
Ornamental plants for micropollutant removal in wetland systems
Upload
independentCategory
view
1download
0
ARTICLE IN PRESS
Ecotoxicology and Environmental Safety 72 (2009) 693–698
Contents lists available at ScienceDirect
Ecotoxicology and Environmental Safety
0147-65
doi:10.1
$ Ani
conduct
animal� Corr
fax: +55
E-m
journal homepage: www.elsevier.com/locate/ecoenv
Copper sensitivity of wild ornamental fish of the Amazon$
Rafael Mendonc-a Duarte, Ana Cristina Leite Menezes, Leonardo da Silveira Rodrigues,Vera Maria Fonseca de Almeida-Val, Adalberto Luis Val �
Laboratory of Ecophysiology and Molecular Evolution, Brazilian National Institute for Amazon Research (INPA), Ave Andre Araujo 2936, 69060-001 Manaus, AM, Brazil
a r t i c l e i n f o
Article history:
Received 28 June 2008
Received in revised form
24 September 2008
Accepted 5 October 2008Available online 6 December 2008
Keywords:
Amazon
Rio Negro
Copper sensitivity
Ornamental fish
LC50
Characids
Catfish
Cichlids
13/$ - see front matter & 2008 Elsevier Inc. A
016/j.ecoenv.2008.10.003
mal welfare: On behalf of co-authors I assur
ed in accordance with Brazilian and Institu
welfare.
esponding author. Tel.: +55 92 3643 3190/+55
92 643 3186.
ail addresses: [email protected], dalval@inter
a b s t r a c t
The copper sensitivity of 10 wild ornamental fish from Rio Negro, an ion-poor river in the Amazon, was
analyzed in the absence of dissolved organic carbon (DOC). No mortality was observed at concentrations
up to 700mg Cu l�1 for two species exposed to CuCl2 �2H2O dissolved in standard EPA-USA water. Copper
sensitivity of fish was also tested in INPA groundwater, which is similar to that of the Rio Negro, but
lacking typical DOC of Rio Negro water. In these experiments, all four characid fish species were more
sensitive to copper than the catfish and the cichlids. LC50 values ranged from 12.8mg Cu l�1 for the
characid Hemigrammus rhodostomus to 74.1mg Cu l�1 for the callichtyid Dianema urostriatum. Copper
sensitivity of Hyphessobrycon socolofi was 52-fold lower in EPA (1405.5mg Cu l�1) than in INPA water
(26.8mg Cu l�1). The differences in values of LC50 may be related to species-specific physiological
abilities to ion-regulate under the original conditions of the habitat of the analyzed species.
& 2008 Elsevier Inc. All rights reserved.
1. Introduction
Copper is an essential element required by all living organismsfor several physiological functions and biochemical reactions(Linder and Hazegh-Azam, 1996). Although copper is an essentialmetal, it can be toxic to fish. The magnitude of its toxicity dependson the fish species and the characteristics of the water (Grosellet al., 2002). Interest in the effects of copper on freshwater fishhave led to numerous studies on copper uptake mechanisms,copper accumulation in the gills, and other target organs,differences in copper sensitivity between species, mechanismsof acute copper toxicity, and the general physiological effects ofcopper (Wood, 2001). However, these studies have been limited totemperate fish species and to waters with similar chemical andphysical compositions. Information is very limited concerning thebiological effects of copper on tropical fish in waters with extremecharacteristics.
The Amazon basin contains the richest fish fauna in the world.The number of fish species exceeds 3000 and includes represen-tatives of almost all freshwater fish groups (Val and Almeida-Val,
ll rights reserved.
e that all experiments were
tional (INPA) guidelines for
92 3643 3189;
next.com.br (A.L. Val).
1995). The dominant fish group is the Ostariophysi, which isby far the largest teleost group of fish living in the Amazon.Among the ostariophysans, Characiformes (scaled fishes) isthe largest order; it contains more than 1200 fish species.Siluriformes (catfishes and armored catfishes) constitute thesecond largest order, comprising more than 1000 fish species.Complex environmental interactions affect the distribution of fishacross the myriad habitats in the Amazon (Almeida-Val et al.,1993; Val et al., 1998), and they determine the ability of animalsto survive acute environmental changes, such as frequentepisodes of hypoxia and anoxia in the waters of the Amazon(Randall et al., 1981; Val et al., 1998). The animals must also copewith the ion-poor, highly acidic pH of many of the water bodies inthe region (Gonzalez et al., 2002; Wilson et al., 1999; Wood et al.,1998).
Amazon waters display a remarkable array of colors dueto characteristics of the drainage area in the sub-basin. Threegeneral types of water are easily recognized: ‘‘white’’, ‘‘black’’, and‘‘clear’’ (Sioli, 1984). Intermediate patterns also occur. The ‘‘white’’water, which carries a large amount of silt, typically occurs in therivers flowing out of the Andes, such as the Solimoes-AmazonasRiver. ‘‘Black’’ water is highly transparent, much more than‘‘white’’ water. The black color results from high levels of humicand fulvic acids and other secondary plant metabolites leachedfrom poorly drained soils. These soils remain swampy for manymonths every year. ‘‘Black’’ water is acidic and found in riversdraining both the Brazilian and the Guiana shields. ‘‘Clear’’ water

ARTICLE IN PRESS
R.M. Duarte et al. / Ecotoxicology and Environmental Safety 72 (2009) 693–698694
contains high mineral levels and much less humic and fulvic acids,even though clear water is found in rivers that drain poor ion soils,such as the Xingu and Tapajos Rivers (Val and Almeida-Val, 1995).Increased levels of metals affect fish differently, depending ontheir physiology and metal bioavailability in different types ofwater.
