
Public Opinion and Senate Confirmation of Supreme Court Nominees

IAWA Journal, Vol. 15 (2),1994: 171-184
CONFIRMATION AND SIGNIFICANCE OF BARTHOLIN'S METHOD FOR THE IDENTIFICATION OF THE WOOD OF PICEA AND IARIX
by
SusanE. Anagnost, RobertW.Meyer and CarideZeeuw1
Department of Wood Products Engineering, SUNY College of Environmental Science and Forestry, Syracuse, New York 13210, U.S.A.
Summary Eighty-seven Picea and Larix samples
from thirty-nine species were examined microscopically to confirm an accurate method of genus separation. The proportions and type of ray tracheid bordered pits as described by Bartholin (1979) were the only wood anatomical feature to correctly identify all specimens.
Picea subgroups closely followed those previously defined by the presence or absence of spiral thickenings in the longitudinal tracheids and ray tracheids. No Larix subgroups were evident.
Key words: Picea, Larix, ray tracheid pit border, spiral thickenings.
Introduction The wood anatomical features of Pice a
A. Dietr. and Larix Mill. are so similar that anatomical characteristics normally used to separate species must be used to distinguish the genera (Bailey 1909). Picea and Larix, genera of the Pinaceae, have axial and transverse resin canals, thick-walled epithelial cells, ray parenchyma cells with piceoid or cupressoid cross-field pits, and ray tracheids (Phillips 1949; Schweingruber 1978; Panshin & de Zeeuw 1980; Wilson & White 1986).
Bartholin (1979) was able to distinguish Picea from Larix using the relative proportions of pit types in the ray tracheids in his study of eleven Picea and eight Larix species. He determined that Picea ray tracheids have thick pit borders with angular extensions into the cell lumen and small pit apertures, while Larix ray tracheids have smooth pit borders and large pit apertures.
1) Deceased.
Bartholin (1979) identified four types of pit borders in the ray tracheids in radial sections of Picea and Larix. Larix (L) type pits, in cross section, have a smooth pit border and a large aperture, while Picea-l (P-l) type pits have a thickened border with a ridge or bump extending into the cell lumen and a small pit aperture. Picea-2 (P-2) pits also have a small pit aperture and a large extension or bump on the border (Bartholin 1979). An intermediate (I) pit type has pit apertures with intermediate diameter and a border that may be slightly thickened.
As reported by Bartholin (1979), in Larix the proportion of L pit borders is high, the proportion of P-l pit border types is very low and no P-2 types are found, whereas in Picea the proportion of L pit borders is low, P-l is high and P-2 types are almost always present but in variable amounts. Some workers have suggested Bartholin's method as a means of separating the genera (Panshin & de Zeeuw 1980; Wilson & White 1986). The main objective of this study is to examine a worldwide distribution of available samples of Picea and Larix to determine whether Bartholin's criteria are consistent throughout each genus. Anatomical features used to separate Picea and Larix, such as earlywood to latewood transition, spiral thickenings, number of epithelial cells and pit seriation may not always be reliable. This study will examine the accuracy of these features to determine their value in separating Picea and Larix.
Another objective of this study is to determine whether these samples form subgroups consistent with previously defined subgroups

172 IAWA Journal, Vo1.l5 (2), 1994
Table 1. Sample identification and origin.
Sample --1 Wood collection Collector and Origin
r Species number number
1 Picea abies (L.) Karst. BWCw3537 unknown Russia 2 BWCw3539 3 BWCw4211 R. Trendelenburg Germany 4 BWCw 5131 PRFw 4016 5 P. ajanensis Fisch. BWCw2836 Fujioka No. 071 Korea 6 P. asperata Mast. BWCw 13002 Tang No. 071 Szechuan, China 7 BWCw 13443 Aeronautical Research Lifan, Szechuan, China
Project No. 148 8 BWCw 13469 9 BWCwSL 2707
10 P. bicolor (Maxim.) Mayr BWCwSL 2784 Fujioka No. 72 Komaba, Tokyo, Japan 11 P. brachytyla (Franch.) Fritz. BWCw 13470 Aeronautical Research Lihaien, Szechuan, China
Project No. 43 12 P. breweriana S. Wats. BWCw8668 BWCw Project I 8668 Oregon, USA 13 P. chihuahuana Mart. BWCw20149 MEXFw Durango, Mexico 14 P. engelmannii (Parr.) Engelrn. BWCw8099 BWCw Project I 8099 Idaho, USA 15 BWCw8109 BWCw Project I 8109 Arizona, USA 16 BWCw8125 BWCw Project I 8125 Idaho, USA 17 BWCw8190 BWCw Project I 8190 Idaho, USA 18 BWCw8334 BWCw Project I 8334 Colorado, USA 19 BWCw sl. 2786 unknown unknown 20 P. glauca (Moench.) Voss BWCw8134 BWCw Project I 8134 Michigan 21 BWCw8253 BWCw Project I 8253 Minnesota 22 BWCw sl. 2785 unknown unknown
per D.A. Kribs 23 P. glauca var. albertiana BWCw4180 William Harlow Black Hills, SD, USA
