
Contribution of Naïve and Memory T-Cell Populations to the Human Alloimmune Response
10
American Journal of Transplantation 2009; 9: 2057–2066 Wiley Periodicals Inc. C ⃝ 2009 The Authors Journal compilation C ⃝ 2009 The American Society of Transplantation and the American Society of Transplant Surgeons doi: 10.1111/j.1600-6143.2009.02742.x Contribution of Na¨ ıve and Memory T-Cell Populations to the Human Alloimmune Response C. Macedo a , E. A. Orkis a , I. Popescu a , B. D. Elinoff a , A. Zeevi a, b, c , R. Shapiro a , F. G. Lakkis a, b, † and D. Metes a, b, ∗, † a Human Immunology Program, Thomas E. Starzl Transplantation Institute, Departments of Surgery, b Immunology and c Pathology, University of Pittsburgh, Pittsburgh, PA * Corresponding author: Diana Metes, [email protected] †These authors contributed equally to the work. T-cell alloimmunity plays a dominant role in allograft rejection. The precise contribution of na¨ ıve and mem- ory T cells to this response however remains un- clear. To address this question, we established an ex vivo flow-cytometric assay that simultaneously mea- sures proliferation, precursor frequency and effector molecule (IFNc , granzyme B/perforin) production of alloreactive T cells. By applying this assay to periph- eral blood mononuclear cells from healthy volunteers, we demonstrate that the CD4 + and CD8 + popula- tions mount similar proliferative responses and con- tain comparable frequencies of alloreactive precursors. Effector molecule expression, however, was signifi- cantly higher among CD8 + T cells. Analysis of sorted na¨ ıve and memory T cells showed that alloreactive precursors were equally present in both populations. The CD8 + effector and terminally differentiated effec- tor memory subsets contained the highest propor- tion of granzyme B/perforin after allostimulation, sug- gesting that these cells present a significant threat to transplanted organs. Finally, we demonstrate that virus-specific lymphocytes contribute significantly to the alloresponse in certain responder–stimulator HLA combinations, underscoring the importance of T-cell cross-reactivity in alloimmunity. These results provide a quantitative assessment of the roles of na¨ ıve and memory T-cell subsets in the normal human alloim- mune response and establish a platform for measuring T-cell alloreactivity pre- and posttransplantation. Key words: Alloimmune response, human, im- munoassay, memory T cells Received 19 March 2009, revised 11 May 2009 and ac- cepted for publication 14 May 2009 Introduction Humans and experimental animals mount a vigorous im- mune response to tissues transplanted from genetically disparate individuals. This response, referred to as alloim- munity, is mediated principally by T lymphocytes that rec- ognize nonself-histocompatibility antigens (alloantigens) including the highly polymorphic human leukocyte anti- gens (HLA) encoded by the major histocompatibility com- plex (MHC) as well as multiple minor histocompatibility antigens encoded by non-MHC genes (1). A salient feature of T-cell alloimmunity is its extraordinary potency which is attributed to the presence of a high fre- quency of host T cells (1–10% of the total T-cell reper- toire) that react to donor alloantigens (2–4). While immune responses to nominal antigens are dependent on a very small proportion of T cells that recognize foreign peptides presented in the context of self-MHC molecules, allore- sponses are in addition generated by a much larger pop- ulation of T cells that recognize peptides complexed to allogeneic MHC on donor cells—a phenomenon known as direct allorecognition (5,6). The high precursor frequency of directly alloreactive T cells is likely the result of the in- herent bias of the prethymic T-cell repertoire toward MHC recognition (7), and the presence of cross-reactive T cells in the postthymic repertoire (6,8). The latter are self-MHC re- stricted mature T cells that are specific to microbial-derived peptides but cross-react with allogeneic MHC. Support for the cross-reactivity hypothesis is provided by molecular evidence that cytotoxic T cells specific to common viruses such as the Epstein-Barr Virus (EBV) exhibit strong allore- activity to particular HLA molecules (9,10). Since humans are exposed to a large array of pathogens beginning very early in life, it is inferred that any given individual harbors a significant number of pathogen-specific T cells that are potentially alloreactive. The development of ex vivo assays to identify, quantitate and analyze alloreactive T cells has been a longstanding interest of human immunologists. It is hoped that such as- says will lead to better understanding of human alloimmu- nity and enable clinicians to predict allograft outcomes and individualize immunosuppression safely. Earlier attempts at studying the human alloimmune response have relied on assays that gauge one parameter or one cell popula- tion at a time; for example, measuring T-cell proliferation in the mixed lymphocyte reaction (MLR), IFNc production in 2057
Transcript of Contribution of Naïve and Memory T-Cell Populations to the Human Alloimmune Response

American Journal of Transplantation 2009; 9: 2057–2066Wiley Periodicals Inc.
C⃝ 2009 The AuthorsJournal compilation C⃝ 2009 The American Society of
Transplantation and the American Society of Transplant Surgeons
doi: 10.1111/j.1600-6143.2009.02742.x
Contribution of Naı̈ve and Memory T-Cell Populationsto the Human Alloimmune Response
C. Macedoa, E. A. Orkisa, I. Popescua,B. D. Elinoffa, A. Zeevia,b,c, R. Shapiroa,F. G. Lakkisa,b,† and D. Metesa,b,∗,†
aHuman Immunology Program, Thomas E. StarzlTransplantation Institute, Departments of Surgery,bImmunology and cPathology, University of Pittsburgh,Pittsburgh, PA*Corresponding author: Diana Metes,[email protected]†These authors contributed equally to the work.
T-cell alloimmunity plays a dominant role in allograftrejection. The precise contribution of naı̈ve and mem-ory T cells to this response however remains un-clear. To address this question, we established an exvivo flow-cytometric assay that simultaneously mea-sures proliferation, precursor frequency and effectormolecule (IFNc , granzyme B/perforin) production ofalloreactive T cells. By applying this assay to periph-eral blood mononuclear cells from healthy volunteers,we demonstrate that the CD4+ and CD8+ popula-tions mount similar proliferative responses and con-tain comparable frequencies of alloreactive precursors.Effector molecule expression, however, was signifi-cantly higher among CD8+ T cells. Analysis of sortednaı̈ve and memory T cells showed that alloreactiveprecursors were equally present in both populations.The CD8+ effector and terminally differentiated effec-tor memory subsets contained the highest propor-tion of granzyme B/perforin after allostimulation, sug-gesting that these cells present a significant threatto transplanted organs. Finally, we demonstrate thatvirus-specific lymphocytes contribute significantly tothe alloresponse in certain responder–stimulator HLAcombinations, underscoring the importance of T-cellcross-reactivity in alloimmunity. These results providea quantitative assessment of the roles of naı̈ve andmemory T-cell subsets in the normal human alloim-mune response and establish a platform for measuringT-cell alloreactivity pre- and posttransplantation.
