
Comparison of two laboratory cultures of Psyttalia concolor (Hymenoptera: Braconidae), as a...
8
Biological Control 39 (2006) 248–255 www.elsevier.com/locate/ybcon 1049-9644/$ - see front matter © 2006 Elsevier Inc. All rights reserved. doi:10.1016/j.biocontrol.2006.06.007 Comparison of two laboratory cultures of Psyttalia concolor (Hymenoptera: Braconidae), as a parasitoid of the olive fruit Xy Karen R. Sime a,¤ , Kent M. Daane a , Russell H. Messing b , Marshall W. Johnson c a Division of Organisms and Environment and Center for Biological Control, University of California, Berkeley, CA 94720-3114, USA b University of Hawaii, Kauai Agricultural Research Center, 7370 Kuamoo Road, Kapaa, Kauai, HI 96746, USA c Department of Entomology, University of California,Riverside, CA 92521, USA Received 2 March 2006; accepted 14 June 2006 Available online 27 June 2006 Abstract Life-history trials were conducted in the laboratory on two cultures of Psyttalia concolor (Szépligeti) (Hymenoptera: Braconidae) reared on olive fruit Xy, Bactrocera oleae (Rossi) (Diptera: Tephritidae). The tested cultures (A and B) had similar histories except that Culture A was maintained in Kenya for four years. Results showed that parasitoids from both cultures preferentially searched on olive fruit containing 8 to 10-day-old (second and third instar) olive Xies, and reproduced most successfully when third-instar Xy larvae were available. The mean longevity of adult female P. concolor was a negative function of temperature. Females from Culture B lived signiW- cantly longer at 15, 22, 25, and 32 °C than females from Culture A. For both cultures, adult female longevity was signiWcantly longer when the parasitoids were provided with honey than when provided water alone, or nothing; the presence of hosts signiWcantly reduced longev- ity, suggesting an energetic cost for reproduction. The parasitoids produced an average of 28.7 § 4.1 and 22.2 § 5.1 oVspring per female in Cultures A and B, respectively. The results are discussed with respect to use of biological control agents held under diVerent rearing con- ditions, and the potential of P. concolor for use as a biological control agent for olive fruit Xy in California. © 2006 Elsevier Inc. All rights reserved. Keywords: Psyttalia concolor; Braconidae; Bactrocera oleae; Tephritidae; Olea; Biological control; Parasitoid biology; Olive 1. Introduction The olive fruit Xy, Bactrocera oleae (Rossi) (Diptera: Tephritidae), has long been a pest of olives in the Mediter- ranean basin (Clausen, 1978; Tzanakakis, 2003; White and Elson-Harris, 1992). It was discovered in southern Califor- nia in 1998 and within four years had spread nearly throughout the state, posing a serious threat to the olive industry (Rice et al., 2003). Current research eVorts empha- size the development of sustainable management practices. Insecticidal baits are currently the principal means of con- trol, but are limited in their eVectiveness. Infested trees in suburban and rural landscaping act as reservoirs for reinva- sion (Collier and Van Steenwyk, 2003), and, furthermore, the successful biological controls that have been established for scale pests in California olives (Daane et al., 2005) might be disrupted by insecticides. As no natural enemies exist in California that can eVectively suppress olive Xy, a classical biological control program was initiated in 2002. EVorts to identify useful natural enemies have to date included foreign exploration in Africa and Central Asia, and evaluation of parasitoids used to control tephritid pests elsewhere (Hoelmer et al., 2004; Sime et al., 2006). Among the olive Xy parasitoids readily available in cul- ture is Psyttalia concolor (Szépligeti) (Hymenoptera: Bra- conidae). Recently, this species was released in California on a limited basis (Collier and Van Steenwyk, 2003), but it has not yet been approved for widespread, open-Weld release, pending evaluation of its potential eVects on non- target species. Although P. concolor has been reared from numerous tephritid species in the laboratory and in agricul- tural settings (Wharton and Gilstrap, 1983), only two * Corresponding author. Fax: +1 510 643 5438. E-mail address: [email protected] (K.R. Sime).
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Comparison of two laboratory cultures of Psyttalia concolor (Hymenoptera: Braconidae), as a...

Biological Control 39 (2006) 248–255
www.elsevier.com/locate/ybcon
Comparison of two laboratory cultures of Psyttalia concolor (Hymenoptera: Braconidae), as a parasitoid of the olive fruit Xy
Karen R. Sime a,¤, Kent M. Daane a, Russell H. Messing b, Marshall W. Johnson c
a Division of Organisms and Environment and Center for Biological Control, University of California, Berkeley, CA 94720-3114, USAb University of Hawaii, Kauai Agricultural Research Center, 7370 Kuamoo Road, Kapaa, Kauai, HI 96746, USA
c Department of Entomology, University of California,Riverside, CA 92521, USA
Received 2 March 2006; accepted 14 June 2006Available online 27 June 2006
Abstract
Life-history trials were conducted in the laboratory on two cultures of Psyttalia concolor (Szépligeti) (Hymenoptera: Braconidae)reared on olive fruit Xy, Bactrocera oleae (Rossi) (Diptera: Tephritidae). The tested cultures (A and B) had similar histories except thatCulture A was maintained in Kenya for four years. Results showed that parasitoids from both cultures preferentially searched on olivefruit containing 8 to 10-day-old (second and third instar) olive Xies, and reproduced most successfully when third-instar Xy larvae wereavailable. The mean longevity of adult female P. concolor was a negative function of temperature. Females from Culture B lived signiW-cantly longer at 15, 22, 25, and 32 °C than females from Culture A. For both cultures, adult female longevity was signiWcantly longer whenthe parasitoids were provided with honey than when provided water alone, or nothing; the presence of hosts signiWcantly reduced longev-ity, suggesting an energetic cost for reproduction. The parasitoids produced an average of 28.7§ 4.1 and 22.2§ 5.1 oVspring per female inCultures A and B, respectively. The results are discussed with respect to use of biological control agents held under diVerent rearing con-ditions, and the potential of P. concolor for use as a biological control agent for olive fruit Xy in California.© 2006 Elsevier Inc. All rights reserved.
Keywords: Psyttalia concolor; Braconidae; Bactrocera oleae; Tephritidae; Olea; Biological control; Parasitoid biology; Olive
1. Introduction
The olive fruit Xy, Bactrocera oleae (Rossi) (Diptera:Tephritidae), has long been a pest of olives in the Mediter-ranean basin (Clausen, 1978; Tzanakakis, 2003; White andElson-Harris, 1992). It was discovered in southern Califor-nia in 1998 and within four years had spread nearlythroughout the state, posing a serious threat to the oliveindustry (Rice et al., 2003). Current research eVorts empha-size the development of sustainable management practices.Insecticidal baits are currently the principal means of con-trol, but are limited in their eVectiveness. Infested trees insuburban and rural landscaping act as reservoirs for reinva-sion (Collier and Van Steenwyk, 2003), and, furthermore,
* Corresponding author. Fax: +1 510 643 5438.E-mail address: [email protected] (K.R. Sime).
1049-9644/$ - see front matter © 2006 Elsevier Inc. All rights reserved.doi:10.1016/j.biocontrol.2006.06.007
the successful biological controls that have been establishedfor scale pests in California olives (Daane et al., 2005)might be disrupted by insecticides. As no natural enemiesexist in California that can eVectively suppress olive Xy, aclassical biological control program was initiated in 2002.EVorts to identify useful natural enemies have to dateincluded foreign exploration in Africa and Central Asia,and evaluation of parasitoids used to control tephritid pestselsewhere (Hoelmer et al., 2004; Sime et al., 2006).