(S. Brown) Sarg. 24 P. glehni (Fr. Schm.) Mast. BWCw4115 Fujioka No.3 Komaba, Tokyo, Japan 25 P. hondoensis Mayr BWCw827 unknown unknown 26 P. jezoensis BWCw4114 Fujioka No. 2 Komaba, Tokyo, Japan
(Sieb. & Zuce.) Carr. 27 BWCw sl. 2783
per D. A. Kribs 28 P. koraiensis Nakai BWCw2838 Fujioka No. 3259 Korea
Kyushu University 29 P. koyamai Shiras. SJRw 51422 Stern & Brizicky No. 718 Brett Pinetum
Fairfield, CT, USA 30 SJRw 51423 Stern & Brizicky No. 719 31 SJRw 51426 Stern & Brizicky No. 723 32 P. likiangensis (Franch.) Fritz. SJRw 51420 Stern & Brizicky No. 716 33 P. mariana (Mill.) BWCw8010 BWCw Project I 8010 New York, USA
Britt., Sterns & Pogg. 34 BWCw8135 BWCw Project I 8135 Michigan, USA
35 BWCw8201 BWCw Project 18201 New York, USA
36 BWCw8254 BWCw Project I 8254 Minnesota, USA 37 P. maximowiczii Reg. MADw4453 TWfw5 Japan 38 P. mexicana Mart. BWCw20155 MEXFwX-lOl Nuevo Leon, Mexico
39 P. montigena Mast. SJRw 23059 Fan Memorial Institute China Field No. 093
40 P. morrisonicola Hayata BWCw3309 R. Kanehira No.9 Taiwan 41 BWCw 5949 Fujioka No. 15 Komaba, Japan 42 BWCw 15935 R. Kanehira No.9 Taiwan 43 P. neovitchii Mast. BWCw 15409 SJRw 24458 Fan China
Memorial Inst., Tang. 44 P. omorika (Paneic.) Purkyne BWCw3137 per Dr. Rusnov Austria 45 P. orienta lis (L.) Carr. SJRw 19862 per Dr. A. H. Graves Asia Minor
Brooklyn Botanical Gardens 46 SJRw 51428 Stern & Brizicky No. 725 Brett Pinetum
Fairfield, CT, USA 47 P. polita (Sieb. & Zucc.) Carr. USw 23686 TOFOw 1250 Japan

Anagnost, Meyer & De Zeeuw - Wood identification of Picea and Larix 173
(Table 1 continued)
Sample J Wood collection Collector and Origin
rSpecies number number
48 P. pofita (Sieb. & Zucc.) Carr. MADw20342 TWTw842 Japan 49 P. pungens Engelm. BWCw8443 BWCw Project I 8443 Utah, USA 50 BWCw8466 BWCw Project I 8466 Colorado, USA 51 P. rubens Sarg. BWCw8011 BWCw Project 18011 New York, USA 52 BWCw8476 BWCw Project I 8476 Maine, USA 53 BWCw8661 BWCw Project I 8661 North Carolina, USA 54 P. sitchensis (Bong.) Carr. BWCw 8154 BWCw Project I 8154 California, USA 55 BWCw8448 BWCw Project I 8448 Washington, USA 56 BWCw8798 BWCw Project I 8798 Vancouver 57 P. smithiana (Wall.) Boiss. BWCw sl. 1442 unknown India
per H. P. Brown 58 P. wilsonii Mast. BWCw4669 C.F. Le, Fan Memorial Tung Ling, China
Institute, Field No. 86 59 Larix decidua Mill. BWC sl. 27802 unknown unknown
per D.A. Kribs 60 BWCw5130 PRFw 4012 Great Britain 61 L. gmelini (Rupr.) BWCw2834 Fujioka No. 3255 Korea
(= L. dahurica) Kyushu University 62 BWCw2869 Fujioka No. 3299 Korea
Kyushu University 63 BWCw 5193 Fujioka NO.3 Karafutu University Forest,
Japan 64 L. kaempleri (Lamb.) Carr. BWCw5180 Fujioka No. 76 Chichibu University Forest,
Japan 65 BWCw 10963 PRFw 4854 Great Britain 66 L. faricina (Du Roi) K. Koch BWCw 130 H.P. Brown Cicero Bogs,
New York, USA 67 BWCw7509 PRFw 2952 Newfoundland 68 BWCw8090 BWCw Project I 8090 New York, USA 69 BWCw8155 BWCw Project I 8155 Minnesota, USA 70 BWCw8652 BWCw Project I 8652 Wisconsin, USA 71 BWCw 18048 MADw 2946 72 BWC sl. 27812 D.A. Kribs 7034 unknown
per D. A. Kribs 73 BWC sl. 22032 A. C. Fegel No. 74 unknown 74 L. leptolepis (Sieb. & Zucc.) BWCw4012 Y. Sugiur Komaba. Japan
Gord. 75 BWCw 14901 TAIw0068 Taiwan
National Taiwan University 76 L. fyallii ParI. BWCw8604 BWCw Project I 8604 Washington, USA 77 BWCw8607 BWCw Project I 8607 Idaho, USA 78 L. occidentalis Nutt. BWCw 196 unknown Wyoming, USA 79 BWCw634 unknown unknown 80 BWCw 1554 unknown Flathead National Forest,
Montana, USA 81 BWCw 8123 BWCw Project I 8123 Idaho, USA 82 BWCw8325 BWCw Project I 8325 Oregon, USA 83 BWCw9688 USw 3684 United States 84 BWCw 11788 University of California California, USA
No.7 85 BWC sl. 27822 D.A. Krib USA
per D.A. Kribs 86 L. potanini Batal. BWCw 15256 Fan Memorial Institute Szechuan, China 87 L. russica (Sab. et Trautv.)
(= L. sibirica Ledeb.) BWCw3538 unknown Russia-Archangel District
BWCw = Brown Wood Collection, SUNY College of Environmental Science and Forestry, Syracuse, NY. MADw = U.S. Forest Products Laboratory, Madison, Wisconsin. SJRw Samuel J. Record Wood Collection, as supplied by the U.S. Forest Products Lab., Madison WI. USw Wood collection of the Smithsonian Institute, Washington, D.C.
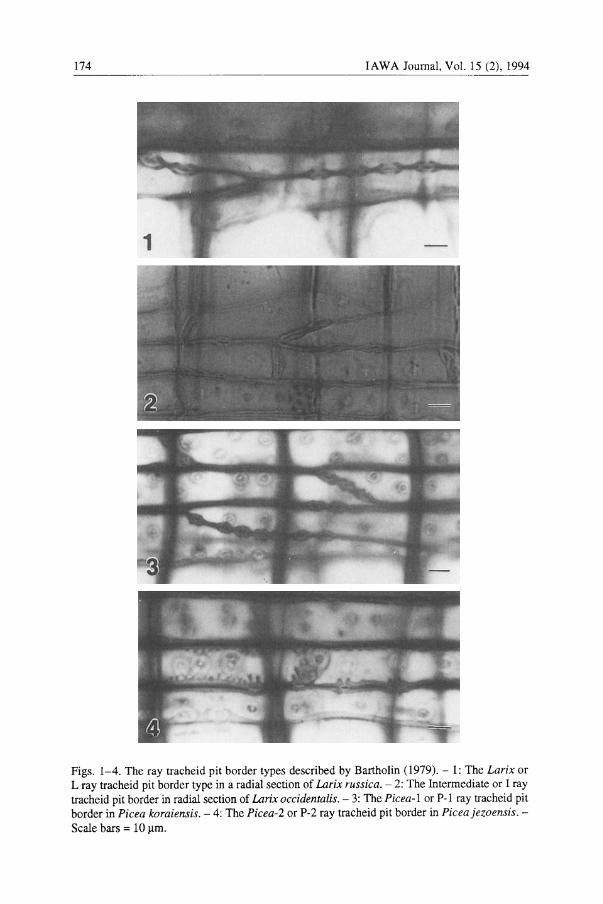
174 IAWA Journal, Vol. 15 (2),1994
Figs. 1-4. The ray tracheid pit border types described by Bartholin (1979). - 1: The Larix or L ray tracheid pit border type in a radial section of Larix russica. - 2: The Intermediate or I ray tracheid pit border in radial section of Larix occidentalis. - 3: The Picea-1 or P-1 ray tracheid pit border in Picea koraiensis. - 4: The Picea-2 or P-2 ray tracheid pit border in Piceajezoensis.Scale bars = 10 ~m.

