
Conflict between insect conservation and public safety: the case study of a saproxylic beetle (...
11
ORIGINAL PAPER Conflict between insect conservation and public safety: the case study of a saproxylic beetle (Osmoderma eremita) in urban parks Giuseppe Maria Carpaneto • Adriano Mazziotta • Giorgia Coletti • Luca Luiselli • Paolo Audisio Received: 21 July 2009 / Accepted: 17 February 2010 / Published online: 4 March 2010 Ó Springer Science+Business Media B.V. 2010 Abstract Urban parks can harbour small populations of saproxylic insects of high conservation concern, such as Osmoderma eremita and other rare beetles. These areas often host old trees which have become very uncommon in rural areas where they are threatened by commercial for- estry management procedures based on frequent tree cut- ting. Nevertheless, old trees of urban parks may represent a hazard for public safety and are sometimes cut by man- agement authorities. The aim of this work was to assess the loss of reproductive sites for saproxylic beetles of the Scarabaeidae, Lucanidae and Cerambycidae, when felling plans are adopted according to a Visual Tree Assessment Procedure (VTA), in a Mediterranean urban park. On July– August 2004, 1,247 holm oaks were surveyed within the border of an urban park of Rome (Villa Borghese). The occurrence of saproxylic beetles (i.e. the presence of frass, living insects or their remains) was verified in 66 old holm oaks, 41% of which were doomed to cutting by VTA. Eleven of these trees (41% of the trees doomed to be cut) held fragments of adults and sometimes living larvae of Osmoderma eremita, and four of them (36%) were inclu- ded in the felling plan. The presence of Osmoderma ere- mita in tree holes was more frequent in deep cavities. The presence of frass in the cavities was positively associated with tree height and a high degree of damage at the root collar and negatively with the presence of hole-nesting birds. Keywords Dead wood Habitat directive Saproxylic beetles Site of community importance Urban ecosystems Visual tree assessment Introduction Saproxylic beetles are fundamental links in the trophic chains of forest ecosystems (Grove 2002; Jonsson et al. 2005; Hanula et al. 2006; Johansson 2006). They supply at least two different services to temperate forest ecosystems: (1) they are an important food resource for forest birds (e.g. woodpeckers, owls, crows) and mammals (e.g. bats, rodents, carnivores) (Beckwith and Bull 1985; Radu 2007); (2) together with fungi, they contribute to fragmentation and breakdown of dead wood (Ausmus 1977; Edmonds and Eglitis 1989). The latter, however, is of great concern to managers of urban parks, because the increase in the decaying rate of trees may represent a danger to the public. For this reason, nature conservation and public safety often conflict in human settlements. Old trees in urban parks rep- resent reservoirs for saproxylic insects of high conservation priority listed on Appendix II of the European Habitat Directive (Anonymous 1992; Luce 1996). Among these Osmoderma eremita (Scopoli 1763) (Family: Scarabaeidae), is also considered Vulnerable by the IUCN Red List of Threatened Species, i.e. it is facing a high risk of extinction in the wild in the medium-term future, because its population is declining in their area of occupancy, extent of occurrence and quality of habitat (World Conservation Monitoring G. M. Carpaneto (&) A. Mazziotta Department of Biology, University ‘‘Roma Tre’’, Viale G. Marconi 446, 00146 Rome, Italy e-mail: [email protected] G. Coletti P. Audisio Department of Animal and Human Biology, ‘‘Sapienza’’ University, Rome, Italy L. Luiselli Centre of Environmental Studies Demetra S.r.l., via Olona 7, 00198 Rome, Italy 123 J Insect Conserv (2010) 14:555–565 DOI 10.1007/s10841-010-9283-5
Transcript of Conflict between insect conservation and public safety: the case study of a saproxylic beetle (...

ORIGINAL PAPER
Conflict between insect conservation and public safety: the casestudy of a saproxylic beetle (Osmoderma eremita) in urban parks
Giuseppe Maria Carpaneto • Adriano Mazziotta •
Giorgia Coletti • Luca Luiselli • Paolo Audisio
Received: 21 July 2009 / Accepted: 17 February 2010 / Published online: 4 March 2010
� Springer Science+Business Media B.V. 2010
Abstract Urban parks can harbour small populations of
saproxylic insects of high conservation concern, such as
Osmoderma eremita and other rare beetles. These areas
often host old trees which have become very uncommon in
rural areas where they are threatened by commercial for-
estry management procedures based on frequent tree cut-
ting. Nevertheless, old trees of urban parks may represent a
hazard for public safety and are sometimes cut by man-
agement authorities. The aim of this work was to assess the
loss of reproductive sites for saproxylic beetles of the
Scarabaeidae, Lucanidae and Cerambycidae, when felling
plans are adopted according to a Visual Tree Assessment
Procedure (VTA), in a Mediterranean urban park. On July–
August 2004, 1,247 holm oaks were surveyed within the
border of an urban park of Rome (Villa Borghese). The
occurrence of saproxylic beetles (i.e. the presence of frass,
living insects or their remains) was verified in 66 old holm
oaks, 41% of which were doomed to cutting by VTA.
Eleven of these trees (41% of the trees doomed to be cut)
held fragments of adults and sometimes living larvae of
Osmoderma eremita, and four of them (36%) were inclu-
ded in the felling plan. The presence of Osmoderma ere-
mita in tree holes was more frequent in deep cavities. The
presence of frass in the cavities was positively associated
with tree height and a high degree of damage at the root
collar and negatively with the presence of hole-nesting
birds.
Keywords Dead wood � Habitat directive �Saproxylic beetles � Site of community importance �Urban ecosystems � Visual tree assessment
Introduction
Saproxylic beetles are fundamental links in the trophic
chains of forest ecosystems (Grove 2002; Jonsson et al. 2005;
Hanula et al. 2006; Johansson 2006). They supply at least
two different services to temperate forest ecosystems: (1)
they are an important food resource for forest birds (e.g.
woodpeckers, owls, crows) and mammals (e.g. bats, rodents,
carnivores) (Beckwith and Bull 1985; Radu 2007); (2)
together with fungi, they contribute to fragmentation and
breakdown of dead wood (Ausmus 1977; Edmonds and
Eglitis 1989). The latter, however, is of great concern to
managers of urban parks, because the increase in the
decaying rate of trees may represent a danger to the public.
For this reason, nature conservation and public safety often
conflict in human settlements. Old trees in urban parks rep-
resent reservoirs for saproxylic insects of high conservation
priority listed on Appendix II of the European Habitat
Directive (Anonymous 1992; Luce 1996). Among these
Osmoderma eremita (Scopoli 1763) (Family: Scarabaeidae),
is also considered Vulnerable by the IUCN Red List of
Threatened Species, i.e. it is facing a high risk of extinction in
the wild in the medium-term future, because its population is
declining in their area of occupancy, extent of occurrence
and quality of habitat (World Conservation Monitoring
G. M. Carpaneto (&) � A. Mazziotta
Department of Biology, University ‘‘Roma Tre’’, Viale G.
Marconi 446, 00146 Rome, Italy
e-mail: [email protected]
G. Coletti � P. Audisio
Department of Animal and Human Biology, ‘‘Sapienza’’
University, Rome, Italy
L. Luiselli
Centre of Environmental Studies Demetra S.r.l., via Olona 7,
00198 Rome, Italy
123
J Insect Conserv (2010) 14:555–565
DOI 10.1007/s10841-010-9283-5

Centre 1996). The status of this species should be updated in
the light of the recent researches conducted in different
European countries (for a review, Ranius et al. 2005). The
larvae of these beetles develop in wood mould of decaying
trunks. In a recent review, Ranius et al. (2005) mentioned
several study cases, e.g. Austria, France, Germany, Latvia,
Lithuania, Poland, Russia, Slovakia, and Switzerland where
Osmoderma eremita occurs in urban parklands. These
authors also pointed out the potential conflict of interests
between the needs of preserving saproxylic insect and the
necessity of assuring public safety.