Among the olive Xy parasitoids readily available in cul-ture is Psyttalia concolor (Szépligeti) (Hymenoptera: Bra-conidae). Recently, this species was released in Californiaon a limited basis (Collier and Van Steenwyk, 2003), but ithas not yet been approved for widespread, open-Weldrelease, pending evaluation of its potential eVects on non-target species. Although P. concolor has been reared fromnumerous tephritid species in the laboratory and in agricul-tural settings (Wharton and Gilstrap, 1983), only two

K.R. Sime et al. / Biological Control 39 (2006) 248–255 249
species (the olive Xy and the Mediterranean fruit Xy(MedXy), Ceratitis capitata (Wiedemann)) are known astypical hosts in its native range. In parts of northern andeastern Africa, P. concolor is regularly reared from olive Xyin both wild and cultivated olives (Clausen, 1978; Copelandet al., 2004; El-Heneidy et al., 2001; Gaouar and Debouzie,1991), and it is also frequently reared from MedXy inKenya (Wharton et al., 2000) and North Africa (Naraya-nan and Chawla, 1962). European programs for olive Xybiological control have focused on P. concolor to the exclu-sion of almost all other parasitoid species, especially afteran eYcient mass-rearing method (using MedXy in artiWcialdiet) for P. concolor was developed in the 1950s (Great-head, 1976). Shortly after it was Wrst identiWed in materialcollected in Tunisia in 1910, P. concolor was introduced toItaly and France, and in subsequent decades was releasedwidely in the olive-growing regions of southern Europe.Few of these introductions resulted in establishment, how-ever, and where it has established, inundative releases arerequired to boost parasitism rates (Clausen, 1978; Great-head, 1976). Wharton (1989) has argued that the nearlycentury-long reliance on P. concolor has had more to dowith the ease of collecting and rearing this species, com-pared to other olive Xy parasitoids, than with its proveneVectiveness as a control agent. He further argues that this“misguided” emphasis on P. concolor has come at theexpense of investigating the potential of other parasitoidspecies.
Despite this history, we considered P. concolor an impor-tant species to screen for use in California’s biological con-trol program against the olive Xy. Several considerationssuggest that P. concolor merits further investigation. First,mass cultures of P. concolor are maintained on MedXy,because this host is easily reared on artiWcial diets. Changesin P. concolor mating and oviposition behavior have beennoted in cultures maintained for many generations, how-ever, and it has been suggested that these and other traitsassociated with the ease of rearing may diminish P. con-color eVectiveness in the Weld (Kimani-Njogu et al., 2001),especially as a parasitoid of the olive Xy (i.e., a novel hostoutside of the culture). ModiWcations of culture protocols,such as increasing the addition of wild stock and rearing onolive Xy rather than MedXy, may thus improve the perfor-mance of P. concolor in an olive Xy program. These modiW-cations are now possible with improvements in rearingmethods. (In California, rearing on MedXy is impossible,because MedXy is not considered to have established and itcannot be legally imported into the continental USA(Headrick and Goeden, 1996).) Second, P. concolor is awidespread species: its native range spans most of aridnorthern and eastern Africa (Kimani-Njogu et al., 2001;Wharton and Gilstrap, 1983). It likely comprises severalcryptic species or genetically diVerentiated populations, andgiven the diversity of habitats and climates encompassed bythis distribution, there may exist races adapted to particularenvironments or hosts (Diehl and Bush, 1984; Hopperet al., 1993; Kimani-Njogu et al., 2001; Wharton and
Gilstrap, 1983). Despite the mixed results in biological con-trol programs, no attempt has yet been made to comparethe biology of P. concolor from diVerent regions speciWcallyto determine whether some populations are diVerentiallyadapted to olive Xy or to MedXy, or whether some popula-tions fare better in certain climates. Finally, the biology ofP. concolor is known almost entirely from studies con-ducted using MedXy in artiWcial diet as the host (Avilla andAlbajes, 1983; Biliotti and Delanoue, 1959; Canale andRaspi, 2000; Canard et al., 1979; Loni, 1997; Stavraki-Pau-lopoulou, 1966; Wang and Messing, 2004). The identiWca-tion of diVerences in its biology when attacking olive Xylarvae reared in whole fruit could lead to improvement ofinsectary and Weld-release protocols.
The goals of this study were to investigate the biology ofP. concolor when using olive Xy as the host, in olive fruit,and to determine whether intra-speciWc variation exists inbiological traits relevant to its performance in the insectaryand in the Weld. To begin our investigation of intra-speciWcvariation in P. concolor, we worked with two longstandinglaboratory cultures of this species. Although both culturescame from stock maintained on MedXy, we switched bothto olive Xy for approximately ten generations prior to thebeginning of this study, and conducted all experiments witholive Xy (reared in picked fruit) as host. To investigate hostadaptation, we compared the oviposition behavior andfecundity of females from the two colonies. To investigatetheir hardiness in diVerent conditions, we compared femalelongevity at diVerent temperatures and given diVerent pro-visions. The short-term goal of these studies was to deter-mine whether either or both of the cultures would provideimproved stock for innoculative release against olive Xy inCalifornia. In the long term, however, determining whetherthere exists genetic variation for host compatibility or cli-mate tolerance in P. concolor will help direct foreign explo-ration for biological control of both the olive Xy and theMedXy, and will help improve mass-production techniques.
2. Materials and methods
2.1. Sources of insects and plants, and culture maintenance
Laboratory cultures of olive Xy were derived frominfested olives collected near Davis, California (YoloCounty). Flies were reared on olive fruit following themethods set forth by Tzanakakis (1989, 2003). Because theXies do not develop on immature fruit less than twomonths old, and olives picked when fully ripe tend to rotbefore the Xy larvae (or their parasitoids) complete devel-opment, we used a variety of olive cultivars (mostly Man-zanillo, Sevillano, and Mission) with varying periods ofripening. These cultivars could be collected at diVerenttimes across a long section of the state (Riverside, Kern,Tulare, Fresno, and Yolo Counties), thereby providingfruit of an acceptable quality for 9 to 10 months out of theyear. Olives held in cold storage were used for the remain-ing period.

250 K.R. Sime et al. / Biological Control 39 (2006) 248–255
Olives were exposed to adult Xies in an ovipositionalchamber (45£ 45£ 45 cm wooden cage with organdy sidesand a glass top) that was kept in a temperature-controlledroom (22§ 2 °C, 16:8 (L:D)h, 40% RH) at the University ofCalifornia Berkeley Insectary and Quarantine Facility(Berkeley I&Q). The adult Xies had free access to water anda mixture of honey and a dry yeast extract (2:1 by volume)(FisherBiotech, Fairlawn, New Jersey). Olives were left inthe cage for 1–2 d or until each had 5–10 oviposition marks.Infested olives were then transferred to plastic boxes(36£ 18£10 cm) with wire-mesh tops. To reduce moldgrowth, olives were placed in the box no more than 2–3 lay-ers deep and were raised about 2 cm oV the bottom of thecontainer by a metal grid. Under these conditions, themature larvae exited the fruit and pupated on the box bot-tom after 10–14 d. Puparia were collected and transferredto the ovipositional chamber to emerge as adults and repeatthe process.
