.
YooSH,YamazakiS,LowreyPL,ShimomuraK,KoCH,BuhrED,SiepkaSM,HongHK,OhWJ,YooOJ,
Menaker M,Takahashi JS. PERIOD2::LUCIFERASEreal-time reporting of circadian dynamics
revealspersistentcircadianoscillatorsinmouseperipheraltissues.ProcNatlAcadSciUSA.
2004Apr13;101(15):5339-46.DOI:10.1073/pnas.0308709101.
Yoshii T, Wulbeck C, Sehadova H, Veleri S, Bichler D, Stanewsky R, Helfrich-Förster C. The
neuropeptide pigment-dispersing factor adjusts period and phase of Drosophila's clock. J
Neurosci.2009Feb25;29(8):2597–610.DOI:10.1523/JNEUROSCI.5439-08.2009.
YuEA,WeaverDR.Disruptingthecircadianclock:gene-specificeffectsonaging,cancer,andother
phenotypes.Aging(AlbanyNY).2011May;3(5):479-93.DOI:10.18632/aging.100323.
Zhou JN,Hofman MA,Swaab DF. VIPneuronsin thehumanSCNinrelationtosex,age, and
Alzheimer’sdisease.NeurobiolAging.1995Jul-Aug;16(4):571-6.
Zhou JN,Swaab DF. Activationanddegenerationduringaging: amorphometricstudyof the
humanhypothalamus. Microsc Res Tech.1999 Jan 1;44(1):36-48. DOI: 10.1002/(SICI)1097-
0029(19990101)44:1<36::AID-JEMT5>3.0.CO;2-F.

The life story of a kingdom in my telling has come to an end.
It arises from the ground as bricks and tiles, stands up high, and fades into
dilapidated walls and dust.
Its beauty and loneliness, glory and shadow, shall be remembered.

APPENDIX AP
210

Contents lists available at ScienceDirect
Ageing Research Reviews
journal homepage: www.elsevier.com/locate/arr
Review
The functional changes of the circadian system organization in aging
Jia Zhao, Guy Robert Warman, James Frederick Cheeseman⁎
Department of Anaesthesiology, School of Medicine, The University of Auckland, Park Road, Grafton, Auckland, 1023, New Zealand
A R T I C L E I N F O
Keywords:Circadian networkCentral and peripheral oscillatorsCircadian rhythmsClock genesSuprachiasmatic nuclei
A B S T R A C T
The circadian clock drives periodic oscillations at different levels of an organism from genes to behavior. Thistiming system is highly conserved across species from insects to mammals and human beings. The question ofhow the circadian clock is involved in the aging process continues to attract more attention. We aim to char-acterize the detrimental impact of aging on the circadian clock organization. We review studies on differentcomponents of the circadian clock at the central and periperal levels, and their changes in aged rodents andhumans, and the fruit fly Drosophila. Intracellular signaling, cellular activity and intercellular coupling in thecentral pacemaker have been found to decline with advancing age. Evidence of degradation of the molecularclockwork reflected by clock gene expression in both central and peripheral oscillators due to aging is in-adequate. The findings on age-associated molecular and functional changes of peripheral clocks are mixed. Weconclude that aging can affect the circadian clock organization at various levels, and the impairment of thecentral network may be a fundamental mechanism of circadian disruption seen in aged species.
1. Introduction
The circadian clock is the endogenous pacemaker that generates the˜24 h daily rhythms in behavior, physiology, metabolism and cellularprocesses, such as body temperature, endocrine secretion, sleep-wakecycles and locomotor activity (Panda et al., 2002). These circadianoutput rhythms and the intrinsic clock mechanism not only synchronizewith environmental rhythms (entrainment) but also persist underconstant conditions (free-running).
Aging is the inevitable process of functional decline and the circa-dian timing system is also susceptible, showing progressive deteriora-tion with advancing age. As a prominent signature of aging, the declineof circadian output rhythms has been widely observed in fruit flies andmammals including humans (Kondratova and Kondratov, 2012). Inhumans, the circadian output changes associated with aging include thereduction of the amplitude and earlier timing of phase of daily rhythms,both evident in body temperature and sleep-wake cycle, as well as adisruption of quantity and quality of nocturnal sleep (Duffy et al., 2015;Hood and Amir, 2017). Mammalian studies on aging demonstratechange in the free-running period, shift in the phase and weakening ofrhythmicity, which have been reported in body temperature, locomotoractivity patterns and drinking behavior (Froy, 2011). Similarly, pro-longed free-running period, decline in sleep consolidation and decreaseof overall rest-activity rhythm strength, have been revealed in Droso-phila (Koh et al., 2006; Umezaki et al., 2012).
It is widely accepted that the molecular mechanism of the circadianclock can be found in nearly all cells of the body (Brown and Azzi,2013). Correspondingly, the circadian timing system is composed of anumber of tissues and organs comprising cell-autonomous circadianoscillators. The well-organized circadian network is formed with in-terconnected cellular oscillators within individual tissues and organs,and these cooperate with each other within the system (Mohawk et al.,2012). Arguably the impact of aging on the circadian clock is likely tobe complex, with the possibility that the clock can be arrested at thecellular, tissue/organ or systemic levels resulting in desynchrony. In thefirst part of this review, we introduce the architecture of the circadianclock system, and in the second, summarize how aging affects thefunction of different parties in the clock organization.
1.1. The functional organization of the circadian clock
The circadian clock consists of the central pacemaker and numerousperipheral oscillators, and each can be dissected into functional unitsshowing circadian properties at the cellular and tissue/organ levels. Inview of this, a circadian pattern of clock gene expression has beenshown in individual cells and the tissue of the mammalian central clock(Honma, 2018). Likewise, evidence of the existence of clock mechanismhas been found in peripheral cells and tissues/organs (Dibner et al.,2010). Ultimately, these circadian components collaboratively con-tribute to the timed functioning of the organism.
https://doi.org/10.1016/j.arr.2019.04.006Received 16 December 2018; Received in revised form 14 April 2019; Accepted 24 April 2019
⁎ Corresponding author.E-mail address: [email protected] (J.F. Cheeseman).
Ageing Research Reviews 52 (2019) 64–71
Available online 29 April 20191568-1637/ © 2019 Elsevier B.V. All rights reserved.
T

1.1.1. Central clock at the tissue, intracellular and intercellular levelsThe central clock in mammals is situated in the suprachiasmatic
nucleus (SCN), which has been identified as both necessary and suffi-cient for controlling circadian physiology and behavior through a seriesof classic lesion and transplantation experiments (Lehman et al., 1987;Ralph et al., 1990; Stephan and Zucker, 1972). The SCN can be dividedinto regions exhibiting sequential spatiotemporal activation patterns ofclock gene expression (Foley et al., 2011; Yamaguchi et al., 2003) andphase adjustment at different rates in response to light stimuli(Nakamura et al., 2005), as well as distinctive distribution of calciumrhythm (Enoki et al., 2012, 2017). The SCN is mainly entrained by thelight, which is perceived by the retina and then transmitted to the SCNvia the retinohypothalamic tract (RHT). The SCN receives direct photicinput from intrinsically photoreceptive retinal ganglion cells (ipRGCs),in this case, the ipRGCs-expressing photo-pigment melanopsin areconsidered the principal circadian photoreceptors (Do and Yau, 2010).
In the Drosophila brain, the central circadian clock in each hemi-sphere is composed of approximate 150 pacemaker neurons, which aredivided into multiple clusters based on their location, size, neuropep-tide expression and function. These are dorsal lateral neurons (LNd),pigment dispersing factor (PDF)-expressing large ventral lateral neu-rons (l-LNv), lateral posterior neurons (LPN), PDF-positive small ventrallateral neurons (s-LNv), PDF-negative s-LNv, anterior dorsal neurons 1(DN1a), posterior dorsal neurons 1 (DN1p), dorsal neurons 2 (DN2) anddorsal neurons 3 (DN3) (Helfrich-Förster, 2005).
First and foremost, the circadian clock is driven by the transcription-translation feedback loops of clock gene expression existing at a cellularlevel. Most clock genes are well conserved from insects to humans and,with few exceptions, play similar roles in the timekeeping mechanism.The details of transcription-translation feedback loops in animals aredescribed elsewhere (e.g. Hardin and Panda, 2013). Briefly, in Droso-phila, the negative feedback loop is mainly composed of gene period(per) and timeless (tim), whose translational products PERIOD (PER) andTIMELESS (TIM) in cytoplasm form a heterodimer. This enters thenucleus to inhibit the transcription of per and tim themselves. The po-sitive feedback loop primarily consists of the gene clock (clk) and cycle(cyc), whose corresponding proteins CLOCK (CLK) and CYCLE (CYC)heterodimerize in the nucleus. They then facilitate transcription ofother clock genes such as per and tim by recognizing their specializedpromoter regions known as the E-box. These two feedback loops workclosely together in an antiphase manner to generate circadian rhythms.The circadian molecular mechanism is mirrored in mammals, with apair of transcriptional factors CLOCK and BMAL1 which form hetero-dimer to activate the transcription of genes encoding their repressors,PERIOD 1–3 (PERs) and CRYPTOCHROME 1–2 (CRYs).
Secondly, in order to produce functional cellular oscillations withinthe SCN, reciprocal interaction between molecular and cytosolic clocksis necessary (O’Neill and Reddy, 2012). The relationships among cel-lular membrane events, intracellular signaling cascades and transcrip-tional-translational regulation have been increasingly studied but nottotally elucidated. The molecular clock machinery in SCN neuronsdrives neural activities such as a daily oscillation in spontaneous firingrate (namely membrane excitability or action potential) (Enoki et al.,2017). This in turn plays an essential role in the cellular expression ofclock genes as well as the coupling between individual neurons withinthe SCN (Colwell, 2011; Yamaguchi et al., 2003). Recent data alsosuggest the essential contribution of intracellular signaling to thetimekeeping mechanisms, and the calcium rhythmicity is one such ex-ample (O’Neill and Reddy, 2012). Found at a single-cell level in theSCN, robust circadian calcium rhythms play an important role tomediate the input signals to the core molecular loop for circadianrhythm generation (Enoki et al., 2012) and regulate the synchroniza-tion of rhythmicity across the population of SCN clock cells (Nahmet al., 2005). In Drosophila as in mammals, membrane depolarizationand cellular signaling are similarly important for clock function at theintracellular level (Harrisingh et al., 2007; Nitabach et al., 2002) and at
the intercellular level (Nitabach and Taghert, 2008).Thirdly, individual cellular oscillators within the SCN are synchro-
nized between each other through intercellular coupling, so that theentire SCN can perform as an efficient network system. In this case, theSCN population of neurons can be constrained to a narrow range ofparameters such as ‘free-running period’ corresponding to the circadianoutput rhythms (Herzog et al., 2004; Honma et al., 2004). For instance,the circadian spontaneous firing rhythms are synchronized throughoutthe SCN slice (Enoki et al., 2017), though dispersed individual neuronsdisplay heterogeneity in periods, phases and strength of oscillation(Honma et al., 1998, 2004; Liu et al., 1997; Welsh et al., 1995). Inaddition, the essential role of intercellular coupling is also emphasizedby the evidence that densely cultured neurons show not only highersynchrony but also stronger rhythmicity than dispersed ones (Atonet al., 2005; Webb et al., 2009).
The specific coupling mechanisms which maintain the SCN neuronssynchronized as a functional unit include synaptic signaling and neu-ropeptides (Evans, 2016) and the latter one is so far believed to beunique to the SCN (Liu and Chang, 2017). Acting as the most importantneurotransmitter in the SCN, vasoactive intestinal polypeptide (VIP) isnecessary to synchronize the pace of timekeeping among individualneurons (Aton et al., 2005; Maywood et al., 2006). It is a given thatinhibition of VIP-mediated coupling leads to loss of synchrony in clockgene expression throughout the SCN tissue (Abraham et al., 2010).Although the role of inhibitory neurotransmitter gamma-aminobutyricacid (GABA) in the SCN neuronal synchronization is still unclear, it ishypothesized that VIP works with GABA to promote synchrony ofheterogeneous oscillators in the SCN. In this model, VIP produced byVIP-secreting cells in the SCN works as an essential driver for syn-chronization while GABA generated from both VIP-secreting and non-VIP-secreting cells serves as a negative feedback mechanism to balancecell coupling (Kingsbury et al., 2016).
Similar to the instability of isolated individual cellular oscillators inthe SCN, single Drosophila central clock neurons are weak oscillatorsthat need to be in an intact circadian circuit to generate robust circa-dian rhythms (Sabado et al., 2017). A sophisticated network is formedby the intercellular coupling among different clusters of clock neuronsin Drosophila, and PDF strongly resembles those functions described forVIP in the SCN. Though only released by s-LNv and l-LNv, PDF also feedsback to non-PDF neurons, playing a key role in communication withinthe central clock neuronal network (Top and Young, 2017). Clockneurons from flies lacking PDF or PDF receptors are uncoupled andshow impaired rhythmicity (Collins et al., 2014; Lin et al., 2004; Penget al., 2003; Yoshii et al., 2009).
In summary, synchronization among the central pacemaker neuronsrequires neuropeptide signaling (VIP in the mammalian SCN and PDF inthe Drosophila brain), regulated by a reciprocal interaction between themolecular clockwork and rhythmic neuronal activity, which in turndepends on intracellular signaling (Herzog et al., 2017).
1.1.2. Peripheral clocks and their connection with the central clockThe ubiquity of the circadian clock mechanism assures that aside
from the central clock, endogenous oscillators exist in most cellsthroughout the body. These individual cells and the functional tissues/organs they comprise, are the origin of peripheral clocks. In many cases,peripheral clocks are specifically referred to peripheral tissues thatshow circadian clock property (Yamazaki et al., 2000; Yoo et al., 2004).Mammalian peripheral oscillators residing in cells/tissues/organs showcircadian properties outside the SCN, including various non-SCN brainregions (such as the hypothalamus, forebrain, olfactory bulb and pinealgland) and non-neuronal tissues/organs (such as the liver, kidney,muscle, adipose tissue and blood cells (O’Neill and Reddy, 2011;Cermakian and Boivin, 2009). Peripheral clocks appear to have a si-milar molecular makeup to the oscillators in the SCN (Dibner et al.,2010; Yagita et al., 2001), despite the fact that there are large differ-ences in the relative contributions of individual clock components as
J. Zhao, et al. Ageing Research Reviews 52 (2019) 64–71
65

well as the largely non-overlapping downstream genes under circadiancontrol (Mohawk et al., 2012). Outside the central clock in the brain,peripheral clocks in Drosophila are found in almost all tissues whichdisplay rhythmic per expression (Liu et al., 1988; Plautz et al., 1997),showing the same core molecular program present in central pacemakerneurons with diverse tissue-specificity (Ceriani et al., 2002; Krishnanet al., 2001).
In normal circumstances, circadian oscillators in individual cells aresynchronized within a tissue and individual tissues are kept in a stablephase-relationship with each other within the circadian system. In viewof this, the SCN is not a generator of peripheral rhythms, instead being acoordinator and synchronizer at the head of a distributed organizationof individual clocks, mainly through neural and humoral pathways(Balsalobre, 2002). Many fundamental properties of circadian oscilla-tions in peripheral clocks in vivo (Tahara et al., 2012) and in vitro(Izumo et al., 2014) have been shown to persist even in the absence ofSCN function. Meanwhile, it has been reported that clock gene ex-pression dampens in mammalian peripheral tissues (such as the liver,spleen, kidney, heart and lung) in vitro within days in constant condi-tions, but sustains in the SCN explants for weeks (Yamazaki et al.,2000). Furthermore, in constant conditions, loss of the SCN functionresults in peripheral clocks that become desynchronized (Izumo et al.,2014; Yoo et al., 2004). Likewise, peripheral tissues in Drosophila can beclearly distinguished from the central oscillators due to their rapiddampening in constant conditions (Peng et al., 2003). These resultsarise from the fact that peripheral clocks are oscillators that requireinternal or external signals (including signals from the central clock) tosynchronize and thus sustain their circadian rhythms, whereas thecentral clock is able to tick independently without entraining signals(self-sustaining). One possible explanation is that peripheral clocks lackthe intercellular coupling mechanism that is unique to the central pa-cemaker (Brown and Azzi, 2013). Deficiency in coupling leads to a lossof synchrony among individual cells and thus dampening of the en-semble rhythm at the peripheral population level, from lacking internalor external cues (Welsh et al., 2004). Taking the argument further, thiscould also be the reason why peripheral tissues are more easily affectedwhen facing challenges such as genetic perturbation in both mammals(Liu et al., 2007) and Drosophila (Weiss et al., 2014).
The coordination among peripheral clocks in Drosophila, which canrespond directly to the same entrainment cues that set the phase of thecentral pacemaker neurons in the brain, is thought to occur mainly vialight and temperature sensitive intracellular pathways (Agrawal et al.,2017; Dubruille and Emery, 2008; Krishnan et al., 2001). In this way,some peripheral clocks can maintain synchrony with external en-vironmental cues independent of input from the central clock, such asthe Malpighian tubules (Giebultowicz and Hege, 1997) and the antenna(Krishnan et al., 1999; Tanoue et al., 2004). Other peripheral oscillatorssuch as the prothoracic gland are driven by the central clock, similar tothose in mammals (Myers et al., 2003; Selcho et al., 2017). Meanwhile,an exceptional case has been reported in the fat body, where the cyclingof some genes depends on the local clock whereas that of other genesoccurs in response to central brain signals (Xu et al., 2008; Erion et al.,2016).
1.2. The circadian clock and aging
Some of the mammalian and Drosophila studies suggest that thefunction of the central and peripheral clocks is weakened by the agingprocess. Conversely, minimal or even no impact has been revealed inother studies. Clear evidence has been found to point at a damagednetwork at the central level which is related to aging process, whereascore molecular clock mechanism seems to stay intact based on currentaging research findings. In this part of the review, we focus on how thefunction of the circadian clock changes with advancing age, includingalterations of the circadian signals at the tissue and cellular levels (suchas the neuronal electrical activity), the neuronal network interactions
(such as neuropeptides in the SCN), the intracellular properties (such asmembrane potential and ionic currents), as well as the expression ofclock genes and their protein products.
1.2.1. The impact of aging on the central clockThe first clue that the central clock is involved in aging process
comes from transplantation studies. Implantation of fetal SCN tissue inold animals rescues circadian output function such as daily behavioralrhythms in hamsters (Viswanathan and Davis, 1995) and diurnalrhythms of hormones in rats (Cai et al., 1997). Furthermore animalsthat have restored higher clock function end up with increased long-evity (Hurd and Ralph, 1998). However, it is still unclear whethercircadian decline observed in aged animals is paralleled with age-as-sociated alterations in the structural organization of the SCN. The re-sults of the changes of the SCN in both volume and total cell numbersunder aging in both humans and rodents are mixed (Hood and Amir,2017). For instance, despite that SCN neuronal cells have been found toreduce with increasing age in rats (Tsukahara et al., 2005), there are anumber of studies showing no loss in the total number of cells (Madeiraet al., 1995; Roozendaal et al., 1987) or the volume (Madeira et al.,1995; Tsukahara et al., 2005) of the SCN.
There is good evidence that with age comes impairment of lighttransmission from eye to the SCN, as well as the reduction of respon-siveness of the SCN to light (Asai et al., 2001; Kolker et al., 2003; Lupiet al., 2012; Zhang et al., 1998). Despite this there is no reported age-related change of the RHT (Lupi et al., 2012; Zhang et al., 1998), butextensive evidence points to the decrease of short-wavelength lightpassage through the yellowing and thickened lens with increasing agein humans (Brainard et al., 1997; Kessel et al., 2010; Najjar et al., 2014)and rodents (Zhang et al., 1998). Aside from the decline of retinalfunction observed in humans (Freund et al., 2011; Gerth et al., 2002),the number of ipRGCs has been reported to decrease with advancingage in mice (Semo et al., 2003). Therefore a plausible mechanism ofcircadian clock decline is weaker entrainment by light leading to de-synchrony among oscillators within the SCN. This in turn feeds back onthe circadian clocks throughout the body worsening the systemic effect.
The results of age-associated decline in SCN neuronal activity at thetissue level are consistent, which may account for behavioral andphysiological impairment. The electrical activity patterns of SCN neu-rons measured by multiunit activity (MUA) recordings indicate that theamplitude of SCN activity decreases with increasing age in brain slicepreparations in vitro (Biello, 2009; Nygård et al., 2005; Satinoff et al.,1993; Watanabe et al., 1995) and in vivo (Nakamura et al., 2011). Thedecrease in neuronal electricity rhythms may reflect a loss of coherencein firing patterns across SCN cells in aged animals, ending up with aphase desynchronization. This hypothesis is supported by the findingsof a wider phase distribution and an antiphase activation of cluster ofcells during the night (Farajnia et al., 2012), as well as an elevatednumber of silent cells during the day (Nygård et al., 2005) with ad-vancing age.
One of the possible explanations to the deterioration of the SCNneuronal signals at the tissue level is the age-related degradation of thecentral network system (Nakamura et al., 2016). Correspondingly,rhythms of clock gene expression recorded at single cell level rapidlybecome desynchronized and dissociated and gradually drift out ofphase with each other in associated SCN neurons from aged animals,while those in young SCN cells stay synchronized (Nakamura et al.,2015), indicating a ‘network fault’ due to aging.
The communication among SCN neurons undergoes significant im-pairment in aged animals due to lacking of effective messengers. It hasbeen reported that the expression level and the amplitude of VIP mRNAand the expression level of its receptor mRNA in the SCN decrease inthe course of mammalian aging (Kalló et al., 2004; Kawakami et al.,1997). Such attenuation is associated with both a decline in the level ofmRNA within each cell as well as in the number of VIP-producing cellsin the SCN (Krajnak et al., 1998). This is well supported by human
J. Zhao, et al. Ageing Research Reviews 52 (2019) 64–71
66

