
Conisbrough_with figures
64
1 The Conisbrough Estate and the southern boundary of Northumbria. Environmental and archaeological evidence from a late sixth/early seventh century structure and a later deer park boundary at Conisbrough, South Yorkshire Paul C. Buckland, David Hey, Richard O’Neill Ian Tyers Abstract Excavations close to the Anglo-Saxon church at Conisbrough, South Yorkshire, UK, revealed a plank-walled construction dated by dendrochronology to the late sixth or early seventh century. It is suggested that this formed part of a stock pond for fish, associated with an elite residence to which a partly surviving Anglo-Saxon church incorporating Northumbrian features is related. Environmental evidence shows a neglected wood pasture landscape associated with the infilling of a ditch which cuts through the structure. This is likely to be related to a deer park established shortly after the Norman Conquest and associated with the castle. Introduction Conisbrough, south Yorkshire (NGR SK512988), lies south of the river Don, 15 km west of Doncaster, at 64m OD, occupying the northeast end of a promontory overlooking the confluence of the rivers Don and Dearne (figs. 1 and 2), at the point where the Don cuts north-eastwards through the Magnesian Limestone cuesta. The village lies at the point where a prehistoric and Roman trackway descends the limestone escarpment to a crossing of the Don at Strafforth Sands and close to the place where the west – east Roman road from the fort at Templeborough near Rotherham to Doncaster crosses this route. At the end of the spur, on a small outlier of limestone surrounded by Coal Measure shales, lies the buttressed cylindrical keep of the Norman castle and its bailey. Excavations on the castle site (Johnson 1980) have failed to produce any evidence for pre-Norman
Transcript of Conisbrough_with figures

1
The Conisbrough Estate and the southern boundary of Northumbria.
Environmental and archaeological evidence from a late sixth/early seventh century
structure and a later deer park boundary at Conisbrough, South Yorkshire
Paul C. Buckland,
David Hey,
Richard O’Neill
Ian Tyers
Abstract
Excavations close to the Anglo-Saxon church at Conisbrough, South Yorkshire, UK,
revealed a plank-walled construction dated by dendrochronology to the late sixth or
early seventh century. It is suggested that this formed part of a stock pond for fish,
associated with an elite residence to which a partly surviving Anglo-Saxon church
incorporating Northumbrian features is related. Environmental evidence shows a
neglected wood pasture landscape associated with the infilling of a ditch which cuts
through the structure. This is likely to be related to a deer park established shortly after
the Norman Conquest and associated with the castle.
Introduction
Conisbrough, south Yorkshire (NGR SK512988), lies south of the river Don, 15 km west
of Doncaster, at 64m OD, occupying the northeast end of a promontory overlooking the
confluence of the rivers Don and Dearne (figs. 1 and 2), at the point where the Don cuts
north-eastwards through the Magnesian Limestone cuesta. The village lies at the point
where a prehistoric and Roman trackway descends the limestone escarpment to a crossing
of the Don at Strafforth Sands and close to the place where the west – east Roman road
from the fort at Templeborough near Rotherham to Doncaster crosses this route. At the
end of the spur, on a small outlier of limestone surrounded by Coal Measure shales, lies
the buttressed cylindrical keep of the Norman castle and its bailey. Excavations on the
castle site (Johnson 1980) have failed to produce any evidence for pre-Norman

2
occupation, although the depth of later accumulation within the bailey precluded detailed
examination of the earliest deposits. It seems probable therefore that the ‘king’s burgh’
implied by the place name, first referred to in the will of the Mercian Wulfric Spott in
1002-4 (Whitelock 1979, no. 125, 586-589; Sawyer 1979, no. 3) lay elsewhere. The
parish church, dedicated to St Peter, lies 0.5km to the southwest of the castle, surrounded
by the modern village. Externally the present building is largely Perpendicular, but this
encapsulates an Anglo-Saxon nave, which Ryder (1980; 44-49; 1982, 45) suggests may
belong to the eighth century. Hey (1979, 32-33; 1986, 14-18), following Glanville Jones’
(1965, 79-81; 1975, 3-27) model of Anglo-Saxon multiple estates, has argued that St
Peter’s at Conisbrough was the minster church for an extended area south of the Don
from Hatfield and Tudworth, to the northeast of Doncaster, towards the borders of
Hallamshire in the southwest.
At the time of Domesday Book, the fee of Conisbrough was the most considerable estate
in south Yorkshire. It had belonged to Harold Godwinsson before the Conquest and was
an early grant to one of the Conqueror’s most trusted barons, William de Warenne. The
unusual form of its entry in Domesday Book, with over twenty sokelands, suggests that
Conisbrough may have been a special liberty or franchise, free from the usual obligations
to the Crown (Dalton 1994, 33-34). Within the fee, 108 plough-teams and about 300
peasants were recorded in 28 south Yorkshire townships, most of which lay south of the
river Don and which stretched as far as Yorkshire’s southern boundary (Faull and Stinson
1986).
Several clues indicate that in previous centuries this estate had belonged to the kings of
Northumbria on their border with Mercia. First, the place-name was recorded as
Cunugesburh in 1002-4 in the will of Wulfric Spott, the only document that survives for
south Yorkshire (Whitelock 1979, no. 125) before the Norman Conquest. Its etymology
is ‘the king’s stronghold’, the same meaning as Konigsberg (modern Kaliningrad) in
Russia and the two Konigsbergs in Germany (Smith 1961). Second, Conisbrough
attracted the attention of medieval chroniclers. Geoffrey of Monmouth wrote of oppidum
Caerconan quod nunc Cunungeburg appellatur and claimed that burge Conane was the

3
tomb of Hengist. In the thirteenth century Peter Langtoft noted that King Egbert and his
men went to Burh-Konan at Whitsuntide (Hunter 1828, 1, 197-215). We do not know
whether these myths had any kernel of truth, but they attest to Conisbrough’s presumed
past importance. Third, the church of St Peter, Conisbrough, is a large Anglo-Saxon
building, whose substantial nave may date from the eighth century. Its size and design
suggest a major minster church in the Northumbrian tradition (Ryder 1980), a royal
foundation for the southern part of the kingdom, beyond the river Don. The medieval
parish that it served extended to Yorkshire’s southern boundary at Harthill and across the
eastern lowlands beyond Doncaster to Hatfield, Thorne and Fishlake. In the late eleventh
century, when William, second Earl of Warenne, granted Conisbrough and its dependent
churches to the Cluniac priory that his father had founded at Lewes, these churches each
lay within the fee (Hunter 1828, 1, 197-215); three of them are recorded in Domesday
Book. Fourth, Conisbrough overlooks an ancient crossing of the river Don at Strafforth
Sands (the meeting place of Strafforth wapentake), where a prehistoric and Roman route,
evident both on aerial photographs and in later parish boundaries (cf. Hey 2003, 17, and
unpublished aerial photograph of Barnburgh Cliff (NGR SE 499038) in Doncaster
Museum collection), ran north-south along the crest of the Magnesian Limestone belt.
Known variously as Packman Lane or Ricknield Street, by late Anglo-Scandinavian
times this highway separated the fee of Conisbrough from the smaller lordship of
Laughton-en-le-Morthen, which the Normans placed within their new honour of Tickhill
(Hunter 1828, 1, 197-215). The strategic importance of Conisbrough was enhanced by
the east-west Roman road that linked the forts at Doncaster to Templeborough and which
must have crossed the north-south route to the south of the church.
A Roman villa has been identified to the south of this junction, within the medieval
hunting area that is still known as Conisbrough Parks (Malcolm Grant and Ian
Macdonald, pers. comm. 2009). As late as 1848, Conisbrough Parks formed a distinct
estate of 1,337 acres that had a separate Tithe Award (Borthwick TA348S). Only three
farms - Conisbrough Lodge in the centre, Birk Lodge in the south-west, and Parks Farm
in the north-east - were named, with two unnamed farms on the northern and southern
boundaries. The boundary (fig. 3), which followed streams wherever possible, appears to

4
pre-date the creation of the deer park, and it is possible that the Norman deer park was
coterminous with apparent boundaries of a Roman estate. The park boundary comes close
to three settlements and obviously prevented their expansion. Clifton, on the eastern
boundary, was recorded in Domesday Book; the etymology of Firsby to the west is
‘farmstead of the Frisians’; and Micklebring to the south has a topographical name,
meaning ‘the great slope’, that is usually considered to be early (Smith 1961). The
boundaries of both, on the east and west sides of the park respectively, appear to have
been cut out of a pre-existing more regular shaped estate, the later deer park, and on the
archaeological evidence presented here, the park once extended further north, having
been truncated by the development of Conisbrough between castle and deer park (figs. 1
and 3). Whether or not the original park estate boundaries extend back to a Roman estate
associated with a known villa, they appear to have been well-established before the
Norman Conquest.
Historians have long thought that, taken together, these clues point to an important royal
estate on the southern border of Northumbria, but it has not been possible to date its
foundation. The translation of the place name as ‘king’s fort’ goes back at least to the
early nineteenth century (Wainwright 1829, vi), whilst Jones (1975) points out the
similarity with villa regia’. The interpretation is also discussed by Buckland, Magilton
and Hayfield (1986, 20-23) in the introduction to the monograph on the excavations in
Doncaster. The dendrochronological and radiocarbon evidence considered here indicates
that a major structure there may date from early in the reign of Aethelfrith (~ AD 590-
615), the king who united Bernicia and Deira to form this kingdom, and this may be
interpreted as evidence for a villa regia, in the area surrounding the later Anglo-Saxon
church. The evidence is, however, insufficiently precise to preclude a structure belonging
to the last years of a preceding British elite site.
Wellgate
St Peter’s church, Conisbrough, lies at the northeast end of an outcrop of Lower
Magnesian Limestone close to the contact with the underlying impermeable Coal

5
Measure shales, and this throws out a series of springs. At Wellgate (fig. 4), north of the
church, a small rectangular well house, probably of late medieval origin, has been
constructed over one of these to cover a water source, which may to relate to supply to
the castle (Miller 1804, 268). The ground slopes steeply to the north and east down to the
floodplain of the Don, and were it not for the large keep of the Norman castle, the church
would dominate the landscape.
The modern village of Conisbrough (figs. 1 and 2) lies immediately west and north of the
church. In 2002, a group of 1960s flats, behind nineteenth century properties fronting
onto Church Street and along Wellgate, was demolished to make way for new housing
development. In advance of construction works, and following recommendations made
by the South Yorkshire Archaeology Service, an archaeological evaluation (Trenches 1-
15) was carried out by ARCUS (formerly Archaeological Research Consultancy at the
University of Sheffield, now part of Wessex Archaeology), the results of which prompted
further detailed excavation in five areas (Trenches A-E) (fig. 4) (NGR SK51179881).
Although much of the stratigraphy had been truncated by post-medieval activity, three
main phases of pre-modern activity were identified in several pockets of archaeology
surviving across the site (O’Neill 2004).
The earliest phase comprised a substantial cut feature of uncertain function (figs. 5 and
6), itself cut into by a broad linear feature. The proximity of the spring has led to the
survival of waterlogged materials, including a linear post and plank-faced structure,
stakes and a wicker fence or track. There was no dateable artefactual record associated
with these structures and the sixth to seventh century date was achieved by
dendrochronological and radiocarbon dates (see below). The original extent of the
planked structure could not be determined, but sufficient was recovered to indicate its
careful construction, and its interpretation and that of its successor form the focus of this
paper. This had been cut into by a broad, shallow ditch, which appeared to be on the same
alignment. Palaeoecological materials from these earlier phases of the site were well
preserved. Despite the site’s proximity to the church, there is little evidence of further
activity until the 11th -12/ 13th centuries, when several pits were dug, possibly within field

6
or property boundaries, in two concentrations north-east and west of the earlier features.
These may have been contemporary with the Norman castle, or a major rebuilding of the
church in the 12th century (Ryder 1982, 45). The steep drop of approximately two metres
between the churchyard and properties on the north side of Church Street indicates some
truncation of the stratigraphy before their construction and the last main phase identified
belongs to these post-medieval structures, the ancillary buildings of which had been
demolished prior to the building of the flats in the 1960s. One other building of note that
survived on the site until demolished in the 1940s was the Priory Manor House; this was
not identified during the investigations. The house, thought to be eighteenth century in
date, had survived in a central area of the site known as Low Fold, and it is possible that
this building was on the site of an earlier structure (Allport 1911), a lodging attached to
Lewes Priory in Sussex as part of its foundation with revenues from the Conisbrough
Estates in 1078 (Hunter 1828, 1, 197-215). The evidence for this location, however, is
equivocal as another post-medieval building called 'The Priory' lies immediately east of
St Peter's Church and High Street (fig. 1).
The Structures
The principal excavated area (Area A/Trench 4; figs. 6-10) lay east of the wellhead,
entirely on the weathered grey clay surface of the Coal Measures, and this impervious
horizon had lead to extensive waterlogging of the archaeological features dug into it.
Beneath the modern tarmac surface associated with the 1960s development, was the
corner of a post-medieval building, a boundary wall, culvert and associated buried topsoil
and subsoil deposits. Pottery of 17th to 18th century date from immediately below the
building provides a terminus post quem for its construction, although the quantity of
primarily 19th to early 20th century utilitarian wares recovered from demolition rubble and
accumulating soils beyond the building to the south and east might indicate a later date.
Beneath the eastern wall of the building was a single short section of stone wall, on a
different alignment to the later building and property boundaries, possibly of late
medieval date. Below the latter wall, and extending across the site above the natural clay,
was a layer, probably a soil, containing 13 -14th century and 15 - 16th century pottery.

7
The ‘Ditch’
Along the southern edge of the site, natural clay was encountered immediately beneath
the modern surface material, but to the north, a broad shallow linear feature [cut 4025]
(figs. 5 and 6), running east-west, had been cut into the clay, and this still took some
subsurface flow from the spring to the west. The feature [cut 432]/[4025]/[4047] was
traced for a length of at least 20m, and 6-9m in width, it survived to up to 1m in depth.
Becoming narrower and shallower with the slope towards the east, this was almost
certainly as the result of later landscaping [4011], perhaps associated with the
construction of 19th century properties in this area. The extent of the feature, interpreted
as a broad, shallow ditch, was not established, and insufficient was excavated to be
certain of its alignment, whether it was straight or curved around the church. Two other
trenches were excavated on the projected alignment of the feature. To the east (Trench 3),
the stratigraphy had been largely removed by the insertion of a post-medieval boundary
and associated reduction of ground levels, although a shallow deposit at the base of
Trench 3, similar to a deposit (413) filling the ditch feature in Area A/Trench 4, may have
been the base of the feature surviving beneath this later truncation. To the west, beyond
an extant 1960s housing block, since demolished, no trace of the feature or associated
deposits were identified in Trench 5, and no access was given to the adjacent area under
the housing block during later building work.
It seems possible that the ditch reflects the northern edge of the deer park extending along
Old Road and Common Lane to the gates of the Norman castle (figs. 1 and 3), a line later
cut back by the development of the village of Conisbrough. Environmental evidence,
presented below, would support this interpretation.
The Planked Structure
The cutting of the linear feature had largely removed deposits associated with an earlier
structure which on the north side was faced with a timber revetment [4048] consisting of