Anagnost, Meyer & De Zeeuw - Wood identification of Picea and Larix 175
(Sudo 1968; Budcevicz 1961). Sudo (1968) divided the genus Picea into two subgroups based on the extent of spiral thickenings in the longitudinal and ray tracheids. The group which has spiral thickenings consistently in both earlywood and latewood axial trachcids also has more pronounced spiral thickenings in the ray tracheids (Sudo 1968). A method of subdividing the genus Larix based on the number of epithelial cells in the horizontal resin canals has been described (Budcevicz 1961).
A third objective of this study is to determine the relationship between the presence or absence of spiral thickenings in Picea and Larix and the types of ray tracheid pit borders. Since spiral thickenings (Bailey 1909; Sudo 1968) and dentations (Phillips 1949) were reported as occurring often in Picea ray tracheids and less often in Larix ray tracheids, there may be a relationship between their presence and the type of ray tracheid pit border present (Bartholin's criteria). Also to be examined is the distinctive structure of the P-l and P-2 pit borders and how it relates to cell wall sculpturing of the ray tracheid.
Materials and Methods Fifty-eight samples representing thirty spe
cies of Picea and twenty-nine samples representing nine Larix species were examined (Table 1). Species determination, origin, wood collection numbers, collector and collector's numbers of each sample when known are indicated (Table 1). Specific names follow the format of A Handbook of Coniferae and Ginkgoaceae (Dallimore & Jackson 1967), and The Checklist of Trees of the United States (Little 1979).
Many of the specimens were prepared slides obtained from the H.P. Brown Memorial Wood Collection of the College of Environmental Science and Forestry, Syracuse, New York (BWCw). For all other samples, slides were prepared of water-soaked blocks by sectioning 10-20 ~m thick with a sliding microtome, staining with hematoxylin and safranin-O, dehydrating and mounting for light microscopic observation.
A light microscope was used to observe those features previously determined to be
useful in separating Picea and Larix: the proportion and type of ray tracheid bordered pits (Bartholin's method), the nature of the earlywood to latewood transition zone, pit seriation of longitudinal tracheids in the earlywood, the number of epithelial cells surrounding the horizontal resin canals and the presence or absence of spiral thickenings in longitudinal and ray tracheids.
The pit border types follow the description of Bartholin (1979) and are shown in Figures 1-5. Fifty pits were counted, beginning at one comer of a radial section and scanning back and forth, skipping those pits not clearly sectioned through their central portion. During examination the species name was covered to prevent any bias. The mean and standard deviation of the number of each ray tracheid pit type observed was obtained to compare the two genera and to compare these results with data obtained by others (Bartholin 1979; Sudo 1968).
A discriminate analysis was used in this study to test the percentages of the four ray tracheid pit types as a means to separate the {wo genera. The linearised discriminate function is based on the multivariate t-test which tests for the equality of means of two or more populations based on Mahalanobis distance (Morrison 1976). In order to test for membership in either genus, the t 2 statistic was
Larix
Intermediate ~
Picea-l
Picea-2
Fig. 5. Diagrams of the ray tracheid pit borders as described by Bartholin (1979).
176 IAWA Journal, VoL 15 (2), 1994
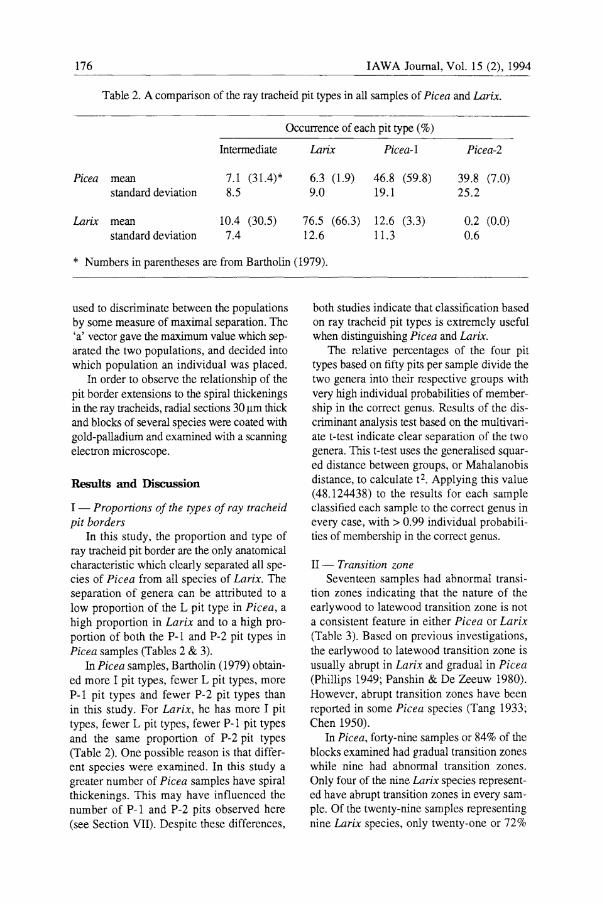
Table 2. A comparison of the ray tracheid pit types in all samples of Picea and Larix.
Occurrence of each pit type (% )
Intermediate
Picea mean 7.1 (31.4)* standard deviation 8.5
Larix mean 10.4 (30.5) standard deviation 7.4
Larix
6.3 (1.9) 9.0
76.5 (66.3) 12.6
Picea-l
46.8 (59.8) 19.1
12.6 (3.3) 11.3
Picea-2
39.8 (7.0) 25.2
0.2 (0.0) 0.6
* Numbers in parentheses are from Bartholin (1979).
used to discriminate between the populations by some measure of maximal separation. The 'a' vector gave the maximum value which separated the two populations, and decided into which population an individual was placed.