The two P. concolor laboratory cultures used in thisstudy originated with material Weld-collected in NorthAfrica in the 1970s and then maintained for decades in Italy(University of Pisa), where they were reared on MedXy(Canale, 1998). This initial “Italian” culture is the same cul-ture referred to by Kimani-Njogu (2001), Loni (1997), andRaspi and Canale (2000). Our Wrst culture, referred to here-after as “Culture A”, was sent from the Italian culture toKenya (International Center of Insect Physiology andEcology (ICIPE), Nairobi) in 1998, from Kenya to Hawaii(Kauai Agricultural Research Center) in 2002, and thenfrom Hawaii to Berkeley I&Q in 2003. The second culture,referred to hereafter as “Culture B”, was sent from the Ital-ian culture to Hawaii (Kauai Agricultural Research Center)in 1999 and then from Hawaii to Berkeley I&Q in 2003. Thecultures thus had similar backgrounds before arriving atBerkeley I&Q for our studies. The important diVerence isthat Culture A left Italy in 1998 and was maintained inKenya for four years. Otherwise, these colonies should rep-resent the same population. Culture techniques were simi-lar. The only diVerence we are aware of is that in Kenya(i.e., Culture A) the culture was maintained at a slightlyhigher temperature (27–28 °C) (Mohamed et al., 2003) thanin Hawaii (20–24 °C) (Wang and Messing, 2002).
The adult parasitoids were held in 45£ 45£ 45 cm3 cages(as described for the olive Xy) that were freely provisionedwith water and a honey–water solution (50% by volume)and kept in a temperature-controlled room (22§2 °C, par-tial natural light augmented by 16:8 (L:D)h, 40% RH). Pre-vious reports describe P. concolor (reared on C. capitata) asa koinobiont that favors third (last) instar larvae for ovipo-sition, with the larva completing its development in thehost’s puparium (Biliotti and Delanoue, 1959; Canale,1998; Wang and Messing, 2004). Olives infested 6–10 d ear-lier and thus containing a mixture of second- and third-instar Xy larvae were exposed to parasitoids for 1–3 d,depending on parasitoid density. The inoculated materialwas then transferred to plastic boxes as described above.The Xy larvae dropped to the bottom of these containers to
pupate, at which time the puparia were collected and trans-ferred to transparent plastic Petri dishes (9-cm diam) thatwere monitored daily for the emergence of adult Xies andparasitoids. Experiments described below were conductedbeginning in August 2004, thus over a year (t10 genera-tions) after the cultures were brought to Berkeley I&Q andswitched to rearing on olive Xy.
2.2. Host stages used for oviposition
Host-stage preference and reproductive success on diVer-ent olive Xy development stages were examined in choicetests. To produce an age series of olive Xies, fresh olives wereexposed to adult Xies for 8h every 2 d and then held at25§1°C. The olives were presented to the parasitoids whenthe Xy larvae were 2, 4, 6, 8, 10, and 12-d-old. A subsample ofolives from each set was dissected shortly before the test todetermine which larval stages were present. Under these con-ditions, 2-d-old olives contained eggs and, rarely (<5%), Wrstinstars; 4-d-old olives contained Wrst instars; 6-d-old olivescontained second instars; 8-d-old olives contained secondand young third instars; 10-d-old olives contained thirdinstars; and 12-d-old olives contained mature third instarsand were accompanied by prepupal larvae (emerging fromthe fruit) and occasionally puparia.
For each replicate, four female parasitoids were held for24 h in an ovipositional chamber (a translucent white plasticcylinder 13 cm deep £20 cm diam, with a Wne mesh top) con-taining 24 olives, of which four each were infested by one ofthe six olive Xy age classes. The olives were placed in the bot-tom of the container, with the olives of each olive Xy age classgrouped in open Petri dishes (5-cm diam). During the Wrst 7–8 h of the exposure period, ten 5-s observations were made, atapproximately 40-min intervals, of activity within the con-tainers. The age class of olives contacted by parasitoids wasrecorded. After the parasitoids were removed from the ovi-positional chamber, the olives were held at 25§1 °C to reareither adult parasitoids or olive Xies. There were 7 replicatesfor Culture A and 10 replicates for Culture B.
2.3. Adult longevity at various temperatures
Adult male and female longevities were measured at sixtemperatures (15.2§0.3, 21.9§0.2, 24.8§0.4, 27.8§0.2,30.1§ 0.2, and 32.0§ 0.5 °C). Newly emerged parasitoidswere placed singly in glass vials (5-cm long£ 1-cm diam,with a mesh lid), provisioned with a streak of honey–water(50% solution by volume), and randomly assigned to a tem-perature cabinet. The parasitoids were checked daily formortality. The honey–water was replenished every 2–3 d oras needed. From each culture, 10 male and 10 female para-sitoids were tested at each temperature.
2.4. Adult longevity given diVerent provisions
Female longevity was compared among Wve treatmentswith access to (1) olives containing hosts, honey–water
K.R. Sime et al. / Biological Control 39 (2006) 248–255 251
(50% by volume), and water; (2) uninfested olives, honey–water, and water; (3) honey–water and water only; (4)water only; and (5) no provisions. Newly emerged femaleswere collected daily, transferred to a small container withmales, supplied with water and honey–water, and held for 2d to mate. The females were then randomly assigned to oneof the Wve treatments, with each parasitoid isolated in asmall plastic container (15 cm diam£ 6 cm deep) with ahole (7 cm diam) cut in the lid and covered with nylon meshfor ventilation. The olives (four per container) werereplaced every other day. Where olives with hosts wereoVered, the Xy larvae were at a suitable stage for parasitoidoviposition (mostly third instars). Each of the four oliveshad 5–10 oviposition marks, and therefore some 20–40 lar-vae were available over each 2 d interval. Honey-water,streaked along the sides of the container, and distilledwater, in a soaked cotton wick, were freely available. Para-sitoids were checked daily for mortality. All treatmentswere kept in a temperature-controlled room (22§2 °C, 40%RH, 16:8 (L:D) h supplemented by natural daylight). Eachtreatment was replicated 10 times for each culture.
2.5. Lifetime fecundity
To determine lifetime reproductive potential, theinfested olives that were collected every other day in theexperiment described previously (Section 2.4) were held inplastic cups for the emergence of adult Xies or parasitoids.The number and sex of the emerging oVspring wererecorded.
2.6. Statistics
Results are presented as means per treatment (§SE).Treatment eVects were analyzed using analysis of variance(ANOVA), with treatment means separated using Tukey’sHSD test (three or more treatments) or t tests (two-waycomparisons).
