
Compensatory thio-redox interactions between DsbA, CcdA and CcmG unveil the apocytochrome c holdase...
15
Compensatory thio–redox interactions between DsbA, CcdA and CcmG unveil the apocytochrome c holdase role of CcmG during cytochrome c maturation Serdar Turkarslan, † Carsten Sanders, † Seda Ekici and Fevzi Daldal* Department of Biology, Plant Science Institute, University of Pennsylvania, Philadelphia, PA 19104, USA. Summary During cytochrome c maturation (Ccm), the DsbA- dependent thio-oxidative protein-folding pathway is thought to introduce a disulphide bond into the haem- binding motif of apocytochromes c. This disulphide bond is believed to be reduced through a thio- reductive pathway involving the Ccm components CcdA (DsbD), CcmG and CcmH. Here, we show in Rhodobacter capsulatus that in the absence of DsbA cytochrome c levels were decreased and CcdA or CcmG or the putative glutathione transporter CydDC was not needed for Ccm. This decrease was not due to overproduction of the periplasmic protease DegP as a secondary effect of DsbA absence. In contrast, CcmH was absolutely necessary regardless of DsbA, indicating that compensatory thio–redox interactions excluded it. Remarkably, the double (DsbA–CcmG) and triple (DsbA–CcmG–CcdA) mutants produced cytochromes c at lower levels than the DsbA-null mutants, unless they contained a CcmG derivative (CcmG*) lacking its thio-reductive activity. Purified CcmG* can bind apocytochrome c in vitro, revealing for the first time a thiol-independent, direct interac- tion between apocytochrome c and CcmG. Further- more, elimination of the thio–redox components does not abolish cytochrome c production, restricting the number of Ccm components essential for haem– apocyt c ligation per se during Ccm. Introduction The cytochromes (cyts) c are membrane-attached or soluble ubiquitous electron carrier proteins essential for energy metabolisms of almost all organisms. They contain Fe-protoporphyrin IX (haem), which is stereo- specifically and covalently attached through two thioet- her bonds between the haem vinyl groups and the cysteine sulphydryls of the apocyt c C 1XXC2H motif. The covalent haem–apocyt c ligation is carried out by a post- translation and post-export process called cyt c matura- tion (Ccm), which involves in most Gram-negative bacteria 10 essential components [CcmABCDEFGHI and CcdA (or DsbD)] (Barker and Ferguson, 1999; Thony-Meyer, 2000). During Ccm, it is thought that the cysteine thiols of apocyt c C1XXC2H motif are first oxi- dized by the DsbA-dependent extracytoplasmic thio- oxidative protein-folding pathway (Fig. 1) (Kranz et al., 1998; Thony-Meyer, 2002; Allen et al., 2003a). Subse- quent reduction of this disulphide bond then becomes a prerequisite for haem ligation (Daltrop et al., 2002). The reduction is accomplished by a thio-reductive pathway consisting of CcdA, CcmG and CcmH (Monika et al., 1997). In Rhodobacter capsulatus, CcdA transfers elec- trons from cytoplasmic thioredoxins to CcmG (Katzen and Beckwith, 2000; Katzen et al., 2002). CcmG is believed to be re-oxidized by shuttling its electrons to CcmH, which then transfers them to apocyts c (Fig. 1) (Monika et al., 1997; Fabianek et al., 1999). Formation of a disulphide bond in the apocyt c C 1XXC2H motif occurs in vitro, and is considered to be an intermediate of Ccm (Daltrop et al., 2002). However, whether this is always the case in vivo is unclear. Ini- tially, DsbA and DsbB mutants in Escherichia coli were identified as unable to produce cyts c (Metheringham et al., 1996). Recent works in E. coli (Allen et al. 2003b; Kojima et al., 2005) and other bacteria (Erlendsson and Hederstedt, 2002; Deshmukh et al., 2003) indicated that DsbA- and DsbB-null mutants are Ccm-proficient. Fur- thermore, the lack of DsbA or DsbB functions sup- presses the cyt c deficiency of CcdA-null mutants in Bacillus subtilis or R. capsulatus (Erlendsson and Hed- erstedt, 2002; Deshmukh et al., 2003), demonstrating that the thio-reductive branch becomes dispensable in the absence of the thio-oxidative pathway. Therefore, why a thio–redox loop (i.e. thio-oxidation followed by thio-reduction) occurs at the apocyt c haem binding Accepted 25 August, 2008. *For correspondence. E-mail fdaldal@ sas.upenn.edu; Tel. (+1) 215 898 4394; Fax (+1) 215 898 8780. † Both authors contributed equally to this work. Molecular Microbiology (2008) 70(3), 652–666 doi:10.1111/j.1365-2958.2008.06441.x First published online 25 September 2008 © 2008 The Authors Journal compilation © 2008 Blackwell Publishing Ltd
-
Upload
systemsbiology -
Category
Documents
-
view
0 -
download
0
Transcript of Compensatory thio-redox interactions between DsbA, CcdA and CcmG unveil the apocytochrome c holdase...

Compensatory thio–redox interactions between DsbA, CcdAand CcmG unveil the apocytochrome c holdase role of CcmGduring cytochrome c maturation
Serdar Turkarslan,† Carsten Sanders,† Seda Ekiciand Fevzi Daldal*Department of Biology, Plant Science Institute,University of Pennsylvania, Philadelphia, PA 19104,USA.
Summary
During cytochrome c maturation (Ccm), the DsbA-dependent thio-oxidative protein-folding pathway isthought to introduce a disulphide bond into the haem-binding motif of apocytochromes c. This disulphidebond is believed to be reduced through a thio-reductive pathway involving the Ccm componentsCcdA (DsbD), CcmG and CcmH. Here, we show inRhodobacter capsulatus that in the absence of DsbAcytochrome c levels were decreased and CcdA orCcmG or the putative glutathione transporter CydDCwas not needed for Ccm. This decrease was not dueto overproduction of the periplasmic protease DegPas a secondary effect of DsbA absence. In contrast,CcmH was absolutely necessary regardless of DsbA,indicating that compensatory thio–redox interactionsexcluded it. Remarkably, the double (DsbA–CcmG)and triple (DsbA–CcmG–CcdA) mutants producedcytochromes c at lower levels than the DsbA-nullmutants, unless they contained a CcmG derivative(CcmG*) lacking its thio-reductive activity. PurifiedCcmG* can bind apocytochrome c in vitro, revealingfor the first time a thiol-independent, direct interac-tion between apocytochrome c and CcmG. Further-more, elimination of the thio–redox components doesnot abolish cytochrome c production, restricting thenumber of Ccm components essential for haem–apocyt c ligation per se during Ccm.
Introduction
The cytochromes (cyts) c are membrane-attached orsoluble ubiquitous electron carrier proteins essential for
energy metabolisms of almost all organisms. Theycontain Fe-protoporphyrin IX (haem), which is stereo-specifically and covalently attached through two thioet-her bonds between the haem vinyl groups and thecysteine sulphydryls of the apocyt c C1XXC2H motif. Thecovalent haem–apocyt c ligation is carried out by a post-translation and post-export process called cyt c matura-tion (Ccm), which involves in most Gram-negativebacteria 10 essential components [CcmABCDEFGHIand CcdA (or DsbD)] (Barker and Ferguson, 1999;Thony-Meyer, 2000). During Ccm, it is thought that thecysteine thiols of apocyt c C1XXC2H motif are first oxi-dized by the DsbA-dependent extracytoplasmic thio-oxidative protein-folding pathway (Fig. 1) (Kranz et al.,1998; Thony-Meyer, 2002; Allen et al., 2003a). Subse-quent reduction of this disulphide bond then becomes aprerequisite for haem ligation (Daltrop et al., 2002). Thereduction is accomplished by a thio-reductive pathwayconsisting of CcdA, CcmG and CcmH (Monika et al.,1997). In Rhodobacter capsulatus, CcdA transfers elec-trons from cytoplasmic thioredoxins to CcmG (Katzenand Beckwith, 2000; Katzen et al., 2002). CcmG isbelieved to be re-oxidized by shuttling its electrons toCcmH, which then transfers them to apocyts c (Fig. 1)(Monika et al., 1997; Fabianek et al., 1999).
Formation of a disulphide bond in the apocyt cC1XXC2H motif occurs in vitro, and is considered to bean intermediate of Ccm (Daltrop et al., 2002). However,whether this is always the case in vivo is unclear. Ini-tially, DsbA and DsbB mutants in Escherichia coli wereidentified as unable to produce cyts c (Metheringhamet al., 1996). Recent works in E. coli (Allen et al. 2003b;Kojima et al., 2005) and other bacteria (Erlendsson andHederstedt, 2002; Deshmukh et al., 2003) indicated thatDsbA- and DsbB-null mutants are Ccm-proficient. Fur-thermore, the lack of DsbA or DsbB functions sup-presses the cyt c deficiency of CcdA-null mutants inBacillus subtilis or R. capsulatus (Erlendsson and Hed-erstedt, 2002; Deshmukh et al., 2003), demonstratingthat the thio-reductive branch becomes dispensable inthe absence of the thio-oxidative pathway. Therefore,why a thio–redox loop (i.e. thio-oxidation followed bythio-reduction) occurs at the apocyt c haem binding
Accepted 25 August, 2008. *For correspondence. E-mail [email protected]; Tel. (+1) 215 898 4394; Fax (+1) 215 898 8780.†Both authors contributed equally to this work.