research, showing decreased number of VIP neurons in the SCN ofmiddle-aged males (Zhou et al., 1995; Zhou and Swaab, 1999). Thedisruption of VIP expression has functional implications in the aging ofthe circadian clock. The extent of age-related changes of circadianamplitude of behavior rhythms are correlated with the number of VIP-expressing SCN neurons in humans (Wang et al., 2015). In addition, thepresence of VIP-expressing transplanted cells successfully restores cir-cadian output behavior and physiology in aged mice (Li and Satinoff,1998).
Aside from neuropeptides, quantification of synaptic terminals inthe SCN has demonstrated an age-related reduction in synaptic spinesand a shortening of dendrites (Palomba et al., 2008). This marked re-duction in the synaptic signaling of the aged SCN affects equally GA-BAergic terminals and the remaining subset. Furthermore, patch-clamprecordings in hypothalamic slices show that GABAergic postsynapticcurrents decrease in frequency (Nygård et al., 2005) and amplitude(Farajnia et al., 2012) in SCN neurons of aged mice.
It is evident that an age-related loss of neuronal connectivity existsat the central level, but the molecular clock in the SCN as a whole isrelatively resistant to aging (Mattis and Sehgal, 2016). Indeed, anumber of studies have compared the expression of core clock genes inthe aged SCN and have shown conflicting results with either normal orreduced expression of various clock genes. This is aside from a con-sistent age-related decline in expression of Bmal1 and a consistent lackof age-related change in the expression of Per1 (Banks et al., 2016). In arecent study reporting the gene profiling in the SCN of a primate model,core clock gene expression at both mRNA and protein level has beenfound to stay unchanged in aged animals (Eghlidi et al., 2018). In viewof this, causal links between clock gene expression patterns in thecentral clock and age-associated disturbance in circadian rhythmscannot be conclusively demonstrated. For instance, with the samepurpose to investigate the intrinsic age-related changes of the centralclock mechanism, results of Per2/PER2 expression (same clock gene)measured in the SCN under constant conditions in aged mice (samespecies) when compared with young counterparts from different studiesare diverse. Age-related reduction has been found in Per2 mRNAmeasured by in situ hybridization in vitro in free-running aged malemice (approx. 15 months) when compared to young mice (approx. 4months) (Weinert et al., 2001). Likewise, the amplitude of PER2::LUCrhythms in vitro drops markedly in aged male mice (13–15 months)when compared to young mice (3–5 months) (Nakamura et al., 2015).Nonetheless, only minor deficits of PER2 expression measured by bothimmunohistochemistry and bioluminescence in vitro have been shownin free-running aged male mice (13–16 months) when compared toyoung mice (3–6 months) (Nakamura et al., 2011). In a recent studywhere animals with more advancing age are used, Per2 rhythms aremeasured in old male and female Per2Luc mice (24–26 months) in vitro,and the difference is not notable when compared to younger animals(8–13 months) (Polidarová et al., 2016).
The molecular clockwork is also believed to be intact at the singlecellular level in the central clock (Nakamura et al., 2016). Previousattempts to measure the clock genes in individual neurons under agingare contradictory and have shown both preservation of the clock geneprofiles (Asai et al., 2001) and lower amplitude oscillations (Weinertet al., 2001). A recent study using an ultra-sensitive camera systemcapable of recording single cell imaging in PER2::LUC reporter mice,has provided strong evidence on the preservation of the intracellularclock mechanism by showing clear circadian rhythms and similar am-plitude in individual cells in both young and aged SCN (Nakamuraet al., 2015).
Despite maintenance of normal clock gene expression, individualneurons in the SCN have shown functional decline resulting from aging.By using long-term single-cell recording in vivo, the effect of aging onfiring rate patterns of individual SCN neurons has been revealed, dis-playing a decreased amplitude of circadian impulse activity at thecellular level (Aujard et al., 2001). Not only the neuronal activity, but
also the intracellular signaling is modified by aging process. For in-stance, the reduction in the circadian amplitude of certain potassiumcurrents has been observed in aged SCN neurons (Farajnia et al., 2012;Itri et al., 2005, 2010). Another study also reports the loss of the cir-cadian modulation of large-conductance Ca2+-activated K+ (BK)channels (Farajnia et al., 2015). Cellular signaling is important in theregulation of the resting membrane potential and the action potential,thus their age-related changes may lead to diminished rhythm of neu-ronal activity in the SCN cells from aged animals (Farajnia et al., 2012).
Findings in aged Drosophila are similar to those in mammals, in-cluding a controversial result on changes of molecular oscillators at thecentral level and a convincing pattern of decline in intercellular syn-chronization among central clock neurons. On one hand, it has beenreported that PER and TIM oscillations in almost all central clockneuron groups (s-LNv, l-LNv, DN1, DN2 and LNd) deteriorate with ad-vancing age (Umezaki et al., 2012). However, this is challenged by thefinding that PER is robustly expressed in synchrony among differentgroups of clock neurons (s-LNv, DN1 and LNd (Luo et al., 2012) andDN1–3 and LNd (Zhao et al., 2018) in aged flies. On the other hand, theevidence showing reduction of the levels of PDF in the central clocknetwork is consistent (Luo et al., 2012; Umezaki et al., 2012). More-over, overexpression of PDF in PDF-positive neurons increases certainclock gene expression in specific central clock neurons, which leads tothe partial rescue of rhythmicity and period changes of output behaviorin old flies (Umezaki et al., 2012).
These results in both mammals and Drosophila suggest the importantrole of age-associated attenuation of intercellular communicationwithin the central neuronal network. With advancing age, it becomesincreasingly difficult for the rhythms of individual neurons to remainsynchronized with each other and thus the rhythmic output of thenetwork is fragmented. Here we predict a hypothesis that the network isserving as a protective mechanism rather than a pure victim under theprocess of aging. Age-induced dampening of circadian rhythms ofneuronal activity is greater at the cellular level when compared to thewhole SCN in vitro (Farajnia et al., 2012) and in vivo (Nakamura et al.,2011), suggesting a compensatory role of the SCN network capable ofcovering for functional deficiency of individual cells under aging(Farajnia et al., 2014).
1.2.2. The impact of aging on the peripheral clocksOne possible mechanism underlying circadian disruption with in-
creasing age is the attenuation of the SCN’s ability to drive local os-cillators, leading to relative desynchrony between the SCN and oscil-lators in the peripheral tissues (Hood and Amir, 2017). There isevidence suggesting that age-related changes in the SCN can affect thefunction of peripheral oscillators, given that the SCN signals such asneuronal activity decline under aging as discussed above (1.2.1). TheSCN regulates peripheral clocks via pathways including neural controlthrough the autonomic nervous system and humoral control throughcirculation (Mohawk et al., 2012). It has been reported that deficiencyin sympathetic input from the central clock is in part responsible forage-related disturbance of the peripheral oscillators. For instance, aweaker response to noradrenergic stimulation as well as decreasedadrenergic receptor mRNA expression in target peripheral tissues havebeen found in aged animals (Tahara et al., 2017).
Other findings also support the idea that an age-related dampeningof signal from the master clock may be responsible for functional de-cline in the periphery. It has been revealed that the entrainment ofperipheral clocks (the esophagus, lung and thymus gland) in aged miceis slower under a shift of the light-dark (LD) schedule in vitro than theresponse in young counterpart, meanwhile a more rapid reaction hasbeen found in the aged SCN (Sellix et al., 2012). Likewise, phase-re-setting in the SCN appears nearly normal in aged rats, yet re-synchronization of peripheral clocks is disrupted in some (such as theliver) but speeds up in others (such as the arcuate nucleus and pinealgland) (Davidson et al., 2008). This study raises the concern that tissue-
J. Zhao, et al. Ageing Research Reviews 52 (2019) 64–71
67

specific changes of entraining speed cannot be simply explained by theweakening signal from the SCN in aged animals, rather, peripheraloscillators themselves may be affected and directly involved in theaging process.
The age-related deterioration of circadian signal from peripheralclocks has been shown in some tissues. For example, the neuronal ac-tivity rhythms are clearly degraded in one of the peripheral tissues (thesub-paraventricular zone) in the brain (Nakamura et al., 2011). Ac-cordingly, studies focusing on the molecular changes in aged peripheralclocks have indicated disruption in clock gene expression in certaincases. In a study using Per1 bioluminescence reporter in vitro, circadianrhythmicity of the lung in aged rats (24–26 months) has been found tobe disturbed (Yamazaki et al., 2002). By using the same technique, Per2expression in aged mice (24–26 months) has also been shown to declinein the lung in vitro (Novosadová et al., 2018). Moreover, a human studyhas found consistent effects of aging on the circadian pattern of PER1and PER2 expression in post-mortem brain samples (the prefrontalcortex) collected from old (> 60 years) and young (< 40 years) sub-jects, showing a phase advance and an amplitude reduction (Chen et al.,2016).
One important point to note is that a number of other studies hasfound no apparent change of peripheral clocks resulting from the courseof aging. A recent study has revealed that neuronal firing is not affectedby aging at either the single cell level or as the overall pattern in themice motor cortex (McKillop et al., 2018). Besides, the circadian ex-pression profiles of clock gene Per1, Per2, or Cry1 mRNA measured byreverse transcription polymerase chain reaction (RT-PCR) in vitro arequite similar between young and aged rats (22–26 months) in theperipheral tissues (the paraventricular nucleus (PVN) and pineal gland)(Asai et al., 2001). This finding is supported by subsequent studiesshowing almost identical clock genes Per2 and Bmal1 expression byusing similar method between young and aged mice (15 months) in thebrain, heart and liver (Oishi et al., 2011), and normal Per1 and Per2mRNA in aged mice (> 18 months) in the kidney, liver and sub-mandibular gland (Tahara et al., 2017). Further, these three peripheralclocks measured in PER2::LUC aged mice (> 18 months) in vivo exhibitnormal oscillation amplitudes (Tahara et al., 2017). Similarly, withbioluminescence reporters in vitro, Per1 expression in some peripheraltissues (the PVN, pineal gland, arcuate nucleus, cornea, pituitary, liverand kidney) in aged rats (24–26 months) (Yamazaki et al., 2002) andPer2 expression of the pancreas in aged mice (24–26 months)(Novosadová et al., 2018) have been found to be unchanged. In a recenthuman study, the phase of Per3 expression in peripheral hair tissuesfrom very old subjects (83–94 years old) has been shown to be similarwith young controls (Yamaguchi et al., 2018).
It is not difficult to tell from these results that there is no clearevidence pointing at an age-associated peripheral disturbance inmammals. Under aging, changes of neuronal signal as well as rhythmicclock gene expression have been observed in some but not other per-ipheral oscillators. The challenge lies in the fact that a strong tissue-dependent manner of molecular mechanism within individual oscilla-tors would complicate the general interpretation when it comes to thediscussion on peripheral clocks and aging. For instance, in addition tostudies mentioned above (Novosadová et al., 2018; Yamazaki et al.,2002), tissue-dependent changes of CLOCK and BMAL1 expression hasbeen reported in a number of extra-SCN regions in aged mice (Wyse andCoogan, 2010).
One potential solution is to look at age-related peripheral alterna-tions at the cellular level, although current results are inadequate todraw a confirmative conclusion. It has been reported that circadianexpression of clock genes PER2 and BMAL1 in serum-stimulated se-nescent cells is significantly impaired compared with that in young cellsboth in vitro and in vivo (Kunieda et al., 2006). Nevertheless, it is alsosuggested that basic clock properties of peripheral cells do not changeduring aging, by showing the identical period length, amplitude, andphase of Bmal1 expression in aged fibroblast in vitro (Pagani et al.,
2011).In recent years, instead of only focusing on damage of the core
molecular mechanism, a novel theory putting emphasis on the age-as-sociated change of the clock-controlled effector genes is rising. Throughanalyzing the whole transcriptome in aged mice (> 18 months), it hasbeen revealed that epidermal and muscle stem cells retain a robust corecircadian machinery, with comparable amplitude and phase both in LDconditions and the following constant darkness. However, although stillactive and rhythmic, the downstream oscillating transcriptome is ex-tensively reprogrammed, switching from genes involved in homeostasisto those involved in tissue-specific stresses, such as inflammation, DNAdamage and inefficient autophagy (Solanas et al., 2017).
Concerning the peripheral clocks in Drosophila, many studies in-dicate age-related molecular decline. First, reduction in amplitude ofcircadian clock gene expression has been found throughout the body inaged animals (Driver, 2000). Likewise, per and tim luciferase reportershas shown decline in expression of peripheral clocks throughout the flybody with advancing age (Luo et al., 2012; Zhao et al., 2018). Further,per mRNA oscillations (Krishnan et al., 2009) as well as per and tim atboth mRNA and protein levels (Luo et al., 2012; Rakshit andGiebultowicz, 2013) have been reported to decrease in heads of oldflies. It has also been shown in a recent study by using both techniquesof RNA sequencing and qPCR that tim expression declines in aged flies’heads (Kuintzle et al., 2017).
Even though many results in Drosophila studies show moleculardecline at the gross peripheral level, different tissues or body parts seemto respond differently. For example, transcriptional oscillations of perand tim have been proved to decrease in heads but not bodies of agedflies (Rakshit et al., 2012). As one of the peripheral clocks, the retinalphotoreceptor cells in the compound eyes of old flies show decline inexpression of PER, yet that is not the case for another peripheral os-cillator, the Malpighian tubules, where strong PER oscillations havebeen observed in aged animals (Giebultowicz and Long, 2015).
In brief, it is known that tissues/organs age at different rates andhave tissue-specific signatures in both mammals and Drosophila(Girardot et al., 2006), adding the complexity of age-related propertieswithin individual oscillators. Besides, various peripheral tissues/organsmay have different functional implication to the entire circadiansystem. Therefore, there is no unifying theory to summarize the com-prehensive changes of the peripheral clocks due to aging at this stage.
2. Conclusions
In this review we compare the changes of various components of thecircadian clock system at different levels during the aging process. Themain impairment lies in the age-related breakdown of the central net-work, the maintenance of which could serve as a protective role againstaging. The intercellular coupling as well as the intracellular signalinghave been proved to be disrupted at the central pacemaker. Althoughage-associated impairment of the molecular clockwork in central os-cillators is currently unconfirmed, it is evident that the neuronal ac-tivity measured from both single-cell and tissue levels declines withadvancing age. The deterioration of the peripheral oscillators, includingmolecular machinery, cellular activity and downstream clock-con-trolled genes, has been proposed in some aging studies. However, thestudy on the peripheral field is still largely unclear and one possiblereason is that diverse tissues/organs change differently under the cir-cumstance of aging. Presumably, the combination of the damage fromvarious functional units is reflected by the layout at the body level,resulting in disrupted behavioral or physiological output rhythms seenin aged animals.
One main limitation of existing studies investigating how the cir-cadian clock changes with aging is that they have failed to confirm thecausal relationship between the two. On the one hand, it is difficult tofully ascertain that the effects on the circadian clock seen in the agedanimals are simply due to aging. One cannot exclude that other factors,
J. Zhao, et al. Ageing Research Reviews 52 (2019) 64–71
68