8
large uprights, between 2m and 2.5m apart, into which horizontal planks had been fitted
into grooves in the uprights (figs. 7-10). This revetment had been constructed of large,
well finished horizontal oak timbers, fitted together with tongue and groove along the
tops of the planks. This planked wall survived over a length of 10.7m in three and one
half ‘bays’, two planks high at its western extent, the largest plank measuring 2.5m by
1.66m and 0.12m in thickness. The one vertical post where it was possible to expose the
base showed it to have been cut off square and the length of timberwork had been
inserted along the edge of the excavated feature, which had been cut into the natural
weathered surface of the Coal Measure shales. At no point did the woodwork survive to
its full original height and it had been hacked through during the digging of the overlying
linear feature [4025], disturbing some of its timber into the filling [413]. This suggests a
significant time gap between the destruction of any surface trace of the timber-revetted
feature and the cutting of the ditch.
The base of this feature had the remains of either a collapsed wattle fence or track [4064],
approximately parallel with the revetment, a small plank-lined pit [4060], a line of stakes
[4065] and other wood [4087]. Little remained of associated deposits [4052; 4059]
infilling this structure, since these had been largely cut away by the large linear feature
[4025].
The only dateable artefactual material recovered from the filling of all the features was a
few sherds of local Roman grey wares and samian, all of which were clearly residual. A
small quantity of worked flint and waste were also recovered, none of which was
diagnostic of any particular period. The quantity of material of all periods before the
medieval settlement is no greater than that which would have been obtained from local
field-walking and perhaps represents manuring scatters.
Some evidence of occupation in the vicinity is indicated, however, by large amounts of
smithing slag and numerous fire-cracked pebbles in the filling of features, particularly
from context 413, the upper fill of the linear feature or ditch, although like the Roman
pottery, none of this needs to be contemporary. Timbers in the ditch also appear to have

9
been redeposited either from the upper part of the planked wall or a structure
contemporary with it. Faunal remains were few in number and showed no evidence of
butchery or working, with the exception of two examples of worked antler, one of which
appeared to have been soaked to aid working.
Dating Evidence: Tree-Ring Analysis of Timbers and Radiocarbon Dating
In the absence of artefactual material from the timber structures, the dendrochronological
evidence represents a crucial element in site interpretation. All the large timbers from the
site were of oak. Complete sequences of growth rings in the samples which were selected
for dating purposes were measured to an accuracy of 0.01mm using a computer-based
travelling stage (Tyers 2004). The ring sequences were plotted onto semi-log graph paper
to enable visual comparisons to be made between sequences. In addition, a cross-
correlation algorithm (Baillie and Pilcher 1973) was employed to search for positions
where the ring sequences were highly correlated. These positions were checked using the
graphs and, where these were satisfactory, new mean sequences were constructed from
the synchronised sequences. The t-values reported below are derived from the original
CROS algorithm. A t-value of 3.5 or over is usually indicative of a good match, although
this is with the proviso that high t-values at the same relative or absolute position must be
obtained from a range of independent sequences, and that these positions are supported
by satisfactory visual matching. All the measured sequences from this assemblage were
compared with each other. Any that were found to cross-match were combined to form a
series of site master curves. These master curves and the remaining unmatched ring
sequences were then tested against a range of reference chronologies, using the same
matching criteria: high t-values, replicated values against a range of chronologies at the
same position, and satisfactory visual matching. Where such positions are found these
provide calendar dates for the ring-sequence.
These tree-ring dates can initially only date the rings present in the timber. Their
interpretation relies upon the nature of the final rings in the sequence. If the sample ends
in the heartwood of the original tree, a terminus post quem (tpq) for the felling of the tree

10
is indicated by the date of the last ring plus the addition of the minimum expected number
of sapwood rings that may be missing. This tpq may be many decades prior to the real
felling date. Where some of the outer sapwood or the heartwood/sapwood boundary
survives on the sample, a felling date range can be calculated using the maximum and
minimum number of sapwood rings likely to have been present. Alternatively, if bark-
edge survives, then a felling date can be directly utilised from the date of the last
surviving ring. The sapwood estimates applied through-out this report are a minimum of
10 and maximum of 46 annual rings, where these figures indicate the 95% confidence
limits of the range. These figures are applicable to oaks from England and Wales (Tyers
1998). The dates obtained by the technique do not by themselves necessarily indicate the
date of the structure or deposit from which they are derived. It is necessary to incorporate
other specialist evidence concerning the re-use of timbers, and the repairs of structures
before the dendrochronological dates given here can be reliably interpreted as reflecting
the construction date of phases within the archaeological sequence.
Results
Initially some samples were examined during November 2002 as part of an assessment of
the potential of the material following the archaeological evaluation. The majority of the
material was analysed during January and September 2003. All the material was assessed
for its potential for dendrochronological analysis and nine timbers were selected for
analysis. The rest of the material had no potential for analysis since they either contained
too few rings, or contained a variety of anatomical anomalies that prevented their reliable
analysis. The tree-ring sequences from seven samples (Fig. 11 and Table 1) were found to
cross-match with each other. A single composite 149-year sequence was calculated from
the overlapping series and a match was located for this sequence indicating the rings
covered parts of the period AD 425-573 inclusive. Sub-samples from a component timber
within the sequence were dispatched for radiocarbon analysis to confirm this dating.
These results provide some independent support for the dating of this sequence and this
match is accepted in the absence of opposing evidence (see below).

11
The Tree ring evidence: Discussion
A summary of the findings is presented in Table 1, and Figure 11. The overlaps in the
dendrochronological sequence leave no doubt that, as the excavated evidence would
suggest, that the plank-built wall [4048], small wooden framed pit [4060] and the wood
and wattle work on the floor of the structure [4064] are part of the same structural
sequence. The single timber from the upper filling of the ditch which cuts across the
earlier feature [413] has a similar ring sequence and was probably therefore disturbed
from it. As a caveat, however, none of the datable samples definitely include bark-edge;
one retains significant quantities of sapwood and a further sample is complete to the
heartwood/sapwood edge. The absence of datable bark-edge effectively prevents the
dendrochronological analysis from determining whether all of this material is precisely
contemporaneous or is the product of a number of temporally close construction events.
Accepting the structural evidence, that the timbers are precisely coeval, and hence the
result of a single event, the results can be combined into the narrowest common period,
and this would indicate the felling and probable initial use of the timbers fell within the
period AD 573-90.
In dendrochronological terms the quality of the cross-matching of the Wellgate sequence
to two contemporaneous sequences is good, but the dating this provides is supported by
relatively few other series, hence the result has poor levels of independent replication.
The two sites that yield highly significant correlations with the Wellgate sequence consist
of a single timber from Coppergate in York (Groves et al. in press), and a multiple series
of samples from a horizontal mill in Tamworth, Staffordshire (Baillie 1992). The
distribution of ‘dark-age’ tree-ring data within England is extremely uneven with the
Conisbrough sequence the first produced from South Yorkshire, whilst there still are
none from the East Midlands or the North-East. The situation has not changed
significantly since the most recent synthesis of the evidence (Tyers et al. 1994), with
most of the contemporaneous tree-ring series derived from London or the southern
counties. The absence of significant correlations between these and Wellgate suggests
that most of these are too far away, themselves aberrant, or too poorly replicated to yield
statistically significant correlations at these distances. Within this overall picture it should

12
be noted that the material from Tamworth provides some additional statistical support for
the dating evidence since there are significant correlations between several individual
series from Wellgate and similar series from Tamworth.
Given the archaeological significance of the findings at Wellgate and the crucial issue of
dating, it was felt that the dendrochronological dating would have to be backed up by a
series of radiocarbon results before they could be considered completely secure. Two
segments of timber 4 413 SF18 were independently submitted for radiocarbon analysis
(Table 2). These are samples Beta-183712, and Beta-185185, which are the inner and
outer halves respectively of this timber. Allowing for the offsets between the sample
locations within the timbers and the end dates of the sequence, these results individually
provide supporting evidence for the growth period of this timber as somewhere in the
third to eighth centuries AD. The original intention was to provide closer support for the
dendrochronological date by wiggle-matching the results obtained from a series of
radiocarbon samples of known decades from within the site sequence (Bronk Ramsey,
van der Plicht and Weninger 2001, 381-389). This sample was the only one still suitable
for radiocarbon analysis when this requirement became clear, its size is small, reducing
the amount of material usable for the analyses, and the relatively short length of the tree-
ring sequence from this sample also works against this aim. Analysis of the results using
the D_Sequence function in the radiocarbon calibration program Oxcal (Bronk Ramsey
1999) does yield a calibrated end-date for the Conisbrough sequence of AD 500-690 (at
95% confidence), but there is a caveat in that the internal correlation of the radiocarbon
data obtained with a known calendar interval is very poor. Given this statistical anomaly,
the radiocarbon results cannot be used to support the dendrochronological dating in the
way they were originally intended, but it should be stressed that the dating does not
disprove the dendrochronological date given here. An additional date (Beta-183713) was
obtained on another timber from the same context [413] and its range, of AD 380-650,
similarly overlaps. In view of the fact that these timbers are almost certainly disturbed
into this layer, it is unfortunate that further dates upon associated plant macrofossil
material are not available.

13
Archaeological Evidence: Discussion
It is unfortunate that radiocarbon dating was restricted to wood from the infilling of the
feature cut through the planked wall, with a view to validating the dendrochronological
sequence rather than dating the features per se, although the absence of evidence for
reworking of the timbers and their preservation suggest that there was a relatively short
interval between any earlier use and re-use. The structure must be dated to the late sixth
or early seventh century, and belongs to the period when locally the British kingdom of
Elmet was being absorbed into Saxon Northumbria.
The well finished nature of the woodwork (figs. 8-10) does raise the possibility of re-use
of timbers from an above ground building, or that the surviving fragment is the north wall
of a substantial cellar to a larger structure utilising this method of construction. The
grooved upright and horizontal plank technique, termed bulhus or bulværk in Sweden, is
known from Viking Age structures in Scandinavia (Bäck 2009; Fallgren 2008, fig. 7.2
from Carlsson 1981), and survived in peasant housing in the Baltic region, particularly on
Gotland, and in Denmark and southern Norway into the post-medieval period (Roede
1999, fig. 9). In England, this technique appears as early as the Late Iron Age at
Glastonbury lake village in Somerset, where it is used to revet a causeway (Coles &
Minnett 1995, fig. 4.13c). Goodburn (1994) has noted its use in mid-eleventh to late
twelfth century structures in London, it appears in a thirteenth century building at Weoley
Castle, Birmingham, where the planks are also slotted into a horizontal sill beam (Oswald
1964, fig. 44; see also Smith 1965), and Meeson (1983) has suggested that some of the
surviving timberwork in the isolated church at Rushton Spencer in north Staffordshire
reflects a similar thirteenth century plank-built structure, although a more recent
dendrochronological assessment indicates a slightly later date of ca. 1318 (Walker 1999,
table1). It is apparent, however, that the intermittent character of the evidence is a
product of both the limited nature of the waterlogged archaeological record and the
chance survival of timber buildings.

14
There is little excavated evidence of Middle Saxon buildings, beyond plans recovered
from the disposition of postholes, and still less of sixth century British structures,
particularly where preservation is sufficient to infer the detailed nature of any
timberwork. The evidence provided by excavated posthole and construction trench is
generally insufficient to indicate whether horizontal planks had been fitted into grooves
in uprights, rather than lodged behind them, or whether panels were infilled with wattle
and daub. A building at Thirlings in Northumberland roughly contemporary with that at
Conisbrough, apparently had planks slotted between vertical posts (Webster and Cherry
1975, 226, fig. 90; O’Brien and Miket 1991), and the closely spaced large postholes of
Saxon halls on sites such as Chalton in Hampshire, and Maxey, near Peterborough, for
example, would have allowed the intervening spaces to be filled with short, horizontal
planks (Rahtz 1976, fig. 2.15). Addyman, Leigh and Hughes’ (1972, fig. 13)
reconstruction of the Chalton evidence, however, suggests semicircular posts either side
of a horizontally planked wall, similar to Thirlings. It is at least as probable that the
spaces would be filled with wattle and daub panels, the ends of laths or wattling being
fixed into grooves in the uprights, a feature apparent in many later medieval buildings.
James, Marshall and Millett (1984, 182-215) provide a discussion of early medieval
buildings using plank and post construction, although in several cases the excavated
evidence, for example, in the putative Roman amphitheatre at Inveresk (Neighbour 2007,
fig. 7) and the pre-rampart structure beneath the castle at Hen Domen (Barker and
Higham 1982, fig. 13), the excavated evidence remains unconvincing.
The construction of the Wellgate structure, with well finished tongued and grooved
horizontal planks fitting into grooved uprights, is curious if all that was intended was a
revetment, since a more effective way of supporting the side of any below ground
structure would have been, as in many medieval wharves, to have fitted planks behind
vertical retaining posts (e.g. Hull, Ayers 1979, fig. 6; Kings Lynn, Clarke and Carter, fig.
46). It is possible that, as in the ninth century horizontal mill at Tamworth, Staffordshire
(Rahtz and Meeson 1992, fig. 82), the joints were intended to provide a more rigid
watertight structure. The catchment of the spring in Wellgate, however, is small, and it is

15
very unlikely that it would have provided more than a trickle of water, insufficient to
power a mill, but perhaps enough to fill a fishpond.
Environmental Samples
In the absence of an artefactual record, the palaeoecological evidence may provide
contextual as well as landscape information (cf. Buckland et al. 2001), and samples were
collected for macrofossil analysis, although because of limited funding this research had
to be limited to examination of a number of samples for their fossil insect faunas.
Samples of approximately five litres each were taken from the base of the filling [4052],
adjacent to the planked structure, and as a column of six samples through contexts 416
and 413/439 in five 10 cm thick slices with a basal sample of 5 cm at the bottom of
context 416, close to the base of the ditch in the western part of the excavation (fig. 7,
Section 10). The matrix of the samples consisted of a dark grey clay with a few evident
plant remains. Sample 4052, associated with the wattle and timberwork adjacent to the
planked wall, was a cleaner, lighter grey. All samples contained occasional well rounded
pebbles of Coal Measure sandstone and quartzite pebbles from the Sherwood Sandstone
group and these derive from patches of local glacial deposits (cf. Mitchell et al. 1947,
136).
The samples were washed out over a 300µm sieve and subjected to paraffin (kerosene)
flotation (Coope 1986). The resultant floats were washed in detergent and stored in
alcohol before sorting under a low power binocular microscope. Insect remains were
identified using the reference collection then housed at the University of Sheffield (now
at the University of Edinburgh) and critical identifications were checked against the more
extensive collections of the Doncaster Museum. Table 3 lists the taxa identified.
Taxonomy for the Coleoptera follows Böhme (2005), and habitat and fossil record data
are derived from the database BugsCEP (Buckland and Buckland 2006), checked against
primary sources where necessary; the latter software was also used to generate the
summary diagram figure 12 (see Appendix 2).