Molecular Microbiology (2008) 70(3), 652–666 � doi:10.1111/j.1365-2958.2008.06441.xFirst published online 25 September 2008
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd

motifs during Ccm remains unclear (Fig. 1). Here, weexamined cyt c production without such a thio–redoxloop in R. capsulatus. We found that mutants lackingDsbA produced less cyts c, but this decrease was notdue to the overproduction of periplasmic protease DegPthat occurs in the absence of DsbA in R. capsulatus(Onder et al., 2008). Moreover, when the DsbA-dependent thio-oxidative pathway was compromised,only CcmH, but neither CcdA nor CcmG, was requiredfor haem–apocyt c ligation. The compensatory thio–redox interactions were also independent of the glu-tathione transporter CydDC (Pittman et al., 2005), unlikein E. coli (Poole et al., 1994). Remarkably though, theDsbA–CcmG double and DsbA–CcmG–CcdA triplemutants produced consistently lower levels of cyt c thanthe DsbA-null mutants, unless they contained a CcmGderivative devoid of its thio-reductive activity. This findingrevealed a second role of CcmG acting as an “apocyt choldase” during Ccm in addition to its thio-reductivefunction. Our findings show that the thio–redox pathwayis important, but not essential, for cyt c production, andreduce the complex Ccm apparatus to those that areabsolutely required for haem–apocyt c ligation per se inR. capsulatus.
Results
R. capsulatus mutants lacking DsbA produce loweramounts of cyts c
Rhodobacter capsulatus mutants lacking DsbA are profi-cient for Ccm, unlike those lacking CcdA (Deshmukh et al.,2000). To further assess the role of DsbA during Ccm, wecompared the total amounts of periplasmic soluble andmembrane-associated cyts c produced in a wild-typestrain of R. capsulatus with an isogenic DsbA-null mutant(Fig. 1). Qualitative SDS-PAGE/TMBZ analyses showedquasi-wild type-like cyts c profiles in DsbA-null mutants(Fig. 2A) (Deshmukh et al., 2003). Although this stainingtechnique is very useful to monitor the overall cyts cprofiles, the band intensities are time-dependent and maynot be directly proportional to cyts c amounts. Thus, weused optical redox difference spectroscopy to obtainmore quantitative estimations. Ascorbate-reduced minusferricyanide-oxidized spectra recorded between 500 and600 nm using membranes and membrane supernatantsrevealed that supernatant fractions of a DsbA-null mutantcontained twice less (~40% per 0.5 mg per ml of totalproteins) 550 nm absorbing materials relative to a wild-type strain (Fig. 2B). In R. capsulatus, this absorbance
Fig. 1. Production of cyts c in the presence of the Ccm thio–redox loop. In a wild-type background, the cysteine thiols of the C1XXC2H motifof reduced apocyts c, secreted through the general secretion pathway (Sec), are oxidized via the DsbA- and DsbB-dependent thio-oxidationpathway. Oxidized apocyts c are then reduced by the CcdA- and CcmG-dependent thio-reduction pathway, and conveyed to the CcmFHIhaem ligation components that use haem from the haem delivery pathway (double arrows) to yield mature cyts c. Thick arrows show the routefollowed by apocyts c, and Cys, His, N and C refer to cysteine, histidine, N-terminal and C-terminal of apocyts c, respectively, and WWDrefers to the tryptophan-rich motif of CcmF component of Ccm.
Ccm in the absence of periplasmic thio–redox pathways 653
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology, 70, 652–666
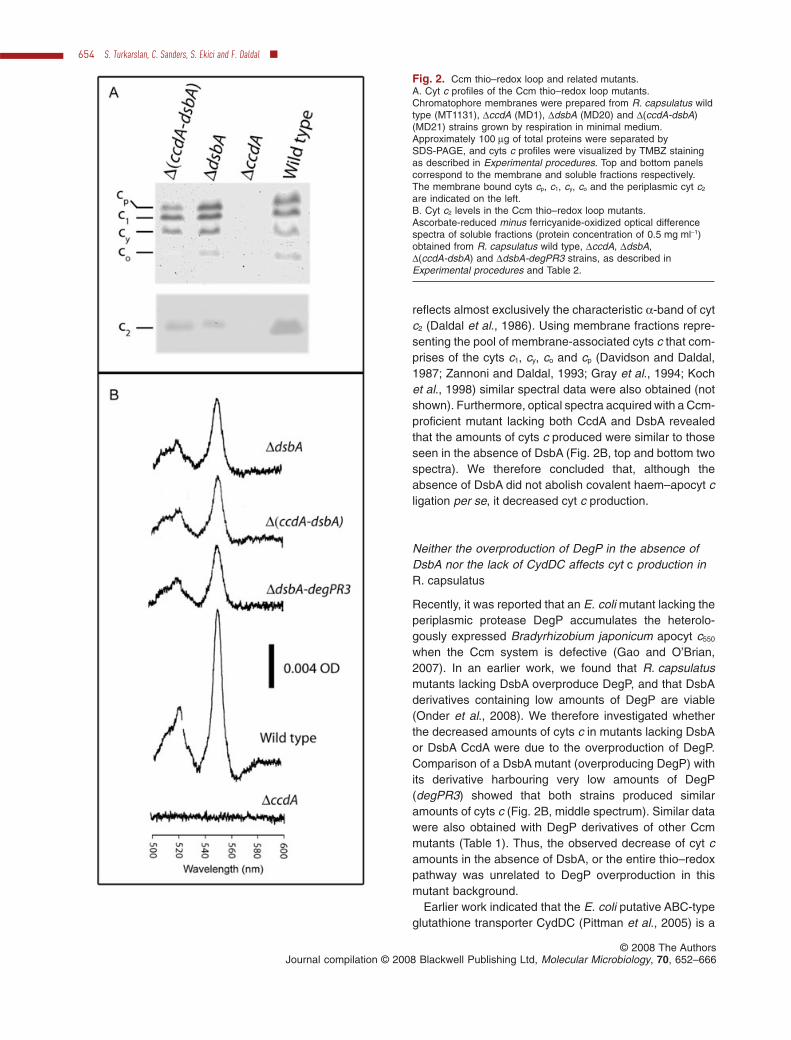
reflects almost exclusively the characteristic a-band of cytc2 (Daldal et al., 1986). Using membrane fractions repre-senting the pool of membrane-associated cyts c that com-prises of the cyts c1, cy, co and cp (Davidson and Daldal,1987; Zannoni and Daldal, 1993; Gray et al., 1994; Kochet al., 1998) similar spectral data were also obtained (notshown). Furthermore, optical spectra acquired with a Ccm-proficient mutant lacking both CcdA and DsbA revealedthat the amounts of cyts c produced were similar to thoseseen in the absence of DsbA (Fig. 2B, top and bottom twospectra). We therefore concluded that, although theabsence of DsbA did not abolish covalent haem–apocyt cligation per se, it decreased cyt c production.
Neither the overproduction of DegP in the absence ofDsbA nor the lack of CydDC affects cyt c production inR. capsulatus
Recently, it was reported that an E. coli mutant lacking theperiplasmic protease DegP accumulates the heterolo-gously expressed Bradyrhizobium japonicum apocyt c550
when the Ccm system is defective (Gao and O’Brian,2007). In an earlier work, we found that R. capsulatusmutants lacking DsbA overproduce DegP, and that DsbAderivatives containing low amounts of DegP are viable(Onder et al., 2008). We therefore investigated whetherthe decreased amounts of cyts c in mutants lacking DsbAor DsbA CcdA were due to the overproduction of DegP.Comparison of a DsbA mutant (overproducing DegP) withits derivative harbouring very low amounts of DegP(degPR3) showed that both strains produced similaramounts of cyts c (Fig. 2B, middle spectrum). Similar datawere also obtained with DegP derivatives of other Ccmmutants (Table 1). Thus, the observed decrease of cyt camounts in the absence of DsbA, or the entire thio–redoxpathway was unrelated to DegP overproduction in thismutant background.
Earlier work indicated that the E. coli putative ABC-typeglutathione transporter CydDC (Pittman et al., 2005) is a
Fig. 2. Ccm thio–redox loop and related mutants.A. Cyt c profiles of the Ccm thio–redox loop mutants.Chromatophore membranes were prepared from R. capsulatus wildtype (MT1131), DccdA (MD1), DdsbA (MD20) and D(ccdA-dsbA)(MD21) strains grown by respiration in minimal medium.Approximately 100 mg of total proteins were separated bySDS-PAGE, and cyts c profiles were visualized by TMBZ stainingas described in Experimental procedures. Top and bottom panelscorrespond to the membrane and soluble fractions respectively.The membrane bound cyts cp, c1, cy, co and the periplasmic cyt c2
are indicated on the left.B. Cyt c2 levels in the Ccm thio–redox loop mutants.Ascorbate-reduced minus ferricyanide-oxidized optical differencespectra of soluble fractions (protein concentration of 0.5 mg ml-1)obtained from R. capsulatus wild type, DccdA, DdsbA,D(ccdA-dsbA) and DdsbA-degPR3 strains, as described inExperimental procedures and Table 2.
654 S. Turkarslan, C. Sanders, S. Ekici and F. Daldal �
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology, 70, 652–666

component required for the bd-type hydroquinone oxidaseassembly and for cyt c production (Georgiou et al., 1987;Poole et al., 1994). We found that R. capsulatus mutantslacking CydC or CydD, or their derivatives lacking DsbA orDsbA CcdA (Table 2) also did not contain any active hyd-roquinone oxidase (K. Zhang and F. Daldal, unpubl. data).However, all CydDC derivatives still produced cyts c, unliketheir E. coli counterparts (Fig. 3A). Thus, in R. capsulatus,the putative ABC-type gluthatione transporter CydDC wasnot involved in cyt c production, and the compensatorythio–redox interactions seen between DsbA and CcdA didnot rely on the periplasmic reductive power that originatefrom the CydDC transporter.