3. Results and discussion
3.1. Host stages used for oviposition
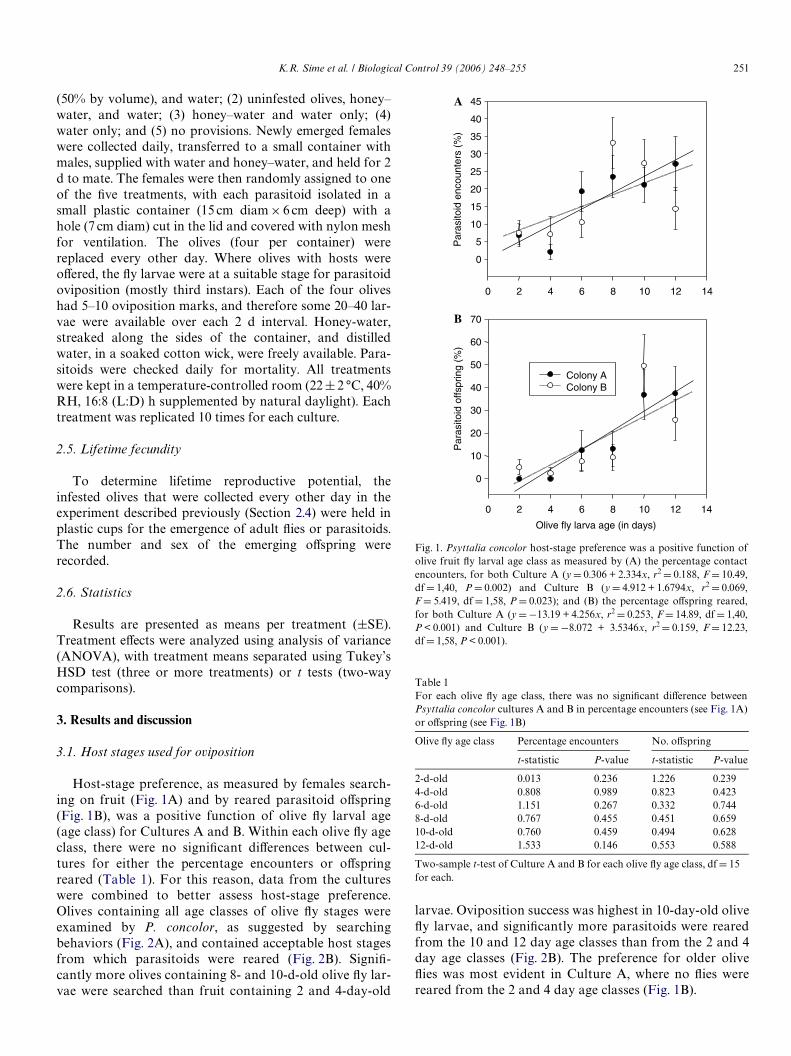
Host-stage preference, as measured by females search-ing on fruit (Fig. 1A) and by reared parasitoid oVspring(Fig. 1B), was a positive function of olive Xy larval age(age class) for Cultures A and B. Within each olive Xy ageclass, there were no signiWcant diVerences between cul-tures for either the percentage encounters or oVspringreared (Table 1). For this reason, data from the cultureswere combined to better assess host-stage preference.Olives containing all age classes of olive Xy stages wereexamined by P. concolor, as suggested by searchingbehaviors (Fig. 2A), and contained acceptable host stagesfrom which parasitoids were reared (Fig. 2B). SigniW-cantly more olives containing 8- and 10-d-old olive Xy lar-vae were searched than fruit containing 2 and 4-day-old
larvae. Oviposition success was highest in 10-day-old oliveXy larvae, and signiWcantly more parasitoids were rearedfrom the 10 and 12 day age classes than from the 2 and 4day age classes (Fig. 2B). The preference for older oliveXies was most evident in Culture A, where no Xies werereared from the 2 and 4 day age classes (Fig. 1B).
Fig. 1. Psyttalia concolor host-stage preference was a positive function ofolive fruit Xy larval age class as measured by (A) the percentage contactencounters, for both Culture A (y D 0.306 + 2.334x, r2 D 0.188, F D 10.49,dfD 1,40, P D 0.002) and Culture B (y D 4.912 + 1.6794x, r2 D 0.069,FD 5.419, df D 1,58, P D 0.023); and (B) the percentage oVspring reared,for both Culture A (y D¡13.19 + 4.256x, r2 D 0.253, FD 14.89, df D 1,40,P < 0.001) and Culture B (y D¡8.072 + 3.5346x, r2 D 0.159, F D 12.23,dfD 1,58, P < 0.001).
Olive fly larva age (in days)
0 2 4 6 8 10 12 14
Par
asito
id o
ffspr
ing
(%)
0
10
20
30
40
50
60
70B
0 2 4 6 8 10 12 14
Par
asito
id e
ncou
nter
s (%
)
0
5
10
15
20
25
30
35
40
45
Colony AColony B
A
Table 1For each olive Xy age class, there was no signiWcant diVerence betweenPsyttalia concolor cultures A and B in percentage encounters (see Fig. 1A)or oVspring (see Fig. 1B)
Two-sample t-test of Culture A and B for each olive Xy age class, dfD 15 for each.
Olive Xy age class Percentage encounters No. oVspring
t-statistic P-value t-statistic P-value
2-d-old 0.013 0.236 1.226 0.2394-d-old 0.808 0.989 0.823 0.4236-d-old 1.151 0.267 0.332 0.7448-d-old 0.767 0.455 0.451 0.65910-d-old 0.760 0.459 0.494 0.62812-d-old 1.533 0.146 0.553 0.588
252 K.R. Sime et al. / Biological Control 39 (2006) 248–255
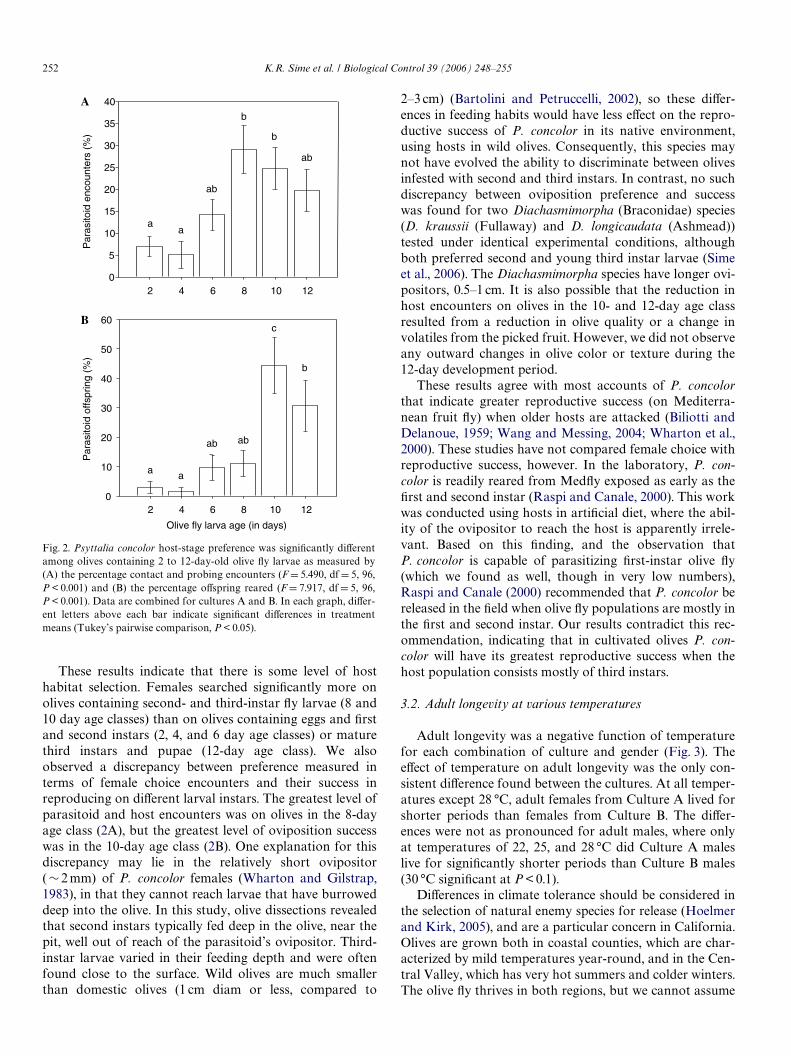
These results indicate that there is some level of hosthabitat selection. Females searched signiWcantly more onolives containing second- and third-instar Xy larvae (8 and10 day age classes) than on olives containing eggs and Wrstand second instars (2, 4, and 6 day age classes) or maturethird instars and pupae (12-day age class). We alsoobserved a discrepancy between preference measured interms of female choice encounters and their success inreproducing on diVerent larval instars. The greatest level ofparasitoid and host encounters was on olives in the 8-dayage class (2A), but the greatest level of oviposition successwas in the 10-day age class (2B). One explanation for thisdiscrepancy may lie in the relatively short ovipositor(»2 mm) of P. concolor females (Wharton and Gilstrap,1983), in that they cannot reach larvae that have burroweddeep into the olive. In this study, olive dissections revealedthat second instars typically fed deep in the olive, near thepit, well out of reach of the parasitoid’s ovipositor. Third-instar larvae varied in their feeding depth and were oftenfound close to the surface. Wild olives are much smallerthan domestic olives (1 cm diam or less, compared to
Fig. 2. Psyttalia concolor host-stage preference was signiWcantly diVerentamong olives containing 2 to 12-day-old olive Xy larvae as measured by(A) the percentage contact and probing encounters (F D 5.490, df D 5, 96,P < 0.001) and (B) the percentage oVspring reared (FD 7.917, df D 5, 96,P < 0.001). Data are combined for cultures A and B. In each graph, diVer-ent letters above each bar indicate signiWcant diVerences in treatmentmeans (Tukey’s pairwise comparison, P < 0.05).
