
Comparison of Adaptive Responses in Familiar and Novel Environments: Modulatory Factors
11
PART II. GROOMING BEHAVIORS DEVELOPMENT & ENVIRONMENTAL MANIPULATION OF Comparison of Adaptive Responses in Familiar and Novel Environments: Modulatory Factors SANDRA E. FILE,' PETER S. MABBUTT, AND JACQUELINE H. WALKER MRC Neuropharmacology Research Group Department of Pharmacology The School of Pharmacy University of London, London WClN IAX, United Kingdom The links among anxiety, stress, and grooming are not clearly established. Yule' found increased grooming when rats were placed in unfamiliar cages. Thompson and Higgins' found more grooming in shocked versus unshocked rats, and Cohen and Price3 found that rats exposed to the sound of rat screams showed more grooming. These findings might suggest that increased grooming occurs in response to an increase in fear or arousal, but Doyle and Yule' found no correlation between grooming and the inci- dence of other indications of fear or anxiety, such as urination, defecation, and freezing. Gispen and IsaacsonS suggested that in fact increased grooming occurred on termina- tion of, or habituation to, a stressful situation. In support of this suggestion, Roth and Katz6found that rats placed in an open field after an hour-long exposure to loud noise and bright light showed higher levels of grooming than their unstressed controls. They found a significant change with repeated testing over days, but unfortunately the direction of the effect was not specified. The purpose of experiment 1 was to study the influence on grooming of factors that are known to modulate anxiety. In experiment 1 the time spent grooming and the type of grooming (face wash, front paw lick, body wash, genital lick, or hind limb scratch) was studied in the home cage and in the test conditions used in two animal tests of anxiety. In the social interaction test of anxiety, rats are placed in a test arena that is either familiar or unfamiliar to them and is lit by high or low light.'." In the elevated plus-maze, anxiety is generated by open elevated maze arms.9 Thus the effects on grooming of manipulating environmental familiarity, light level, and elevation were studied. Both of these tests generate behavioral changes that indicate increased anxiety and both raise plasma corticosterone concentrations. Strain and sex differences have been found in the social interaction and elevated maze and therefore compar- isons were made between male Wistar and male and female hooded Lister rats. The relationship between anxiety and grooming can also be explored by examining the effects of drugs. ACTH and CRF have both been reported to increase gro~ming,'~-'~ and these peptides also have an anxiogenic effect (shown by a specific reduction in social interaction) in the social interaction However, Roth and Katz6 found that although adrenalectomy increased grooming in the open field, hypophysectomy SEF is a Wellcome Trust Senior Lecturer. 69
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Comparison of Adaptive Responses in Familiar and Novel Environments: Modulatory Factors

PART II. GROOMING BEHAVIORS
DEVELOPMENT & ENVIRONMENTAL MANIPULATION OF
Comparison of Adaptive Responses in Familiar and Novel Environments:
Modulatory Factors SANDRA E. FILE,' PETER S. MABBUTT,
AND JACQUELINE H. WALKER MRC Neuropharmacology Research Group
Department of Pharmacology The School of Pharmacy
University of London, London WClN IAX, United Kingdom
The links among anxiety, stress, and grooming are not clearly established. Yule' found increased grooming when rats were placed in unfamiliar cages. Thompson and Higgins' found more grooming in shocked versus unshocked rats, and Cohen and Price3 found that rats exposed to the sound of rat screams showed more grooming. These findings might suggest that increased grooming occurs in response to an increase in fear or arousal, but Doyle and Yule' found no correlation between grooming and the inci- dence of other indications of fear or anxiety, such as urination, defecation, and freezing. Gispen and IsaacsonS suggested that in fact increased grooming occurred on termina- tion of, or habituation to, a stressful situation. In support of this suggestion, Roth and Katz6 found that rats placed in an open field after an hour-long exposure to loud noise and bright light showed higher levels of grooming than their unstressed controls. They found a significant change with repeated testing over days, but unfortunately the direction of the effect was not specified.