The status of these insects is often critical in forested
areas because of forestry management procedures consist-
ing of premature cutting and removal of trees (Ranius et al.
2005; Fontaine et al. 2007). This hampers the natural
ageing of trees and precludes the formation of natural
cavities needed by insects and other animals such as
rodents, birds and lizards (Lindenmayer et al. 2006; Ranius
et al. 2009). By contrast, old trees are often left alive in
some urban parks because they (1) have an aesthetical and
symbolic value in recreational areas; (2) provide people
with shadow and coolness; (3) are not prioritized for timber
exploitation. For this reason, urban parks can harbour
populations of saproxylic insects and have the role of small
biodiversity reservoirs for this insect community. However,
old trees in urban parks may become a public danger,
because diseased branches can fall and jeopardize life.
Therefore, cut and removal procedures are carried out in
the management of urban green areas to reduce human risk.
The occurrence of beetles protected by the Habitat Direc-
tive (not only Osmoderma eremita, but also Lucanus cervus
and Cerambyx cerdo) requires management authorities of
the urban green areas to carry out a study of Environmental
Impact Assessment, before any intervention.
We surveyed all the species belonging to Scarabaeidae,
Lucanidae and Cerambycidae (hereafter named ‘‘target
species’’) at an historical green area in the centre of Rome
(Villa Borghese). These families include species of con-
servation concern, including Osmoderma eremita. More-
over, anecdotic information gathered from Italian
entomologists suggests a marked decline of most of the
saproxylic species belonging to these three families. The
aim of this work was to assess the loss of reproductive sites
for saproxylic beetles of these families when felling plans
are adopted, according to a Visual Tree Assessment Pro-
cedure (VTA), in a Mediterranean urban park. We mea-
sured different characteristics of the trees and analysed
which of them were the best predictors of presence/absence
of the surveyed species (verified through the occurrence of
live specimens, larval frass, exoskeletal fragments) and
species richness. Finally, suggestions are given for pre-
serving both hollow trees and public safety, without
affecting insect diversity.
Materials and methods
Study area
This research was conducted in Villa Borghese, one of the
green areas of Rome (159 ha including also the Pincio
Garden, mean altitude: 60 m a.s.l.), located in the centre of
the city (coordinates for the core area of the study site:
projection WGS84 UTM Zone 33 N; 41�5404500N,
12�2901200E). The habitat is a continuous parkland, com-
prising an open woodland formed by scattered trees and
several clearings. Dominant trees are either native or
introduced species of oaks and other hardwood trees,
interspersed with conifers (cedars, pines, cypresses). The
most common landmark trees are old holm oaks (Quercus
ilex), an evergreen component that is the dominant tree
species of Mediterranean sclerophyllous scrub forest.
Villa Borghese was included in the list of the proposed
Sites of Community Importance (pSCIs) (IT6030052) of
the European Commission, together with another urban
park of Rome (Villa Pamphili), because of the presence of
Osmoderma eremita (Ranius et al. 2005). The management
authorities of Villa Borghese (Servizio Giardini di Roma)
divided the park into eight sectors, and four of these sectors
were subject to a Visual Tree Assessment procedure (see
below for details), in order to assess their status and the risk
for public safety due to falling trunks or branches. We
inspected all the 1,247 holm oaks included in those sectors,
focusing on those potentially harbouring saproxylic beetles
associated with tree hollows.
The visual tree assessment procedure (VTA)
The visual tree assessment (VTA) procedure was devel-
oped by Mattheck and Breloer (1994) to assess the risk for
public safety by falling trees. It was applied in urban areas
by Matheny and Clark (1993) and was used in the study
area to minimise the risk of tree failure by planning
interventions. These precautionary measures ranged from
cutting the decaying branches of a given tree to the
removal of the whole tree. A Falling Risk Category (FRC)
was assigned to each tree, ranging from A (healthy tree) to
D (seriously damaged), as an increasing scale of risk cat-
egory. Both C and D categories were divided into seven
subcategories (C1-3, CD, D1-3). These categories and
subcategories were assigned by evaluating the status of the
collar, trunk, and canopy. This information was obtained
using a resistograph, an instrument measuring the resis-
tance to a needle inserted into the wood under constant
drive, and a fractometer, an instrument measuring the
strength loss of the tree due to decay. This method is an
alternative to the method developed by Pacyniak (1992) to
assess tree health, and also used by Oleksa et al. (2006,
556 J Insect Conserv (2010) 14:555–565
123

2007) to verify the presence of the target saproxylic beetles
by means of tree features. This was based on a five steps
scale: (1) trunk and crown healthy; (2) hollows present, up
to 25% of crown damaged (loss); (3) 25–50% damaged
(loss); (4) 50–75% damaged (loss); (5) above 75% dam-
aged (loss) or a dead tree).
Detection of adult beetles
The search for adult saproxylic beetles of the families
Scarabaeidae, Lucanidae and Cerambycidae (hereafter
quoted as ‘‘target beetles’’) was conducted during the after-
noons (from 2 to 7 p.m.), from 19 July 2004 to 28 August
2004. Each sector (see ‘‘Study area’’) was visited once every
4 days, for five periods throughout the duration of the study.
A torch, ladder and climbing equipment were used to explore
each hollow located up to 5 m above the ground. Target
beetles were initially searched for over all the trees, subse-
quently searching was confined to trees belonging to C and D
categories, i.e. to the trees showing hollows or a decaying
status. These were the trees both potentially harbouring
saproxylic beetles and affected by a higher falling risk,
included in ‘‘cut and removal’’ procedures.
The following procedure was applied each day:
• accurate visual checking of all the trees, to verify the
presence of hollows suitable for the development of
Scarabaeidae and Lucanidae, and the occurrence
of adult target beetles on the bark and on the surface
of wood mould in the cavities;
• olfactory searching of Osmoderma eremita following
the strong scent track left by male pheromones (Larsson
et al. 2003; Svensson et al. 2003); no other saproxylic
beetle have this scent which can be smelt by humans at
a distance of some meters;
• searching of specimens and/or fragments of adult body
parts, examining the inner surface of the cavity and
gently extracting by hand the wood mould from the
hole, then scattering it over a white oilcloth to detect
body fragments and frass (larval excrement of lamelli-
corn beetles), and finally replacing the wood mould into
the hollow;
• placing of 10 empty pitfall traps in each sector and
checking them every 3 days, in order to capture adult
beetles. Pitfall traps were buried in the wood mould to
the brim, inside the tree hollows.
Detection of larvae and frass
The search for larvae of saproxylic scarab beetles and their
excrement (frass), together with the description of tree
features, was conducted from 1 to 30 September 2004
(from 9 a.m. to 6 p.m.). Each sector was visited 3 times.
The wood mould inside hollows was accurately examined
with the same procedure adopted for the search of adult
body fragments, i.e. scattering it over a white oilcloth, in
order to detect larvae and frass of saproxylic scarab beetles
(Stegner 2002), and immediately relocated in the same tree.
Larvae of darkling beetles (Tenebrionidae) and click bee-
tles (Elateridae) were found but not identified.