Key words: Alloimmune response, human, im-munoassay, memory T cells
Received 19 March 2009, revised 11 May 2009 and ac-cepted for publication 14 May 2009
Introduction
Humans and experimental animals mount a vigorous im-mune response to tissues transplanted from geneticallydisparate individuals. This response, referred to as alloim-munity, is mediated principally by T lymphocytes that rec-ognize nonself-histocompatibility antigens (alloantigens)including the highly polymorphic human leukocyte anti-gens (HLA) encoded by the major histocompatibility com-plex (MHC) as well as multiple minor histocompatibilityantigens encoded by non-MHC genes (1).
A salient feature of T-cell alloimmunity is its extraordinarypotency which is attributed to the presence of a high fre-quency of host T cells (1–10% of the total T-cell reper-toire) that react to donor alloantigens (2–4). While immuneresponses to nominal antigens are dependent on a verysmall proportion of T cells that recognize foreign peptidespresented in the context of self-MHC molecules, allore-sponses are in addition generated by a much larger pop-ulation of T cells that recognize peptides complexed toallogeneic MHC on donor cells—a phenomenon known asdirect allorecognition (5,6). The high precursor frequencyof directly alloreactive T cells is likely the result of the in-herent bias of the prethymic T-cell repertoire toward MHCrecognition (7), and the presence of cross-reactive T cells inthe postthymic repertoire (6,8). The latter are self-MHC re-stricted mature T cells that are specific to microbial-derivedpeptides but cross-react with allogeneic MHC. Support forthe cross-reactivity hypothesis is provided by molecularevidence that cytotoxic T cells specific to common virusessuch as the Epstein-Barr Virus (EBV) exhibit strong allore-activity to particular HLA molecules (9,10). Since humansare exposed to a large array of pathogens beginning veryearly in life, it is inferred that any given individual harborsa significant number of pathogen-specific T cells that arepotentially alloreactive.
The development of ex vivo assays to identify, quantitateand analyze alloreactive T cells has been a longstandinginterest of human immunologists. It is hoped that such as-says will lead to better understanding of human alloimmu-nity and enable clinicians to predict allograft outcomes andindividualize immunosuppression safely. Earlier attemptsat studying the human alloimmune response have reliedon assays that gauge one parameter or one cell popula-tion at a time; for example, measuring T-cell proliferation inthe mixed lymphocyte reaction (MLR), IFNc production in
2057

Macedo et al.
the enzyme-linked immunosorbent spot (ELISPOT) assayor cytotoxicity in the limiting dilution analysis (11). Althoughthese assays have been instrumental in elucidating someof the fundamental features of human alloimmunity (12–15), several questions remain unanswered. Importantly, itis still unclear what the relative contributions of naı̈ve andmemory CD4+ and CD8+ T cells are, and which memoryT cell subsets play the most prominent role in the alloim-mune response.
In this manuscript, we established a reproducible, ex vivoflow cytometric multiparameter assay that allows simulta-neous measurement of proliferation, precursor frequencyand effector molecules of CD4+ and CD8+ T-cell subsetssorted from healthy volunteers, investigated the relativecontribution of the naı̈ve and memory T-cell pools to thealloimmune response, and directly tested whether virus-specific T cells exhibit alloreactivity.
Materials and Methods
Human subjectsTwenty-nine healthy volunteers were recruited following informed con-sent under an IRB-approved protocol at the University of Pittsburgh(IRB#00608014). The mean age ± SD of healthy volunteers was 39.8 ± 10.6years and included 12 males and 17 females. In addition, nine leukocyteconcentrates from healthy volunteers were purchased from the PittsburghBlood Bank.
Peripheral blood mononuclear cells (PBMC) isolationand cell sortingPBMC were isolated by density gradient centrifugation (16). To obtain CD3-depleted PBMC, magnetic cell separation was performed using CD3 mi-crobeads (Miltenyi-Biotech, Auburn, CA). For T-cell memory subset sepa-ration, CD3+ T cells were sorted by negative selection using a pan T-cellisolation kit II (Miltenyi-Biotech). These cells were further exposed to eitherCD45RA or CD45RO and subsequently to CD62L microbeads (Miltenyi-Biotech). Accordingly, naı̈ve (TN-CD45ROloCD62Lhi), effector mem-ory (TEM-CD45ROhiCD62Llo), terminally differentiated effector memory(TEMRA-CD45ROloCD62Llo) and central memory (TCM-CD45ROhiCD62Lhi)cell subsets were obtained. The purity of TN cells was >93% (<7% con-tamination with TEMRA cells), TEM cells >91% (<9% TCM contamination),TCM cells >89% (<11% TEM contamination) and TEMRA cells >83% (<17%TN cell contamination).
HLA typingMolecular HLA typing for class I (A and B loci) and class II (DR locus)was done on all participants in the Tissue Typing Laboratory, University ofPittsburgh Medical Center.
Media and reagentsRPMI-1640 (Cellgro, Manassas, VA) was supplemented with 2 mM L-glutamine, 10 mM HEPES, 100 IU/mL penicillin/streptomycin (Gemini Bio-products, West Sacramento, CA) and 5% normal human AB serum (NHS)(Nabi, Boca Raton, FL). Carboxyfluorescein succinimidyl ester (CFSE) waspurchased from Invitrogen (Eugene, OR) and PKH-26 kit from Sigma (St.Louis, MO). HLA-B08-restricted EBV peptide EBNA-3A (FLRGRAYGL) andHIV peptide (SLYNTVATL) were synthesized at the Peptide Synthesis Facility(University of Pittsburgh, Pittsburgh, PA). These peptides were employed at10 lg/mL to stimulate T cells in the functional assays. PE-HLA-B08 tetramer
incorporating EBNA-3A peptide was generated at the NIAID MHC-TetramerCore Facility (Emory University, Atlanta, GA) and used to detect EBV-specificCD8+ T cells by flow cytometry. Negative tetramer was purchased fromBeckman Coulter (Fullerton, CA). Tacrolimus was purchased from Sigma.