The three major sources of copper in the waters of the Amazonare substrate lithology, mining activities, and modern anthropo-genic activities. In general, the copper content in the watersreflects variations in the chemical composition of the soil andsediment. The copper content is very low in the waters of all threegeological provinces of the Amazon, as compared to the copperlevels in the world’s rivers. Within the Amazon, the largestdifference in copper content exists between the whitewatersof the Rio Solimoes and the blackwaters of the Rio Negro(Kornhauser et al., 1994). The waters of Rio Negro have copperconcentrations ranging from 0.03 to 0.1mg/l. These values areabout two orders of magnitude lower than those in the waters ofRio Solimoes.
Mining activities occur in relatively small areas, but theynevertheless can have a large local impact on the environment.In the Amazon, petroleum and copper mining are the majormining activities that can release copper to the environment.Petroleum mining at the upper reaches of the Urucu River, 700 kmnorthwest of Manaus, generates nine barrels of productionwater per barrel of crude oil. Copper concentrations in thisproduction water are approximately 250mg/l. Most of the water isre-injected into the wells, so it has minimal environmentalimpact, unless a leak occurs. In contrast, significant copperdeposits occur in the Mineral Province of Carajas, where threedeposits together contain 12,000 MT of copper. These depositsextend over an area of 25,000 ha on one side of the Salobo River,which is 20 km long (Damous et al., 2002). Currently, ametallurgic project has just begun to extract copper, iron, andgold. These are the three major elements found in Carajasprovince. The natural background copper concentration in watersof the Salobo River is 3.2–3.8mg/l. Near the mining areas, thecopper concentration exceeds 15mg/l. The chemical characteris-tics of Salobo River water include reduced levels of carbonate andchloride ligands, slightly acid pH, low calcium levels, and lowlevels of inorganic anions compared to the water of the AmazonRiver. These characteristics limit the ability of Salobo River waterto buffer the possible effects of anthropogenic copper on localbiota.
The third source of copper in Amazonian waters is a result ofthe recent increase in the industrial use of copper. In the majorcities of the Amazon, copper use has increased only slightly, but ithas increased dramatically in the waters of the streams crossingthe industrial pole in Manaus, where it has raised majorenvironmental concerns. Copper levels in the waters of the majorstream in this basin, the ‘‘Igarape do 40’’, exceed 1000mg/l(Sampaio, 2000).
The form of copper toxic to fish is the free divalent cation form,Cu2+ (Campbell, 1995). In fresh water fish, copper disrupts Na+ andCl� active uptake mechanisms in the fish gills. Fish exposed tohigh copper concentrations experience an increase in gill perme-ability that results in net ion loss and subsequent death (Laurenand McDonald, 1985). The sensitivity of fish to copper is expressedas the concentration of dissolved copper that kills 50% of thesample population (LC50). The lower the LC50, the higher thesensitivity of the species is to copper. Amazon fish are increasinglyexposed to environmental changes of anthropogenic origin,including increased background levels of copper. In this study,the sensitivity of 10 fish species to copper was measured in termsof the LC50 after 96 h of exposure to ion-poor water undercontrolled conditions.
2. Materials and methods
2.1. Experimental animals
Ten fish species were acquired from two ornamental fish exporters (Turky’s
and K2 aquariums) and transported to the Laboratory of Ecophysiology and
Molecular Evolution at the Brazilian National Institute for Amazon Research
(LEEM-INPA), located in Manaus. The fish were collected from their natural
environment around Barcelos city (�500 km from Manaus) near the banks of the
Rio Negro. The fish were kept for 2 weeks in aerated tanks (150 l) supplied with a
flow-through INPA groundwater ([Na+], 0.83; [K+], 0.45; [Ca2+], 0.10; [Mg2+], 0.040;
[Cl�], 0.90 mg l�1; [Cu2+], 7.0mg l�1; hardness ¼ 1.33 mg CaCO3 l�1; pH ¼ 6.80).
Fish were fed commercial dry food pellets once a day. Feeding was suspended for
48 h before the start of experiments. The tanks were checked daily for fish
mortality and siphoned every other day to avoid residue accumulation. The levels
of ammonia were measured every other day. There was no mortality observed
during the acclimation period. All experiments were conducted in accordance with
applicable current national guidelines for care and use of laboratory animals.
2.2. Experimental series
Ten animals were transferred to each of the 12 plastic test tanks and allowed to
acclimate for at least 24 h prior to beginning the tests. For these experiments, the
volume of the tanks was adjusted to keep a ratio of 0.8 g of fish per liter of water in
each test tank. All tests were performed in a flow-through system. The first
series of tests was conducted with standard soft water with the composition
recommended by the US Environmental Protection Agency (see Table 1 for soft
standard water composition). Specimens of two characid species, Paracheirodon
axelrodi weighing in average 0.24570.010 g and Hyphessobrycon socolofi weighing
in average 0.79570.020 g, were placed in water containing copper in the form of
CuCl2 �2H2O, dissolved at concentrations ranging from 15.2 to 2602mg Cu l�1.