In order to observe the relationship of the pit border extensions to the spiral thickenings in the ray tracheids, radial sections 30 /lm thick and blocks of several species were coated with gold-palladium and examined with a scanning electron microscope.
Results and Discussion
I - Proportions of the types of ray tracheid pit borders
In this study, the proportion and type of ray tracheid pit border are the only anatomical characteristic which clearly separated all species of Picea from all species of Larix. The separation of genera can be attributed to a low proportion of the L pit type in Picea, a high proportion in Larix and to a high proportion of both the P-l and P-2 pit types in Picea samples (Tables 2 & 3).
In Picea samples, Bartholin (1979) obtained more I pit types, fewer L pit types, more P-l pit types and fewer P-2 pit types than in this study. For Larix, he has more I pit types, fewer L pit types, fewer P-l pit types and the same proportion of P-2 pit types (Table 2). One possible reason is that different species were examined. In this study a greater number of Picea samples have spiral thickenings. This may have influenced the number of P-l and P-2 pits observed here (see Section VII). Despite these differences,
both studies indicate that classification based on ray tracheid pit types is extremely useful when distinguishing Picea and Larix.
The relative percentages of the four pit types based on fifty pits per sample divide the two genera into their respective groups with very high individual probabilities of membership in the correct genus. Results of the discriminant analysis test based on the multivariate t-test indicate clear separation of the two genera. This t-test uses the generalised squared distance between groups, or Mahalanobis distance, to calculate t2. Applying this value (48.124438) to the results for each sample classified each sample to the correct genus in every case, with> 0.99 individual probabilities of membership in the correct genus.
II - Transition zone Seventeen samples had abnormal transi
tion zones indicating that the nature of the early wood to latewood transition zone is not a consistent feature in either Picea or Larix (Table 3). Based on previous investigations, the earlywood to latewood transition zone is usually abrupt in Larix and gradual in Pice a (Phillips 1949; Panshin & De Zeeuw 1980). However, abrupt transition zones have been reported in some Picea species (Tang 1933; Chen 1950).
In Picea, forty-nine samples or 84% of the blocks examined had gradual transition zones while nine had abnormal transition zones. Only four of the nine Larix species represented have abrupt transition zones in every sample. Of the twenty-nine samples representing nine Larix species, only twenty-one or 72%

Anagnost, Meyer & De Zeeuw - Wood identification of Picea and Larix 177
have abrupt transition zones while eight samples have abnormal transition zones.
Since compression wood alters the nature of the transition zone to the opposite condition (Panshin & de Zeeuw 1980), those Picea samples with abrupt transition zones and Larix samples with gradual transition zones were checked for compression wood. Of these, only Picea polita USw 23686 and MADw 20342, Larix gmelini BWCw 5193, and Larix laricina BWCw 8090 were found to have compression wood in the material examined. Thirteen of the eighty-seven samples remained misclassified using the transition zone as the sole criterion.
III - Pit seriation of longitudinal tracheids Based on previous studies, intertracheid
pitting is commonly uniseriate in Picea and biseriate in Larix, with the exception of Picea sitchensis (Bong.) Carr. in which biseriate rows of pits are a regular feature separating it from other North American Picea species (Panshin & De Zeeuw 1980). Also, several Chinese species of Picea are reported as having biseriate rows of pits (Tang 1933; Chen 1950). In this study, the seriation of the intertracheid pitting is another inconsistent feature in both Picea and Larix (Table 3). In three of the thirty Pice a species the intertracheid pitting is biseriate: Pieea ajanensis, P. koraiensis, and P. morrisonieola. Ten Pieea species have at least one sample with part biseriate pitting and part uniseriate pitting. They are P. abies, P. asperata, P. bieolor, P. braehytyla, P. engelmanni, P. neovitehii, P. omorika, P. polita, P. sitehensis, and P. wilsonii. Sixteen of fifty-eight or 28% of all Pieea samples examined had some biseriate pitting. These samples would be misclassified using pit seriation as the sole criterion for identification.
Nine out of twenty-nine or 31 % of the Larix samples have uniseriate intertracheid pitting. These samples would be misclassified using this method alone. For some species, pit seriation was consistent. Both samples of Larix lyalli have uniseriate pitting, while samples of L. decidua, L. gmelini, and L. russica have biseriate pitting. The pit seriation of the other Larix species is inconsistent in the samples studied here (Table 3).
IV - Epithelial cells in the horizontal resin canals
In this study, of the eighty-five samples examined (two slides lacked tangential sections with no wood available) fifty-four (64%) were classified correctly using the number of epithelial cells as the discriminating factor. Sixteen of fifty-eight Picea samples (28%) and fifteen of twenty-seven Larix samples (56%) were misclassified using this method. The number of epithelial cells surrounding the resin canals is reported to be seven to nine in Picea and nine to twelve in Larix (Jane 1956). The overlapping ranges make this method inherently unreliable.
Twenty of thirty Picea species (represented by forty-two samples) were classified correctly (Table 3). Those species misclassified are: two of four P. abies samples, P. ajanensis, P. bicolor, one of five P. engelmannii samples, one of two P. glauea samples, P. glauca var. albertiana, both P. jezoensis samples, P. koraiensis, P. neovitehii and one of three P. sitchensis samples.
For two Larix species, all samples were classified correctly: L. leptolepis (2 samples) and L. sibirica (l sample) (Table 3). Seven of eight L. laricina samples were also correctly classified based on the number of epithelial cells. For L. decidua, L. kaempferi and L. lyalli, one of two samples were classified correctly. Only two often L. occidentalis samples were correctly classified. The three L. gmelini samples were misclassified.
Sudo (1968) states that Budcevicz (1961) divides Larix into two groups based on the number of epithelial cells: those having 8-12 and those having greater than 12 epithelial cells per resin canal. In this study, most Larix samples had fewer than twelve epithelial cells. Four of the seven L. laricina samples had an average greater than twelve epithelial cells. Three Larix samples had from 7 to 17 epithelial cells but an average fewer than twelve. No similar subdivisions could be defined; either the number of epithelial cells overlapped both ranges, or were inconsistent within species.