Compensatory thio–redox interactions during Ccm isnot restricted to CcdA and DsbA
CcmG and CcmH are also thought to be parts of the Ccmthio-reductive branch (Deshmukh et al., 2003). Therefore,we probed whether the compensatory thio–redox interac-tions seen between DsbA and CcdA also extended toCcmG and CcmH. Like CcdA-null mutants, CcmG-null orCcmH-null mutants cannot grow under photosyntheticconditions (Ps-) due to the absence of cyts c (Beckman
et al., 1992). However, unlike the CcdA-null mutants thatrevert readily to Ps+ by DsbA inactivation (Deshmukhet al., 2003), CcmG-null mutants required supplementa-tion of the growth medium with thiol-reactive reagents(0.33 mM L-cysteine and 0.165 mM L-cystine) in the caseof MD11, and without supplement in the case of anSB1003 derivative on enriched medium, to obtain Ps+
revertants. In contrast, CcmH-null mutants never yieldedany Ps+ revertants under the conditions tested. Analysesof several Ps+ revertants of CcmG-null mutants indicatedthat they all produced cyts c (e.g. ccmG-Rev2 and -Rev3in Fig. 3B), and that their Ps+ growth was suppressedupon introduction of a wild-type allele of dsbA in trans.DNA sequence analyses revealed that the revertantscarried chromosomal mutations inactivating dsbA. Thesemutations included a deletion encompassing the basepairs 55–68 in MD11R2, 5 and 7, a G insertion at position135 in MD11R1, 3, 4, 6, 8, and DhelXR5, 2, an A insertionat position 363 in DhelXR6 and 7, and a T to G substitutionat position 629 in DhelXR1, 3, 4 and 8 (Fig. 4 and Table 2).
Indeed, construction of CcmG-null, DsbA-null, andCcmH-null DsbA-null double mutants confirmed that theloss of DsbA suppressed the Ccm deficiency of CcmG-null, but not that of CcmH-null, mutants (Table 2). The
Table 1. Ps growth properties and cyt c2 contents of R. capsulatus thio–redox mutants.
Pathway Strain PlasmidMedAa
PsbMedA*Ps
MPYEPs
MPYE*Ps
Cyt c2
(%)
Wild type + + + + 100Thio-oxidation mutants DdsbA + + + + ~44
DdsbA-degPR3 + + + + ~40
Thio-reduction mutants DccdA or DccmG or DccmH - - - - 0
Thio–redox loop mutants D(ccdA-dsbA) + + + + ~50D(ccmG-dsbA) - + - - ~10–15D(ccmH-dsbA) - - - - 0DccmG-Rev2 - + - - ~10–15D(ccmG-dsbA) degPR3 - + - - ~10D(ccdA-ccmG-dsbA) - + - - ~10–15D(ccdA-ccmH-dsbA) - - - - 0DccmG pCcmGWT + + + + 100D(ccmG-dsbA) pCcmGWT + + - + ~65DccmG pCcmGC75S or
pCcmGC78S orpCcmGC75S/78S
- - - - 0
D(ccmG-dsbA) pCcmGC75S s + - + ~40–50pCcmGC78S - + - + ~40–50pCcmGC75S/C78S + + - + ~48
D(ccmG-dsbA)-degPR3 pCcmGWT + + + + ~70pCcmGC75S/C78S + + + + ~41
DccmH pCcmHWT + + + + 100pCcmHC42S/C45S - - - - 0
D(ccmH-dsbA) pCcmHWT + + + + ~90pCcmHC42S/C45S - - - - 0
a. MedA and MPYE refer to minimal and enriched growth medium, respectively, and the asterisk (*) indicates that they were supplemented with0.33 mM cysteine and 0.165 mM cystine.b. Ps designate photosynthetic growth conditions, and +, s and - refer to normal, slow and no growth respectively. All strains are Res+ in bothMedA* and MPYE*, but those lacking DsbA often exhibit variable degrees of Res or Ps growth defects (slow to no growth) in the absence of thethio-reactive supplements.
Ccm in the absence of periplasmic thio–redox pathways 655
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology, 70, 652–666

Table 2. Bacterial strains and plasmids used in this study.
Strain or plasmid Description Relevant phenotype Source or reference
StrainsE. coli
HB101 F- D(gpt-proA)62 araC14 leuB6(Am) supE44 galK2(Oc)lacY1 D(mcrC-mrr) rpsL20(Strr) xylA5 mtl-1 thi-1
Sambrook and Russel (2001)
XL1-Blue F′::Tn10 proA +B + lacIq D(lacZ)M15/recA1 endA1gyrA96 (Nalr) thi hsdR17 (rK
– mK+) supE44 relA1 lac
Stratagene
R. capsulatusMT1131a crtD121 Rifr Wild type Scolnik et al. (1980)
Res+, Ps+, cyts c +
MD1 D(ccdA::spe) Res+/Ps-, cyts c - Deshmukh et al. (2000)MD14 ccmH::spe ccmF + Res+/Ps-, cyts c - Deshmukh et al. (2002)MD20 D(dsbA::kan) Res+/Ps+ on MedA,
Res–/Ps+ on MPYE, cyts c +Deshmukh et al. (2003)
MD20R3 D(dsbA::kan) degPR3 Res+/Ps+, cyts c +, low DegP Onder et al. (2008)MD21 D(ccdA::spe) D(dsbA::kan) Res+/Ps+ on MedA,
Res–/Ps+ on MPYE, cyts c +Deshmukh et al. (2003)
MD11 D(ccmG::kan) Res+/Ps-, cyts c - Sanders et al. (2007)SB1003-DhelX D(ccmG::kan) Res+/Ps-, cyts c - Beckman and Kranz (1993)ST21 D(ccmG::kan) D(dsbA::spe) Res+/Ps– on MPYE and
MPYE*, Ress/Ps– on MedA,Res+/Ps+ on MedA*
This work
ST38 D(dsbA::spe) degPR3 Res+/Ps+, cyts c +, low DegP This workST40 D(ccmG::kan) D(dsbA::spe) degPR3 Res+/Ps- on MPYE and
MPYE*, Res+/Ps– on MedA,Res+/Ps+ on MedA*, lowDegP
This work
ST34 D(ccmG::kan) D(dsbA::spe) D(ccdA::gen) Res+/Ps- on MPYE andMPYE*, Ress/Ps– on MedA,Res+/Ps+ on MedA*
This work
ST27 ccmH::spe D(dsbA::kan) Res+/Ps-, cyts c - This workST36 ccmH::spe D(dsbA::kan) D(ccdA::gen) Res+/Ps-, cyts c - This workMD11Ri or
SB1003-DhelXRi
(i = 1–10)
D(ccmG::kan)-Revi (i.e. dsbAi) Res+/Ps- on MPYE andRes+/Ps+ on MedA*
This workMPYE*, Ress/Ps– on MedA
STD1 D(cydD::gen) Res+/Ps+, cyts c + This workSTD3 D(cydD::gen) D(dsbA::kan) Res+/Ps+, cyts c + This workSTD5 D(cydD::gen) D(dsbA::kan) D(ccdA::spe) Res+/Ps+, cyts c + This work
PlasmidspBluescript pBluescript II KS(+) (pBSK) Ampr StratagenepRK2013 Helper Kanr Ditta et al. (1985)pRK415 Broad host-range vector with E. coli lacZ promoter Tetr Keen et al. (1988)pCHB500 Broad host-range vector with R. capsulatus cycA
promoterTetr Benning and Somerville (1992)
pHP45W-Spc W-spectinomycin (spe) cassette Sper Prentki and Krisch (1984)pBS-DsbAwt Phosphorylated 700 bp PCR product with dsbA cloned
into the SmaI site of pBSKAmpr Deshmukh et al. (2003)
pTC4-1K 4.2 kb insert carrying dsbA interrupted by kanamycincasette in pRK415
Tetr, Kanr Deshmukh et al. (2003)
pK6 ccdA with engineered BglII site at position 749 inpBluescript
Ampr Deshmukh et al. (2000)
pCS1540 570 bp PCR product containing ccmG cloned intoXbaI–KpnI site of pCHB500
Tetr Sanders et al. (2005a)
pCS1545 Cysteine-less ccmG-C75S/C78S allele of ccmG inpCS1540
Tetr Sanders et al. (2005a)
pYZ1 2.82 kb fragment carrying ccmFH operon expressedfrom G488A up-promoter mutation in pBluescript
Ampr Deshmukh et al. (2002)
pHX1 pRK415 with a DNA fragment containing theD(ccmG::kan) [previously called D(helX::kan)]
Tetr, Kanr Katzen et al. (2002)
pQE60 Expression vector (Qiagen) Ampr, PT5 QiagenpQE60-helX R. capsulatus ccmG without its signal peptide cloned
at the NcoI and BamHI sites of pQE60 to yield aC-terminal His-tagged derivative of CcmG
Ampr, PT5 J.F. Collet (unpublished)
pCS905 pET-3a derivative (Novagen) with T7 promoter regionreplaced by a DNA fragment encoding lacI and thetac promoter region
Ampr, Ptac, LacI+ Sanders et al. (2001)
656 S. Turkarslan, C. Sanders, S. Ekici and F. Daldal �
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology, 70, 652–666

DsbA-null CcmG-null double mutants became Ps+ inminimal medium supplemented with thiol-reactivereagents and produced cyts c (Fig. 3B and C). In contrast,the DsbA-null CcmH-null double mutants remained Ps-,independent of thiol-reactive reagents addition to thegrowth medium, and produced no cyt c (Fig. 5A and B).Thus, the compensatory thio–redox interactions seenbetween DsbA and CcdA also occurred between DsbAand CcmG, but not between DsbA and CcmH. In agree-ment with these findings, the triple mutant lacking DsbACcdA CcmG (but not DsbA CcdA CcmH) (Table 2) wasPs+ in minimal medium supplemented with thiol-reactivereagents, and exhibited cyts c profiles similar to those
seen with the CcmG-null DsbA-null double mutant(Fig. 3B and C). We concluded that haem–apocyt c liga-tion occurred in the absence of the Ccm thio–redox loopencompassing DsbA, DsbB, CcdA and CcmG (Fig. 1).
An additional role for CcmG as an apocyt c holdaseduring Ccm
The observation that the CcmG-null DsbA-null doublemutants required thiol-reactive supplements to growunder Ps conditions, unlike the CcdA-null DsbA-nullmutants, suggested that absence of CcmG hamperedCcm efficiency more severely than CcdA. Examination of
Table 2. cont.