Olive fly larva age (in days)
2 4 6 8 10 120
5
10
15
20
25
30
35
40
2 4 6 8 10 120
10
20
30
40
50
60
Par
asito
id o
ffspr
ing
(%)
B
Par
asito
id e
ncou
nter
s (%
)
A
a a
ab ab
b
c
aa
ab
b
ab
b
2–3 cm) (Bartolini and Petruccelli, 2002), so these diVer-ences in feeding habits would have less eVect on the repro-ductive success of P. concolor in its native environment,using hosts in wild olives. Consequently, this species maynot have evolved the ability to discriminate between olivesinfested with second and third instars. In contrast, no suchdiscrepancy between oviposition preference and successwas found for two Diachasmimorpha (Braconidae) species(D. kraussii (Fullaway) and D. longicaudata (Ashmead))tested under identical experimental conditions, althoughboth preferred second and young third instar larvae (Simeet al., 2006). The Diachasmimorpha species have longer ovi-positors, 0.5–1 cm. It is also possible that the reduction inhost encounters on olives in the 10- and 12-day age classresulted from a reduction in olive quality or a change involatiles from the picked fruit. However, we did not observeany outward changes in olive color or texture during the12-day development period.
These results agree with most accounts of P. concolorthat indicate greater reproductive success (on Mediterra-nean fruit Xy) when older hosts are attacked (Biliotti andDelanoue, 1959; Wang and Messing, 2004; Wharton et al.,2000). These studies have not compared female choice withreproductive success, however. In the laboratory, P. con-color is readily reared from MedXy exposed as early as theWrst and second instar (Raspi and Canale, 2000). This workwas conducted using hosts in artiWcial diet, where the abil-ity of the ovipositor to reach the host is apparently irrele-vant. Based on this Wnding, and the observation thatP. concolor is capable of parasitizing Wrst-instar olive Xy(which we found as well, though in very low numbers),Raspi and Canale (2000) recommended that P. concolor bereleased in the Weld when olive Xy populations are mostly inthe Wrst and second instar. Our results contradict this rec-ommendation, indicating that in cultivated olives P. con-color will have its greatest reproductive success when thehost population consists mostly of third instars.
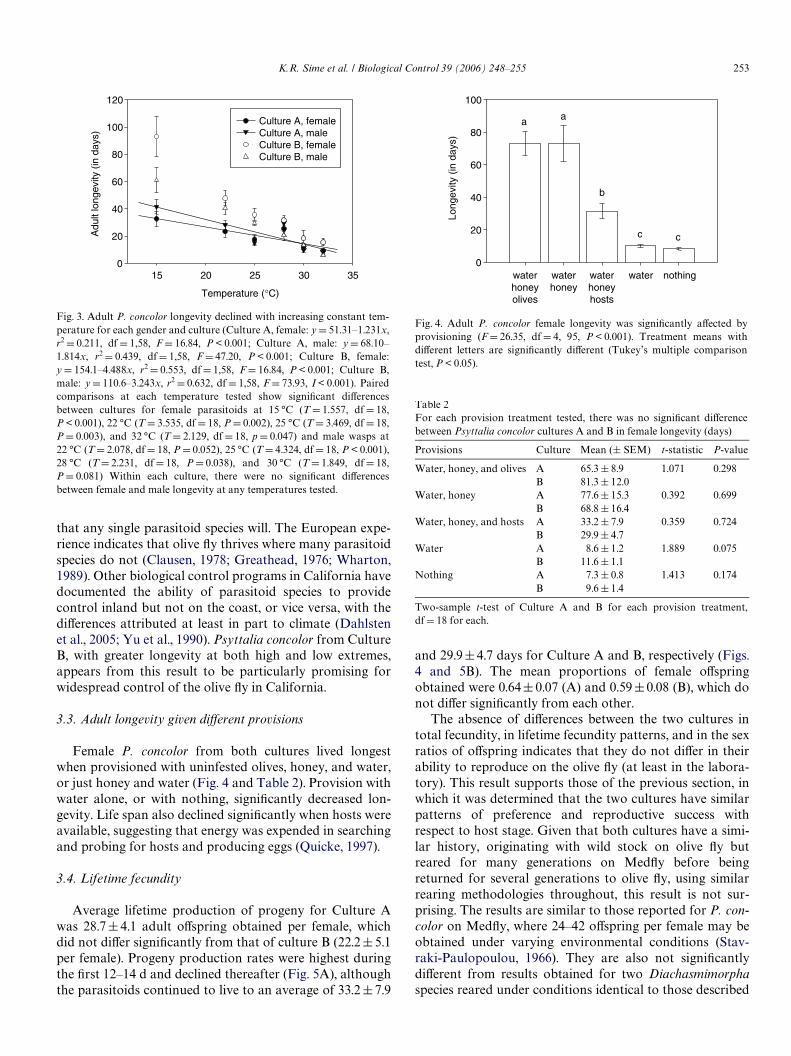
3.2. Adult longevity at various temperatures
Adult longevity was a negative function of temperaturefor each combination of culture and gender (Fig. 3). TheeVect of temperature on adult longevity was the only con-sistent diVerence found between the cultures. At all temper-atures except 28 °C, adult females from Culture A lived forshorter periods than females from Culture B. The diVer-ences were not as pronounced for adult males, where onlyat temperatures of 22, 25, and 28 °C did Culture A maleslive for signiWcantly shorter periods than Culture B males(30 °C signiWcant at P < 0.1).