One-way MLRCFSE-labeled (2 lM) bulk PBMC or sorted memory T-cell subsets (17)used as responders were incubated in with c -irradiated allogeneic PKH-26-labeled (according to manufacture’s protocol) stimulator PBMC (1:1 ratio)for 5 days. To ascertain allogeneic cross-reactivity to an EBV epitope, CFSE-labeled bulk PBMC from HLA-B08+ EBV+ healthy subjects were incubatedwith c -irradiated allogeneic stimulator PBMC from HLA-B44+ or HLA-B44−
subjects.
Flow cytometric analysisProliferation of allo-activated CD4+ or CD8+ T cells was identified after cellsurface staining with mAbs anti-CD3-PeCy7, anti-CD8-PerCP-Cy5.5, anti-CD45RO-APC, anti-CD14-PB (BD Biosciences, San Jose, CA) and with fix-able dead cell stain kit-PB (Invitrogen) and by quantifying CFSE dilution. Todetect EBV-specific CD8+ T cells, staining with HLA-B08 EBNA-3A tetramerwas performed prior to other cell surface staining (18). For intracellular stain-ing, aliquots of surface stained and fixed cells were further permeabilizedand incubated with antigranzyme B-Alexa700 (BD) and antiperforin-APC(Biolegend, San Diego, CA). For IFNc -Alexa700 (BD) intracellular staining,cultured cells were re-stimulated in vitro for 4 h with either 4 lg/mL anti-CD3 (BD) and 2 lg/mL anti-CD28 (R&D, Minneapolis, MN) mAbs or withPKH26-labeled, CD3-depleted, c -irradiated allogeneic PBMC (ratio 1:1) inthe presence of Golgi-Plug (BD). Data acquisition was performed on a LSRII flow cytometer (BD) and data analysis was carried out using FlowJo (TreeStar, Ashland, OR) or ModFit LT R⃝ (Verity Software, USA).
Cytotoxicity assayAllostimulated T cells were tested in a standard 51Cr release assay. Astargets, autologous (HLA-B08+HLA-B44−) and allogeneic (HLA-B08−HLA-B44+ or HLA-B08−HLAB44−) PHA blasts were generated as previouslydescribed (9). 51Cr-labeled targets were incubated with 10 lg/mL HLA-B08EBV EBNA3A peptide (or negative control HIV peptide) (9). Effector:targetratios (100:1, 50:1, 25:1, 12.5:1) were plated in triplicates in 96-well plates(Costar Corning Inc, Corning, NY), and 51Cr release was measured in agamma counter (LKB, Helsinki, Finland). The percentage of specific lysiswas calculated using a previously published formula (19).
Statistical analysisMedians and means ± standard deviations (SD) were calculated and com-pared using the two-tail Student t-test. Results were considered statisticallysignificant if the p-value was ≤ 0.05.
Results
Assessment of alloantigen-induced T-cell proliferationby a flow cytometry-based MLRAlloantigen-induced T-cell proliferation has traditionallybeen measured by thymidine incorporation in the MLR.A shortcoming of this method is that it does not iden-tify which lymphocyte population or T-cell subset (CD4+
or CD8+) is proliferating or quantify the precursor frequen-cies of alloreactive CD4+ and CD8+ T cells unless tediouslimiting dilution assays are set up in parallel. We there-fore implemented a flow cytometry-based MLR methodto analyze the alloreactivity of human T cells. The results
2058 American Journal of Transplantation 2009; 9: 2057–2066

Memory T Cells in Alloimmunity
Figure 1: Identification and quantification of T-cell proliferation by flow cytometry-based one-way MLR. (A) Responder T cellswere identified at the end of the 5-day MLR by gating on CD3+ cells followed by exclusion of dead cells and PKH26+ stimulator cells.(B) Proliferation of the CD4+ and CD8+ populations was then quantified by CFSE dilution (% CFSElow cells) (C) and by histogram plots ofgenerations of divided cells as determined by ModFit-LT R⃝ software. (D) The specificity of T-cell proliferation in this assay was determinedby measuring CFSE dilution after co-culture with either autologous or allogeneic PBMC in the presence or absence of tacrolimus (TAC)(100 ng/mL). Results shown are one example of 41 independent responder–stimulator pairs. Suppression of T-cell proliferation bytacrolimus was demonstrated in 10 independent pairs. The absence of significant proliferative response to autologous cells was confirmedin 24 independent pairs.
of one representative experiment are shown in Figure 1to illustrate the analysis method. Gating strategy is shownin Figure 1A, while the CFSE dilution of the CD4+ andCD8+ T-cell subsets was analyzed separately to mea-sure their proliferation. Results are depicted either as dotplots (Figure 1B) or histograms (Figure 1C). As shown inFigure 1C, both the parent nonproliferating CD4+ and CD8+
T cells as well as 11 generations of dividing cells could beidentified using ModFit LT R⃝ software. The histogram plotsconsistently showed that the bulk of proliferated CD4+ andCD8+ T cells clustered in peaks representing, on the aver-age, cells that had divided between 4 and 10 times.
To test the specificity of the assay, CFSE-labeled respon-der PBMC were co-cultured with either autologous PBMCor with allogeneic PBMC in the presence of the T-cell im-munosuppressive drug tacrolimus (TAC). The results de-picted in Figure 1D show minimal proliferation in responseto autologous cells and significant inhibition of both CD4+
and CD8+ T-cell proliferation in response to allostimulationif TAC (100 ng/mL) was added to the MLR. These data indi-cate that proliferation measured by flow cytometry-basedMLR is both alloantigen and T-cell dependent.