The purpose of experiment 1 was to study the influence on grooming of factors that are known to modulate anxiety. In experiment 1 the time spent grooming and the type of grooming (face wash, front paw lick, body wash, genital lick, or hind limb scratch) was studied in the home cage and in the test conditions used in two animal tests of anxiety. In the social interaction test of anxiety, rats are placed in a test arena that is either familiar or unfamiliar to them and is lit by high or low light.'." In the elevated plus-maze, anxiety is generated by open elevated maze arms.9 Thus the effects on grooming of manipulating environmental familiarity, light level, and elevation were studied. Both of these tests generate behavioral changes that indicate increased anxiety and both raise plasma corticosterone concentrations. Strain and sex differences have been found in the social interaction and elevated maze and therefore compar- isons were made between male Wistar and male and female hooded Lister rats.
The relationship between anxiety and grooming can also be explored by examining the effects of drugs. ACTH and CRF have both been reported to increase gro~ming, '~- '~ and these peptides also have an anxiogenic effect (shown by a specific reduction in social interaction) in the social interaction However, Roth and Katz6 found that although adrenalectomy increased grooming in the open field, hypophysectomy
SEF is a Wellcome Trust Senior Lecturer.
69

70 ANNALS NEW YORK ACADEMY OF SCIENCES
was without effect. Opioid antagonists have an anxiogenic effect in the social interac- tion test,lP but Roth and Katz6 found that naltrexone reduced grooming of previously stressed and of unstressed rats placed in the open field. The anxiolytic drug ethanol also increased grooming in mice,’O and so the relationship between drug-induced changes in anxiety and grooming may not be a simple one. In experiment 2, the effects of three anxiogenic drugs were examined on grooming in the home cage, in the social interac- tion test, and in the elevated maze. The three drugs were chosen because each exerts its anxiogenic effects by acting at a different receptor (see ref. 21 for review). Pen- tylenetetrazole acts at the chloride ionophore on the GABA-benzodiazepine receptor complex, FG 7142 acts at the benzodiazepine binding site, and yohimbine acts at a’ adrenoceptors.
METHODS
Animals
Male Wistar rats (home bred from Bantin and Kingman stock), male hooded Lister rats (Olac Ltd., Bicester) and female hooded Lister rats (home bred from Olac stock) were housed singly with food and water freely available. They were housed in an 11-h light : 13-h dark cycle with lights on at O600 h. The cages were opaque plastic, 35 x 24 x 17 cm, with wire grid floor and lids. The animals were housed in low light (30 scotopic lux).
Apparatus
The arena normally used for the social interaction test was a wooden box, 60 x 60 x 35 cm, with a solid floor. A camera was mounted vertically above the arena and self-grooming was scored from a monitor in an adjacent room. The observer, who was blind to the drug treatment of the rats, recorded the duration and incidence of each type of grooming. All sessions were recorded on videotape to permit rescoring. For the high light test conditions the illuminance on the floor of the box was 300 sco- topic lux and for the low light test conditions it was 30 scotopic lux.
The elevated open arm was wooden, 50 x 10 cm, and was raised 50 crn above the floor. The light level on the arm was 185 lux.
Drugs
Pentylenetetrazole (Sigma) and yohimbine (Sigma) were dissolved in water, and FG 7142 (Ferrosan) was suspended in water with a drop of ’been-20, to various con- centrations to give a constant injection volume of 2 ml/kg. Control animals received equal volume injections of water or the water-’been vehicle. All injections were in- traperitoneal.
Procedure
Experiment I
Each group of rats was randomly allocated (n = 10 per group) among the test

FILE ef a/.: FAMILIAR VS. NOVEL ENVIRONMENTS 71
conditions: home cage; elevated arm; low light, familiar arena; low light, unfamiliar arena; high light, familiar arena; high light, unfamiliar arena. In order to avoid any circadian changes all testing was conducted between 0630 and 1200 h. Each test period lasted 10 rnin and the frequency and duration of each type of grooming was recorded separately in four 2.5-min time bins. The following categories of grooming were scored: face wash; front paw lick; body wash; genital lick; hind limb scratch. Home Cage Test. In this test the rats were left in their home cages, but these were placed so that the observer could clearly see the rat. The observer sat 64 cm from the home cage and observed from above. Each rat was observed for 10 min starting im- mediately after the cage was moved. Elevated Arm. Each rat was placed on the elevated open arm for 10 min. The observer sat 64 cm away from the arm. Wooden Test Arena. Rats to be tested in the familiar arenas received the two 10-min familiarization sessions on the 2 days preceding the test, in the appropriate light level. On the test day, each rat was placed in the center of the arena and observed for 10 min.