Strain or plasmid Description Relevant phenotype Source or reference
pCS1302 pCS905 derivative, Strep-tag II sequence (IBA) fusedto GFP, rendering GFP replaceable by cloning anygene of interest in frame into NdeI and BamHI sites
Ampr, Ptac, LacI+
Strep-GFP+Sanders et al. (2008)
pCS1726 R. capsulatus cycA derivative encoding the matureform of cyt c2 with an N-terminal Strep-tag clonedinto pCS1302 using NdeI and BamHI sites
Ampr, Ptac, LacI+
Strep-cyt c2+
Sanders et al. (2008)
pCS1757 R. capsulatus cycA derivative encoding the matureform of cyt c2 with an N-terminal Strep-tag and aC-terminal 6xHis-tag cloned into pCS1302 usingNdeI and BamHI sites
Ampr, Ptac, LacI+, Strep-cytc2-6xHis+
This work
pCS1554 Cysteine 75 and 78 of R. capsulatus ccmG inpQE60-helX mutated to serines
Ampr, PT5 This work
pST6 ccmH-Strep from pST8 cloned into the XbaI–KpnI sitesof pCHB500
Tetr This work
pST7 PCR-amplified 0.48 kb ccmH fragment from pYZ1cloned into the XbaI–KpnI sites of pBluescript
Ampr This work
pST8 PCR-amplified and 3′-end Strep-tagged 0.53 kb ccmHfragment from pST7 cloned into the XbaI–KpnI sitesof pBSK
Ampr This work
pST14 ccmH-C42S/C45S allele in pCHB500 Tetr This workpKZ66 A 4.8 kb PCR fragment containing cydDC cloned into
BamHI–EcoRI sites of pBSKAmpr This work
pKZ68 pKZ66 with Genr cartridge replacing the 1.2 kb PstIfragment in cydC
Ampr Genr This work
pKZ69 XbaI–KpnI fragment of pKZ68 containing D(cydC::gen)in pRK415
Tetr Genr This work
pKZ7 HindIII-digested and religated pKZ66 without its BbsIsite downstream from cydC
Ampr This work
pKZ71 pKZ7 with Genr cartridge replacing the 320 bp BbsIfragment in cydD
Ampr Genr This work
pKZ72 XbaI–KpnI fragment of pKZ71 containing D(cydD::gen)in pRK415
Tetr, Genr This work
pCHB::Gm pCHB500 carrying a Genr cassette (gen) driven by thecycA promoter
Tetr, Genr K. Zhang and F. Daldal
pBS-STA3 The Sper cartridge of pHP45W-Spe cloned into theBglII–SfiI sites of pBS-DsbAwt
Ampr, Sper This work
pSTA1 2.1 kb XbaI–KpnI fragment of pBS-STA3 cloned intothe same sites of pRK415
Tetr, Sper This work
pST1 Phosphorylated 1.2 kb PCR fragment of Genr cassettefrom pCHB::Gm cloned into pBluescript
Genr, Ampr This work
pST3 BamHI fragment of pST1 cloned into the BglII sites ofpK6
Genr, Ampr This work
pST4 D(ccdA::gen) from pST3 cloned into pRK415 Genr, Tetr This workpCS1543 ccmG-C75S allele of ccmG in pCS1540 Tetr This workpCS1544 ccmG-C78S allele of ccmG in pCS1540 Tetr This work
a. R. capsulatus MT1131 is a derivative of SB1003 and referred to as wild type with respect to its growth properties and cyts c profile.The asterisk (*) refers to minimal (MedA) and enriched (MPYE) growth medium supplemented with 0.33 mM cysteine and 0.165 mM cystine.
Ccm in the absence of periplasmic thio–redox pathways 657
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology, 70, 652–666

the optical redox difference spectra indicated that theCcmG-null DsbA-null double or CcdA-null CcmG-nullDsbA-null triple mutants indeed contained much less(~10–15% of wild type) (Fig. 3C) 550 nm absorbingmaterials than the CcdA-null DsbA-null double mutants(~40–50% of wild type) (Fig. 2B). As the only differencebetween these thio–redox loop mutants was the absenceof CcmG, we probed whether the decrease of cyt c pro-duction was linked to the absence of the CcmG proteinper se. CcmG has two active-site cysteines (C75 and C78in R. capsulatus) (Monika et al., 1997), which face theperiplasm and accept electrons from CcdA (Katzen andBeckwith, 2000; Katzen et al., 2002). Substitution of theseactive-site cysteines with serine yielded the CcmGC75S,CcmGC78S and CcmGC75S/C78S (the latter dubbed here asCcmG*) mutants. As expected, these mutants wereunable to complement a CcmG-null strain for Ps growthin any growth medium, regardless of thiol-reactivesupplement. They also did not produce any cyt c that canbe detected by SDS-PAGE/TMBZ (Fig. 6A and B, top)or optical redox difference spectra analyses (Fig. 7A)(Table 1). Immunoblot data indicated that all strains(except CcmGC78S in enriched medium) produced mutantforms of CcmG at wild type or higher amounts (Fig. 6Aand B, bottom). We also noted that the amounts of CcmGwere variable in various mutants grown under differentconditions, and this point deserves future studies.
Interestingly, the cysteine-less derivative of CcmGcomplemented even in the absence of thiol-reactivesupplements the DsbA-null CcmG-null mutant for Psgrowth in minimal medium. Moreover, it also producedmore cyts c as shown by both SDS-PAGE/TMBZ (Fig. 6Aand B) and optical redox difference spectra analyses(Fig. 7B) (Table 1). Furthermore, the DsbA-null CcmG-nullmutant carrying the CcmG* derivative was, like the DsbA-null or the DsbA-null CcdA-null mutants, Ps+ in minimalmedium regardless of any thiol-reactive supplement, andcontained the highest amounts of cyts c among the CcmGmutants (Fig. 6). Optical spectra again showed that theamounts of the 550 nm absorbing materials found in theDsbA-null CcmG-null mutants harbouring CcmG* were
Fig. 3. Cyt c profiles of R. capsulatus thio–redox mutants.A and B. The cyts c production in the putative glutathionetransporter mutant DcydD (STD1) and its derivatives D(cydD-dsbA)(STD3) and D(cydD-dsbA-ccdA) (STD5) (A), in the wild type(MT1131) and Ccm thio-reduction mutant DccmG (MD11), its Ps+
revertants (DccmG-Rev2) (MD11R2) and (DccmG-Rev3) (MD11R3)and its derivatives D(ccmG-dsbA) (ST21) and D(ccdA-ccmG-dsbA)(ST34) (B), grown by respiration in minimal medium weredetermined by SDS-PAGE/TMBZ as described in Fig. 2 andExperimental procedures. Different cyts c are indicated on the left.C. Cyt c2 levels in the Ccm thio-reduction mutant CcmG and itsderivatives. Ascorbate-reduced minus ferricyanide-oxidized opticaldifference spectra of soluble fractions (protein concentration of0.5 mg ml-1) obtained from R. capsulatus strains DccmG,D(ccmG-dsbA), D(ccmG-dsbA)degPR3 (ST40) andD(ccdA-ccmG-dsbA) (Table 2) were determined as in Fig. 2.
Fig. 4. Ps+ revertants of R. capsulatus CcmG-null mutants are located in DsbA. Ps+ revertants from a CcmG-null mutant (MD11Ri fromMT1131 or DhelXRi from SB1003-DhelX) (Table 2) were isolated as described in the text, and DNA sequence of the chromosomal dsbA locusof each mutant was determined after PCR amplification. The mutant names, positions and type of the mutations found are indicated on top ofthe dsbA gene, and asterisk (*) indicates the active-site cysteine residues of DsbA.
658 S. Turkarslan, C. Sanders, S. Ekici and F. Daldal �
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology, 70, 652–666

comparable (~40–50% of wild type) to those found inthe DsbA-null or DsbA-null CcdA-null mutants (Fig. 7B).Thus, the presence of CcmG* devoid of its thio-reductiveactivity increased drastically cyt c production in theabsence of DsbA, indicating that CcmG had an additional,thio-reduction-independent function referred to as apocytc holdase.
Similar studies were also conducted with CcmH andits cysteine-less derivatives. R. capsulatus CcmH hastwo functionally essential cysteine residues at positions42 and 45 facing the periplasm (Monika et al., 1997).CcmH mutants lacking them (CcmHC42S, CcmHC45S and
CcmHC42S/C45S, the latter dubbed CcmH*) did not comple-ment for Ps growth the CcmH-null or DsbA-null CcmH-null mutants, and produced no cyt c (Fig. 5A and B)(Table 1) (unlike in E. coli; see Robertson et al., 2008)although they had similar amounts of wild type or mutantCcmH proteins, as indicated by immunoblot analyses(Fig. 5A, bottom). Overall comparison of the dataobtained with CcmH and CcmG indicated that the twothio-reductive components behaved very differently inrespect to the DsbA- and CcdA-dependent thio–redoxloop, suggesting that their reductive roles during Ccmmight be different.
Fig. 5. Cyt c profiles and CcmH immunoblots of wild type and cysteine-less CcmH mutants.A. Cyts c profiles of R. capsulatus DccmH (MD14) and D(ccmH-dsbA) (ST27) strains harbouring empty plasmid (vector) or wild type(WT/pST6) or cysteine-less (C42S C45S/pST14) alleles of ccmH grown in minimal medium were determined by SDS-PAGE/TMBZ analyses(top) as in Fig. 2. The same preparations were also subjected to SDS-PAGE using 50 mg of total proteins per lane, and the amounts of CcmHwere detected by immunoblot analyses (bottom) using anti-CcmH polyclonal antibodies, as described in Experimental procedures.B. Cyt c2 levels in the CcmH-null mutant and its DsbA-null derivatives. Ascorbate-reduced minus ferricyanide-oxidized optical differencespectra of soluble fractions (protein concentration of 0.5 mg ml-1) obtained from R. capsulatus DccmH (left) and D(ccmH-dsbA) (right) mutantsharbouring empty plasmid (vector) or wild type (+ CcmHWT) or cysteine-less (+ CcmHC42S/C45S) alleles of ccmH were determined as described inFig. 2.