CD4+ and CD8+ T cells proliferate at comparablelevels in response to allostimulation and containsimilar alloreactive precursor frequenciesTo characterize and quantify the normal human T-cell al-loimmune response, we obtained PBMC from 29 HLA-typed, healthy, nontransplanted volunteers and performedflow cytometry-based MLR on 41 different responder–stimulator pairs. In each case, the proportion (%) of eitherCD4+ or CD8+ T cells that had undergone at least one di-vision (second-generation cells or greater) was measured.The precision of this measurement was determined by per-forming five replicate assays on four responder–stimulatorpairs (data not shown) that yielded a low mean coefficientof variability of 14% (range = 6–23%), indicating that theassay is precise. As summarized in Figure 2A, overall CD4+
and CD8+ T-cell subsets proliferated equally in response toallostimulation. CD4+ T cells that had divided ranged be-tween 4% and 63% of total CD4+ T-cells (mean ± SD =19 ± 11%; median = 17%), and CD8+ T cell proliferationranged between 2% and 88% of total CD8+ T cells (18 ±15%, 15%). We further calculated the precursor frequencyof alloreactive T cells present in the CD4+ and CD8+ sub-set using ModFit LT R⃝. This software generates histograms
American Journal of Transplantation 2009; 9: 2057–2066 2059

Macedo et al.
Figure 2: Alloantigen-induced proliferative response and alloreactive precursor frequencies of human T cells. (A) T-cell proliferationin response to allogeneic PBMC was measured in the one-way MLR by CFSE dilution, and the proportion of CD4+ and CD8+ T cellsthat had proliferated was calculated as % of total CD4+ or CD8+ T cells, respectively. (B) Alloreactive precursor frequencies present inthe CD4+ and CD8+ populations were calculated by ModFit LT R⃝ software based on histogram plots similar to that shown in Figure 1C.Horizontal lines are of mean values (n = 41 responder–stimulator pairs).
based on CFSE intensity (Figure 1C) and applies deconvo-lution algorithms to calculate the precursor frequency ofcells that divided, reported as% of parent population. Asshown in Figure 2B, the overall CD4+ and CD8+ subsetscontained comparable frequencies of alloreactive T cells:3.9 ± 3.5% (range = 0.9–20.4%; median = 3%) and 2.5 ±3.7% (range = 0.1–25%; 1.6%), respectively. These dataunderscore the heterogeneity of the human alloimmuneresponse and are consistent with alloreactive T-cell precur-sor frequencies reported by others (2–4). In addition, ourresults clearly indicate that CD4+ and CD8+ populationsproliferate equally in response to allostimulation.
We then analyzed the effect of responder–stimulator HLAdisparities on CD4+ and CD8+ T-cell alloresponses. Allresponder–stimulator pairs had at least 3 antigen mis-matches except for one pair that had one antigen mismatchrestricted to the HLA-DR (class II) locus and another thathad one class I and one class II HLA mismatches. We alsoanalyzed the response of any given individual to multipleHLA-mismatched stimulators. The overall results suggestthat cumulative differences in HLA mismatches do not in-fluence T-cell proliferation in the one-way MLR when atleast one HLA-DR mismatch is present and underscorethe importance of the ‘quality’, rather than ‘quantity’, ofthe HLA mismatch in defining the strength of the alloim-mune response (Supporting Figure S1A and B).
Assessment of type-1 T-cell effector moleculeexpression using the flow cytometry-based MLRUnlike traditional immunological assays that measure oneparameter at a time, modern flow cytometry provides theopportunity to measure multiple parameters simultane-ously. Therefore, we employed the flow cytometry-basedMLR method to investigate whether T cells that prolif-erated in response to alloantigens also acquired effec-tor molecules necessary for their function. We chose tomeasure IFNc , granzyme B and perforin because these
molecules were showed to contribute to allograft destruc-tion (15,20). A representative experiment that illustratesIFNc measurement relative to cell proliferation (CFSE dilu-tion) is shown in Figure 3A for both CD4+ and CD8+ T cells.Allogeneic re-stimulation induced IFNc production only inthose responder cells that had already undergone multi-ple proliferation cycles (Figure 3A), indicating that they haddifferentiated into effector T lymphocytes. Cumulative anal-ysis of the data revealed that the CD8+ T-cell subset had asignificantly higher proportion of IFNc -producing cells thanthe CD4+ subset following either allospecific or polyclonalre-stimulation (Figure 3B). Moreover, IFNc production didnot correlate with the magnitude of proliferation of eitherCD4+ or CD8+ T cells (Figure 3C). The latter result indi-cates that measuring proliferation alone, as has been tradi-tionally done in MLR assays, does not guarantee accurateinferences about T-cell effector functions.
Intracellular granzyme B and perforin were measured inresponder CD4+ and CD8+ T cells at the end of the 5-day MLR without further re-stimulation (Figure 4A). Aswith IFNc , only T cells that had proliferated in the MLRexpressed granzyme B and/or perforin. Cumulative analy-sis of the data demonstrated that the proportion of cellsthat acquired either granzyme B or both granzyme B andperforin expression was significantly higher among CD8+
than CD4+ T cells (Figure 4B). We also observed an in-verse correlation between cell proliferation and granzymeB/perforin double expression in the CD8+, but not in theCD4+ T-cell subset (Figure 4C). These results further un-derscore the advantage of the flow cytometry-based MLRover other assays in that it simultaneously measures pro-liferation and functional attributes of alloreactive T cells.They also clearly illustrate that a strong proliferative T-cellresponse in the MLR does not necessarily imply a po-tent effector response. This may explain why MLR meth-ods that only measure proliferation have failed to correlatewith or predict clinical outcomes in transplant recipients(21–23).
2060 American Journal of Transplantation 2009; 9: 2057–2066

Memory T Cells in Alloimmunity
Figure 3: IFNc production by alloantigen-activated T cells. (A) IFNc -producing CD4+ and CD8+ T cells were quantified by flowcytometry at the end of the 5-day one-way MLR after 4 h of restimulation with anti-CD3 plus anti-CD28, CD3-depleted allogeneic PBMCor media alone. Number displayed in the left upper quadrant of each dot plot represents the proportion (%) of proliferating (CFSElow) CD4+
or CD8+ T cells that produced IFNc . Results are of one representative experiment. (B) Proportion of IFNc -producing cells, measuredas in (A), is greater in the CD8+ than that in CD4+ T-cell population (n = 11). (C) Lack of correlation between cell proliferation and IFNcproduction in either the CD4+ or CD8+ T-cell population. Each symbol represents a single responder–stimulator pair (n = 11).