DiVerences in climate tolerance should be considered inthe selection of natural enemy species for release (Hoelmerand Kirk, 2005), and are a particular concern in California.Olives are grown both in coastal counties, which are char-acterized by mild temperatures year-round, and in the Cen-tral Valley, which has very hot summers and colder winters.The olive Xy thrives in both regions, but we cannot assume
K.R. Sime et al. / Biological Control 39 (2006) 248–255 253
that any single parasitoid species will. The European expe-rience indicates that olive Xy thrives where many parasitoidspecies do not (Clausen, 1978; Greathead, 1976; Wharton,1989). Other biological control programs in California havedocumented the ability of parasitoid species to providecontrol inland but not on the coast, or vice versa, with thediVerences attributed at least in part to climate (Dahlstenet al., 2005; Yu et al., 1990). Psyttalia concolor from CultureB, with greater longevity at both high and low extremes,appears from this result to be particularly promising forwidespread control of the olive Xy in California.
3.3. Adult longevity given diVerent provisions
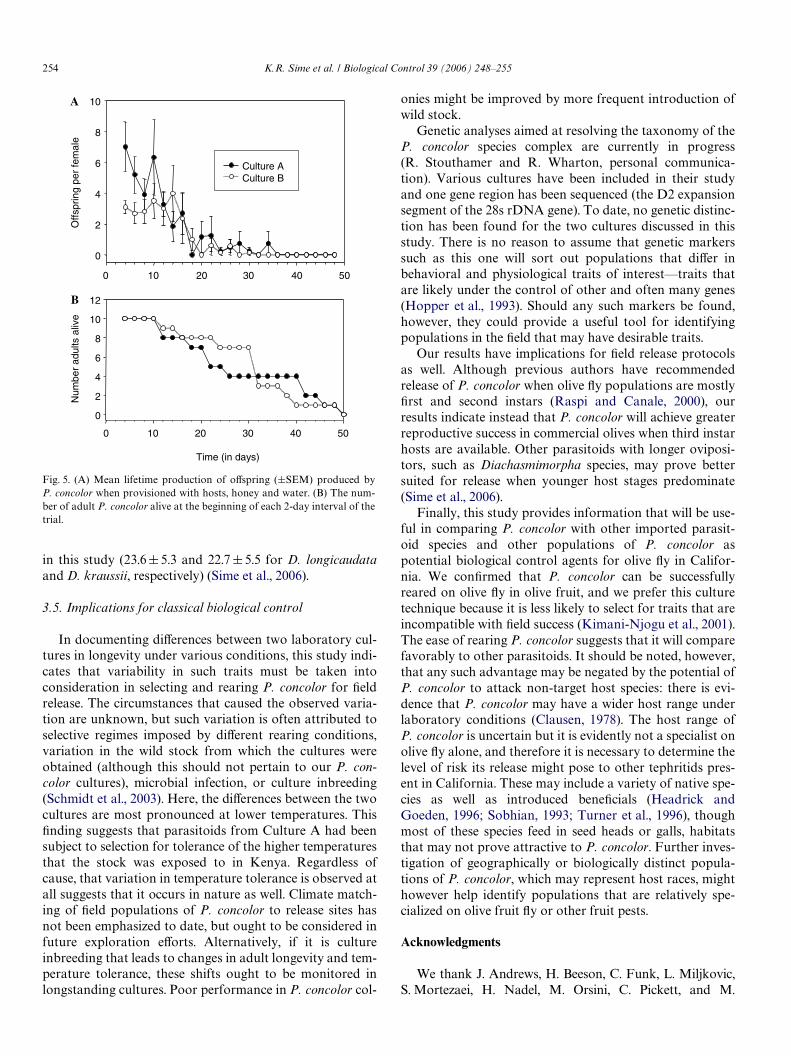
Female P. concolor from both cultures lived longestwhen provisioned with uninfested olives, honey, and water,or just honey and water (Fig. 4 and Table 2). Provision withwater alone, or with nothing, signiWcantly decreased lon-gevity. Life span also declined signiWcantly when hosts wereavailable, suggesting that energy was expended in searchingand probing for hosts and producing eggs (Quicke, 1997).
3.4. Lifetime fecundity
Average lifetime production of progeny for Culture Awas 28.7§ 4.1 adult oVspring obtained per female, whichdid not diVer signiWcantly from that of culture B (22.2§5.1per female). Progeny production rates were highest duringthe Wrst 12–14 d and declined thereafter (Fig. 5A), althoughthe parasitoids continued to live to an average of 33.2§7.9
Fig. 3. Adult P. concolor longevity declined with increasing constant tem-perature for each gender and culture (Culture A, female: y D 51.31–1.231x,r2 D 0.211, df D 1,58, F D 16.84, P < 0.001; Culture A, male: y D 68.10–1.814x, r2 D 0.439, dfD 1,58, F D 47.20, P < 0.001; Culture B, female:y D 154.1–4.488x, r2 D 0.553, dfD 1,58, FD 16.84, P < 0.001; Culture B,male: y D 110.6–3.243x, r2 D 0.632, df D 1,58, F D 73.93, I < 0.001). Pairedcomparisons at each temperature tested show signiWcant diVerencesbetween cultures for female parasitoids at 15 °C (T D 1.557, df D 18,P < 0.001), 22 °C (T D 3.535, df D 18, P D 0.002), 25 °C (T D 3.469, df D 18,P D 0.003), and 32 °C (T D 2.129, df D 18, p D 0.047) and male wasps at22 °C (TD 2.078, dfD 18, P D 0.052), 25 °C (T D 4.324, df D 18, P < 0.001),28 °C (T D 2.231, dfD 18, P D 0.038), and 30 °C (T D 1.849, dfD 18,P D 0.081) Within each culture, there were no signiWcant diVerencesbetween female and male longevity at any temperatures tested.
Temperature (°C)
15 20 25 30 35
Adu
lt lo
ngev
ity (
in d
ays)
0
20
40
60
80
100
120
Culture A, femaleCulture A, maleCulture B, femaleCulture B, male
and 29.9§4.7 days for Culture A and B, respectively (Figs.4 and 5B). The mean proportions of female oVspringobtained were 0.64§ 0.07 (A) and 0.59§0.08 (B), which donot diVer signiWcantly from each other.
The absence of diVerences between the two cultures intotal fecundity, in lifetime fecundity patterns, and in the sexratios of oVspring indicates that they do not diVer in theirability to reproduce on the olive Xy (at least in the labora-tory). This result supports those of the previous section, inwhich it was determined that the two cultures have similarpatterns of preference and reproductive success withrespect to host stage. Given that both cultures have a simi-lar history, originating with wild stock on olive Xy butreared for many generations on MedXy before beingreturned for several generations to olive Xy, using similarrearing methodologies throughout, this result is not sur-prising. The results are similar to those reported for P. con-color on MedXy, where 24–42 oVspring per female may beobtained under varying environmental conditions (Stav-raki-Paulopoulou, 1966). They are also not signiWcantlydiVerent from results obtained for two Diachasmimorphaspecies reared under conditions identical to those described
Fig. 4. Adult P. concolor female longevity was signiWcantly aVected byprovisioning (F D 26.35, df D 4, 95, P < 0.001). Treatment means withdiVerent letters are signiWcantly diVerent (Tukey’s multiple comparisontest, P < 0.05).
waterhoneyolives
waterhoney
waterhoneyhosts
water nothing0
20
40
60
80
100
Long
evity
(in
day
s)
a a
b
c c
Table 2For each provision treatment tested, there was no signiWcant diVerencebetween Psyttalia concolor cultures A and B in female longevity (days)
Two-sample t-test of Culture A and B for each provision treatment,dfD 18 for each.