16
Interpretation
The Planked Structure
The one sample [4052] contemporary with the planked structure provides evidence of
permanent water conditions, with the remains of larval caddis flies (Trichoptera) and
several water beetles. The absence of strictly synanthropic species implies a natural
deposit accumulating in a shallow pond with muddy shores and some plant litter
accumulation in a grassland environment. Whilst the accumulation may post-date the
actual use of the structure, the implication is of an outdoor construction rather than the
basement of a larger building. The few xylophagous taxa, the click beetle Dalopius
marginatus, woodworm, Anobium sp. and the bark beetles Hylesinus oleiperda and
Leperisinus sp., both normally associated with ash, Fraxinus excelsior, are widespread at
the present day and may reflect individual trees in an otherwise open landscape. The
fauna contains one particularly rare and notable species, the colydiid Myrmechixenus
subterraneus. Usually associated with ants of the genera Formica, particularly F. rufa
(Fowles 1994), and Lasius (Päivinen et al. 2003), the beetle has few recent records from
Britain, the most northerly being from Cromford in Derbyshire in 1956 (Hyman 1992,
223), although it is probable that because of its subterranean habitat it is under-recorded.
The ‘Ditch’
It is evident from the Bugstats summary bar diagram (fig. 12) that the infilling of the
broad feature cut through the planked structure relates to a natural landscape, perhaps
best described as wood pasture, or hay meadow with old tree pollards (cf. Hæggström
1998). All the species in the faunas, which are associated with human activity, are also
found in natural situations, particularly in association with old trees, and obligate
synanthropes are absent. The succession, whilst it has evidence for damp ground and wet
mud, contains only one dytiscid water beetle, the large Dytiscus circumcinctus, a strong
flier (Nilsson and Holmen 1995, 197), which is probably adventitious in the deposit. The
insect faunas are remarkably consistent, and despite proximity to the parish church, only

17
50m to the south of the site, and any adjacent settlement, they provide little evidence for
human occupation. Although the cucujid beetle Cryptolestes ferrugineus is one of the
most common pests of stored products, and in most fossil assemblages would be regarded
as an indicator of stored grain, it is cold hardy, able to maintain populations outdoors in
England, and has also been recorded from under bark (Halstead 1993, 99-197). The
species has numerous fossil records, largely from Roman grain deposits (Buckland 1991,
updated in Buckland and Buckland 2006), and it may be significant that there is then a
gap in records until the period around the Norman Conquest when it re-appears on the
Parliament Street site in York (Carrott et al. 1995). The species was perhaps largely
restricted to ‘natural’, or at least wood pasture habitats, during periods of a reduction in
the large scale movement and storage of grain (cf. Kenward, in Hall and Kenward 2004,
424). The large assemblage of other species which can be associated with wood pasture
at Conisbrough suggests that this is the most likely origin for the specimens. Alexander
(2002, 48) also notes that early records of the nitidulid Carpophilus sexpustulatus are
from stored products, but that it has become increasingly common in wood pasture, and
Garland (1983, 9) regards it as an old woodland species, noting several records from the
Doncaster region. Continental records also indicate that it occurs under damp bark on
damaged deciduous trees, and this is likely to be its primary habitat (Koch 1989, 143).
The often synanthropic endomycid Mycetaea subterranea is also found in the wild in the
well rotted interiors of trees, as well as in compost (idem, 234). Overall, the image which
the faunas from the broad ditch produce is one of a wood pasture environment, in keeping
with the deer park, known to have been attached to the castle throughout the medieval
period.
The assemblages are dominated by two elements, the one associated with fresh hay and
the other with old moribund trees, the carrion element perhaps reflecting a long
established raptor’s or large corvid’s nest in an adjacent tree. The fauna associated
directly with ruderals, docks, clovers and grasses (Table 4), is outnumbered by species
occurring with the early stages of decay of plant material, part of what Kenward (in
Kenward et al. 1978, 67) has termed the ‘sweet compost’ assemblage. This includes the
hydrophilids, Cercyon spp., Megasternum boletophagum and Cryptopleurum minutum,

18
many of the staphylinids, including Micropeplus, Omalium and Xylodromus spp., as well
as rhizophagids, cryptophagids and latridiids. There is significant overlap between this
community and those of herbivore dung, and some of the histerids and scarabaeids may
have been attracted to any heaps of decaying plant debris. Indeed, most of this element in
the fauna could be recovered from a summer heap of ‘grass’ cuttings from a rather weedy
lawn at the present day in south Yorkshire.
The dead wood fauna from the ditch samples is extensive and includes several species,
which are currently rare or endangered in Britain. Although the woodworm beetle,
Anobium punctatum/inexpectatum, could have been associated with structural or
discarded timber, the bulk of the xylophagous element reflects trees, rather than worked
timber. The bark beetle assemblage indicates the proximity of several living deciduous
trees, including ash, probably oak and perhaps alder. The ostomid Nemosoma elongatum
is a predator on these scolytids, often in elm, but also in a range of other timbers; it has
not been recorded over the past century north of Nottinghamshire and Cheshire (Hyman
1992, 421). The fauna from beneath peeling mouldy bark includes the cucujid Pediacus
dermestoides, widespread in southern England, and extending northwards to North
Yorkshire. The much rarer pselaphid Trichonyx sulcicollis, whilst essentially a dead
wood species (cf. Alexander 2002), is also recorded from compost heaps, but has no
modern records outside central southern England (Hyman 1994, 78-79). The fossil
record, however, includes several specimens from Roman to Anglo-Scandinavian York,
as well as a Roman record from Ribchester in north Lancashire (Hall and Kenward 1990;
Kenward and Hall 1995; Carrott et al. 2000). The minute histerid Aeletes atomarius is
usually associated with the borings of the lesser stag beetle Dorcus parallelipipedus, but
may also occur with Sinodendron cylindricum, which is present in the Conisbrough
assemblage. Abraeus perpusillus is also associated with soft, well rotted wood; both these
small histerid beetles are now restricted to scattered localities where there is sufficient
dead wood (Hyman 1992, 335-336).
The chafer beetles in the assemblage provide some evidence of the nature of any
woodland, for whilst the rose chafer Cetonia aurata may breed in either wood mould or

19
compost, Melolontha hippocastani is a woodland species, whose larvae probably feed on
roots. It is present as a fossil, however, in the otherwise open landscapes of the Bronze
Age Wiltshire Chalk at Wilsford and in the similarly open Iron Age landscape around
Tattershall Thorpe in south-east Lincolnshire (Osborne 1989; Chowne et al. 1986); both
sites lie well outside its modern distribution, which is essentially upland and north-
western (Hyman 1992, 393) The Conisbrough site is similarly marginal to its modern
distribution, although there are old records from Derbyshire, and an association with a
habitat that is now rare in the lowlands is probable. The chafer may rely on unimproved
grassland in woodland clearings, and whilst some elements of the old woodland fauna
have found temporary sanctuaries in the old trees of lowland wood pastures, any
associated pasture has often been ploughed, fertilized and otherwise improved, destroying
the species’ habitat and its continuity. Buckland (2005, 95) further suggests that M.
hippocastani may have benefited from the former presence of free ranging swine and
wild boar in parkland, opening up the otherwise rather densely grazed sward.
The suggestion of either a raptor or corvid nesting in an adjacent old tree rests on the
carrion elements in the fauna, on the relative numbers of Trox scaber, Omosita discoidea,
and several histerids. There were few bones in the deposits, and since all of these taxa are
recorded from bird nests, this appears a probable explanation. The indeterminate fleas,
Siphonaptera, unfortunately insufficiently well preserved to identify, may also have
dropped from the same source, although they could equally relate to small rodents
foraging in the compost.
Discussion
The ‘Bulhus’ Structure
The limited nature of the evidence, constrained by standing buildings to the south and
west (fig. 4) and the truncation of the stratigraphy to the east, does allow the possibility
that the plank-walled structure, with gentle slope from the south side and vertical
revetment to the north, did not extend much beyond the excavated area. The fossil insect
evidence from sample 4052 would support an interpretation involving standing water,
rather than a cellar beneath a building. It is possible that it formed the middle section of

20
an elongate pond, at least 11m long by 7m wide, perhaps utilising the water from the
spring for a fishpond or other agricultural function, such as retting of flax or hemp.
Retting (Higham 1989), however, produces particularly foul residues and any water
surface or wet mud would have attracted a characteristic eutrophic insect fauna (Gearey
et al. 2005). Another possible explanation for the Wellgate structure is that it relates to
water management for either a fish or stock pond, trapping the water just below the
source, with the planked revetment along the northern, downslope side of the feature. The
stake and wattle structures in the base of the linear feature have parallels with the
numerous fish weirs, dating from the seventh to twelfth centuries, identified along the
River Trent and the Humber foreshore (e.g. Elliott, Jones and Howard 2004, fig. 7.3), but
it is unlikely that there was sufficient flow for large scale water management. The limited
environmental evidence provides a little support for the presence of standing water, but
there were no bones of aquatic vertebrates, fish or amphibian, in the small assemblage
from the site. At Conisbrough, any fishponds – and the dating evidence is particularly
early for such structures in the post-Roman, pre-Norman period - are more likely to have
been located along either the River Don or Dearne, 0.5km to the north, or in the valley a
similar distance north-east of the site, where a millpond existed into the last century. The
catchment of the Wellgate springs, close to the top of the slope is also relatively slight
and there would be insufficient water to power a mill, and the scale of the work would
suggest a more substantial purpose than a watering hole for stock. A more prominent
obligate dung fauna would also have been expected if livestock had drunk directly from
the feature.
Truncation of the structure by the later broad ditch makes interpretation difficult and
allows the possibility that it was asymmetric, with a gentle cut slope southwards towards
the present church and the downhill side strongly revetted to provide a vertical wall.
Despite the limited length excavated, the scale of the structure is clearly quite significant
and the question has to be asked as to whether it formed part of an enclosure around a
site, perhaps the temenos of a monastery or the original burh of Conisbrough, centred on
the later church. The form is without parallel in Anglo-Saxon defences, however, and it
is probable that the feature, if linear, was demarcatory rather than defensive. In the latter

21
case, it could only have been effective where strong enfilading power would have
enabled the trapping of an enemy against the wooden wall; it is therefore not surprising
that such form appears with the development of firearms in late and post-medieval
architecture, and this suggestion can perhaps be discounted.
A stock pond for fish, with the wood-lined central pit functioning as a trap after draining
appears the most probable interpretation of the structure. Fishponds are known from
Roman sites in England (Zeepvat 1988) and fisheries figure in Anglo-Saxon monastic
charters, as well as Domesday, although there is insufficient detail to define particular
structures. The will of Wulfric Spott (1002 x 4; Whitelock 1979, no. 125, 586-589;
Sawyer 1979, no. 3) refers to renders of fish to Burton Abbey, although the source of
these is likely to have been in the eastern part of the Conisbrough estate on the Don at
Tudworth, where fisheries belonging to the lord of Conisbrough, providing 20,000 eels,
are recorded in Domesday (Hunter 1828, I, 97; Hey 2003, 44). There is no convincing
evidence of fishponds in the period between the end of the Roman period and the Late
Saxon, and Currie (1991) in his study of the spread of carp suggests that the earliest large
scale ponds are a creation of the Norman aristocracy. Whatever the Conisbrough
construction is, its location suggests association with other elite buildings, either
monastery or palace, close to the later minster church.
The Ditch
The insect assemblages from the ditch quite clearly indicate an environment, wherein
decaying plant material, perhaps intended as fodder, accumulated, or was disposed of in
the wet, grassed over linear feature, which continued to silt up. The stratigraphy (fig. 7)
indicates at least one re-cutting along the same alignment (4054 cut into 4055/4027 and
4090 into 4084/4046], suggesting the need to renew a structure which, even allowing for
its truncation is more likely to be demarcatory than defensive, a point reinforced by the
environmental evidence. Nearby, probably along any associated bank, were old trees,
perhaps pollards with much dead and decaying wood, with possibly the nest of a corvid
or raptor. The fact that the earthwork is oblique to the existing street plan and appears to
have had little if any influence on the layout of the village suggests that any significant

22
late Saxon and Norman settlement lay elsewhere, and that the feature had largely silted
up before the present village plan developed. The paucity of any artefactual evidence also
supports this. There is little dating evidence and the radiocarbon and dendrochronological
dates on oak timbers seem to relate to material disturbed from the plank-walled structure
or a contemporary building during the cutting of the ditch. The extent of truncation
means that it is not possible to be certain when the ditch was abandoned, although the
environmental evidence suggests a process of silting up in a wood pasture environment,
perhaps with some disposal of plant litter, fodder as opposed to any domestic debris. The
possibility has to be considered that the structure relates to the primary northern boundary
of the medieval deer park known to be associated with the castle.
The park at Conisbrough survived into the sixteenth century, and Hey (2003, 70) notes
that as late as 1539 there were 440 fallow deer within the park. The known bounds of the
medieval deer park (fig. 3), however, still shown as the civil parish of Conisbrough Parks
on the Ordnance Survey map, can be defined as a north-south irregular, elongated oval,
south of the Doncaster Tinsley turnpike road to Hill Top, and then the parish boundary to
Firsby and Ravenfield, east to Micklebring, and north through Clifton back into
Conisbrough, a circuit which excludes St Peter’s church, castle and village, but does
include the site of a Roman villa close to its eastern boundary. It is possible that the
excavated feature reflects an earlier park boundary, extending along Old Road and
Clifton Hill north-eastwards to the gatehouse of the Norman castle, but this would place
the existing parish church within the park. Although the evidence would suggest that the
excavated feature did not influence the layout of the medieval and later village, the south
eastward deflection of Old Road/Church Street may be a reflection of the course of the
excavated feature. It is probable that any remaining feature had been levelled or had
largely silted up before the present village plan developed, perhaps in the late twelfth
century when the Saxon church was enlarged (cf. Ryder 1982). The grant of a weekly
market and annual fair in 1201 may provide a context for this (Hey, 2003, 70). The ditch
would therefore date from shortly after the Conquest, when the initial Norman ringwork
or motte and bailey was constructed on the site of the later stone castle, itself rebuilt in
stone in the late twelfth century (Johnson 1980), and it provides no evidence for a pre-

23
Conquest deer park, although the plank-walled structure must relate to a significant
residence on the site in the latest sixth or early seventh century.
An immediately post-Conquest date is itself interesting and the evidence does suggest an
earlier boundary to either park or estate. Both Hooke (1986, 159-163; 1988, 123-156;
1998, 21) and Rackham (2003, 189-191) note the problems over the meaning of the place
name element haga, the root of both hay and hedge, and the probability of enclosures,
predating the earliest reference to a park (derhage), that at Ongar in Essex in an Anglo-
Saxon Will of 1045. Fallow deer, that archetypal park animal, appear not to have been re-
introduced until after the Norman Conquest, probably directly from Sicily (Sykes 2004,
75-84; see also now evidence for Roman fallow deer in Sykes et al. 2006, 948-959) and
the idea of a formalized enclosed park may have come with it. Rowley (1984) has
suggested that the idea for Henry I’s (1100-1135) park at Woodstock in Oxfordshire
derived directly from Sicily, where the Normans, under Robert Guiscard de Hauteville,
had been established since 1060. Liddiard (2003, 4-23) has recently argued that the
absence of references to deer parks in Domesday is an artefact of that document in they
were not places where the king could derive revenue directly and were therefore not
included, and Hooke (1985, 159-163; 1988, 123-152; 1998, 19-32), on field and place
name evidence, has argued for the existence of pre-Norman parks in Wessex and
southern Mercia. On the Continent, Nitz (1988, 272) and Hauck (1963, 30-74) have both
noted the existence of hunting parks attached to Carolingian palaces, and it is apparent
from sources from the mid-seventh century onwards (cf. Kiess 1998, 11) that it remains
possible that hunting reserves may have been attached to the palace sites of peripatetic
English Saxon kings.
With only a single section, the projected extent of a boundary at least in part coeval with
a later deer park can only be hypothetical, and any enclosed area at Conisbrough may
have been much smaller. The ditch may have encircled a site centred on the later Saxon
church, although the environmental evidence would imply an old, established boundary
with mature trees and grassland, which fits better with the wood pasture of a deer park.