such as disease status, may also actively affect the circadian clock. Onthe other hand, it also raises the question of whether the dampening ofthe circadian clock rhythms contributes to the aging process. System-wide knockouts of clock genes creating “accelerated aging” phenotypesin both Drosophila and mice suggest a causal role of the central andperipheral molecular clockwork on aging (Hood and Amir, 2017). Forinstance, null mutation of the per gene (Krishnan et al., 2009) or timgene (Vaccaro et al., 2017) in Drosophila is associated with significantlyshortened lifespan, as well as impaired locomotor rhythm. Likewise,deficiency of the BMAL1 or CLOCK protein significantly affects long-evity and results in the development of age-specific pathologies, such assarcopenia, osteoporosis and cataracts (Kondratov et al., 2006).
Given that not a particular part but instead the diverse componentsof the clock can be affected by aging, rescue strategies should aim atpreservation of the circadian system organization. Caloric restriction(CR) or dietary restriction (DR) seems to be a desirable candidate toreset different levels of the clock system to slow down the deteriorationcaused by aging. To date, CR/DR is the only known intervention cap-able of consistently and reproducibly prolonging lifespan in severalanimal models from yeast to mammals (Cornélissen and Otsuka, 2017).The effects of CR/DR are a coordinated systemic response, throughregulation of both central and peripheral oscillators (Guarente, 2013).At the cellular level, CR/DR has the capacity to prevent the age-asso-ciated gene reprogramming and to restore homeostasis (Sato et al.,2017; Solanas et al., 2017). At the central level, CR/DR is implicated inthe regulation of the central pacemaker to maintain robust circadiancontrol against aging (Chang and Guarente, 2013). At the peripherallevel, CR/DR preserves the cycling of most circadian clock genes whichmediates the lifespan-extending effects (Katewa et al., 2016).
Research on human beings also indicates the protective effect of CR/DR against age-associated metabolic disarrangement, inflammation andoxidative stress, sharing similar mechanisms that have been shown inmammalian studies (Fontana et al., 2010). Although there is no directevidence showing that CR/DR prolongs human lifespan, a promisingprospect comes from a more-than two-decade longitudinal study con-ducted in non-human primate rhesus monkeys (Colman et al., 2009,2014). It suggests that long-term CR/DR significantly delays the onsetof age-related disorders such as diabetes, cardiovascular diseases andcancer, and what’s more, reduces the incidence of age-related and all-cause deaths.
While the link between aging and the circadian clock has attractedgrowing interest there is still much to learn. Future studies are war-ranted to find out answers to questions such as what is the causal re-lationship between the clock and aging, whether individual clock cells/tissues/organs contribute to the systemic decline of the circadian or-ganization, and how the alterations of circadian mechanism contributeto the behavioral and physiological changes seen in aged species. Withthe development of advancing techniques, such as next-generation se-quencing approaches (Takahashi, 2017) and single-cell circadian re-porter technology (Hong et al., 2018; Nakamura et al., 2015), theknowledge will continue to expand towards the broader and deeperfields. If more is known about the circadian system and its interactionwith aging, we will be able to investigate potential strategies withwhich to approach age-associated problems.
Declarations of interest
None.
Acknowledgements
No acknowledgements.
References
Abraham, U., Granada, A.E., Westermark, P.O., Heine, M., Kramer, A., Herzel, H., 2010.
Coupling governs entrainment range of circadian clocks. Mol. Syst. Biol. 6, 438.Agrawal, P., Houl, J.H., Gunawardhana, K.L., Liu, T., Zhou, J., Zoran, M.J., Hardin, P.E.,
2017. Drosophila CRY entrains clocks in body tissues to light and maintains passivemembrane properties in a non-clock body tissue independent of light. Curr. Biol. 27,2431–2441.
Asai, M., Yoshinobu, Y., Kaneko, S., Mori, A., Nikaido, T., Moriya, T., Akiyama, M.,Shibata, S., 2001. Circadian profile of per gene mRNA expression in the su-prachiasmatic nucleus, paraventricular nucleus, and pineal body of aged rats. J.Neurosci. Res. 66, 1133–1139.
Aton, S.J., Colwell, C.S., Harmar, A.J., Waschek, J., Herzog, E.D., 2005. Vasoactive in-testinal polypeptide mediates circadian rhythmicity and synchrony in mammalianclock neurons. Nat. Neurosci. 8, 476–483.
Aujard, F., Herzog, E.D., Block, G.D., 2001. Circadian rhythms in firing rate of individualsuprachiasmatic nucleus neurons from adult and middle-aged mice. Neuroscience106, 255–261.
Balsalobre, A., 2002. Clock genes in mammalian peripheral tissues. Cell. Tissue. Res. 309,193–199.
Banks, G., Nolan, P.M., Peirson, S.N., 2016. Reciprocal interactions between circadianclocks and aging. Mamm. Genome 27, 332–340.
Biello, S.M., 2009. Circadian clock resetting in the mouse changes with age. Age (Dordr.)31, 293–303.
Brainard, G.C., Rollag, M.D., Hanifin, J.P., 1997. Photic regulation of melatonin in hu-mans: ocular and neural signal transduction. J. Biol. Rhythms 12, 537–546.
Brown, S.A., Azzi, A., 2013. Peripheral circadian oscillators in mammals. Handb. Exp.Pharmacol. 45–66.
Cai, A., Scarbrough, K., Hinkle, D.A., Wise, P.M., 1997. Fetal grafts containing su-prachiasmatic nuclei restore the diurnal rhythm of CRH and POMC mRNA in agingrats. Am. J. Physiol. 273, R1764–1770.
Ceriani, M.F., Hogenesch, J.B., Yanovsky, M., Panda, S., Straume, M., Kay, S.A., 2002.Genome-wide expression analysis in Drosophila reveals genes controlling circadianbehavior. J. Neurosci. 22, 9305–9319.
Cermakian, N., Boivinlk, D.B., 2009. The regulation of central and peripheral circadianclocks in humans. Obes. Rev. 10 (Suppl. 2), 25–36.
Chang, H.C., Guarente, L., 2013. SIRT1 mediates central circadian control in the SCN by amechanism that decays with aging. Cell 153, 1448–1460.
Chen, C.Y., Logan, R.W., Ma, T., Lewis, D.A., Tseng, G.C., Sibille, E., McClung, C.A., 2016.Effects of aging on circadian patterns of gene expression in the human prefrontalcortex. Proc. Natl. Acad. Sci. U. S. A. 113, 206–211.
Collins, B., Kaplan, H.S., Cavey, M., Lelito, K.R., Bahle, A.H., Zhu, Z., Macara, A.M.,Roman, G., Shafer, O.T., Blau, J., 2014. Differentially timed extracellular signalssynchronize pacemaker neuron clocks. PLoS Biol. 12, e1001959.
Colman, R.J., Anderson, R.M., Johnson, S.C., Kastman, E.K., Kosmatka, K.J., Beasley,T.M., Allison, D.B., Cruzen, C., Simmons, H.A., Kemnitz, J.W., Weindruch, R., 2009.Caloric restriction delays disease onset and mortality in rhesus monkeys. Science.325, 201–204.
Colman, R.J., Beasley, T.M., Kemnitz, J.W., Johnson, S.C., Weindruch, R., Anderson,R.M., 2014. Caloric restriction reduces age-related and all-cause mortality in rhesusmonkeys. Nat Commun. 5, 3557.
Colwell, C.S., 2011. Linking neural activity and molecular oscillations in the SCN. Nat.Rev. Neurosci. 12, 553–569.
Cornélissen, G., Otsuka, K., 2017. Chronobiology of aging: a mini-review. Gerontology63, 118–128.
Davidson, A.J., Yamazaki, S., Arble, D.M., Menaker, M., Block, G.D., 2008. Resetting ofcentral and peripheral circadian oscillators in aged rats. Neurobiol. Aging 29,471–477.
Dibner, C., Schibler, U., Albrecht, U., 2010. The mammalian circadian timing system:organization and coordination of central and peripheral clocks. Annu. Rev. Physiol.72, 517–549.
Do, M.T., Yau, K.W., 2010. Intrinsically photosensitive retinal ganglion cells. Physiol.Rev. 90, 1547–1581.
Driver, C., 2000. The circadian clock in old Drosophila melanogaster. Biogerontology 1,157–162.
Dubruille, R., Emery, P., 2008. A plastic clock: how circadian rhythms respond to en-vironmental cues in Drosophila. Mol. Neurobiol. 38, 129–145.
Duffy, J.F., Zitting, K.M., Chinoy, E.D., 2015. Aging and circadian rhythms. Sleep. Med.Clin. 10, 423–434.
Eghlidi, D., Luna, S.L., Brown, D., Garyfallou, V., Kohama, S., Urbanski, H.F., 2018. Geneexpression profiling of the SCN in young and old rhesus macaques. J. Mol.Endocrinol. 61, 57–67.
Enoki, R., Kuroda, S., Ono, D., Hasan, M.T., Ueda, T., Honma, S., Honma, K., 2012.Topological specificity and hierarchical network of the circadian calcium rhythm inthe suprachiasmatic nucleus. Proc. Natl. Acad. Sci. U. S. A. 109, 21498–21503.
Enoki, R., Oda, Y., Mieda, M., Ono, D., Honma, S., Honma, K.I., 2017. Synchronous cir-cadian voltage rhythms with asynchronous calcium rhythms in the suprachiasmaticnucleus. Proc. Natl. Acad. Sci. U. S. A. 114, E2476–E2485.
Erion, R., King, A.N., Wu, G., Hogenesch, J.B., Sehgal, A., 2016. Neural clocks and neu-ropeptide F/Y regulate circadian gene expression in a peripheral metabolic tissue.Elife 5 pii, e13552.
Evans, J.A., 2016. Collective timekeeping among cells of the master circadian clock. J.Endocrinol. 230, R27–49.
Farajnia, S., Michel, S., Deboer, T., vanderLeest, H.T., Houben, T., Rohling, J.H.,Ramkisoensing, A., Yasenkov, R., Meijer, J.H., 2012. Evidence for neuronal desyn-chrony in the aged suprachiasmatic nucleus clock. J. Neurosci. 32, 5891–5899.
Farajnia, S., Deboer, T., Rohling, J.H., Meijer, J.H., Michel, S., 2014. Aging of the su-prachiasmatic clock. Neuroscientist 20, 44–55.
Farajnia, S., Meijer, J.H., Michel, S., 2015. Age-related changes in large-conductance
J. Zhao, et al. Ageing Research Reviews 52 (2019) 64–71
69

calcium-activated potassium channels in mammalian circadian clock neurons.Neurobiol. Aging 36, 2176–2183.
Foley, N.C., Tong, T.Y., Foley, D., Lesauter, J., Welsh, D.K., Silver, R., 2011.Characterization of the orderly spatiotemporal patterns of clock gene activation inmammalians suprachiasmatic nucleus. Eur. J. Neurosci. 33, 1851–1865.
Fontana, L., Partridge, L., Longo, V.D., 2010. Extending healthy life span–from yeast tohumans. Science 328, 321–326.
Freund, P.R., Watson, J., Gilmour, G.S., Gaillard, F., Sauvé, Y., 2011. Differential changesin the retina function with normal aging in humans. Doc. Ophthalmol. 122, 177–190.
Froy, O., 2011. Circadian rhythms, aging, and life span in mammals. Physiology(Bethesda) 26, 225–235.
Gerth, C., Garcia, S.M., Ma, L., Keltner, J.L., Werner, J.S., 2002. Multifocal electro-retinogram: age-related changes for different luminance levels. Graefes. Arch. Clin.Exp. Ophthalmol. 240, 202–208.
Giebultowicz, J.M., Hege, D.M., 1997. Circadian clock in malpighian tubules. Nature 386,664.
Giebultowicz, J.M., Long, D.M., 2015. Ageing and circadian rhythms. Curr. Opin. Insect.Sci. 7, 82–86.
Girardot, F., Lasbleiz, C., Monnier, V., Tricoire, H., 2006. Specific age-related signaturesin Drosophila body parts transcriptome. BMC. Genomics 7, 69.
Guarente, L., 2013. Calorie restriction and sirtuins revisited. Genes. Dev. 27, 2072–2085.Hardin, P.E., Panda, S., 2013. Circadian timekeeping and output mechanisms in animals.
Curr. Opin. Neurobiol. 23, 724–731.Harrisingh, M.C., Wu, Y., Lnenicka, G.A., Nitabach, M.N., 2007. Intracellular Ca2+
regulates free-running circadian clock oscillation in vivo. J. Neurosci. 27,12489–12499.
Helfrich-Förster, C., 2005. Neurobiology of the fruit fly’s circadian clock. Genes. Brain.Behav. 4, 65–76.
Herzog, E.D., Aton, S.J., Numano, R., Sakaki, Y., Tei, H., 2004. Temporal precision in themammalian circadian system: a reliable clock from less reliable neurons. J. Biol.Rhythms 19, 35–46.
Herzog, E.D., Hermanstyne, T., Smyllie, N.J., Hastings, M.H., 2017. Regulating the su-prachiasmatic nucleus (SCN) circadian clockwork: interplay between cell-autono-mous and circuit-level mechanisms. Cold. Spring. Harb. Perspect. Biol. 9.
Hong, G., Fu, T.M., Qiao, M., Viveros, R.D., Yang, X., Zhou, T., Lee, J.M., Park, H.G.,Sanes, J.R., Lieber, C.M., 2018. A method for single-neuron chronic recording fromthe retina in awake mice. Science 360, 1447–1451.
Honma, S., 2018. The mammalian circadian system: a hierarchical multi-oscillatorstructure for generating rhythm. J Physiol Sci. 68, 207–219.
Honma, S., Shirakawa, T., Katsuno, Y., Namihira, M., Honma, K., 1998. Circadian periodsof single suprachiasmatic neurons in rats. Neurosci. Lett. 250, 157–160.
Honma, S., Nakamura, W., Shirakawa, T., Honma, K., 2004. Diversity in the circadianperiods of single neurons of the rat suprachiasmatic nucleus depends on nuclearstructure and intrinsic period. Neurosci. Lett. 358, 173–176.
Hood, S., Amir, S., 2017. The aging clock: circadian rhythms and later life. J. Clin. Invest.127, 437–446.
Hurd, M.W., Ralph, M.R., 1998. The significance of circadian organization for longevityin the golden hamster. J. Biol. Rhythms 13, 430–436.
Itri, J.N., Michel, S., Vansteensel, M.J., Meijer, J.H., Colwell, C.S., 2005. Fast delayedrectifier potassium current is required for circadian neural activity. Nat. Neurosci. 8,650–656.
Itri, J.N., Vosko, A.M., Schroeder, A., Dragich, J.M., Michel, S., Colwell, C.S., 2010.Circadian regulation of a-type potassium currents in the suprachiasmatic nucleus. J.Neurophysiol. 103, 632–640.
Izumo, M., Pejchal, M., Schook, A.C., Lange, R.P., Walisser, J.A., Sato, T.R., Wang, X.,Bradfield, C.A., Takahashi, J.S., 2014. Differential effects of light and feeding oncircadian organization of peripheral clocks in a forebrain Bmal1 mutant. Elife 19, 3.
Kalló, I., Kalamatianos, T., Piggins, H.D., Coen, C.W., 2004. Ageing and the diurnal ex-pression of mRNAs for vasoactive intestinal peptide and for the VPAC2 and PAC1receptors in the suprachiasmatic nucleus of male rats. J. Neuroendocrinol. 16,758–766.
Katewa, S.D., Akagi, K., Bose, N., Rakshit, K., Camarella, T., Zheng, X., Hall, D., Davis, S.,Nelson, C.S., Brem, R.B., Ramanathan, A., Sehgal, A., Giebultowicz, J.M., Kapahi, P.,2016. Peripheral circadian clocks mediate dietary restriction-dependent changes inlifespan and fat metabolism in Drosophila. Cell. Metab. 23, 143–154.
Kawakami, F., Okamura, H., Tamada, Y., Maebayashi, Y., Fukui, K., Ibata, Y., 1997. Lossof day-night differences in VIP mRNA levels in the suprachiasmatic nucleus of agedrats. Neurosci. Lett. 222, 99–102.
Kessel, L., Lundeman, J.H., Herbst, K., Andersen, T.V., Larsen, M., 2010. Age-relatedchanges in the transmission properties of the human lens and their relevance tocircadian entrainment. J. Cataract. Refract. Surg. 36, 308–312.
Kingsbury, N.J., Taylor, S.R., Henson, M.A., 2016. Inhibitory and excitatory networksbalance cell coupling in the suprachiasmatic nucleus: a modeling approach. J. Theor.Biol. 397, 135–144.
Koh, K., Evans, J.M., Hendricks, J.C., Sehgal, A., 2006. A Drosophila model for age-as-sociated changes in sleep:wake cycles. Proc. Natl. Acad. Sci. U. S. A. 103,13843–13847.
Kolker, D.E., Fukuyama, H., Huang, D.S., Takahashi, J.S., Horton, T.H., Turek, F.W.,2003. Aging alters circadian and light-induced expression of clock genes in goldenhamsters. J. Biol. Rhythms 18, 159–169.
Kondratov, R.V., Kondratova, A.A., Gorbacheva, V.Y., Vykhovanets, O.V., Antoch, M.P.,2006. Early aging and age-related pathologies in mice deficient in BMAL1, the corecomponent of the circadian clock. Genes. Dev. 20, 1868–1873.
Kondratova, A.A., Kondratov, R.V., 2012. The circadian clock and pathology of the ageingbrain. Nat. Rev. Neurosci. 13, 325–335.
Krajnak, K., Kashon, M.L., Rosewell, K.L., Wise, P.M., 1998. Aging alters the rhythmic
expression of vasoactive intestinal polypeptide mRNA but not arginine vasopressinmRNA in the suprachiasmatic nuclei of female rats. J. Neurosci. 18, 4767–4774.
Krishnan, B., Dryer, S.E., Hardin, P.E., 1999. Circadian rhythms in olfactory responses ofDrosophila melanogaster. Nature 400, 375–378.
Krishnan, B., Levine, J.D., Lynch, M.K., Dowse, H.B., Funes, P., Hall, J.C., Hardin, P.E.,Dryer, S.E., 2001. A new role for cryptochrome in a Drosophila circadian oscillator.Nature 411, 313–317.
Krishnan, N., kretzschmar, D., Rakshit, K., Chow, E., Giebultowicz, J.M., 2009. The cir-cadian clock gene period extends healthspan in aging Drosophila melanogaster.Aging (Albany NY) 1, 937–948.
Kuintzle, R.C., Chow, E.S., Westby, T.N., Gvakharia, B.O., Giebultowicz, J.M., Hendrix,D.A., 2017. Circadian deep sequencing reveals stress-response genes that adopt ro-bust rhythmic expression during aging. Nat. Commun. 8, 14529.
Kunieda, T., Minamino, T., Katsuno, T., Tateno, K., Nishi, J., Miyauchi, H., Orimo, M.,Okada, S., Komuro, I., 2006. Cellular senescence impairs circadian expression ofclock genes in vitro and in vivo. Circ. Res. 98, 532–539.
Lehman, M.N., Silver, R., Gladstone, W.R., Kahn, R.M., Gibson, M., Bittman, E.L., 1987.Circadian rhythmicity restored by neural transplant. Immunocytochemical char-acterization of the graft and its integration with the host brain. J. Neurosci. 7,1626–1638.
Li, H., Satinoff, E., 1998. Fetal tissue containing the suprachiasmatic nucleus restoresmultiple circadian rhythms in old rats. Am. J. Physiol. 275, R1735–1744.
Lin, Y., Stormo, G.D., Taghert, P.H., 2004. The neuropeptide pigment-dispersing factorcoordinates pacemaker interactions in the Drosophila circadian system. J. Neurosci.24, 7951–7957.
Liu, F., Chang, H.C., 2017. Physiological links of circadian clock and biological clock ofaging. Protein Cell 8, 477–488.
Liu, X., Lorenz, L., Yu, Q.N., Hall, J.C., Rosbash, M., 1988. Spatial and temporal ex-pression of the period gene in Drosophila melanogaster. Genes Dev. 2, 228–238.
Liu, C., Weaver, D.R., Strogatz, S.H., Reppert, S.M., 1997. Cellular construction of acircadian clock: period determination in the suprachiasmatic nuclei. Cell 91,855–860.
Liu, A.C., Welsh, D.K., Ko, C.H., Tran, H.G., Zhang, E.E., Priest, A.A., Buhr, E.D., Singer,O., Meeker, K., Verma, I.M., Doyle 3rd, F.J., Takahashi, J.S., Kay, S.A., 2007.Intercellular coupling confers robustness against mutations in the SCN circadianclock network. Cell 129, 605–616.
Luo, W., Chen, W.-F., Yue, Z., Chen, D., Sowcik, M., Sehgal, A., Zheng, X., 2012. Old flieshave a robust central oscillator but weaker behavioral rhythms that can be improvedby genetic and environmental manipulations. Aging Cell 11, 428–438.
Lupi, D., Semo, M., Foster, R.G., 2012. Impact of age and retinal degeneration on the lightinput to circadian brain structures. Neurobiol. Aging 33, 383–392.
Madeira, M.D., Sousa, N., Santer, R.M., Paula-Barbosa, M.M., Gundersen, H.J., 1995. Ageand sex do not affect the volume, cell numbers, or cell size of the suprachiasmaticnucleus of the rat: and unbiased stereological study. J. Comp. Neurol. 361, 585–601.
Mattis, J., Sehgal, A., 2016. Circadian rhythms, sleep, and disorders of aging. TrendsEndocrinol. Metab. 27, 192–203.
Maywood, E.S., Reddy, A.B., Wong, G.K., O’Neill, J.S., O’Brien, J.A., McMahon, D.G.,Harmar, A.J., Okamura, H., Hastings, M.H., 2006. Synchronization and maintenanceof timekeeping in suprachiasmatic circadian clock cells by neuropeptidergic sig-naling. Curr. Biol. 16, 599–605.
McKillop, L.E., Fisher, S.P., Cui, N., Peirson, S.N., Foster, R.G., Wafford, K.A., Vyazovskiy,V.V., 2018. Effects of aging on cortical neural dynamics and local sleep homeostasisin mice. J. Neurosci. 38, 3911–3928.
Mohawk, J.A., Green, C.B., Takahashi, J.S., 2012. Central and peripheral circadian clocksin mammals. Annu. Rev. Neurosci. 35, 445–462.
Myers, E.M., Yu, J., Sehgal, A., 2003. Circadian control of eclosion: interaction between acentral and peripheral clock in Drosophila melanogaster. Curr. Biol. 13, 526–533.
Nahm, S.S., Farnell, Y.Z., Griffith, W., Earnest, D.J., 2005. Circadian regulation andfunction of voltage-dependent calcium channels in the suprachiasmatic nucleus. J.Neurosci. 25, 9304–9308.
Najjar, R.P., Chiquet, C., Teikari, P., Cornut, P.L., Claustrat, B., Denis, P., Cooper, H.M.,Gronfier, C., 2014. Aging of non-visual spectral sensitivity to light in humans: com-pensatory mechanisms? PLoS One 9, e85837.
Nakamura, W., Yamazaki, S., Takasu, N.N., Mishima, K., Block, G.D., 2005. Differentialresponse of period 1 expression within the suprachiasmatic nucleus. J. Neurosci. 25,5481–5487.
Nakamura, T.J., Nakamura, W., Yamazaki, S., Kudo, T., Cutler, T., Colwell, C.S., Block,G.D., 2011. Age-related decline in circadian output. J. Neurosci. 31, 10201–10205.
Nakamura, T.J., Nakamura, W., Tokuda, I.T., Ishikawa, T., Kudo, T., Colwell, C.S., Block,G.D., 2015. Age-related changes in the circadian system unmasked by constantconditions. eNeuro 2.
Nakamura, T.J., Takasu, N.N., Nakamura, W., 2016. The suprachiasmatic nucleus: age-related decline in biological rhythms. J. Physiol. Sci. 66, 367–374.
Nitabach, M.N., Taghert, P.H., 2008. Organization of the Drosophila circadian controlcircuit. Curr. Biol. 18, R84–93.
Nitabach, M.N., Blau, J., Holmes, T.C., 2002. Electrical silencing of Drosophila pacemakerneurons stops the free-running circadian clock. Cell 109, 485–495.
Novosadová, Z., Polidarová, L., Sládek, M., Sumová, A., 2018. Alteration in glucosehomeostasis and persistence of the pancreatic clock in aged mPer2Luc mice. Sci. Rep.8, 11668.
Nygård, M., Hill, R.H., Wikström, M.A., Kristensson, K., 2005. Age-related changes inelectrophysiological properties of the mouse suprachiasmatic nucleus in vitro. BrainRes. Bull. 65, 149–154.
O’Neill, J.S., Reddy, A.B., 2011. Circadian clocks in human red blood cells. Nature 469,498–503.
O’Neill, J.S., Reddy, A.B., 2012. The essential role of cAMP/Ca2+ signalling in
J. Zhao, et al. Ageing Research Reviews 52 (2019) 64–71
70