Experiment 2
The same three test conditions were used as in experiment 1. Within each condi- tion male hooded rats were allocated randomly to control, pentylenetetrazole (15 or 30 mglkg), FG 7142 (2.5 or 5 mg/kg) or yohimbine (1.25 or 2.5 mg/kg). There were 10 rats in each condition. Injections were given 30 min before observations started for the yohimbine and FG 7142 groups and 5 rnin before for the pentylenetetrazole groups.
RESULTS
It can be seen in FIGURE 1 that, whereas there were no differences among the different rat groups in the time they spent grooming in the home cage, there were both strain and sex differences in the increase in grooming in the elevated arm. The male Wistars spent significantly longer grooming in the elevated arm than did the male hooded rats (test x strain F[1,36] = 5.48, p < 0.03), and the female hooded rats also spent longer grooming than their male counterparts (test x sex F[1,36] = 5.07, p < 0.04). FIGURE 1 also shows the distribution of grooming over the 10-min time period. There was a significant effect of time for the two male strains (F[3,108] = 5.0, p < 0.005) and a significant time x strain x test interaction (F[3,108] = 2.82, p < 0.05), because the male hooded rats showed the steepest rise at the end of the home cage test, whereas the Wistars showed more change over time in the elevated maze. The female hooded rats showed the same change over time as their male counterparts, with the steepest rise being in the last 2.5 rnin of the home cage test (see FIG. 1).
FIGURE 2 shows the time spent grooming in each of the four types of wooden test arena. There was a significant effect of light level (F[1$5] = 9.41, p < 0.005). with less grooming in the high light test conditions. The familiarity of the test arena was without effect on the time spent grooming (F[1,35] = l.Ol), nor were any of the inter- actions with strain, sex, or light significant. There was a significant effect of the time spent in the test situation for the two male strains (F[3,105] = 11.16, p < 0.OOOl). There were also significant interactions for time x strain (F[3,105] = 2.85, p < 0.05) and time x strain x familiarity x light (F[3.105] = 3.57, p < 0.02); this was because the
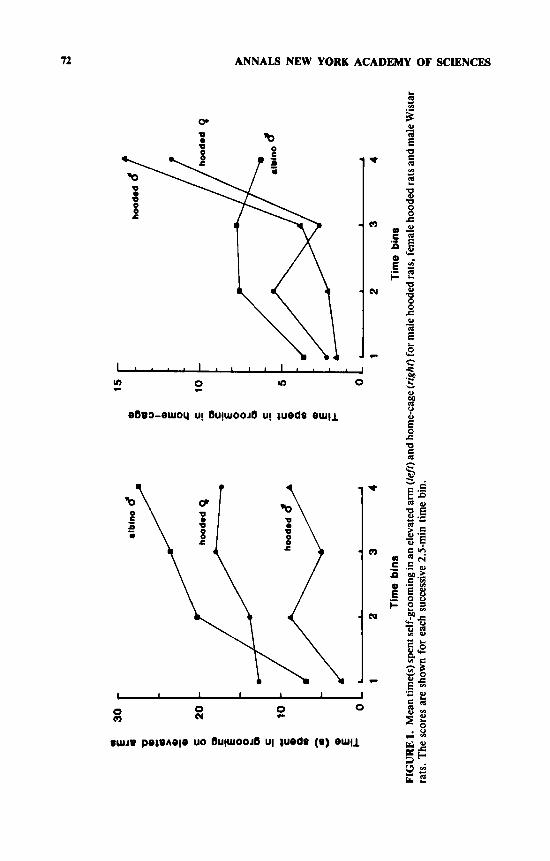
72 ANNALS NEW YORK ACADEMY OF SCLENCES
0 5 0 (Y 0
(3
Y) 3
e

FILE et ul.: FAMILIAR VS. NOVEL ENVIRONMENTS
50
40 - Y P '$ 30
i
* 2 0 - a
- a a
i=
10
0 -
13
-
-
-
- J 11.I - 1 2 3 4 1 2 3 4 T l m blni
LOW ugh1 * V d
Unfrmlllar Funllkr
FIGURE 2. Mean time(s) spent in self-grooming by rats placed for 10 min in an unfamiliar or familiar test arena that was lit by high or by low light. The scores are shown for each succes- sive 2.5-min time bin and for male Wistar. male hooded Lister, and female hooded Lister rats.