Relative contribution of naı̈ve and memory T-cellpopulations to the normal human alloimmuneresponseLimiting dilution assays performed more than 15 yearsago provided the first evidence that alloreactive lympho-cytes are not restricted to the naı̈ve T cell pool but in-clude an equal number of memory T cells (12,13). Sincethese publications appeared, newer markers that differen-tiate naı̈ve from memory T cells have been developed, andevidence that distinct memory T-cell subpopulations existin humans has emerged: the TCM subset lacks immedi-ate effector function and homes to secondary lymphoidorgans where they proliferate and differentiate into effec-tors upon proper antigenic re-challenge. In contrast, TEM
cells are found mostly in the peripheral inflamed tissuesand readily secrete effector molecules in response to anti-geneic stimulation. The TEMRA subset has similar func-tional properties as TEM; however, they are more potenteffectors and are more susceptible to death (24,25). Wetherefore purified CD3+ cells from leukapheresis productsof nine healthy subjects and separated them into TN, TCM,TEM and TEMRA (24,25). The phenotype and frequency ofeach subset are shown in Figure 5A. The T-cell subsetswere then tested individually in the flow cytometry-based
MLR. The results shown in Table 1 indicate that alloreac-tive T-cell precursors are present in all four populations withthe naı̈ve and memory subsets contributing equally. WhileTN and TCM cells proliferated the most in response to al-lostimulation, the TEM and TEMRA subpopulations trendedtoward higher proportion of IFNc , and significant highergranzyme B/perforin expressing CD8+ T cells (Figure 5B).CD4+ T cell proliferation was comparable to that of CD8+
T cells, but their lytic molecules expression was signifi-cantly lower (Figure 5B). These results indicate that healthysubjects harbor alloreactive T cells within both the naı̈veand memory lymphocyte pools, with TEM and TEMRA sub-populations containing alloreactive T cells that readily ex-press IFNc and lytic molecules upon allostimulation.
Contribution of virus-specific CD8+ T cells to thehuman T-cell alloimmune responseThe presence of IFNc - and granzyme B/perforin-producingalloreactive CD8+ T cells in the memory pools of all healthysubjects studied points to the possibility that these arecross-reactive T cells generated by prior exposure to com-mon pathogens. To test directly the contribution of virus-specific CD8+ T cells to the human alloimmune response,
American Journal of Transplantation 2009; 9: 2057–2066 2061

Macedo et al.
Figure 4: Granzyme B and perforin expression by alloantigen-activated T cells. (A) Granzyme B and/or perforin-expressing CD4+
and CD8+ T cells were quantified by flow cytometry at the end of the 5-day one-way MLR. Granzyme B and perforin expression areshown for both undivided (CFSEhigh) and proliferating (CFSElow) cells. Numbers displayed in the quadrants of each dot plot represent theproportion (%) of CD4+ or CD8+ T cells expressing granzyme B, perforin, both or neither. (B) Overall % of granzyme B, perforin or bothgranzyme B and perforin-expressing cells (n = 14). (C) Correlation between cell proliferation and granzyme B/perforin double expressionin the CD4+and CD8+ T-cell population. Each symbol represents a single responder–stimulator pair (n = 14).
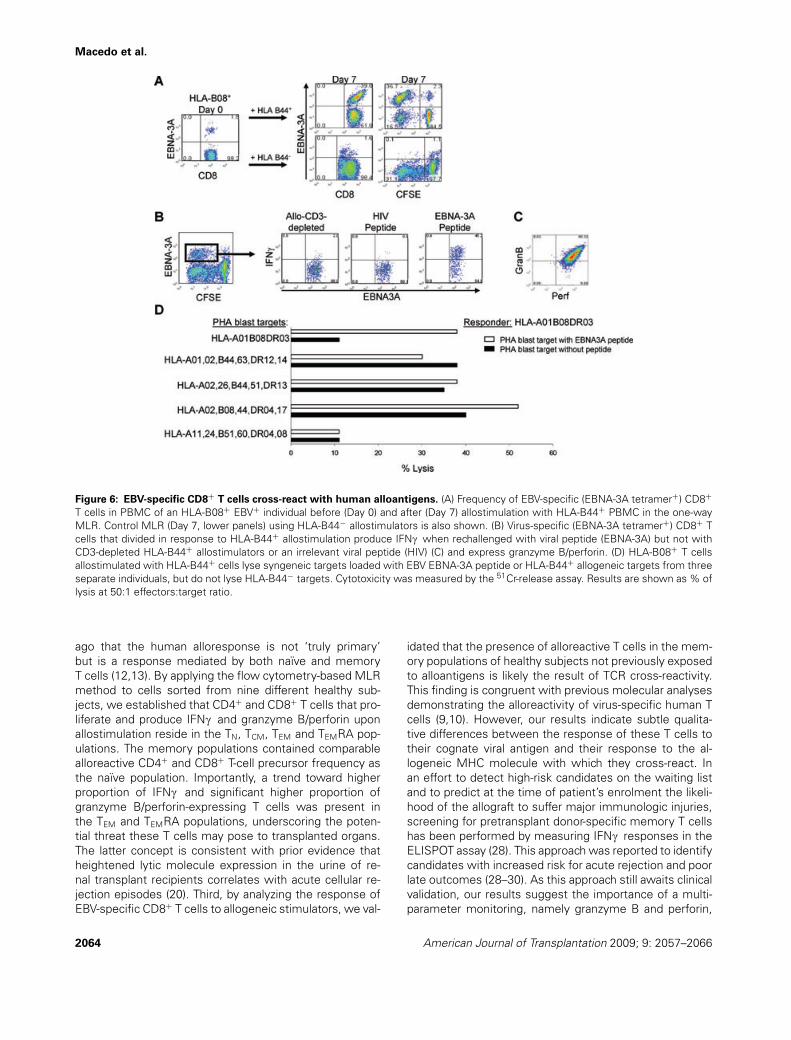
we took advantage of tetramer technology to track Ag-specific CD8+ T cells in the flow cytometry-based MLR as-say. Burrows et al. had demonstrated that HLA-B08+/EBV+
adults express T-cell receptors specific to the EBV-epitopeEBNA-3A that cross-react with HLA-B44 (9). Accordingly,we investigated the response of PBMC from two HLA-B08+/EBV+ healthy controls against irradiated PBMC fromHLA-B44+ mismatched individuals or HLA-B44− allogeneiccontrols. EBNA-3A tetramer+ cells constituted 1.8% of cir-culating CD8+ T cells (96% displaying memory phenotype)at baseline (Figure 6A). Their frequency increased dramat-ically to 39% 7 days after allostimulation with HLA-B44+
PBMC but remained at 1.6% when allostimulated withHLA-B44− PBMC. Importantly, EBNA-3A tetramer+ T cellsrepresented approximately 70% of CD8+ T cells that haddivided in response to HLA-B44+ allogeneic cells. Inter-estingly, a significant proportion of these cross-reactiveEBNA-3A tetramer+ CD8+ T cells produced IFNc in re-sponse to the anti-CD3/anti-CD28 polyclonal stimulation(data not shown) and to the EBV-peptide (Figure 6B), butfailed to do so in response to HLA-B44+ allostimulation(Figure 6B). These cells also expressed granzyme B andperforin (Figure 6C), and were cytotoxic against HLA-B08+