Provisions Culture Mean (§ SEM) t-statistic P-value
Water, honey, and olives A 65.3 § 8.9 1.071 0.298B 81.3 § 12.0
Water, honey A 77.6 § 15.3 0.392 0.699B 68.8 § 16.4
Water, honey, and hosts A 33.2 § 7.9 0.359 0.724B 29.9 § 4.7
Water A 8.6 § 1.2 1.889 0.075B 11.6 § 1.1
Nothing A 7.3 § 0.8 1.413 0.174B 9.6 § 1.4
254 K.R. Sime et al. / Biological Control 39 (2006) 248–255
in this study (23.6§5.3 and 22.7§5.5 for D. longicaudataand D. kraussii, respectively) (Sime et al., 2006).
3.5. Implications for classical biological control
In documenting diVerences between two laboratory cul-tures in longevity under various conditions, this study indi-cates that variability in such traits must be taken intoconsideration in selecting and rearing P. concolor for Weldrelease. The circumstances that caused the observed varia-tion are unknown, but such variation is often attributed toselective regimes imposed by diVerent rearing conditions,variation in the wild stock from which the cultures wereobtained (although this should not pertain to our P. con-color cultures), microbial infection, or culture inbreeding(Schmidt et al., 2003). Here, the diVerences between the twocultures are most pronounced at lower temperatures. ThisWnding suggests that parasitoids from Culture A had beensubject to selection for tolerance of the higher temperaturesthat the stock was exposed to in Kenya. Regardless ofcause, that variation in temperature tolerance is observed atall suggests that it occurs in nature as well. Climate match-ing of Weld populations of P. concolor to release sites hasnot been emphasized to date, but ought to be considered infuture exploration eVorts. Alternatively, if it is cultureinbreeding that leads to changes in adult longevity and tem-perature tolerance, these shifts ought to be monitored inlongstanding cultures. Poor performance in P. concolor col-
Fig. 5. (A) Mean lifetime production of oVspring (§SEM) produced byP. concolor when provisioned with hosts, honey and water. (B) The num-ber of adult P. concolor alive at the beginning of each 2-day interval of thetrial.
0 10 20 30 40 50
0
2
4
6
8
10
Culture ACulture B
Time (in days)
0 10 20 30 40 50
0
2
4
6
8
10
12
Offs
prin
g pe
r fe
mal
eN
umbe
r ad
ults
aliv
e
A
B
onies might be improved by more frequent introduction ofwild stock.
Genetic analyses aimed at resolving the taxonomy of theP. concolor species complex are currently in progress(R. Stouthamer and R. Wharton, personal communica-tion). Various cultures have been included in their studyand one gene region has been sequenced (the D2 expansionsegment of the 28s rDNA gene). To date, no genetic distinc-tion has been found for the two cultures discussed in thisstudy. There is no reason to assume that genetic markerssuch as this one will sort out populations that diVer inbehavioral and physiological traits of interest—traits thatare likely under the control of other and often many genes(Hopper et al., 1993). Should any such markers be found,however, they could provide a useful tool for identifyingpopulations in the Weld that may have desirable traits.
Our results have implications for Weld release protocolsas well. Although previous authors have recommendedrelease of P. concolor when olive Xy populations are mostlyWrst and second instars (Raspi and Canale, 2000), ourresults indicate instead that P. concolor will achieve greaterreproductive success in commercial olives when third instarhosts are available. Other parasitoids with longer oviposi-tors, such as Diachasmimorpha species, may prove bettersuited for release when younger host stages predominate(Sime et al., 2006).
Finally, this study provides information that will be use-ful in comparing P. concolor with other imported parasit-oid species and other populations of P. concolor aspotential biological control agents for olive Xy in Califor-nia. We conWrmed that P. concolor can be successfullyreared on olive Xy in olive fruit, and we prefer this culturetechnique because it is less likely to select for traits that areincompatible with Weld success (Kimani-Njogu et al., 2001).The ease of rearing P. concolor suggests that it will comparefavorably to other parasitoids. It should be noted, however,that any such advantage may be negated by the potential ofP. concolor to attack non-target host species: there is evi-dence that P. concolor may have a wider host range underlaboratory conditions (Clausen, 1978). The host range ofP. concolor is uncertain but it is evidently not a specialist onolive Xy alone, and therefore it is necessary to determine thelevel of risk its release might pose to other tephritids pres-ent in California. These may include a variety of native spe-cies as well as introduced beneWcials (Headrick andGoeden, 1996; Sobhian, 1993; Turner et al., 1996), thoughmost of these species feed in seed heads or galls, habitatsthat may not prove attractive to P. concolor. Further inves-tigation of geographically or biologically distinct popula-tions of P. concolor, which may represent host races, mighthowever help identify populations that are relatively spe-cialized on olive fruit Xy or other fruit pests.
Acknowledgments
We thank J. Andrews, H. Beeson, C. Funk, L. Miljkovic,S. Mortezaei, H. Nadel, M. Orsini, C. Pickett, and M.

K.R. Sime et al. / Biological Control 39 (2006) 248–255 255
Yamamoto for their assistance and advice. Funds providedby the California Specialty Crop Block Grant (administeredby the California Department of Food and Agriculture andthe United States Department of Agriculture) and the Cali-fornia Olive Committee are gratefully acknowledged.
References
Avilla, J., Albajes, R., 1983. Preliminary studies on mutual interference inOpius concolor Szépl. (Hymenoptera, Braconidae). Zeit. Angew. Ento-mol. 96, 27–32.
Bartolini, G., Petruccelli, R., 2002. ClassiWcation, origin, diVusion and his-tory of the olive. Food and Agriculture Organization of the UnitedNations, Rome.
Biliotti, E., Delanoue, P., 1959. Contribution a l’etude biologique d’Opiusconcolor Szépl. (Hym.: Braconidae) en elevage de laboratoire. Entom-ophaga 4, 7–14.
Canale, A., 1998. EVect of parasitoid/host ratio on superparasitism of Cerati-tis capitata (Wiedemann) larvae (Diptera, Tephritidae) by Opius concolorSzépligeti (Hymenoptera, Braconidae). Frustula Entomol. 21, 137–148.
Canale, A., Raspi, A., 2000. Host location and oviposition behaviour inOpius concolor Szépligeti (Hymenoptera: Braconidae). Entomol. Prob-lems 31, 25–32.
Canard, M., Liaropoulos, C., Laudeho, Y., 1979. Development of Opiusconcolor (Hym.: Braconidae) during the hypogeal phase of Dacus oleae(Dipt.: Trypetidae). Ann. Zool. Ecol. Anim. 11, 13–18.
Clausen, C.P., 1978. Introduced parasites and predators of arthropod pestsand weeds: a world review. United States Department of Agriculture,Agriculture Handbook No. 480, Washington, DC.
Collier, T.R., Van Steenwyk, R.A., 2003. Prospects for integrated controlof olive fruit Xy are promising in California. Calif. Agric. 57, 28–31.
Copeland, R.S., White, I.M., Okumu, M., Machera, P., Wharton, R.A.,2004. Insects associated with fruits of the Oleaceae (Asteridae, Lami-ales) in Kenya, with special reference to the Tephritidae (Diptera).Bishop Mus. Bull. Entomol. 12, 135–164.
Daane, K.M., Rice, R.E., Barnett, W.W., Zalom, F.G., Johnson, M.W.,2005. Arthropod pests. In: Sibbett, G., Ferguson, L. (Eds.), Olive Pro-duction Manual. U.C. Division of Agriculture and Natural Resources,Berkeley, pp. 105–114.