mammalian circadian timekeeping. Biochem. Soc. Trans. 40, 44–50.Oishi, K., Koyanagi, S., Ohkura, N., 2011. Circadian mRNA expression of coagulation and
fibrinolytic factors is organ-dependently disrupted in aged mice. Exp. Gerontol. 46,994–999.
Pagani, L., Schmitt, K., Meier, F., Izakovic, J., Roemer, K., Viola, A., Cajochen, C., Wirz-Justice, A., Brown, S.A., Eckert, A., 2011. Serum factors in older individuals changecellular clock properties. Proc. Natl. Acad. Sci. U. S. A. 108, 7218–7223.
Palomba, M., Nygård, M., Florenzano, F., Bertini, G., Kristensson, K., Bentivoglio, M.,2008. Decline of the presynaptic network, including GABAergic terminals, in theaging suprachiasmatic nucleus of the mouse. J. Biol. Rhythms 23, 220–231.
Panda, S., Hogenesch, J.B., Kay, S.A., 2002. Circadian rhythms from flies to human.Nature 417, 329–335.
Peng, Y., Stoleru, D., Levine, J.D., Hall, J.C., Rosbash, M., 2003. Drosophila free-runningrhythms require intercellular communication. PLoS Biol. 1, E13.
Plautz, J.D., Kaneko, M., Hall, J.C., Kay, S.A., 1997. Independent photoreceptive circa-dian clocks throughout Drosophila. Science 278, 1632–1635.
Polidarová, L., Sládek, M., Novosadová, Z., Sumová, A., 2016. Aging does not compromisein vitro oscillation of the suprachiasmatic nuclei but makes it more vulnerable toconstant light. Chronobiol. Int. 34, 105–117.
Rakshit, K., Giebultowicz, J.M., 2013. Cryptochrome restores dampened circadianrhythms and promotes healthspan in aging Drosophila. Aging Cell 12, 752–762.
Rakshit, K., Krishnan, N., Guzik, E.M., Pyza, E., Giebultowicz, J.M., 2012. Effects of agingon the molecular circadian oscillations in Drosophila. Chronobiol. Int. 29, 5–14.
Ralph, M.R., Foster, R.G., Davis, F.C., Menaker, M., 1990. Transplanted suprachiasmaticnucleus determines circadian period. Science 247, 975–978.
Roozendaal, B., van Gool, W.A., Swaab, D.F., Hoogendijk, J.E., Mirmiran, M., 1987.Changes in vasopressin cells of the rat suprachiasmatic nucleus with aging. Brain Res.409, 259–264.
Sabado, V., Vienne, L., Nagoshi, E., 2017. Evaluating the autonomy of the Drosophilacircadian clock in dissociated neuronal culture. Front. Cell. Neurosci. 11, 317.
Satinoff, E., Li, H., Tcheng, T.K., Liu, C., McArthur, A.J., Medanic, M., Gillette, M.U.,1993. Do the suprachiasmatic nuclei oscillate in old rats as they do in young ones?Am. J. Physiol. 265, R1216–1222.
Sato, S., Solanas, G., Peixoto, F.O., Bee, L., Symeonidi, A., Schmidt, M.S., Brenner, C.,Masri, S., Benitah, S.A., Sassone-Corsi, P., 2017. Circadian reprogramming in the liveridentifies metabolic pathways of aging. Cell 170, 664–677.
Selcho, M., Millán, C., Palacios-Muñoz, A., Ruf, F., Ubillo, L., Chen, J., Bergmann, G., Ito,C., Silva, V., Wegener, C., Ewer, J., 2017. Central and peripheral clocks are coupledby a neuropeptide pathway in Drosophila. Nat. Commun. 8, 15563.
Sellix, M.T., Evans, J.A., Leise, T.L., Castanon-Cervantes, O., Hill, D.D., DeLisser, P.,Block, G.D., Menaker, M., Davidson, A.J., 2012. Aging differentially affects the re-entrainment response of central and peripheral circadian oscillators. J. Neurosci. 32,16193–16202.
Semo, M., Peirson, S., Lupi, D., Lucas, R.J., Jeffery, G., Foster, R.G., 2003. Melanopsinretinal ganglion cells and the maintenance of circadian and pupillary responses tolight in aged rodless/coneless (rd/rd cl) mice. Eur. J. Neurosci. 17, 1793–1801.
Solanas, G., Peixoto, F.O., Perdiguero, E., Jardí, M., Ruiz-Bonilla, V., Datta, D.,Symeonidi, A., Castellanos, A., Welz, P.S., Caballero, J.M., Sassone-Corsi, P., Muñoz-Cánoves, P., Benitah, S.A., 2017. Aged stem cells reprogram their daily rhythmicfunctions to adapt to stress. Cell 170, 678–692.
Stephan, F.K., Zucker, I., 1972. Circadian rhythms in drinking behavior and locomotoractivity of rats are eliminated by hypothalamic lesions. Proc. Natl. Acad. Sci. U. S. A.69, 1583–1586.
Tahara, Y., Kuroda, H., Saito, K., Nakajima, Y., Kubo, Y., Ohnishi, N., Seo, Y., Otsuka, M.,Fuse, Y., Ohura, Y., Komatsu, T., Moriya, Y., Okada, S., Furutani, N., Hirao, A.,Horikawa, K., Kudo, T., Shibata, S., 2012. In vivo monitoring of peripheral circadianclocks in the mouse. Curr. Biol. 22, 1029–1034.
Tahara, Y., Takatsu, Y., Shiraishi, T., Kikuchi, Y., Yamazaki, M., Motohashi, H., Muto, A.,Sasaki, H., Haraguchi, A., Kuriki, D., Nakamura, T.J., Shibata, S., 2017. Age-relatercircadian disorganization caused by sympathetic dysfunction in peripheral clockregulation. NPJ Aging Mech. Dis. 3, 16030.
Takahashi, J.S., 2017. Transcriptional architecture of the mammalian circadian clock.Nat. Rev. Genet. 18, 164–179.
Tanoue, S., Krishnan, P., Krishnan, B., Dryer, S.E., Hardin, P.E., 2004. Circadian clocks inantennal neurons are necessary and sufficient for olfaction rhythms in Drosophila.Curr. Biol. 14, 638–649.
Top, D., Young, M.W., 2017. Coordination between differentially regulated circadianclocks generates rhythmic behavior. Cold Spring Harb. Perspect. Biol. 10.
Tsukahara, S., Tanaka, S., Ishida, K., Hoshi, N., Kitagawa, H., 2005. Age-related changeand its sex differences in histoarchitecture of the hypothalamic suprachiasmaticnucleus of F344/N rats. Exp. Gerontol. 4, 147–155.
Umezaki, Y., Yoshii, T., Kawaguchi, T., Helfrich-Förster, C., Tomioka, K., 2012. Pigment-dispersing factor is involved in age-dependent rhythm changes in Drosophila mela-nogaster. J. Biol. Rhythms 27, 423–432.
Vaccaro, A., Issa, A.R., Seugnet, L., Birman, S., Klarsfeld, A., 2017. Drosophila clock isrequired in brain pacemaker neurons to prevent premature locomotor aging in-dependently of its circadian function. PLoS Genet. 13, e1006507.
Viswanathan, N., Davis, F.C., 1995. Suprachiasmatic nucleus grafts restore circadianfunction in aged hamsters. Brain Res. 686, 10–16.
Wang, J.L., Lim, A.S., Chiang, W.Y., Hsieh, W.H., Lo, M.T., Schneider, J.A., Buchman,A.S., Bennett, D.A., Hu, K., Saper, C.B., 2015. Suprachiasmatic neuron numbers andrest-activity circadian rhythms in older humans. Ann. Neurol. 78, 317–322.
Watanabe, A., Shibata, S., Watanabe, S., 1995. Circadian rhythm of spontaneous neuronalactivity in the suprachiasmatic nucleus of old hamster in vitro. Brain Res. 695,237–239.
Webb, A.B., Angelo, N., Huettner, J.E., Herzog, E.D., 2009. Intrinsic, nondeterministiccircadian rhythm generation in identified mammalian neurons. Proc. Natl. Acad. Sci.U. S. A. 106, 16493–16498.
Weinert, H., Weinert, D., Schurov, I., Maywood, E.S., Hastings, M.H., 2001. Impairedexpression of the mPer2 circadian clock gene in the suprachiasmatic nuclei of agingmice. Chronobiol. Int. 18, 559–565.
Weiss, R., Bartok, O., Mezan, S., Malka, Y., Kadener, S., 2014. Synergistic interactionsbetween the molecular and neuronal circadian networks drive robust behavioralcircadian rhythms in Drosophila melanogaster. PLoS Genet. 10, e1004252.
Welsh, D.K., Logothetis, D.E., Meister, M., Reppert, S.M., 1995. Individual neurons dis-sociated from rat suprachiasmatic nucleus express independently phased circadianfiring rhythms. Neuron 14, 697–706.
Welsh, D.K., Yoo, S.H., Liu, A.C., Takahashi, J.S., Kay, S.A., 2004. Bioluminescenceimaging of individual fibroblasts reveals persistent, independently phased circadianrhythms of clock gene expression. Curr. Biol. 14, 2289–2295.
Wyse, C.A., Coogan, A.N., 2010. Impact of aging on diurnal expression patterns of CLOCKand BMAL1 in the mouse brain. Brain Res. 1337, 21–31.
Xu, K., Zheng, X., Sehgal, A., 2008. Regulation of feeding and metabolism by neuronaland peripheral clocks in Drosophila. Cell Metab. 8, 289–300.
Yagita, K., Tamanini, F., van Der Horst, G.T., Okamura, H., 2001. Molecular mechanismsof the biological clock in cultured fibroblasts. Science 292, 278–281.
Yamaguchi, S., Isejima, H., Matsuo, T., Okura, R., Yagita, K., Kobayashi, M., Okamura, H.,2003. Synchronization of cellular clocks in the suprachiasmatic nucleus. Science 302,1408–1412.
Yamaguchi, A., Tatsumoto, M., Matsumura, R., Endo, T., Hirata, K., Tokuda, I., Akashi,M., 2018. Normal peripheral circadian phase in the old-old with abnormal circadianbehavior. Genes Cells 23, 849–859.
Yamazaki, S., Numano, R., Abe, M., Hida, A., Takahashi, R., Ueda, M., Block, G.D., Sakaki,Y., Menaker, M., Tei, H., 2000. Resetting central and peripheral circadian oscillatorsin transgenic rats. Science 288, 682–685.
Yamazaki, S., Straume, M., Tei, H., Sakaki, Y., Menaker, M., Block, G.D., 2002. Effects ofaging on central and peripheral mammalian clocks. Proc. Natl. Acad. Sci. U. S. A. 99,10801–10806.
Yoo, S.H., Yamazaki, S., Lowrey, P.L., Shimomura, K., Ko, C.H., Buhr, E.D., Siepka, S.M.,Hong, H.K., Oh, W.J., Yoo, O.J., Menaker, M., Takahashi, J.S., 2004.PERIOD2::LUCIFERASE real-time reporting of circadian dynamics reveals persistentcircadian oscillators in mouse peripheral tissues. Proc. Natl. Acad. Sci. U. S. A. 101,5339–5346.
Yoshii, T., Wulbeck, C., Sehadova, H., Veleri, S., Bichler, D., Stanewsky, R., Helfrich-Förster, C., 2009. The neuropeptide pigment-dispersing factor adjusts period andphase of Drosophila’s clock. J. Neurosci. 29, 2597–2610.
Zhang, Y., Brainard, G.C., Zee, P.C., Pinto, L.H., Takahashi, J.S., Turek, F.W., 1998.Effects of aging on lens transmittance and retinal input to the suprachiasmatic nu-cleus in golden hamsters. Neurosci. Lett. 258, 167–170.
Zhao, J., Warman, G.R., Cheeseman, J.F., 2018. Clock gene expression and locomotoractivity predict death in the last days of life in Drosophila melanogaster. Sci. Rep. 8,11923.
Zhou, J.N., Swaab, D.F., 1999. Activation and degeneration during aging: a morphometricstudy of the human hypothalamus. Microsc. Res. Tech. 44, 36–48.
Zhou, J.N., Hofman, M.A., Swaab, D.F., 1995. VIP neurons in the human SCN in relationto sex, age, and Alzheimer’s disease. Neurobiol. Aging 16, 571–576.
J. Zhao, et al. Ageing Research Reviews 52 (2019) 64–71
71


https://doi.org/10.1177/0748730419836818
JOURNAL OF BIOLOGICAL RHYTHMS, Vol. XX No. X, Month 201X 1 –11DOI: 10.1177/0748730419836818© 2019 The Author(s)Article reuse guidelines: sagepub.com/journals-permissions
1
The circadian clock is the endogenous pace-maker, which generates the daily output rhythms in behavior, physiology, metabolism and cellular pro-cesses, and persists in the absence of environmental cues (Chang, 2006; Peschel and Helfrich-Förster, 2011). Entrainment is the synchronization of the endogenous clock to fluctuating environmental fac-tors, which are called zeitgebers, and light is the most important one (Peschel and Helfrich-Förster, 2011). A functional circadian system requires 3 basic components: 1) the endogenous circadian oscillator, which is temperature compensated, comprising key
clock genes that regulate their own expression; 2) input pathways, which transmit environmental information such as light and entrain the circadian oscillator; and 3) output pathways, which manifest timed behavior and physiology (Yu and Hardin, 2006). In Drosophila, the central clock is defined as a cluster of approximately 150 clock gene-expressing neurons in the brain, which regulates circadian rhythmicity, such as locomotor activity. These cen-tral clock neurons are divided into dorsal lateral neurons (LNd), large ventral lateral neurons (l-LNv), lateral posterior neurons (LPN), small ventral lateral
836818JBRXXX10.1177/0748730419836818Journal of Biological RhythmsZhao et al. / Clock Develops in Embryogenesisresearch-article2019
1. To whom all correspondence should be addressed: James Frederick Cheeseman, Department of Anaesthesiology, School of Medicine, University of Auckland, Support Building 599, Auckland City Hospital, Park Road, Grafton, Auckland, 1142,New Zealand; e-mail: [email protected].
Development of the Molecular Circadian Clock and Its Light Sensitivity in Drosophila Melanogaster
Jia Zhao*, Guy Robert Warman*, Ralf Stanewsky†, and James Frederick Cheeseman*,1
*Department of Anaesthesiology, School of Medicine, University of Auckland, Auckland, 1142 New Zealand,and †Institute for Neuro- and Behavioral Biology, Westfälische Wilhelms University, 48149 Münster, Germany
Abstract The importance of the circadian clock for the control of behavior and physiology is well established but how and when it develops is not fully under-stood. Here the initial expression pattern of the key clock gene period was recorded in Drosophila from embryos in vivo, using transgenic luciferase report-ers. PERIOD expression in the presumptive central-clock dorsal neurons started to oscillate in the embryo in constant darkness. In behavioral experi-ments, a single 12-h light pulse given during the embryonic stage synchronized adult activity rhythms, implying the early development of entrainment mecha-nisms. These findings suggest that the central clock is functional already during embryogenesis. In contrast to central brain expression, PERIOD in the periph-eral cells or their precursors increased during the embryonic stage and peaked during the pupal stage without showing circadian oscillations. Its rhythmic expression only initiated in the adult. We conclude that cyclic expression of PERIOD in the central-clock neurons starts in the embryo, presumably in the dorsal neurons or their precursors. It is not until shortly after eclosion when cyclic and synchronized expression of PERIOD in peripheral tissues com-mences throughout the animal.
Keywords clock gene reporter, period gene, locomotor activity, central oscillator, periph-eral oscillators