24
An interpretation to be considered is that the structure forms part of the enceinte of the
‘king’s burh’ implied by the place name, a ‘villa regia’.
Of the extensively excavated palace sites, any boundary at Northampton (Williams et al.
1985) has not been examined, and that at Cheddar (Rahtz 1979) shows no enclosure ditch
on the scale of the Conisbrough feature. At Yeavering, at the other end of the kingdom of
Northumbria from Conisbrough, the palace complex includes a large enclosure,
surrounded by what appears to have been a walkway supported on two rows of large
posts, a construction for which Hope-Taylor could find no contemporary parallels.
Initially interpreted as a ‘fort’, in the final report, he is less certain, and opts for the
neutral term ‘enclosure’ (Hope-Taylor 1977). It appears particularly massive for a simple
enclosure for domestic animals, and it is possible that any walkway faced inwards rather
than outwards, and like the segmental structure on the same site interpreted as a ‘theatre’,
it may owe its origins to the imitation of Rome and the ritualized animal hunts in the
circus. At both Yeavering and Conisbrough, a church and cemetery later came to occupy
part of the enclosure.
The construction of the large church at Conisbrough in an evidently Northumbrian style
(Ryder 1982, 45), in a position which must lie within any enclosure defined by the ditch,
implies that the site remained in royal hands at least into the eighth century, and there is
some evidence to suggest that the site had earlier significance. Miller (1804, 262) notes
the discovery of what must have been a cist grave in the area of the churchyard. Sub-
Roman, or Anglian burials of this form have been discussed by Faull (1979, 5-9, see also
Buckland 1984, 55), and the burial may have lain on the boundary of an estate centred on
the Conisbrough Park Roman villa (For similar examples, see the discussion in Zadora-
Rio 2003, 1-20). Some deliberate attempt to reinforce the connection with Rome may be
evident in the building materials of the early church for although the church lies
immediately adjacent to excellent freestone in the Lower Magnesian Limestone, it
incorporates large, evidently reused amounts of Coal Measure sandstones, as well as
reused blocks of the limestone. Both may have come from the villa site, particularly its
bath house, but like the sandstones in the nearby churches of Laughton-en-le-Morthen,

25
Arksey, Bolton-on-Dearne and Burghwallis, it may have travelled further, from the ruins
of the Roman forts at Templeborough, near Rotherham, and Doncaster. From the material
surviving in Rotherham Museum, the fort at Templeborough appears largely to have
utilised the Upper Carboniferous Rotherham Red Rock (May 1922), whilst the surviving
stretch of the late Roman fort wall in Doncaster is of Lower Magnesian Limestone,
although Upper Carboniferous sandstones were recovered from the Roman urban
defences (Buckland and Magilton 1986, 102). Such deliberate transport of Roman
materials (cf. Morris 1988, 191-200) may have been seen as enforcing the legitimacy of
Northumbrian rule, although it has to be admitted that it has been argued elsewhere
(Buckland 2010) that there is evidence that such redistribution took place in the Anglo-
Scandinavian period.
On the dendrochronology, the plank-built structure belongs to the late sixth century or
early seventh century and this allows alternative explanations, either as a fishpond or
building associated with a British stronghold or monastic site, later taken over by
Northumbrian kings, a southern parallel for Yeavering in Northumberland.
The historical context of the Conisbrough estate
The region either side of the River Don appears to have been a frontier zone from at least
the late Iron Age onwards. The most northerly pre-Vespanianic Roman forts, at
Templeborough, west of Rotherham, and Rossington Bridge, south of Doncaster, lay
close to the river, and Buckland (in Buckland, Hartley and Rigby 2001, 86-87) has
argued that the concentration of Roman pottery kilns in the area south of Doncaster
reflects the movement of potters to the northern limits of their own, or at least their
owners’ civitas, that of the Corieltauvi; to the north of the river lay the Brigantes. How
this division persisted into the post-Roman period is uncertain. Both Hunter Blair (1948,
98-126) and Buckland, Magilton and Hayfield (1986, 16-19) have argued that the so-
called Roman Rig, a linear earthwork running from the Don near Sheffield to both the
Don and Dearne, north of Conisbrough, reflects the border of Northumbria, either the
delimitation of the northern frontier of Mercia, perhaps under Offa, or the southern limit

26
of the British kingdom of Elmet, although Higham (2006) has recently argued otherwise
and Roberts (2010) in a study of cropmarks on the Magnesian limestone outcrop prefers
an Iron Age origin and dates on linear earthworks elsewhere in west Yorkshire would
support this (cf. Roberts et al. 2001). Whatever its date, its eastern terminus was close to
the Don at Strafforth Sands, the ford over the river below Conisbrough, where Rickneild
Street, a prehistoric route along the limestone escarpment, descends to the crossing.
Strafforth is also the name of the Wapentake. As Buckland and others (1986, 19),
following older authors note, Conisbrough may be the Villa Regia of Bede, one of the
centres with Doncaster, if this is his Campodunum, within the large multiple estate
extending eastwards to Hatfield. At Doncaster, the late Roman fort was refortified at least
twice before the construction of the Norman castle and the earlier of these has been
described as ‘Anglian’, although it lacks convincing dating evidence (Buckland, Magilton
and Hayfield 1986, 72-83).
To the west, the region of Hallamshire has its origins in a further estate centred on the
church at Ecclesfield and a probable civil centre at Sheffield; it was bounded on the
southern side by the river Sheaf, a tributary of the Don (Hey 2000, 31-47). A meeting
took place in 828 at Dore on this river between the Northumbrians and the victorious
Egbert of Wessex, and Dore is also mentioned as the northern limits of Mercia in 942,
when Edmund of Wessex incorporated the Danelaw into his expanding kingdom
(Rollason 2003, 262).
The dating evidence, both dendrochronological and radiometric, points to an origin for
the structure either at the very end of the sixth or early in the seventh century, and the
historical context is significant. In 606, the Bernician king Æthelfrith took over Deira,
killing its king Ælle and driving the latter’s son Edwin into exile with Cearl, King of
Mercia. In ca. 615, Æthelfrith, now king of a temporarily united Northumbria, defeated
the Britons at Chester. Higham (1993) has suggested that Bede (HE II, 2), with his
concern for the moral superiority of the Roman church over the British, conveniently fails
to mention Mercian involvement in the conflict, which seems to have led to the collapse
of Mercia and Edwin having to seek refuge with Rædwald of East Anglia, where Bede

27
alleges that Æthelfrith attempted to engineer his murder. The subsequent events leading
to the death of Æthelfrith in 616 at the Battle of the Idle, presumably close to the place
where the Roman road from Lincoln to Doncaster crossed the river, on the south-east
boundary of the Conisbrough multiple estate, should be seen not as an altruistic move by
Rædwald in support of Edwin, but as a reaction to the perceived threat to East Anglian
hegemony of an expanding Northumbria. Edwin appears to have remained Rædwald’s
client until the latter’s death in ca. 624-5, but his ability to strike at Cuichelm of Wessex
shortly thereafter without regard to opposition from Mercia suggests that the damage
inflicted on any Midland warbands at Chester was not yet repaired. Within ten years
however, Penda had successfully rebuilt Mercia and in alliance with Cadwallon of
Gwynedd had defeated Edwin at the Battle of Heathfield in 633. Although Revill (1975)
has argued that this conflict took place near Cuckney in Nottinghamshire, the inclusion of
Hatfield with Lindsey in the Tribal Hidage, which following Brooks (1989, 159-171),
Higham (2000, 46-47) has interpreted as a Northumbrian document of the time of Edwin,
might support a site on the eastern boundary of the Conisbrough estate, between Thorne
and Hatfield Moors. The order of places in the Tribal Hidage suggests that Elmet had
previously been tributary to Mercia, and it is tempting to see the takeover or creation of
the multiple estates of Conisbrough and Ecclesfield as a development of the defeat at
Chester and the need for Æthelfrith to drive a wedge between Mercia and its sometime
ally of Elmet to the north. The incorporation of Elmet into Northumbria by Edwin and the
expulsion of its British king Ceretic in 617 was a natural consequence of its isolation.
Blair (2005, 275-279) has argued recently that it is ‘hard to recognise a category of royal
palaces during the seventh to early ninth centuries which were inherently deep-rooted and
stable’ and that the seventh- and eighth-century sites with large timber halls that have
been identified by aerial photography were largely or wholly unenclosed. Instead, a king
and his retinue would move around, living off food-renders from the various territories
that made up his kingdom. If the Conisbrough structure is interpreted as a stock pond for
fish, and it currently has no parallels, it might relate to provisioning of king or monastery.
The latter may have developed into a minster, sites, which by contrast, had a more
permanent character and, unlike nearly all the secular sites that are known, continued

28
through into the high middle ages and afterwards as stable central places. They were
often known as burhs, which before the mid eighth century seems to have been a
vernacular alternative to mynster. Once a minster had been founded within a regio it
might become ‘so much its natural centre as to become synonymous with the territory’.
From the mid ninth century onwards, minsters were annexed by kings, lords, and
religious corporations as secular centres. By the time of the Domesday survey old minster
sites, such as the Yorkshire ones of Ripon, Beverley, Otley, Gilling, Whitby, and
Dewsbury, had become ‘secular places containing residually important churches’. Blair
(2005, 326) draws attention to the various “Kingsburys’, another form of the name
Conisbrough, such as those in St Albans and Aylesbury, which developed in this manner.
Can the development of Conisbrough be seen in this way? The place-name first appears
in surviving documents in 1002-4, when it took the Old Norse form of Cunugesburh,
which may have replaced an Old English cyning, ‘king’. Was an old minster site
demarcated, if not fortified in the Anglo-Saxon period long before the Norman castle was
built at the end of the promontory? St Peter’s Church, which stands on the crest of the
hill above the archaeological feature being discussed, is a large church, whose nave
measures internally c. 13.4 m by 5.28 m, comparable to the 14 m by 5.3 m of Ledsham
church (21 miles to the north), which Taylor and Taylor (1965; see also Faull 1986)
considered to be of eighth-century date. This similarity was first raised by Davies (1969)
in a local history pamphlet and developed by Ryder (1982). Both churches have large,
side-alternate quoins in the Northumbrian style. The surviving architecture, the
dedication, and the large medieval parish of Conisbrough suggest that it was an important
minster, and the burh element of the place-name may have been acquired without
fortification; only further excavation can reveal whether this is the case.
Could the late sixth / early-seventh century structure described above be associated with
an even earlier minster, perhaps an eccles within the British kingdom of Elmet? If so, it
would have been comparable with Ecclesfield, 10 miles to the west, where St Mary’s
Church served the whole of Hallamshire. This Northumbrian shire (which may have been
an even earlier territory) covered 71,526 acres on the edge of the Pennines and included

29
chapels at Sheffield and Bradfield. In 1620 Roger Dodsworth noted that, ‘This church is
called (and that deservedly) by the vulger the Mynster of the Moores’ (Clay 1904, 4-9).
Sheffield, not Ecclesfield, was developed as a secular, fortified site after (and perhaps
before) the Norman Conquest, but the name of this distinctive shire, on the south-western
border of Northumbria, still survives. Conisbrough seems to have been the central place
of a similar territory, with Doncaster perhaps as the equivalent secular centre, and
stretching from the river Don to Northumbria’s southern border.
Conclusion
Whilst the hypothesis that multiple estates had origins in the Middle Saxon period, if not
earlier, has been the subject of much criticism, (cf. Hadley 1996) - and the view that sees
Doncaster as the centre of a late Roman civitas (Whitwell 1977) is perhaps even more
extreme, some sort of organisation must have persisted in the region after the end of the
Roman period. Conisbrough provides a little evidence, which, whilst it is open to
different interpretations, reinforces the belief that it was an important site long before it
was recorded in surviving documents. We conclude, not without reservations, that the
massive oak planked structure and associated wooden structures is most likely to be a
stock pond for fish belonging to the end of the sixth or early seventh century. After a
period of disuse sufficiently long to obscure the buried timberwork, a broad shallow
ditch, orientated towards the castle was cut across it and this was itself re-cut at least
once. The archaeoentomological data suggest a wood pasture landscape with neglected
old trees and perhaps hay fodder in the ditch. The feature is interpreted as the primary
course of the deer park boundary before the development of the modern, presumably later
medieval village.
Acknowledgements
This report owes much to discussion with the late John Magilton to whom primary
acknowledgement must be made. Charles Frederick provided useful commentary on
sediments in the field, and Eva Panagiotakopulu, Peter Robinson and two un-named

30
referees commented on an earlier draft leading to extensive revision. Martin Limbert at
Doncaster Museum is thanked for access to modern reference material of insects, and
Colin Howes of the same institution provided data upon Yorkshire deer parks. The site
drawings are the work of Cathy Speight and Jo Mincher of the former archaeological
consultancy ARCUS. The initial assessment and analysis of this material was funded by
ARCUS, on behalf of Doncaster Metropolitan Borough Council, Hallam Housing
Society, and South Yorkshire Housing Association.
Appendix 1. Pollen analysis of samples from the ditch
Dr B. Gearey5
A monolith sample was collected from the filling of the linear feature [4047] in
Excavation Trench A/Trial Trench 4 for pollen analysis. Three sub-samples of 1cm3
were extracted from the monolith sequence from contexts 4056, 4085, and 4084.
Samples were prepared using standard techniques including KOH digestion, HF
treatment and Acetylation (Moore et al. 1991) with the addition of exotic spores to
permit the calculation of pollen concentrations (Stockmarr 1971). The samples
were mounted in silicon oil and counted at a magnification of x400 using a Leica
DMLB microscope. Samples would normally be counted to a minimum o f 150
land pol len grains , but pollen concentrations were very low and thus counting
proceeded until one complete slide had been counted.
The raw data counts are presented in Table 5. Pollen concentration was very low for
all three samples, which is perhaps unsurprising given the high inorganic
component of the sediment. Due to the low numbers of grains present, the data are
presented as raw counts rather than percentages. Preservation of the palynomorphs
was in general good although most grains displayed some signs of mechanical damage.
4056: In the basal silt of the ditch, the most abundant grains identified are spores of
bracken, Pteridium aquilinum, although the group including dandelions, Lactuceae

31
undiff., hazel, Corylus avellana-type, pine, Pinus sylvestris, and heaths, Ericaceae
undiff., are also present.
4085: The grasses, Poaceae undiff., and bracken Pteridium aquilinum are the most
abundant taxa, with tree and shrubs including ash, Fraxinus excelsior, alder, Alnus
glutinosa , oak, Quercus, birch, Betula and Corylus avellana-type also evidence. The
herbs sorrel, Rumex acetosa, narrow-leaved plantain, Plantago lanceolata, the
bedstraws, Rubiaceae and Lactuceae undiff. occur.
4084: The shrub Corylus avellana-type is the most common taxon, with other tree
taxa, including alder, birch, elm, Ulmus, oak, and ash present. Bracken is again
also relatively well represented. The highest counts for herbs include grasses and
sedges, Cyperaceae, but others namely the sorrels, Rumex acetosa and R. obtusifolius-
type, fat hen, Chenopodiaceae, Plantago lanceolata and Lactuceae undiff. are also
present.
Given the very low concentrations of pollen in the samples, it is difficult to draw any
secure conclusions from the data. However, a few general comments may be made. It
seems unlikely that the feature was used for retting - no Cannabis-type pollen is
recorded and concentrations of this grain are generally found to be high in
sediments from these contexts (cf. Gearey et al. 2005). Little other positive information
regarding the function of the feature can be drawn from the data. As regards the
character of the surrounding landscape, the most abundant grains in the primary and
secondary fill contexts include bracken and grasses, with other herbs including
sorrel, plantain and the dandelion group present. These grains all reflect open,
disturbed grassland contexts but tree and shrub pollen is present albeit in low
quantities. Given the fact that the pollen source area for a feature of this sort would be
expected to be relatively small, some scattered mixed woodland might have been
present near to the site. The sample from [4084] is distinguished by higher counts for
Corylus avellana-type which may reflect more hazel cover in the vicinity of the site in
the period during the later infilling of the pond/channel. This may also be supported
by the recovery of hazel shells from the feature.
In general, the pollen assemblages are not inconsistent with the interpretation from the

32
insect faunas of a managed landscape of trees set among mown hayfields, but the low
counts preclude much more detailed comment.
Appendix 2. Explanation of Bugstats summary diagram (Fig. 12)
Dr P I Buckland6
The Bugstats module of the Bugs Coleopteran Ecology Package (BugsCEP; Buckland &
Buckland 2006) has the facility to produce graphical, bar graph, illustrations of the
habitats represented by beetle faunas. It includes a variety of options for limiting, refining
and scaling the visualisation, including the ability to proportionally scale reconstructions
for increased comparability between samples and sites. By scaling the indications for
each habitat as a proportion of the total number of habitat indications, the differential
influence of varying sample size and diversity are effectively removed. This allows for a
more comparable visualisation of the habitats represented in the fauna of each sample. It
is especially useful where sample volumes are not known, but also has the advantage
over volume or weight standardisation of sidestepping a number of issues related to
sedimentation rates and sample richness and the representivity of insect assemblages. The
software can also be configured to exclude identifications above species level, increasing
the reliability of the reconstruction but potentially losing some useful information which
can be gained from genus level identifications of stenotopic genera.
The habitat classifications used in BugsCEP are derived from those commonly used by
palaeoentomologists, and species have been assigned to habitats by the authors in
reference to ecology data stored in BugsCEP (Buckland 2007). The significantly more
detailed classification system of Koch (1989-92) was also used as reference, as was the
advice of a number of species area experts.
Bugstats can be set to ignore abundance values (numbers of individuals per taxon),
which, although environmentally significant, can sometimes be misleading due to both
ecological and taphonomic issues. Different species occur naturally at different
population densities, and may occur at different densities within their habitat ranges;
some species are more likely to be deposited in certain sediments than others; some