The number of epithelial cells is often valuable for identification, being nearly constant in each genus; however, in the first 30-40 growth rings and depending on the growth rate, the number of epithelial cells does vary
178 IAWA Journal, Vol. 15 (2),1994
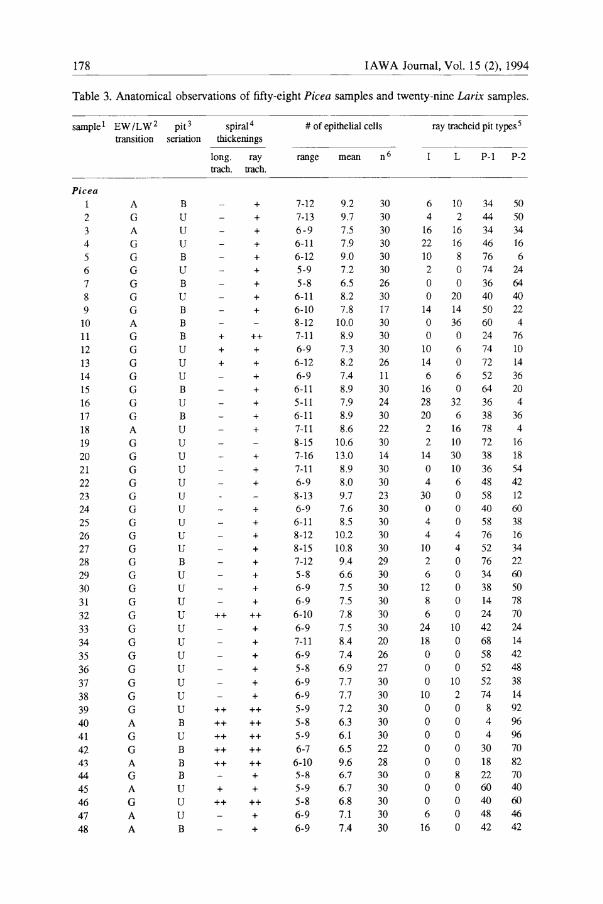
Table 3. Anatomical observations of fifty-eight Picea samples and twenty-nine Larix samples.
sample 1 EW IL W 2 pit 3 spiral 4
Picea 1 2 3 4
5 6 7 8 9
10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39
40 41 42 43
44 45 46 47 48
transition seriation thickenings
A G A G G G G G G A G G G G G G G A G G G G G G G G G G G G G
G G G G G G G G A G G A G A G A A
B U U U B U B U B B B U U U B U B U U U U U U U U U U B U U U U U U U U U U U B U B B B U U U B
long. ray trach. trach.
+ + +
++
++ ++ ++ ++ ++
+
++
+
+ + + +
+ + + +
++ + + + + + +
+
+
+ +
+
+
+
+ + + +
+
++
+ + + + + +
++ ++ ++ ++ ++ +
+
++ + +
# of epithelial cells
range mean n 6
7-12 7-13 6-9 6-11 6-12 5-9 5-8 6-11 6-10 8-12 7-11 6-9 6-12 6-9 6-11 5-11 6-11 7-11
8-15 7-16 7-11
6-9 8-13 6-9 6-11 8-12 8-15 7-12 5-8 6-9 6-9 6-10 6-9 7-11 6-9 5-8 6-9 6-9 5-9 5-8 5-9 6-7
6-10 5-8
5-9 5-8 6-9 6-9
9.2 9.7 7.5 7.9 9.0 7.2 6.5 8.2 7.8
10.0 8.9 7.3 8.2 7.4 8.9 7.9 8.9 8.6
10.6 13.0 8.9 8.0 9.7 7.6 8.5
10.2 10.8 9.4
6.6 7.5 7.5 7.8 7.5 8.4 7.4 6.9 7.7 7.7 7.2 6.3 6.1 6.5 9.6 6.7
6.7 6.8 7.1 7.4
30 30 30 30 30 30 26 30 17 30 30 30 26 11 30 24 30 22 30 14 30 30 23 30 30 30 30 29 30 30 30 30 30 20 26 27 30 30 30 30 30 22 28 30
30 30 30 30
ray tracheid pit types 5
L P-l P-2
6 10 34 50 4 2 44 50
16 16 34 34 22 16 46 16 10 8 76 6 2 0 74 24 o 0 36 64 o 20 40 40
14 14 50 22 o 36 60 4 o 0 24 76
10 6 74 10
14 0 72 14 6 6 52 36
16 0 64 20 28 32 36 4 20 6 38 36 2 16 78 4 2 10 72 16
14 30 38 18 o 10 36 54 4 6 48 42
30 0 58 12 o 0 40 60 4 0 58 38 4 4 76 16
10 4 52 34 2 0 76 22 6 0 34 60
12 0 38 50 8 0 14 78 6 0 24 70
24 10 42 24 18 0 68 14 o 0 58 42 o 0 52 48 o 10 52 38
10 2 74 14 o 0 8 92 o 0 4 96 o 0 4 96 o 0 30 70 o 0 18 82 o 8 22 70 o 0 60 40 o 0 40 60 6 0 48 46
16 0 42 42
Anagnost, Meyer & De Zeeuw - Wood identification of Picea and Larix 179
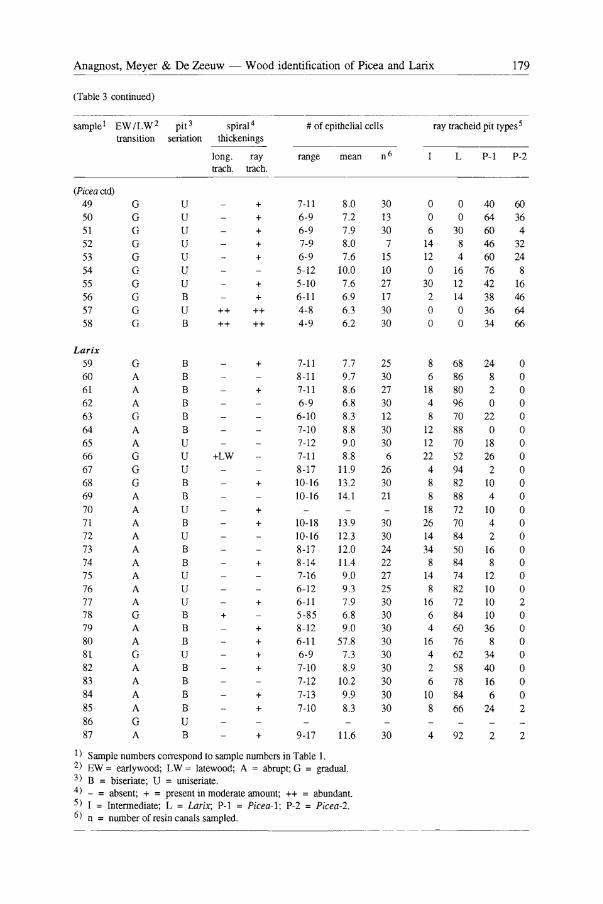
(Table 3 continued)
sample! EW ILW2 pit 3 spiral 4 # of epithelial cells transition seriation thickenings
long. ray range mean n 6
(Piceactd) 49 G 50 G 51 G 52 G 53 G 54 G 55 G 56 57
58
Larix 59 60 61
62
63
64 65 66 67
68 69
70
71 72 73
74 75
76 77 78 79
80 81 82
83
84
85
86
87
G
G G
G A A A G
A A G
G G A A A A A
A A A A G A A G A A A A
G A
u u u u u u U B U B
B B B B B B U U U B B U B U B B U U U B B B U B
B B B
U B
trach. trach.