2 JOURNAL OF BIOLOGICAL RHYTHMS / Month 201X
neurons (s-LNv), and dorsal neurons 1-3 (DN1-3) (Helfrich-Förster, 2005; Liu et al., 2015; Peschel and Helfrich-Förster, 2011). In addition to the central clock, peripheral clocks reside in various organs and tissues, characterized by the detection of clock gene or protein oscillations throughout the animal (Ito and Tomioka, 2016).
The circadian clock has profound significance in maintaining health throughout life in a range of organisms, including humans. The circadian clock machinery is largely conserved between humans and fruit flies, and Drosophila has been an effective tool for elucidating the cellular and biochemical mechanisms underlying the circadian system (Panda, 2002). The questions of when, where, and how these rhythms take place have attracted growing attention in circa-dian biology. A number of studies have worked on the developmental process of the circadian system in Drosophila (Glossop et al., 2014; Houl et al., 2008; James et al., 1986; Kaneko et al., 1997; Kaneko and Hall, 2000; Liu et al., 2015; Malpel et al., 2002; Malpel et al., 2004; Ruiz et al., 2010; Sehgal et al., 1992), yet the underlying mechanisms have not been fully described. Of particular interest is determining the timing of when the central molecular clock starts to oscillate and when the peripheral clocks are synchro-nized in the whole animal.
Adult activity rhythms are the most detailed and commonly described output of the Drosophila circa-dian clock. Sehgal et al. reported that adult behav-ioral rhythms could be synchronized by a 12-h light pulse administered as early as the first-instar larval stage (but not during embryonic stages), implying that the circadian clock might function continuously from the first larval stage onward (Sehgal et al., 1992). Subsequently, it has been demonstrated that a 10-min light pulse, different light regimes, and ethanol expo-sure, in the larvae can affect the adult free-running period or shift the phase of adult rhythms (Kaneko et al., 2000; Seggio et al., 2012; Sheeba et al., 2002). In summary, this supports the idea that a functional cir-cadian clock is present from the first larval stage.
Driving behavioral rhythms is the “molecular oscillator,” which is based on the transcription-trans-lation feedback loops of several clock genes, includ-ing period and timeless (per and tim) (Hardin and Panda, 2013). The expression of core clock genes and their product proteins, including per/PER and tim/TIM, as well as their regulators, are found in the embryonic central nervous system of Drosophila, sug-gesting that brain oscillator neurons may begin their development during embryogenesis. However, the restricted 24-h duration of the embryonic stage at 25°C (typical environmental rearing temperature) limits our ability to conclude the existence of circa-dian pattern of clock gene expression (Glossop et al.,
2014; Houl et al., 2008; James et al., 1986; Liu et al., 2015; Ruiz et al., 2010).
Immunohistochemistry studies by Kaneko in 1997 demonstrated that the cyclic expression of PER and TIM occurs in the Drosophila larval central nervous system (CNS). This required animals to be sacrificed every 6 h, with data pooled from different brain sam-ples (Kaneko et al., 1997). The question remains whether this technique has sufficient resolution to represent faithfully the key parameters of circadian oscillations. Several studies (Glossop et al., 2014; Houl et al., 2008; James et al., 1986; Liu et al., 2015; Ruiz et al., 2010) that put more emphasis on the spa-tial expression than temporal expression of clock genes, have concentrated on where the circadian clock may start to “tick” instead of when. Using lucif-erase as a real-time reporter of gene expression in vivo makes it possible to monitor clock-gene levels in living individual flies (Stanewsky, 1997). The advan-tages of this technique when compared with other methods include short-interval and long-term mea-surements, as well as the ability to collect time-series data from a single live animal.
Until now, there has been no definite conclusion on whether an entrainable clock exits at the end of embryonic development. The negative correlation between temperature and developmental timing is well defined in Drosophila (Kuntz and Eisen, 2014), offering the possibility to extend the embryonic stage to potentially reveal rhythmicity. A higher tempera-ture speeds up development, whereas a lower tem-perature—within the range of 16°C and 29°C—slows it down; outside of this range leads to an increase in mortality (Economos and Lints, 1986). Thus, 16°C is an optimal choice to prolong maximally the embry-onic stage, and lead to an approximate doubling of the developmental time as compared with 25°C; these results are based on our previous observations (data not shown) and the findings of other studies (e.g., Ghosh et al., 2013). In contrast, circadian clocks are temperature compensated; i.e. the frequency of molecular and behavioral rhythms remains constant at different temperatures (Hall, 1997). Therefore, extending developmental time by lowering the ambi-ent temperature may help to clarify the timing of the emergence of an entrainable molecular clock beyond the 24-h temporal constraint.
In this study, we aimed to determine when the cen-tral and peripheral components of the circadian clock start to oscillate. First, PER expression was measured continuously from the beginning of the embryonic stage in individual flies at 25°C and 16°C using lucif-erase clock-gene reporters. Second, we investigated the adult locomotor activity response to timed light treatment during pre-adult stages. Third, although it is known that the establishment of behavioral

Zhao et al. / CLOCK DEVELOPS IN EMBRYOGENESIS 3
rhythms in individual flies does not require exposure to light during development, we investigated here if this is also true for molecular rhythms of clock gene expression.
MatErials anD MEthoDs
Measurement of Clock Gene Expression
Two transgenic reporter lines 8.0-luc and XLG-luc were used. XLG-luc contains the per promoter sequences plus almost the entire per coding region, whereas 8.0-luc contains the same coding region without the 5’-UTR sequences (Veleri et al., 2003). Flies were reared on growth-media loaded in stan-dard plastic Drosophila vials and kept on a 12-12 h LD cycle with a constant temperature of 25°C and used for real-time bioluminescence recording of per, as described previously (Stanewsky, 2007). Individual eggs were collected within 4 h of being laid in DD and transferred into a single well of a white opaque 96-well microplate (Greiner Bio-one) under 100-luxwhite light. Minimal light was used because eggscannot be distinguished under DD even with the aidof infrared night vision equipment or under red light.This short-term operation falls in the first half of theembryonic stage, when neither the molecular clocknor its light-response has established, according toour and previous findings (Houl et al., 2008). Eachwell contained growth-media fortified with d-lucif-erin, and 15 mM potassium salt; wells without ani-mals were used as controls. Plates were loaded into aMultimode Plate Reader (EnSpire, PerkinElmer) keptinside an incubator (Percival Scientific Inc, USA) andsubjected to DD or 12-12 LD cycles. Bioluminescencelevels emanating from each well were recorded fromthe embryonic stage onward. Experiments were con-ducted for 20 days at 25°C and 10 days at 16°C. Theshorter duration at the lower temperature encom-passed the embryonic and early larval stages, whichwere the primary focus of the study.
Monitoring of locomotor activity
For the locomotion assay, wild-type flies were maintained as above, and eggs were collected and loaded into standard vials all under DD with the aid of infrared night vision equipment. Fruit flies from one group were kept in DD during the entire experi-ment, and other groups were given a 12-h 1000-lux light pulse at different developmental times within the embryonic and early larval stages. When a par-ticular group of flies was under light treatment, the other groups were kept in darkness (as shown in
Figure 3). Fly locomotor activity was monitored auto-matically using Trikinetics Drosophila Activity Monitors (Waltham, MA, USA), as previously described (Zhao et al., 2018). On the days before eclo-sion, with the aid of infrared night vision equipment in DD, individual pupae were transferred into glass tubes (8-cm long and 5-mm diameter) with growth media at one end, sealed with plastic plugs, and plugged with cotton at the other end. Locomotor activity, temperature and relative humidity were recorded every 5 min for at least 7 days in DD.
statistical analysis
Data are shown as mean and standard error (mean [SEM]). Periodogram with Chi-square in ClockLab (Actimetrics, USA) was used to analyze free running periods (tau), phases, and rhythmicity strength of wild-type fly locomotor activity in DD. Lomb-Scargle periodograms in ClockLab and MetaCycle (Wu et al., 2016) were used to analyze periodicity and rhythmic-ity of bioluminescence signals. P values < 0.05 were considered rhythmic.
As the phases of PER expression in individual flies (both 8.0-luc and XLG-luc) raised in DD do not neces-sarily synchronize, rhythms of PER expression were aligned post hoc at the data analysis stage. This was achieved by manually aligning the acrophase of flies across the recording period. To ensure that no larval signal was incorrectly assumed to be an embryonic signal, we chose a cut-off of embryonic expression of 24 h (this is the standard embryonic developmental time at 25°C).
rEsults
Period Expression in the Central Clock during Early Development
To measure PER expression in subsets of the cen-tral-clock neurons, we used 8.0-luc flies, which express a PER-luciferase fusion protein in subsets of the dorsal neuronal clusters (DN1-3) and the LNd (Veleri et al., 2003; Yoshii et al., 2009). Over the 20 days of continuous recording at 25°C, starting with 8.0-luc embryos, we found that 81.5% of 8.0-luc indi-viduals (n = 108) commenced rhythmic PER expres-sion from the embryonic stage onward in constant darkness (DD) (p < 0.05); oscillations in the remaining individuals did not reach significance (Figure 1A, B). The acrophases of rhythmic individuals occurred at different developmental times, with an average of 16.0 ± 0.6 h after egg collection, corresponding to embryogenic stages 16 and 17. Similarly, over the 10

4 JOURNAL OF BIOLOGICAL RHYTHMS / Month 201X
Figure 1. Bioluminescence time-series data of individual 8.0-luc fruit flies measured in DD at 25°C (a, B) and 16°C (C, D). individual bioluminescence (counts per second) is plotted as grey lines and synchronized by acrophases, averaged as black lines. the control is the average signal from wells containing only luciferin-media without any flies under the same conditions, represented as black dashed lines. Corresponding developmental stages are shown at different temperatures. (a) synchronized PEr expression in 8.0-luc (n = 88) during embryonic and early larval stages of the first 72 h after fertilization. the periodogram displays significant circadian rhythm (lomb-scargle, p < 0.05) with a 23.6 h tau, and (B) from the embryo until adult (the first 20 days after fertilization), the periodogram shows cyclic expression, with a mean period of 24.2 ± 0.1 h, as shown in the box-and-whisker plot. (C) synchronized PEr expression in 8.0-luc (n = 31) during the embryonic and early larval stages of the first 96 h after fertilization. the periodogram displays significant circadian rhythm (lomb-scargle, p < 0.05) with a 23.6-hour tau, and (D) from the embryonic to the larval stages (the first 10 days after fertilization), the periodogram shows cyclic expression, with a mean period of 24.1 ± 0.2 h, as shown in the box-and-whisker plot.

Zhao et al. / CLOCK DEVELOPS IN EMBRYOGENESIS 5
days of continuous recording at 16°C, 75.6% of 8.0-luc flies (n = 41) showed rhythmic PER expression onset from the embryonic stage onward in DD (p < 0.05); oscillations in the remaining individuals did not reach significance (Figure 1C, D). The acrophases of rhythmic individuals occurred at 36.7 ± 1.3 h after egg collection, with levels falling in the second half of embryogenesis. At this lower temperature, where embryonic developmental time is doubled, clear peaks of PER expression occurred after 24 h and 48 h, demonstrating that the molecular clock starts operat-ing during embryogenesis (Figure 1C). Because rhythmic PER expression in DD flies was not syn-chronized at either temperature, we manually syn-chronized the signals by aligning the acrophases, grouping them as an average signal to visualize rhythmicity (Figure 1). At 25°C, grouped PER expres-sion during the embryonic (24 h) and early larval (stages 1 and 2) stages was emphasized (Figure 1A), with a free-running period of 23.6 h. At 16°C, grouped PER expression during the embryonic (48 h) and 1st larval (48 h) stages was emphasized (Figure 1C), with a free-running period of 23.6 h.
Period Expression in the Peripheral Clocks during Early Development
After observing these per expression patterns at the central clock level, we decided to further study how PER oscillations develop at the peripheral-clock level. To distinguish the effect from the central clock, a second per transgenic line, XLG-luc was used, which is most likely expressed in many, if not all, per-expressing cells throughout the adult body, including the central and peripheral clock tissues (Stanewsky et al., 1997; Veleri et al., 2003). However, it mainly reports per signals in peripheral clocks because of the small number of central brain clock neurons com-pared with the thousands of clock cells in the rest of the body.
Again, because rhythmic PER expression in flies reared in DD was not synchronized, we manually synchronized the signals by aligning their acrophases at 25°C. During the continuous 20-day and 10-day XLG-luc recordings at 25°C or 16°C, respectively, we found that PER started to rise during the embryonic stage. All of the embryos kept in DD (n = 68) and in LD (n = 67) at 25°C showed this increase in PER sig-nal (Figure 2A, B). The rise in PER peaked at the pupa and then decreased. Phase differences between DD and LD records most likely were caused by manual phase sorting of the DD records to aid recognition of rhythmic PER expression (see Methods for details). Importantly, no oscillations were evident during this first rise in PER. Subsequently, PER began to rise again after eclosion and, at this time, also began to
cycle in DD with a period of 24 h and in LD with a period of 23.5 h. The strength of rhythmicity of the bioluminescence oscillations was more robust in LD compared with DD, as expected. At 16°C, PER expres-sion in XLG-luc (n = 14 in DD and n = 82 in LD) begins to rise from the embryonic stage until the lar-val stage, without oscillations (Figure 2C, D).
synchronized Behavioral rhythms in adult Flies Exposed to light Early in Development
The results from the above experiments indicated the existence of a functional clock in the embryo. Thus, we attempted to confirm this by testing whether light given at such early developmental stages influ-enced the phase of adult locomotor activity. Groups of wild-type flies kept in DD were given light treat-ment at different times of development. We found that the locomotor activity of individual flies raised in DD was not synchronized at either 25°C or 16°C. The probability that the distribution of individual phases within a population is random was calculated by Rayleigh’s test. Probabilities for flies reared in DD being synchronized were not significant, indicating a random distribution (p > 0.05). Flies that received light-treatment at the first half of the embryonic stage showed desynchrony at both temperatures. In con-trast, those that had received light treatment at the second half of the embryonic stage showed synchro-nized distributions of phases (Figure 3). Even though the group of flies kept in DD was never exposed to light during their early development, they showed rhythmic locomotor activity as adults, similar to all the light-treated groups (Table 1).
DisCussion
In this study, we analyzed the development of the circadian clock in Drosophila by utilizing trans-genic reporters that record the essential clock com-ponent PER in vivo. These tools allowed us to monitor the expression of clock genes in real time and to track the measurement throughout the life of the flies. Here, we have shown that the expression of PER starts in the embryo but does not begin to cycle until the adult stage. However, a reporter that reflects PER expression only in subsets of central clock neurons starts to show rhythmic circadian patterns during embryogenesis, coinciding with the time when light begins to affect the clock. In addi-tion, and in agreement with previous studies, clock gene oscillations of PER as well as adult locomotor activity rhythms (Sehgal et al., 1992) initiate with-out the requirement of light.

6 JOURNAL OF BIOLOGICAL RHYTHMS / Month 201X
Figure 2. Bioluminescence time-series data of individual XLG-luc fruit flies measured at 25°C (a, B) and at 16°C (C, D). individual bioluminescence CPs is plotted as grey lines and averaged as black lines. the control is the average signal from wells containing only luciferin media, without any flies under the same condition, noted as black dashed lines. the y-axis is rescaled to better show the first signal increase in the embryo and the pattern of rhythmicity in the adult. Corresponding developmental stages are shown at different temperatures. (a) synchronized PEr expression in XLG-luc in DD (n = 68) begins to rise from the embryo until it reaches a peak at the pupa. the signal decreases in the pupal stage but rises again and begins cycling with a period of 24.0 h (lomb-scargle, p < 0.05) from the adult stage. (B) synchronized PEr expression in XLG-luc in lD (n = 67) shows a similar pattern to that in DD, with oscillations begin-ning in the adults with a period of 23.5 h (lomb-scargle, p < 0.05). no oscillation is detected in the increasing signal from the embryonic and larval stages in DD or lD. PEr expression at 16°C in XLG-luc in (C) DD and (D) lD (n = 14 and n = 82, respectively) begins to rise from the embryonic stage until the larval stage without oscillations (lomb-scargle, p > 0.05). Periodogram in lD is calculated as the average signal minus the control signal to counteract the pseudo-rhythm caused by the light-induced background (stanewsky, 2007).