Dahlsten, D.L., Daane, K.M., Paine, T.D., Sime, K.R., Lawson, A.B., Row-ney, D.L., Roltsch, W., Andrews, J.W., Kabashima, J.N., Shaw, D.A.,Robb, K.L., Geisel, P.M., Chaney, W.E., Ingels, C.A., Varela, L.G.,Bianchi, M.L., 2005. Imported parasitoid helps control red gum lerppsyllid. Calif. Agric. 59, 229–234.
Diehl, S.R., Bush, G.L., 1984. An evolutionary and applied perspective oninsect biotypes. Annu. Rev. Entomol. 29, 471–504.
El-Heneidy, A.H., Omar, A.H., El-Sherif, H., El-Khawas, M.A., 2001. Sur-vey and seasonal abundance of the parasitoids of the olive fruit Xy,Bactrocera (Dacus) oleae Gmel. (Diptera: Trypetidae) in Egypt. Arab.J. Plant Prot. 19, 80–85.
Gaouar, N., Debouzie, D., 1991. Olive fruit Xy, Dacus oleae Gmel. (Dip-tera, Tephritidae), damage in Tlemcen region of Algeria. J. Appl. Ento-mol. 112, 288–297.
Greathead, D.J., 1976. A review of biological control in western and south-ern Europe. Farnham Royal, Commonwealth Agricultural Bureaux.
Headrick, D.H., Goeden, R.D., 1996. Issues concerning the eradication orestablishment and biological control of the Mediterranean fruit Xy,Ceratitis capitata (Wiedemann) (Diptera: Tephritidae), in California.Biol. Control 6, 412–421.
Hoelmer, K.A., Kirk, A., Wharton, R.A., Pickett, C.H., 2004. Foreign explo-ration for parasitoids of the olive fruit Xy, Bactrocera oleae. In: Woods,D. (Ed.), Biological control program annual summary, 2003. CDFAPlant Health and Pest Prevention Services, Sacramento, CA, pp. 12–14.
Hoelmer, K.A., Kirk, A.A., 2005. Selecting arthropod biological controlagents against arthropod pests: can the science be improved to decreasethe risk of releasing ineVective agents? Biol. Control 34, 255–264.
Hopper, K.R., Roush, R.T., Powell, W., 1993. Management of genetics ofbiological-control introductions. Annu. Rev. Entomol. 38, 27–51.
Kimani-Njogu, S.W., Trostle, M.K., Wharton, R.A., Woolley, J.B., Raspi,A., 2001. Biosystematics of the Psyttalia concolor species complex(Hymenoptera: Braconidae: Opiinae): the identity of populationsattacking Ceratitis capitata (Diptera: Tephritidae) in coVee in Kenya.Biol. Control 20, 167–174.
Loni, A., 1997. Developmental rate of Opius concolor (Hym: Braconidae)at various constant temperatures. Entomophaga 42, 359–366.
Mohamed, S.A., Overholt, W.A., Wharton, R.A., Lux, S.A., Eltoum, E.M.,2003. Host speciWcity of Psyttalia cosyrae (Hymenoptera: Braconidae)and the eVect of diVerent host species on parasitoid Wtness. Biol. Con-trol 28, 155–163.
Narayanan, E.S., Chawla, S.S., 1962. Parasites of fruit Xy pests of theworld. Beitr. Entomol. 12, 437–476.
Quicke, D.L.J., 1997. Parasitic wasps. Chapman & Hall, London.Raspi, A., Canale, A., 2000. EVect of superparasitism on Ceratitis capitata
(Wiedemann) (Diptera Tephritidae) second instar larvae by Psyttaliaconcolor (Szepligeti) (Hymenoptera Braconidae). Redia 83, 123–131.
Rice, R.E., Phillips, P.A., Stewart-Leslie, J., Sibbett, G.S., 2003. Olive fruitXy populations measured in central and southern California. Calif.Agric. 57, 122–127.
Schmidt, V.B., Linker, H.M., Orr, D.B., Kennedy, G.G., 2003. Variation inbiological parameters of Trichogramma spp. purchased from commer-cial suppliers in the United States. Biocontrol 48, 487–502.
Sime, K.R., Daane, K.M., Nadel, H., Funk, C.S., Messing, R.H., Andrews,J.W., Johnson, M.W., Pickett, C.H., 2006. Diachasmimorpha longicau-data and D. kraussii (Hymenoptera: Braconidae), potential parasitoidsof the olive fruit Xy. Biocontrol Sci. Tech. 16, 169–179.
Sobhian, R., 1993. Life-history and host-speciWcity of Urophora sirunaseva(Hering) (Dipt, Tephritidae), a candidate for biological control of yellowstarthistle, with remarks on the host plant. J. Appl. Entomol. 116, 381–390.
Stavraki-Paulopoulou, H.G., 1966. Contribution a l’etude de la capacitereproductrice et de la fecondite reelle d’Opius concolor Szépl. (Hyme-noptera-Braconidae). Annu. Epiphyties 17, 391–435.
Turner, C.E., Piper, G.L., Coombs, E.M., 1996. Chaetorellia australis (Dip-tera: Tephritidae) for biological control of yellow starthistle, Centaureasolstitialis (Compositae), in the western USA: establishment and seeddestruction. Bull. Entomol. Res. 86, 177–182.
Tzanakakis, M., 1989. Small scale rearing: Dacus oleae. In: Robinson, A.,Hooper, G. (Eds.), Fruit Flies: Their Biology, Natural Enemies andControl, Vol. 3B. Elsevier, Amsterdam, pp. 105–118.
Tzanakakis, M.E., 2003. Seasonal development and dormancy of insectsand mites feeding on olive: a review. Neth. J. Zool. 52, 87–224.
Wang, X.G., Messing, R.H., 2002. Newly imported larval parasitoids poseminimal competitive risk to extant egg-larval parasitoid of tephritidfruit Xies in Hawaii. Bull. Entomol. Res. 92, 423–429.
Wang, X.G., Messing, R.H., 2004. Potential interactions between pupaland egg- or larval-pupal parasitoids of tephritid fruit Xies. Environ.Entomol. 33, 1313–1320.
Wharton, R.A., 1989. Classical biological control of fruit-infesting Tephriti-dae. In: Robinson, A., Hooper, G. (Eds.), Fruit Flies: Their Biology, Nat-ural Enemies and Control, Vol. 3B. Elsevier, Amsterdam, pp. 303–313.
Wharton, R.A., Gilstrap, F., 1983. Key to and status of opiine braconid(Hymenoptera) parasitoids used in biological control of Ceratitis andDacus s.l. (Diptera: Tephritidae). Annu. Entomol. Soc. America 76,721–742.
Wharton, R.A., Trostle, M.K., Messing, R.H., Copeland, R.S., Kimani-Njogu, S.W., Lux, S., Overholt, W.A., Mohamed, S., Sivinski, J., 2000.Parasitoids of medXy, Ceratitis capitata, and related tephritids inKenyan coVee: a predominantly koinobiont assemblage. Bull. Ento-mol. Res. 90, 517–526.
White, I., Elson-Harris, M., 1992. Fruit Xies of economic signiWcance: theiridentiWcation and bionomics. CAB International, Oxon, UK.
Yu, D.S., Luck, R.F., Murdoch, W.W., 1990. Competition, resource parti-tioning and coexistence of an endoparasitoid Encarsia perniciosi andan ectoparasitoid Aphytis melinus of the California red scale. Ecol.Entomol. 15, 469–480.




