++ ++
+LW
+
+ +
+ + +
+
+ ++ ++
+
+
+
+
+
+
+
+
+
+
+
+
+
+
7-11 6-9
6-9 7-9 6-9
5-12 5-10
6-11
4-8 4-9
8.0 7.2
7.9
8.0 7.6
10.0
7.6 6.9
6.3 6.2
7-11 7.7 8-11 9.7
7-11 8.6 6-9 6.8
6-10 8.3 7-10 8.8 7-12 9.0
7-11 8.8
8-17 11.9 10-16 13.2
10-16 14.1
10-18 13.9 10-16 12.3
8-17 12.0 8-14 11.4 7-16 9.0 6-12 9.3 6-11 7.9 5-85 6.8 8-12 9.0
6-11 57.8 6-9 7.3
7-10 8.9
7-12 10.2
7-13 9.9
7-10 8.3
9-17 11.6
1) Sample numbers correspond to sample numbers in Table 1. 2) EW = earlywood; LW = latewood; A = abrupt; G = gradual. 3) B = biseriate; U = uniseriate. 4) _ = absent; + = present in moderate amount; ++ = abundant. 5) I = Intermediate; L = Larix; P-1 = Picea-1; P-2 = Picea-2. 6) n = number of resin canals sampled.
30 13 30
7 15
10
27 17
30
30
25
30 27
30 12
30 30 6
26
30 21
30
30 24 22 27 25
30 30 30 30 30
30
30
30
30
30
ray tracheid pit types 5
o o 6
14 12
o 30 2 o o
L
o o
30 8 4
16 12
14
o o
8 68 6 86
18 80
4 96 8 70
12 88
12 70 22 52
4 94
8 82
8 88 18 72 26 70 14 84
34 50 8 84
14 74
8 82 16 72
6 84 4 60
16 76 4 62
2 58
6 78
10 84
8 66
4 92
P-1 P-2
40 64
60 46
60
76 42
38
36 34
24
8 2
o 22 o
18
26
2 10
4 10
4 2
16 8
12
10 10 10 36 8
34
40
16
6 24
2
60 36 4
32 24
8 16
46 64
66
o o o o o o o o o o o o o o o o o o 2
o o o o o o o 2
2
180 IAWA Journal, VoL 15 (2), 1994
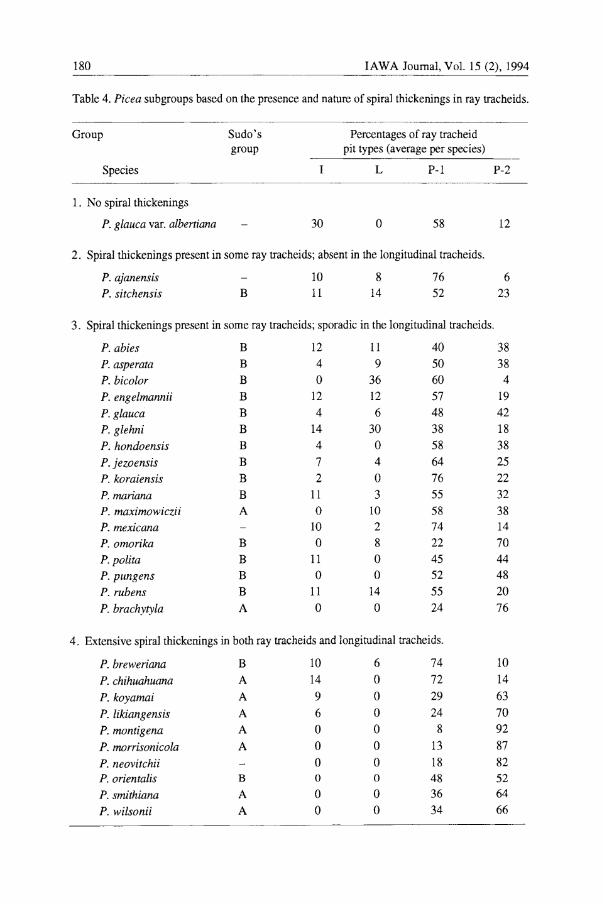
Table 4. Picea subgroups based on the presence and nature of spiral thickenings in ray tracheids.
Group Sudo's Percentages of ray tracheid group pit types (average per species)