33
genera are more easily identified than others; and there may be some bias in the recovery
of different species from samples. There are also potential problems associated with the
intercomparability of habitat indications between samples with very different total
abundances. The software, however, provides an abundance scaling option as a research
and data exploration tool for those wishing to explore these factors.
The visualisation used in this paper uses the most stringent settings for removing
uncertainty: no abundance data, habitat sum proportional scaling (%SumRep) and species
level identifications only. This provides the most probable reconstruction results as can
be expected from a semi-quantitative classification based statistic, and an excellent
complement to a species by species indicator based interpretation. The reconstruction has
the advantage of being universally comparable to any other site or sample run through the
system with the same settings. It allows broad scale environmental reconstruction
comparisons to be made irrespective of site type, age or location, and the system is freely
available and transparent in terms of its data and methods (see Buckland 2007 for more
details of the calculation methods). The Conisbrough species list and abundance data are
also freely available in the BugsCEP database.
The specific methods used for this diagram (fig 12) are as follows:
- Beetle taxa have been assigned to habitat classes (EcoCodes in BugsCEP) by the
authors as part of the Bugs database project. A taxon may represent more than one
habitat (e.g. Pasture/dung and Foul habitat/dung), but only ever one indicator class
(e.g. Dung, Deciduous, Standing water), these being narrow subsets of the general
classes.
- The software, configured as described above, tallies the habitat classes represented
by each taxon in a sample and stores the totals for each class. These numbers are
then divided by the sum of all habitat totals and divided by 100 to produce a
standardised graph. Indicator classes are excluded from this standardisation total as
their inclusion would dilute the impact of the other habitats represented in samples
where these habitats were present.
- This process is repeated for each sample, and the results graphed. The graph, raw
and calculated data are exported in an MS Excel workbook.
A report showing a sample by sample breakdown of the habitats represented by each

34
taxon can be generated using the Bugstats module in BugsCEP for a more detailed
investigation of the fauna and exactly what represents what. The software also allows the
user to explore source data and form their own interpretations should they wish to.
BugsCEP is available for download from http://www.bugscep.com and the database is
browsable at http://www.sead.se

35
FOOTNOTES
1. Paul C. Buckland, 20 Den Bank Close, Crosspool, Sheffield, S10 5PA, England, UK.
2. David Hey, Institute of Lifelong Learning, University of Sheffield, Sheffield, S10
2TN, UK. [email protected]
3. Richard O’Neill, Wessex Archaeology, Unit R6, Sheaf Bank Business Park, Prospect
Road, Sheffield S2 3EN. [email protected]
4. Ian Tyers, Dendrochronology Laboratory, Sheffield University, West Court, 2 Mappin
Street, Sheffield S1 4DT.
5. Ben Gearey, Birmingham Archaeoenvironmental, University of Birmingham,
Birmingham, B15 2TT, England, UK. [email protected]
6. Philip I. Buckland, Environmental Archaeology Lab., Department of Historical,
Philosophical and Religious Studies, Umeå University, SE90187, Umeå, Sweden.

36
REFERENCES
Borthwick Institute of Historical Research, York, TA348S
Bede, Hist. Ecc. Bede, Historia Ecclesiastica Gentis Anglorum (ed. C. Plummer), 1896, Oxford University Press, Oxford.
Addyman, P. V., Leigh, D. and Hughes, M. J. (1972). Anglo-Saxon houses at Chalton,
Hampshire. Medieval Archaeology, 16, 13-32.
Alexander, K. N. A. 2002. The invertebrates of living and decaying timber in Britain and
Ireland. A provisional annotated checklist. Peterborough: English Nature
Allport, C. H. 1913. History of Conisbrough from the earliest ages to the present day
Ayers, B. 1979. Excavations at Chapel Lane Staithe 1978. Hull Old Town Report Series
3. East Riding Archaeologist, 5, 1-97.
Bäck, M. 2009. Vikingatida bebyggelse funnen utanför Birkas stadvall. Fornvännen, 104,
262-273.
Baillie, M. G. L. 1992 Dendrochronology, in P. Rahtz, and R. Meeson (eds.), An Anglo-
Saxon Watermill at Tamworth: Excavations in the Bolebridge Street area of
Tamworth, Staffordshire in 1971 and 1978. London: Counc. Brit. Archaeol.
Res.Rep. 83, 122-4
Baillie, M. G. L. and Pilcher, J. R. 1973. A simple crossdating program for tree-ring
research, Tree Ring Bulletin, 33, 7-14
Barker, P. and Higham, R. 1982. Hen Domen, Montgomery. A timber castle on the
English Welsh Border. London: Royal Archaeol. Institute
Blair, J. 2005. The Church in Anglo-Saxon Society. Oxford: Oxford University Press
Böhme, J. 2005. Die Käfer Mitteleuropas. K. Katalog (Faunistiche Übersicht) (2nd ed.).
Munich: Spektrum Academic
Bronk Ramsey, C. 1999. OxCal: A Radiocarbon Calibration and Analysis Program,
Oxford Radiocarbon Accelerator Unit <http://c14.arch.ox.ac.uk/>.
Bronk Ramsey, C., van der Plicht, J. and Weninger, B. 2001. 'Wiggle Matching'
Radiocarbon Dates, Radiocarbon, 43, 381-9

37
Brooks, N. 1989. The formation of the Mercian kingdom, in S. R. Bassett (ed.) The
origins of the Anglo-Saxon kingdoms, 159-170. Leicester: Leicester University
Press.
Buckland, P. C. 1984. The Anglian Tower and the Use of Jurassic Limestone in York, in
P. V. Addyman, and V. E. Black (eds.) Archaeological Papers from York
presented to M. W. Barley, 51-57. York: York Archaeological Trust.
Buckland, P. C. 1991. Granaries, stores and insects. The archaeology of insect
synanthropy, in D. Fournier, and F. Sigaut (eds.) La preparation alimentaire des
cereales, 69-81. Rixenart: PACT.
Buckland, P. C. 2005. Palaeoecological evidence for the Vera hypothesis? In K. M.
Hodder, J. M., Bullock, P. C. Buckland and K. J. Kirby Large herbivores in the
wildwood and modern naturalistic grazing systems. Peterborough: English Nature
Research Report 648, 62-116.
Buckland, P. C. 2010. Ragnarök and the stones of York, in J. Sheehan, and D. Ó Corráin,
(eds.) The Viking Age. Ireland and the West. 47-59. Dublin: Four Courts Press.
Buckland, P. C., Hartley, K. F. and Rigby, V. 2001. The Roman Pottery Industry at
Rossington Bridge: Excavations 1956-61. J. Roman Pottery Stud., 9
Buckland, P. C. and Magilton, J. R. 1986. The archaeology of Doncaster. 1. The Roman
civil settlement. Oxford: Brit. Archaeol. Rep. Brit. Ser., 148
Buckland, P. C., Magilton, J. R. and Hayfield, C. 1989. The Archaeology of Doncaster. 2.
The medieval and later town. Oxford: Brit. Archaeol. Rep. Brit. Ser., 202
Buckland, P. C., Parker Pearson, M., Wigley, A. and Girling, M. A. 2001. Is there
anybody out there? A reconsideration of the environmental evidence from the
Breiddin Hillfort, Powys, Wales. Antiq. J., 81, 51-76
Buckland, P. I. 2007. The Development and Implementation of Software for
Palaeoenvironmental and Palaeoclimatological Research: The Bugs Coleopteran
Ecology Package (BugsCEP). PhD thesis, Environmental Archaeology Lab.
Department of Archaeology & Sámi Studies. University of Umeå, Sweden.
Archaeology and Environment 23, 236 pp + CD. URL: http://www.diva-
portal.org/umu/abstract.xsql?dbid=1105

38
Buckland, P. I. & Buckland, P. C. 2006. BugsCEP Coleopteran Ecology Package. IGBP
PAGES/World Data Center for Paleoclimatology Data Contribution Series #
2006-116. NOAA/NCDC Paleoclimatology Program, Boulder CO, USA. URL:
http://www.ncdc.noaa.gov/paleo/insect.html (Versions: BugsCEP v7.63;
Bugsdata v7.11; BugsMCR v2.02; BugStats v1.22) [Downloaded 1 1 11 from
www.bugscep.com]
Carlsson, D. 1981. Från stengrund til bulhus - gotländska husformer under yngre
järnalder-tidig medeltid. Ett rekonstruktionsförlag utifrån Fjäle i Ala.
Bebyggelsehistorisk tidskrift, 2, 37-47
Carrott, J., Dobney, K., Hall, A., Issitt, M., Jaques, D., Johnstone, C., Kenward, H. and
Milles, A. 1995. An evaluation of biological remains from excavations at 44-45
Parliament Street, York (site code: 1994.3210). Reports from the Environmental
Archaeology Unit, York 95/08.
Carrott, J., Fell, V., Large, F., Kenward, H. and Kent, P. 2000. Insects and other
invertebrate remains, in K. Buxton and C. Howard-Davies (eds.) Bremetenacum:
excavations at Roman Ribchester 1980, 1989-1990. Lancaster Imprint Ser. 9, 387-
400
Chowne, P., Girling, M. and Greig, J. 1986. Excavations at an Iron Age defended
enclosure at Tattershall Thorpe, Lincolnshire. Proc. Prehist. Soc., 52, 159-188
Clarke, H. and Carter, A. 1977. Excavations in Kings Lynn 1963-1970. London:
Medieval Archaeology Monograph 7
Clay, J. W. 1904. Yorkshire Church Notes, 1619-1631, by Roger Dodsworth. Yorkshire
Archaeol. Soc. Record Ser., 34. 4-9
Coope, G. R. 1986. Coleoptera analysis, in B. E. Berglund (ed.) Handbook of Holocene
Palaeoecology and Palaeohydrology, 703-713. Chichester: J. Wiley & Sons
Currie, C. K. 1991. The early history of the carp and its economic significance in
England. Agricultural History Rev., 39, 97-107
Dalton, P. 1994. Conquest, Anarchy and Lordship: Yorkshire 1066-1154. Cambridge:
Cambridge University Press
Davies, G. 1969, St Peter's church, Conisbrough, in Anon (ed.) Sprotbrough in local
history. Part 2. Doncaster: Workers’Educational Association Local History Group

39
Elliott, L., Jones, H., and Howard A. J. 2004. The medieval landscape, in D. Knight and
A. J. Howard (eds.) Trent Valley Landscapes, 153-182. Kings Lynn: Heritage
Marketing & Publications Ltd.
Fallgren, J.-H. 2008. Farm and village in the Viking age, in S. Brink, and N. Price (eds.)
The Viking World, 67-76. London: Routledge
Faull, M. L. 1977. British survival in Anglo-Saxon Northumbria, in L. Laing (ed.) Studies
in Celtic survival, 1-56. Oxford: Brit. Archaeol. Rep. Brit. Ser. 37
Faull, M. L. 1986. The decoration of the south doorway of Ledsham church tower. J.
Brit. Archaeol. Assoc., 139, 143-147
Faull, M. and Stinson, M. 1986. Domesday Book: Yorkshire (2 vols.). Chichester:
Philimore
Fowles, A. P. 1994. A review of the ecology of the red wood ant Formica rufa L.
(Hymenoptera, Formicidae) and its status in Wales. Cardiff: Countryside Council
for Wales
Garland, S. 1983. Beetles as Primary Woodland Indicators. Sorby Rec., 21, 3-38
Gearey, B. R., Hall, A. R., Kenward, H. K., Bunting, M. J., Lillie, M. C. and Carrott, J.
2005. Recent palaeoenvironmental evidence for the processing of hemp
(Cannabis sativa L.) in eastern England during the medieval period. Medieval
Archaeology, 49, 317-322
Goodburn, D. 1994. Beyond the postholes. Notes on stratigraphy and timber buildings
from a London perspective, in L. Shepherd (ed.) Interpreting stratigraphy 5.
Proceedings of a conference held at Norwich Castle Museum on Thursday 16th
June 1994, 43-52. Huntington: Witley Press.
Groves, C. and Hillam, J. (forthcoming). Tree-ring analysis of timbers excavated at York
Coppergate: Periods 3-5, in Hall, R. A. and Hunter-Mann, K., Early Medieval
Occupation at 16-22 Coppergate Archaeology of York. York: Counc. Brit.
Archaeol. for York Archaeol. Trust
Hadley, D. M. 1996. Multiple estates and the manorial structure of the northern
Danelaw'. J. Hist. Geog., 22, 3-15

40
Hæggström, C.-A. 1998. Pollard meadows: multiple use of human-made nature, in K. J.
Kirby and C. Watkins (eds.) The ecological history of European forests, 33-41.
Wallingford: CAB International.
Hall, A. R. and Kenward, H. K. 1990. Environmental evidence from the Colonia
Archaeology of York, 14/6. London: Counc. Brit. Archaeol. for York
Archaeological Trust
Hall, A. R. and Kenward, H. K. 2004. Setting people in their environment : plant and
animal remains from Anglo-Scandinavian York, in R. A. Hall, D. W. Rollason,
M. Blackburn, D. N. Parsons, G. Fellows Jensen, A. R. Hall, H. K. Kenward, T.
P. O'Connor, D. Tweddle, A. J. Mainman and N. S. H. Rogers (eds.), Aspects of
Anglo-Scandinavian York. York: Counc.Brit. Archaeol. for York Archaeol. Trust.
8/4: 372-426
Hall, A. R., Rowland, S., Kenward, H. K., Jaques, D. and Carrott, J. 2000. Evaluation of
biological remains from excavations on the land to the rear of 7-15 Spurriergate,
York (site code: 2000.584). Reports from the Environmental Archaeology Unit,
York 2000/80
Halstead, D. G. H. 1993. Keys for the identification of beetles associated with stored
products. II. Laemophloeidae, Passandrinae and Silvanidae. J. stored Prod. Res.,
29, 99-195
Hauck, K. 1963. Tiergärten im Pfalzbereich. Deutsche Königspfalzen Beiträge sur ihrer
historischen und archäologischen Erforschung 1, 30-74. Göttingen:
Veröffentlichtungen des Max-Planck-Institut für Geschichte.
Hey, D. 1979. The making of South Yorkshire. Ashbourne: Moorland Publ. Co.
Hey, D. 1986. Yorkshire from AD 1000. London: Longman
Hey, D. 2000. Yorkshire's southern boundary. Northern History, 37, 31-47
Hey, D. 2002. Historic Hallamshire. Ashbourne: Landmark Publishing
Hey, D. 2003. Medieval South Yorkshire. Ashbourne: Landmark Publishing
Higham, M. C. 1989. Some evidence for 12th- and 13th century linen and woollen textile
processing. Medieval Archaeology, 33: 38-52
Higham, N. J. 1993. The kingdom of Northumbria AD 350-1100. Stroud: Alan Sutton

41
Higham, N. J. 2000. King Edwin of the Deiri: rhetoric and the reality of power in early
England, in H. Geake and J. Kenny (eds.) Early Deira: archaeological studies of
the East Riding in the 4th to 9th centuries AD, 41-50. Oxford: Oxbow Books
Higham, N. J. 2006. Northumbria's southern frontier: a review. Early Medieval Europe,
14, 391-418
Hooke, D. 1985. The Anglo-Saxon Landscape: the Kingdom of the Hwicce. Manchester:
Manchester University Press
Hooke, D. 1988. Regional variation in southern and central England in the Anglo-Saxon
period and its relationship to land units and settlement, in D. Hooke (ed.) Anglo-
Saxon Settlements, 123-152. Oxford: Blackwell
Hooke, D. 1998. Medieval forests and parks in southern and central England, in C.
Watkins (ed.) European woods and forests. Studies in cultural history, 19-32.
Wallingford: CAB International
Hope-Taylor, B. 1977. Yeavering: An Anglo-British Centre of Northumbria. London:
HMSO
Hunter Blair, P. 1948. The Northumbrians and their southern frontier. Archaeol. Aeliana,
fourth ser., 26, 98-126
Hunter, J. 1828-31. SouthYorkshire: the history and topography of the Deanery of
Doncaster (2 vols.). London: J. B. Nichols & son for the author.
Hyman, P. S. 1992. Review of the scarce and threatened Coleoptera of Great Britain.
Part 1. Peterborough: Joint Nature Conservancy Council
Hyman, P. S. 1994. Review of the scarce and threatened Coleoptera of Great Britain.
Part 2. Peterborough: Joint Nature Conservancy Council
James, S., Marshall, A. and Millett, M. 1984. An early medieval building tradition.
Archaeol. J., 141, 182-215
Johnson, S. 1980. Excavations at Conisbrough Castle 1973-1977. Yorks. Archaeol. J., 52,
59-88
Jones, G. R. J. 1965. Early territorial organization in Northern England and its bearing on
the Scandinavian settlement, in A. Small (ed.) Fourth Viking Congress, York
1961, 67-84. Edinburgh: Oliver & Boyd for University of Aberdeen