Fig. 6. Cyt c profiles and CcmG immunoblots of cysteine-less CcmG mutants. Cyts c profiles of R. capsulatus DccmG and D(ccmG-dsbA)strains harbouring wild type [pCcmGWT (WT)] or different cysteine-less [pCcmGC75S (C75S), pCcmGC78S (C78S) and pCcmGC75S/C78S (C75SC78S)] alleles of ccmG grown in either minimal (A) or enriched (B) medium were determined by SDS-PAGE/TMBZ analyses (top), asdescribed in Fig. 2. The same preparations were subjected to SDS-PAGE using 50 mg of total proteins per lane, and the amounts of CcmGwere detected by immunoblot analyses (bottom) using anti-CcmG polyclonal antibodies, as described in Experimental procedures.
Ccm in the absence of periplasmic thio–redox pathways 659
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology, 70, 652–666

R. capsulatus apocyt c2 physically interacts withCcmG* in vitro
The additional, thioredoxin-independent apocyt c holdasefunction of CcmG during Ccm was investigated in vitro byprobing the direct physical interactions between purifiedapocyt c and partially purified CcmG*. A C-terminally Hisepitope-tagged mature form of R. capsulatus CcmG* wasexpressed in E. coli, and partially purified by affinitychromatography. Similarly, a double (N-terminal His andC-terminal Strep) epitope-tagged derivative of R. capsula-tus apocyt c2 devoid of its signal sequence was expressedin E. coli cytoplasm, and tandem affinity purified usingHis-bind and then Strep-Tactin affinity columns, asdescribed in Experimental procedures. As the apocyt c2
was highly unstable, use of tandem affinity tags facilitated
the isolation of intact apocyt c2. Purified apocyt c2 andCcmG*-containing extracts (50 and 40 mg total proteinsrespectively) were incubated together for 2 h at 4°C, andapocyt c2 was re-isolated using Strep-Tactin affinity chro-matography. Column fractions were probed for the pres-ence of apocyt c2 using anti-Strep and anti-His (not shown)antibodies, as well as for CcmG using anti-CcmG antibod-ies (Fig. 8). A similar mock experiment using partially puri-fied CcmG* but no apocyt c2 was also conducted to showthat CcmG* itself was not retained by the Strep-Tactinaffinity column (Fig. 8A). However, when an incubationmixture containing both CcmG* and apocyt c2 were loadedonto the same column, and eluted with desthiobiotin(DTB), apparently CcmG* was retained by the column asboth apocyt c2 and CcmG* were co-eluted in the samefractions (mainly E3) (Fig. 8B and C). We therefore con-
Fig. 7. Cyt c2 levels in R. capsulatuscysteine-less CcmG mutants and itsDsbA-null derivatives. Ascorbate-reducedminus ferricyanide-oxidized optical differencespectra of soluble fractions (0.5 mg of totalproteins ml-1) prepared from R. capsulatusDccmG (A) and D(ccmG-dsbA) orD(ccmG-dsbA)degPR3 (B) strains harbouring[+ pCcmGWT (WT)] or different cysteine-less[+ pCcmGC75S (C75S), +pCcmGC78S (C78S)and +pCcmGC75S/C78S (C75S C78S)] allelesof ccmG, grown on enriched medium, weredetermined as described in Fig. 2. In (B),the spectra labelled degPR3 were from aCcmG-null DsbA-null mutant carrying theD(ccmG-dsbA)degPR3 mutations lowering thelevels of DegP in this background (Onderet al., 2008).
660 S. Turkarslan, C. Sanders, S. Ekici and F. Daldal �
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology, 70, 652–666

cluded that a CcmG* lacking its active-site cysteines canrecognize and bind apocyt c2 in vitro as an apocyt choldase.
Discussion
This work was initiated in order to assess whether thecompensatory thio–redox interactions previously seenbetween DsbA and CcdA mutants (Deshmukh et al.,2003) also extended to other thio-reductive Ccm com-ponents. By doing so we hoped to dissect apart the Ccmcomponents involved in preparing apocyt c as anadequate substrate from those catalysing the subsequenthaem–apocyt c ligation per se. Searches for spontaneousPs+ revertants of CcmG-null and CcmH-null strains, andconstruction of appropriate double and triple knock-out mutants, demonstrated for the first time that theDsbA-dependent compensatory thio–redox interactionsinvolved solely CcdA and CcmG, and not CcmH (or CcmI,data not shown). Earlier, it was observed that Ps+ rever-tants of CcmG-null mutants, unlike CcdA-null mutants,
could only be obtained if the growth medium was supple-mented with thiol-reactive chemicals (Monika et al., 1997).Similarly, we noted that CcmG DsbA and CcdA CcmGDsbA knock-out mutants were Ps+ only upon supplemen-tation with thiol-reactive reagents. The basis for this redoxconstraint is unclear. A possibility is that, in the absence ofCcmG (in contrast to that of CcdA), R. capsulatus cellsmight be metabolically more limited to reduce their growthenvironment to initiate anoxygenic photosynthesis.
Although the cysteine-less derivatives (Monika et al.,1997; Fabianek et al., 1999) of the unusual thioredoxin-like CcmH (Di Matteo et al., 2007) were indeed Ccm-deficient, the thio-reductive function of CcmH could not becompensated by a thio-oxidative defect. These findingslimited the thio–redox loop to DsbA, DsbB, CcdA andCcmG (Fig. 1) establishing that these components werenot essential for cyt c production in vivo. Moreover, theysimplified the number of Ccm components required forhaem–apocyt c ligation per se (Fig. 9). Importantly, thedata also suggested that both CcmG and CcmH mighthave additional function(s) distinct from their knownthio-reductive roles (Monika et al., 1997). With respect toCcmH, a possibility is that its oxidized form might interactwith CcmG-reduced apocyt c during its transfer to haemligation complex containing CcmF. Indeed, CcmH isknown to co-immunoprecipitate with CcmF (Ren et al.,2002), and its overproduction together with CcmFbypasses the Ccm deficiency of CcmI-null mutants (Desh-mukh et al., 2002; Sanders et al., 2005a). Furthermore,our recent data indicate that CcmH is a component of acomplex also containing CcmF and CcmI in R. capsulatusmembranes (Sanders et al., 2008).
Unlike other thioredoxins, CcmG is highly specific forCcm (Monika et al., 1997; Fabianek et al., 1998; Edelinget al., 2002) with an active site unusually acidic, proposedto provide its substrate specificity (Edeling et al., 2004).Our finding that the cysteine-less CcmG derivative CcmG*produced high amounts of cyts c and exhibited Ps growthregardless of the thio-reactive supplements indicated thatCcmG protein per se also had a thioredoxin-independentrole(s) referred to as apocyt c holdase during Ccm (Fig. 9).Earlier, the active-site cysteines of E. coli CcmG was foundto be important, but not essential, for Ccm in the presenceof reducing agents (Fabianek et al., 1998). In the light of thecompensatory thio–redox interactions seen between DsbAand CcmG, this finding could also be interpreted as coun-teracting the DsbA oxidative power, and not necessarily asan additional activity of CcmG. The significant increase ofcyts c production in the presence of CcmG* in the absenceof DsbA, as seen here, established unambiguously athioredoxin-independent role for the CcmG protein per se(Fig. 9). This additional function of CcmG was further sup-ported by the fact that purified apocyt c and CcmG* devoidof its cysteines directly interact with each other in vitro.
Fig. 8. Co-purification of CcmG* with apocyt c2. Purified apocytc2 (50 mg) and CcmG*-enriched extracts (40 mg) were incubatedin the presence of 20 mM Tris-HCl pH = 7.9, 50 mM NaCl, 50 mgof DTB and 10 mM imidazole for 2 h at 4°C and loaded onto aStrepTactin® Sepharose affinity chromatography column, washed,and eluted with 2.5 mM DTB as described in Experimentalprocedures. Aliquots (300 ml of 1 ml total) from various purificationsteps (W: column wash, and E1 to E6: DTB elution fractions 1–6)were precipitated with ice-cold Acetone/Methanol and analysedby SDS-PAGE and immunoblots. Antibodies against CcmG andStrep-tag are as indicated on the right, and molecular weightmarkers (in kDa) are as shown on the left of each panel. (A)illustrates that CcmG*, which does not contain a Strep-tag, is notretained by the StrepTactin® Sepharose column in the absenceof apocyt c2. (B) and (C) show that in the presence of apocyt c2,CcmG* is retained by the column and co-eluted with apocyt c2,documenting that the two proteins interact with each other underthe conditions used.
Ccm in the absence of periplasmic thio–redox pathways 661
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology, 70, 652–666

Growing evidence suggests that CcmG acts as a holdase,conveying apocyt c to the haem ligation core complex. Thecrystal structure of B. subtilis CcmG homologue ResArevealed a cavity, close to its active-site cysteines, specu-lated to form a binding surface for apocyt c (Crow et al.,2004; Colbert et al., 2006). Consistent with this proposal,we observed that overproduction of CcmG or apocyt c(Sanders et al., 2007) in addition to CcmF and CcmH alsoincreased cyt c production in R. capsulatus mutantslacking CcmI (Deshmukh et al., 2002; Sanders et al.,2005a).
It should be emphasized that DsbA is not essential forCcm, but its absence decreases cyt c production irrespec-tive of the thio-reductive pathway (i.e. CcdA and CcmG).Although this observation could rationalize why the Ccmthio–redox loop might be conserved evolutionarily, whyit occurs remains unknown. That the thio–redox loopreactions might protect some apocyts c intermediates(Sanders et al., 2005a), or improve the fidelity of stereo-specific haem ligation (Sanders et al., 2005b) has beensuggested. Unfolded apocyts c might be more vulnerableto degradation unless protected by chaperones such asCcmG* (depicted as dashed outline in Fig. 9). Formationof a disulphide bond at the apocyt c C1XXC2H motif,although requiring subsequent reduction by the thio-reductive pathway, might be beneficial by promotingpartial folding and decreasing degradation of apocyt c.