PHA blasts that carry the EBNA-3A peptide and against al-logeneic PHA blasts that share HLA-B44 with the stimula-tor PBMC, but not against HLA-B08−HLA-B44− PHA blasts(Figure 6D). Similar EBV-specific CD8+ T cell proliferation,cytotoxicity and IFNc expression were observed in a sec-ond HLA-B08+/EBV+ subject (Supporting Figure S2). Theseresults demonstrate that virus-specific T cells contributeto the human alloresponse if the appropriate responder–stimulator HLA combination is present. The response toalloantigens by these cross-reactive CD8+ T cells, how-ever, may be qualitatively different from their response tothe viral epitope when cytotoxicity and IFNc productionare measured.
Discussion
T-cell alloimmunity is considered one of the dominant fac-tors mediating graft injury after transplantation. However,the precise contribution of naı̈ve versus memory T-cell sub-sets to human alloimmunity has remained elusive. Here weestablished an accurate and specific flow cytometry-basedex vivo assay that measures simultaneously proliferation,
2062 American Journal of Transplantation 2009; 9: 2057–2066

Memory T Cells in Alloimmunity
Figure 5: Contribution ofnaı̈ve versus memory T-cell subsets to human al-loreactivity. (A) Naı̈ve andmemory T-cell populationspresent in the peripheralblood of nine healthy sub-jects were identified andquantitated (% of total Tcells) by flow cytometry.TN = naı̈ve; TEM = effec-tor memory; TCM = cen-tral memory; and TEMRA =terminally differentiated ef-fector memory. (B) Prolifer-ation, IFNc production andgranzyme B/perforin doubleexpression of CD4+ andCD8+ purified naı̈ve andmemory T-cell subsets weredetermined in the 5-dayone-way MLR. IFNc datashown is after 4 h restimula-tion with anti-CD3 plus anti-CD28 mAbs. ∗One-tail Stu-dent’s t-test analysis.
precursor frequencies and effector molecule (IFNc andgranzyme B/perforin) expression of alloreactive CD4+ andCD8+ T cells present in the peripheral blood of human sub-jects. We then utilized this assay to assess the contribu-tions of naı̈ve and memory T-cell subsets to the alloimmuneresponse.
Our results provide new insights into the human T-cell al-loimmune response. First, we found that CD4+ and CD8+
T cells proliferate equally in response to allostimulationand contain comparable alloreactive precursor frequenciesdetermined by CFSE dilution (approximately 4%), not de-tected previously by thymidine incorporation MLR. In ad-dition, allostimulated CD4+ T cells acquired effector medi-ators, including lytic molecules, albeit at lower levels thanCD8+ T cells. Prior attempts at quantitating alloreactiveCD4+ T-cell precursors by limiting dilution assays and IL-2
measurement resulted in much lower frequencies (<0.2%)(26,27), suggesting that alloreactivity of the CD4+ com-partment may have been underestimated. Second, our re-sults extended the observation made more than 15 years
Table 1: Precursor frequency of alloreactive T cells in the naı̈veand memory populations
Precursor frequency (%)
CD4+A CD8+A
TEM 4.6 ± 1.4 (n = 7) 3.1 ± 1.5 (n = 8)TCM 4.9 ± 2.2 (n = 5) 5.1 ± 2.3 (n = 7)TEMRA 4.4 ± 3.0 (n = 2) 3.2 ± 1.1 (n = 4)TN 3.7 ± 2.6 (n = 6) 4.0 ± 1.9 (n = 6)AMean ± SD; TEM = effector memory; TCM = central memory;TEMRA = terminally differentiated effector memory; TN = naı̈ve.
American Journal of Transplantation 2009; 9: 2057–2066 2063
Macedo et al.
Figure 6: EBV-specific CD8+ T cells cross-react with human alloantigens. (A) Frequency of EBV-specific (EBNA-3A tetramer+) CD8+
T cells in PBMC of an HLA-B08+ EBV+ individual before (Day 0) and after (Day 7) allostimulation with HLA-B44+ PBMC in the one-wayMLR. Control MLR (Day 7, lower panels) using HLA-B44− allostimulators is also shown. (B) Virus-specific (EBNA-3A tetramer+) CD8+ Tcells that divided in response to HLA-B44+ allostimulation produce IFNc when rechallenged with viral peptide (EBNA-3A) but not withCD3-depleted HLA-B44+ allostimulators or an irrelevant viral peptide (HIV) (C) and express granzyme B/perforin. (D) HLA-B08+ T cellsallostimulated with HLA-B44+ cells lyse syngeneic targets loaded with EBV EBNA-3A peptide or HLA-B44+ allogeneic targets from threeseparate individuals, but do not lyse HLA-B44− targets. Cytotoxicity was measured by the 51Cr-release assay. Results are shown as % oflysis at 50:1 effectors:target ratio.