42
Jones, G. R. J. 1975. Early territorial organisation in Gwynedd and Elmet. Northern
History, 10, 3-27
Kenward, H. K., Williams, D., Spencer, P. J., Greig, J. R. A., Rackham, D. J. and
Brinklow, D. A. 1978. The environment of Anglo-Scandinavian York, in R. Hall
(ed.) Viking Age York and the North. London: Counc. Brit. Archaeol.Res. Rep.
27, 58-70
Kenward, H. K. and Hall, A. R. 1995. Biological evidence from 16-22 Coppergate.
Archaeology of York, 14/7. York: Counc. Brit. Archaeol. for York Archaeol.
Trust
Kiess, R. 1998. The word 'Forst/forest' as an indicator of fiscal property and possibly
consequences for the history of Western European forests, in C. Watkins (ed.)
European woods and forests. Studies in cultural history, 11-18. Wallingford:
CAB International
Koch, K. 1989-92. Die Käfer Mitteleuropas. Ökologie, 1-3. Krefeld: Goecke & Evers.
Liddiard, R. 2003. The deer parks of Domesday Book. Landscapes, 4, 4-23
May, T. 1922. The Roman Forts of Templeborough near Rotherham. Rotherham:
Rotherham Borough Council
Meeson, R. 1983. Plank-walled building techniques and the church of St. Lawrence,
Rushton Spencer. Vernacular Architecture, 14, 29-35
Miller, E. 1804. The history and antiquities of Doncaster and its vicinity. Doncaster: W
Sheardown
Mitchell, G. H., Stephens, J. V., Bromehead, C. E. N. and Wray, D. A. 1947. Geology of
the country around Barnsley. Explanation of Sheet 87. London: British
Geological Survey, HMSO
Morris, R. K. 1988. Churches in York and its hinterland: building patterns and stone
sources in the 11th and 12th centuries, in J. Blair (ed.) Minsters and Parish
Churches: the local church in transition 950-1200, 191-200. Oxford: Oxford
University Press
Neighbour, T. 2007. A semi-elliptical timber-framed structure at Inveresk (the most
northerly amphitheatre in the Empire?). Britannia, 38, 125-140

43
Nilsson, A. N. and Holmen, M. 1985. The aquatic Adephaga (Coleoptera) of
Fennoscandia and Denmark. II. Dytiscidae. Leiden: E. J. Brill
Nitz, H.-J. 1988. Settlement structures and settlement systems of the Frankish central
state in Carolingian and Ottonian times, in D. Hooke (ed.) Anglo-Saxon
Settlements, 249-274. Oxford: Blackwell.
O'Brien, C. and Miket, R. 1991. The early medieval settlement at Thirlings,
Northumberland. Durham Archaeol. J., 7, 57-91
O’Neill, R. 2004. Final report of archaeological investigations at Wellgate (Areas A, B
and C), Conisbrough, Doncaster, South Yorkshire. ARCUS unpublished report
670c.6. Sheffield: South Yorkshire Archaeology Service Sites and Monuments
Record, Sheffield and Doncaster Museum
Osborne, P. J. 1989. Insects, in P. Ashbee, M. Bell and E. Proudfoot (eds.) Wilsford shaft
excavations 1960-62, 96-99. London: English Heritage
Oswald, A. 1964. Excavation of a thirteenth-century wooden building at Weoley Castle,
Birmingham, 1960-61. Medieval Archaeology, 6-7, 109-134
Päivinen, J., Ahlroth, P., Kaitala, V., Kotiaho, J. S., Suhonen, J. and Virola, T. 2003.
Species richness and regional distribution of myrmecophilous beetles. Oecologia,
134, 587-595
Rackham, O. 2003. Ancient woodland - its history, vegetation and uses in England. (new
ed) Dalbeattie: Castlepoint Press
Rahtz, P. 1976. Buildings and rural settlement, in D. M. Wilson (ed.) The archaeology of
Anglo-Saxon England, 49-98. Cambridge: Cambridge University Press
Rahtz, P. 1979. The Saxon and medieval palaces at Cheddar. Oxford: Brit. Archaeol.
Rep. Brit. Ser. 65
Rahtz, P. and Meeson, R. 1992. An Anglo-Saxon Watermill at Tamworth: Excavations in
the Bolebridge Street area of Tamworth, Staffordshire in 1971 and 1978. London:
Counc. Brit. Archaeol. Res. Rep. 83
Revill, S. 1975. King Edwin and the Battle of Heathfield. Trans. Thoroton Soc. Notts.,
79, 40-49
Roberts, I. 2010. Understanding the cropmark landscapes of the Magnesian Limestone.
Wakefield: Archaeological Services WYAS and English Heritage

44
Roberts, I., Burgess, A. and Berg, D. (eds.) 2001. A new link to the past. The
archaeological landscape of the M1-A1 link road. Yorkshire Archaeology 7.
Wakefield: West Yorkshire Archaeology Service.
Roede, L. 1999. Grindbygg og bindingsverk, in H. Schjelderup and O. Storstetten (eds.)
Grindbygde hus i Vest-Norge, 113-121. Oslo: Byggens Museum
Rollason, D. W. 2002. Northumbria. The rise and fall of a kingdom 500-1100.
Cambridge: Cambridge University Press
Rowley, T. 1984. The Norman heritage 1066-1200. London: Granada
Ryder, P. 1980. St Peter's Church, Conisbrough (SK 512988). Programme of the Summer
Meeting at Wentworth Woodhouse, Monday 14 July to Saturday 19 July, 44-52.
London: Royal Archaeol. Inst.
Ryder, P. F. 1982. Saxon Churches in South Yorkshire. Sheffield: South Yorkshire
Archaeological Service
Sawyer, P. H. 1979. The charters of Burton Abbey. Oxford: Oxford University Press
Smith, A. H. (ed.) 1961. The placenames of the West Riding of Yorkshire. Lower and
Upper Strafforth and Staincross Wapentakes. 1. Survey of English Place Names
30. Cambridge: Cambridge University Press.
Smith, J. T. 1965. The structure of the timber kitchen at Weoley Castle, Birmingham.
Medieval Archaeology, 9, 82-93
Sykes, N. 2004. The introduction of fallow deer to Britain: a zooarchaeological
perspective. Environmental Archaeol., 9, 75-84
Taylor, H. M. and Taylor, J. 1965. Anglo-Saxon Architecture. I and II. Cambridge:
Cambridge University Press
Tyers, I. 1998. Tree-ring analysis and wood identification of timbers excavated on the
Magistrates Court Site, Kingston upon Hull, East Yorkshire, Sheffield: ARCUS
Rep. 410
Tyers, I. 2004 Dendro for Windows program guide 3rd edn, Sheffield: ARCUS Rep.
500b
Tyers, I., Hillam, J. and Groves, C. 1994. Trees and woodland in the Saxon period: the
dendrochronological evidence, in J. Rackham (ed.) Environment and Economy in
Anglo-Saxon England, London: Counc. Brit. Archaeol. Res. Rep. 89, 12-22

45
Wainwright, J. 1829. An historical and topographical introduction to a knowledge of the
ancient state of the wapentake of Strafford and Tickhill: with ample accounts of
Doncaster and Conisbrough.... Sheffield
Walker, J. 1999. Late-Twelfth and Early-Thirteenth-Century Aisled Buildings: A
Comparison. Vernacular Architecture, 30, 21-53
Webster, L. E. and Cherry, J. 1975. Medieval Britain in 1974. Medieval Archaeology, 19,
220-260
Whitelock, D. 1979. English Historical Documents. I. 500-1042 (2nd ed.). Oxford:
Oxford University Press.
Whitwell, J. B. 1982. The Coritani. Some aspects of the Iron Age tribe and the Roman
civitas. Oxford: Brit. Archaeol. Rep. Brit. Ser. 99
Williams, J., Shaw, M. and Denham, V. 1985. Middle Saxon Palaces at Northampton.
Northampton: Northampton Development Corporation
Zadora-Rio, E. 2003. The making of churchyards and parish territories in the early
medieval landscape of France and England in the 7th - 12th centuries: a
reconsideration. Medieval Archaeology, 47, 1-20
Zeepvat, R. J. 1988. Fishponds in Roman Britain, in M. Aston (ed.) Medieval fish,
fisheries and fishponds in England, 17-26. Oxford: Brit. Archaeol. Rep Brit. Ser.
182

46
List of Figures
Fig. 1. Conisbrough, South Yorkshire location map. The base map is the First Edition
Ordnance Survey Map of 1843, with the Doncaster New Road (Tinsley Turnpike) of
1764 removed to emphasise the line of the Old Road and Clifton Hill marking the
northern limit of the medieval deer park. The northern edge of the park boundary, as
indicated on the tithe award map of 1848, is indicated by a dotted line in the lower left of
the map (see fig. 3).
Fig. 2. Conisbrough, South Yorkshire, May 2011: The village from the north east, viewed
across the river Don. Although the foreground has been infilled with modern housing, the
siting of the castle and parish church on a long, narrow promontory is still evident.
Conisbrough Parks, the site of the medieval deer park, lies to the south.
Fig. 3. The outline of Conisbrough Parks from the 1848 tithe award superimposed on the
modern map. The modern estate of Conanby, west of Conisbrough, and the lines of
dismantled railways have been removed to make the park limits clearer, although the line
of the M18 motorway cutting through the Lower Magnesian Limestone escarpment
below Micklebring in the south-east corner of the map has been retained. Conisbrough
parish church and the castle are the top right hand corner of the map, linked to the park
by the line of Old Road (see fig. 1). Note the intakes of Firsby to the west and Clifton to
the east cutting into the Park. (Reproduced with the permission of the Controller of Her
Majesty’s Stationary Office. © Crown Copyright Licence No. AL 50228A).
Fig. 4. Conisbrough, South Yorkshire: detailed trench location plan. The structures
discussed in this paper were located in Trench 4 (reproduced with the permission of the
Controller of Her Majesty’s Stationary Office. © Crown Copyright Licence No. AL
50228A).

47
Fig. 5. Excavations in Wellgate, Conisbrough in progress, October 2002, Trench 4
looking west. Work was constrained by 1960s housing units, since demolished. The cut
and dark, organic infill of the medieval ditch is apparent in the section and the post and
plank structure occurs centre right.
Fig. 6. Wellgate. Conisbrough, South Yorkshire: Site plan (Trench A/4)
Fig. 7. Wellgate, Conisbrough, South Yorkshire: Sections 10, 43 and 50 in Trench 4
showing the post and plank structure [4048] cut through by the broad ditch [4055 et
seq.].
Fig. 8. Wellgate, Conisbrough, South Yorkshire: Oblique photograph looking northwest.
The thick, horizontal oak planks have been fitted into grooves cut in the upright oak posts
and are fitted to the overlying planks by a continuous tongue locating into a groove
(Scale 2m).
Fig. 9. Wellgate, Conisbrough, South Yorkshire: Face view of plank and upright at west
end of trench. A fragment of a second layer of planking sits over a tongue in the upper
edge of the lower plank (Scale 1m).
Fig. 10. Conisbrough, South Yorkshire: Plank and post structure elevation (Section 48)
Fig. 11.Bar diagram showing the relative and absolute positions of the dated
dendrochronological samples from the Wellgate site, Conisbrough. Each bar is annotated
with an interpretation based on the date of the ring sequence and the presence of
sapwood.
Fig 12. Bugs bar chart of habitat frequency of species in the Conisbrough insect
assemblages (see Appendix 2 for explanation).
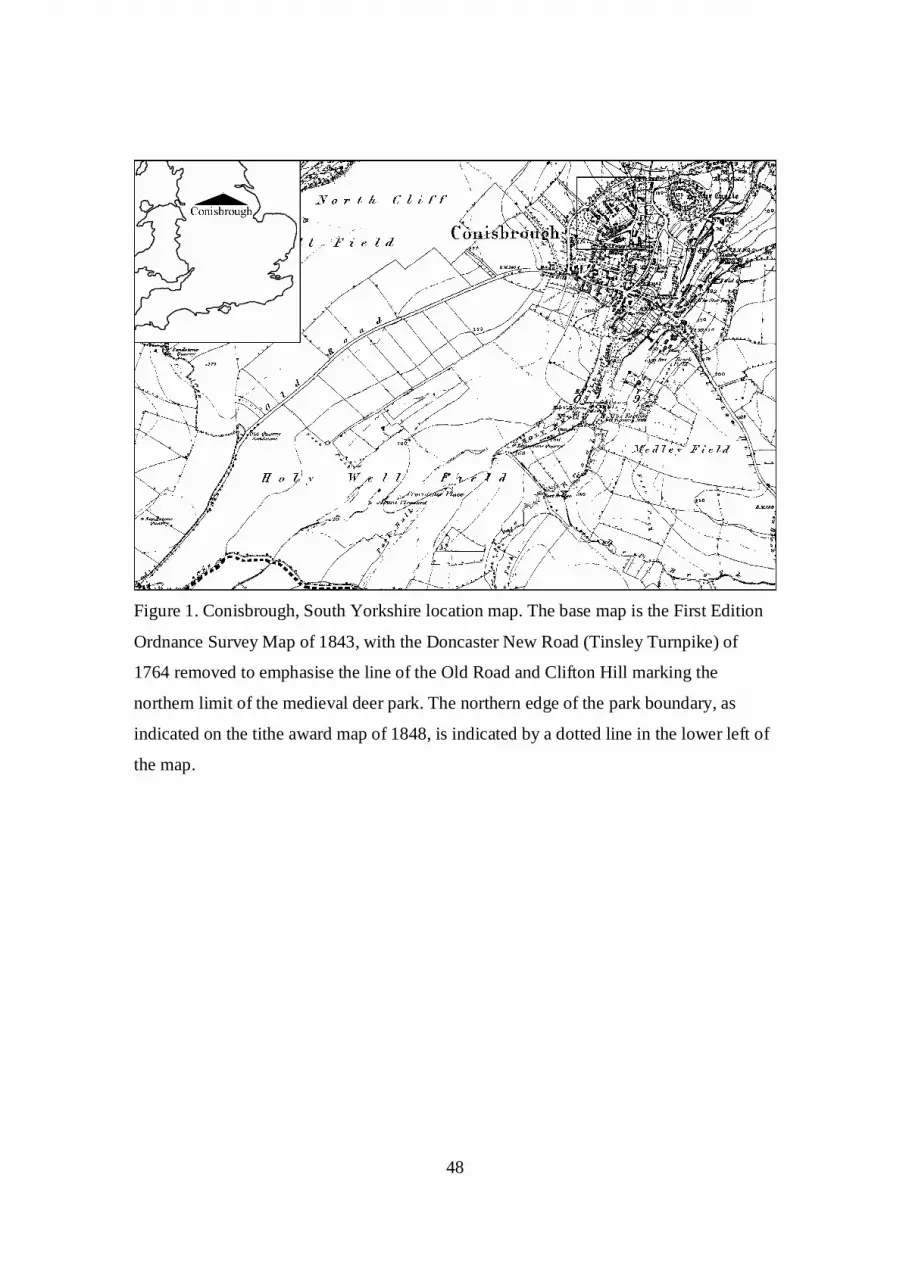
48
Figure 1. Conisbrough, South Yorkshire location map. The base map is the First Edition
Ordnance Survey Map of 1843, with the Doncaster New Road (Tinsley Turnpike) of
1764 removed to emphasise the line of the Old Road and Clifton Hill marking the
northern limit of the medieval deer park. The northern edge of the park boundary, as
indicated on the tithe award map of 1848, is indicated by a dotted line in the lower left of
the map.