Consequently, in the absence of the thio–redox loop andCcmG*, cyt c production would be much lower than nativeamounts. Absence of the Ccm thio–redox loop might alsocompromise stereo-specific haem ligation (Sanders et al.,2005b), yielding non-native cyts c that are prone todegradation. Indeed the DsbA, CcdA DsbA, CcmG* DsbAor CcdA CcmG* DsbA mutants produced lower amountsof cyts c than a wild-type strain of R. capsulatus. Insummary, this study established that the Ccm thio–redoxloop components DsbA, CcdA and CcmG are not essen-tial for cyt c production, hence decreasing the number ofCcm components absolutely required for haem–apocytc ligation. Furthermore, it revealed a second, thio-reduction-independent role of CcmG, which is critical forcyt c production, and raised the possibility that the oxi-dized form of CcmH might be required for transferringCcmG-reduced apocyt c to a haem ligation complex con-taining CcmF and CcmI. Future studies would hopefullybetter define the interactions between apocyt c and theessential Ccm components during cyt c production inbacteria.
Experimental procedures
Bacterial strains, plasmids and growth conditions
The bacterial strains and plasmids used in this work aredescribed in Table 2. R. capsulatus strains were grown at
Fig. 9. Production of cyts c in the absence of the Ccm thio–redox loop. In the absence of the DsbA–DsbB, CcdA and CcmG thio–redox loopshown in Fig. 1, reduced apocyts c, although more prone to degradation, are matured at lower levels by the CcmFHI ligation components. Thepresence of a thio-reduction inactive CcmG derivative (CcmG* shown in dashed line) increases the amounts of mature cyts c, by acting as anapocyt c holdase improving delivery of the apocyts c to the CcmFHI haem ligation components. Thick arrows show the route followed by theapocyts c, dashed arrows indicate the interactions of apocyts c and haem ligation components via CcmG*, and Cys, His, N, C and WWD areas described in Fig. 1.
662 S. Turkarslan, C. Sanders, S. Ekici and F. Daldal �
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology, 70, 652–666

35°C on enriched (Daldal et al., 1986) or minimal (Sistrom,1960) media, supplemented with appropriate antibiotics (2.5,10, 10 and 1 mg ml-1 tetracycline, kanamycin, spectinomycinand gentamicin respectively), either chemoheterotrophically(Res growth) or photoheterotrophically (Ps growth) in anaero-bic jars with H2 + CO2 generating gas packs from BBL Micro-biology Systems (Cockeyville, MD). All growth media weresupplemented with 0.33 mM L-cysteine and 0.165 mML-cystine when appropriate. E. coli strains were grown onLuria–Bertani (LB) broth supplemented with appropriate anti-biotics (12.5, 50, 10, 100 and 12 mg ml-1 tetracycline, kana-mycin, spectinomycin, ampicilin and gentamicin respectively)as needed.
Molecular genetics techniques
Standard molecular genetic techniques were performed asdescribed (Sambrook and Russel, 2001). Plasmid pBS-DsbAwt (Deshmukh et al., 2003) carrying the R. capsulatuswild-type dsbA was digested with BglII and SfiI to replace its0.5 kb internal fragment with a 2.0 kb DNA polymeraseblunted BamHI spectinomycin resistance cassette frompHP45Wspec to yield pBS-STA3 harbouring D(dsbA::spe)(Table 2). The XbaI–KpnI fragment of pBS-STA3 was thencloned into the respective sites of pRK415 to yield pSTA1. A1.2 kb gentamicin resistance cassette under the control of thecyt c2 gene promoter (PcycA-gen) was first amplified frompCHB::Gm (Table 2) with M13-forward and M13-reverseprimers, phosphorylated using T4 polynucleotide kinase andligated into the EcoRV-digested pBluescript to yield pST1(Table 2). To obtain a D(ccdA::PcycA-gen) allele pST1 wasrestricted with BamHI and the 1.2 kb fragment harbouringPcycA-gen ligated into BglII restricted pK6, eliminating a0.3 kb internal fragment of ccdA. The 2.7 kb insert from theresulting plasmid pST3 was then transferred into the BamHIsite of pRK415 to yield pST4. Plasmids pCS1540 andpCS1545 containing wild-type ccmG and the ccmG* variantwith both cysteines (C75 and C78) to serine substitutions,respectively, were constructed as described (Sanders et al.,2005a) (Table 2). To obtain pCS1543 and pCS1544, the C75or C78 of ccmG in pCS1540 was substituted with serineusing the QuikChange® XL Site-Directed Mutagenesis Kitaccording to the manufacturer’s instructions (Stratagene, LaJolla, CA) and the mutagenic primers HelX-C75Ss (5′-CTTCTG GGC TTC CTG GAG CGC GCC CTG TCG-3′) andHelX-C75Sas (5′-ACC CGA CAG GGC GCG CTC CAG GAAGCC CAG-3′) for pCS1543, and HelX-C78Ss (5′-TCC TGGTGC GCG CCC AGT CGG GTC GAA CA-3′) and HelX-C78Sas (5′-GGA TGT TCG ACC CGA CTG GGC GCG CACCAG-3′) for pCS1544 (Table 2). PCR amplification of ccmHusing pYZ1 as a template and CcmH/XbaI-Fwd (5′-CTC TCTAGA GTT GCG GCG GAG TGA GCG ATG CTG AAA CG-3′)and CcmH/KpnI-Rev (5′-GAG GGT ACC CTA GTC CTT CAGCAG GTC TTT CAG CCG CGC CTC-3′) as primers gener-ated a 0.48 kb DNA fragment, which was digested with XbaIand KpnI and ligated into the respective sites of pBluescript IIKS(+) (pBSK) to yield pST7 (Table 2). Plasmid pST8 wasconstructed by engineering a Strep-tag sequence at the3′-end of ccmH via PCR amplification using pST7 as a tem-plate and the primers CcmH-StrepC/XbaI-Fwd (5′-CTC TCTAGA GTT GCG GCG GAG TGA GCG ATG CTG AAA CG-3′)
and CcmH-StrepC/KpnI-Rev (5′-GAG GGT ACC CTA TTTTTC GAA CTG CGG GTG GCT CCA AGC GCT GTC CTTCAG CAG GTC TTT CAG CCG CGC CTC-3′). The 0.51 kbDNA fragment thus generated was cut with XbaI and KpnIand ligated into the same sites of pBSK. Plasmid pST8 wasdigested with XbaI and KpnI and the fragment carrying ccmH-Strep was ligated into the respective sites of pCHB500 toyield pST6. The cysteine-less derivative of CcmH wasobtained by substituting C42 and C45 of ccmH (with anin-frame Strep-tag sequence at its 3′-end) with serine asdescribed above by using pST8 as a template and CcmH-C42S/C45Ss (5′-TCG CAG GTG CTG CGC AGC CCG GTCAGT CAG GGC GAG A-3′) and CcmH-C42S/C45Sas (5′-GAT ATT CTC GCC CTG ACT GAC CGG GCT GCG CAGCAC CTG C-3′) as mutagenic primers to yield pST11(Table 2). Plasmid pST11 was digested with XbaI and KpnI,and the fragment carrying mutant ccmH was ligated intothe respective sites of pCHB500 to yield pST14. A 4.8 kbPCR fragment encompassing cydDC genes was amplifiedwith primers CydDCF (5′-GCA CTT GCT CGT CGA ACAGGA TCC AG-3′) and CydDCR (5′-CCG GAA TTC GAAACA GAC GCG CGA CA-3′), digested with BamHI and EcoRI(introduced sites in the primers are underlined) and clonedinto respective sites of pBluescript to yield pKZ66. In order tocreate deletion–insertion allele of cydC, a 1.2 kb PstI frag-ment of pKZ66 was replaced with 1.2 kb gentamicin resis-tance cartridge from pCHB::Gm yielding pKZ68. XbaI–KpnIfragment from pKZ68 carrying cydD +DC::Gm allele wasinserted into the corresponding sites of pRK415 to yieldpKZ69. Plasmid pKZ66 was digested with HindIII and ligatedback, eliminating BbsI site immediately downstream of cydCto yield pKZ7. To create deletion–insertion allele of cydD, a320 bp BbsI fragment from pKZ7 was excised, and theremaining vector was made blunt and ligated to Gm cartridgefrom pCHB::Gm to yield pKZ71. XbaI–KpnI fragment ofpKZ71 was then cloned into XbaI–KpnI site of pRK415 tocreate pKZ72 (Table 2). Plasmid pQE60-helX was obtainedby cloning into pQE60 (Qiagen, Valencia, CA) R. capsulatusccmG lacking its N-terminal signal peptide/transmembranehelix by using the NcoI and BamHI sites. Plasmid pCS1554expressing the C-terminally His-tagged mature form ofCcmG* was obtained by mutating to serine the cysteines 75and 78 of ccmG in pQE60-helX using the primers HelX-C75S/C78Ss (5′-CTT CTG GGC TTC CTG GAG CGC GCCCAG TCG GGT CGA ACA-3′) and HelX-C75S/C78Sas(5′-GGA TGT TCG ACC CGA CTG GGC GCG CTC CAGGAA GCC CAG-3′) according to the instruction of theQuikChange® XL Site-Directed Mutagenesis Kit from Strat-agene (La Jolla, CA). The mature form of R. capsulatus cytc2 gene (cycA mat) was PCR-amplified using the primersCS24 (5′-GTT GAC AAT TAA TCA TCG GC-3′) andRccycACtHis6BamHIr (5′-GCC GGA TCC TAG TGA TGGTGA TGG TGA TGT TTC ACG ACC GAG GCC AGA TAGGC-3′) and pCS1726 (Sanders et al., 2008) as a template.The 0.5 kb fragment thus generated was digested with NdeIand BamHI, and cloned into the same sites of pCS1302(Sanders et al., 2008) to yield pCS1757 that carries cycA mat
with an in-frame Strep-tag sequence at its 5′-end and a6xHis-tag at its 3′-end. The expression of the double-taggedcycA mat derivative is controlled by a Ptac-lac promoter–operator system in E. coli.