Zhao et al. / CLOCK DEVELOPS IN EMBRYOGENESIS 7
Figure 3. the circular distributions of the locomotor activity phases (acrophases), average actograms and periodograms at (a) 25°C (left panel) and (B) 16°C (right panel). one group per panel was under DD throughout the entire experiment (top). the12-h light pulses were administrated to other groups at different stages of early development. Each clock face shows a 24-h clock time. Black dots around clock faces indicate the timing of acrophase of every single fly in the same group. the mean phase of each population is shown by the angle of the radius with 95% confidence intervals. Double-plotted actograms were averaged from flies in each group. (a) the animals kept in DD for the entire experiment (n = 28) and given light pulses at 0 to 12 h (embryo, n = 34) show random distributions (p = 0.48 and p = 0.35, respectively), with arrhythmic average actograms supported by periodograms. animals given light pulses at 12 to 24 h (embryo, n = 38), and at 24 to 36 h (larva, n = 35) show significantly concentrated distributions (p < 0.001), with rhythmic average actograms supported by periodograms (Chi-square, p < 0.05). (B) animals kept in DD for the entire experiment (n = 43) show random distributions (p = 0.08). animals given light pulses at 0 to 12 h (embryo, n = 48) and at 12 to 24 h (embryo, n = 45) show random distributions (p = 0.82 and p = 0.67, respectively), with arrhythmic average actograms supported by periodograms. animals given light pulses at 24 to 36 h (embryo, n = 42), 36 to 48 h (embryo, n = 54), or 48 to 72 h (larva, n = 47) all show significantly concentrated distributions (p = 0.003, p = 0.01 and p = 0.03, respectively), with rhythmic average actograms supported by periodograms (Chi-square, p < 0.05).
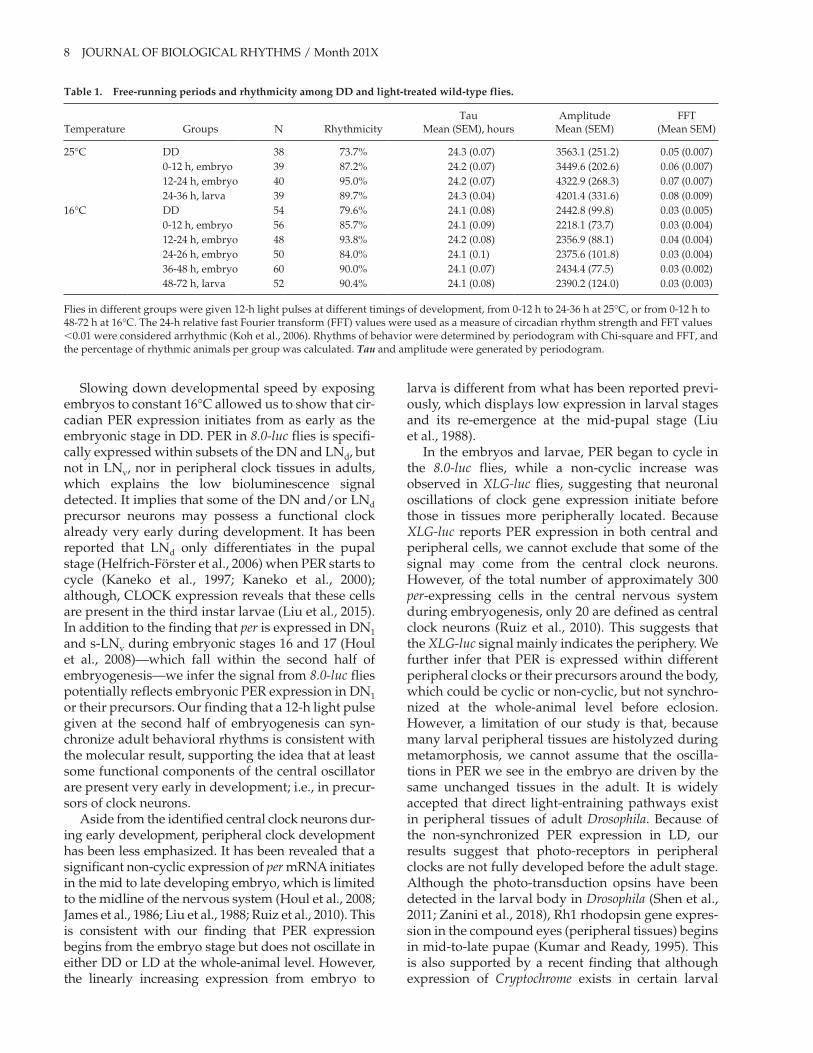
8 JOURNAL OF BIOLOGICAL RHYTHMS / Month 201X
Slowing down developmental speed by exposing embryos to constant 16°C allowed us to show that cir-cadian PER expression initiates from as early as the embryonic stage in DD. PER in 8.0-luc flies is specifi-cally expressed within subsets of the DN and LNd, but not in LNv, nor in peripheral clock tissues in adults, which explains the low bioluminescence signal detected. It implies that some of the DN and/or LNd precursor neurons may possess a functional clock already very early during development. It has been reported that LNd only differentiates in the pupal stage (Helfrich-Förster et al., 2006) when PER starts to cycle (Kaneko et al., 1997; Kaneko et al., 2000); although, CLOCK expression reveals that these cells are present in the third instar larvae (Liu et al., 2015). In addition to the finding that per is expressed in DN1 and s-LNv during embryonic stages 16 and 17 (Houl et al., 2008)—which fall within the second half of embryogenesis—we infer the signal from 8.0-luc flies potentially reflects embryonic PER expression in DN1 or their precursors. Our finding that a 12-h light pulse given at the second half of embryogenesis can syn-chronize adult behavioral rhythms is consistent with the molecular result, supporting the idea that at least some functional components of the central oscillator are present very early in development; i.e., in precur-sors of clock neurons.
Aside from the identified central clock neurons dur-ing early development, peripheral clock development has been less emphasized. It has been revealed that a significant non-cyclic expression of per mRNA initiates in the mid to late developing embryo, which is limited to the midline of the nervous system (Houl et al., 2008; James et al., 1986; Liu et al., 1988; Ruiz et al., 2010). This is consistent with our finding that PER expression begins from the embryo stage but does not oscillate in either DD or LD at the whole-animal level. However, the linearly increasing expression from embryo to
larva is different from what has been reported previ-ously, which displays low expression in larval stages and its re-emergence at the mid-pupal stage (Liu et al., 1988).
In the embryos and larvae, PER began to cycle in the 8.0-luc flies, while a non-cyclic increase was observed in XLG-luc flies, suggesting that neuronal oscillations of clock gene expression initiate before those in tissues more peripherally located. Because XLG-luc reports PER expression in both central and peripheral cells, we cannot exclude that some of the signal may come from the central clock neurons. However, of the total number of approximately 300 per-expressing cells in the central nervous system during embryogenesis, only 20 are defined as central clock neurons (Ruiz et al., 2010). This suggests that the XLG-luc signal mainly indicates the periphery. We further infer that PER is expressed within different peripheral clocks or their precursors around the body, which could be cyclic or non-cyclic, but not synchro-nized at the whole-animal level before eclosion. However, a limitation of our study is that, because many larval peripheral tissues are histolyzed during metamorphosis, we cannot assume that the oscilla-tions in PER we see in the embryo are driven by the same unchanged tissues in the adult. It is widely accepted that direct light-entraining pathways exist in peripheral tissues of adult Drosophila. Because of the non-synchronized PER expression in LD, our results suggest that photo-receptors in peripheral clocks are not fully developed before the adult stage. Although the photo-transduction opsins have been detected in the larval body in Drosophila (Shen et al., 2011; Zanini et al., 2018), Rh1 rhodopsin gene expres-sion in the compound eyes (peripheral tissues) begins in mid-to-late pupae (Kumar and Ready, 1995). This is also supported by a recent finding that although expression of Cryptochrome exists in certain larval
table 1. Free-running periods and rhythmicity among DD and light-treated wild-type flies.
Temperature Groups N RhythmicityTau
Mean (SEM), hoursAmplitude
Mean (SEM)FFT
(Mean SEM)
25°C DD 38 73.7% 24.3 (0.07) 3563.1 (251.2) 0.05 (0.007)0-12 h, embryo 39 87.2% 24.2 (0.07) 3449.6 (202.6) 0.06 (0.007)12-24 h, embryo 40 95.0% 24.2 (0.07) 4322.9 (268.3) 0.07 (0.007)24-36 h, larva 39 89.7% 24.3 (0.04) 4201.4 (331.6) 0.08 (0.009)
16°C DD 54 79.6% 24.1 (0.08) 2442.8 (99.8) 0.03 (0.005)0-12 h, embryo 56 85.7% 24.1 (0.09) 2218.1 (73.7) 0.03 (0.004)12-24 h, embryo 48 93.8% 24.2 (0.08) 2356.9 (88.1) 0.04 (0.004)24-26 h, embryo 50 84.0% 24.1 (0.1) 2375.6 (101.8) 0.03 (0.004)36-48 h, embryo 60 90.0% 24.1 (0.07) 2434.4 (77.5) 0.03 (0.002)48-72 h, larva 52 90.4% 24.1 (0.08) 2390.2 (124.0) 0.03 (0.003)
Flies in different groups were given 12-h light pulses at different timings of development, from 0-12 h to 24-36 h at 25°C, or from 0-12 h to 48-72 h at 16°C. The 24-h relative fast Fourier transform (FFT) values were used as a measure of circadian rhythm strength and FFT values <0.01 were considered arrhythmic (Koh et al., 2006). Rhythms of behavior were determined by periodogram with Chi-square and FFT, and the percentage of rhythmic animals per group was calculated. Tau and amplitude were generated by periodogram.

Zhao et al. / CLOCK DEVELOPS IN EMBRYOGENESIS 9
peripheral tissues but not others, it lacks the function to mediate light-entrainment in both the central and peripheral clocks in adult Drosophila (Agrawal et al., 2017). This is different from the vertebrate system, where peripheral cells in zebrafish are synchronized by light at early embryonic stages, whereas in the adult peripheral clocks of both Drosophila and zebraf-ish, they are directly light-sensitive (Carr et al., 2005).
One of the important features of the circadian clock is that it is endogenous. The current develop-mental study shows that, in certain species, such as Drosophila, the circadian clock is able to develop and mature without the requirement of entraining stim-uli, such as light (Sehgal et al., 1992). In our study light exposure did not initiate or speed up the devel-opment of the circadian clock or change the clock-gene expression level, but it synchronized the phases among individual flies. This is consistent with mam-malian clocks that can start synchronized oscilla-tions within an individual independently of zeitgebers, and independently of a functional mater-nal circadian clock (Jud and Albrecht, 2006). It also shares similarity with clocks in zebrafish, which can initiate but not synchronize without light (Dekens and Whitmore, 2008).
A single light pulse given during the embryonic stage in the current study, or during the larval stage in previous studies (Kaneko et al., 2000; Sehgal et al., 1992), can synchronize adult rhythmic behavior. One possible explanation is a “time-memory,” which implies the special ability of the clock to memorize such stimuli and manifest it later in life when the clock becomes more mature (Kaneko et al., 2000). The direction and magnitude of phase shifts of locomotor activity were different between wild-type and fast-clock pershort mutants, suggesting the involve-ment of per in the regulation of developmental “time-memory” (Kaneko et al., 2000). However, a reasonable assumption is that, at some point during early adult stages, synchronization between different clock cells is under the control of an as yet undescribed intrinsic mechanism. So in our behavioral experiment, rather than a time-memory, it is possible that the clock is “set” by the initial light and free runs in the central clock neurons (as we observe in 8.0-luc in DD) but not in most peripheral clock tissues.
An important role of the circadian clock in the regulation of development has been reported (Kumar et al., 2006; Kyriacou et al., 1990; Vallone et al., 2007). In this study, we placed emphasis on the development of the circadian clock itself. It is assumed that the development of the circadian clock should be in accordance with sequential and precise developmental processes. Consequently, the circadian clock is thought to develop as the whole body develops, in a gradual way (Vallone et al.,
2007). Through the new application of established luciferase clock gene reporter techniques, we show cyclic clock-gene expression, presumably in precur-sors of dorsal clock neurons during the embryonic stages, which is earlier than previously thought. Light is not required to start the clock, but it can, to some extent, synchronize the clock as early as in the embryo, which is also earlier than previously thought. In total, the early cyclic expression of PER and light-entrainable behavior in this study sug-gests that key components of the circadian clock are present very early in life. Future studies may put emphasis on developmental details in individual pacemaker neurons, perhaps by performing simul-taneous recordings of behavioral rhythms and clock-gene expression in the same animal.
authors’ ContriButions
J.Z., G.R.W., and J.F.C. designed research; J.Z. performedresearch; J.Z. and J.F.C. analyzed data; and J.Z., G.R.W.,R.S., and J.F.C. wrote the paper.
ConFliCt oF intErEst statEMEnt
The author(s) have no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
orCiD iD
James Frederick Cheeseman https://orcid.org/0000 -0001-5590-1808
rEFErEnCEs
Agrawal P, Houl JH, Gunawardhana KL, Liu T, Zhou J, Zoran MJ, and Hardin PE (2017) Drosophila CRY entrains clocks in body tissues to light and maintains passive membrane properties in a non-clock body tis-sue independent of light. Curr Biol 27:2431-2441.e3.
Carr AJ, Tamai TK, and Whitmore D (2005) Zebrafish cir-cadian clocks: cells that see light. Biochem Soc Trans 33:962.
Chang DC (2006) Neural circuits underlying circadian behaviour in Drosophila melanogaster. Behav Processes 71:211-225.
Dekens MP and Whitmore D (2008) Autonomous onset of the circadian clock in the zebrafish embryo. EMBO J 27:2757-2765.

10 JOURNAL OF BIOLOGICAL RHYTHMS / Month 201X
Economos AC and Lints FA (1986) Developmental tem-perature and life span in Drosophila melanogaster. I. Constant developmental temperature: evidence for physiological adaptation in a wide temperature range. Gerontology 32:18-27.
Ghosh SM, Testa ND, and Shingleton AW (2013) Temperature-size rule is mediated by thermal plastic-ity of critical size in Drosophila melanogaster. Proc Biol Sci 280:20130174.
Glossop NRJ, Gummadova JO, Ghangrekar I, Hardin PE, and Coutts GA (2014) Effects of TWIN-OF-EYELESS on clock gene expression and central-pacemaker neuron development in Drosophila. J Biol Rhythms 29:151-166.
Hall JC (1997) Circadian pacemakers blowing hot and cold–but they’re clocks, not thermometers. Cell 90:9-12.
Hardin PE and Panda S (2013) Circadian timekeeping and output mechanisms in animals. Curr Opin Neurobiol 23:724-731.
Helfrich-Förster C (2005) Neurobiology of the fruit fly’s cir-cadian clock. Genes Brain Behav 4:65-76.
Helfrich-Förster C, Shafer OT, Wülbeck C, Grieshaber E, Rieger D, and Taghert P (2006) Development and mor-phology of the clock-gene-expressing lateral neurons of Drosophila melanogaster. J Comp Neurol 500:47-70.
Houl JH, Ng F, Taylor P, and Hardin PE (2008) CLOCK expression identifies developing circadian oscilla-tor neurons in the brains of Drosophila embryos. BMC Neurosci 9:119.
Ito C and Tomioka K (2016) Heterogeneity of the peripheral circadian systems in Drosophila melanogaster: A review. Front Physiol 7:8.
James AA, Ewer J, Reddy P, Hall JC, and Rosbash M (1986) Embryonic expression of the period clock gene in the central nervous system of Drosophila melanogaster. EMBO J 5:2313-2320.
Jud C and Albrecht U (2006) Circadian rhythms in murine pups develop in absence of a functional maternal circa-dian clock. J Biol Rhythms 21:149-154.
Kaneko M and Hall JC (2000) Neuroanatomy of cells expressing clock genes in Drosophila: transgenic manipulation of the period and timeless genes to mark the perikarya of circadian pacemaker neurons and their projections. J Comp Neurol 422:66-94.
Kaneko M, Hamblen MJ, and Hall JC (2000) Involvement of the period gene in developmental time-memory: effect of the perShort mutation on phase shifts induced by light pulses delivered to Drosophila larvae. J Biol Rhythms 15:13-30.
Kaneko M, Helfrich-Förster C, and Hall JC (1997) Spatial and temporal expression of the period and timeless genes in the developing nervous system of Drosophila: newly identified pacemaker candidates and novel features of clock gene product cycling. J Neurosci 17: 6745-6760.
Koh K, Evans JM, Hendricks JC, and Sehgal A (2006) A Drosophila model for age-associated changes in
sleep:wake cycles. Proc Natl Acad Sci U S A. 103:13843-13847.
Kumar JP and Ready DF (1995) Rhodopsin plays an essen-tial structural role in Drosophila photoreceptor develop-ment. Development 121:4359-4370.
Kumar S, Vaze KM, Kumar D, and Sharma VK (2006) Selection for early and late adult emergence alters the rate of pre-adult development in Drosophila melanogas-ter. BMC Dev Biol 6:57.
Kuntz SG and Eisen MB (2014) Drosophila embryogenesis scales uniformly across temperature in developmen-tally diverse species. PLoS Genet 10:e1004293.
Kyriacou CP, Oldroyd M, Wood J, Sharp M, and Hill M (1990) Clock mutations alter developmental timing in Drosophila. Heredity 64:395-401.
Liu T, Mahesh G, Houl JH, and Hardin PE (2015) Circadian activators are expressed days before they initiate clock function in late pacemaker neurons from Drosophila. J Neurosci 35:8662-8671.
Liu X, Lorenz L, Yu QN, Hall JC, and Rosbash M (1988) Spatial and temporal expression of the period gene in Drosophila melanogaster. Genes Dev 2:228-238.
Malpel S, Klarsfeld A, and Rouyer F (2002) Larval optic nerve and adult extra-retinal photoreceptors sequen-tially associate with clock neurons during Drosophila brain development. Development 129:1443-1453.
Malpel S, Klarsfeld A, and Rouyer F (2004) Circadian syn-chronization and rhythmicity in larval photopercep-tion-defective mutants of Drosophila. J Biol Rhythms 19:10-21.
Panda S, Hogenesch JB, and Kay SA (2002) Circadian rhythms from flies to human. Nature 417:329-335.
Peschel N and Helfrich-Förster C (2011) Setting the clock–by nature: circadian rhythm in the fruitfly Drosophila melanogaster. FEBS Lett 585:1435-1442.
Ruiz S, Rickert C, Berger C, Technau GM, and Cantera R (2010) Spatio-temporal pattern of cells expressing the clock genes period and timeless and the lineages of period expressing neurons in the embryonic CNS of Drosophila melanogaster. Gene Expr Patterns 10: 274-282.
Seggio JA, Possidente B, and Ahmad ST (2012) Larval ethanol exposure alters adult circadian free-running locomotor activity rhythm in Drosophila melanogaster. Chronobiol Int 29:75-81.
Sehgal A, Price J, and Young MW (1992) Ontogeny of a bio-logical clock in Drosophila melanogaster. Proc Natl Acad Sci U S A 89:1423-1427.
Sheeba V, Chandrashekaran MK, Joshi A, and Sharma VK (2002) Developmental plasticity of the locomotor activ-ity rhythm of Drosophila melanogaster. J Insect Physiol 48:25-32.
Shen WL, Kwon Y, Adegbola AA, Luo J, Chess A, and Montell C (2011) Function of rhodopsin in tem-perature discrimination in Drosophila. Science 331: 1333-1336.

Zhao et al. / CLOCK DEVELOPS IN EMBRYOGENESIS 11
Stanewsky R (2007) Analysis of rhythmic gene expression in adult Drosophila using the firefly luciferase reporter gene. Methods Mol Biol 362:131-142.
Stanewsky R, Jamison CF, Plautz JD, Kay SA, and Hall JC (1997) Multiple circadian-regulated elements contrib-ute to cycling period gene expression in Drosophila. EMBO J 16:5006-5018.
Vallone D, Lahiri K, Dickmeis T, and Foulkes NS (2007) Start the clock! Circadian rhythms and development. Dev Dyn 236:142-155.
Veleri S, Brandes C, Helfrich-Förster C, Hall JC, and Stanewsky R (2003) A self-sustaining, light-entrainable circadian oscillator in the Drosophila brain. Curr Biol 13:1758-1767.
Wu G, Anafi RC, Hughes ME, Kornacker K, and Hogenesch JB (2016) MetaCycle: an integrated R package to
evaluate periodicity in large scale data. Bioinformatics 32:3351-3353.
Yoshii T, Wulbeck C, Sehadova H, Veleri S, Bichler D, Stanewsky R, and Helfrich-Förster C (2009) The neuro-peptide pigment-dispersing factor adjusts period and phase of Drosophila’s clock. J Neurosci 29:2597-2610.
Yu W and Hardin PE (2006) Circadian oscillators of Drosophila and mammals. J Cell Sci 119:4793-4795.
Zanini D, Giraldo D, Warren B, Katana R, Andrés M, Reddy S, Pauls S, Schwedhelm-Domeyer N, Geurten BRH, and Göpfert MC (2018) Proprioceptive opsin functions in Drosophila larval locomotion. Neuron 98:67-74.e4.
Zhao J, Warman GR, and Cheeseman JF (2018) Clock gene expression and locomotor activity predict death in the last days of life in Drosophila melanogaster. Sci Rep 8:11923.