Regarding the environmental variables that may affect
the occurrence of the target beetles in tree hollows, we
chose the physical characteristics of individual hollow trees
that previous authors considered to be associated with
microclimate, nutrient supply and successional stages of
the trees in northern Europe (Ranius and Nilsson 1997), in
order to test the same correlations in Mediterranean habi-
tats (Table 1). We added the distance between a hollow
tree and the nearest hollow tree as a surrogate of connec-
tivity. The continuous tree cover did not permit identifi-
cation of individual stand level features. We also added a
variable adapted to the specific objective of this work, the
Falling Risk Category (FRC).
Statistical analyses
For the analyses, the tree features (Table 1) were consid-
ered as predictor variables. The response variables were:
(a) presence/absence of excrement of saproxylic scarab
beetles in tree hollows (frass); (b) presence/absence of
fragments of Osmoderma eremita in tree hollows (frag-
ments); (c) the species richness of target beetles (species).
Some continuous variables were transformed into ordinal
variables by assigning categorical scores for each of them
(see Table 1 for the list of all covariates and their codes). In
order to satisfy homogeneity of variances, all data were
log-transformed, and in this case parametric statistics were
used. When data were not normalizable, nonparametric
tests were used. Spearman rank correlation coefficient (for
non-normal variables) were used to explore pairwise
association among variables, with alpha set at 5%.
Sixty-one cavities (a single cavity randomly selected for
each tree) were analysed twice by a logistic regression (by
alternatively using the Forward and Backward entry Step-
wise design models) to predict the way in which the asso-
ciation of environmental predictor variables may influence
significantly the presence/absence of both larval frass (frass)
and adult fragments (fragments) of Osmoderma eremita in
hollow trees (Hosmer and Lemeshow 2000). For this anal-
ysis, categorical covariates were treated with the Polynomial
contrast method because their values were equally spaced.
For these analyses, we considered only those variables that
were not significantly associated with each other in pairwise
comparison (i.e. bird_nest, fungus, aspect, canopy_cover,
wood_mould, root_status, trunk_status, dist_hollow_tree,
height, DBH, width, dist_hollow_surface).
J Insect Conserv (2010) 14:555–565 557
123

In order to analyse the influence of the joint contribution
of the other covariates in predicting species richness of the
target beetles (species), we also performed independent
regressions, and then assessed their significance using the
Bonferroni correction for multiple simultaneous tests.
As for the logistic regression, the way in which inde-
pendent predictors were introduced into the analysis was
chosen according to the lowest alpha value of the full
models. All statistical elaborations were carried out using
two software packages: SPSS 8.0 (SPSS Inc.) and STAT-
ISTICA 7.0 (Statsoft).
Results
Saproxylic beetles and hazardous trees
Ten species of target beetles (Cerambycidae, Lucanidae
and Scarabaeidae) were found (Table 2), including
Cerambyx cerdo and Osmoderma eremita which are
included in Appendix 2 of the Habitat Directive.
Among the 1,247 trees occurring in the four sectors
examined, 76 trees belonged to the D3 FRC category (i.e.
6.1% of the total), representing a hazard for humans and
therefore requiring a urgent cut and removal procedure.
Sixty-six trees (5.3% of the total) belonging to different
Falling Risk Categories had signs of target beetles (i.e.
fragments, frass, living adult insects or larvae). Of these
trees, 27 (41%) belonged to D3 category and contained
fragments, living adult insects or larvae; only 39 trees
contained frass (51.3% D3, 23.1% C–D, 10.3% C3, 10.3%
D1, 5.1% D2) (Fig. 1C). Only the occurrence of larvae
and/or frass gave us the evidence that a tree was a breeding