The second series of tests was conducted using INPA groundwater (see Table 1
for water composition). Initially, the LC50 values for the two species previously
mentioned were compared between the two types of water. Subsequent tests were
performed with two more characid species, Carnegiella strigata (weighing
in average 0.28070.011 g) and Hemigrammus rhodostomus (0.33870.019 g);
three catfishes, Corydoras schwartzi (0.94070.019 g), Dianema urostriatum
(3.03670.101 g), and Otocinclus hasemani (0.38970.017 g); and three ciclids,
Dicrossus maculatus (0.68070.009 g), Apistogramma agassizzi (1.26770.015 g),
and Apistogramma hyppolytae (1.21570.084 g). Groups of 10 test tanks were
prepared with five copper concentrations (10–400mg Cu l�1), with the copper
supplied by CuCl2 �2H2O. Finer range of concentrations was additionally used to
obtain more precise measurements of LC50.
An appropriate aliquot of 1 M CuCl2 �2H2O stock solution was added to the
head water tanks (50 or 150 l, depending of the volume of the test tanks), which
were then stirred for at least 6 h. At the start of the two sets of tests, the water of
the test tanks was carefully replaced with the appropriate experimental solution
and water recirculation was launched using a system set to a flow rate that
replaced 80% of the water volume every 24 h. Test tanks were continuously aerated
during the experiments.
Water samples were collected every 12 h from both test tanks and the head tank.
The samples were acidified with 100ml of concentrated HNO3 and stored at �20 1C
for subsequent copper analysis. Copper concentrations were measured by atomic
absorption spectophotometry using a graphite furnace technique for a Perkin-Elmer
Analyst 800 AA spectrophotometer. The concentrations of the major cations, Na+, K+,
Ca2+, and Mg2+, were measured by flame technique on this instrument. Aqueous
concentrations of Cl� were measured using the colorimetric assay described by Zall
et al. (1956). For DOC analysis, water samples were collected in darkened bottles and
acidified with concentrated H2SO4 until the pH was below 2, then shipped cooled at
4 1C to the environmental sanitation company CETESB (Sao Paulo). The DOC
concentrations were determined using a total carbon analyzer (Shimadzu Model
5000) using combustion method with infrared detection. Water pH, dissolved
oxygen, and water temperature were measured using a YSI water analyzer. Hardness
was measured by the standard titration method (APHA, 1998).
2.3. Calculations
Estimates of LC50 and associated 95% confidence limits were determined by
the trimmed Spearman–Karber method (Hamilton et al., 1977), using measured
concentrations of copper. Mean fish mortality was recorded every 12 h. The LC50
values were determined after 24, 48, 72, and 96 h.
3. Results
Table 1 summarizes the water characteristics in the test tanksfor all 10 species analyzed. When INPA groundwater was used, no
ARTICLE IN PRESS
Table 1Water composition in the test tanks
Species Na+ (mg l�1) K+ (mg l�1) Ca2+ (mg l�1) Mg2+ (mg l�1) Cl�1 (mg l�1) pH DOCa (mg C l�1) Hardness
C. strigatta 0.8370.04 0.7270U06 0.2070.02 0.0670.002 1.7970.011 6.7370.09 3.1770.45 5.0870.70
H. rhodostomus 1.2070.08 1.4270.13 0.3970.04 0.0970.006 1.6670.009 7.0570.04 3.1770.45 5.6070.20
H. socolofi 1.3970.02 1.1070.24 0.3770.03 0.0770.002 1.7870.014 6.8470.07 3.1770.45 5.0870.70
P. axelrodi 1.4670.03 1.3870.02 0.6970.20 0.0770.02 1.8270.060 6.4670.18 3.1770.45 5.1270.63
D. urostriatum 1.3570.10 1.1970.12 0.5170.08 0.1070.01 2.2770.23 6.8170.04 3.1770.45 5.4070.30
C. schwartzi 1.2570.08 1.1470.07 0.4870.04 0.0870.005 1.9470.11 7.3870.05 3.1770.45 4.9870.30
O. hasemani 1.0370.13 0.7970.03 0.3470.09 0.0770.01 1.6970.09 6.8170.04 3.1770.45 5.6070.20
A. agassizzi 1.6170.03 0.9670.03 1.3270.05 0.1570.03 1.4770.12 6.8970.03 3.1770.45 5.5070.20
A. hyppolytae 1.5270.02 2.2170.05 0.7770.02 0.1170.002 1.3970.01 7.1270.04 3.1770.45 5.6070.02
D. maculatus 1.6170.03 0.9670.03 1.3270.05 0.1570.03 2.1570.17 6.6570.13 3.1770.45 5.4070.03
Standard soft water 3.2370.06 1.2570.02 5.6570.14 6.4570.48 1.7970.08 6.6470.23 1.1870.23 10.5070.99
Rio Negro watera 0.3870.12 0.3370.11 0.2170.07 0.1170.04 1.7070.70 5.170.6 10.571.3 –
The values in the first 10 rows are averages (mean7SEM) of water samples collected every 12 h from each test tank. For DOC the value represents the average of 10 samples.