49
Fig. 2. Conisbrough, South Yorkshire, May 2011: The village from the north east, viewed
across the river Don. Although the foreground has been infilled with modern housing, the
siting of the castle and parish church on a long, narrow promontory is still evident.
Conisbrough Parks, the site of the medieval deer park, lies to the south.

50
Fig. 3. The outline of Conisbrough Parks from the 1848 tithe award superimposed on the
modern map. The modern estate of Conanby, west of Conisbrough, and the lines of
dismantled railways have been removed to make the park limits clearer, although the line
of the M18 motorway cutting through the Lower Magnesian Limestone escarpment
below Micklebring in the south-east corner of the map has been retained. Conisbrough
parish church and the castle are in the top right hand corner of the map, linked to the park
by the line of Old Road (see fig. 1). Note the intakes of Firsby to the west and Clifton to
the east cutting into the Park. (Reproduced with the permission of the Controller of Her
Majesty’s Stationary Office. © Crown Copyright Licence No. AL 50228A).

51
Fig. 4. Conisbrough, South Yorkshire: detailed trench location plan. The structures
discussed in this paper were located in Trench 4 (reproduced with the permission of the
Controller of Her Majesty’s Stationary Office. © Crown Copyright Licence No. AL
50228A).

52
Fig. 5. Excavations in Wellgate, Conisbrough in progress, October 2002, Trench 4
looking west. Work was constrained by 1960s housing units, since demolished. The cut
and dark, organic infill of the medieval ditch is apparent in the section and the post and
plank structure occurs centre right.
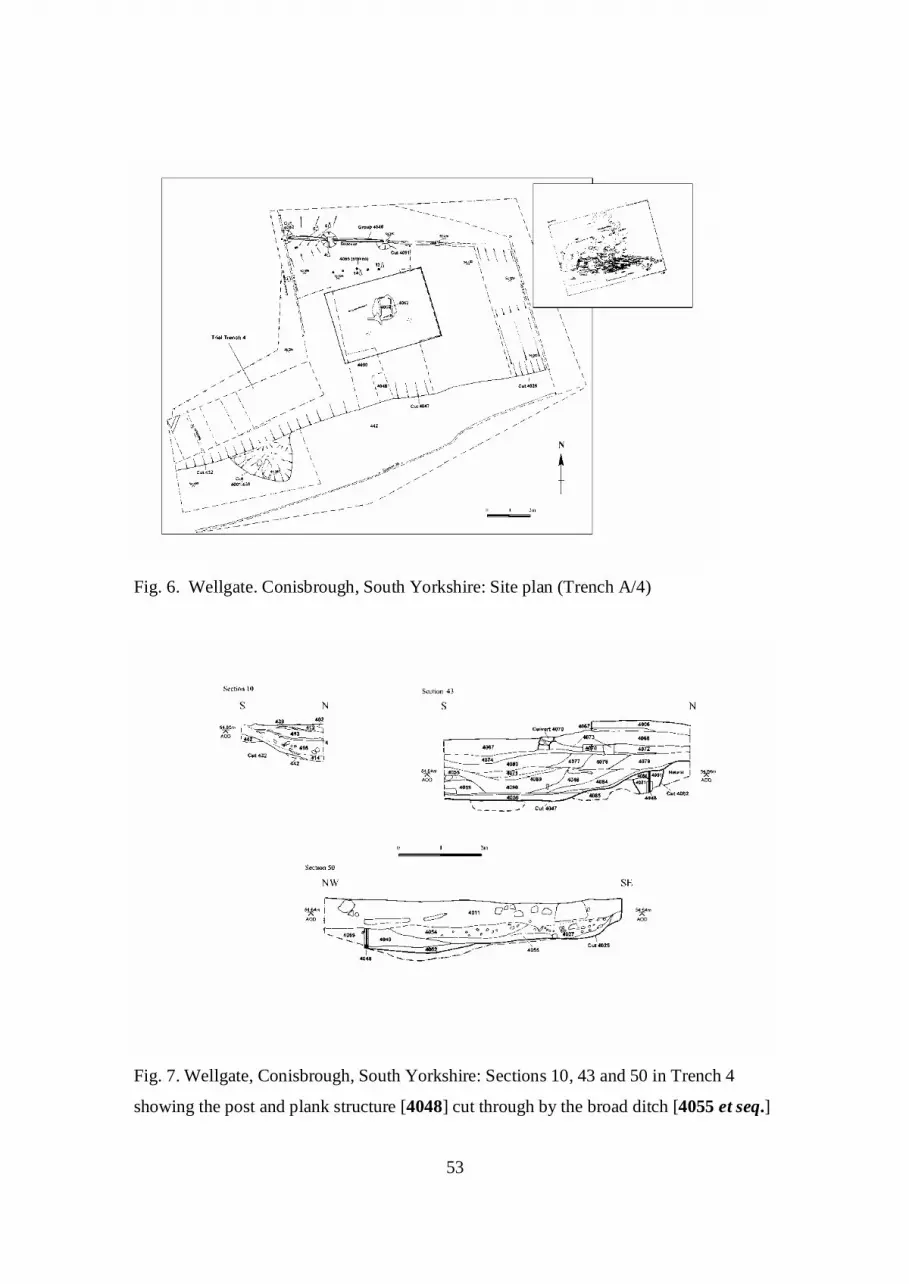
53
Fig. 6. Wellgate. Conisbrough, South Yorkshire: Site plan (Trench A/4)
Fig. 7. Wellgate, Conisbrough, South Yorkshire: Sections 10, 43 and 50 in Trench 4
showing the post and plank structure [4048] cut through by the broad ditch [4055 et seq.]

54
Fig. 8. Wellgate, Conisbrough, South Yorkshire: Oblique photograph looking northwest.
The thick, horizontal oak planks have been fitted into grooves cut in the upright oak posts
and are fitted to the overlying planks by a continuous tongue locating into a groove
(Scale 2m).

55
Fig. 9. Wellgate, Conisbrough, South Yorkshire: Face view of plank and upright at west
end of trench. A fragment of a second layer of planking sits over a tongue in the upper
edge of the lower plank (Scale 1m).
Fig. 10. Conisbrough, South Yorkshire: Fence elevation (Section 48)

56
Fig. 11. Bar diagram showing the relative and absolute positions of the dated samples
from the Wellgate site, Conisbrough. Each bar is annotated with an interpretation based
on the date of the ring sequence and the presence of sapwood.
Wellgate, Conisbrough
Calendar Years
Span of ring sequences
AD500 AD450 AD550
Trench 4 413 SF18 after AD554
Trench A 4048 SF4 after AD522 SF5 after AD558
SF6 after AD567
Trench A 4062 SF7 after AD547 SF15 AD573-90
Trench A 4064 SF14 AD565-601?
KEY
heartwood sapwood felled after felled between

57
Fig. 12. Bugs bar chart of habitat frequency of species in the Conisbrough insect
assemblages (see Appendix 2 for explanation).

58
Trench/Context/S
F
Rings Sap or
bark
ARW
Date of sequence Interpretation
A 4047 SF12 66 - 0.82 undated -
A 4048 SF4 88 - 1.64 AD425-AD512 after AD522
A 4048 SF5 119 - 1.52 AD430-AD548 after AD558
A 4048 SF6 103 - 1.73 AD455-AD557 after AD567
A 4062 SF15 89 29 0.74 AD485-AD573 AD573-90
A 4062 SF7 92 - 2.41 AD446-AD537 after AD547
A 4064 SF1 87 21+Bw 1.48 undated -
A 4065 SF14 83 ?H/S 1.15 AD473-AD555 AD565-601?
4 413 SF18 61 - 1.33 AD484-AD544 after AD554
KEY
ARW: average ring width (mm/year)
Bw: winter felled
?H/S: ends at the heartwood/sapwood boundary
The material is Oak: = Quercus spp.
Table 1. Dendrochronological details of the analysed samples from the Wellgate site
(A670), Conisbrough
Radiocarbon
Age BP
Calibrated Age
range d1
Calibrated Age
range d2
Delta 13C
rel. PBD
(%)
Lab code
1430±60 AD580-660 (68%) AD530-690 (95%) -25.0‰ Beta-183712
1550±70 AD420-600 (68%) AD380-650 (95%) -25.0‰ Beta-183813
1660±70 AD330-440 (68%) AD230-550 (95%) -25.0‰ Beta-185185
Table 2. Radiocarbon dates from Context 413
59
Sample no. s413 s439 s416/s0 s416/s10 s416/s20 s416/s30 s416/s35 s4052
Coleoptera Carabidae Nebria brevicollis (F.) 0 0 0 1 0 0 1 0 Clivina fossor (L.) 1 0 1 0 0 0 0 0 C. collaris (Hbst.) 0 0 0 0 0 1 0 0 Clivina sp. 0 0 0 1 0 0 0 0 Trechus quadristriatus (Schr.) 0 0 1 0 0 0 1 0 T. obtusus Er. 0 0 0 0 0 1 0 0 T. obtusus/quadristriatus Er./(Schr.) 0 0 0 0 0 0 0 1 Bembidion lampros (Hbst.) 1 0 0 0 0 0 0 0 B. clarkii (Dawson) 0 0 0 1 0 0 0 0 B. mannerheimi Sahl. 0 1 0 0 0 0 0 0 Bembidion sp. 0 0 0 0 1 0 0 0 Pseudoophonus rufipes (Deg.) 0 0 1 0 0 0 0 0 Poecilus sp. 0 0 0 0 0 0 0 1 Pterostichus melanarius (Ill.) 1 0 0 0 0 1 0 0 Amara sp. 1 0 0 0 0 0 0 0 Dytiscidae Hydroporus spp. 0 0 0 0 0 0 0 3 Agabus bipustulatus (L.) 0 0 0 0 0 0 0 1 Dytiscus circumcinctus Ahr. 1 0 0 0 0 0 0 0 Hydraenidae Hydraena sp. 0 0 0 0 0 0 0 1 Ochthebius sp. 0 0 0 0 0 0 0 5 Limnebius truncatellus (Thun.) 0 0 0 0 0 0 0 2 Hydrophilidae Helophorus spp. 1 0 1 0 0 0 0 3 Sphaeridium sp. 1 0 0 0 0 0 0 0 Cercyon atricapillus (Marsh.) 0 0 1 1 0 1 0 0 C. analis (Payk.) 9 3 5 5 2 4 2 0 Cercyon spp. 4 1 2 4 3 1 1 0 Megasternum obscurum (Marsh.) 4 0 1 3 1 3 2 4 Cryptopleurum minutum (F.) 2 0 0 4 1 1 0 0 Enochrus sp. 0 0 0 0 0 0 0 2 Histeridae Onthophilus striatus (Müll.) 0 0 0 0 1 0 0 0 Abraeus perpusillus (Marsh.) 1 0 3 2 1 2 2 0 Aeletes atomarius (Aubé) 0 0 0 1 0 0 0 0 Acritus nigricornis (Hoff.) 5 1 3 4 0 1 1 0 Hister impressus F. 0 0 0 0 1 0 0 0 Hister sp. 1 0 0 0 0 0 0 0 Atholus duodecimstriatus (Schrank) 0 0 0 1 0 0 0 0 Silphidae Ablattaria laevigata (F.) 0 0 0 0 0 0 1 0 Catopidae Choleva/Catops sp. 1 0 1 0 1 1 0 1 Clambidae Clambus sp. 0 0 1 0 0 0 1 0