ago that the human alloresponse is not ‘truly primary’but is a response mediated by both naı̈ve and memoryT cells (12,13). By applying the flow cytometry-based MLRmethod to cells sorted from nine different healthy sub-jects, we established that CD4+ and CD8+ T cells that pro-liferate and produce IFNc and granzyme B/perforin uponallostimulation reside in the TN, TCM, TEM and TEMRA pop-ulations. The memory populations contained comparablealloreactive CD4+ and CD8+ T-cell precursor frequency asthe naı̈ve population. Importantly, a trend toward higherproportion of IFNc and significant higher proportion ofgranzyme B/perforin-expressing T cells was present inthe TEM and TEMRA populations, underscoring the poten-tial threat these T cells may pose to transplanted organs.The latter concept is consistent with prior evidence thatheightened lytic molecule expression in the urine of re-nal transplant recipients correlates with acute cellular re-jection episodes (20). Third, by analyzing the response ofEBV-specific CD8+ T cells to allogeneic stimulators, we val-
idated that the presence of alloreactive T cells in the mem-ory populations of healthy subjects not previously exposedto alloantigens is likely the result of TCR cross-reactivity.This finding is congruent with previous molecular analysesdemonstrating the alloreactivity of virus-specific human Tcells (9,10). However, our results indicate subtle qualita-tive differences between the response of these T cells totheir cognate viral antigen and their response to the al-logeneic MHC molecule with which they cross-react. Inan effort to detect high-risk candidates on the waiting listand to predict at the time of patient’s enrolment the likeli-hood of the allograft to suffer major immunologic injuries,screening for pretransplant donor-specific memory T cellshas been performed by measuring IFNc responses in theELISPOT assay (28). This approach was reported to identifycandidates with increased risk for acute rejection and poorlate outcomes (28–30). As this approach still awaits clinicalvalidation, our results suggest the importance of a multi-parameter monitoring, namely granzyme B and perforin,
2064 American Journal of Transplantation 2009; 9: 2057–2066

Memory T Cells in Alloimmunity
proliferation and precursor frequencies in addition to IFNc ,as screening tools for detecting T-cell alloreactivity and,therefore, as a strategy to optimize pretransplant risk as-sessment.
Flow cytometry-based MLR assays have been previouslyemployed to measure alloimmune responses in trans-planted patients (22,23,31). Tanaka et al. quantified CD4+
and CD8+ T cell proliferation in living-donor liver transplantrecipients by this method in response to either donor orthird-party allogeneic cells, but no significant correlationbetween proliferation and acute rejection could be found(22). Precursor frequencies and cytokine or lytic moleculeexpression were not measured in this study. Using a simi-lar assay, Kreijveld et al. calculated CD4+ and CD8+ allore-active T-cell precursor frequency in renal transplant recip-ients who had been withdrawn from tacrolimus therapy.They found that frequencies were stable over time anddid not correlate with the occurrence of acute rejection.IFNc and granzyme B/perforin expression by alloreactiveT cells were not determined (23). These reports, there-fore, suggest that measuring T-cell proliferation alone doesnot provide an adequate reflection of human alloimmunity.Our analysis of PBMC from healthy subjects provided di-rect proof that proliferation does not correlate with thefunctional potential of alloreactive CD4+ or CD8+ T cells(Figures 3C and 4C). In fact, we found an inverse rela-tion between granzyme B/perforin production and CD8+
T cell proliferation (Figure 4C). Therefore, we propose thatsimultaneous monitoring of donor-specific alloreactivity us-ing multiple functional parameters is a superior approachto single parameter measurements and may allow an ac-curate prediction of graft outcomes posttransplantation.In addition, this multiparameter analysis could be slightlymodified to use allogeneic PBMC lysates instead of allo-geneic irradiated PBMC stimulation to also quantify theindirect pathway of allorecognition.
In summary, the data presented in this manuscript providedirect evidence for the importance of memory T cells inthe normal human alloimmune response. Specifically, wehave identified the CD8+ TEM and TEMRA subsets as likelythreats to transplanted organs because of their heightenedexpression of effector molecules upon stimulation. Moni-toring these T-cell subsets in pre- and postsolid organ orbone marrow transplantation may prove useful for predict-ing adverse immunological events such as allograft rejec-tion and graft-versus-host disease.
Acknowledgments
We thank Dr Igor Dvorchik, PhD, and Dr Hongmei Shen, PhD, for theirvaluable advice with the establishment and validation of the flow cytometry-based MLR assay. We also acknowledge the NIH Tetramer Facility (EmoryUniversity, Atlanta, GA) for generating the PE-HLA-B08 EBNA-3A tetramer.This work was supported in part by NIH grant AI049466 (FGL).
References
1. Afzali B, Lechler RI, Hernandez-Fuentes MP. Allorecognition andthe alloresponse: Clinical implications. Tissue Antigens 2007; 69:545–556.
2. Lindahl KF, Wilson DB. Histocompatibility antigen-activated cyto-toxic T lymphocytes: II. Estimates of the frequency and specificityof precursors. J Exp Med 1977; 145: 508–522.
3. Suchin EJ, Langmuir PB, Palmer E, Sayegh MH, Wells AD, TurkaLA. Quantifying the frequency of alloreactive T cells in vivo: Newanswers to an old question. J Immunol 2001; 166: 973–981.
4. Baker RJ, Hernandez-Fuentes MP, Brookes PA, Chaudhry AN,Lechler RI. The role of the allograft in the induction of donor-specificT cell hyporesponsiveness. Transplantation 2001; 72: 480–485.
5. Colf LA, Bankovich AJ, Hanick NA et al. How a single T cell receptorrecognizes both self and foreign MHC. Cell 2007; 129: 135–146.
6. Ely LK, Burrows SR, Purcell AW, Rossjohn J, McCluskey J. T-cellsbehaving badly: Structural insights into alloreactivity and autoim-munity. Curr Opin Immunol 2008; 20: 575–580.
7. Zerrahn J, Held W, Raulet DH. The MHC reactivity of the T cellrepertoire prior to positive and negative selection. Cell 1997; 88:627–636.
8. Archbold JK, Macdonald WA, Burrows SR, Rossjohn J, McCluskeyJ. T-cell allorecognition: A case of mistaken identity or deja vu?Trends Immunol 2008; 29: 220–226.