The values in the last row are averages of six water samples collected from test tanks containing standard EPA soft water.a Val and Almeida-Val (1995).
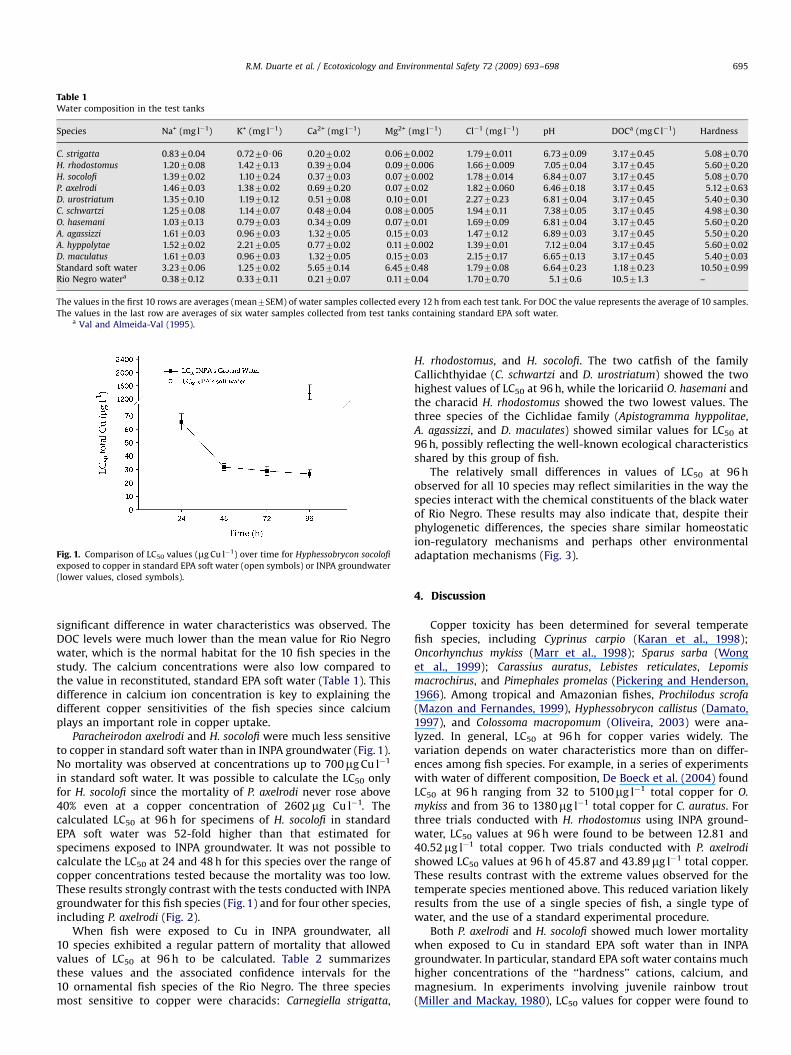
Fig. 1. Comparison of LC50 values (mg Cu l�1) over time for Hyphessobrycon socolofi
exposed to copper in standard EPA soft water (open symbols) or INPA groundwater
(lower values, closed symbols).
R.M. Duarte et al. / Ecotoxicology and Environmental Safety 72 (2009) 693–698 695
significant difference in water characteristics was observed. TheDOC levels were much lower than the mean value for Rio Negrowater, which is the normal habitat for the 10 fish species in thestudy. The calcium concentrations were also low compared tothe value in reconstituted, standard EPA soft water (Table 1). Thisdifference in calcium ion concentration is key to explaining thedifferent copper sensitivities of the fish species since calciumplays an important role in copper uptake.