site for at least one species. Larvae of the tribe Cetoniinae
(Osmoderma, Cetonia, Potosia, Eupotosia, Netocia) were
only found in five trees. Larvae of Osmoderma eremita
were found only in two trees and some remains of dead
adults of Osmoderma eremita were found in eleven trees.
Only one live adult was found. The eleven trees where we
found Osmoderma eremita (larvae or adults and their
remains) belonged to different categories: four trees
belonged to D3, one tree to D1 and six trees to C–D
(Fig. 1).
Features of the hollow trees
All the 66 hollow trees examined (i.e. the trees which
showed at least one cavity and signs of presence of target
beetles) were alive (living) (Fig. 2). The median value of
the distances among two hollow trees (dist_hollow_tree)
Table 1 Code, classes, typology and description of the variables used in the regression analysis
Code Class Type Description of variables
FRC CT TC Falling Risk Category (C3 = 1; C-D = 2; D1 = 3; D2 = 4, D3 = 5)
Living CT TC Living tree (No = 0, Yes = 1)
Cracks CT TC Cracks (absent = 0; present = 1)
Dist_hollow_tree CN TC Distance from the nearest hollow tree (m)
Height CN TC Tree height (m)
DBH CN TC Trunk diameter at breast height (DBH), i.e. 1.30 m above ground (cm)
Dist_ground_hollow CN TC Distance between the ground and the hollow entrance (m)
Width CN TC Maximum width of the hollow (cm)
Dist_hollow_surface CN TC Vertical distance between the entrance hollow and the surface of the wood mould (cm)
Bird_nest CT TC Bird nest remnants (absence = 0; presence = 1)
Fungus CT TC Fungus infestation (less than 10% of the interior cavity surface = 0; more than 10% = 1)
Direction CT TC Direction of the entrance hole respect to the ground (horizontal = 0; angulated or vertical = 1)
Aspect CT TC Aspect of entrance hole (NE = 1; N o E = 2; NW o SE = 3, W o S = 4; SW = 5)
Canopy_cover CT TC Canopy cover, i.e. % of the sky covered by canopy over the hollow (coverage B 50% = 0;
coverage [ 50% = 1)
Wood_mould CT TC Would mould quantity in the cavity (volume B 2 litres = 0; volume [ 2 litres = 1)
Frass CT SD Excrement of lamellicorn beetles (absence = 0; presence = 1)
Fragments CT SD Fragments of Osmoderma eremita (absence = 0, presence = 1)
Species CN SD Number of saproxylic target species in the cavity
Root_status CT TC Status of the root collar (good = 1, discrete = 2, sufficient = 3, mediocre = 4, scarce = 5)
Trunk_status CT TC Status of the trunk (good = 1, discrete = 2, sufficient = 3, mediocre = 4, scarce = 5)
Canopy_status CT TC Status of the canopy (good = 1, discrete = 2, sufficient = 3, mediocre = 4, scarce = 5)
Class: categorical (CT) or continuous (CN). Type: species data = SD, tree characteristics = TC
558 J Insect Conserv (2010) 14:555–565
123
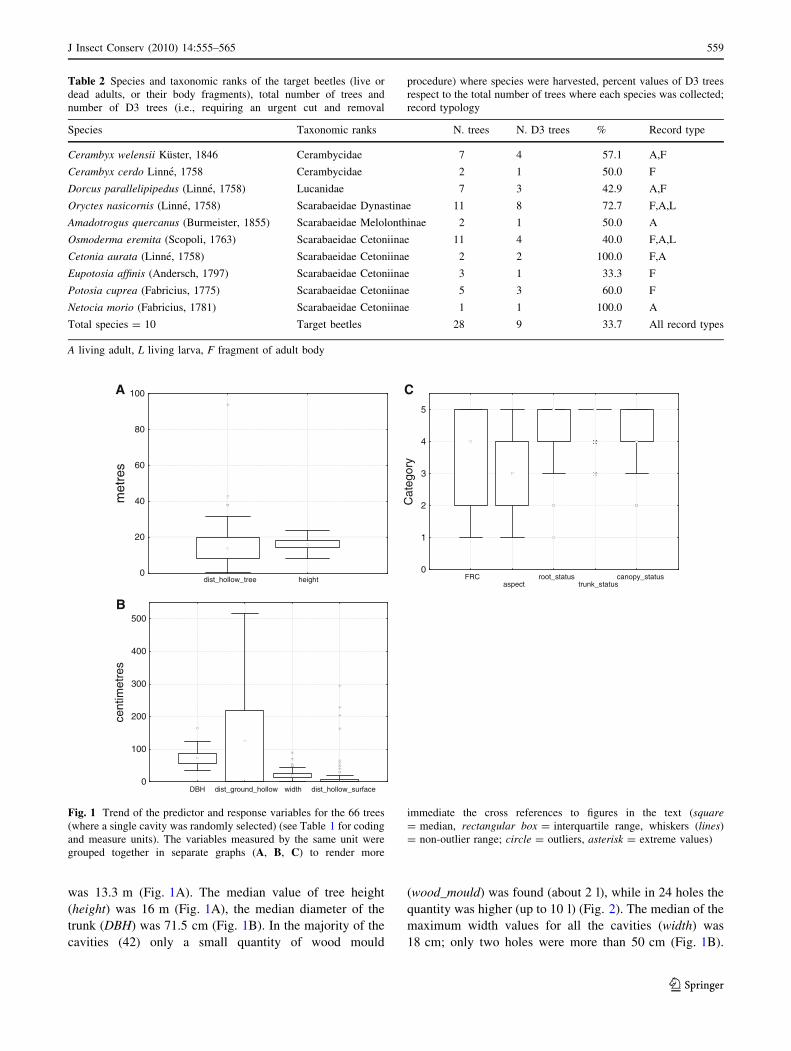
was 13.3 m (Fig. 1A). The median value of tree height
(height) was 16 m (Fig. 1A), the median diameter of the
trunk (DBH) was 71.5 cm (Fig. 1B). In the majority of the
cavities (42) only a small quantity of wood mould
(wood_mould) was found (about 2 l), while in 24 holes the
quantity was higher (up to 10 l) (Fig. 2). The median of the
maximum width values for all the cavities (width) was
18 cm; only two holes were more than 50 cm (Fig. 1B).
Table 2 Species and taxonomic ranks of the target beetles (live or
dead adults, or their body fragments), total number of trees and
number of D3 trees (i.e., requiring an urgent cut and removal
procedure) where species were harvested, percent values of D3 trees
respect to the total number of trees where each species was collected;
record typology
Species Taxonomic ranks N. trees N. D3 trees % Record type
Cerambyx welensii Kuster, 1846 Cerambycidae 7 4 57.1 A,F
Cerambyx cerdo Linne, 1758 Cerambycidae 2 1 50.0 F
Dorcus parallelipipedus (Linne, 1758) Lucanidae 7 3 42.9 A,F
Oryctes nasicornis (Linne, 1758) Scarabaeidae Dynastinae 11 8 72.7 F,A,L
Amadotrogus quercanus (Burmeister, 1855) Scarabaeidae Melolonthinae 2 1 50.0 A
Osmoderma eremita (Scopoli, 1763) Scarabaeidae Cetoniinae 11 4 40.0 F,A,L
Cetonia aurata (Linne, 1758) Scarabaeidae Cetoniinae 2 2 100.0 F,A