9. Burrows SR, Khanna R, Burrows JM, Moss DJ. An alloresponsein humans is dominated by cytotoxic T lymphocytes (CTL) cross-reactive with a single Epstein-Barr virus CTL epitope: Implicationsfor graft-versus-host disease. J Exp Med 1994; 179: 1155–1161.
10. Archbold JK, Macdonald WA, Gras S et al. Natural micropolymor-phism in human leukocyte antigens provides a basis for geneticcontrol of antigen recognition. J Exp Med 2009; 206: 209–219.
11. Dinavahi R, Heeger PS. T-cell immune monitoring in organ trans-plantation. Curr Opin Organ Transplant 2008; 13: 419–424.
12. Merkenschlager M, Terry L, Edwards R, Beverley PC. Limitingdilution analysis of proliferative responses in human lymphocytepopulations defined by the monoclonal antibody UCHL1: Implica-tions for differential CD45 expression in T cell memory formation.Eur J Immunol 1988; 18: 1653–1661.
13. Lombardi G, Sidhu S, Daly M, Batchelor JR, Makgoba W, LechlerRI. Are primary alloresponses truly primary? Int Immunol 1990; 2:9–13.
14. Poggio ED, Augustine JJ, Clemente M et al. Pretransplant cellularalloimmunity as assessed by a panel of reactive T cells assaycorrelates with acute renal graft rejection. Transplantation 2007;83: 847–852.
15. Hricik DE, Rodriguez V, Riley J et al. Enzyme linked immunosor-bent spot (ELISPOT) assay for interferon-gamma independentlypredicts renal function in kidney transplant recipients. Am J Trans-plant 2003; 3: 878–884.
16. Boyum A, Lovhaug D, Tresland L, Nordlie EM. Separation of leuco-cytes: Improved cell purity by fine adjustments of gradient mediumdensity and osmolality. Scand J Immunol 1991; 34: 697–712.
17. Popescu I, Macedo C, Abu-Elmagd K et al. EBV-specific CD8+ Tcell reactivation in transplant patients results in expansion of CD8+type-1 regulatory T cells. Am J Transplant 2007; 7: 1215–1223.
18. Macedo C, Donnenberg A, Popescu I et al. EBV-specific memoryCD8+ T cell phenotype and function in stable solid organ trans-plant patients. Transplant Immunol 2005; 14: 109–116.
19. Dal Col RH, Zeevi A, Rabinowich H, Herlan DB, Yousem SA, GriffithBP. Donor-specific cytotoxicity testing: An advance in detectingpulmonary allograft rejection. Ann Thorac Surg 1990; 49: 754–758.
American Journal of Transplantation 2009; 9: 2057–2066 2065

Macedo et al.
20. Li B, Hartono C, Ding R et al. Noninvasive diagnosis of renal-allograft rejection by measurement of messenger RNA for per-forin and granzyme B in urine. New Engl J Med 2001; 344: 947–954.
21. Segall M, Noreen H, Edwins L, Haake R, Shu XO, Kersey J. Lack ofcorrelation of MLC reactivity with acute graft-versus-host diseaseand mortality in unrelated donor bone marrow transplantation.Hum Immunol 1996; 49: 49–55.
22. Tanaka Y, Ohdan H, Onoe T et al. Low incidence of acute rejectionafter living-donor liver transplantation: Immunologic analyses bymixed lymphocyte reaction using a carboxyfluorescein diacetatesuccinimidyl ester labeling technique. Transplantation 2005; 79:1262–1267.
23. Kreijveld E, Koenen HJ, van Cranenbroek B, van Rijssen E, JoostenI, Hilbrands LB. Immunological monitoring of renal transplant re-cipients to predict acute allograft rejection following the discontin-uation of tacrolimus. PLoS ONE 2008; 3: e2711.
24. Sallusto F, Lenig D, Forster R, Lipp M, Lanzavecchia A. Two sub-sets of memory T lymphocytes with distinct homing potentialsand effector functions. Nature 1999; 401: 708–712.
25. Appay V, van Lier RA, Sallusto F, Roederer M. Phenotype andfunction of human T lymphocyte subsets: Consensus and issues.Cytometry A 2008; 73: 975–983.
26. Orosz CG, Adams PW, Ferguson RM. Frequency of humanalloantigen-reactive T lymphocytes: II. Method for limiting dilutionanalysis of alloantigen-reactive helper T cells in human peripheralblood. Transplantation 1987; 43: 718–724.
27. Young NT, Roelen DL, Dallman MJ, Wood KJ, Morris PJ, Welsh KI.Enumeration of human alloreactive helper T lymphocyte precursorfrequencies by limiting dilution analysis of interleukin-2 production.J Immunol Methods 1996; 195: 33–41.
28. Augustine JJ, Siu DS, Clemente MJ, Schulak JA, Heeger PS, Hri-cik DE. Pre-transplant IFN-gamma ELISPOTs are associated with
post-transplant renal function in African American renal transplantrecipients. Am J Transplant 2005; 5: 1971–1975.
29. Poggio ED, Roddy M, Riley J et al. Analysis of immune markers inhuman cardiac allograft recipients and association with coronaryartery vasculopathy. J Heart Lung Transplant 2005; 24: 1606–1613.
30. Najafian N, Salama AD, Fedoseyeva EV, Benichou G, Sayegh MH.Enzyme-linked immunosorbent spot assay analysis of peripheralblood lymphocyte reactivity to donor HLA-DR peptides: potentialnovel assay for prediction of outcomes for renal transplant recipi-ents. J Am Soc Nephrol 2002; 13: 252–259.
31. Bloom DD, Hu H, Fechner JH, Knechtle SJ. T-lymphocyte allore-sponses of Campath-1H-treated kidney transplant patients. Trans-plantation 2006; 81: 81–87.
Supporting Information
The following supporting information is available for thisarticle online:
Figure S1: Influence of HLA mismatch on alloantigen-induced T-cell proliferation.
Figure S2: 56 EBV-specific CD8+ T cells cross-react withhuman alloantigens.
Please note: Wiley-Blackwell are not responsible for thecontent or functionality of any supporting materials sup-plied by the authors. Any queries (other than missing mate-rial) should be directed to the corresponding author for thearticle.
2066 American Journal of Transplantation 2009; 9: 2057–2066




