Wistars showed more change with time in the low light conditions and the hoodeds showed more change in the high light unfamiliar condition (see FIG. 2). The significant familiarity x time interaction (F[3,105] = 4.97, p < 0.005) reflects the different time course seen in unfamiliar and familiar conditions. In the unfamiliar arena there was a steady increase over each successive time period, whereas in the familiar test arenas the peak time spent grooming was the second or third time period (see FIGURE 2). The significant time x light interaction (F[3,105] = 5 . 5 4 , ~ < 0.005) reflects the much greater rise of grooming over the test period in the low light conditions (see FIG. 2). The fe- male hooded rats showed significantly more grooming than the males (F[1,35] = 4.49, p < 0.05) and there was a significant familiarity x time x sex X light interaction (F[3,105] = 5.20, p < 0.005), because the females showed the greatest rise with time in the high,light, unfamiliar condition (see FIG. 2).
RGURE 3 shows the frequency with which each type of grooming occurred for the three groups of rats in all the test conditions. It is clear from this figure that the most common forms of grooming in all the conditions were face wash and forepaw lick.
FIGURE 4 shows the time spent grooming in the home cage and in the elevated arm for male hooded rats injected with vehicle or with pentylenetetrazole or FG 7142 5 or 30 min prior to observation. As can be seen, pentylenetetrazole increased the time spent grooming in both the home cage and the elevated arm (Ft2.54) = 13.38, p <

I,
!! 0 a
ME
AN
FR
EQ
UE
NC
Y
OF
EA
CH
TY
PE
A
ow
*a
O
NP
dD
0,
O
II
11
11
11
1
11
11
11
11
.1
1
OF
GR
OO
MIN
G 9
- cr a 0
4
P

FILE ef 01.: FAMILIAR VS. NOVEL ENVIRONMENTS 75
50
40
30
20
10
Home cage Elevated arm
PTZ FG PTZ FG
0 1530 2.5 5
.... ... .... .... .:.;.:. *.*.*I
I ..*.'. ............. ......... .......... ........... ....
....... .............. .............. ....... .:.;.; ..*.*.* .:.:.;. .:.:.; ;.;.;.; ... ............ :.;.;. ....... ............ ............ ...... .;I.;
.......... .........
FIGURE 4. Mean time(s) spent in self-grooming in the home cage ( lef) a n d in the elevated arm (right) for rats injected 5 min before with pentylenetetrazole (PTZ, 15 o r 30 mg/kg) o r 30 min before with FG 7142 (FG, 2.5 or 5 mglkg).
0.o00l). There was a significant effect of time (F[3,162] = 3.88, p < 0.02) and a time x pentylenetetrazole interaction (F[6,162] = 2.35, p < 0.05), because the higher dose group showed a particularly marked increase with time in both the home cage and the elevated arm. FG 7142 also caused a significant increase in the time spent grooming (F[2,54] = 5.63, p < 0.01), and there was a significant time x FG x test condition interaction (F[6,162] = 2.20, p < 0.05), because the increase in grooming in the rats treated with FG 7142 occurred particularly in the last time period of the home cage test. There was no overall effect of yohimbine on the time spent grooming, however there was a test x yohimbine interaction of borderline significance, because yohim- bine increased the time spent grooming only in the home cage (F[2,53] = 2.39, p = 0.10). Although the increase in time spent grooming was not significant, there was a significant increase in the frequencies of front paw licking and body washing in the home cage after yohimbine (p < 0.05, see FIGURE 5) . Pentylenetetrazole and FG 7142 also selectively increased these forms of grooming in the home cage (p < 0.05). leaving unchanged the incidence of other types of grooming.