Ccm in the absence of periplasmic thio–redox pathways 663
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology, 70, 652–666

Automated DNA sequencing with the Big-DyeTM terminatorcycle sequencing kit (AmpliTaq FS; Applied Biosystems) wasperformed according to the manufacturer by using the M13-forward and M13-reverse primers to verify all plasmids con-structed in this study. DNA sequence analyses, homologysearches and genome sequence comparisons were per-formed using MacVector (IBI; Kodak) and BLAST softwarepackages (Altschul et al., 1990).
Conjugal transfers of plasmids from E. coli to R. capsulatusand chromosomal allele replacement via interposonmutagenesis using the gene transfer agent (Yen et al., 1979)were carried out as described earlier (Daldal et al., 1986).Briefly, the mutant strain ST40 (ccmG::kan dsbA::spedegPR3) mutant was obtained using pHX-1 (ccmG::kan)as a donor and ST38 (dsbA::spe, degPR3), which is aSpecR derivative of MD20R3 (Onder et al., 2008) as a re-cipient strain. The CcmH-null DsbA-null double mutant ST27,and CcdA-null CcmH-null DsbA-null triple mutant ST36,were constructed using pTC4-1K (dsbA::kan) and pST4(ccdA::gen) as donors and MD14 (ccmH::spe) and ST27(ccmH::spe dsbA::kan) as recipient strains respectively.
Biochemical techniques
Intracytoplasmic membrane vesicles (chromatophore mem-branes) were prepared in 50 mM MOPS (pH 7.0) containing100 mM KCl for spectral, or 1 mM KCl for SDS-PAGE, analy-ses as described earlier (Gray et al., 1994). All preparationscontained 10 mM EDTA, 1 mM PMFS and 0.1 mM e-amino-caproic acid as protease inhibitors. Protein concentra-tions were determined according to Lowry et al. (1951) andSDS-PAGE was used as in Schagger and von Jagow (1987)using 16.5% polyacrylamide gels. Cyts c were revealed basedon the endogenous peroxidase activity of covalently attachedhaem by using 3,3′,5,5′-tetramethylbenzidine (TMBZ)(Thomas et al., 1976). For CcmG and CcmH immunodetec-tion, protein samples were separated using 15% SDS-PAGE(Laemmli, 1970). Following electrophoresis, gels were elec-troblotted onto Immobilon-P PVDF membranes (Millipore, Bil-lerica, MA). CcmG and CcmH were detected using anti-CcmGand anti-CcmH polyclonal antibodies (Monika et al., 1997),ECL anti-rabbit IgG/horseradish peroxidase-conjugated sec-ondary antibodies from Amersham-Pharmacia (Piscataway,NJ) and SuperSignal West Pico Chemiluminescent substratefrom Pierce (Rockford, IL).
Expression and purification of apocytochromec2 and CcmG*
Escherichia coli strains containing pCS1757 (for apocyt c2) orpCS1554 (for CcmG*) were grown in Luria Broth (LB) liquidmedium (4 l each) containing 100 mg ml-1 ampicillin at 37°Cwith shaking (200 r.p.m.) until an OD600 of approximately 0.8.Cultures were then induced under the same growth condi-tions for 2 h by adding isopropyl-thio-b-D-galactoside (IPTG)to a final concentration of 10 mM. Cells were harvested bycentrifugation for 10 min at 10 000 g and re-suspended inTris Buffer (50 mM Tris-HCl pH 8.0, 100 mM NaCl and 1 mMPMSF), and disrupted on ice by using a French pressure cellas described in Daldal et al. (1986). Cell lysates were centri-
fuged at 25 500 g for 1 h, followed by at 118 000 g for 1 h.Cleared supernatants were filtered through a 0.45 mm filterand applied to a His-bind purification chromatography column(His-bind Kit from Novagen) following the manufacturer’sinstructions. Final concentrations of 25 and 100 mM imida-zole in the washing and elution buffers, respectively, wereused, and column fractions monitored by SDS-PAGE andWestern blot analyses. His-bind chromatography fractioncontaining apocyt c2 was pooled, concentrated by ultrafiltra-tion (3000 MWCO, Amicon Ultra, Millipore) to eliminate imi-dazole, diluted 10-fold in Strep Wash buffer before loadingonto the StrepTactin® Sepharose affinity column, and purifiedaccording to manufacturer’s instruction (IBA). Direct interac-tions between purified apocyt c2 and CcmG* were probed bymixing 50 mg of His and Strep-purified apocyt c2 with 40 mg ofCcmG*-enriched extracts and incubating for 2 h at 4°C, in thepresence of 20 mM Tris-HCl pH = 7.9, 50 mM NaCl, 50 mg ofDTB and 10 mM imidazole (final concentrations). After incu-bation, the reaction mixture was applied to the Strep-Tactinaffinity chromatography column and purification was per-formed according to the manufacturer’s instructions (IBA).Column fractions were subjected to SDS-PAGE and Westernblot analyses, and apocyt c2 was detected with anti-His andanti-Strep antibodies whereas CcmG was identified withCcmG antisera.
Spectroscopic analysis
Optical absorbance redox difference spectra were recordedusing a Hitachi U-3210 UV/vis spectrophotometer between500 and 600 nm. Chromatophore membrane supernatantswere centrifuged at 120 000 g for 2 h to eliminate membranedebris. The supernatants thus obtained were adjusted with50 mM MOPS/100 mM KCl to a protein concentration of0.5 mg ml-1 and oxidized by adding a crystal of potassiumferricyanide, and subsequently reduced by either solidsodium ascorbate or sodium dithionite to measure the totalamounts of reduced cyts c.
Chemicals
All chemicals were of reagent grade and obtained from com-mercial sources.
Acknowledgements
This work was supported by grants from DOE 91ER20052and NIH GM 38237 to F.D. The authors thank Drs R.G. Kranzfor providing anti-CcmG and anti-CcmH antibodies, J.F.Collet for plasmid pQE60-helX, and K. Zhang and M. Desh-mukh for construction of various pKZ plasmids and MDstrains respectively.
References
Allen, J.W., Daltrop, O., Stevens, J.M., and Ferguson, S.J.(2003a) C-type cytochromes: diverse structures and bio-genesis systems pose evolutionary problems. Philos TransR Soc Lond B Biol Sci 358: 255–266.
664 S. Turkarslan, C. Sanders, S. Ekici and F. Daldal �
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology, 70, 652–666

Allen, J.W., Barker, P.D., and Ferguson, S.J. (2003b) A cyto-chrome b562 variant with a c-type cytochrome CXXCHheme-binding motif as a probe of the Escherichia coli cyto-chrome c maturation system. J Biol Chem 278: 52075–52083.
Altschul, S.F., Gish, W., Miller, W., Myers, E.W., and Lipman,D.J. (1990) Basic local alignment search tool. J Mol Biol215: 403–410.
Barker, P.D., and Ferguson, S.J. (1999) Still a puzzle: why ishaem covalently attached in c-type cytochromes? Struc-ture 7: 281–290.
Beckman, D.L., and Kranz, R.G. (1993) Cytochromes c bio-genesis in a photosynthetic bacterium requires a periplas-mic thioredoxin-like protein. Proc Natl Acad Sci USA 90:2179–2183.
Beckman, D.L., Trawick, D.R., and Kranz, R.G. (1992) Bac-terial cytochromes c biogenesis. Genes Dev 6: 268–283.
Benning, C., and Somerville, C.R. (1992) Isolation andgenetic complementation of a sulfolipid-deficient mutant ofRhodobacter sphaeroides. J Bacteriol 174: 2352–2360.
Colbert, C.L., Wu, Q., Erbel, P.J., Gardner, K.H., and Deisen-hofer, J. (2006) Mechanism of substrate specificity in Bacil-lus subtilis ResA, a thioredoxin-like protein involved incytochrome c maturation. Proc Natl Acad Sci USA 103:4410–4415.
Crow, A., Acheson, R.M., Le Brun, N.E., and Oubrie, A.(2004) Structural basis of Redox-coupled protein substrateselection by the cytochrome c biosynthesis protein ResA.J Biol Chem 279: 23654–23660.
Daldal, F., Cheng, S., Applebaum, J., Davidson, E., andPrince, R.C. (1986) Cytochrome c(2) is not essential forphotosynthetic growth of Rhodopseudomonas capsulata.Proc Natl Acad Sci USA 83: 2012–2016.
Daltrop, O., Allen, J.W., Willis, A.C., and Ferguson, S.J.(2002) In vitro formation of a c-type cytochrome. Proc NatlAcad Sci USA 99: 7872–7876.
Davidson, E., and Daldal, F. (1987) Primary structure of thebc1 complex of Rhodopseudomonas capsulata. Nucleotidesequence of the pet operon encoding the Rieske cyto-chrome b, and cytochrome c1 apoproteins. J Mol Biol 195:13–24.
Deshmukh, M., Brasseur, G., and Daldal, F. (2000) NovelRhodobacter capsulatus genes required for the biogenesisof various c-type cytochromes. Mol Microbiol 35: 123–138.
Deshmukh, M., May, M., Zhang, Y., Gabbert, K.K., Karberg,K.A., Kranz, R.G., and Daldal, F. (2002) Overexpression ofccl1-2 can bypass the need for the putative apocytochromechaperone CycH during the biogenesis of c-typecytochromes. Mol Microbiol 46: 1069–1080.