Species L P-l P-2
1. No spiral thickenings
P. glauca var. albertiaJ1a 30 0 58 12
2. Spiral thickenings present in some ray tracheids; absent in the longitudinal tracheids.
P. ajanensis 10 8 76 6 P. sitchensis B 11 14 52 23
3. Spiral thickenings present in some ray tracheids; sporadic in the longitudinal tracheids.
P. abies B 12 11 40 38
P. asperata B 4 9 50 38
P. bicolor B 0 36 60 4
P. engelmannii B 12 12 57 19
P. glauca B 4 6 48 42
P. glehni B 14 30 38 18
P. hondoensis B 4 0 58 38
P. jezoensis B 7 4 64 25
P. koraiensis B 2 0 76 22
P. mariana B II 3 55 32
P. maximowiczii A 0 10 58 38 P. mexicana 10 2 74 14
P.omorika B 0 8 22 70
P.polita B 11 0 45 44
P. pungens B 0 0 52 48
P. rubens B 11 14 55 20
P. brachytyla A 0 0 24 76
4. Extensive spiral thickenings in both ray tracheids and longitudinal tracheids.
P. breweriaJ1a B 10 6 74 10
P. chihuahuaJ1a A 14 0 72 14
P. koyamai A 9 0 29 63
P. likiangensis A 6 0 24 70
P. montigena A 0 0 8 92
P. morrisonicola A 0 0 13 87
P. neovitchii 0 0 18 82 P. orientalis B 0 0 48 52
P. smithiana A 0 0 36 64
P. wilsonii A 0 0 34 66

Anagnost, Meyer & De Zeeuw - Wood identification of Picea and Larix 181
(Sudo 1968). According to Sudo (1968), in Picea the number of epithelial cells per canal increases with increasing distance from the pith, the average number at the thirtieth to fortieth ring for each species being 6.5 to 8. The average number increases rapidly to the thirtieth or fortieth ring and then the rate levels off. Growth rate also affects the rate of increase in the number of epithelial cells; slow growth results in less of an increase in the number of epithelial cells (Sudo 1968). In a small wood sample of unknown age and origin the number of epithelial cells could be unreliable. Also, for routine identification this feature is not of practical use; the number of correct classifications is not large enough for the feature to be reliable.
v - Spiral thickenings In this study, spiral thickenings are found
in the ray tracheids of all but one Picea species (Table 4). This species was placed in group I (Table 4). Two Picea species (four samples) have spiral thickenings in ray tracheids only (Group II). Six species (fifteen samples) have spiral thickenings in ray tracheids with sporadic occurrence in longitudinal tracheids (Group III), while the remaining twelve species (fifteen samples) have extensive spiral thickenings in both ray and longitudinal tracheids (Group IV) (Table 4). Sudo (1968) divides Picea into two subgroups, A and B. Group A has extensive spiral thickenings in all longitudinal tracheids and has well developed spiral thickenings in the ray tracheids. In contrast, group B has spiral thickenings only in the latewood longitudinal tracheids and less prominently in the ray tracheids. In this study, twelve of sixteen samples with extensive spiral thickenings in both longitudinal and ray tracheids (Group IV) are in Sudo's Group A (Table 4). Thirty-three of thirty-five samples in Group III (less extensive spiral thickenings in the ray tracheids and few or none in the longitudinal tracheids) are also in Sudo's Group B (Table 4).
The presence of spiral thickenings in Picea has been reported by others. Bailey (1909) reported spiral thickenings in the ray and longitudinal tracheids of Picea sitchensis (Bong.) Carr. and Picea maxirrwwiczii Reg. ex Mast., and concluded that their presence in the lon-
gitudinal tracheids coincided with their presence in the ray tracheids. Wilson and White (1986) reported that spiral thickenings occur in longitudinal tracheids of both Picea and Larix. Record (1925) reported that the fine dentations occasionally present in ray tracheids of Picea and Larix are actually spiral thickenings. The difference in prevalence of spiral thickenings in ray tracheids of Picea and Larix has been recognised. What were referred to as dentate ray tracheid walls have been reported in both Picea and Larix (Record 1925; Sudo 1968; Core et al. 1979), but they are reported as being less prevalent in Larix (Phillips 1949; Jane 1956).
The results of this study confirm previous observations (Phillips 1949; Jane 1956) that spiral thickenings occur less often in Larix than in Picea. Only one Larix sample, L. occidentalis (BWCw No. 196) has spiral thickenings in the longitudinal tracheids. Spiral thickenings are present in fourteen samples in the latewood ray tracheids (Table 3).
The presence or absence of spiral thickenings appears to influence the proportions of pit types. In Picea, most Group IV species (extensive spiral thickenings in longitudinal and ray tracheids) have high proportions of P-2 pit types and few or no L pit types (Table 4). Most of these samples are in Sudo's subgroup A (Sudo 1968). An example of a Group IV species is Picea rrwntigena which was found to have 92% P-2 pit borders and 8% P-l pit borders (Fig. 6; Table 3). Most Group III species (spiral thickenings in some ray tracheids; sporadic in longitudinal tracheids) have a high proportion of P-I pits and a greater number of I pits. Those Larix samples with the greatest proportions of P-l pits also have spiral thickenings in the ray tracheids. Spiral thickenings were also observed in ray tracheids of the three Larix samples which also contained P-2 pits.
VI - The value of these anatomical characteristics in identification of wood samples
Considering the five anatomical features: earlywood to latewood transition zone, pit seriation of longitudinal tracheids, the number of epithelial cells surrounding the horizontal resin canals, the presence or absence of spiral thickenings and the proportion of

182 IAWA Journal, Vol. 15 (2), 1994
Fig. 6. An example of a Group IV Picea species with spiral thickenings in both the longitudinal tracheids and ray tracheids of Picea nwntigena. Arrows point to spiral thickenings in ray tracheids. The double arrow indicates a P-2 bit border. Bar = 100 ~m.
ray tracheid pit border types, it is obvious that they are all of some value in identification, especially when used in combination. In forty-one of the fifty-eight Picea samples (71 %) and seventeen of twenty-nine Larix samples (59%) correct genus identification could be made without looking at the ray tracheid pit types. However, there are also some samples with highly doubtful determinations based on only the first four characteristics (see samples 10, 43, 66, 67, 68, 81; Table 3). In this study, the only feature to correctly classify every sample is the ray tracheid pit type. Other features are often correct, but none are definitive enough to insure correct classification.