Pentylenetetrazole (30 mg/kg) significantly decreased the time spent grooming in the social interaction test (F[2,53] = 3.69, p < 0.05). but neither FG 7142 nor yohim- bine had a significant effect (in both cases F < 1.0). There was no significant change in the frequency of any type of grooming with administration of anxiogenics in this test.
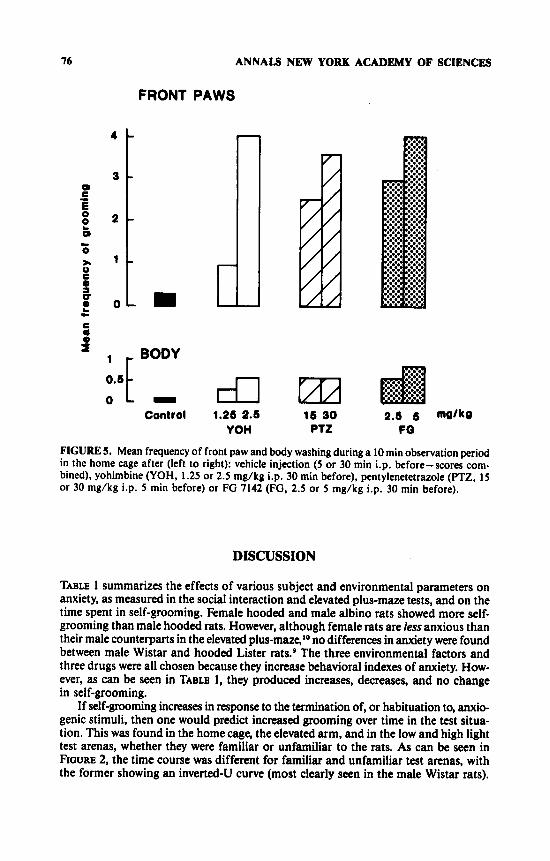
76 ANNALS NEW YORK ACADEMY OF SCIENCES
0 1
FRONT PAWS
r m L
r O * . " t 0 m
Control 1.26 2.6 YOH
16 30 2.6 6 malkg PTZ FQ
FIGURE 5. Mean frequency of front paw and body washing during a 10 min observation period in the home cage after (left to right): vehicle injection (5 or 30 min i.p. before-scores com- bined), yohimbine (YOH, 1.25 or 2.5 mg/kg i.p. 30 min before), pentylenetetrazole (PTZ, 15 or 30 mg/kg i.p. 5 min before) or FG 7142 (FG, 2.5 or 5 mg/kg i.p. 30 min before).
DISCUSSION
TABLE 1 summarizes the effects of various subject and environmental parameters on anxiety, as measured in the social interaction and elevated plus-maze tests, and on the time spent in self-grooming. Female hooded and male albino rats showed more self- grooming than male hooded rats. However, although female rats are less anxious than their male counterparts in the elevated plus-maze,*O no differences in anxiety were found between male Wistar and hooded Lister rats.s The three environmental factors and three drugs were all chosen because they increase behavioral indexes of anxiety. How- ever, as can be seen in TULR l, they produced increases, decreases, and no change in self-grooming.
If self-grooming increases in response to the termination of, or habituation to, anxio- genic stimuli, then one would predict increased grooming over time in the test situa- tion. This was found in the home cage, the elevated arm, and in the low and high light test arenas, whether they were familiar or unfamiliar to the rats. As can be seen in FIGURE 2, the time course was different for familiar and unfamiliar test arenas, with the former showing an inverted4 curve (most clearly seen in the male Wistar rats).