Paracheirodon axelrodi and H. socolofi were much less sensitiveto copper in standard soft water than in INPA groundwater (Fig. 1).No mortality was observed at concentrations up to 700mg Cu l�1
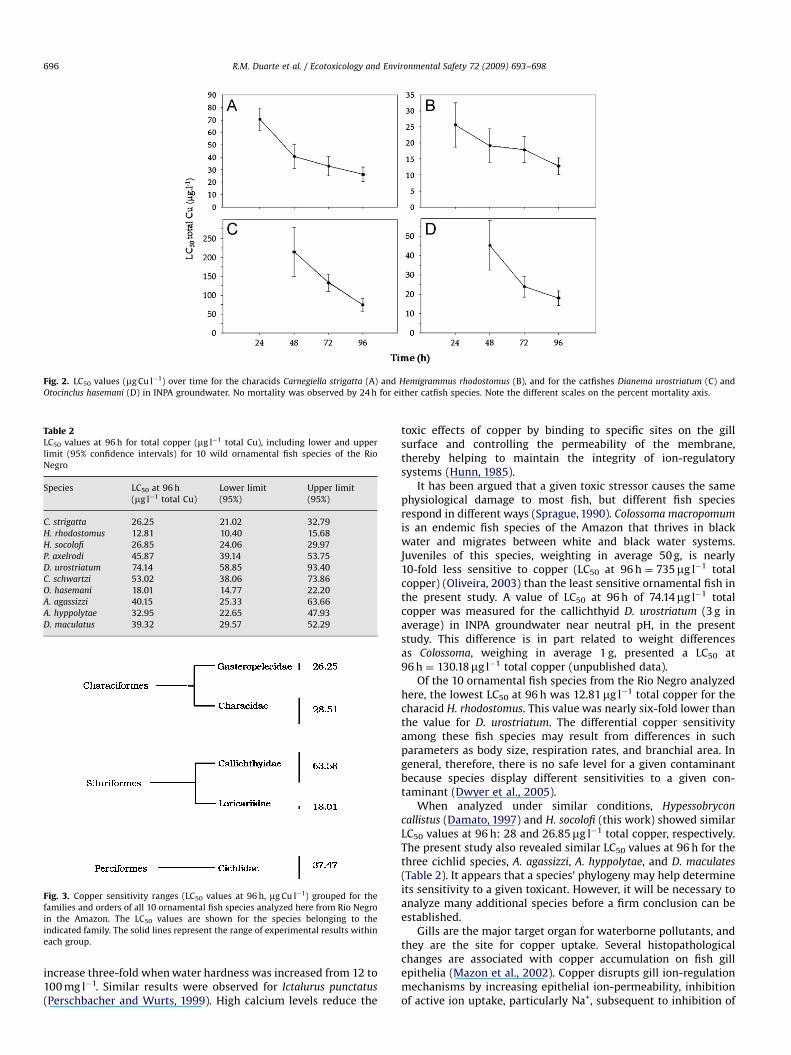
in standard soft water. It was possible to calculate the LC50 onlyfor H. socolofi since the mortality of P. axelrodi never rose above40% even at a copper concentration of 2602mg Cu l�1. Thecalculated LC50 at 96 h for specimens of H. socolofi in standardEPA soft water was 52-fold higher than that estimated forspecimens exposed to INPA groundwater. It was not possible tocalculate the LC50 at 24 and 48 h for this species over the range ofcopper concentrations tested because the mortality was too low.These results strongly contrast with the tests conducted with INPAgroundwater for this fish species (Fig. 1) and for four other species,including P. axelrodi (Fig. 2).
When fish were exposed to Cu in INPA groundwater, all10 species exhibited a regular pattern of mortality that allowedvalues of LC50 at 96 h to be calculated. Table 2 summarizesthese values and the associated confidence intervals for the10 ornamental fish species of the Rio Negro. The three speciesmost sensitive to copper were characids: Carnegiella strigatta,
H. rhodostomus, and H. socolofi. The two catfish of the familyCallichthyidae (C. schwartzi and D. urostriatum) showed the twohighest values of LC50 at 96 h, while the loricariid O. hasemani andthe characid H. rhodostomus showed the two lowest values. Thethree species of the Cichlidae family (Apistogramma hyppolitae,A. agassizzi, and D. maculates) showed similar values for LC50 at96 h, possibly reflecting the well-known ecological characteristicsshared by this group of fish.
The relatively small differences in values of LC50 at 96 hobserved for all 10 species may reflect similarities in the way thespecies interact with the chemical constituents of the black waterof Rio Negro. These results may also indicate that, despite theirphylogenetic differences, the species share similar homeostaticion-regulatory mechanisms and perhaps other environmentaladaptation mechanisms (Fig. 3).
4. Discussion
Copper toxicity has been determined for several temperatefish species, including Cyprinus carpio (Karan et al., 1998);Oncorhynchus mykiss (Marr et al., 1998); Sparus sarba (Wonget al., 1999); Carassius auratus, Lebistes reticulates, Lepomis
macrochirus, and Pimephales promelas (Pickering and Henderson,1966). Among tropical and Amazonian fishes, Prochilodus scrofa
(Mazon and Fernandes, 1999), Hyphessobrycon callistus (Damato,1997), and Colossoma macropomum (Oliveira, 2003) were ana-lyzed. In general, LC50 at 96 h for copper varies widely. Thevariation depends on water characteristics more than on differ-ences among fish species. For example, in a series of experimentswith water of different composition, De Boeck et al. (2004) foundLC50 at 96 h ranging from 32 to 5100mg l�1 total copper for O.
mykiss and from 36 to 1380mg l�1 total copper for C. auratus. Forthree trials conducted with H. rhodostomus using INPA ground-water, LC50 values at 96 h were found to be between 12.81 and40.52mg l�1 total copper. Two trials conducted with P. axelrodi
showed LC50 values at 96 h of 45.87 and 43.89mg l�1 total copper.These results contrast with the extreme values observed for thetemperate species mentioned above. This reduced variation likelyresults from the use of a single species of fish, a single type ofwater, and the use of a standard experimental procedure.
Both P. axelrodi and H. socolofi showed much lower mortalitywhen exposed to Cu in standard EPA soft water than in INPAgroundwater. In particular, standard EPA soft water contains muchhigher concentrations of the ‘‘hardness’’ cations, calcium, andmagnesium. In experiments involving juvenile rainbow trout(Miller and Mackay, 1980), LC50 values for copper were found to
ARTICLE IN PRESS
Fig. 2. LC50 values (mg Cu l�1) over time for the characids Carnegiella strigatta (A) and Hemigrammus rhodostomus (B), and for the catfishes Dianema urostriatum (C) and
Otocinclus hasemani (D) in INPA groundwater. No mortality was observed by 24 h for either catfish species. Note the different scales on the percent mortality axis.