VII - The ray tracheid pit border structure in relation to wall sculpturing
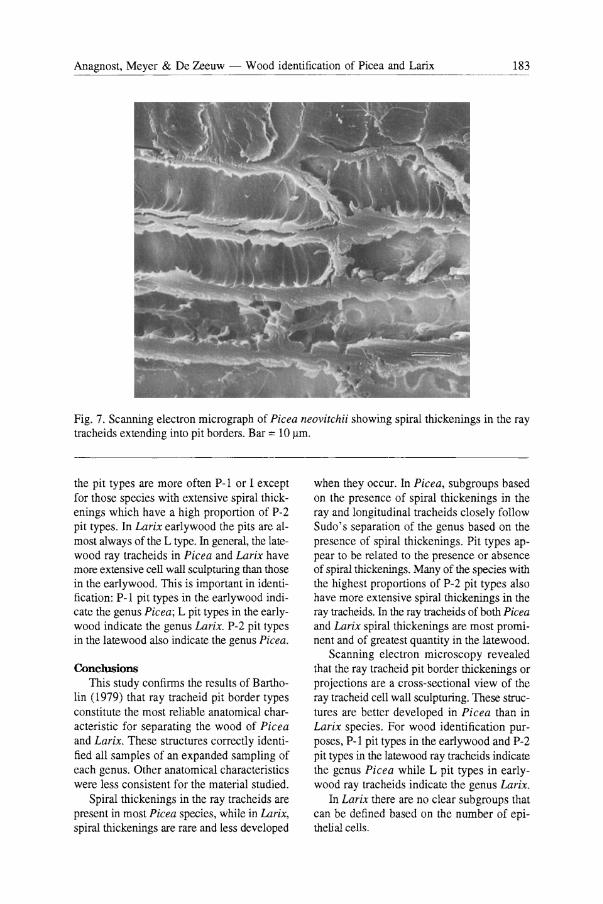
In the radial section of Picea species the projections extending into the cell lumen from the ray tracheid pit borders are actually cell wall thickenings which often appear as small dentations when viewed with the light microscope. Scanning electron microscopy revealed that the pit border projections are actually cross sections of spiral thickenings (Fig. 7).
The extent of wall sculpturing varies within each genus from earlywood to latewood. In Picea species the latewood ray tracheids have a greater proportion of P-2 pit types. In Larix species, the latewood ray tracheids can have L, I, or P-1 pit types. In Picea earlywood,
Anagnost, Meyer & De Zeeuw - Wood identification of Picea and Larix 183
Fig. 7. Scanning electron micrograph of Picea neovitchii showing spiral thickenings in the ray tracheids extending into pit borders. Bar = 10 ~m.
the pit types are more often P-I or I except for those species with extensive spiral thickenings which have a high proportion of P-2 pit types. In Larix earlywood the pits are almost always of the L type. In general, the latewood ray tracheids in Picea and Larix have more extensive cell wall sculpturing than those in the earlywood. This is important in identification: P-I pit types in the earlywood indicate the genus Picea; L pit types in the earlywood indicate the genus Larix. P-2 pit types in the latewood also indicate the genus Picea.
Conclusions This study confirms the results of Bartho
lin (1979) that ray tracheid pit border types constitute the most reliable anatomical characteristic for separating the wood of Picea and Larix. These structures correctly identified all samples of an expanded sampling of each genus. Other anatomical characteristics were less consistent for the material studied.
Spiral thickenings in the ray tracheids are present in most Picea species, while in Larix, spiral thickenings are rare and less developed
when they occur. In Pice a, subgroups based on the presence of spiral thickenings in the ray and longitudinal tracheids closely follow Sudo's separation of the genus based on the presence of spiral thickenings. Pit types appear to be related to the presence or absence of spiral thickenings. Many of the species with the highest proportions of P-2 pit types also have more extensive spiral thickenings in the ray tracheids. In the ray tracheids of both Picea and Larix spiral thickenings are most prominent and of greatest quantity in the latewood.
Scanning electron microscopy revealed that the ray tracheid pit border thickenings or projections are a cross-sectional view of the ray tracheid cell wall sculpturing. These structures are better developed in Picea than in Larix species. For wood identification purposes, P-l pit types in the earlywood and P-2 pit types in the latewood ray tracheids indicate the genus Picea while L pit types in earlywood ray tracheids indicate the genus Larix.
In Larix there are no clear subgroups that can be defined based on the number of epithelial cells.

184
Acknowledgements The authors thank Richard Eyde of the
Smithsonian Institution and Donna Christensen of the USDA Forest Products Laboratory at Madison, Wisconsin for their contributions of wood samples. This study was completed as part of the requirement for a MSc degree in the Department of Wood Products Engineering, SUNY College of Environmental Science and Forestry, Syracuse, New York.
References Bailey, I.W. 1909. The structure of wood in
the Pinae. Bot. Gaz. 48: 47-55. Bartholin, T. 1979. The Picea-Larix problem.
IAWABull. n.s.I: 68-70. Budcevicz, E.V. 1961. The wood of the Pina
ceae. Anatomical structure and keys for the identification of the genera and species: 68-70. Leningrad.
Chen, c.L. 1950. Anatomical structure and identification of thirty Chinese woods. MSc Thesis. SUNY College of Forestry, Syracuse, New York.
Core, H.A., W.A. Cote & A.c. Day. 1979. Wood structure and identification. 2nd Ed. Syracuse University Press, New York.
Dallimore, W. & A.B. Jackson. 1967. A handbook of Coniferae and Ginkgoaceae. St. Martin's Press, New York.
IAWA Journal, Vol. 15 (2),1994
Jane, F.W. 1956. The structure of wood. The MacMillan Co., New York.
Little, E.L., Jr. 1979. Checklist of United States trees (native and naturalized). Agric. Handbook 541. USDA Forest Service, Washington, D.C.
Morrison, D. F. 1976. Multivariate statistical methods. Ed. 2. McGraw-Hill, New York.
Panshin, A.J. & c. de Zeeuw. 1980. Textbook of wood technology. McGraw-Hill, New York.
Phillips, E.W.J. 1949. Identification of softwoods by their microscopic structure. For. Prod. Res. Bull. 22. HMSO, London.
Record, S.J. 1925. Spiral tracheids and fiber tracheids. Tropical Woods 3: l3.
Schweingruber, F. H. 1978. Microscopic wood anatomy. Swiss Federal Inst. For. Res., Zurich.
Sudo, S. 1968. Anatomical studies on the wood of species of Picea with some considerations on their geographical distribution and taxonomy. Bull. Govt. For. Exp. Sta. No. 215, Tokyo.
Tang, Y. 1933. Timber studies of Chinese trees. IV. Anatomical studies and identification of Chinese softwoods I. Bull. Fan Mem. Inst. BioI. 4: 7.
Wilson, K. & D.J.B. White. 1986. The anatomy of wood. Stobart & Son Ltd., London.
Copyright © 2022 FDOKUMEN











![genetic variation of norway spruce (picea abies [l.] karst ...](https://static.fdokumen.com/doc/165x107/633d6582b988f9753c06267d/genetic-variation-of-norway-spruce-picea-abies-l-karst-.jpg)