Eupotosia affinis (Andersch, 1797) Scarabaeidae Cetoniinae 3 1 33.3 F
Potosia cuprea (Fabricius, 1775) Scarabaeidae Cetoniinae 5 3 60.0 F
Netocia morio (Fabricius, 1781) Scarabaeidae Cetoniinae 1 1 100.0 A
Total species = 10 Target beetles 28 9 33.7 All record types
A living adult, L living larva, F fragment of adult body
dist_hollow_tree height0
20
40
60
80
100
met
res
A
DBH dist_ground_hollow width dist_hollow_surface0
100
200
300
400
500
cent
imet
res
B
FRCaspect
root_statustrunk_status
canopy_status0
1
2
3
4
5C
ateg
ory
C
Fig. 1 Trend of the predictor and response variables for the 66 trees
(where a single cavity was randomly selected) (see Table 1 for coding
and measure units). The variables measured by the same unit were
grouped together in separate graphs (A, B, C) to render more
immediate the cross references to figures in the text (square= median, rectangular box = interquartile range, whiskers (lines)
= non-outlier range; circle = outliers, asterisk = extreme values)
J Insect Conserv (2010) 14:555–565 559
123

A large number of holes had a northward (or NE) or
eastward (SE) sun exposure (aspect) (Fig. 1C).
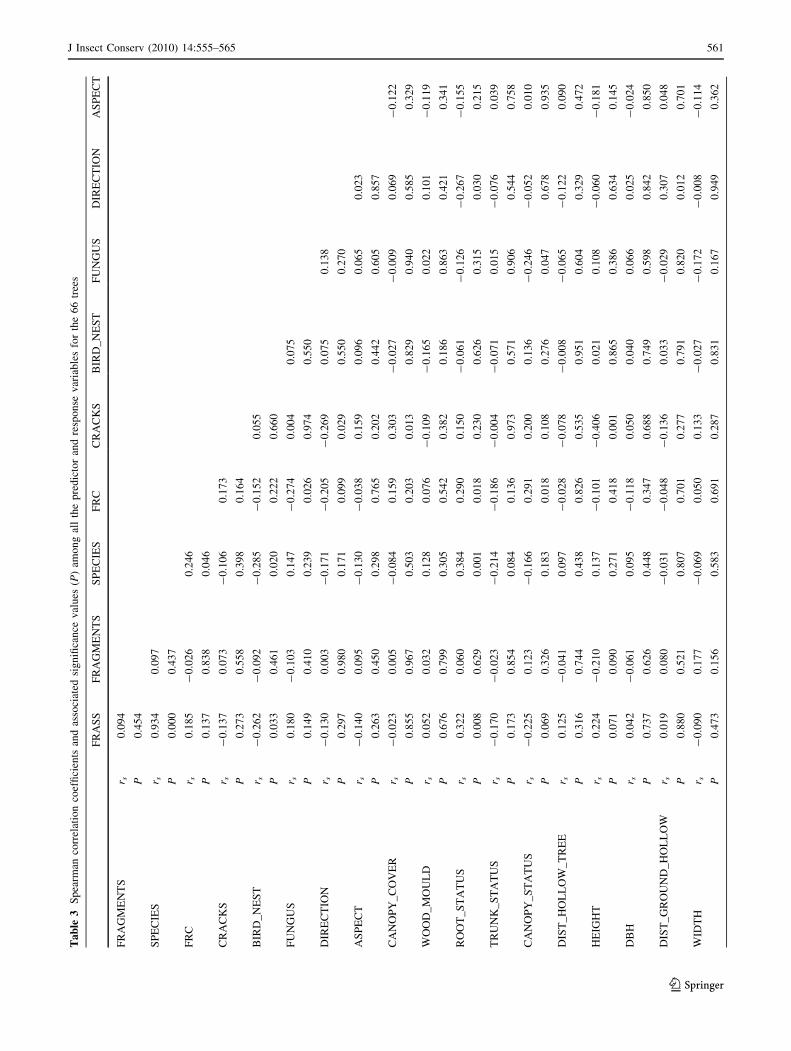
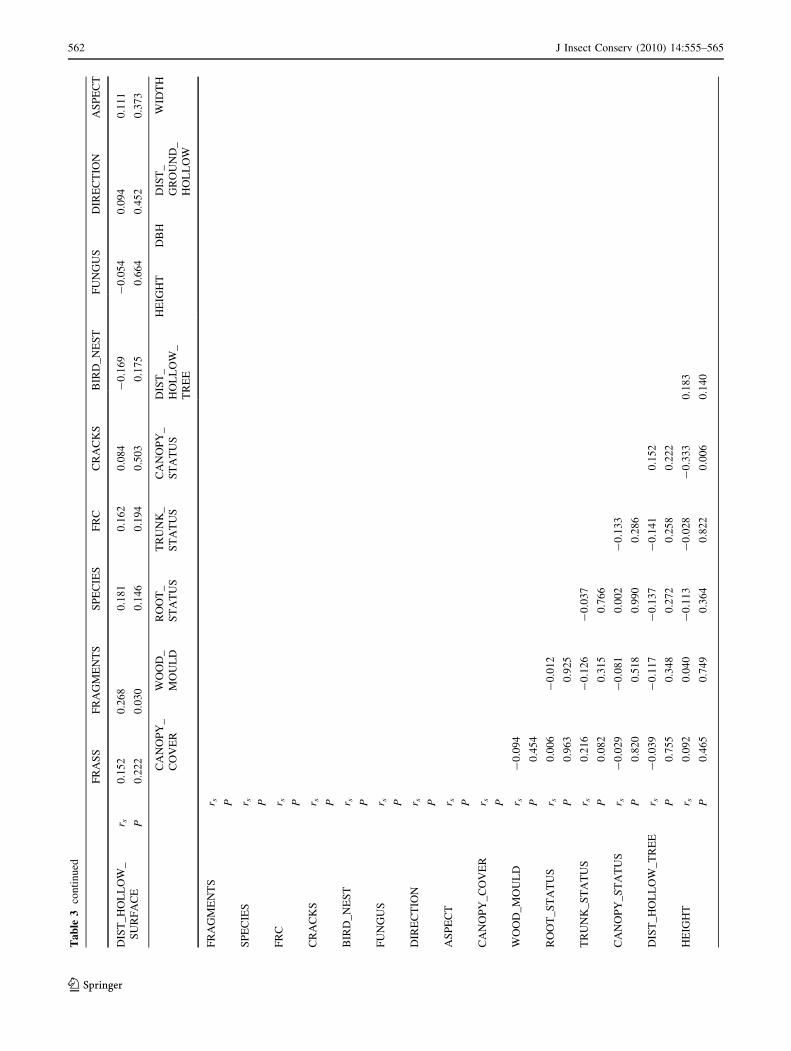
The Spearman correlation coefficients were calculated
for all the predictor variables of 66 full-data cavities
occurring in the 66 trees selected from all the sectors
(Table 3). The following significant results (at least
P \ 0.05) were obtained:
(1) the hazard degree (FRC categories score) increased
with the degradation of the root collar (root) and with
the degradation of the status of the canopy
(canopy_status);
(2) cracks in the trees (cracks) took place preferentially
in short trees (height), with a good canopy cover
(canopy_cover);
(3) tall trees (height) frequently had a poor status of the
canopy (canopy_status);
(4) the status of the canopy (canopy_status) got worse in
trees with fungus infestations in the cavity (fungus);
(5) the quantity of wood mould (wood_mould) existing in
the trunks was higher in trees with larger holes
(width).
Environmental features and beetle occurrence
The presence of fragments of Osmoderma eremita (frag-
ments) was associated with hollows characterized by a
higher vertical distance between the surface of wood mould
and the entrance hole (dist_hollow_surface) (see Table 3).
Excrement (frass) was found in the majority of the trees
where bird nests or feathers were not found in their cavities
(bird_nest) or the root collar was damaged (root_status)
(see Table 3). The higher richness of target species
(species) was found in trees with a high Falling Risk
Category (FRC) whose cavities did not contain bird nests
or feathers (bird_nest), the root collar was damaged
(root_status), and the presence of frass was verified (frass)
(see Table 3). The presence of frass (frass) and the degree
of damage for root collar (root_status) were also jointly
related with the richness of target species, accounting for
the Bonferroni correction (P \ 0.0167).
Logistic regression
The logistic regressions performed for the 66 trees pointed
out a positive significant predictive power of the tree height
(height) and of the fungus infestation (fungus) on the pres-
ence of frass in the cavities (frass) (best model selection
method: Backward; v2 = 37.6; df = 11; P \ 0.001; logit
link function: log (frass ? 1) = (9.144 ± 3.975) 9 log
(height ? 1) ? (2.229 ± 0.905) 9 log (fungus ? 1). The
observed presence of frass in the cavities matched the
expected presence in 82.5% (33/40) of the cavities, while
the observed absence corresponded to the expected absence
in 76.9% (20/26) of the cavities. On the whole, 80.3% of the
observed cavities matched the expected values.
Osmoderma eremita remains (fragments) were predicted
to occur more frequently in cavities with a high dis-
tance between the entrance hollow and the surface of
wood mould (dist_hollow_surface) (best model selection
method: Forward; v2 = 8.127; df = 1; P = 0.004; logit
link function: log (fragments ? 1) = (-2.578 ± 0.542) ?