1Scientific RepoRTs | (2018) 8:11923 | DOI:10.1038/s41598-018-30323-x
www.nature.com/scientificreports
Clock gene expression and locomotor activity predict death in the last days of life in Drosophila melanogasterJia Zhao , Guy Robert Warman & James Frederick Cheeseman
The importance of the circadian clock for the regulation of behaviour and physiology, and the molecular control of these rhythms by a set of clock genes are well defined. The circadian clock deteriorates with advancing age but the mechanism underlying is unclear. Here we recorded the expression of two key clock genes in young, middle-aged and old Drosophila using transgenic luciferase lines reporting period and timeless in vivo. We report a novel marker of imminent death in the expression of TIMELESS. In the days immediately preceding death TIMELESS expression increased to at least 150% of previous acrophase values (88.0% of n = 217) and lost circadian rhythmicity, which predicted death equally well in flies of different ages and under light and temperature cycles. We suggest this transient aberrant clock-gene expression is central to the mechanism of the disturbance in circadian behaviour before death (82.7% of n = 342). We also find that PERIOD expression in central-clock neurons remained robust with age, however PERIOD and TIMELESS in peripheral clocks showed a reduction in both expression level and rhythmicity. In conclusion, as flies age the molecular clock gradually declines at the peripheral level but continues to function at the central until days before death.
Aging is the intrinsic and inevitable process of functional deterioration which ultimately leads to death1. The cir-cadian clock is the endogenous pacemaker that generates the daily rhythms among most organisms. Th s clock is affected by time, showing progressive disturbance with advancing age2. Th s effect of aging on the circadian clock is highly conserved among humans and other animals3. In humans, the changes in circadian output signals associated with aging include the reduction of the amplitude and earlier timing of circadian rhythms, both evident in body temperature and sleep-wake cycles4–6. Mammalian studies on aging demonstrate shifts in the phase of rhythms and weakening of rhythmicity reported in body temperature and locomotor activity patterns2,7. Similar to humans and mammals, Drosophila show a lengthened free-running period and decline in the overall rest-activity rhythm strength and sleep consolidation with increasing age8,9. In addition, Drosophila provide a robust and powerful way to investigate clock changes across life time because of their short lifespan (50–80 days)10.
It is interesting that clock-controlled behavioural changes before death have also been observed. For instance, aging flies in their last days of life have been found to exhibit arrhythmia11. And in laboratory mammals, impair-ment of several parameters of circadian rhythms, such as body temperature and locomotor activity, is considered a marker for imminent death12. Several mammalian studies have shown a marked decrease in the amount and amplitude of activity as well as a loss of ability to maintain entrainment under light-dark (LD) cycles as the ani-mals approach death. Some animals lose their circadian rhythm completely13–15. With general acceptance that decline of the circadian clock is a reliable manifestation of aging, it is not hard to imagine that collapse of the clock could be a potential indicator of death. Nonetheless, there has been very little investigation into the mechanism of this phenomenon prior to death.
As well as the decrease in circadian output rhythms16, there is some evidence for age-related changes in circa-dian organization at other levels. The circadian system is composed of multiple circadian oscillators, controlled by a central pacemaker in the suprachiasmatic nucleus (SCN) in mammals17 or a cluster of approximate 150 clock-gene expressing neurons in the brain in Drosophila (dorsal lateral neurons (LNd), large ventral lateral
Department of Anaesthesiology, School of Medicine, the University of Auckland, Park Road, Grafton, Auckland, 1023, New Zealand. Correspondence and requests for materials should be addressed to J.F.C. (email: [email protected])
Received: 16 February 2018
Accepted: 26 July 2018
Published: xx xx xxxx
OPEN

www.nature.com/scientificreports/
2Scientific RepoRTs | (2018) 8:11923 | DOI:10.1038/s41598-018-30323-x
neurons (l-LNv), lateral posterior neurons (LPN), small ventral lateral neurons (s-LNv), and dorsal neurons 1–3 (DN1–3)18). There are oscillators in other regions of the brain and peripheral tissues, referred to as peripheral clocks19. Currently, whether aging is associated with defects in the central clock, or weakening of the synchroni-zation of peripheral clocks, or both, remains to be seen.
The molecular oscillators within both the central clock and peripheral clocks are driven by the transcription-translation feedback loops of several clock genes, including period and timeless (per and tim) in Drosophila20–22. How the mechanism of the molecular clock and its gene properties change during aging is less well known and the existing evidence is confli ting10,23. It has been reported that in the central clock neurons per is robustly expressed in aging flies11, but this fi ding has been challenged by a study showing per reduction in cen-tral clock with age9. Studies in aging mammals also showed controversial results with either normal or reduced expression of various clock genes2,6,7.
In this study we aim to investigate how the molecular clock, in the brain and in the peripheral tissues, changes intrinsically with increasing age and before death under constant conditions. We also examine whether strong entrainment cues improve age-related functional decline of the clock. By using transgenic luciferase lines, we are able to measure real-time expression of clock genes products PER and TIM in Drosophila in three age cohorts (10-, 30- and 50-day-old) for an extended period of time in vivo. We use both 8.0-luc and XLG-luc lines to meas-ure PER expression. 8.0-luc specifi ally refl cts PER expression in subsets of the central-clock dorsal neuronal clusters (DN1–3) and LNd
22,24. XLG-luc refl cts PER expression in all known per-expressing cells throughout the body, including central and peripheral tissues22. XLG-luc is mainly indicative of peripheral clocks because of the small number of central clock neurons compared to thousands of clock cells in the rest of the body. A similar rule applies to tim-luc, which therefore refl cts TIM expression in peripheral clocks.
ResultsAge-related changes of PER in central and peripheral clocks and TIM in peripheral clocks. We analysed the amplitude and acrophase of bioluminescence signal of each day to show the rhythmicity and level of clock-gene expression, respectively. In constant darkness (DD) the rhythmic PER expression at central level in 8.0-luc middle-aged and old flies was comparable to that in the young flies with robust oscillations (Fig. 1A), showing no decrease in both rhythmicity and expression level. In contrast, both amplitude and acrophase of PER at peripheral level in XLG-luc (Fig. 1B) and TIM in tim-luc (Fig. 1C) middle-aged and old flies were signifi antly lower than corresponding young ones, showing age-dependent decline in both rhythmicity and expression level. Despite that the consumption of luciferin (substrate) led to the decay of signal within each age group, by compar-ing the signal from different age groups at the same time point, the effect of aging on clock-gene expression was still evident.
In order to test the influence of entrainment cues on this age-related effect, flies were kept under regular LD cycles with constant temperature and LD coupled with temperature cycles respectively. In LD and LD/tempera-ture cycles the rhythmic PER expressions at central level in 8.0-luc middle-aged and old flies were comparable to that in the young flies with robust oscillations (Figs 2 and 3A). Similar to that shown in DD, both amplitude and acrophase of PER expression at peripheral level in XLG-luc (Fig. 2B) and TIM expression at peripheral level in tim-luc (Fig. 2C) middle-aged and old flies were significantly lower than corresponding young ones in LD. Both amplitude and acrophase of PER expression in XLG-luc (Fig. 3B) and TIM expression in tim-luc (Fig. 3C) old flie were signifi antly lower than corresponding middle-aged and young ones in LD/temperature cycles.
Molecular marker of imminent death in both PER and TIM in peripheral clocks. We found an unexpected increase of TIM signal in tim-luc flies (Fig. 4) and PER signal in XLG-luc (Fig. 5) at the peripheral level which lasted for several days before eventually vanishing and the animals died. The increase of signal was defi ed as at least 50% higher than the acrophase of the preceding cycle. Prior to death, 87.8% (n = 82) tim-luc flies reared in DD showed an increase in signal over a period of 86.2 ± 2.2 hours, 93.5% (n = 62) in LD cycles showed 98.4 ± 3.4 hours, and 83.6% (n = 73) in LD/temperature cycles showed 90.0 ± 3.0 hours. The age of death of these flies ranged from 20 to 60 days. Prior to death, 34.2% (n = 79) XLG-luc flies reared in DD showed an increase in signal over a period of 84.3 ± 5.2 hours, 54.3% (n = 46) in LD cycles showed 81.4 ± 3.8 hours, and 56.3% (n = 48) in LD/temperature cycles showed 81.3 ± 4.6 hours. The age of death of these flies ranged from 20 to 50 days.
In order to confirm that the signal increase occurred before death, we tested how long the bioluminescence signal would last after death under the same condition in a number of fl es. Decapitated tim-luc fl es showed a 11.4 ± 1.6 hour after-death signal (n = 40) and XLG-luc flies showed 7.8 ± 1.0 hours (n = 41). In summary, imme-diately preceding death most tim-luc flies (88.0%, n = 217) demonstrated an increase in TIM signal that lasted for 91.2 ± 1.7 hours on average independent of age or environment. At its height this elevated TIM signal was 3.8 times as high as the previous acrophase on average (Fig. 6A), and not concentrated at any particular Zeitgeber time (ZT) calculated by Rayleigh Test (Fig. 6B). In contrast this increase in TIM was not refl cted in PER as not as many flies in XLG-luc (45.7%, n = 173) showed an increase of 82.3 ± 2.6 hours on average before death. At its height this elevated PER signal was 2.4 times as high as the previous acrophase on average (Fig. 6C), and concen-trated with mean phase around ZT18 to ZT20, similar to previous cycles (Fig. 6D).
Downstream behavioural changes of imminent death. Locomotor activity changed in the last few days before death, in accordance to the aberrant PER and TIM expression during a similar period of time. 82.7% of well entrained flies (n = 342) at different ages lost the circadian pattern of behavioural rhythms and became arrhythmic over a period of 3.7 ± 0.1 days before death (Fig. 7). Arrhythmic locomotor activity was observed in 82.9% in flies which started to be entrained and recorded from 10-day-old (n = 146), 85.1% in flie from 30-day-old (n = 148), and 75.0% in flies from 50-day-old (n = 48), showing no significant difference by

www.nature.com/scientificreports/
3Scientific RepoRTs | (2018) 8:11923 | DOI:10.1038/s41598-018-30323-x
Chi-square test (p = 0.27). The age of death of these flies ranged from 20 to 70 days. 24-hour relative fast Fourier transform (FFT) values was used as a measure of circadian rhythm strength and FFT value <0.01 was considered arrhythmic. Flies showed a FFT value of 0.050 ± 0.002 under LD entrainment and 0.039 ± 0.002 under following free-running in DD, indicating the existence of circadian rhythms of locomotor activity. FFT value dropped to 0.007 ± 0.0003 (p < 0.001) in the fi al days preceding death, showing behavioural arrhythmicity.
DiscussionIn our investigation of the effect of aging on the endogenous clock we found a reduction in both amplitude and acrophase of PER and TIM expression rhythms throughout the body in Drosophila with advancing age (see Fig. 1). We suggest this reduction in clock gene production and expression rhythmicity indicates the decline of circadian clock function. It has been reported that this reduction in expression as well as rhythmicity is very likely to be the main cause for the changes in the circadian pattern of activity in aging flie 25. Th s theory is further supported by gene manipulation studies in which over-expression of clock gene in specific clock neurons can partially rescue behavioural rhythms and shortened free-running periods in old flie 9,10.
There are two possible reasons for this result in overall reduction of clock-gene expression with age, a weak-ened signal from the central clock or the disruption of the peripheral oscillators. We found that robust molecular oscillations of PER persist in the dorsal neurons (DN1–3 and LNd) of the central clocks. Th s is consistent with a previous fi ding of robust PER cycling in s-LNv, DN1 and LNd
11, but different to Umezaki’s 2012 study in which the amplitude of PER and TIM expression deteriorated with age in all central-clock cell groups (s-LNv, l-LNv, DN1, DN2 and LNd)9. Our fi ding supports the idea that a functional central oscillator is still intact in aged Drosophila. We infer that the effect of age is on the peripheral level rather than the central level. We showed reduction in PER and TIM expression without the total loss of rhythmicity in peripheral clocks throughout the fly
Figure 1. Mean bioluminescence time-series data from young (10-day), middle-aged (30-day) and old (50-day) flies in DD. Bioluminescence counts per second (CPS) is plotted as average of raw counts at 30-minute intervals. Data are shown as mean ± SEM with error bars at each time point. 10-day cohort equals black lines and columns, 30-day cohort equals mid-grey lines and columns, and 50-day cohort equals light-grey lines and columns. (A)8.0-luc (reporting PER in the dorsal neurons, central clock). The 30-day cohort shows higher PER amplitudeand acrophase than 10-day and 50-day (p < 0.001 in all), and there is no difference between 10-day and 50-day (p = 1.00 in both). (B) XLG-luc (reporting PER in the whole animal, peripheral clocks). The 30-day and 50-daycohort show lower PER amplitude and acrophase compared to 10-day (p < 0.001 in all), 50-day is lower than 30-day (p = 0.02 in amplitude and p = 0.001 in acrophase). (C) tim-luc (reporting TIM in the whole animal, peripheralclocks). The 30-day and 50-day cohort show lower TIM amplitude and acrophase compared to 10-day (p < 0.001 in all), and 50-day is lower than 30-day in acrophase (p < 0.001) but not in amplitude (p = 0.70).
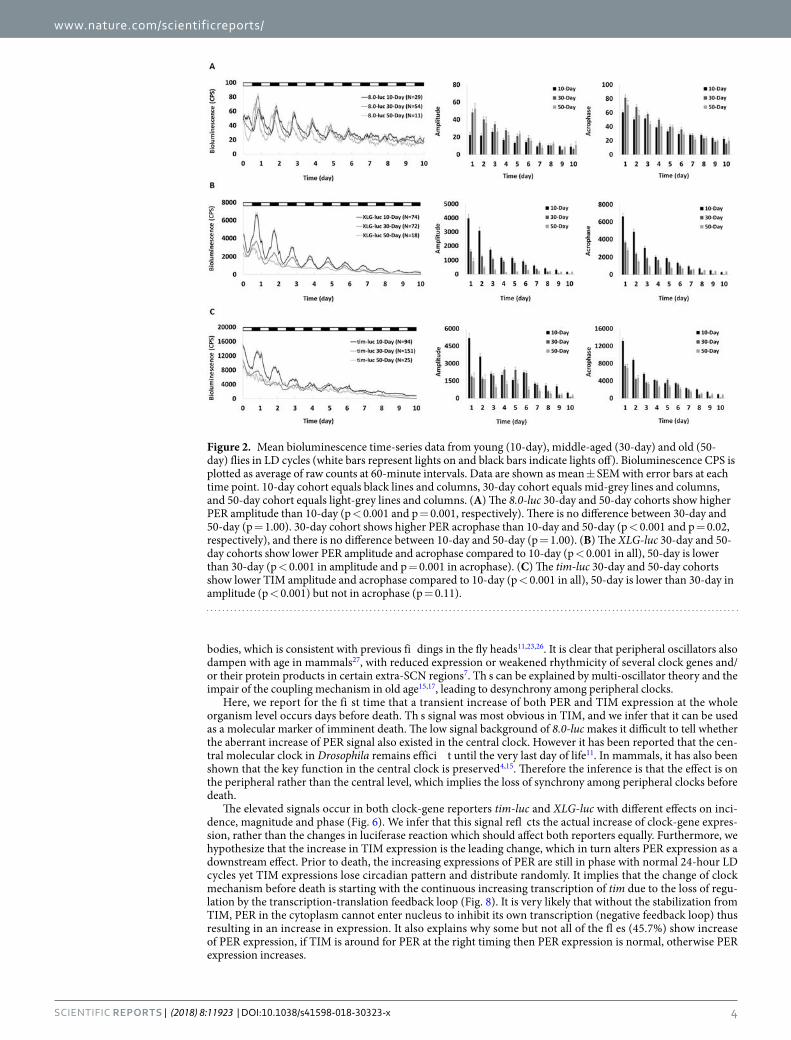
www.nature.com/scientificreports/
4Scientific RepoRTs | (2018) 8:11923 | DOI:10.1038/s41598-018-30323-x
bodies, which is consistent with previous fi dings in the fly heads11,23,26. It is clear that peripheral oscillators also dampen with age in mammals27, with reduced expression or weakened rhythmicity of several clock genes and/or their protein products in certain extra-SCN regions7. Th s can be explained by multi-oscillator theory and the impair of the coupling mechanism in old age15,17, leading to desynchrony among peripheral clocks.
Here, we report for the fi st time that a transient increase of both PER and TIM expression at the whole organism level occurs days before death. Th s signal was most obvious in TIM, and we infer that it can be used as a molecular marker of imminent death. The low signal background of 8.0-luc makes it difficult to tell whether the aberrant increase of PER signal also existed in the central clock. However it has been reported that the cen-tral molecular clock in Drosophila remains effici t until the very last day of life11. In mammals, it has also been shown that the key function in the central clock is preserved4,15. Therefore the inference is that the effect is on the peripheral rather than the central level, which implies the loss of synchrony among peripheral clocks before death.
The elevated signals occur in both clock-gene reporters tim-luc and XLG-luc with different effects on inci-dence, magnitude and phase (Fig. 6). We infer that this signal refl cts the actual increase of clock-gene expres-sion, rather than the changes in luciferase reaction which should affect both reporters equally. Furthermore, we hypothesize that the increase in TIM expression is the leading change, which in turn alters PER expression as a downstream effect. Prior to death, the increasing expressions of PER are still in phase with normal 24-hour LD cycles yet TIM expressions lose circadian pattern and distribute randomly. It implies that the change of clock mechanism before death is starting with the continuous increasing transcription of tim due to the loss of regu-lation by the transcription-translation feedback loop (Fig. 8). It is very likely that without the stabilization from TIM, PER in the cytoplasm cannot enter nucleus to inhibit its own transcription (negative feedback loop) thus resulting in an increase in expression. It also explains why some but not all of the fl es (45.7%) show increase of PER expression, if TIM is around for PER at the right timing then PER expression is normal, otherwise PER expression increases.
Figure 2. Mean bioluminescence time-series data from young (10-day), middle-aged (30-day) and old (50-day) flies in LD cycles (white bars represent lights on and black bars indicate lights off). Bioluminescence CPS is plotted as average of raw counts at 60-minute intervals. Data are shown as mean ± SEM with error bars at each time point. 10-day cohort equals black lines and columns, 30-day cohort equals mid-grey lines and columns, and 50-day cohort equals light-grey lines and columns. (A) The 8.0-luc 30-day and 50-day cohorts show higher PER amplitude than 10-day (p < 0.001 and p = 0.001, respectively). There is no difference between 30-day and 50-day (p = 1.00). 30-day cohort shows higher PER acrophase than 10-day and 50-day (p < 0.001 and p = 0.02,respectively), and there is no difference between 10-day and 50-day (p = 1.00). (B) The XLG-luc 30-day and 50-day cohorts show lower PER amplitude and acrophase compared to 10-day (p < 0.001 in all), 50-day is lowerthan 30-day (p < 0.001 in amplitude and p = 0.001 in acrophase). (C) The tim-luc 30-day and 50-day cohortsshow lower TIM amplitude and acrophase compared to 10-day (p < 0.001 in all), 50-day is lower than 30-day inamplitude (p < 0.001) but not in acrophase (p = 0.11).
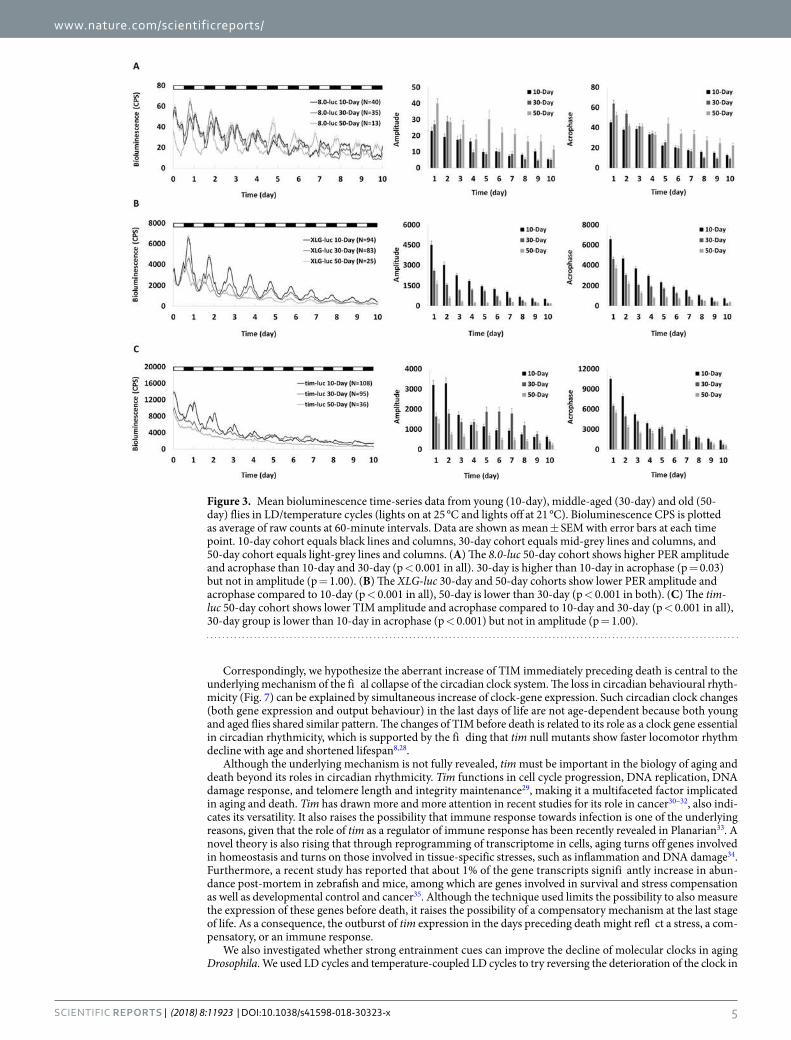
www.nature.com/scientificreports/
5Scientific RepoRTs | (2018) 8:11923 | DOI:10.1038/s41598-018-30323-x
Correspondingly, we hypothesize the aberrant increase of TIM immediately preceding death is central to the underlying mechanism of the fi al collapse of the circadian clock system. The loss in circadian behavioural rhyth-micity (Fig. 7) can be explained by simultaneous increase of clock-gene expression. Such circadian clock changes (both gene expression and output behaviour) in the last days of life are not age-dependent because both young and aged flies shared similar pattern. The changes of TIM before death is related to its role as a clock gene essential in circadian rhythmicity, which is supported by the fi ding that tim null mutants show faster locomotor rhythm decline with age and shortened lifespan8,28.
Although the underlying mechanism is not fully revealed, tim must be important in the biology of aging and death beyond its roles in circadian rhythmicity. Tim functions in cell cycle progression, DNA replication, DNA damage response, and telomere length and integrity maintenance29, making it a multifaceted factor implicated in aging and death. Tim has drawn more and more attention in recent studies for its role in cancer30–32, also indi-cates its versatility. It also raises the possibility that immune response towards infection is one of the underlying reasons, given that the role of tim as a regulator of immune response has been recently revealed in Planarian33. A novel theory is also rising that through reprogramming of transcriptome in cells, aging turns off genes involved in homeostasis and turns on those involved in tissue-specific stresses, such as inflammation and DNA damage34. Furthermore, a recent study has reported that about 1% of the gene transcripts signifi antly increase in abun-dance post-mortem in zebrafish and mice, among which are genes involved in survival and stress compensation as well as developmental control and cancer35. Although the technique used limits the possibility to also measure the expression of these genes before death, it raises the possibility of a compensatory mechanism at the last stage of life. As a consequence, the outburst of tim expression in the days preceding death might refl ct a stress, a com-pensatory, or an immune response.
We also investigated whether strong entrainment cues can improve the decline of molecular clocks in aging Drosophila. We used LD cycles and temperature-coupled LD cycles to try reversing the deterioration of the clock in
Figure 3. Mean bioluminescence time-series data from young (10-day), middle-aged (30-day) and old (50-day) flies in LD/temperature cycles (lights on at 25 °C and lights off at 21 °C). Bioluminescence CPS is plotted as average of raw counts at 60-minute intervals. Data are shown as mean ± SEM with error bars at each time point. 10-day cohort equals black lines and columns, 30-day cohort equals mid-grey lines and columns, and 50-day cohort equals light-grey lines and columns. (A) The 8.0-luc 50-day cohort shows higher PER amplitudeand acrophase than 10-day and 30-day (p < 0.001 in all). 30-day is higher than 10-day in acrophase (p = 0.03)but not in amplitude (p = 1.00). (B) The XLG-luc 30-day and 50-day cohorts show lower PER amplitude andacrophase compared to 10-day (p < 0.001 in all), 50-day is lower than 30-day (p < 0.001 in both). (C) The tim-luc 50-day cohort shows lower TIM amplitude and acrophase compared to 10-day and 30-day (p < 0.001 in all),30-day group is lower than 10-day in acrophase (p < 0.001) but not in amplitude (p = 1.00).