FILE el a&: FAMILIAR VS. NOVEL ENVIRONMENTS 77
TABLE 1. Summary of Effects of Sex, Strain, Light, Familiarity, Elevation and Three Anxiogenic Drugs on Self-Grooming and on Behavioral Indexes of Anxiety
Grooming Anxiety Sex: females t 4 Strain: Wistar t - Light: high .1 t
t Familiarity: unfamiliar - Elevation t t PTZ t t FG 7142 t t
t Yohimbine -
-
-
-
t, Increase. 4, Decrease. = , No change.
This suggests that with increased time in the situation and hence habituation to stress, self-grooming eventually decreases. It also suggests that the habituation occurs both within and between test sessions. Thus, although the overall time spent grooming in the familiar and unfamiliar arenas did not differ, the distribution of grooming over time was different.
If grooming is elicited in response to habituation to stress, it is extremely difficult to make any overall predictions about the effects on grooming of any particular anxio- genic stimulus. The overall level of grooming will be the joint result of the level of stress generated, the rate of habituation to the stressful stimuli, and the duration of the test period. Thus there was greater self-grooming in the elevated arm than in the home cage, because the former was more stressful and sufficient within-session habit- uation to the stress occurred for the rise in grooming to be manifest. The difference in distribution over time of grooming in the high and low light test arenas suggests that the male rats were habituating more slowly to the high light. If the test session had been prolonged to, say 1 hour, then one would predict a gradual rise in self-grooming in the high light, and a decrease in the low light. However, what is interesting is that the within-session decrease in grooming occurs (at least for the male Wistar rats) in the high light, familiar condition without high levels of grooming ever being reached. This suggests between-session habituation may have different effects on grooming than within-session habituation. In general, the changes in grooming as a result of our en- vironmental manipulations are consistent with self-grooming being a response to habit- uation to anxiogenic stimuli. With relatively low levels of stress the rat can habituate to the stress within the Wmin test session. Thus for stimuli within this range there is more grooming as the stress is increased. Anxiogenic conditions enhance self-grooming because there is within-session habituation to the stimuli. With higher levels of stress the overall increase in self-grooming is less because of slower within-session habituation.
Can this hypothesis account for our sex and strain differences? We would have to predict that female hooded and male Wistar rats would show more rapid habitua- tion to anxiogenic or stressful stimuli than male hooded rats. Unfortunately there are no data yet on this question. On the basis of habituation to stress, what can be made of the data on self-grooming in response to anxiogenic drugs? First it must be assumed that the anxiogenic effects of the drug and the test session are additive. PTZ was in- jected only 5 minutes before the test began, and therefore there was little opportunity for the rats to habituate to the anxiogenic effects of the drug before the test. The in-

78 ANNALS NEW YORK ACADEMY OF SCIENCES
creased grooming in PTZ-injected rats, compared with controls in the home cage and elevated arm, suggests that considerable within-session habituation occurred to the anxiogenic stimuli generated by the drug. The significant decrease in grooming seen with the combination of the higher dose of PTZ and the social interaction test arena could be explained if the combined stress of PTZ and the test arenas was sufficient to slow habituation. Both yohimbine and FG 7142 were injected 30 min before testing began. The failure of yohimbine to change self-grooming could be accounted for if complete habituation had occurred in this time to the anxiogenic effects of the drug. From previous experiments we know that it is still possible to detect anxiogenic effects of yohimbine 30 min after injection,I6 but it is possible that self-grooming is triggered by a higher level of stress than is needed to reduce social interaction. The results with FG 7142 would suggest that habituation to its anxiogenic effects was not complete by the time the tests began. Thus, the combination of FG 7142 and the wooden test arenas was stressful enough to decrease the rate of habituation, whereas habituation to the drug could still occur in the elevated arm and the home cage.
In conclusion. the results of these experiments suggest that there is no simple rela- tionship between anxiogenic conditions and the incidence of self-grooming. The results can be reasonably well explained if self-grooming is a response to habituation to anxio- genic stimuli. Given equal rates of habituation, within a given time period the more anxiogenic stimuli will generate more self-grooming. But, at least above a certain level of stress, there seems to be an interaction between the level of anxiety and the rate of habituation. Studies of the time course of grooming over a prolonged period are necessary to verify this hypothesis. Additionally, further studies are needed to clarify the relationship between between-session habituation and grooming.