(1.320 ± 0.481) 9 log (dist_hollow_surface ? 1). The
observed presence of fragments in the cavities matched the
expected presence in 100.0% (3/3) of the cavities, while
the observed absence corresponded to the expected absence
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
living
crac
ks
bird_
nest
fung
us
direc
tion
cano
py_c
over
wood_
mou
ldfra
ss
fragm
ents
1 0
Fig. 2 Percent values of two-states categorical variables for the 66 trees (see Table 1 for the code)
560 J Insect Conserv (2010) 14:555–565
123
Ta
ble
3S
pea
rman
corr
elat
ion
coef
fici
ents
and
asso
ciat
edsi
gn
ifica
nce
val
ues
(P)
amo
ng
all
the
pre
dic
tor
and
resp
on
sev
aria
ble
sfo
rth
e6
6tr
ees
FR
AS
SF
RA
GM
EN
TS
SP
EC
IES
FR
CC
RA
CK
SB
IRD
_N
ES
TF
UN
GU
SD
IRE
CT
ION
AS
PE
CT
FR
AG
ME
NT
Sr s
0.0
94
P0
.45
4
SP
EC
IES
r s0
.93
40
.09
7
P0
.00
00
.43
7
FR
Cr s
0.1
85
-0
.02
60
.24
6
P0
.13
70
.83
80
.04
6
CR
AC
KS
r s-
0.1
37
0.0
73
-0
.10
60
.17
3
P0
.27
30
.55
80
.39
80
.16
4
BIR
D_
NE
ST
r s-
0.2
62
-0
.09
2-
0.2
85
-0
.15
20
.05
5
P0
.03
30
.46
10
.02
00
.22
20
.66
0
FU
NG
US
r s0
.18
0-
0.1
03
0.1
47
-0
.27
40
.00
40
.07
5
P0
.14
90
.41
00
.23
90
.02
60
.97
40
.55
0
DIR
EC
TIO
Nr s
-0
.13
00
.00
3-
0.1
71
-0
.20
5-
0.2
69
0.0
75
0.1
38
P0
.29
70
.98
00
.17
10
.09
90
.02
90
.55
00
.27
0
AS
PE
CT
r s-
0.1
40
0.0
95
-0
.13
0-
0.0
38
0.1
59
0.0
96
0.0
65
0.0
23
P0
.26
30
.45
00
.29
80
.76
50
.20
20
.44
20
.60
50
.85
7
CA
NO
PY
_C
OV
ER
r s-
0.0
23
0.0
05
-0
.08
40
.15
90
.30
3-
0.0
27
-0
.00
90
.06
9-
0.1
22
P0
.85
50
.96
70
.50
30
.20
30
.01
30
.82
90
.94
00
.58
50
.32
9
WO
OD
_M
OU
LD
r s0
.05
20
.03
20
.12
80
.07
6-
0.1
09
-0
.16
50
.02
20
.10
1-
0.1
19
P0
.67
60
.79
90
.30
50
.54
20
.38
20
.18
60
.86
30
.42
10
.34
1
RO
OT
_S
TA
TU
Sr s
0.3
22
0.0
60
0.3
84
0.2
90
0.1
50
-0
.06
1-
0.1
26
-0
.26
7-
0.1
55
P0
.00
80
.62
90
.00
10
.01
80
.23
00
.62
60
.31
50
.03
00
.21
5
TR
UN
K_
ST
AT
US
r s-
0.1
70
-0
.02
3-
0.2
14
-0
.18
6-
0.0
04
-0
.07
10
.01
5-
0.0
76
0.0
39
P0
.17
30
.85
40
.08
40
.13
60
.97
30
.57
10
.90
60
.54
40
.75
8
CA
NO
PY
_S
TA
TU
Sr s
-0
.22
50
.12
3-
0.1
66
0.2
91
0.2
00
0.1
36
-0
.24
6-
0.0
52
0.0
10
P0
.06
90
.32
60
.18
30
.01
80
.10
80
.27
60
.04
70
.67
80
.93
5
DIS
T_
HO
LL
OW
_T
RE
Er s
0.1
25
-0
.04
10
.09
7-
0.0
28
-0
.07
8-
0.0
08
-0
.06
5-
0.1
22
0.0
90
P0
.31
60
.74
40
.43
80
.82
60
.53
50
.95
10
.60
40
.32
90
.47
2
HE
IGH
Tr s
0.2
24
-0
.21
00
.13
7-
0.1
01
-0
.40
60
.02
10
.10
8-
0.0
60
-0
.18
1
P0
.07
10
.09
00
.27
10
.41
80
.00
10
.86
50
.38
60
.63
40
.14
5
DB
Hr s
0.0
42
-0
.06
10
.09
5-
0.1
18
0.0
50
0.0
40
0.0
66
0.0
25
-0
.02
4
P0
.73
70
.62
60
.44
80
.34
70
.68
80
.74
90
.59
80
.84
20
.85
0
DIS
T_
GR
OU
ND
_H
OL
LO
Wr s
0.0
19
0.0
80
-0
.03
1-
0.0
48
-0
.13
60
.03
3-
0.0
29
0.3
07
0.0
48
P0
.88
00
.52
10
.80
70
.70
10
.27
70
.79
10
.82
00
.01
20
.70
1
WID
TH
r s-
0.0
90
0.1
77
-0
.06
90
.05
00
.13
3-
0.0
27
-0
.17
2-
0.0
08
-0
.11
4
P0
.47
30
.15
60
.58
30
.69
10
.28
70
.83
10
.16
70
.94
90
.36
2
J Insect Conserv (2010) 14:555–565 561
123
Ta
ble
3co
nti
nu
ed
FR
AS
SF
RA
GM
EN
TS
SP
EC
IES
FR
CC
RA
CK
SB
IRD
_N
ES
TF
UN
GU
SD
IRE
CT
ION
AS
PE
CT
DIS
T_
HO
LL
OW
_
SU
RF
AC
E
r s0
.15
20
.26
80
.18
10
.16
20
.08
4-
0.1
69
-0
.05
40
.09
40
.11
1
P0
.22
20
.03
00
.14
60
.19
40
.50
30
.17
50
.66
40
.45
20
.37
3
CA
NO
PY
_
CO
VE
R
WO
OD
_
MO
UL
D
RO
OT
_
ST
AT
US
TR
UN
K_
ST
AT
US
CA
NO
PY
_
ST
AT
US
DIS
T_
HO
LL
OW
_
TR
EE
HE
IGH
TD
BH
DIS
T_
GR
OU
ND
_
HO
LL
OW
WID
TH
FR
AG
ME
NT
Sr s P
SP
EC
IES
r s P
FR
Cr s P
CR
AC
KS
r s P
BIR
D_
NE
ST
r s P
FU
NG
US
r s P
DIR
EC
TIO
Nr s P
AS
PE
CT
r s P
CA
NO
PY
_C
OV
ER
r s P
WO
OD
_M
OU
LD
r s-
0.0
94
P0
.45
4
RO
OT
_S
TA
TU
Sr s
0.0
06
-0
.01
2
P0
.96
30
.92
5
TR
UN
K_
ST
AT
US
r s0
.21
6-
0.1
26
-0
.03
7
P0
.08
20
.31
50
.76
6
CA
NO
PY
_S
TA
TU
Sr s
-0
.02
9-
0.0
81
0.0
02
-0
.13
3
P0
.82
00
.51
80
.99
00
.28
6
DIS
T_
HO
LL
OW
_T
RE
Er s
-0
.03
9-
0.1
17
-0
.13
7-
0.1
41
0.1
52
P0
.75
50
.34
80
.27
20
.25
80
.22
2
HE
IGH
Tr s
0.0
92
0.0
40
-0
.11
3-
0.0
28
-0
.33
30
.18
3
P0
.46
50
.74
90
.36
40
.82
20
.00
60
.14
0
562 J Insect Conserv (2010) 14:555–565
123

in 88.9% (56/63) of the cavities. On the whole, 89.4% of
the observed cavities matched the expected values.
Discussion
Saproxylic diversity in the urban park
Eight species of saproxylic scarab beetles (Scarabaeidae
and Lucanidae) were found in Villa Borghese (urban area,
159 ha) (Table 2). This richness corresponded to 47% of
the number of species (17) observed in the Presidential
Estate of Castelporziano, the nearest forest, at about 25 km
of distance, 5,892 ha), during a previous intensive research
for a faunistic inventory (Carpaneto et al., unpublished
data). Moreover, Villa Borghese is one of just nine local-
ities of Osmoderma eremita known for Latium (Ruffo and
Stoch 2004), the administrative region of Rome. Latium
has an area of approximately 17,227 km2, of which 13.6%
is included in protected areas (reserves and parks). In this
region, woodlands cover 35% of the whole surface (IFNC
2005, and official data from: www.regione.lazio.it), but old
living hollow trees are poorly represented across the
landscape. Therefore, urban parklands like Villa Borghese,
whose purpose is not wood production, harbour a consid-
erable number of veteran trees and may therefore have a
role of source areas for saproxylic beetles.
In the study area, 41% of the trees inhabited by target
beetles were labelled as dangerous for citizens, represent-
ing a clash of interests between biodiversity conservation
and public safety. Moreover, a considerable amount of the
trees inhabited by Osmoderma eremita were included in a
felling project.
Suitability of hollow trees for saproxylic beetles
The field detection of Osmoderma eremita in central and
southern Europe is very difficult because of the secluded
life of the species inside hollow trees, its limited dispersal
activity, and its low population density (Bubler and Muller
2009; Carpaneto et al., unpublished data). Therefore, the
definition and use of predictor variables could be an
important tool to make provision for the occurrence of the
species and for monitoring protected areas.