www.nature.com/scientificreports/
6Scientific RepoRTs | (2018) 8:11923 | DOI:10.1038/s41598-018-30323-x
Figure 4. Representative bioluminescence time-series data of tim-luc flies showing aberrant TIM signal at the last days of life. X-axis indicates the age of each fly. Bioluminescence CPS is plotted as raw counts (grey lines) and as a 6-hour moving average (black lines). (A) A 10-day-old and (B) a 30-day-old fly in DD display a rhythmic but descending TIM signal until several days before their deaths at 22 days and 42 days, increases of TIM lasted for 87 hours and 78 hours are observed, respectively. (C) A 10-day-old and (D) a 30-day-old fly in LD cycles show increases of TIM lasted for 91 hours and 75 hours before their deaths at 27 days and 44 days, respectively. (E) A 10-day-old and (F) a 30-day-old fly in LD/temperature cycles show increases of TIM lasted for 91 hours and 77 hours before their deaths at 38 days and 48 days, respectively.
Figure 5. Representative bioluminescence time-series data of XLG-luc flies showing aberrant PER signal at the last days of life. X-axis indicates the age of each fly. Bioluminescence CPS is plotted as raw counts (grey lines) and as a 6-hour moving average (black lines). (A) A 10-day-old and (B) a 30-day-old fly in DD display a rhythmic but descending PER signal until several days before their deaths at 21 days and 42 days, increases of PER lasted for 67 hours and 74 hours are observed, respectively. (C) A 10-day-old and (D) a 30-day-old fly in LD cycles show increases of PER lasted for 64 hours and 98 hours before their deaths at 28 days and 41 days, respectively. (E) A 10-day-old and (F) a 30-day-old fly in LD/temperature cycles show increases of PER lasted for 56 hours and 106 hours before their deaths at 31 days and 48 days, respectively.

www.nature.com/scientificreports/
7Scientific RepoRTs | (2018) 8:11923 | DOI:10.1038/s41598-018-30323-x
peripheral oscillators. The impairment of the input pathway may be a possible explanation and is consistent with the finding that in these older flies the clock cannot be effectively entrained by light or temperature. A previous study has reported that LD cycles alone is not sufficient to rescue disturbance of locomotor activity in aging flies, but the use of LD (12:12) coupling temperature cycles (25 °C/21 °C) turned out to be effective11. Our fi ding suggests a clock-independent temperature effect may be involved. Lower temperatures (such as 21 °C) prolong longevity in Drosophila and slow down the age-associated sleep decline, likely through non-circadian mechanisms8,36.
Circadian coordination has a pronounced impact on physiological functions, overall health, and disease sus-ceptibility37, it is reasonable to infer how lifespan and health would suffer when the circadian system is challenged. Therefore robust and entrained circadian rhythms could potentially lead to better health and increased longevity7. However aiming at entraining the central clock alone is not effici t enough as we fi d that the peripheral clocks are more seriously affected by aging. A number of studies have shown that regular routines of mealtime and exer-cise improve circadian consolidation in elderly individuals, addition with light routines, these timing cues might help substitute for the loss of internal desynchrony5. We also report a remarkable disruption of the circadian clock during the last days of life at both the molecular (Fig. 6) and behavioural levels (Fig. 7 and diagrammatic illustra-tion Fig. 8). Our fi ding indicates not only the underlying cause of the circadian clock disruption before death, but also the exciting potential of tim to be a marker of imminent death, a link for further mechanism exploration and even a basis for intervention.
MethodsFly stock. Drosophila transgenic reporter lines 8.0-luc, XLG-luc and tim-luc, and wild type fl es were reared on growth-media (Carolina, USA) loaded in standard plastic vials. Flies were kept at 12:12 LD cycles and at temper-ature 25 °C, and transferred to fresh vials once a week. Flies were collected within 2 days of eclosion and survival of mated male flies in vials were monitored until their death, showing average longevity of 50–70 days. Young (10-day-old), middle-aged (30-day-old) and old (50-day-old) mated male flies were used. All flies of different ages were kept under identical circumstances.
Measurement of clock-gene expression. Transgenic flies for real-time bioluminescence recording of clock gene per and tim expression were used in this study, as described previously38. Individual flies were transferred into a single well of a white opaque 96-well microplates (Greiner Bio-one). Plates were loaded into
Figure 6. Mean bioluminescence time-series data from the last ten days of life in flies showing elevated signals before death in LD and LD/temperature cycles. X-axis indicates the days before death. Black solid line is the average signal from the last five-days of life, and grey solid line is from the last six to ten days of life with an exponential trendline shown as grey dash line. (A) tim-luc flies (n = 119) show circadian rhythms of TIM expression from day 10 to day 6 before death with a descending trend. The aberrant signal from the last five days (average height value shown in black column) breaks the circadian pattern and shows a remarkable increase (p < 0.001, compared to the average acrophase from the preceding cycle shown in grey column). (C) XLG-luc flie (n = 52) show circadian rhythms of PER expression from day 10 to day 6 before death with a descending trend. The aberrant signal from the last five days follows the circadian pattern and shows a remarkable increase (p < 0.001). The circular distributions of the elevated signals before death in a 24-hour clock face. Black solid dots around clock face indicate individual flies in LD cycles and hollow dots in LD/temperature cycles. ZT (where ZT0 is lights on and ZT12 is lights off) is labelled. (B) tim-luc flies show random distribution in LD (n = 58, p = 0.37) and in LD/temperature (n = 61, p = 0.44). (D) XLG-luc flies show signifi ant concentrated distribution with mean phase pointing at ZT18 in LD (n = 25, p < 0.001) and at ZT20 in LD/temperature (n = 27, p = 0.007).

www.nature.com/scientificreports/
8Scientific RepoRTs | (2018) 8:11923 | DOI:10.1038/s41598-018-30323-x
a Multimode Plate Reader (EnSpire, PerkinElmer) kept inside an incubator (Percival Scientific Inc, USA). Experiments were conducted in DD, LD cycles (12:12) at constant temperature (25 °C), and LD cycles (12:12) coupled with temperature cycles (light on at 25 °C and light off at 21 °C).
Figure 7. Locomotor activity of representative wild type flies showing arrhythmic behaviour before death. Actogram during a period of LD 12:12 followed by a free running rhythms in DD (the white area marks the light phase, the grey shaded area marks the dark phases). Left y-axis indicates the age of each fly and right y-axis indicates the days before death. Dagger is used as label of death. Well-entrained flies of different ages (A) died at 66-day-old, showing a 2-day arrhythmic activity with a FFT value of 0.0001 before death (0.049 in LD and 0.011in DD), (B) died at 56-day-old, showing a 4-day arrhythmic activity with a FFT value of 0.008 before death(0.021 in LD and 0.017 in DD), and (C) died at 31-day-old, showing a 3-day arrhythmic activity with a FFTvalue of 0.0006 before death (0.046 in LD and 0.099 in DD).

www.nature.com/scientificreports/
9Scientific RepoRTs | (2018) 8:11923 | DOI:10.1038/s41598-018-30323-x
Monitoring of locomotor activity. Locomotor activity of wild type flies was monitored automatically using Trikinetics Drosophila Activity Monitors (Waltham, MA, USA), as previously described39. Individual flies with known age (10-day-old, 30-day-old and 50-day-old) were transferred into glass tubes (8 cm long and 5 mm diameter) with grow media at one end, sealed with plastic plugs, and plugged with cotton at the other end. Locomotor activity, temperature and relative humidity were recorded every five minutes for an entraining period of 13 days in LD and then in DD until most flies died.
Statistical analysis. Data are shown in mean and standard error (Mean ± SEM). Statistical signifi ance of the amplitude and acrophase of time series data among young, middle-aged and old transgenic flies were determined using two-way ANOVA with Bonferonni’s post hoc test23. ClockLab (Actimetrics, USA) was used for the analysis of periodicity and rhythmicity in locomotor activity of wild type flies. Periodicity was tested by Chi-square periodogram and rhythm strength was measured by FFT values8.
References1. Fonseca Costa, S. S. & Ripperger, J. A. Impact of the circadian clock on the aging process. Front Neurol. 6, 43 (2015).2. Nakamura, T. J. et al. Age-related changes in the circadian system unmasked by constant conditions (1, 2, 3). eNeuro. 2 (2015).3. Kondratova, A. A. & Kondratov, R. V. The circadian clock and pathology of the ageing brain. Nat Rev Neurosci. 13, 325–335 (2012).4. Weinert, D. & Waterhouse, J. The circadian rhythm of core temperature: effects of physical activity and aging. Physiol Behav. 90,
246–256 (2007).
Figure 8. Schematic illustration showing the circadian clock disruption immediately preceding death at both the behavioural and molecular levels (A) and the hypothesised pathways responsible (B). The loss in the rhythmicity of locomotor activity in the last few days of life is mirrored by a simultaneous increase of clock-gene expression. The negative-regulated transcription-translation feedback loop is composed mainly of gene per and tim, their translational products PER and TIM in cytoplasm form a heterodimer which enters the nucleus to inhibit the transcription of per and tim themselves. The change of clock mechanism before death starts with the increasing transcription of tim without following a circadian pattern (here shown at day 5). We hypothesise this is due to the loss of regulation by the transcription-translation feedback loop. The increase of PER is likely to be a downstream effect of the aberrant expression of TIM. The changes of TIM may be related to its role in clock rhythmicity, cell cycle, DNA replication, DNA damage response, telomere length and integrity maintenance, and immune response.

www.nature.com/scientificreports/
1 0Scientific RepoRTs | (2018) 8:11923 | DOI:10.1038/s41598-018-30323-x
5. Brown, S. A., Pagani, L., Cajochen, C. & Eckert, A. Systemic and cellular refl ctions on ageing and the circadian oscillator: a mini-review. Gerontology. 57, 427–434 (2011).
6. Banks, G., Nolan, P. M. & Peirson, S. N. Reciprocal interactions between circadian clocks and aging. Mamm Genome. 27, 332–340 (2016). 7. Froy, O. Circadian rhythms, aging, and life span in mammals. Physiology (Bethesda). 26, 225–235 (2011). 8. Koh, K., Evans, J. M., Hendricks, J. C. & Sehgal, A. A Drosophila model for age-associated changes in sleep:wake cycles. Proc Natl
Acad Sci USA 103, 13843–13847 (2006). 9. Umezaki, Y., Yoshii, T., Kawaguchi, T., Helfrich-Förster, C. & Tomioka, K. Pigment-dispersing factor is involved in age-dependent
rhythm changes in Drosophila melanogaster. J Biol Rhythms. 27, 423–432 (2012). 10. Giebultowicz, J. M. & Long, D. M. Ageing and circadian rhythms. Curr Opin Insect Sci. 7, 82–86 (2015). 11. Luo, W. et al. Old flies have a robust central oscillator but weaker behavioral rhythms that can be improved by genetic and
environmental manipulations. Aging Cell. 11, 428–438 (2012). 12. Basso, A. et al. Circadian rhythms of body temperature and locomotor activity in aging BALB/c mice: early and late life span
predictors. Biogerontology. 17, 703–714 (2016). 13. Morin, L. P. Age-related changes in hamster circadian period, entrainment, and rhythm splitting. J Biol Rhythms. 3, 237–248 (1988). 14. Davis, F. C. & Viswanathan, N. Stability of circadian timing with age in Syrian hamsters. Am J Physiol. 275, R960–968 (1998). 15. Weinert, H. & Weinert, D. Circadian activity rhythms of laboratory mice during the last weeks of their life. Biol Rhythm Res. 29,
159–178 (1998). 16. Nakamura, T. J., Takasu, N. N. & Nakamura, W. The suprachiasmatic nucleus: age-related decline in biological rhythms. J Physiol Sci.
66, 367–374 (2016). 17. Yamazaki, S. et al. Effects of aging on central and peripheral mammalian clocks. Proc Natl Acad Sci USA 99, 10801–10806 (2002). 18. Helfrich-Förster, C. Neurobiology of the fruit fly’s circadian clock. Genes Brain Behav. 4, 65–76 (2005). 19. Ito, C. & Tomioka, K. Heterogeneity of the peripheral circadian systems in Drosophila melanogaster: a review. Front Physiol. 7, 8 (2016). 20. Sehgal, A., Ousley, A. & Hunter-Ensor, M. Control of circadian rhythms by a two-component clock. Mol Cell Neurosci. 7, 165–172 (1996). 21. Stanewsky, R., Lynch, K. S., Brandes, C. & Hall, J. C. Mapping of elements involved in regulating normal temporal period and
timeless RNA expression patterns in Drosophila melanogaster. J Biol Rhythms. 17, 293–306 (2002). 22. Veleri, S., Brandes, C., Helfrich-Förster, C., Hall, J. C. & Stanewsky, R. A self-sustaining, light-entrainable circadian oscillator in the
Drosophila brain. Curr Biol. 13, 1758–1767 (2003). 23. Rakshit, K., Krishnan, N., Guzik, E. M., Pyza, E. & Giebultowicz, J. M. Effects of aging on the molecular circadian oscillations in
Drosophila. Chronobiol Int. 29, 5–14 (2012). 24. Yoshii, T. et al. The neuropeptide pigment-dispersing factor adjusts period and phase of Drosophila’s clock. J Neurosci. 29, 2597–2610
(2009). 25. Driver, C. The circadian clock in old Drosophila melanogaster. Biogerontology. 1, 157–162 (2000). 26. Krishnan, N., kretzschmar, D., Rakshit, K., Chow, E. & Giebultowicz, J. M. The circadian clock gene period extends healthspan in.
aging Drosophila melanogaster. Aging (Albany NY). 1, 937–948 (2009). 27. Mattis, J. & Sehgal, A. Circadian rhythms, sleep, and disorders of aging. Trends Endocrinol Metab. 27, 192–203 (2016). 28. Vaccaro, A., Issa, A.-R., Seugnet, L., Birman, S. & Klarsfeld, A. Drosophila clock is required in brain pacemaker neurons to prevent
premature locomotor aging independently of its circadian function. Plos Genet. 13, e1006507 (2017). 29. Mazzoccoli, G., Laukkanen, M. O., Vinciguerra, M., Colangelo, T. & Colantuoni, V. A Timeless link between circadian patterns and
disease. Trends Mol Med. 22, 68–81 (2016). 30. Chi, L. et al. TIMELESS contributes to the progression of breast cancer through activation of MYC. Breast Cancer Res. 19, 53 (2017). 31. Liu, S. L. et al. TIMELESS confers cisplatin resistance in nasopharyngeal carcinoma by activating the Wnt/β-catenin signaling
pathway and promoting the epithelial mesenchymal transition. Cancer Lett. 402, 117–130 (2017). 32. Zhang, W. et al. Aberrant TIMELESS expression is associated with poor clinical survival and lymph node metastasis in early-stage
cervical carcinoma. Int J Oncol. 50, 173–184 (2017). 33. Gutiérrez-Gutiérrez, Ó., Felix, D. A. & González-Estévez, C. Planarian fi ds time(less) to fi ht infection. Virulence. 2, 1–6 (2017). 34. Solanas, G. et al. Aged stem cells reprogram their daily functions to adapt to stress. Cell. 170, 678–692 (2017). 35. Pozhitkov, A. E. & Noble, P. A. Gene expression in the twilight of death: The increase of thousands of transcripts has implications to
transplantation, cancer, and forensic research. Bioessays. 39 (2017). 36. Shaw, P., Ocorr, K., Bodmer, R. & Oldham, S. Drosophila aging 2006/2007. Exp Gerontol. 43, 5–10 (2008). 37. Gibson, E. M., Williams, W. P. & Kriegsfeld, L. J. Aging in the circadian system: considerations for health, disease prevention and
longevity. Exp Gerontol. 44, 51–56 (2009). 38. Stanewsky, R. Analysis of rhythmic gene expression in adult Drosophila using the fi efly luciferase reporter gene. Methods Mol Biol.
362, 131–142 (2007). 39. Zordan, M. A., Benna, C. & Mazzotta, G. Monitoring and analyzing Drosophila circadian locomotor activity. Methods Mol Biol. 362,
67–81 (2007).
Author ContributionsJ.Z. and J.F.C. designed research and analysed data; J.Z. performed research; and J.Z., G.R.W. and J.F.C. wrote the paper.
Additional InformationCompeting Interests: The authors declare no competing interests.Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or
format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Cre-ative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not per-mitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/. © The Author(s) 2018




