1. 2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
12.
13.
REFERENCES
YULE, E. P. 1957. The open field test. Proc. S. Afr. Psychol. Assoc. THOMPSON, W. R. & W. H. HIOOINS. 1958. Emotion and organised behavior. Can. J. Psy-
COHEN, J. A. & E. 0. PRICE. 1979. Grooming in the Norway rat: Displacement activity or “boundary shift”? Behav. Neural Biol. M: 177-188.
DOYLE, G. & E. P. YULE. 1959. Grooming activities and freezing behaviour in relation to emotionality in albino rats. Anim. Behav. 7: 18-22.
GISPEN, W. H. & R. L. ISMCSON. 1981. ACTH-induced grooming in the rat. Pharmacol. Ther. 12: 209-224.
ROTH, K. A. & R. J . KATZ. 1979. Stress, behavioral arousal, and open field activity-a reexamination of emotionality in the rat. Neurosci. Biobehav. Rev. 3: 247-263.
FILE, S.E. 1980. The use of social interaction as a method for detecting anxiolytic activity of chlordiazepoxide-like drugs. J. Neurosci. Methods 2: 219-238.
FILE, S. E. 1985. Animal models for predicting clinical efficacy of anxiolytic drugs: Social behaviour. Neuropsychobiology 13: 55-62.
PELLOW, S., P. CHOPIN, S. E. FILE & M. BRILEY. 1985. The validation of open:closed arm entries in an elevated plus-maze as a measure of anxiety in the rat. J . Neurosci. Methods
FILE, S. E. & S. V. VELLUCCI. 1979. Behavioural and biochemical measures of stress in hooded rats obtained from different sources. Physiol. Behav. 22: 31-35.
JOHNSTON, A. L., S. E. FILE, F. FARAEIOLLINI & C. A. Wilson. 1986. The advantages of a female brain: Decreased anxiety and increased activity. Horm. Behav. Submitted.
DUNN, A. J., E. J. GREEN & R. L. ISMCSON. 1979. Intracerebral adrenocorticotropic hor- mone mediates novelty-induced grooming in the rat. Science 203: 281-283.
GISPEN, W. H., V. M. WIEGANT, H. M. GREVEN & D. DE WIED. 1975. The induction of
chol. 12: 61-68.
14: 149-167.

FILE el al.: FAMILIAR VS. NOVEL ENVIRONMENTS 79
14.
15.
16.
17.
18.
19.
20.
21.
excessive grooming in the rat by intraventricular application of peptides derived from ACTH: Structure-activity studies. Life Sci. 17: 645-652.
MORLEY, J. & A. S. LEVINE. 1982. Corticotropin-releasing factor, grooming and ingestive behavior. Life Sci. 31: 1459-1464.
VELDHUIS, H. D. & D. DE WIED. 1984. Differential behavioral actions of corticotropin- releasing factor (CRF). Pharmacol. Biochem. Behav. 21: 707-713.
DUNN, A. J . & S. E. FILE. 1986. Corticotropin-releasing factor displays an anxiogenic ac- tion in the social interaction test. Horm. Behav. Submitted.
FILE, S. E. & A . CLARKE. 1980. lntraventricular ACTH reduces social interaction in male rats. Pharmacol. Biochem. Behav. 12: 855-859.
FILE, S. E. & S. V. VELLUCCI. 1978. Studies on the role of ACTH and of 5-HT in anxiety using an animal model. J . Pharm. Pharmacol. 30: 105-110.
FILE, S. E. 1980. Naloxone reduces social and exploratory activity in the rat. Psychophar- macology 71: 41-44.
ALLAN, A. M. & R. L. ISAACSON. 1985. Ethanol-induced grooming in mice selectively bred for differential sensitivity to ethanol. Behav. Neural Biol. 44: 386-392.
PELLOW, S. & S. E. FILE. 1984. Multiple sites of action for anxiogenic drugs: Behavioural, electrophysiological and biochemical correlations. Psychopharmacology 83: 304-315.




