Table 2LC50 values at 96 h for total copper (mg l�1 total Cu), including lower and upper
limit (95% confidence intervals) for 10 wild ornamental fish species of the Rio
Negro
Species LC50 at 96 h
(mg l�1 total Cu)
Lower limit
(95%)
Upper limit
(95%)
C. strigatta 26.25 21.02 32.79
H. rhodostomus 12.81 10.40 15.68
H. socolofi 26.85 24.06 29.97
P. axelrodi 45.87 39.14 53.75
D. urostriatum 74.14 58.85 93.40
C. schwartzi 53.02 38.06 73.86
O. hasemani 18.01 14.77 22.20
A. agassizzi 40.15 25.33 63.66
A. hyppolytae 32.95 22.65 47.93
D. maculatus 39.32 29.57 52.29
Fig. 3. Copper sensitivity ranges (LC50 values at 96 h, mg Cu l�1) grouped for the
families and orders of all 10 ornamental fish species analyzed here from Rio Negro
in the Amazon. The LC50 values are shown for the species belonging to the
indicated family. The solid lines represent the range of experimental results within
each group.
R.M. Duarte et al. / Ecotoxicology and Environmental Safety 72 (2009) 693–698696
increase three-fold when water hardness was increased from 12 to100 mg l�1. Similar results were observed for Ictalurus punctatus
(Perschbacher and Wurts, 1999). High calcium levels reduce the
toxic effects of copper by binding to specific sites on the gillsurface and controlling the permeability of the membrane,thereby helping to maintain the integrity of ion-regulatorysystems (Hunn, 1985).
It has been argued that a given toxic stressor causes the samephysiological damage to most fish, but different fish speciesrespond in different ways (Sprague, 1990). Colossoma macropomum
is an endemic fish species of the Amazon that thrives in blackwater and migrates between white and black water systems.Juveniles of this species, weighting in average 50 g, is nearly10-fold less sensitive to copper (LC50 at 96 h ¼ 735mg l�1 totalcopper) (Oliveira, 2003) than the least sensitive ornamental fish inthe present study. A value of LC50 at 96 h of 74.14mg l�1 totalcopper was measured for the callichthyid D. urostriatum (3 g inaverage) in INPA groundwater near neutral pH, in the presentstudy. This difference is in part related to weight differencesas Colossoma, weighing in average 1 g, presented a LC50 at96 h ¼ 130.18mg l�1 total copper (unpublished data).
Of the 10 ornamental fish species from the Rio Negro analyzedhere, the lowest LC50 at 96 h was 12.81mg l�1 total copper for thecharacid H. rhodostomus. This value was nearly six-fold lower thanthe value for D. urostriatum. The differential copper sensitivityamong these fish species may result from differences in suchparameters as body size, respiration rates, and branchial area. Ingeneral, therefore, there is no safe level for a given contaminantbecause species display different sensitivities to a given con-taminant (Dwyer et al., 2005).
When analyzed under similar conditions, Hypessobrycon
callistus (Damato, 1997) and H. socolofi (this work) showed similarLC50 values at 96 h: 28 and 26.85mg l�1 total copper, respectively.The present study also revealed similar LC50 values at 96 h for thethree cichlid species, A. agassizzi, A. hyppolytae, and D. maculates
(Table 2). It appears that a species’ phylogeny may help determineits sensitivity to a given toxicant. However, it will be necessary toanalyze many additional species before a firm conclusion can beestablished.
Gills are the major target organ for waterborne pollutants, andthey are the site for copper uptake. Several histopathologicalchanges are associated with copper accumulation on fish gillepithelia (Mazon et al., 2002). Copper disrupts gill ion-regulationmechanisms by increasing epithelial ion-permeability, inhibitionof active ion uptake, particularly Na+, subsequent to inhibition of

ARTICLE IN PRESS
R.M. Duarte et al. / Ecotoxicology and Environmental Safety 72 (2009) 693–698 697
Na+–K+-ATPase and a decrease in the number of mitochondrial-rich cells in the gills (Lauren and McDonald, 1985). This disruptioncauses fish death. Metal toxicity in aquatic organisms has beenshown to relate directly to the chemical composition of the water,particularly the levels of Ca2+, Na+, and Mg2+; the pH; and thedissolved organic carbon (DOC) content (Niyogi and Wood, 2004).The ions can compete with copper for the gill’s ion uptake sites,and the pH and DOC can alter the bioavailability of copper forbinding to gill epithelia. In fact, the ability of DOC to reducecopper toxicity has been reported for the characid C. macropomum
(Matsuo et al., 2005). INPA groundwater is very low in both ionsand DOC, which contributes to the high copper sensitivity in thiswater observed for all 10 species analyzed in the present study.