The analysis of predictor variables confirms the impor-
tance for target beetles of hollow trees which are still alive, as
their cavities represent a suitable microhabitat and probably
will maintain this role until the death of the trees (cf. Ranius
and Nilsson 1997; Schaffrath 2003a, b). The analysis of the
66 trees with different signs of presence of target beetles also
indicates that good connectivity (i.e. short distance among
hollow trees) favours the target beetles. For example Os-
moderma eremita, has a dispersal range varying from 30 toTa
ble
3co
nti
nu
ed
CA
NO
PY
_
CO
VE
R
WO
OD
_
MO
UL
D
RO
OT
_
ST
AT
US
TR
UN
K_
ST
AT
US
CA
NO
PY
_
ST
AT
US
DIS
T_
HO
LL
OW
_
TR
EE
HE
IGH
TD
BH
DIS
T_
GR
OU
ND
_
HO
LL
OW
WID
TH
DB
Hr s
-0
.17
00
.23
40
.05
9-
0.0
90
0.1
75
0.0
62
0.1
51
P0
.17
20
.05
90
.64
10
.47
20
.16
00
.62
00
.22
5
DIS
T_
GR
OU
ND
_
HO
LL
OW
r s-
0.0
56
-0
.12
5-
0.2
90
-0
.15
70
.18
20
.05
4-
0.0
99
0.1
82
P0
.65
70
.31
70
.01
80
.20
90
.14
30
.66
70
.42
80
.14
4
WID
TH
r s0
.09
90
.41
70
.02
7-
0.0
43
0.1
85
-0
.19
2-
0.0
98
0.1
14
-0
.12
2
P0
.42
70
.00
00
.82
70
.72
90
.13
80
.12
30
.43
20
.36
30
.32
9
DIS
T_
HO
LL
OW
_
SU
RF
AC
E
r s-
0.0
07
-0
.05
1-
0.0
01
0.0
47
0.2
36
-0
.05
7-
0.1
99
0.0
89
0.4
08
-0
.12
9
P0
.95
30
.68
60
.99
30
.70
50
.05
60
.64
80
.10
80
.47
80
.00
10
.30
3
Th
ev
aria
ble
‘‘li
vin
g’’
was
del
eted
bec
ause
itw
asco
nst
ant.
See
the
leg
end
of
abb
rev
iati
on
sin
Tab
le1
J Insect Conserv (2010) 14:555–565 563
123

200 m in Northern Europe (for Sweden: Ranius and Hedin
2001; Ranius 2006; Hedin et al. 2008) and reaches up to
700 m in Southern Europe (for France: Dubois and Vignon
2008). Almost all the trees which were hosts of the target
beetles were big trees; height and diameter had a positive
influence on the quantity of wood mould found in each
hollow. Most of the tree cavities with target beetles had a
small entrance hole, and were northward or eastward sun-
exposed, all factors that maintain a stable and humid
microclimate (Ranius and Nilsson 1997).
The pairwise comparison of predictor variables showed
that Falling Risk Categories for hollow trees were really
associated with different indications of decay and these
signs were related to each other. Substantially, the state of
the variables determining the frequency of occurrence of
the target beetles in Southern Europe were practically the
same detected by other authors in Northern Europe (Ranius
and Nilsson 1997).
According to the results of pairwise comparison and
logistic regression, the frass of saproxylic scarab beetles
and the species richness had a positive response to old trees
with signs of ageing (in particular a damaged root collar)
and without bird nesting activity, in agreement with
the results obtained by other authors in Northern Europe
(Ranius and Nilsson 1997). Osmoderma eremita fragments
were preferentially found in deep cavities. However, a
more extensive analysis of the relations between the
occurrence of Osmoderma eremita and environmental
variables should be carried out in different ecogeographical
contexts to assess the ecological requirements of this spe-
cies throughout its range. The importance of high trees in
preserving the target beetles was related with the high
probability that a tall tree harbours more hollows and
sheltered cavities in its high trunks.
Conclusions and management proposals
An urban park entrapped inside a big city can be a reservoir
for threatened saproxylic beetles and can host a diverse
community of these insects, owing to the occurrence of
monumental trees. On the contrary, out of cities, aged
veteran trees represent a rare and scattered resource either
in managed woodlands or in the forest remnants of a
fragmented landscape. Thus, the deadwood resource of
urban parks may be more suitable to host species of the
genus Osmoderma, than those of the countryside. Never-
theless, these species are at risk of removal from these
urban refuges because of the clash of interests between
biodiversity conservation and public safety. In the study
area, the presence of Osmoderma eremita together with a
guild of saproxylic beetles was confirmed in old trees and
predicted by some variables associated with a category of
high Falling Risk, as designated by the Visual Tree
Assessment Procedure (VTA).
The following reference list summarizes the main
requirements for the life cycle of saproxylic scarab beetles,
especially for Osmoderma eremita: (1) living trees with
abundant wood mould in the hollows; (2) cavities adequate
to maintain high humidity and cool temperatures in sum-
mer; (3) old hollow trees located at short distance each
other (maximum 100 m), to favour adult dispersion and
encounter between them; (4) cavities not affected by bird
nesting activity. The latter requirement may be explained
by the current increasing population of the starling (Sturnus
vulgaris), an invasive hole-nesting bird which usually feeds
on large insects, and could represent a direct threat for
Osmoderma eremita and other saproxylic scarab beetles.
The designation of Falling Risk Categories associated
with trees allows wildlife managers to identify conserva-
tion hotspots for threatened saproxylic beetles. The fol-
lowing recommendations are given to promote the
conservation of saproxylic insects in urban green areas, in
agreement with actions for public safety: (1) reducing cuts
of secondary tree branches to conserve a canopy cover and
optimal microclimate conditions within the holes; (2)
leaving the largest branches and trunks on the ground after
cutting, in order to increase development sites for sapr-
oxylic larvae; (3) using steel ropes and other shores to hold
collapsing trees; (4) applying Natural Fracture Techniques
such as pruning methods used to mimic fractured ends
which naturally occur on trunks and branches; (5) remov-
ing inadequate reinforcement materials used in the past to
strengthen old trees or to occlude hollows; (6) removing
trash and other objects left in hollows by people, in order to
recover habitat conditions.
Acknowledgments We are grateful to Bruno Cignini, executive
manager of the Rome Municipality (10th Department: Environment
and Agriculture Policies/Conservation and management of vast areas
and biodiversity) for providing facilities and funding. A special thank
is due for Fabio Nicolai (Zoological Museum, Rome) for field
assistance and help.