Deshmukh, M., Turkarslan, S., Astor, D., Valkova-Valchanova, M., and Daldal, F. (2003) The dithiol:disulfideoxidoreductases DsbA and DsbB of Rhodobacter capsula-tus are not directly involved in cytochrome c biogenesis,but their inactivation restores the cytochrome c biogenesisdefect of CcdA-null mutants. J Bacteriol 185: 3361–3372.
Di Matteo, A., Gianni, S., Schinina, M.E., Giorgi, A., Altieri, F.,Calosci, N., et al. (2007) A strategic protein in cytochromec maturation: three-dimensional structure of CcmH andbinding to apocytochrome c. J Biol Chem 282: 27012–27019.
Ditta, G., Schmidhauser, T., Yakobson, E., Lu, P., Liang,
X.W., Finlay, D.R., et al. (1985) Plasmids related to thebroad host range vector, pRK290, useful for gene cloningand for monitoring gene expression. Plasmid 13: 149–153.
Edeling, M.A., Guddat, L.W., Fabianek, R.A., Thony-Meyer,L., and Martin, J.L. (2002) Structure of CcmG/DsbE at1.14 A resolution: high-fidelity reducing activity in an indis-criminately oxidizing environment. Structure 10: 973–979.
Edeling, M.A., Ahuja, U., Heras, B., Thony-Meyer, L., andMartin, J.L. (2004) The acidic nature of the CcmG redox-active center is important for cytochrome c maturation inEscherichia coli. J Bacteriol 186: 4030–4033.
Erlendsson, L.S., and Hederstedt, L. (2002) Mutations in thethiol-disulfide oxidoreductases BdbC and BdbD can sup-press cytochrome c deficiency of CcdA-defective Bacillussubtilis cells. J Bacteriol 184: 1423–1429.
Fabianek, R.A., Hennecke, H., and Thony-Meyer, L. (1998)The active-site cysteines of the periplasmic thioredoxin-likeprotein CcmG of Escherichia coli are important but notessential for cytochrome c maturation in vivo. J Bacteriol180: 1947–1950.
Fabianek, R.A., Hofer, T., and Thony-Meyer, L. (1999) Char-acterization of the Escherichia coli CcmH protein revealsnew insights into the redox pathway required for cyto-chrome c maturation. Arch Microbiol 171: 92–100.
Gao, T., and O’Brian, M.R. (2007) Control of DegP-dependent degradation of c-type cytochromes by hemeand the cytochrome c maturation system in Escherichiacoli. J Bacteriol 189: 6253–6259.
Georgiou, C.D., Fang, H., and Gennis, R.B. (1987) Identifi-cation of the cydC locus required for expression of thefunctional form of the cytochrome d terminal oxidasecomplex in Escherichia coli. J Bacteriol 169: 2107–2112.
Gray, K.A., Grooms, M., Myllykallio, H., Moomaw, C., Slaugh-ter, C., and Daldal, F. (1994) Rhodobacter capsulatus con-tains a novel cb-type cytochrome c oxidase without a CuAcenter. Biochemistry 33: 3120–3127.
Katzen, F., and Beckwith, J. (2000) Transmembrane electrontransfer by the membrane protein DsbD occurs via a dis-ulfide bond cascade. Cell 103: 769–779.
Katzen, F., Deshmukh, M., Daldal, F., and Beckwith, J.(2002) Evolutionary domain fusion expanded the substratespecificity of the transmembrane electron transporterDsbD. EMBO J 21: 3960–3969.
Keen, N.T., Tamaki, S., Kobayashi, D., and Trollinger, D.(1988) Improved broad-host-range plasmids for DNAcloning in gram-negative bacteria. Gene 70: 191–197.
Koch, H.G., Hwang, O., and Daldal, F. (1998) Isolationand characterization of Rhodobacter capsulatus mutantsaffected in cytochrome cbb3 oxidase activity. J Bacteriol180: 969–978.
Kojima, N., Yamanaka, M., Ichiki, S., and Sambongi, Y.(2005) Unexpected elevated production of Aquifex aeoli-cus cytochrome c555 in Escherichia coli cells lackingdisulfide oxidoreductases. Biosci Biotechnol Biochem 69:1418–1421.
Kranz, R., Lill, R., Goldman, B., Bonnard, G., and Merchant,S. (1998) Molecular mechanisms of cytochrome c biogen-esis: three distinct systems. Mol Microbiol 29: 383–396.
Laemmli, U.K. (1970) Cleavage of structural proteins duringthe assembly of the head of bacteriophage T4. Nature 227:680–685.
Ccm in the absence of periplasmic thio–redox pathways 665
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology, 70, 652–666

Lowry, O.H., Rosebrough, N.J., Farr, A.L., and Randall, R.J.(1951) Protein measurement with the Folin phenol reagent.J Biol Chem 193: 265–275.
Metheringham, R., Tyson, K.L., Crooke, H., Missiakas, D.,Raina, S., and Cole, J.A. (1996) Effects of mutations ingenes for proteins involved in disulphide bond formation inthe periplasm on the activities of anaerobically inducedelectron transfer chains in Escherichia coli K12. Mol GenGenet 253: 95–102.
Monika, E.M., Goldman, B.S., Beckman, D.L., and Kranz,R.G. (1997) A thioreduction pathway tethered to the mem-brane for periplasmic cytochromes c biogenesis; in vitroand in vivo studies. J Mol Biol 271: 679–692.
Onder, O., Turkarslan, S., Sun, D., and Daldal, F. (2008)Overproduction or absence of the periplasmic proteaseDegP severely compromises bacterial growth in theabsence of the dithiol: disulfide oxidoreductase DsbA. MolCell Proteomics 7: 875–890.
Pittman, M.S., Robinson, H.C., and Poole, R.K. (2005) Abacterial glutathione transporter (Escherichia coli CydDC)exports reductant to the periplasm. J Biol Chem 280:32254–32261.
Poole, R.K., Gibson, F., and Wu, G. (1994) The cydD geneproduct, component of a heterodimeric ABC transporter, isrequired for assembly of periplasmic cytochrome c and ofcytochrome bd in Escherichia coli. FEMS Microbiol Lett117: 217–223.
Prentki, P., and Krisch, H.M. (1984) In vitro insertionalmutagenesis with a selectable DNA fragment. Gene 29:303–313.
Ren, Q., Ahuja, U., and Thony-Meyer, L. (2002) A bacterialcytochrome c heme lyase. CcmF forms a complex with theheme chaperone CcmE and CcmH but not with apocyto-chrome c. J Biol Chem 277: 7657–7663.
Robertson, I.B., Stevens, J.M., and Ferguson, S.J. (2008)Dispensable residues in the active site of the cytochrome cbiogenesis protein CcmH. FEBS Lett 582: 3067–3072.
Sambrook, J., and Russel, D.W. (2001) Molecular Cloning: ALaboratory Manual. Cold Spring Harbor, NY: Cold SpringHarbor Laboratory Press.
Sanders, C., Wethkamp, N., and Lill, H. (2001) Transport ofcytochrome c derivatives by the bacterial Tat protein trans-location system. Mol Microbiol 41: 241–246.
Sanders, C., Deshmukh, M., Astor, D., Kranz, R.G., andDaldal, F. (2005a) Overproduction of CcmG and CcmFH(Rc) fully suppresses the c-type cytochrome biogenesis
defect of Rhodobacter capsulatus CcmI-null mutants.J Bacteriol 187: 4245–4256.
Sanders, C., Turkarslan, S., and Daldal, F. (2005b) Periplas-mic oxidative folding and cytochrome c maturation: amechanistic view of stereo-selective heme attachment. InPhotosynthesis: Fundamental Aspects to Global Perspec-tives. Van der Est, A., and Bruce, D. (eds.). Lawrence, KS:Allen Press, pp. 421–424.
Sanders, C., Boulay, C., and Daldal, F. (2007) Membrane-spanning and periplasmic segments of CcmI have distinctfunctions during cytochrome c biogenesis in Rhodobactercapsulatus. J Bacteriol 189: 789–800.
Sanders, C., Turkarslan, S., Lee, D.-W., Onder, O., Kranz,R.G., and Daldal, F. (2008) The cytochrome c maturationcomponents CcmF, CcmH and CcmI form a membrane-integral multisubunit heme ligation complex. J Biol Chem(in press). [Epub ahead of print], PMID 18753134.
Schagger, H., and von Jagow, G. (1987) Tricine-sodiumdodecyl sulfate-polyacrylamide gel electrophoresis for theseparation of proteins in the range from 1 to 100 kDa. AnalBiochem 166: 368–379.
Scolnik, P.A., Walker, M.A., and Marrs, B.L. (1980) Biosyn-thesis of carotenoids derived from neurosporene inRhodopseudomonas capsulata. J Biol Chem 255: 2427–2432.
Sistrom, W.R. (1960) A requirement for sodium in the growthof Rhodopseudomonas spheroides. J Gen Microbiol 22:778–785.
Thomas, P.E., Ryan, D., and Levin, W. (1976) An improvedstaining procedure for the detection of the peroxidaseactivity of cytochrome P-450 on sodium dodecyl sulfatepolyacrylamide gels. Anal Biochem 75: 168–176.
Thony-Meyer, L. (2000) Haem–polypeptide interactionsduring cytochrome c maturation. Biochim Biophys Acta1459: 316–324.
Thony-Meyer, L. (2002) Cytochrome c maturation: a complexpathway for a simple task? Biochem Soc Trans 30: 633–638.
Yen, H.C., Hu, N.T., and Marrs, B.L. (1979) Characterizationof the gene transfer agent made by an overproducermutant of Rhodopseudomonas capsulata. J Mol Biol 131:157–168.
Zannoni, D., and Daldal, F. (1993) The role of c-type cyto-chromes in catalyzing oxidative and photosynthetic elec-tron transport in the dual functional plasmamembrane offacultative phototrophs. Arch Microbiol 160: 413–423.
666 S. Turkarslan, C. Sanders, S. Ekici and F. Daldal �
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology, 70, 652–666




