Although Rio Negro and its tributaries are considered anextreme environment for aquatic life, a broad diversity of fishthrives there. Many of these fish species show specializedadaptations (Val and Almeida-Val, 1995). One example of adapta-tion is ion homeostasis: species of the Characidae family from theRio Negro show a very high Na+ binding affinity that ensures ahigh rate of Na+ uptake (Gonzalez et al., 2002). However, in theabsence of DOC, these species are highly vulnerable to dissolvedcopper, possibly because of a slowing in this normally high rate ofuptake. In future studies, the putative protective role of Rio NegroDOC as an antagonist of copper toxicity will be examined. Themechanisms of copper toxicity in native fish species will also bestudied.
5. Conclusion
All 10 wild ornamental fish species of the Rio Negro showed ahigh copper sensitivity when assayed in the absence of DOC. LC50
values at 96 h ranged from 12.8mg Cu l�1 for the characidH. rhodostomus to 74.1mg Cu l�1 for the callichtyid D. urostriatum.
The analyzed fish groups seem to present a discrete mean value.However, there is no clear phylogenetic trend from less specia-lized to more advanced fish groups. When assayed in the presenceof high levels of calcium and magnesium, a substantially lowercopper sensitivity—in other words, much higher LC50 values at96 h was observed. To our knowledge, this study is the firstcomprehensive examination of the effect of water constituents ona wide range of tropical fish species from the same habitat. Ithelps to refine our understanding of copper sensitivity among thefish of the Amazon. The findings show that fish species of RioNegro are extremely sensitive to the toxic effects of copper whenexposed to INPA well DOC-free ion-poor water.
Acknowledgements
This work was supported by a research grant from theInternational Copper Association (COPPER) and a PRONEX grantfrom the Brazilian National Research Council (CNPq) and theAmazonas State Research Foundation (FAPEAM). We thank Turky’sand K2 Aquariums for the donation of all experimental animals.RMD, LSR, and ACLM were recipients of Master’s fellowships fromCNPq during the experimental period. ALV and VMFAV wererecipients of research fellowships from CNPq.
References
Almeida-Val, V.M.F., Val, A.L., Hochachka, P.W., 1993. Hypoxia tolerance in Amazonfishes: status of an under-explored biological ‘‘goldmine’’. In: Hochachka, P.W.,et al. (Eds.), Surviving Hypoxia: Mechanisms of Control and Adaptation. CRCPress, Boca Raton, FL, pp. 435–445.
APHA, 1998. Standard Methods for the Examination of Water and Wastewater.APHA, Washington, DC.
Campbell, P.G.C., 1995. Interactions between trace metals and aquatic organisms: acritique of the free-ion activity model. In: Tessier, A., Tunrer, D.R. (Eds.),Metal Speciation and Bioavailability in Aquatic Systems. Wiley, Chichester,pp. 45–102.
Damato, M., 1997. Estudo da influencia do nıvel de tratamento de efluentes derefinarias de petroleo na sua toxicidade, empregando diferentes especiesindicadoras. Ph.D. Dissertation, Universidade de Sao Paulo, Sao Paulo.
Damous, N.R., Wagener, A.L.R., Patchineelam, S.R., Wagener, K., 2002. Baselinestudies on water and sediments in the copper mining region of Salobo-3a,Carajas-Amazon, Brazil. J. Braz. Chem. Soc. 13, 140–150.
De Boeck, G., Meeus, W., De Coen, W., Blust, R., 2004. Tissue-specific accumulationCu bioaccumulation patterns and differences in sensitivity to waterborne Cu inthree freshwater fish: rainbow trout (Oncorhynchus mykiss), common carp(Cyprinus carpio), and gibel carp (Carassius auratus gibelio). Aquat. Toxicol. 70,179–188.
Dwyer, F.J., Mayer, F.L., Sappington, L.C., Buckler, D.R., Bridges, C.M., Greer, I.E.,Hardesty, D.K., Henke, C.E., Ingersoll, C.G., Kunz, J.L., Whites, D.W., Augspurger,T., Mount, D.R., Hattala, K., Neuderfer, G.N., 2005. Assessing contaminantsensitivity of endangered and threatened aquatic species: part I. Acute toxicityof five chemicals. Arch. Environ. Contam. Toxicol. 48, 143–154.
Gonzalez, R.J., Wilson, R.W., Wood, C.M., Patrick, M.L., Val, A.L., 2002. Diversestrategies for ion regulation in fish collected from the ion-poor, acidic RioNegro. Physiol. Biochem. Zool. 75, 37–47.
Grosell, M., Nielsen, C., Bianchini, A., 2002. Sodium turnover rate determinessensitivity to acute copper and silver exposure in freshwater animals. Comp.Biochem. Physiol. 133C, 287–303.
Hamilton, M.A., Russo, R.C., Thurston, R.V., 1977. Trimmed Spearman–Karbermethod for estimating lethal concentrations in toxicity bioassays. Environ. Sci.Technol. 11, 714–719.
Hunn, J.B., 1985. Role of calcium in gill function in freshwater fishes. Comp.Biochem. Physiol. 82A, 543–547.
Karan, V., Vitorovic, S., Tutundzic, V., Poleksic, V., 1998. Functional enzymesactivity and gill histology of carp after copper sulfate exposure and recovery.Ecotoxicol. Environ. Saf. 40, 49–55.
Kornhauser, K.O., Fyfe, W.S., Kronberg, B.I., 1994. Multi-element chemistry of someAmazonian waters and soils. Chem. Geol. 111, 155–175.