References
Anonymous (1992) Directive 92/43 of the Council of the European
Community on the conservation of habitats and wild fauna and
flora. Brussels, European Community
Ausmus BS (1977) Regulation of wood decomposition rates by
arthropod and annelid populations. Ecol Bull 25:180–192
Beckwith RC, Bull EL (1985) Scat analysis of the arthropod
component of pileated woodpecker diet. The Murrelet 66:90–92
Bubler H, Muller J (2009) Vacuum cleaning for conservationists: a
new method for inventory of Osmoderma eremita (Scop., 1763)
(Coleoptera: Scarabaeidae) and other inhabitants of hollow trees
in Natura 2000 areas. J Insect Conserv 13:355–359
564 J Insect Conserv (2010) 14:555–565
123

Dubois G, Vignon V (2008) First results of radio-tracking of
Osmoderma eremita (Coleoptera: Cetoniidae) in French chestnut
orchards. Rev Ecol (Terre Vie) 63:123–130
Edmonds RL, Eglitis A (1989) The role of the Douglas-fir beetle and
wood borers in the decomposition of and nutrient release from
Douglas-fir logs. Can J For Res 19:853–859
Fontaine B, Bouchet P, Van Achterberg K, Alonso-Zarazaga MA,
Araujo R, Asche M, Aspock U, Audisio P, Aukema B, Baill N,
Balsamo M, Bank RA, Barnard P, Belfiore C, Bogdanowicz W,
Bongers T, Boxshall G, Burckhardt D, Camicas J-L, Chylarecki
P, Crucitti P, Deharveng L, Dubois A, Enghoff H, Faubel A,
Fochetti R, Gargominy O, Gibson D, Gibson R, Gomez Lopez
MS, Goujet D, Harvey MS, Heller K-G, Van Helsdingen P, Hoch
H, De Jong H, De Jong Y, Karsholt O, Los W, Lundqvist L,
Magowski W, Manconi R, Martens J, Massard JA, Massard-
Geimer G, Mcinnes SJ, Mendes LF, Mey E, Michelsen V,
Minelli A, Nielsen C, Nieto Nafrıa JM, Van Nieukerken EJ,
Noyes J, Pape T, Pohl H, De Prins W, Ramos M, Ricci C,
Roselaar C, Rota E, Schmidt-Rhaesa A, Segers H, Strassen RZ,
Szeptycki A, Thibaud J-M, Thomas A, Timm T, Van Tol J,
Vervoort W, Willmann R (2007) The European union’s 2010
target: putting rare species in focus. Biol Cons 139:167–185
Grove SJ (2002) Saproxylic insect ecology and the sustainable
management of forest. Ann Rev Ecol Syst 33:1–23
Hanula JL, Horn S, Wade DD (2006) The role of dead wood in
maintaining arthropod diversity on the forest floor. In: Insect
biodiversity and dead wood. Proceedings of a symposium for the
22nd international congress of entomology. Gen Tech Rep SRS-
93. Asheville, NC: US Department of Agriculture Forest
Service, Southern Research Station
Hedin J, Ranius T, Nilsson SG G, Smith H (2008) Restricted dispersal
in a flying beetle assessed by telemetry. Biodiv Conserv 17:675–
684
Hosmer DW, Lemeshow S (2000) Applied logistic regression, 2nd
edn. Wiley, New York
IFNC (2005) Inventario Forestale Nazionale e dei serbatoi di
Carbonio. Corpo Forestale dello Stato. Ispettorato Generale.
Trento
Johansson T (2006) The conservation of saproxylic beetles in boreal
forest: importance of forest management and dead wood
characteristics. Doctoral thesis, Swedish University of Agricul-
tural Science
Jonsson BG, Kruys N, Ranius T (2005) Ecology of species living on
dead wood—lessons for dead wood management. Silva Fennica
39:289–309
Larsson MC, Hedin J, Svensson GP, Tolasch T, Francke W (2003)
Characteristic odour of Osmoderma eremita identified as a male-
released pheromone. J Chem Ecol 29:575–587
Lindenmayer DB, Franklin JF, Fischer J (2006) General management
principles and a checklist of strategies to guide forest biodiver-
sity conservation. Biol Conserv 131:433–445
Luce JM (1996) Osmoderma eremita (Scopoli, 1763). In: van
Helsdingen PJ, Willemse L, Speight MCD (eds) Background
information on invertebrates of the habitats directive and the
Bern Convention. Part 1: Crustacea, Coleoptera and Lepidoptera.
Council of Europe, Strasbourg, pp 64–69
Matheny NP, Clark JR (1993) A photographic guide to the evaluation
of hazard trees in urban areas. International Society of Arbor-
iculture, USA
Mattheck C, Breloer H (1994) The body language of trees—a
handbook for failure analysis. Research for amenity trees Series
4. The British Department of the Environment. HMSO editions
Oleksa A, Ulrich W, Gawronski R (2006) Occurrence of the marbled
rose-chafer (Protaetia lugubris Herbst, Coleoptera, Cetoniidae)
in rural avenues in northern Poland. J Insect Conserv 10:241–
247
Oleksa A, Ulrich W, Gawronski R (2007) Host tree preferences of
hermit beetles (Osmoderma eremita scop., Coleoptera: Scara-
baeidae) in a network of rural avenues in Poland. Pol J Ecol
55:315–323
Pacyniak C (1992) Najstarsze drzewa w Polsce [The oldest trees in
Poland]—Wydawnictwo PTTK ,,Kraj’’, Warszawa (in Polish)
Radu S (2007) The ecological role of deadwood in natural forests. In:
Gafta D, Akeroyd J (eds) Nature conservation. Concepts and
practice. Springer, Heidelberg, pp 137–141
Ranius T (2006) Measuring the dispersal of saproxylic insects: a key
characteristic for their conservation. Popul Ecol 48:177–188
Ranius T, Hedin J (2001) The dispersal rate of a beetle, Osmodermaeremita, living in tree hollows. Oecologia 126:363–370
Ranius T, Nilsson SG (1997) Habitat of Osmoderma eremita Scop.
(Coleoptera: Scarabaeidae), a beetle living in hollow trees. J
Insect Conserv 1:193–204
Ranius T, Aguado LO, Antonsson K, Audisio P, Ballerio A,
Carpaneto GM, Chobot K, Gjurasin B, Hanssen O, Huijbregts
H, Lakatos F, Martin O, Neculiseanu Z, Nikitsky NB, Paill W,
Pirnat A, Rizun V, Ruicnescu A, Stegner J, Suda I, Szwako P,
Tamutis V, Telnov D, Tsinkevich V, Versteirt V, Vignon V,
Vogeli M, Zach P (2005) Osmoderma eremita (Coleoptera,
Scarabaeidae, Cetoniinae) in Europe. Anim Biodiv Conserv
28:1–44
Ranius T, Niklasson M, Berg N (2009) Development of tree hollows
in pedunculate oak (Quercus robur). Forest Ecol Manag
257:303–310
Ruffo S, Stoch F (2004) CKMap—5.1 Version. Copyright: Ministero
dell’Ambiente e della Tutela del Territorio—Direzione per la
Protezione della Natura
Schaffrath U (2003a) Osmoderma eremita (Scopoli, 1763). In:
Bundesamt fur Naturschutz (ed) Das Europaische Schutzgebi-
etssystem Natura 2000. Schriftenreihe fur Landschaftspflege und
Naturschutz 69(1):415–425
Schaffrath U (2003b) Zu Lebensweise, Verbreitung und Gefahrdung
von Osmoderma eremita (Scopoli, 1763) (Coleoptera, Scarab-
aeoidea, Cetoniidae, Trichiinae). Philippia 10:290–298
Stegner J (2002) Der Eremit, Osmoderma eremita (Scopoli, 1763)
(Col. Scarabaeidae), in Sachsen: Anforderungen an Schutzmaß-
nahmen fur eine prioritare Art der FFH-Richtlinie. Entomolog-
ische Nachrichten und Berichte
Svensson GP, Larsson MC, Hedin J (2003) Air sampling of its
pheromone to monitor the occurrence of Osmoderma eremita, a
threatened beetle inhabiting hollow trees. J Insect Conserv
7:189–198
World Conservation Monitoring Centre (1996) Cerambyx cerdoand Osmoderma eremita. In: IUCN 2006. 2006 IUCN Red List
of Threatened Species. \http://www.iucnredlist.org www.iucnred
list.org[. Downloaded on 21 January 2010
J Insect Conserv (2010) 14:555–565 565
123




