60
Scydmaenidae Neuraphes sp. 0 0 1 0 0 1 1 0 Stenichnus sp. 0 0 2 0 0 0 0 0 Scydmaenus tarsatus Müll. & Kunze 1 0 0 1 0 0 0 0 Orthoperidae Sericoderus lateralis (Gyll.) 2 0 1 1 1 2 0 0 Ptiliidae Ptenidium fuscicorne Er. 1 0 0 0 0 0 0 0 Ptenidium sp. 2 1 0 4 0 2 0 0 Oligella sp. 0 1 0 0 0 0 0 0 Acrotrichis sp. 5 0 2 0 0 0 2 2 Staphylinidae Scaphisoma agaricinum (L.) 0 0 1 0 0 0 0 0 Micropeplus tesserula Curt. 2 2 3 2 0 0 3 2 M. staphylinoides (Marsh.) 1 0 2 5 0 0 0 0 M. fulvus Er. 2 0 7 2 0 2 1 1 Megarthrus prosseni Schatz. 0 1 0 0 0 0 0 0 M. depressus (Payk.) 0 1 0 0 0 0 0 0 Proteinus ovalis Steph. 0 0 1 0 0 0 0 0 Acrulia inflata (Gyll.) 0 0 0 0 1 0 0 0 Phyllodrepa ioptera (Steph.) 0 0 0 0 0 0 0 1 P. vilis (Er.) 0 0 0 0 0 0 1 0 Omalium rivulare (Payk.) 1 0 0 3 0 1 0 1 O. caesum Grav. 0 1 1 0 1 0 0 0 O. excavatum Steph. 0 0 0 0 0 0 0 1 Omalium spp. 0 0 0 0 0 1 0 2 Xylodromus depressus (Grav.) 0 1 0 0 0 0 0 0 X. concinnus (Marsh.) 2 2 4 6 2 3 2 0 Acidota cruentata Mann. 0 0 0 0 0 0 0 1 Lesteva punctata Er. 0 0 0 0 0 0 0 2 L. pubescens Mann. 0 0 0 0 0 0 0 1 Carpelimus bilineatus (Steph.) 3 0 2 2 2 0 2 0 C. fuliginosus (Grav.) 0 0 0 1 0 0 0 0 Carpelimus spp. 0 2 1 0 1 0 2 0 Oxytelus sculptus Grav. 0 0 0 1 0 0 1 0 O. laqueatus (Marsh.) 0 0 0 0 0 0 1 0 Anotylus rugosus (F.) 3 2 0 3 1 2 1 0 A. inustus (Grav.) 0 0 2 0 0 0 0 0 A. sculpturatus (Grav.) 5 0 1 2 0 0 0 1 A. nitidulus (Grav.) 2 2 9 4 1 1 1 4 A. tetracarinatus Block 1 0 0 3 0 0 2 0 Platystethus arenarius (Geoff.) 1 0 1 1 1 2 1 0 Stenus spp. 2 0 1 1 1 1 2 3 Rugilus rufipes (Germ.) 0 1 0 0 0 0 0 0 R.. orbiculatus (Payk.) 1 0 1 2 0 1 0 0 Lithocharis ochracea (Grav.) 3 0 3 1 1 3 0 0 Lithocharis sp. 0 0 0 1 0 0 0 0 Lathrobium (s.l.) sp. 0 0 0 0 0 0 0 1 Leptacinus intermedius Donis. 0 0 0 0 0 1 0 0 L. batychrus (Gyll.) 0 0 2 2 0 0 0 0 L. pusillus (Steph.) 4 0 0 2 2 0 1 0 Phacophallus parumpunctatus (Gyll.) 0 0 0 1 0 0 0 0
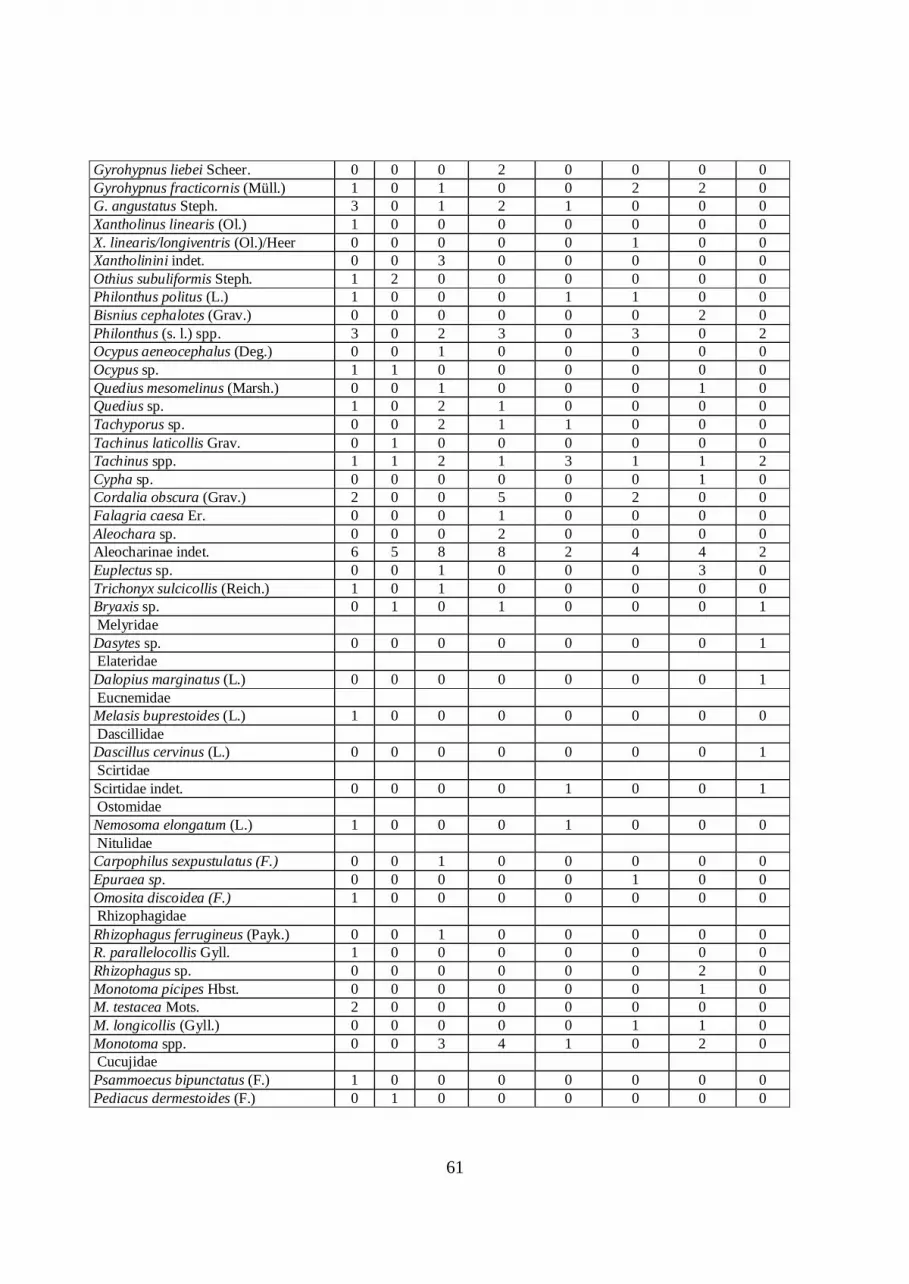
61
Gyrohypnus liebei Scheer. 0 0 0 2 0 0 0 0 Gyrohypnus fracticornis (Müll.) 1 0 1 0 0 2 2 0 G. angustatus Steph. 3 0 1 2 1 0 0 0 Xantholinus linearis (Ol.) 1 0 0 0 0 0 0 0 X. linearis/longiventris (Ol.)/Heer 0 0 0 0 0 1 0 0 Xantholinini indet. 0 0 3 0 0 0 0 0 Othius subuliformis Steph. 1 2 0 0 0 0 0 0 Philonthus politus (L.) 1 0 0 0 1 1 0 0 Bisnius cephalotes (Grav.) 0 0 0 0 0 0 2 0 Philonthus (s. l.) spp. 3 0 2 3 0 3 0 2 Ocypus aeneocephalus (Deg.) 0 0 1 0 0 0 0 0 Ocypus sp. 1 1 0 0 0 0 0 0 Quedius mesomelinus (Marsh.) 0 0 1 0 0 0 1 0 Quedius sp. 1 0 2 1 0 0 0 0 Tachyporus sp. 0 0 2 1 1 0 0 0 Tachinus laticollis Grav. 0 1 0 0 0 0 0 0 Tachinus spp. 1 1 2 1 3 1 1 2 Cypha sp. 0 0 0 0 0 0 1 0 Cordalia obscura (Grav.) 2 0 0 5 0 2 0 0 Falagria caesa Er. 0 0 0 1 0 0 0 0 Aleochara sp. 0 0 0 2 0 0 0 0 Aleocharinae indet. 6 5 8 8 2 4 4 2 Euplectus sp. 0 0 1 0 0 0 3 0 Trichonyx sulcicollis (Reich.) 1 0 1 0 0 0 0 0 Bryaxis sp. 0 1 0 1 0 0 0 1 Melyridae Dasytes sp. 0 0 0 0 0 0 0 1 Elateridae Dalopius marginatus (L.) 0 0 0 0 0 0 0 1 Eucnemidae Melasis buprestoides (L.) 1 0 0 0 0 0 0 0 Dascillidae Dascillus cervinus (L.) 0 0 0 0 0 0 0 1 Scirtidae Scirtidae indet. 0 0 0 0 1 0 0 1 Ostomidae Nemosoma elongatum (L.) 1 0 0 0 1 0 0 0 Nitulidae Carpophilus sexpustulatus (F.) 0 0 1 0 0 0 0 0 Epuraea sp. 0 0 0 0 0 1 0 0 Omosita discoidea (F.) 1 0 0 0 0 0 0 0 Rhizophagidae Rhizophagus ferrugineus (Payk.) 0 0 1 0 0 0 0 0 R. parallelocollis Gyll. 1 0 0 0 0 0 0 0 Rhizophagus sp. 0 0 0 0 0 0 2 0 Monotoma picipes Hbst. 0 0 0 0 0 0 1 0 M. testacea Mots. 2 0 0 0 0 0 0 0 M. longicollis (Gyll.) 0 0 0 0 0 1 1 0 Monotoma spp. 0 0 3 4 1 0 2 0 Cucujidae Psammoecus bipunctatus (F.) 1 0 0 0 0 0 0 0 Pediacus dermestoides (F.) 0 1 0 0 0 0 0 0

62
Cryptolestes ferrugineus (Steph.) 0 0 1 0 1 0 1 0 Cryptophagidae Cryptophagus scutellatus Newm. 1 0 4 1 0 1 1 0 C. lycoperdi (Scop.) 0 0 0 1 0 0 0 0 Cryptophagus spp. 3 0 2 3 1 0 1 1 Atomaria spp. 2 2 0 0 0 3 0 1 Ootypus globosus (Waltl) 1 0 0 0 0 0 0 0 Latridiidae Latridius minutus (L.) (grp.) 1 0 4 2 2 0 1 0 L. pseudominutus (Strand) 0 0 0 0 0 1 0 0 Latridius sp. 0 0 0 0 0 1 0 0 Enicmus fungicola Thom. 0 1 0 0 0 0 0 0 E. transversus (Ol.) 0 0 0 0 0 0 1 0 Enicmus sp. 0 0 0 0 0 0 2 0 Corticaria punctulata Marsh. 1 0 0 0 0 0 0 0 C. crenulata (Gyll.) 1 2 1 1 0 0 0 0 C. fulva (Com.) 0 0 0 1 0 0 0 0 Corticaria spp. 0 0 0 3 0 0 0 0 Corticariinae indet. 3 0 3 2 1 2 0 1 Colydiidae Myrmechoxenus subterraneus (Chevr.) 0 0 0 0 0 0 0 1 Cerylon histeroides (F.) 0 0 0 1 0 0 0 0 Mycetaea subterranea (Marsh.) 0 0 0 2 0 0 0 0 Anobiidae Grynobius planus (F.) 0 0 1 1 0 0 0 0 Anobium sp. 1 1 0 0 2 0 1 1 Hemicoelus fulvicornis (Strm.) 0 0 1 0 0 0 0 0 Ptinus fur (L.) 1 0 1 1 0 0 0 0 Ptinus sp. 1 0 1 1 1 0 2 0 Anthicidae Omonadus formicarius (Goeze) 5 0 3 3 0 0 0 0 Anthicus sp. 1 1 0 0 0 1 1 0 Scarabaeidae Trox scaber (L.) 4 0 1 3 1 0 0 0 Geotrupes stercorarius (L.) 0 0 0 1 0 0 0 0 Geotrupes (s.l.) sp. 0 1 0 0 0 0 1 1 Oxyomus sylvestris (Scop.) 0 0 0 0 0 1 0 0 Aphodius prodromus (Brahm) 1 0 0 0 0 0 0 1 A. ater (Deg.) 1 0 0 0 0 0 0 0 Aphodius spp. 0 0 2 1 1 0 0 2 Melolontha hippocastani F. 0 0 0 0 0 0 3 0 Cetonia aurata (L.) 0 0 1 0 0 0 0 0 Lucanidae Sinodendron cylindricum (L.) 0 0 1 0 0 0 0 0 Chrysomelidae Gastrophysa viridula (Deg.) 0 0 1 0 0 1 0 0 Phyllotreta sp. 0 1 0 0 0 0 0 0 Longitarsus sp. 0 0 1 0 0 0 0 1 Chaetocnema concinna (Marsh.) 1 0 0 1 0 0 0 0 C. hortensis (Geoff.) 0 0 0 1 0 1 0 0 Chaetocnema sp. 0 0 1 0 0 0 0 0 Scolytidae

63
Scolytus multistriatus (Marsh.) 0 0 1 1 0 0 0 0 Scolytus sp. 1 0 0 0 0 0 0 0 Hylesinus oleiperda (F.) 0 0 0 0 0 0 0 1 Leperisinus sp. 1 0 0 1 2 0 1 1 Dryocoetinus villosus (F.) 0 0 0 1 0 0 0 0 D. alni (Georg) 0 0 0 0 0 0 1 0 Xyleborus dryographus (Ratz.) 3 0 4 1 2 2 1 0 Platypodidae Platypus cylindrus (F.) 0 0 1 0 0 0 0 0 Curculionidae Apion frumentarium L. 0 0 0 1 0 0 0 0 Apion (s.l.) sp. 0 1 0 0 0 0 0 0 Phyllobius/Polydrusus sp. 0 0 0 0 0 0 0 1 Trachyphloeus sp. 0 0 0 0 0 0 0 1 Polydrusus cervinus (L.) 1 0 0 0 0 0 0 0 P. mollis (Ström.) 0 0 0 0 0 0 0 1 Barynotus moerens (F.) 0 0 1 0 0 0 0 0 Sitona lineellus (Bons.) 0 0 0 1 0 0 0 0 Sitona sp. 0 1 0 0 0 0 0 0 Phloeophagus lignarius (Marsh.) 1 0 0 0 1 1 0 0 Tychius sp. 0 0 0 1 0 0 0 0 Hypera meles (F.) 0 0 0 0 0 0 1 0 Nedyus quadrimaculatus (L.) 1 0 0 0 0 0 0 1 Trichoptera Trichoptera larvae indet. 0 0 0 0 0 0 0 2 Siphonaptera Siphonaptera indet. 1 0 2 2 0 0 0 0 Hymenoptera Formicidae Myrmica sp. 1 0 1 0 0 0 0 0
Table 3: Insect remains from Wellgate, Conisbrough
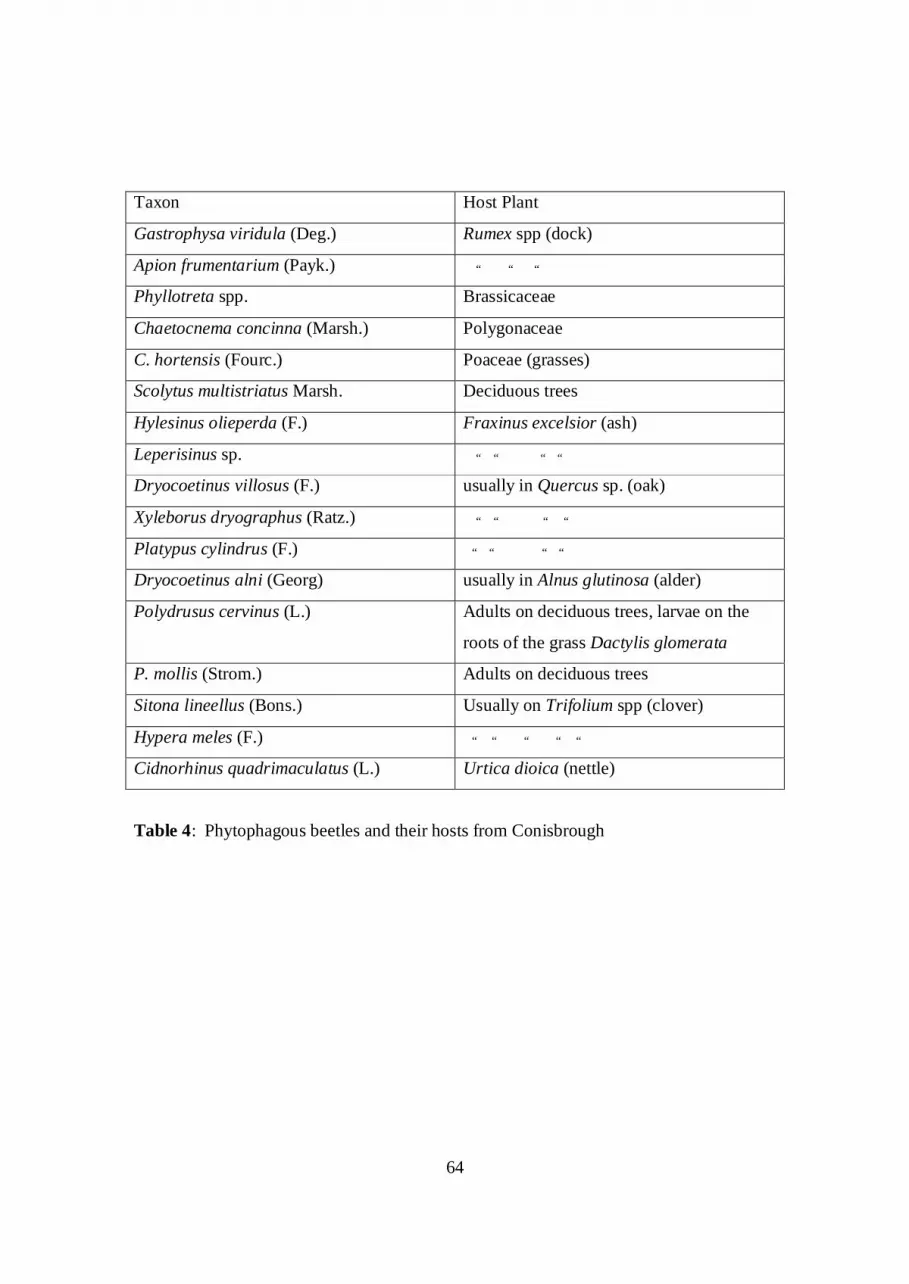
64
Taxon Host Plant
Gastrophysa viridula (Deg.) Rumex spp (dock)
Apion frumentarium (Payk.) “ “ “
Phyllotreta spp. Brassicaceae
Chaetocnema concinna (Marsh.) Polygonaceae
C. hortensis (Fourc.) Poaceae (grasses)
Scolytus multistriatus Marsh. Deciduous trees
Hylesinus olieperda (F.) Fraxinus excelsior (ash)
Leperisinus sp. “ “ “ “
Dryocoetinus villosus (F.) usually in Quercus sp. (oak)
Xyleborus dryographus (Ratz.) “ “ “ “
Platypus cylindrus (F.) “ “ “ “
Dryocoetinus alni (Georg) usually in Alnus glutinosa (alder)
Polydrusus cervinus (L.) Adults on deciduous trees, larvae on the
roots of the grass Dactylis glomerata
P. mollis (Strom.) Adults on deciduous trees
Sitona lineellus (Bons.) Usually on Trifolium spp (clover)
Hypera meles (F.) “ “ “ “ “
Cidnorhinus quadrimaculatus (L.) Urtica dioica (nettle)
Table 4: Phytophagous beetles and their hosts from Conisbrough




