Lauren, D.J., McDonald, D.G., 1985. Effects of copper on branchial ionoregulation inthe rainbow trout, Salmo gairdneri, Richardson. J. Comp. Physiol. B 155,635–644.
Linder, M.C., Hazegh-Azam, M., 1996. Copper biochemistry and molecular biology.Am. J. Clin. Nutr. 63, 797s–811s.
Marr, J.C.A., Hansen, J.A., Meyer, J.S., Cacela, D., Podrabsky, T., Lipton, J., Bergman,H.L., 1998. Toxicity of cobalt and copper to rainbow trout: application of amechanistic model for predicting survival. Aquat. Toxicol. 43, 225–238.
Matsuo, A.Y.O., Wood, C.M., Val, A.L., 2005. Effects of copper and cadmium on iontransport and gill metal binding in the Amazonian teleost tambaqui (Colossomamacropomum) in extremely soft water. Aquat. Toxicol. 74, 351–364.
Mazon, A.F., Fernandes, M.N., 1999. Toxicity and differential tissue accumulation ofcopper in the tropical freshwater fish, Prochilodus scrofa (Prochilodontidae).Bull. Environ. Contam. Toxicol. 63, 797–804.
Mazon, A.F., Cerqueira, C.C., Fernandes, M.N., 2002. Gill cellular changes induced bycopper exposure in the South American tropical freshwater fish Prochilodusscrofa. Environ. Res. 88, 52–63.
Miller, T.G., Mackay, W.C., 1980. The effects of hardness, alkalinity and pH of testwater on the toxicity of copper to rainbow trout (Salmo gairdneri). Water Res.14, 129–133.
Niyogi, S., Wood, C.M., 2004. Biotic ligand model, a flexible tool for developingsite-specific water quality guidelines for metals. Environ. Sci. Technol. 38,6177–6192.
Oliveira, C.P.F., 2003. Efeito de dois metais pesados presentes na agua de formac- aoderivada da extrac- ao do petroleo da provıncia petroleira do Urucu-AM sobre otambaqui, Colossoma macropomum (Cuvier, 1818). Master Dissertation, INPA,UFAM, Manaus.
Perschbacher, P.W., Wurts, W.A., 1999. Effects of calcium and magnesium hardnesson acute copper toxicity to juvenile channel catfish Ictalurus punctatus.Aquaculture 172, 275–280.
Pickering, Q.H., Henderson, C., 1966. The acute toxicity of some heavy metals todifferent species of warm water fishes. Int. J. Air Water Pollut. 10, 453–463.
Randall, D.J., Burggren, W.W., Farrell, A.P., Haswell, M.S., 1981. The Evolutionof Air-Breathing Vertebrates. Cambridge University Press, Cambridge.
Sampaio, A.Q., 2000. Caracterizac- ao fısica e quımica dos sedimentos na area doDistrito Industrial de Manaus (AM). PPG em Ciencias Ambientais. UniversidadeFederal do Amazonas, Manaus.
Sioli, H., 1984. The Amazon. Limnology and Landscape Ecology of a Might TropicalRiver and Its Basin. Dr. W. Junk Publishers, Dordrecht.
Sprague, J.B., 1990. Aquatic toxicology. In: Schrench, C.B., Moyle, P.B.(Eds.), Methods for Fish Biology. American Fisheries Society, Bethesda,pp. 491–528.
Val, A.L., Almeida-Val, V.M.F., 1995. Fishes of the Amazon and Their Environments.Physiological and Biochemical Features. Springer, Heidelberg.
Val, A.L., Silva, M.N.P., Almeida-Val, V.M.F., 1998. Hypoxia adaptation in fish of theAmazon: a never-ending task. S. Afr. J. Zool. 33, 107–114.
Wilson, R.W., Wood, C.M., Gonzalez, R.J., Patrick, M.L., Bergman, H.L., Narahara, A.,Val, A.L., 1999. Ion and acid–base balance in three species of Amazonian fishduring gradual acidification of extremely soft water. Physiol. Biochem. Zool. 72,277–285.

ARTICLE IN PRESS
R.M. Duarte et al. / Ecotoxicology and Environmental Safety 72 (2009) 693–698698
Wong, P.P.K., Chu, L.M., Wong, C.K., 1999. Study of toxicity and bioaccumulation ofcopper in the silver sea bream Sparus sarba. Environ. Int. 25, 417–422.
Wood, C.M., 2001. The toxic response of the gill. In: Benson, H.W., Schlenk, D.W.(Eds.), Target Organ Toxicity in Marine and Freshwater Teleost. Taylor &Francis, Washington, pp. 1–87.
Wood, C.M., Wilson, R.W., Gonzalez, R.J., Patrick, M.L., Bergman, H.L., Narahara, A.,Val, A.L., 1998. Responses of an Amazonian teleost, the tambaqui (Colossomamacropomum) to low pH in extremely soft water. Physiol. Zool. 71, 658–670.
Zall, D.M., Fisher, D., Garner, M.Q., 1956. Photometric determination of chloride inwater. Anal. Chem. 28, 1665–1668.
Copyright © 2022 FDOKUMEN




















